
The vagus nerve mediates behavioural depression, but not fever, in response to peripheral immune...
13
The vagus nerve mediates behavioural depression, but not fever, in response to peripheral immune signals; a functional anatomical analysis Jan Pieter Konsman, 1 Giamal N. Luheshi, 2 Rose-Marie Bluthe ´ 1 and Robert Dantzer 1 1 INSERM U394, Neurobiologie Inte ´ grative, Institut Franc ¸ ois Magendie, Rue Camille Saint-Sae ¨ ns, 33077 Bordeaux Cedex, France 2 Douglas Hospital Research Center, 6875, Boulevard LaSalle, Verdun, Quebec, Canada H4H 1R3 Keywords: Fos, interleukin-1, neuro–immune interactions, rat, Stat3 Abstract Cytokines act on the brain to induce fever and behavioural depression after infection. Although several mechanisms of cytokine- to-brain communication have been proposed, their physiological significance is unclear. We propose that behavioural depression is mediated by the vagus nerve activating limbic structures, while fever would primarily be due to humoral mechanisms affecting the preoptic area, including interleukin-6 (IL-6) action on the organum vasculosum of the laminae terminalis (OVLT) and induction of prostaglandins. This study assessed the effects of subdiaphragmatic vagotomy in rats on fever, behavioural depression, as measured by the social interaction test, and Fos expression in the brain. These responses were compared with induction of the prostaglandin-producing enzyme cyclooxygenase-2 and the transcription factor Stat3 that translocates after binding of IL-6. Vagotomy blocked behavioural depression after intraperitoneal injection of recombinant rat IL-1b (25 mg/kg) or lipopolysaccharide (250 mg/kg; LPS) and prevented Fos expression in limbic structures and ventromedial preoptic area, but not in the OVLT. Fever was not affected by vagotomy, but associated with translocation of Stat3 in the OVLT and cyclooxygenase-2 induction around blood vessels. These results indicate that the recently proposed vagal link between the immune system and the brain activates limbic structures to induce behavioural depression after abdominal inflammation. Although the vagus might play a role in fever in response to low doses of LPS by activating the ventromedial preoptic area, it is likely to be overridden during more severe infection by action of circulating IL-6 on the OVLT or prostaglandins induced along blood vessels of the preoptic area. Introduction Detection of bacterial lipopolysaccharide (LPS) by immune cells leads to interleukin-1b (IL-1b) and interleukin-6 (IL-6) release. These cytokines act on the brain to provoke fever and behavioural depression (Rothwell & Hopkins, 1995). Fever is an adaptive response to infection (Kluger, 1991) and behavioural depression contributes to it by reducing energy expenditure (Hart, 1988). Because of their size and hydrophilic nature, cytokines cannot diffuse across the blood–brain barrier (BBB). Three possible mechanisms of cytokine-to-brain signalling have been proposed. Classically, cytokines are thought to act on circumventricular organs where the BBB is absent (Blatteis, 1992). IL-6, for example, rapidly rises in plasma after LPS administration (Luheshi et al., 1996) and induces the cellular activation marker c-fos exclusively in circum- ventricular organs (Vallie `res et al., 1997). Alternatively, LPS and IL- 1b induce cyclooxygenase-2 (COX-2), the limiting enzyme for prostaglandin synthesis, in endothelial cells (Cao et al., 1996; Laflamme et al., 1999). Lipophilic prostaglandins freely diffuse across the BBB and might thus act on receptors found on neuronal populations (Zhang & Rivest, 1999). Recently, the vagus nerve was proposed as an immune-to-brain pathway (Dantzer, 1994). Although evidence exists for each of these mechanisms, their possible intervention has usually been tested separately and by addressing only one physiological response. Transection of the vagus nerves inhibits fever only in response to low doses of LPS or IL-1b (Hansen & Krueger, 1997; Romanovsky et al., 1997). Clearly, higher amounts of circulating cytokines are more prone to act on circumventricular organs or induce COX-2 around blood vessels. However, vagotomy consistently blocks LPS- and IL-1b-induced behavioural depression, as measured by the social interaction test, even in response to high doses (Bluthe ´ et al., 1994, 1996b). Since higher doses of IL-1b are needed to induce behavioural depression than fever (Anforth et al., 1998), it is possible that fever and behavioural depression are mediated by distinct pathways of immune-to-brain communication, resulting in activation of different brain structures. In the case of fever, the preoptic hypothalamus is the target structure (Boulant, 1981). The neural structures implicated in behavioural changes are still unknown, but limbic structures are good candidates. This study was designed to test the hypothesis that fever and behavioural depression are due to different mechanisms of immune- to-brain signalling. In addition to fever and behavioural depression, we evaluated LPS-induced Fos expression in rats that underwent subdiaphragmatic vagotomy and compared it with COX-2 induction. Brain targets of IL-6 were investigated by studying expression of Stat3 that translocates into the nucleus after activation by the signal Correspondence: Dr Jan Pieter Konsman, Neuroscience, Floor 11, Division of Cell Biology, Department of Biomedicine and Surgery, Faculty of Health Sciences, 58185 Linko ¨ping, Sweden. E-mail: [email protected] Received 4 February 2000, revised 7 September 2000, accepted 25 September 2000 European Journal of Neuroscience, Vol. 12, pp. 4434–4446, 2000 ª Federation of European Neuroscience Societies
Transcript of The vagus nerve mediates behavioural depression, but not fever, in response to peripheral immune...

The vagus nerve mediates behavioural depression, butnot fever, in response to peripheral immune signals; afunctional anatomical analysis
Jan Pieter Konsman,1 Giamal N. Luheshi,2 Rose-Marie BlutheÂ1 and Robert Dantzer1
1INSERM U394, Neurobiologie InteÂgrative, Institut FrancËois Magendie, Rue Camille Saint-SaeÈns, 33077 Bordeaux Cedex, France2Douglas Hospital Research Center, 6875, Boulevard LaSalle, Verdun, Quebec, Canada H4H 1R3
Keywords: Fos, interleukin-1, neuro±immune interactions, rat, Stat3
Abstract
Cytokines act on the brain to induce fever and behavioural depression after infection. Although several mechanisms of cytokine-
to-brain communication have been proposed, their physiological signi®cance is unclear. We propose that behavioural depression
is mediated by the vagus nerve activating limbic structures, while fever would primarily be due to humoral mechanisms affectingthe preoptic area, including interleukin-6 (IL-6) action on the organum vasculosum of the laminae terminalis (OVLT) and induction
of prostaglandins. This study assessed the effects of subdiaphragmatic vagotomy in rats on fever, behavioural depression, as
measured by the social interaction test, and Fos expression in the brain. These responses were compared with induction of the
prostaglandin-producing enzyme cyclooxygenase-2 and the transcription factor Stat3 that translocates after binding of IL-6.Vagotomy blocked behavioural depression after intraperitoneal injection of recombinant rat IL-1b (25 mg/kg) or lipopolysaccharide
(250 mg/kg; LPS) and prevented Fos expression in limbic structures and ventromedial preoptic area, but not in the OVLT. Fever
was not affected by vagotomy, but associated with translocation of Stat3 in the OVLT and cyclooxygenase-2 induction aroundblood vessels. These results indicate that the recently proposed vagal link between the immune system and the brain activates
limbic structures to induce behavioural depression after abdominal in¯ammation. Although the vagus might play a role in fever in
response to low doses of LPS by activating the ventromedial preoptic area, it is likely to be overridden during more severeinfection by action of circulating IL-6 on the OVLT or prostaglandins induced along blood vessels of the preoptic area.
