
Mechanism for alternating access in neurotransmitter transporters
Upload
independentCategory
view
1download
0
1 3
Exp Brain Res (2013) 226:153–163DOI 10.1007/s00221-013-3421-8
RESEARCH ARTICLE
Time-dependent modulation of AMPA receptor phosphorylation and mRNA expression of NMDA receptors and glial glutamate transporters in the rat hippocampus and cerebral cortex in a pilocarpine model of epilepsy
Mark William Lopes · Flávia Mahatma Schneider Soares · Nelson de Mello · Jean Costa Nunes · Aurilene Gomes Cajado · Daniel de Brito · Fabiano Mendes de Cordova · Rodrigo Maranguape Silva da Cunha · Roger Walz · Rodrigo Bainy Leal
Received: 13 June 2012 / Accepted: 15 January 2013 / Published online: 8 February 2013 © Springer-Verlag Berlin Heidelberg 2013
the hippocampus (Hip) and cerebral cortex (Ctx) at differ-ent time points after pilocarpine-induced status epilepticus (Pilo-SE) in male adult Wistar rats. Biochemical analy-sis was performed in the Hip and Ctx at 1, 3, 12 h (acute period), 5 days (latent period), and 50 days (chronic period) after Pilo-SE. Key findings include an increase in the phos-phorylation of GluR1-Ser845 in the Ctx and GluR1-Ser831 in the Hip at different times during the acute period, and a decrease in the total content of the GluR1 subunit in the Ctx in the latent period. There was a down-regulation of the mRNA expression and protein levels of EAAT1 and EAAT2, and a decrease of the NR1 mRNA expression, in the Ctx during the latent period. Notably, during the chronic period, the EAAT2 mRNA expression and protein levels decreased while the NR1 mRNA levels increased in the Hip. Taken together, our findings suggest a time- and structure-dependent imbalance of glutamatergic transmission in response to Pilo-SE, which might be associated with either epileptogenesis or the seizure threshold in MTLE-HS.
Keywords Epilepsy · Pilocarpine · Signal transduction · AMPA and NMDA receptors · EAATs
Introduction
Epilepsies are one of the most common neurological condi-tions, affecting approximately 50 million people worldwide (Sander 2003). Twenty to thirty percent of epilepsies are medically intractable. Mesial temporal lobe epilepsy related to hippocampal sclerosis (MTLE-HS) is the foremost form of surgically remediable epileptic syndrome (Babb 1999).
Systemic administration of pilocarpine in rodents repro-duces the main features of human MTLE-HS (Cavalheiro et al. 1991; Hollmann et al. 1991). Pilocarpine induces
Abstract The pilocarpine model in rodents reproduces the main features of mesial temporal lobe epilepsy related to hippocampus sclerosis (MTLE-HS) in humans. It has been demonstrated in this model that the phosphorylation of the alpha-amino-3-hydroxy-5-methyl-4-isoxazolepro-pionic acid (AMPA) receptor GluR1 subunit is increased 1 h after pilocarpine treatment. Moreover, alterations in the levels of glutamate transporters have been associated with chronic epilepsy in humans. Despite these studies, the profile of these changes has not yet been addressed. We analyzed the protein content and phosphorylation profile of the AMPA receptor GluR1 subunit by western blotting. We also used quantitative real-time polymerase chain reac-tion to analyze the expression of glial glutamate transport-ers and the N-methyl-d-aspartate receptor NR1 subunit in
M. W. Lopes · F. M. de Cordova · R. B. Leal (*) Departamento de Bioquímica, Centro de Ciências Biológicas, Universidade Federal de Santa Catarina, Florianópolis, SC 88040-900, Brazile-mail: [email protected]
F. M. S. Soares · N. de Mello · J. C. Nunes · R. Walz Centro de Neurociências Aplicadas (CeNAp), HU, Universidade Federal de Santa Catarina, Florianópolis, SC, Brazil
A. G. Cajado · D. de Brito · R. M. S. da Cunha Centro de Ciências Agrárias e Biológicas, Universidade Estadual Vale do Acaraú, Fortaleza, CE, Brazil
F. M. de Cordova Escola de Medicina Veterinária e Zootecnia, Universidade Federal do Tocantins, Araguaína, TO, Brazil
R. Walz Departamento de Clínica Médica, Hospital Universitário (HU), Centro de Ciências da Saúde, Universidade Federal de Santa Catarina, Florianópolis, SC, Brazil

154 Exp Brain Res (2013) 226:153–163
1 3
limbic seizures that become secondarily generalized, evolv-ing to status epilepticus (SE), which lasts for several hours (acute period). The SE is followed by a latent “seizure-free” period and by a chronic period characterized by the presence of spontaneous recurrent seizures (Cavalheiro et al. 1991, 1994; Mello et al. 1993). Hippocampal changes following SE include an increase in glutamate in the synap-tic cleft (Costa et al. 2004), cell loss, gliosis and mossy fiber sprouting (Mello et al. 1993). Neurochemical changes such as an imbalance between inhibitory (γ-aminobutyric acid (GABA)-mediated) and excitatory (glutamate-mediated) neurotransmission (in favor of the latter) have been iden-tified in brain tissue obtained from animal models and humans treated surgically for MTLE-HS. However, the in vivo cellular and molecular events that occur during the time course of the epileptogenic process and in the chronic period are still poorly understood (Houser et al. 2008; Li et al. 2010), because the studies have been predominantly performed only during the acute period. With respect to this point, we have recently described a time-dependent profile of mitogen-activated protein kinases (MAPKs) and AKT modulation in the rat hippocampus and cerebral cortex in the pilocarpine model (Lopes et al. 2012). This profile could represent some of the biochemical alterations that lead to the synaptic reorganization and cell death observed in epilepsy.
Alpha-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) and N-methyl-d-aspartate (NMDA) receptors mediate excitatory synaptic transmissions that are crucial for physiological neuroplasticity; however, these receptors are also involved in a range of pathologic conditions, including epileptogenesis (Jerusalinsky et al. 1992; Quillfeldt et al. 1994; Sanderson and Dell’Acqua 2011). AMPA and NMDA receptors are integrated in a complex signaling network that also involves the activity of a variety of protein kinases, such as the cAMP-dependent protein kinase (PKA) (Bernabeu et al. 1997), Ca2+-calmodulin-dependent protein kinases II (CaMKII) (Cammarota et al. 1998), protein kinase C (PKC) (Bernabeu et al. 1995), and MAPKs (Walz et al. 2000; Sweatt 2004; Thomas and Huganir 2004). We have previ-ously demonstrated that extracellular signal-regulated kinases (ERK) and p38MAPK can be activated rapidly after pilocar-pine-induced status epilepticus (Pilo-SE) (Lopes et al. 2012). The activation mechanism was not addressed by our study, but this process could result from the modulation of the glu-tamatergic system (Walz et al. 2000; Thomas and Huganir 2004). Therefore, despite the well-known physiologic role of these signaling systems in learning and memory, their altera-tion may be associated with a pathologic synaptic plasticity leading to epileptogenesis (Sanderson and Dell’Acqua 2011).
High-frequency presynaptic stimulation (HFS) induces NMDAR-dependent long-term potentiation (LTP) by increas-ing synaptic AMPAR number and activity, whereas low-frequency stimulation (LFS) induces NMDAR-dependent
long-term depression (LTD) by decreasing the AMPAR activity and number. During NMDAR-induced LTP, brief but strong elevations in postsynaptic Ca2+ promote the phosphorylation-regulated insertion of AMPARs contain-ing GluR1 into extrasynaptic and perisynaptic plasma membrane sites (Sanderson and Dell’Acqua 2011). The mechanism may involve a PKA-dependent phosphoryla-tion of the Ser845 residue on the GluR1 subunit of AMPARs that induces its insertion into the plasma membrane in the extrasynaptic region, followed by the GluR1-Ser831 phos-phorylation by CaMKII or PKC that drives the receptor into the synaptic region (Esteban et al. 2003; Sanderson and Dell’Acqua 2011). In contrast, NMDAR-induced LTD is triggered by prolonged but low-level Ca2+ elevations that promote GluR1-Ser845 dephosphorylation, decreas-ing synaptic AMPAR activity and number (Lee et al. 1998; Banke et al. 2000). Modulation of synaptic strength may be involved in epileptogenesis (Leite et al. 2005), as well as in compensatory mechanisms in the distant regions of the epileptogenic zone.