Introduction
Detection of bacterial lipopolysaccharide (LPS) by immune cells
leads to interleukin-1b (IL-1b) and interleukin-6 (IL-6) release. These
cytokines act on the brain to provoke fever and behavioural
depression (Rothwell & Hopkins, 1995). Fever is an adaptive
response to infection (Kluger, 1991) and behavioural depression
contributes to it by reducing energy expenditure (Hart, 1988).
Because of their size and hydrophilic nature, cytokines cannot
diffuse across the blood±brain barrier (BBB). Three possible
mechanisms of cytokine-to-brain signalling have been proposed.
Classically, cytokines are thought to act on circumventricular organs
where the BBB is absent (Blatteis, 1992). IL-6, for example, rapidly
rises in plasma after LPS administration (Luheshi et al., 1996) and
induces the cellular activation marker c-fos exclusively in circum-
ventricular organs (VallieÁres et al., 1997). Alternatively, LPS and IL-
1b induce cyclooxygenase-2 (COX-2), the limiting enzyme for
prostaglandin synthesis, in endothelial cells (Cao et al., 1996;
La¯amme et al., 1999). Lipophilic prostaglandins freely diffuse
across the BBB and might thus act on receptors found on neuronal
populations (Zhang & Rivest, 1999). Recently, the vagus nerve was
proposed as an immune-to-brain pathway (Dantzer, 1994). Although
evidence exists for each of these mechanisms, their possible
intervention has usually been tested separately and by addressing
only one physiological response.
Transection of the vagus nerves inhibits fever only in response to
low doses of LPS or IL-1b (Hansen & Krueger, 1997; Romanovsky
et al., 1997). Clearly, higher amounts of circulating cytokines are
more prone to act on circumventricular organs or induce COX-2
around blood vessels. However, vagotomy consistently blocks LPS-
and IL-1b-induced behavioural depression, as measured by the social
interaction test, even in response to high doses (Bluthe et al., 1994,
1996b). Since higher doses of IL-1b are needed to induce behavioural
depression than fever (Anforth et al., 1998), it is possible that fever
and behavioural depression are mediated by distinct pathways of
immune-to-brain communication, resulting in activation of different
brain structures. In the case of fever, the preoptic hypothalamus is the
target structure (Boulant, 1981). The neural structures implicated in
behavioural changes are still unknown, but limbic structures are good
candidates.
This study was designed to test the hypothesis that fever and
behavioural depression are due to different mechanisms of immune-
to-brain signalling. In addition to fever and behavioural depression,
we evaluated LPS-induced Fos expression in rats that underwent
subdiaphragmatic vagotomy and compared it with COX-2 induction.
Brain targets of IL-6 were investigated by studying expression of
Stat3 that translocates into the nucleus after activation by the signal
Correspondence: Dr Jan Pieter Konsman, Neuroscience, Floor 11, Division ofCell Biology, Department of Biomedicine and Surgery, Faculty of HealthSciences, 58185 LinkoÈping, Sweden.E-mail: [email protected]
Received 4 February 2000, revised 7 September 2000, accepted 25 September2000
European Journal of Neuroscience, Vol. 12, pp. 4434±4446, 2000 ã Federation of European Neuroscience Societies

transducer gp130 common to the IL-6 receptor family (Hirano et al.,
1994; Heinrich et al., 1998).
The mechanisms behind the effects of vagotomy are still unclear.
Vagotomy may interrupt transmission of a peripheral IL-1b signal to
the nucleus of the solitary tract (NTS) where vagal ®bres terminate.
Alternatively, attenuated IL-1b induction in the brain of vagotomized
animals has been proposed to underlie its attenuating effects (LayeÂ
et al., 1995). Therefore, both LPS-induced Fos expression in the NTS
and IL-1b immunoreactivity in the brain were assessed.
Materials and methods
Animals
All experiments were performed on adult male Wistar rats (Charles
River, Saint Aubin les Elbeuf, France) of 225±250 g body weight.
The animals were housed in a temperature-controlled room
(21 6 2 °C) arti®cially lit between 08.00 h and 20.00 h, and were
provided with food (Extralabo, Provins, France) and water ad libitum.
Juvenile male rats (21±35 days of age) of the same strain served as
stimulus animals for the behavioural studies. All animal experiments
were conducted in accordance with French legislation.
Surgery
One hundred and sixteen animals were food deprived for 24 h prior to
surgery and randomly assigned to vagotomy or sham operations, as
previously described (Bluthe et al., 1996a). Brie¯y, animals were
anaesthetized with a mixture of ketamine and xylazine (61 and 9 mg/
kg, respectively; RhoÃne MeÂrieux, France and Bayer Pharma, Sens,
France), and the main ventral and dorsal trunks of the vagus nerve
were transected immediately above the stomach. All connective
tissue 2±3 cm between the gastric artery and the oesophagus was
removed to ensure the transection of accessory vagal branches. Sham
animals underwent the same surgical procedure excluding nerve
transection. All rats received a single intramuscular injection of
amoxilin (75 mg/kg) and were allowed to recover for 3 weeks.
During the ®rst week of recovery, animals were offered highly
palatable food, in addition to normal chow. Palatable food has
previously been shown to reduce body weight loss (Kraly et al., 1986)
and differences in basal body temperature in vagotomized animals
(Romanovsky et al., 1997). Thirty-®ve animals which continued to
lose weight or which showed apparent signs of sickness, such as
piloerection, were euthanized.
In the 34 animals assigned to fever and behavioural studies,
vagotomy was veri®ed by blockade of the anorexic effect of
intraperitoneal (i.p.) injection of cholecystokinin (4 mg/kg; Sigma,
St Fallavier, France) (Smith et al., 1981) 2 weeks after the end of the
studies. The rats were deprived of food for 18 h, then injected with
cholecystokinin or saline and presented with food 5 min later. Food
intake was measured over the following 30 min. In the 38 animals
assigned to immunocytochemical studies, vagotomy was veri®ed by
incubating NTS sections with isolectin I-B4 (1 mg/mL phosphate-
buffered saline; L5391, Sigma, St Fallavier, France) which was
previously shown to bind to vagal visceral afferents in the NTS (Li
et al., 1997).