The glutamate released at the synaptic cleft is captured by excitatory amino acid transporter (EAAT) proteins (Beart and O’Shea 2007). These transporters are expressed differ-entially throughout the central nervous system (CNS), but EAAT1/GLAST and EAAT2/GLT-1 are normally expressed by astrocytes. These proteins are considered to play a criti-cal role in the maintenance of low extracellular concentra-tions of glutamate, which protects neurons from excitotoxic injury (Lee and Pow 2010). The disturbance of glutamate transporter activity appears to be involved in various neuro-logical disorders, including epileptogenesis (Rakhade and Loeb 2008).
Studies regarding the phosphorylation profile of GluR1 and the mRNA expression of the NR1 subunit, as well as the glial glutamate transporters (in the acute, latent and chronic period of pilocarpine model), may be relevant to charac-terize possible key modulatory elements for each phase in the epileptogenic processes. Therefore, the main aims of our study were twofold: to determine the phosphorylation profile of Ser831 and Ser845 on the GluR1 subunit of AMPA receptors, and to analyze the expression of glial glutamate transporters EAAT1 and EAAT2, and the NR1 subunit of the NMDA receptor in the hippocampus and cerebral cortex of rats during different periods in the pilocarpine model of epilepsy.
Materials and methods
Chemicals
HEPES, Triton X-100, SDS, acrylamide, and bis-acryla-mide, were obtained from GE Healthcare Life Science

155Exp Brain Res (2013) 226:153–163
1 3
(Piscataway, NJ, USA). Glycine, Tris, TEMED, and β mercaptoethanol were obtained from Amresco Life Sci-ence (Solon, OH, USA). Bovine serum albumin (BSA) was from Inlab (São Paulo, SP, Brazil). Immobilon nitrocellu-lose, goat anti-mouse IgG HRP (horseradish peroxidase)-conjugated (AP308P), goat anti-rabbit IgG HRP (AP132P), and rabbit anti-phospho-GluR1 Ser831 (04-823) and Ser845 (04-1073) antibodies were purchased from Millipore (Billerica, MA, USA). Ammonium persulfate (APS) was from the Sigma Chemical (St. Louis, MO, USA). Lumi-GLO reagent (luminol chemiluminescent substrate), rabbit anti-EAAT1 (#4166), and anti-EAAT2 (#3838) antibodies were obtained from Cell Signaling Technology (Beverly, MA, USA). Mouse anti-GluR1 (sc-13152) and anti-β actin (sc-47778) antibodies were obtained from Santa Cruz Bio-technology (Santa Cruz, CA, USA). All other reagents were of analytical grade.
Experimental procedures
Adult male Wistar rats (90 days old) from the Federal Uni-versity of Santa Catarina (UFSC) breeding colony were maintained in groups of 4 per cage, at a room temperature of between 22 °C and 25 °C, on a 12 h light/dark cycle, with water and food available ad libitum. All experimental procedures were approved by the UFSC Ethics Committee for Animal Experimentation. The induction of status epilep-ticus (SE) by pilocarpine was performed as described previ-ously (Cavalheiro et al. 1991; Mello et al. 1993; Bonan et al. 2000). Scopolamine methyl nitrate (dissolved in saline 0.9 %; Sigma Chemical) was administered subcutaneously at 1 mg/kg for 30 min before pilocarpine administration to reduce peripheral cholinergic effects. Animals received a single intraperitoneal dose of pilocarpine (280 mg/kg dis-solved in 0.9 % saline; Sigma Chemical). Following pilo-carpine administration, most animals became hypoactive, a state that in most cases evolves to generalized convulsions and limbic SE within 40–80 min after the injection. Two hours after the seizure onset, animals received diazepam (5 mg/kg) to decrease the mortality rate. Assisted feeding and hydration were undertaken during the recovery period to improve general clinical state and to reduce mortality. In this model, after a latent period of 14 days (ranging from 4 to 44 days), all animals developed spontaneous seizures (2–15 per month), characterizing the chronic period. In the 80 animals injected with pilocarpine, the mortality rate was 20 % (n = 16), and 5 % (4 animals) did not evolve to SE. Only animals that evolved to SE (n = 60, 75 %) were used in the biochemical analysis.
Survivor animals were divided in 5 groups: (1) the acute group in which animals were euthanized 1 h after status epilepticus (SE) onset (Acute 1 h Group; n = 12); (2) the acute group in which animals were euthanized 3 h after SE
onset (Acute 3 h Group; n = 12); (3) the acute group in which animals were euthanized 12 h after SE onset (Acute 12 h Group; n = 12); (4) the latent group in which animals were euthanized 5 days after SE onset (Latent 5 days Group; n = 12), a period during which seizures were not observed; and (5) the chronic group in which animals were eutha-nized 50 days after SE induction (Chronic 50 days Group; n = 12). The animals from the latent group were behavio-rally evaluated for 6 h per day (0800–1400 hours) between days 3–5 and no clinically visible seizure were documented. The control animals were sacrificed 1, 3, and 12 h, and 5 and 50 days (n = 12 per group) after scopolamine, diazepam, and saline injections. However, the animals from the Acute 1 h Group and their respective controls did not receive diaz-epam. The comparisons were performed only within each group (between control and treated animals in the respective period) because the treatments were performed on different days. Therefore, Student’s t test was applied for independ-ent samples in the statistical analysis. The hippocampus and cerebral cortex were dissected from animals in the control and experimental groups. Analysis of EAAT1/GLAST, EAAT2/GLT1, and NR1 mRNA expression was performed by quantitative real-time polymerase chain reaction (qRT-PCR) (n = 4 animals per group). The total levels of the AMPA receptor GluR1 subunit, EAATs and phosphorylation at the Serine831 (GluR1-Ser831) and Serine845 (GluR1-Ser845) sites were analyzed by western blotting (n = 8 animals per group).
Western blot analysis
For quantification of phosphorylation on the Ser831 and Ser845 sites on the AMPA receptor GluR1 subunit and immunocontent of EAATs, western blot analysis was per-formed as previously described (Leal et al. 2002; Cordova et al. 2004; Posser et al. 2007; Figueiredo et al. 2011; Lopes et al. 2012). Animals were euthanized by decapitation and the brains were excised from the skull. The hippocampus and cerebral cortex were dissected (4 °C) into “cutting solu-tion” (110 mM sucrose, 60 mM NaCl, 3 mM KCl, 1.25 mM KH2PO4, 0.5 mM CaCl2, 7 mM Mg2SO4, 5 mM glucose; 25 mM HEPES, pH 7.4) and placed in liquid nitrogen for storage at −80 °C until use. The samples were prepared as previously described (Oliveira et al. 2008). Briefly, the samples were mechanically homogenized in 400 μl of 50 mM Tris, pH 7.0, 1 mM EDTA, 100 mM NaF, 0.1 mM PMSF, 2 mM Na3VO4, 1 % Triton X-100, 10 % glycerol, and Protease Inhibitor Cocktail (P2714; Sigma) and were then incubated on ice for 10 min. Lysates were centrifuged (10,000g at 4 °C for 10 min) to eliminate cellular debris. The supernatants were diluted 1/1 (v/v) in 100 mM Tris, pH 6.8, 4 mM EDTA, and 8 % SDS and were then boiled for 5 min. Thereafter, the loading buffer (40 % glycerol,

156 Exp Brain Res (2013) 226:153–163
1 3
100 mM Tris, bromophenol blue, pH 6.8) in the ratio 25:100 (v/v) and β-mercaptoethanol (final concentration 8 %) were added to the samples. Protein content was estimated by the method described by Peterson (1977). The same amount of protein (70 μg per lane) for each sample was electro-phoresed in 10 % sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) minigels and transferred to nitrocellulose membranes using a semidry blotting appa-ratus (1.2 mA/cm2; 1.5 h). To verify the transfer efficiency process, the gels were stained with Coomassie blue and the membranes were stained with Ponceau S.