Experimental protocol
Injections were administered i.p. (1.0 mL/kg) to hand-held, lightly
restrained rats. Rats were injected with either 25 mg/kg recombinant
rat IL-1b (rrIL-1b; speci®c activity 317 IU/mg; Dr S. Poole, NIBSC,
Potters Bar, UK) dissolved in pyrogen-free saline containing 0.1%
endotoxin-free bovine serum albumin (BSA; A-8806, Sigma) or
250 mg/kg of LPS (Escherichia coli O127:B8, Sigma) dissolved in
pyrogen-free saline. Control injections consisted of pyrogen-free
saline alone. These doses of rrIL-1b and LPS were previously shown
to reliably induce behavioural depression and robust fever responses
from which animals recover within 24 h (Luheshi, unpublished
observations) (Bluthe et al., 1992; Anforth et al., 1998; Konsman
et al., 1999).
Fever
Fever studies were performed at an ambient temperature of
21 6 2 °C, that is below the thermal neutrality for rats (28 °C).
Hypothermia can occur at ambient temperatures below thermal
neutrality when high doses (1000 mg/kg) of LPS are administrated
intravenously (Romanovsky et al., 1997), raising the possibility that
fever phases are masked. However, hypothermia does not occur at
lower doses of LPS (10 mg/kg intravenously) (Romanovsky et al.,
1998). Since i.p. administration of LPS results in 10±100-fold lower
circulating levels compared with intravenous administration (Yasui
et al., 1995), fever phases are very unlikely to be masked after i.p.
injection of 250 mg/kg LPS when monitoring body temperature at
21 °C. Core body temperature was measured continuously in
conscious, undisturbed, individually housed animals by remote
radio-telemetry, via battery-operated biotelemetry transmitters (Data
Sciences, St Paul, Minnesota, USA), previously implanted in the
abdominal cavity during vagotomy or sham surgery. The output
frequency (Hz) of each transmitter was monitored by an antenna
mounted in a receiver board situated beneath the cage of each animal,
and channelled to a peripheral processor (Dataquest IV, Data
Sciences). Frequencies were sampled at 10-min intervals and
converted to degrees Celsius (°C). Since the pyrogenic effects of
IL-1b and LPS are most pronounced during the light phase (Opp &
Toth, 1998), animals received a single injection of rrIL-1b, LPS or
vehicle at 10.00 h and their temperatures were measured every
10 min for 7 h. The mean of three consecutive measurements for
each individual animal was used for further analysis.
Behavioural depression
Following a 2-week recovery period from the fever study, social
interaction was tested in the same group of animals. In this
experiment, rats were injected with LPS, rrIL-1b or vehicle. All
animals received an injection of vehicle and the treatment groups
were arranged such that rats that had received LPS in the fever study
were injected with rrIL-1b, whereas those rats that had received rrIL-
1b previously were injected with LPS. This experimental design was
adopted to avoid the development of tolerance to LPS. Behavioural
depression was induced by injection of rrIL-1b, or LPS assessed by
reduction in the duration of social interaction with a conspeci®c
juvenile introduced into the home cage of the test animal for a 4-min
observation session, immediately before and 2 h after treatment. This
time-point was chosen because it corresponds to the appearance of
effects of IL-1b and LPS on behaviour (Bluthe et al., 1992). Social
interaction was measured by the duration of ano-genital snif®ng and
active interaction of the adult with the juvenile. Different juveniles
were presented on each occasion to avoid habituation. Durations of
social interaction measured in animals treated with saline were used
as baseline values. Social interaction studies were carried out during
the dark phase of the cycle, between 20:00 h and 01:00 h, since rats
display increased social behaviour during the dark phase.
Tissue processing
Rats assigned to immunocytochemical experiments were killed 2 h
after injection of LPS or saline with an overdose of sodium
Distinct mechanisms mediate fever and sickness behaviour 4435
ã 2000 Federation of European Neuroscience Societies, European Journal of Neuroscience, 12, 4434±4446

pentobarbital administered i.p. Once the hind paw re¯ex upon plantar
pinching had disappeared, brains were ®xed by intracardiac perfusion
of saline via the ascending aorta followed by 4% paraformaldehyde in
0.1 M borate buffer (pH 9.5 at 10 °C). Brains were post-®xed for 4 h,
and then cryoprotected in 30% sucrose in 0.1 M phosphate buffer
(pH 7.4) for 48 h. Series of 12 frontal 30 mm cryostat sections
through the whole brain were collected in cold cryoproectant (0.05 M
phosphate buffer, 30% sucrose, 30% ethylene glycol) and stored at
±20 °C until immunocytochemical processing.
Immunocytochemistry
Immunocytochemistry was performed as previously described
(Konsman et al., 1999). Commercially available antisera to Fos,
COX-2 and Stat3 (Santa Cruz Biotechnology, Santa Cruz, CA, USA)
diluted, respectively, 1 : 2000, 1 : 500 and 1 : 2000 were used. The
Fos antiserum (cat. number sc-52, lot. J297) was raised against a
synthetic peptide corresponding to amino acids 3±16 at the N-
terminal of human and mouse Fos and did not cross-react with Fos-
related antigens according to the manufacturer. The COX-2 antiserum
(cat. number sc-1747, lot. M19) was raised against a synthetic peptide
corresponding to amino acids 586±604 at the carboxy terminus of the
COX-2 precursor of rat origin. For preabsorption the corresponding
peptide was used following the manufacturer's instructions (sc-
1747P, lot. L288). Stat3 antiserum (cat. number sc-482X, lot. K247)
was generated against a peptide corresponding to amino acids 750±
769 of the carboxy terminus of the mouse and did not cross-react with
Stat1, Stat2, Stat4, Stat5 or Stat6, according to the manufacturer. The
preabsorption test was performed by incubating the antibody with the
synthetic peptide (cat. number sc-485P, lot. B028), as speci®ed by the
manufacturer. A sheep antiserum generated to rrIL-1b (NIBSC) was
used at a ®nal dilution of 1 : 1000.
After washing off the cryoprotectant solution, immunocytochem-
ical processing was performed on a one in four series of free ¯oating
sections using the streptavidin-biotin-immunoperoxidase technique.
Brie¯y, nonspeci®c binding sites were blocked by a 1-h incubation of
sections in Tris-buffered saline (pH 7.4; TBS) containing 0.3% Triton
X-100 and 0.2% casein. The ®rst antibody was added for 60 h at 4 °C
in the same buffer. After four rinses in TBS, sections were treated for
30 min in 0.3% (v/v) hydrogen peroxide, followed by rinsing in TBS.
Sections were incubated for 2 h at room temperature with
biotinylated donkey antisheep/goat or biotinylated donkey antirabbit
immunoglobulins G (Amersham, Les Ulis, France, 1 : 1000) depend-
ing on the ®rst antibody, and stained using the ABC protocol
(Vectastain Elite, Vector Laboratories, Burlingame, CA, USA,
1 : 1000) with nickel-enhanced diaminobenzidine as a chromogene.