The membranes were blocked with 5 % skim milk in Tris-buffered saline (TBS) (10 mM Tris, 150 mM NaCl, pH 7.5). GluR1 phosphorylated, total forms, EAAT1 and EAAT2 were detected after overnight incubation with specific anti-bodies diluted in TBS with tween (TBS-T) that contained 2 % BSA in a 1:1,000 dilution (anti-phospho-GluR1-Ser831, anti-phospho-GluR1-Ser845, anti-total-GluR1, anti-EAAT1, and anti-EAAT2). Moreover, the membranes were incu-bated for 1 h at room temperature with horseradish peroxi-dase (HRP)-conjugated anti-rabbit or anti-mouse secondary antibodies for the detection of the phosphorylated sites or the total form of GluR1, respectively. The reactions were developed by chemiluminescence substrate (LumiGLO). All blocking and incubation steps were followed by wash-ing three times (5 min) with TBS-T (10 mM Tris, 150 mM NaCl, 0.1 % Tween-20, pH 7.5). All membranes were incu-bated with mouse anti-β-actin (1:2,000) antibody to verify that equal amounts of protein for each sample were loaded on the gel. The phosphorylation level of GluR1 and immu-nocontent of EAATs were determined as a ratio of the optic density (OD) of the phosphorylated band (GluR1) and pro-tein band (EAATs)/the OD of the total band (GluR1) and β-actin band (EAATs) (Calloni et al. 2005; Posser et al. 2007). The bands were quantified using the Scion Image® software, which is a derivative of NIH Image (Frederick, MD, USA).
The antibodies against GluR1, GluR1-Ser831, and GluR1-Ser845 detected a single band of approximately 100 kDa, anti-EAAT1 and Anti-EAAT2 antibodies detected a single band of approximately 58 and 65 kDa, respectively, and the anti-β-actin antibody detected a single band of approxi-mately 45 kDa.
Total RNA isolation
For determination of mRNA levels of EAAT1, EAAT2, and the NR1 subunit, animals were euthanized by decapitation, the brains were excised from the skulls, and the hippocam-pus and cerebral cortex were quickly dissected and placed in liquid nitrogen for storage at −80 °C until use. Thawed tis-sues were homogenized in 1 ml (cerebral cortex) and 500 μl (hippocampus) volumes of TRIzol® reagent (Invitrogen/
Life Technologies, Grand Island, NY, USA), and total RNA was extracted accordingly to the manufacturer’s instructions and the methods of RNA isolation developed by Chomczyn-ski and Sacchi (1987).
Quantitative real-time RT-PCR
One microgram of total RNA was used for complementary DNA (cDNA) synthesis and qRT-PCR gene expression analysis. Reverse transcription (RT) was then carried out in a 20-μl reaction in the presence of 2 μl, 5 mM deoxy-nucleotide triphosphates (dNTPs), 1 μl oligo primer, and SuperScript® III RT (Invitrogen) (200 U/μl). The reactions conditions were as follows: 65 °C for 5 min, cool at 20 °C for 1 min, 50 °C for 60 min and 70 °C for 15 min.
The reaction product was amplified by qRT-PCR on the thermocycler RealPlex 4S (Eppendorf, Hauppauge, NY, USA) using the SYBR Green core reaction kit (Applied Biosystems/Life Technologies, Grand Island, NY, USA). The thermal cycling conditions were 95 °C for 10 min, fol-lowed by 40 cycles at 95 °C for 15 s, 55 °C for 15 s, and 60 °C for 20 s. This procedure was followed by the melt-ing curve at 95 °C for 15 s, 55 °C for 15 s, and the gradual increase of temperature for 20 min ending with 95 °C for 15 s. Experiments were performed in quadruplicates for each data point. The abundance of EAAT1, EAAT2, and NR1 mRNAs was quantified as relative values compared with an internal reference, glyceraldehyde 3-phosphate dehydrogenase (GAPDH), whose abundance was believed not to change between the varying experimental conditions. The sequences for the primers used for qRT-PCR are as follows: rat EAAT1 (Gen-Bank™ accession number NM 148938), forward primer 5′-CTCGTCACAGGAATGGCG GCC-3′ and reverse primer 5′-TGCCCTTTCCGGGGTG GATGA-3′; rat EAAT2 (Gen-Bank™ accession number NM 0113932), forward primer 5′-CCGTGATGATAGGCAC CGTG-3′ and reverse primer 5′-CTCCAGCAGCGTC CATTCAA-3′; rat NR1 (GenBank™ accession number NM 0081692), forward primer 5′-TGGTAGAGCAGAGCC CGACCC-3′ and reverse primer 5′-CCCCGGTGCTCGT GTCTTTGG-3′; and rat GAPDH (GenBank™ accession number NM 017008) forward primer 5′-GGGGGCTCTCT GCTCCTCCC-3′ and reverse primer 5′-CGGCCAAATC CGTTCACACCG-3′. A 2-μl RT reaction was used for qRT-PCR.
Quantitative values for the transcription of EAAT1, EAAT2, NR1, and GAPDH mRNA were obtained from the threshold cycle number, where the increase in the sig-nal associated with an exponential growth of PCR products begins to be detected. Melting curves were generated at the end of every run to ensure product uniformity. The levels of the target gene were normalized using GAPDH expres-sion as an endogenous RNA control. The ΔCt values of the

157Exp Brain Res (2013) 226:153–163
1 3
samples were determined by subtracting the average Ct val-ues of the target gene mRNAs from the average Ct values of the internal GAPDH control. The ΔΔCt was determined by subtracting the average ΔCt values of the treated group from the average ΔCt values of the control group. As it is uncommon to use ΔΔCt as relative data due to this logarith-mic characteristic, the 2−ΔΔCt parameter was used to express the relative expression data (Livak and Schmittgen 2001).
The efficiency assessment for the primers used in SYBR Green-based qRT-PCR was 0.9 for EAAT1, EAAT2 and NR1. Thus, the Pfaffl method was also used to evaluate the relative expression variations. The mRNA expression of NR1 retained the same profile of the ΔΔCt method of Livak and Schmittgen. The results of glial glutamate trans-porters (EAATs) exhibited only a tendency when analyzed by Pfaffl method, and then the data that were being sug-gested by PCR by ΔΔCt method (Livak and Schmittgen) were confirmed by analysis of the proteins by western blot-ting (see “Results”).
Statistical analysis
Phosphorylation and expression of the GluR1 subunit and the levels of EAAT1, EAAT2, and the NR1 subunit were compared within each group, during different experimental periods, using Student’s t test for independent samples. The data are presented as the mean ± SEM.; a p value <0.05 was considered to be statistically significant.