Microscopy
The stained sections were examined with a microscope (Leica
Microsystems, Cambridge, UK) and the images were captured by a
high-resolution CCD video camera image and fed into a personal
computer. Subsequently, the Quantimet 600 Image Analysis System
(Leica Microsystems) generated a digitized signal proportional to the
intensity of illumination. Photomicrographs of labelled structures
were obtained using this system and saved as TIFF ®les. Image-
editing software (Adobe Photoshop, Adobe Systems, San Jose, CA,
USA) was used to adjust contrast and brightness only. Image
processing was performed on the grey image by de®ning, for
example, brightness and surface above which labelling has to be
taken into account. Once established, these parameters remained
unchanged. The image was then converted to a binary image and
measurements were taken. The number of Fos-immunoreactive cells
was measured in at least four sections through the ventromedial
preoptic area, the parvocellular part of the paraventricular nucleus of
the hypothalamus, the central nucleus of the amygdala, the
dorsolateral part of the bed nucleus of the stria terminalis and the
nucleus of the solitary tract. These structures were previously shown
to contain Fos-positive cells after i.p. injection of the same dose and
serotype of LPS (Konsman et al., 1999). In the same study, LPS was
shown to induce IL-1b immunoreactivity in circumventricular
organs. To test the hypothesis that vagotomy reduces the induction
of IL-1b in the forebrain after LPS injection, the relative surface of
IL-1b-immunoreactive cells in sections of the subfornical organ
relative to the total surface of this organ was used as a measure of IL-
1b induction. The subfornical organ was chosen among circumven-
tricular organs for its size and regular shape.
Statistical analysis
In order to verify that surgery did not affect behaviour or body
temperature of vagotomized animals compared with sham-operated
animals, duration of social investigation and body temperature were
measured before injection and submitted to a one-way ANOVA. The
duration of social investigation 2 h after injection is expressed as a
percentage of baseline and submitted to a one-way, repeated-measure
ANOVA (surgery as a between-subjects factor and treatment as a
within-subject factor). Temperature data from fever experiments are
presented every 30 min as means 6 SEM and were analysed by a
two-way, repeated-measure ANOVA (surgery and treatment as
between-subject factors and time within-subject factor, respectively).
Post hoc comparisons of individual group means were carried out by
the Newman±Keuls test. Data from immunocytochemistry experi-
ments are expressed as means 6 SEM and were analysed by a two-
way ANOVA (surgery and treatment). In all tests a level of P < 0.05
was considered as statistically signi®cant.
Results
Veri®cation of vagotomy
In animals assigned to fever and behavioural studies, vagotomy
was veri®ed by blockade of the anorexic effect of i.p. injected
cholecystokinin. A two-way ANOVA on the amount of food intake
over 30 min after 18 h of food deprivation revealed a signi®cant
effect of cholecystokinin treatment (F1,17 = 15.1; P < 0.01), sur-
gery (F1,17 = 14.6; P < 0.01) and a signi®cant interaction between
cholecystokinin treatment and surgery (F1,17 = 27.56; P < 0.001).
Cholecystokinin (4 mg/kg, i.p.) signi®cantly inhibited food intake
in sham-operated animals (P < 0.001), but had no effect on food
consumption of vagotomized animals (P > 0.10), as previously
shown (Hansen & Krueger, 1997). As abdominal vagotomy blocks
the satiety effect of 4 mg/kg of cholecystokinin injected intraper-
itoneally (Smith et al., 1981), this indicates that the animals used
in the fever and behavioural studies were successfully vagoto-
mized.
Vagotomy was veri®ed by isolectin I B4 binding in the NTS of
those animals used in immunocytochemical studies. Isolectin I B4
binds vagal afferents in the NTS (Li et al., 1997) and its
distribution in the NTS of sham-operated rats was similar to that
reported previously. High isolectin I B4 binding was found in the
medial and commissural nuclei, as well as in the tractus solitarius
of sham-operated animals, as shown in Fig. 1A. After subdiaph-
ragmatic vagotomy, isolectin I B4 binding in these structures was
highly reduced in most animals (Fig. 1C and D). In two animals,
VGX 12, shown in Fig. 1B, and VGX 24, however, isolectin I B4
binding was intermediate between sham-operated animals and the
4436 J. P. Konsman et al.
ã 2000 Federation of European Neuroscience Societies, European Journal of Neuroscience, 12, 4434±4446

remainder of the rats that underwent vagotomy. These animals
were considered to be incompletely vagotomized and excluded
from further analysis.
Vagotomy blocks i.p. IL-1b- and LPS -induced behaviouraldepression but not fever
Behavioural depression
A one-way ANOVA on the duration of social investigation measured
before injection did not reveal any difference between vagotomized
and sham-operated rats (F1,32 = 2.45; P > 0.10). A one-way, repeated
measures ANOVA on the duration of social interaction 2 h after
injection of rrIL-1b or saline revealed a signi®cant effect of rrIL-1binjection (F1,8 = 16.6; P < 0.01) and a signi®cant interaction between
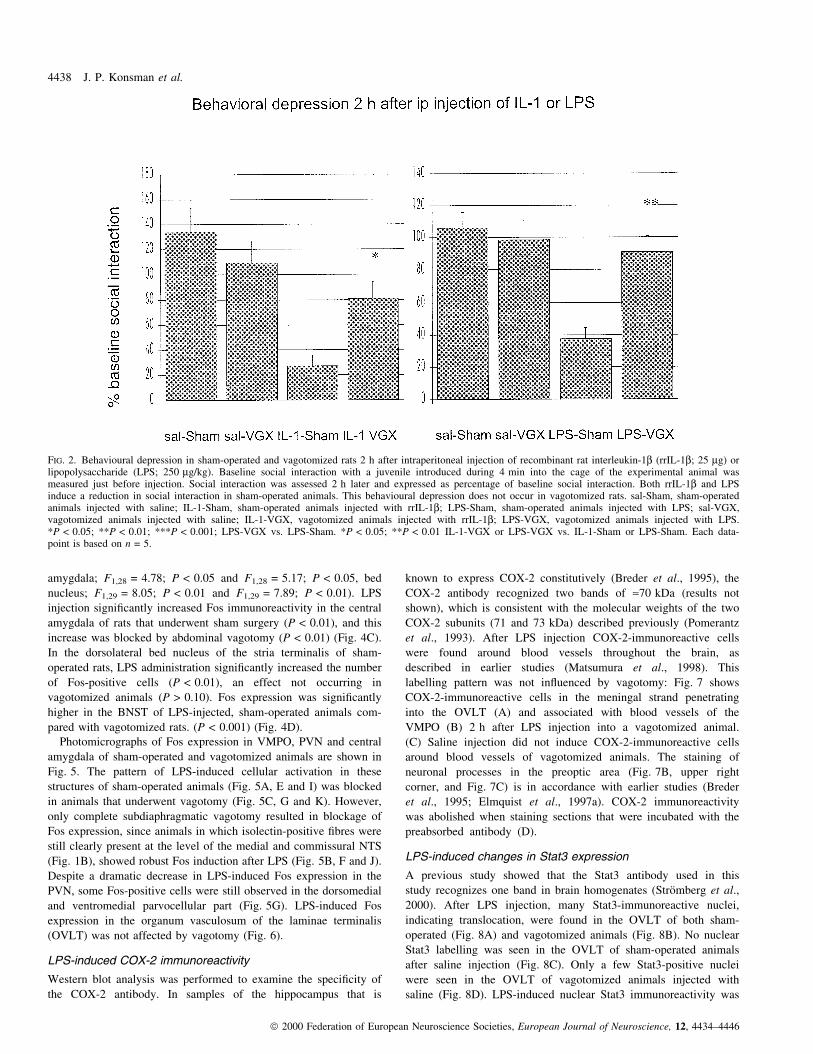
rrIL-1b injection and surgery (F1,8 = 5.71; P < 0.05). rrIL-1binjection signi®cantly reduced social interaction in sham-operated
animals (P < 0.01), but not in vagotomized animals (Fig. 2A). These
animals were immobile, showed piloerection and curled up to
minimize heat loss (Hart, 1988). Vagotomy signi®cantly attenuated
this effect of rrIL-1b (P < 0.05).