Results
Phosphorylation levels of the AMPA receptor GluR1 subunit in the hippocampus and cerebral cortex after Pilo-SE induction
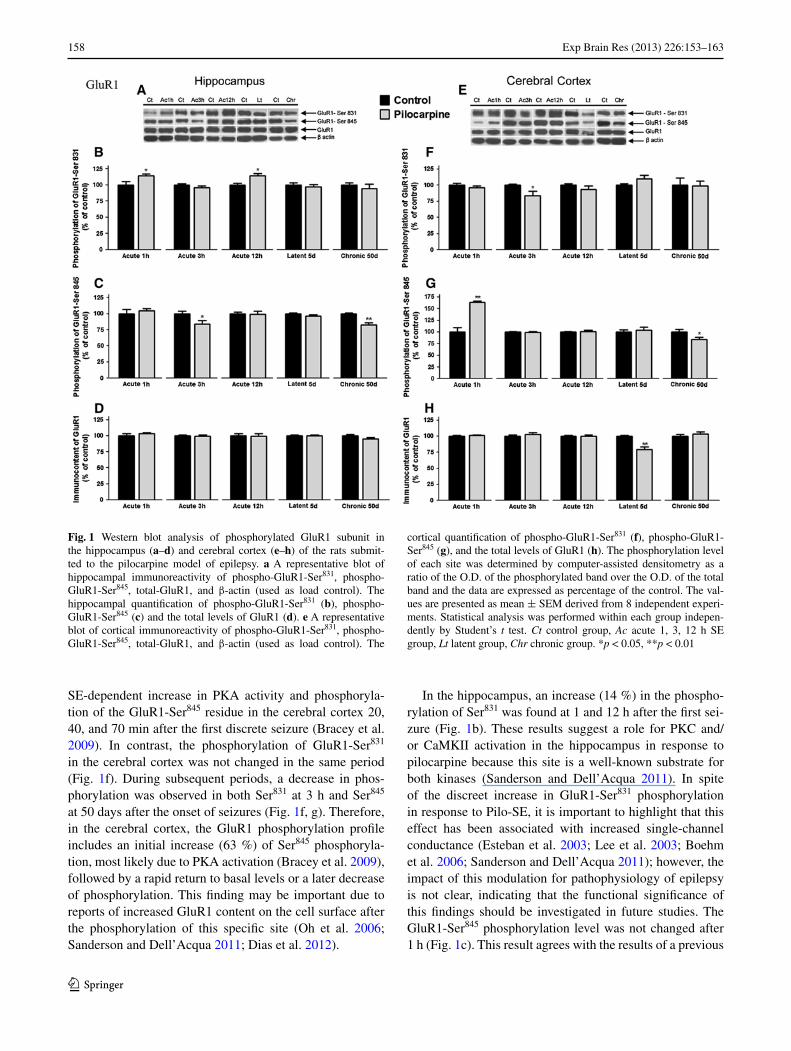
The evaluation of the GluR1 subunit of AMPA receptors in the hippocampus showed a significant increase (p < 0.05) in GluR1-Ser831 phosphorylation at 1 h (14.46 ± 2.82 %) and 12 h (14.00 ± 3.78 %) after SE onset, as compared to the control group (Fig. 1b). The GluR1-Ser831 phosphorylation levels returned to baseline in the latent and chronic periods. In contrast, the GluR1-Ser845 phosphorylation decreased (16.23 ± 5.94 %, p < 0.05) at 3 h after SE onset, returned to the baseline at 12 h and 5 days, and decreased in the chronic period (17.74 ± 3.67 %, p < 0.01) (Fig. 1c). No significant changes were detected in the amount of total form of GluR1 subunit.
In contrast with the hippocampus, the GluR1 subunit phosphorylation profile of AMPA receptors was modulated differently in the cerebral cortex after the Pilo-SE onset. The GluR1-Ser831 phosphorylation decreased (16.21 ± 6.75 %, p < 0.05) at 3 h after Pilo-SE onset, returning to baseline in
the other periods (Fig. 1f). GluR1-Ser845 phosphorylation increased significantly (63.32 ± 2.72 %, p < 0.01) at 1 h after Pilo-SE onset, returning to the controls levels between 3 h and 5 days (Fig. 1g). In the chronic period, there was a decrease (17.74 ± 3.67 %, p < 0.05) of GluR1-Ser845 phosphorylation (Fig. 1g). The total content of the AMPA receptor GluR1 subunit levels remained unchanged in the acute and chronic periods but showed a significant decrease (20.74 ± 4.04 %, p < 0.01) in the latent period (Fig. 1h).
EAAT1, EAAT2 and NR1 expression in the hippocampus and cerebral cortex after Pilo-SE induction
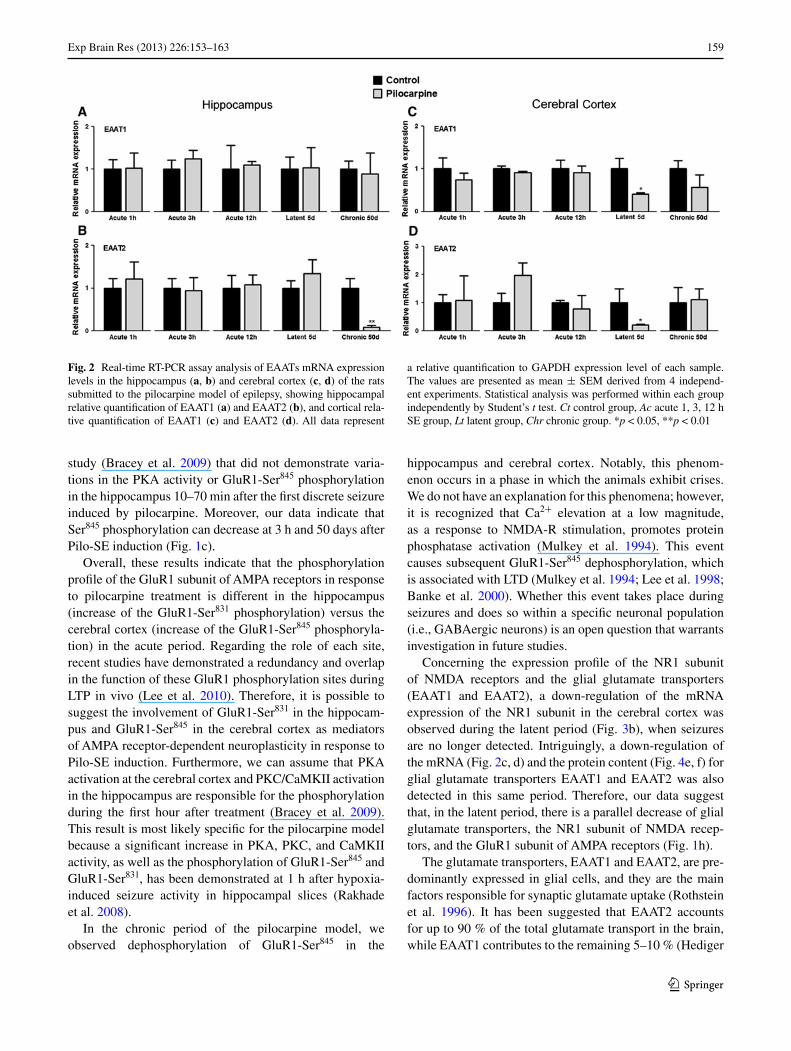
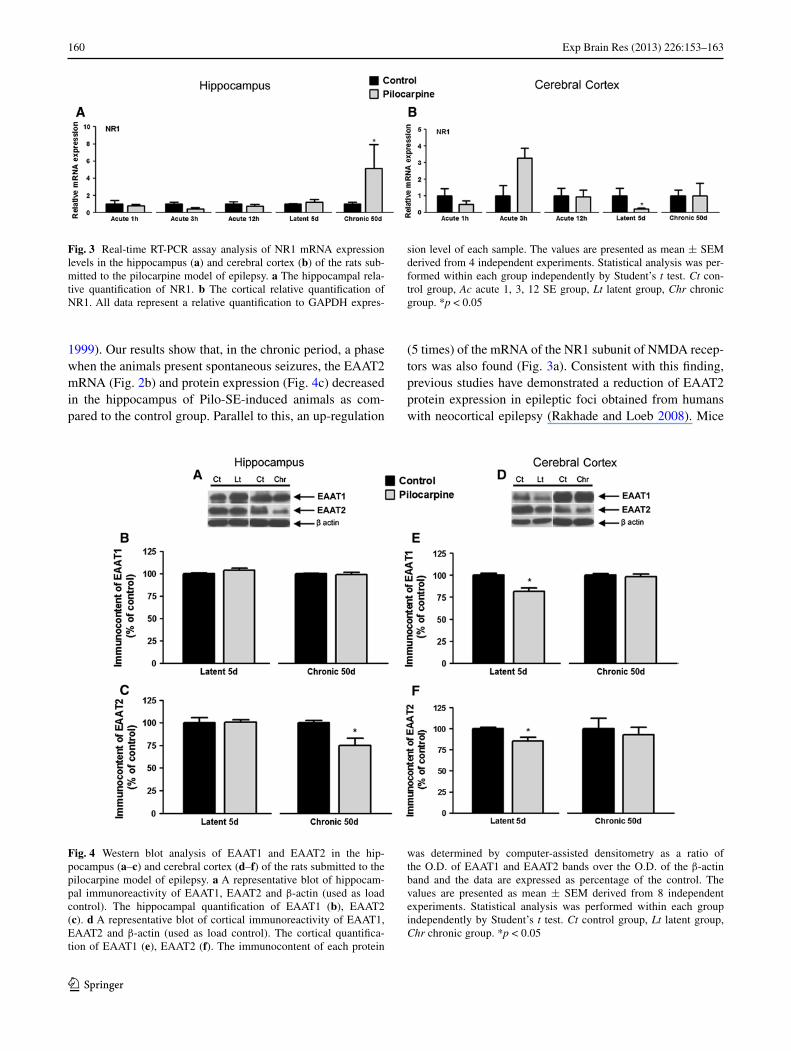
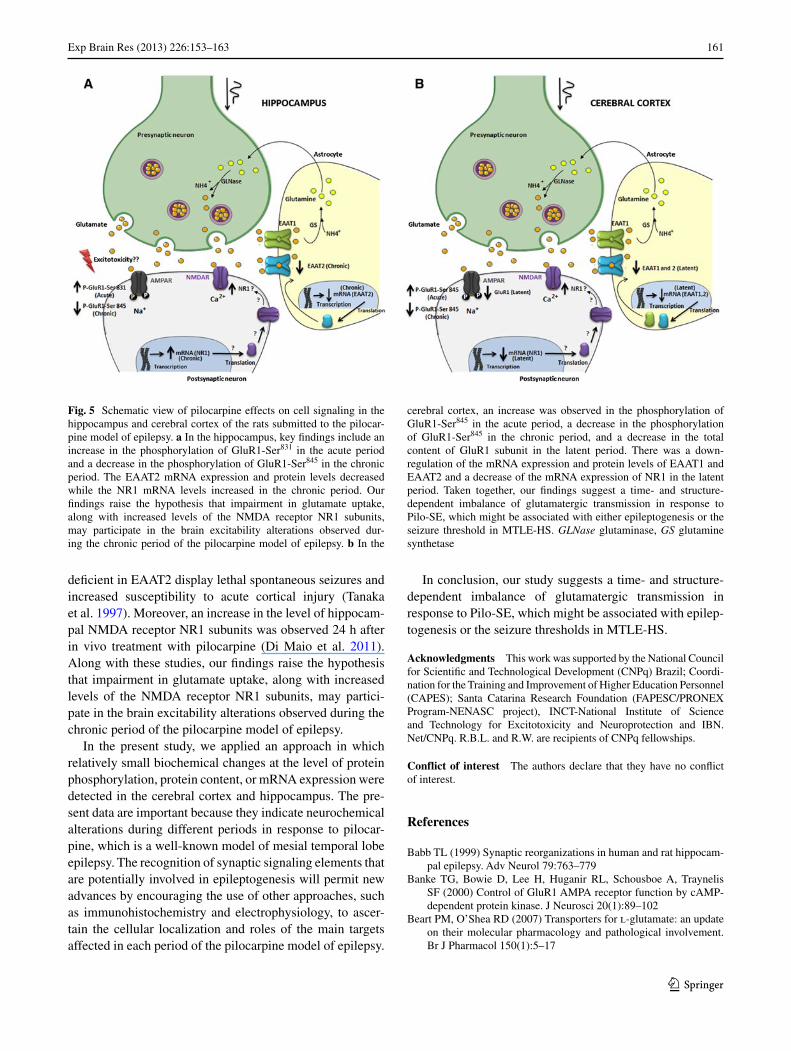
No significant changes were detected in EAAT1 mRNA expression levels (Fig. 2a) in the hippocampus after the Pilo-SE induction. However, compared to the control group, EAAT2 mRNA expression decreased 91 % (Fig. 2b) in the chronic period. This down-regulation was confirmed by pro-tein content analysis by western blotting which showed a decreasing of EATT2 content by 25.07 % ± 7.88 (Fig. 4c). In contrast, the mRNA expression of the NR1 subunit of NMDA receptors increased 5 times in the chronic period, as compared to the respective controls (Fig. 3a).