A one-way, repeated-measures ANOVA on social interaction 2 h
after injection of saline or LPS, revealed a signi®cant effect of
surgery (F1,8 = 5.63; P < 0.05), LPS injection (F1,8 = 14.5;
P < 0.01) and a signi®cant interaction between surgery and LPS
injection (F1,8 = 9.21; P < 0.05). LPS injection signi®cantly reduced
social interaction in sham-operated animals (P < 0.01), but not in
vagotomized animals (Fig. 2B). Vagotomy signi®cantly attenuated
this effect of LPS (P < 0.01).
Fever
A one-way ANOVA on basal temperatures was measured over the
second hour after light onset revealed that body temperature was
slightly, but consistently lower in vagotomized animals (mean
36.64 6 0.28 °C) compared with sham-operated rats (mean
36.89 6 0.21 °C; F1,68 = 17.0; P < 0.001). This is in accordance
with earlier studies conducted at an ambient temperature of 22 °C (Lin
& Chern, 1985; Watkins et al., 1995; Fleshner et al., 1998) and may be
due to sectioning of thermosensitive afferent ®bres in the hepatic
branch of the vagus nerve (Adachi, 1984). However, in contrast to an
earlier study (Watkins et al., 1995), vagotomy did not prevent the rise
in body temperature after handling stress associated with the i.p.
injection. This indicates that temperature control mechanisms were
largely intact in the present study. This initial rise in temperature
normally persisted for 1 h, after which it returned to basal levels.
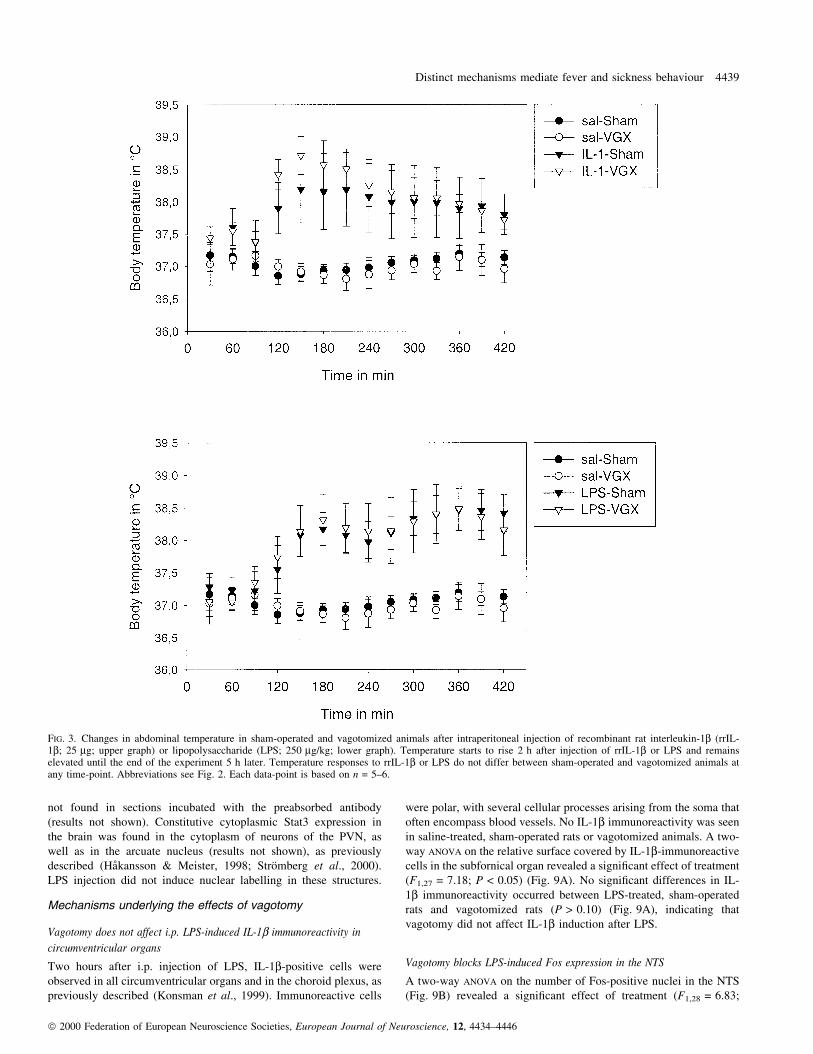
A two-way, repeated measures ANOVA revealed a signi®cant effect
of treatment between subjects (F2,28 = 10.9; P < 0.001) and a
signi®cant effect of time within subjects (F13,364 = 12.5; P < 0.001)
as well as a signi®cant interaction between treatment and time
(F26,364 = 7.21; P < 0.001). Both i.p. injection of rrIL-1b and LPS
induced robust fevers, compared with saline injection (P < 0.001 and
P < 0.01, respectively). Body temperature started to rise 2 h after
rrIL-1b or LPS injection (P < 0.01), and was still elevated at the end
of the experiment (P < 0.001) (Fig. 3). Apart from the early stress-
related transient rise, there were no differences in febrile responses to
rrIL-1b and LPS between vagotomized and sham-operated animals
(P > 0.10).
Effects of vagotomy on i.p. LPS-induced Fos expression in theforebrain
As 25 mg/kg of rrIL-1b and 250 mg/kg of LPS gave rise to similar
febrile responses and behavioural depression, immunocytochemical
studies were not performed for rrIL-1b-injected rats. In addition, it
has previously been shown that IL-1b and LPS induce Fos expression
in the same brain structures (Ericsson et al., 1994; Elmquist et al.,
1996).