The evaluation of the expression of the glutamate trans-porters (EAAT1, EAAT2) in the cerebral cortex showed a down-regulation of both EAAT1 (60 %) (Fig. 2c) and EAAT2 (80 %) mRNA (Fig. 2d) levels during the latent period. This down-regulation of both transporters was also confirmed by protein content analysis by western blotting which showed a decreasing of EAAT1 (18.27 % ± 3.71) and EATT2 (14.79 % ± 4.53) in the latent period (Fig. 4e, f, respectively). The mRNA levels of the NR1 subunit of NMDA receptors decreased by 80 % (Fig. 3b) during the same period.
To display the effects of the pilocarpine model of epi-lepsy in an integrated manner, Fig. 5 was created based on the data presented in Figs. 1, 2, 3 and 4. Interestingly, an increase in the expression of the hippocampal NR1 subunit and a decrease in the expression of the glutamate transporter EAAT2 were observed in the chronic period.
Discussion
Our results provide a GluR1-Ser831 and GluR1-Ser845 phos-phorylation profile for AMPA receptors, as well as demon-strating the mRNA expression of NR1, EAAT1, and EAAT2 in the pilocarpine model of epilepsy. These observations were performed in the hippocampus and cerebral cortex at different periods following Pilo-SE induction.
The GluR1-Ser845 site had an increase in phosphoryla-tion in the cerebral cortex 1 h after SE onset (Fig. 1g). This finding agrees with a previous study that demonstrated an
158 Exp Brain Res (2013) 226:153–163
1 3
SE-dependent increase in PKA activity and phosphoryla-tion of the GluR1-Ser845 residue in the cerebral cortex 20, 40, and 70 min after the first discrete seizure (Bracey et al. 2009). In contrast, the phosphorylation of GluR1-Ser831 in the cerebral cortex was not changed in the same period (Fig. 1f). During subsequent periods, a decrease in phos-phorylation was observed in both Ser831 at 3 h and Ser845 at 50 days after the onset of seizures (Fig. 1f, g). Therefore, in the cerebral cortex, the GluR1 phosphorylation profile includes an initial increase (63 %) of Ser845 phosphoryla-tion, most likely due to PKA activation (Bracey et al. 2009), followed by a rapid return to basal levels or a later decrease of phosphorylation. This finding may be important due to reports of increased GluR1 content on the cell surface after the phosphorylation of this specific site (Oh et al. 2006; Sanderson and Dell’Acqua 2011; Dias et al. 2012).