A two-way ANOVA on the number of Fos-positive cells in the
paraventricular nucleus of the hypothalamus (PVN) revealed a
signi®cant effect of treatment (saline or LPS) (F1,29 = 12.1;
P < 0.01), surgery (sham or vagotomy) (F1,29 = 10.1; P < 0.01)
and a signi®cant interaction between treatment and surgery
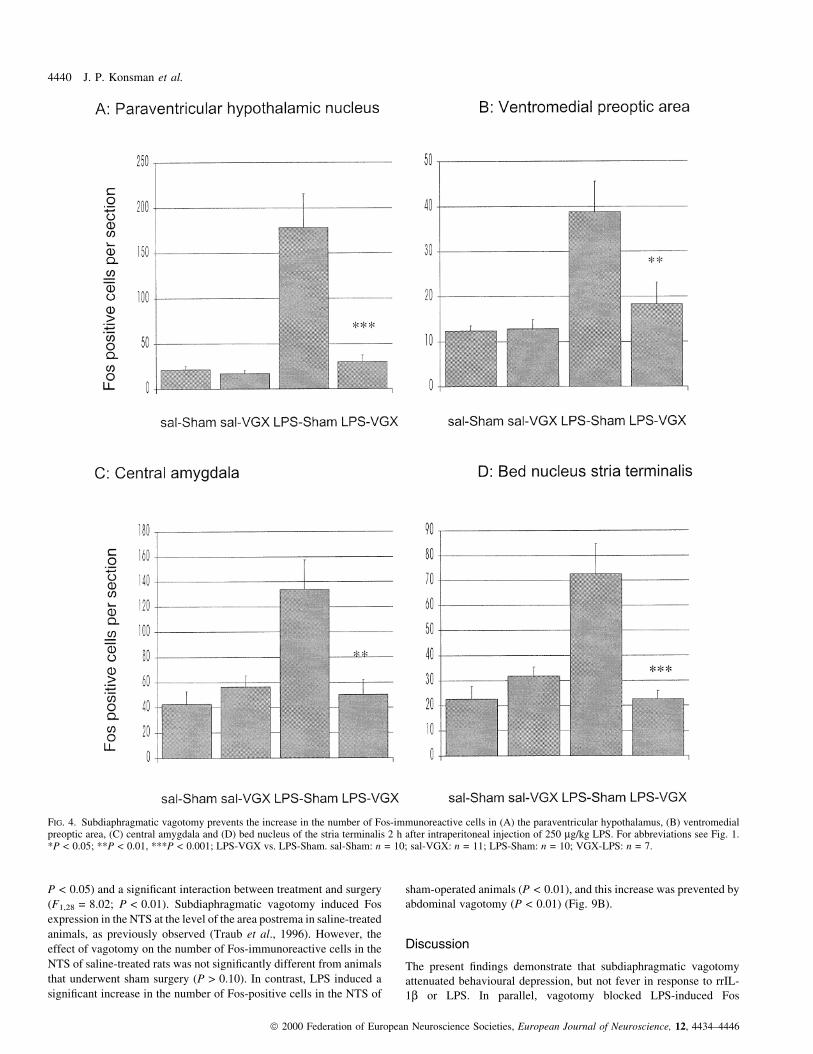
(F1,29 = 8.64; P < 0.01). LPS signi®cantly increased the number of
Fos-positive cells in the PVN of sham-operated animals (P < 0.001).
The PVN of LPS-injected, sham-operated animals showed signi®-
cantly more Fos-immunoreactive cells (P < 0.001) (Fig. 4A) com-
pared with LPS-treated vagotomized animals in which no signi®cant
Fos induction was found compared with saline-injected rats
(P > 0.10). A two-way ANOVA on the number of Fos-immunoreactive
cells in the ventromedial preoptic area (VMPO) revealed a signi®cant
effect of treatment and a signi®cant interaction between treatment and
surgery (F1,29 = 10.7; P < 0.01 and (F1,29 = 4.80; P < 0.05), respect-
ively). The VMPO of LPS-treated sham-operated animals contained
signi®cantly greater numbers of Fos-immunoreactive cells compared
with saline injection (P < 0.01). This was not the case in animals that
underwent vagotomy (P > 0.10). As a result, Fos expression was
signi®cantly higher in sham-operated rats treated with LPS compared
with vagotomized animals (P < 0.01, Fig. 4B).
A two-way ANOVA on the number of Fos-positive nuclei in the
central amygdala (Fig. 4C) and dorsolateral bed nucleus of the stria
terminalis (BNST; Fig. 4D) revealed a signi®cant effect of surgery
and a signi®cant interaction between treatment and surgery (central
FIG. 1. Isolectin I B4 binding to vagal ®bres in the nucleus of the solitarytract (NTS) as a means to verify the extent of subdiaphragmatic vagotomy.(A) Isolectin binding in the commissural and medial NTS of a sham-operated animal (Sham 48). (B) NTS sections of an animal that underwentsubdiaphragmatic vagotomy (VGX 12), but still shows considerableisolectin binding compared to the vagotomized animals shown in C and D(VGX 32 and VGX 30, respectively). Animal VGX 12 was therefore judgedto be incompletely vagotomized. Note also the high isolectin binding in thearea postrema (AP) in A and B compared with C and D (cc, central canal).Scale bar, 100 mm.
Distinct mechanisms mediate fever and sickness behaviour 4437
ã 2000 Federation of European Neuroscience Societies, European Journal of Neuroscience, 12, 4434±4446
amygdala; F1,28 = 4.78; P < 0.05 and F1,28 = 5.17; P < 0.05, bed
nucleus; F1,29 = 8.05; P < 0.01 and F1,29 = 7.89; P < 0.01). LPS
injection signi®cantly increased Fos immunoreactivity in the central
amygdala of rats that underwent sham surgery (P < 0.01), and this
increase was blocked by abdominal vagotomy (P < 0.01) (Fig. 4C).
In the dorsolateral bed nucleus of the stria terminalis of sham-
operated rats, LPS administration signi®cantly increased the number
of Fos-positive cells (P < 0.01), an effect not occurring in
vagotomized animals (P > 0.10). Fos expression was signi®cantly
higher in the BNST of LPS-injected, sham-operated animals com-
pared with vagotomized rats. (P < 0.001) (Fig. 4D).
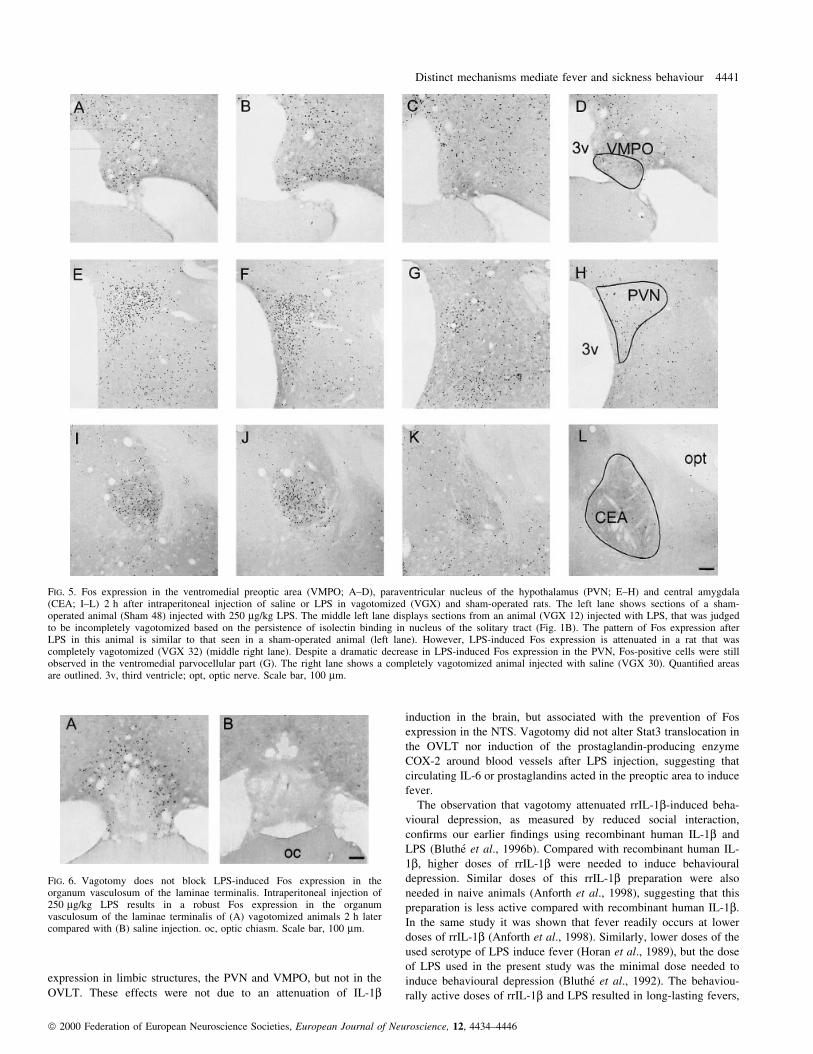
Photomicrographs of Fos expression in VMPO, PVN and central
amygdala of sham-operated and vagotomized animals are shown in
Fig. 5. The pattern of LPS-induced cellular activation in these
structures of sham-operated animals (Fig. 5A, E and I) was blocked
in animals that underwent vagotomy (Fig. 5C, G and K). However,
only complete subdiaphragmatic vagotomy resulted in blockage of
Fos expression, since animals in which isolectin-positive ®bres were
still clearly present at the level of the medial and commissural NTS
(Fig. 1B), showed robust Fos induction after LPS (Fig. 5B, F and J).
Despite a dramatic decrease in LPS-induced Fos expression in the
PVN, some Fos-positive cells were still observed in the dorsomedial
and ventromedial parvocellular part (Fig. 5G). LPS-induced Fos
expression in the organum vasculosum of the laminae terminalis
(OVLT) was not affected by vagotomy (Fig. 6).
LPS-induced COX-2 immunoreactivity
Western blot analysis was performed to examine the speci®city of
the COX-2 antibody. In samples of the hippocampus that is
known to express COX-2 constitutively (Breder et al., 1995), the
COX-2 antibody recognized two bands of »70 kDa (results not
shown), which is consistent with the molecular weights of the two
COX-2 subunits (71 and 73 kDa) described previously (Pomerantz
et al., 1993). After LPS injection COX-2-immunoreactive cells
were found around blood vessels throughout the brain, as
described in earlier studies (Matsumura et al., 1998). This
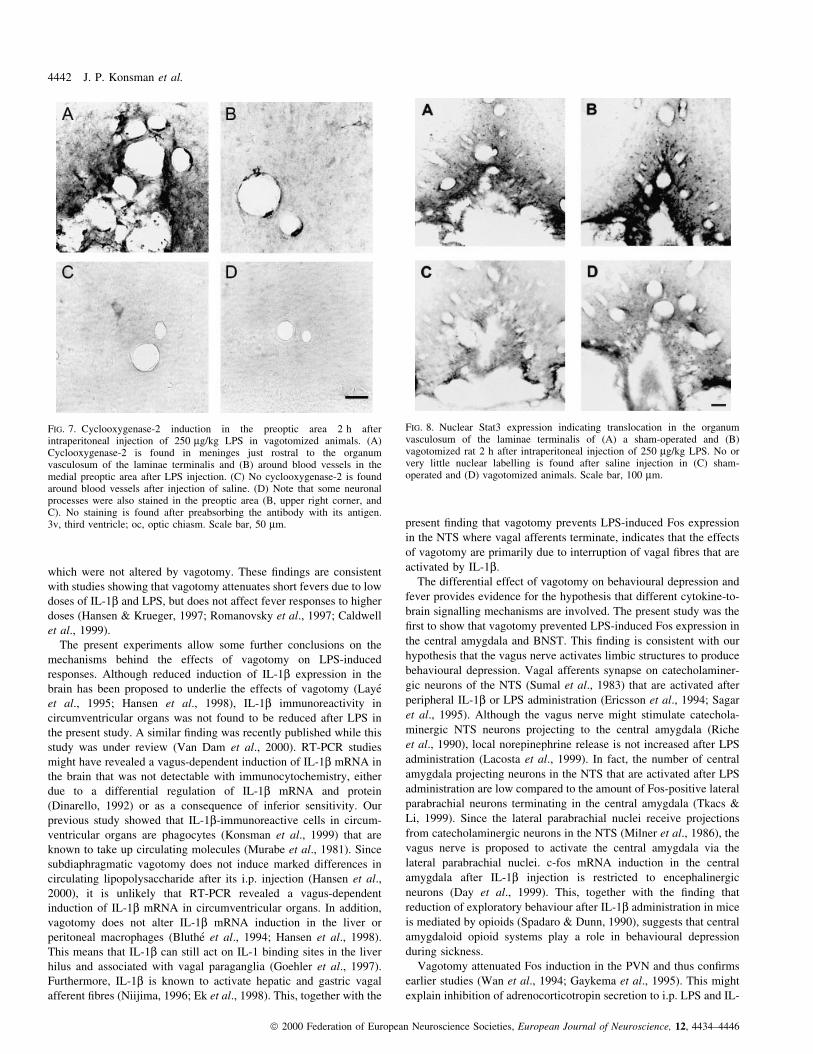
labelling pattern was not in¯uenced by vagotomy: Fig. 7 shows
COX-2-immunoreactive cells in the meningal strand penetrating
into the OVLT (A) and associated with blood vessels of the
VMPO (B) 2 h after LPS injection into a vagotomized animal.
(C) Saline injection did not induce COX-2-immunoreactive cells
around blood vessels of vagotomized animals. The staining of
neuronal processes in the preoptic area (Fig. 7B, upper right
corner, and Fig. 7C) is in accordance with earlier studies (Breder
et al., 1995; Elmquist et al., 1997a). COX-2 immunoreactivity
was abolished when staining sections that were incubated with the
preabsorbed antibody (D).