In the hippocampus, an increase (14 %) in the phospho-rylation of Ser831 was found at 1 and 12 h after the first sei-zure (Fig. 1b). These results suggest a role for PKC and/or CaMKII activation in the hippocampus in response to pilocarpine because this site is a well-known substrate for both kinases (Sanderson and Dell’Acqua 2011). In spite of the discreet increase in GluR1-Ser831 phosphorylation in response to Pilo-SE, it is important to highlight that this effect has been associated with increased single-channel conductance (Esteban et al. 2003; Lee et al. 2003; Boehm et al. 2006; Sanderson and Dell’Acqua 2011); however, the impact of this modulation for pathophysiology of epilepsy is not clear, indicating that the functional significance of this findings should be investigated in future studies. The GluR1-Ser845 phosphorylation level was not changed after 1 h (Fig. 1c). This result agrees with the results of a previous
Fig. 1 Western blot analysis of phosphorylated GluR1 subunit in the hippocampus (a–d) and cerebral cortex (e–h) of the rats submit-ted to the pilocarpine model of epilepsy. a A representative blot of hippocampal immunoreactivity of phospho-GluR1-Ser831, phospho-GluR1-Ser845, total-GluR1, and β-actin (used as load control). The hippocampal quantification of phospho-GluR1-Ser831 (b), phospho-GluR1-Ser845 (c) and the total levels of GluR1 (d). e A representative blot of cortical immunoreactivity of phospho-GluR1-Ser831, phospho-GluR1-Ser845, total-GluR1, and β-actin (used as load control). The
cortical quantification of phospho-GluR1-Ser831 (f), phospho-GluR1-Ser845 (g), and the total levels of GluR1 (h). The phosphorylation level of each site was determined by computer-assisted densitometry as a ratio of the O.D. of the phosphorylated band over the O.D. of the total band and the data are expressed as percentage of the control. The val-ues are presented as mean ± SEM derived from 8 independent experi-ments. Statistical analysis was performed within each group indepen-dently by Student’s t test. Ct control group, Ac acute 1, 3, 12 h SE group, Lt latent group, Chr chronic group. *p < 0.05, **p < 0.01
159Exp Brain Res (2013) 226:153–163
1 3
study (Bracey et al. 2009) that did not demonstrate varia-tions in the PKA activity or GluR1-Ser845 phosphorylation in the hippocampus 10–70 min after the first discrete seizure induced by pilocarpine. Moreover, our data indicate that Ser845 phosphorylation can decrease at 3 h and 50 days after Pilo-SE induction (Fig. 1c).
Overall, these results indicate that the phosphorylation profile of the GluR1 subunit of AMPA receptors in response to pilocarpine treatment is different in the hippocampus (increase of the GluR1-Ser831 phosphorylation) versus the cerebral cortex (increase of the GluR1-Ser845 phosphoryla-tion) in the acute period. Regarding the role of each site, recent studies have demonstrated a redundancy and overlap in the function of these GluR1 phosphorylation sites during LTP in vivo (Lee et al. 2010). Therefore, it is possible to suggest the involvement of GluR1-Ser831 in the hippocam-pus and GluR1-Ser845 in the cerebral cortex as mediators of AMPA receptor-dependent neuroplasticity in response to Pilo-SE induction. Furthermore, we can assume that PKA activation at the cerebral cortex and PKC/CaMKII activation in the hippocampus are responsible for the phosphorylation during the first hour after treatment (Bracey et al. 2009). This result is most likely specific for the pilocarpine model because a significant increase in PKA, PKC, and CaMKII activity, as well as the phosphorylation of GluR1-Ser845 and GluR1-Ser831, has been demonstrated at 1 h after hypoxia-induced seizure activity in hippocampal slices (Rakhade et al. 2008).
In the chronic period of the pilocarpine model, we observed dephosphorylation of GluR1-Ser845 in the
hippocampus and cerebral cortex. Notably, this phenom-enon occurs in a phase in which the animals exhibit crises. We do not have an explanation for this phenomena; however, it is recognized that Ca2+ elevation at a low magnitude, as a response to NMDA-R stimulation, promotes protein phosphatase activation (Mulkey et al. 1994). This event causes subsequent GluR1-Ser845 dephosphorylation, which is associated with LTD (Mulkey et al. 1994; Lee et al. 1998; Banke et al. 2000). Whether this event takes place during seizures and does so within a specific neuronal population (i.e., GABAergic neurons) is an open question that warrants investigation in future studies.
Concerning the expression profile of the NR1 subunit of NMDA receptors and the glial glutamate transporters (EAAT1 and EAAT2), a down-regulation of the mRNA expression of the NR1 subunit in the cerebral cortex was observed during the latent period (Fig. 3b), when seizures are no longer detected. Intriguingly, a down-regulation of the mRNA (Fig. 2c, d) and the protein content (Fig. 4e, f) for glial glutamate transporters EAAT1 and EAAT2 was also detected in this same period. Therefore, our data suggest that, in the latent period, there is a parallel decrease of glial glutamate transporters, the NR1 subunit of NMDA recep-tors, and the GluR1 subunit of AMPA receptors (Fig. 1h).
The glutamate transporters, EAAT1 and EAAT2, are pre-dominantly expressed in glial cells, and they are the main factors responsible for synaptic glutamate uptake (Rothstein et al. 1996). It has been suggested that EAAT2 accounts for up to 90 % of the total glutamate transport in the brain, while EAAT1 contributes to the remaining 5–10 % (Hediger
Fig. 2 Real-time RT-PCR assay analysis of EAATs mRNA expression levels in the hippocampus (a, b) and cerebral cortex (c, d) of the rats submitted to the pilocarpine model of epilepsy, showing hippocampal relative quantification of EAAT1 (a) and EAAT2 (b), and cortical rela-tive quantification of EAAT1 (c) and EAAT2 (d). All data represent
a relative quantification to GAPDH expression level of each sample. The values are presented as mean ± SEM derived from 4 independ-ent experiments. Statistical analysis was performed within each group independently by Student’s t test. Ct control group, Ac acute 1, 3, 12 h SE group, Lt latent group, Chr chronic group. *p < 0.05, **p < 0.01
160 Exp Brain Res (2013) 226:153–163
1 3
1999). Our results show that, in the chronic period, a phase when the animals present spontaneous seizures, the EAAT2 mRNA (Fig. 2b) and protein expression (Fig. 4c) decreased in the hippocampus of Pilo-SE-induced animals as com-pared to the control group. Parallel to this, an up-regulation
(5 times) of the mRNA of the NR1 subunit of NMDA recep-tors was also found (Fig. 3a). Consistent with this finding, previous studies have demonstrated a reduction of EAAT2 protein expression in epileptic foci obtained from humans with neocortical epilepsy (Rakhade and Loeb 2008). Mice
Fig. 3 Real-time RT-PCR assay analysis of NR1 mRNA expression levels in the hippocampus (a) and cerebral cortex (b) of the rats sub-mitted to the pilocarpine model of epilepsy. a The hippocampal rela-tive quantification of NR1. b The cortical relative quantification of NR1. All data represent a relative quantification to GAPDH expres-
sion level of each sample. The values are presented as mean ± SEM derived from 4 independent experiments. Statistical analysis was per-formed within each group independently by Student’s t test. Ct con-trol group, Ac acute 1, 3, 12 SE group, Lt latent group, Chr chronic group. *p < 0.05
Fig. 4 Western blot analysis of EAAT1 and EAAT2 in the hip-pocampus (a–c) and cerebral cortex (d–f) of the rats submitted to the pilocarpine model of epilepsy. a A representative blot of hippocam-pal immunoreactivity of EAAT1, EAAT2 and β-actin (used as load control). The hippocampal quantification of EAAT1 (b), EAAT2 (c). d A representative blot of cortical immunoreactivity of EAAT1, EAAT2 and β-actin (used as load control). The cortical quantifica-tion of EAAT1 (e), EAAT2 (f). The immunocontent of each protein
was determined by computer-assisted densitometry as a ratio of the O.D. of EAAT1 and EAAT2 bands over the O.D. of the β-actin band and the data are expressed as percentage of the control. The values are presented as mean ± SEM derived from 8 independent experiments. Statistical analysis was performed within each group independently by Student’s t test. Ct control group, Lt latent group, Chr chronic group. *p < 0.05
161Exp Brain Res (2013) 226:153–163
1 3
deficient in EAAT2 display lethal spontaneous seizures and increased susceptibility to acute cortical injury (Tanaka et al. 1997). Moreover, an increase in the level of hippocam-pal NMDA receptor NR1 subunits was observed 24 h after in vivo treatment with pilocarpine (Di Maio et al. 2011). Along with these studies, our findings raise the hypothesis that impairment in glutamate uptake, along with increased levels of the NMDA receptor NR1 subunits, may partici-pate in the brain excitability alterations observed during the chronic period of the pilocarpine model of epilepsy.
In the present study, we applied an approach in which relatively small biochemical changes at the level of protein phosphorylation, protein content, or mRNA expression were detected in the cerebral cortex and hippocampus. The pre-sent data are important because they indicate neurochemical alterations during different periods in response to pilocar-pine, which is a well-known model of mesial temporal lobe epilepsy. The recognition of synaptic signaling elements that are potentially involved in epileptogenesis will permit new advances by encouraging the use of other approaches, such as immunohistochemistry and electrophysiology, to ascer-tain the cellular localization and roles of the main targets affected in each period of the pilocarpine model of epilepsy.
In conclusion, our study suggests a time- and structure-dependent imbalance of glutamatergic transmission in response to Pilo-SE, which might be associated with epilep-togenesis or the seizure thresholds in MTLE-HS.
Acknowledgments This work was supported by the National Council for Scientific and Technological Development (CNPq) Brazil; Coordi-nation for the Training and Improvement of Higher Education Personnel (CAPES); Santa Catarina Research Foundation (FAPESC/PRONEX Program-NENASC project), INCT-National Institute of Science and Technology for Excitotoxicity and Neuroprotection and IBN.Net/CNPq. R.B.L. and R.W. are recipients of CNPq fellowships.
Conflict of interest The authors declare that they have no conflict of interest.
References
Babb TL (1999) Synaptic reorganizations in human and rat hippocam-pal epilepsy. Adv Neurol 79:763–779
Banke TG, Bowie D, Lee H, Huganir RL, Schousboe A, Traynelis SF (2000) Control of GluR1 AMPA receptor function by cAMP-dependent protein kinase. J Neurosci 20(1):89–102
Beart PM, O’Shea RD (2007) Transporters for l-glutamate: an update on their molecular pharmacology and pathological involvement. Br J Pharmacol 150(1):5–17
Fig. 5 Schematic view of pilocarpine effects on cell signaling in the hippocampus and cerebral cortex of the rats submitted to the pilocar-pine model of epilepsy. a In the hippocampus, key findings include an increase in the phosphorylation of GluR1-Ser831 in the acute period and a decrease in the phosphorylation of GluR1-Ser845 in the chronic period. The EAAT2 mRNA expression and protein levels decreased while the NR1 mRNA levels increased in the chronic period. Our findings raise the hypothesis that impairment in glutamate uptake, along with increased levels of the NMDA receptor NR1 subunits, may participate in the brain excitability alterations observed dur-ing the chronic period of the pilocarpine model of epilepsy. b In the
cerebral cortex, an increase was observed in the phosphorylation of GluR1-Ser845 in the acute period, a decrease in the phosphorylation of GluR1-Ser845 in the chronic period, and a decrease in the total content of GluR1 subunit in the latent period. There was a down-regulation of the mRNA expression and protein levels of EAAT1 and EAAT2 and a decrease of the mRNA expression of NR1 in the latent period. Taken together, our findings suggest a time- and structure-dependent imbalance of glutamatergic transmission in response to Pilo-SE, which might be associated with either epileptogenesis or the seizure threshold in MTLE-HS. GLNase glutaminase, GS glutamine synthetase

162 Exp Brain Res (2013) 226:153–163
1 3
Bernabeu R, Izquierdo I, Cammarota M, Jerusalinsky D, Medina JH (1995) Learning-specific, time-dependent increase in [3H]phorbol dibutyrate binding to protein kinase C in selected regions of the rat brain. Brain Res 685(1–2):163–168
Bernabeu R, Bevilaqua L, Ardenghi P, Bromberg E, Schmitz P, Bianchin M, Izquierdo I, Medina JH (1997) Involvement of hippocampal cAMP/cAMP-dependent protein kinase signaling pathways in a late memory consolidation phase of aversively moti-vated learning in rats. Proc Natl Acad Sci USA 94(13):7041–7046
Boehm J, Kang MG, Johnson RC, Esteban J, Huganir RL, Malinow R (2006) Synaptic incorporation of AMPA receptors during LTP is controlled by a PKC phosphorylation site on GluR1. Neuron 51(2):213–225
Bonan CD, Walz R, Pereira GS, Worm PV, Battastini AM, Cavalheiro EA, Izquierdo I, Sarkis JJ (2000) Changes in synaptosomal ecto-nucleotidase activities in two rat models of temporal lobe epilepsy. Epilepsy Res 39(3):229–238
Bracey JM, Kurz JE, Low B, Churn SB (2009) Prolonged seizure activity leads to increased Protein Kinase A activation in the rat pilocarpine model of status epilepticus. Brain Res 1283:167–176
Calloni GW, Penno CA, Cordova FM, Trentin AG, Neto VM, Leal RB (2005) Congenital hypothyroidism alters the phosphorylation of ERK1/2 and p38MAPK in the hippocampus of neonatal rats. Brain Res Dev Brain Res 154(1):141–145
Cammarota M, Bernabeu R, Levi De Stein M, Izquierdo I, Medina JH (1998) Learning-specific, time-dependent increases in hip-pocampal Ca2+/calmodulin-dependent protein kinase II activ-ity and AMPA GluR1 subunit immunoreactivity. Eur J Neurosci 10(8):2669–2676
Cavalheiro EA, Leite JP, Bortolotto ZA, Turski WA, Ikonomidou C, Turski L (1991) Long-term effects of pilocarpine in rats: structural damage of the brain triggers kindling and spontaneous recurrent seizures. Epilepsia 32(6):778–782
Cavalheiro EA, Fernandes MJ, Turski L, Naffah-Mazzacoratti MG (1994) Spontaneous recurrent seizures in rats: amino acid and monoamine determination in the hippocampus. Epilepsia 35(1):1–11
Chomczynski P, Sacchi N (1987) Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal Biochem 162(1):156–159
Cordova FM, Rodrigues AL, Giacomelli MB, Oliveira CS, Posser T, Dunkley PR, Leal RB (2004) Lead stimulates ERK1/2 and p38MAPK phosphorylation in the hippocampus of immature rats. Brain Res 998(1):65–72
Costa MS, Rocha JB, Perosa SR, Cavalheiro EA, Naffah-Mazzacoratti MdaG (2004) Pilocarpine-induced status epilepticus increases glutamate release in rat hippocampal synaptosomes. Neurosci Lett 356(1):41–44
Di Maio R, Mastroberardino PG, Hu X, Montero L, Greenamyre JT (2011) Pilocapine alters NMDA receptor expression and function in hippocampal neurons: NADPH oxidase and ERK1/2 mecha-nisms. Neurobiol Dis 42(3):482–495
Dias RB, Ribeiro JA, Sebastiao AM (2012) Enhancement of AMPA currents and GluR1 membrane expression through PKA-coupled adenosine A(2A) receptors. Hippocampus 22(2):276–291
Esteban JA, Shi SH, Wilson C, Nuriya M, Huganir RL, Malinow R (2003) PKA phosphorylation of AMPA receptor subunits con-trols synaptic trafficking underlying plasticity. Nat Neurosci 6(2):136–143
Figueiredo CP, Antunes VL, Moreira EL, de Mello N, Medeiros R, Di Giunta G, Lobao-Soares B, Linhares M, Lin K, Mazzuco TL, Prediger RD, Walz R (2011) Glucose-dependent insulinotropic peptide receptor expression in the hippocampus and neocortex of mesial temporal lobe epilepsy patients and rats undergoing pilo-carpine induced status epilepticus. Peptides 32(4):781–789
Hediger MA (1999) Glutamate transporters in kidney and brain. Am J Physiol 277(4 Pt 2):F487–F492
Hollmann M, Hartley M, Heinemann S (1991) Ca2+ permeability of KA-AMPA—gated glutamate receptor channels depends on subu-nit composition. Science 252(5007):851–853
Houser CR, Huang CS, Peng Z (2008) Dynamic seizure-related changes in extracellular signal-regulated kinase activation in a mouse model of temporal lobe epilepsy. Neuroscience 156(1):222–237
Jerusalinsky D, Ferreira MB, Walz R, Da Silva RC, Bianchin M, Ruschel AC, Zanatta MS, Medina JH, Izquierdo I (1992) Amnesia by post-training infusion of glutamate receptor antagonists into the amygdala, hippocampus, and entorhinal cortex. Behav Neural Biol 58(1):76–80
Leal RB, Cordova FM, Herd L, Bobrovskaya L, Dunkley PR (2002) Lead-stimulated p38MAPK-dependent Hsp27 phosphorylation. Toxicol Appl Pharmacol 178(1):44–51
Lee A, Pow DV (2010) Astrocytes: glutamate transport and alternate splicing of transporters. Int J Biochem Cell Biol 42(12):1901–1906
Lee HK, Kameyama K, Huganir RL, Bear MF (1998) NMDA induces long-term synaptic depression and dephosphorylation of the GluR1 subunit of AMPA receptors in hippocampus. Neuron 21(5):1151–1162
Lee HK, Takamiya K, Han JS, Man H, Kim CH, Rumbaugh G, Yu S, Ding L, He C, Petralia RS, Wenthold RJ, Gallagher M, Huganir RL (2003) Phosphorylation of the AMPA receptor GluR1 subunit is required for synaptic plasticity and retention of spatial memory. Cell 112(5):631–643
Lee HK, Takamiya K, He K, Song L, Huganir RL (2010) Specific roles of AMPA receptor subunit GluR1 (GluA1) phosphorylation sites in regulating synaptic plasticity in the CA1 region of hippocam-pus. J Neurophysiol 103(1):479–489
Leite JP, Neder L, Arisi GM, Carlotti CG Jr, Assirati JA, Moreira JE (2005) Plasticity, synaptic strength, and epilepsy: what can we learn from ultrastructural data? Epilepsia 46(Suppl 5):134–141
Li Y, Peng Z, Xiao B, Houser CR (2010) Activation of ERK by spon-taneous seizures in neural progenitors of the dentate gyrus in a mouse model of epilepsy. Exp Neurol 224(1):133–145
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expres-sion data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 25(4):402–408
Lopes MW, Soares FM, de Mello N, Nunes JC, de Cordova FM, Walz R, Leal RB (2012) Time-dependent modulation of mito-gen activated protein kinases and AKT in rat hippocampus and cortex in the pilocarpine model of epilepsy. Neurochem Res 37(9):1868–1878
Mello LE, Cavalheiro EA, Tan AM, Kupfer WR, Pretorius JK, Babb TL, Finch DM (1993) Circuit mechanisms of seizures in the pilocarpine model of chronic epilepsy: cell loss and mossy fiber sprouting. Epilepsia 34(6):985–995
Mulkey RM, Endo S, Shenolikar S, Malenka RC (1994) Involvement of a calcineurin/inhibitor-1 phosphatase cascade in hippocampal long-term depression. Nature 369(6480):486–488
Oh MC, Derkach VA, Guire ES, Soderling TR (2006) Extrasynaptic membrane trafficking regulated by GluR1 serine 845 phospho-rylation primes AMPA receptors for long-term potentiation. J Biol Chem 281(2):752–758
Oliveira CS, Rigon AP, Leal RB, Rossi FM (2008) The activation of ERK1/2 and p38 mitogen-activated protein kinases is dynamically regulated in the developing rat visual system. Int J Dev Neurosci 26(3–4):355–362
Peterson GL (1977) A simplification of the protein assay method of Lowry et al. which is more generally applicable. Anal Biochem 83(2):346–356
Posser T, de Aguiar CB, Garcez RC, Rossi FM, Oliveira CS, Trentin AG, Neto VM, Leal RB (2007) Exposure of C6 glioma cells to

163Exp Brain Res (2013) 226:153–163
1 3
Pb(II) increases the phosphorylation of p38(MAPK) and JNK1/2 but not of ERK1/2. Arch Toxicol 81(6):407–414
Quillfeldt JA, Schmitz PK, Walz R, Bianchin M, Zanatta MS, Medina JH, Izquierdo I (1994) CNQX infused into entorhinal cortex blocks memory expression, and AMPA reverses the effect. Phar-macol Biochem Behav 48(2):437–440
Rakhade SN, Loeb JA (2008) Focal reduction of neuronal gluta-mate transporters in human neocortical epilepsy. Epilepsia 49(2):226–236
Rakhade SN, Zhou C, Aujla PK, Fishman R, Sucher NJ, Jensen FE (2008) Early alterations of AMPA receptors mediate syn-aptic potentiation induced by neonatal seizures. J Neurosci 28(32):7979–7990
Rothstein JD, Dykes-Hoberg M, Pardo CA, Bristol LA, Jin L, Kuncl RW, Kanai Y, Hediger MA, Wang Y, Schielke JP, Welty DF (1996) Knockout of glutamate transporters reveals a major role for astro-glial transport in excitotoxicity and clearance of glutamate. Neu-ron 16(3):675–686
Sander JW (2003) The epidemiology of epilepsy revisited. Curr Opin Neurol 16(2):165–170
Sanderson JL, Dell’Acqua ML (2011) AKAP signaling complexes in regulation of excitatory synaptic plasticity. Neuroscientist 17(3):321–336
Sweatt JD (2004) Mitogen-activated protein kinases in synaptic plas-ticity and memory. Curr Opin Neurobiol 14(3):311–317
Tanaka K, Watase K, Manabe T, Yamada K, Watanabe M, Takahashi K, Iwama H, Nishikawa T, Ichihara N, Kikuchi T, Okuyama S, Kawashima N, Hori S, Takimoto M, Wada K (1997) Epilepsy and exacerbation of brain injury in mice lacking the glutamate trans-porter GLT-1. Science 276(5319):1699–1702
Thomas GM, Huganir RL (2004) MAPK cascade signalling and syn-aptic plasticity. Nat Rev Neurosci 5(3):173–183
Walz R, Roesler R, Quevedo J, Sant’Anna MK, Madruga M, Rodrigues C, Gottfried C, Medina JH, Izquierdo I (2000) Time-dependent impairment of inhibitory avoidance retention in rats by posttrain-ing infusion of a mitogen-activated protein kinase kinase inhibitor into cortical and limbic structures. Neurobiol Learn Mem 73(1): 11–20
Copyright © 2022 FDOKUMEN




