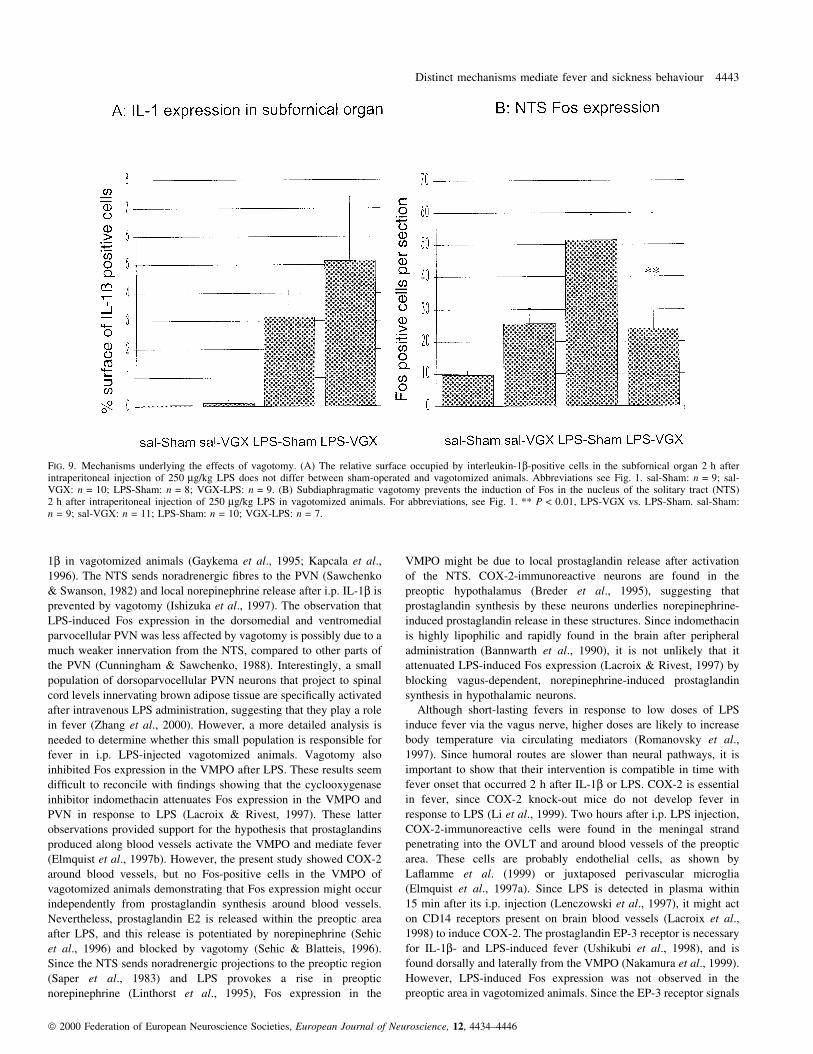
LPS-induced changes in Stat3 expression
A previous study showed that the Stat3 antibody used in this
study recognizes one band in brain homogenates (StroÈmberg et al.,
2000). After LPS injection, many Stat3-immunoreactive nuclei,
indicating translocation, were found in the OVLT of both sham-
operated (Fig. 8A) and vagotomized animals (Fig. 8B). No nuclear
Stat3 labelling was seen in the OVLT of sham-operated animals
after saline injection (Fig. 8C). Only a few Stat3-positive nuclei
were seen in the OVLT of vagotomized animals injected with
saline (Fig. 8D). LPS-induced nuclear Stat3 immunoreactivity was
FIG. 2. Behavioural depression in sham-operated and vagotomized rats 2 h after intraperitoneal injection of recombinant rat interleukin-1b (rrIL-1b; 25 mg) orlipopolysaccharide (LPS; 250 mg/kg). Baseline social interaction with a juvenile introduced during 4 min into the cage of the experimental animal wasmeasured just before injection. Social interaction was assessed 2 h later and expressed as percentage of baseline social interaction. Both rrIL-1b and LPSinduce a reduction in social interaction in sham-operated animals. This behavioural depression does not occur in vagotomized rats. sal-Sham, sham-operatedanimals injected with saline; IL-1-Sham, sham-operated animals injected with rrIL-1b; LPS-Sham, sham-operated animals injected with LPS; sal-VGX,vagotomized animals injected with saline; IL-1-VGX, vagotomized animals injected with rrIL-1b; LPS-VGX, vagotomized animals injected with LPS.*P < 0.05; **P < 0.01; ***P < 0.001; LPS-VGX vs. LPS-Sham. *P < 0.05; **P < 0.01 IL-1-VGX or LPS-VGX vs. IL-1-Sham or LPS-Sham. Each data-point is based on n = 5.
4438 J. P. Konsman et al.
ã 2000 Federation of European Neuroscience Societies, European Journal of Neuroscience, 12, 4434±4446
not found in sections incubated with the preabsorbed antibody
(results not shown). Constitutive cytoplasmic Stat3 expression in
the brain was found in the cytoplasm of neurons of the PVN, as
well as in the arcuate nucleus (results not shown), as previously
described (HaÊkansson & Meister, 1998; StroÈmberg et al., 2000).
LPS injection did not induce nuclear labelling in these structures.
Mechanisms underlying the effects of vagotomy
Vagotomy does not affect i.p. LPS-induced IL-1b immunoreactivity in
circumventricular organs
Two hours after i.p. injection of LPS, IL-1b-positive cells were
observed in all circumventricular organs and in the choroid plexus, as
previously described (Konsman et al., 1999). Immunoreactive cells
were polar, with several cellular processes arising from the soma that
often encompass blood vessels. No IL-1b immunoreactivity was seen
in saline-treated, sham-operated rats or vagotomized animals. A two-
way ANOVA on the relative surface covered by IL-1b-immunoreactive
cells in the subfornical organ revealed a signi®cant effect of treatment
(F1,27 = 7.18; P < 0.05) (Fig. 9A). No signi®cant differences in IL-
1b immunoreactivity occurred between LPS-treated, sham-operated
rats and vagotomized rats (P > 0.10) (Fig. 9A), indicating that
vagotomy did not affect IL-1b induction after LPS.
Vagotomy blocks LPS-induced Fos expression in the NTS
A two-way ANOVA on the number of Fos-positive nuclei in the NTS
(Fig. 9B) revealed a signi®cant effect of treatment (F1,28 = 6.83;
FIG. 3. Changes in abdominal temperature in sham-operated and vagotomized animals after intraperitoneal injection of recombinant rat interleukin-1b (rrIL-1b; 25 mg; upper graph) or lipopolysaccharide (LPS; 250 mg/kg; lower graph). Temperature starts to rise 2 h after injection of rrIL-1b or LPS and remainselevated until the end of the experiment 5 h later. Temperature responses to rrIL-1b or LPS do not differ between sham-operated and vagotomized animals atany time-point. Abbreviations see Fig. 2. Each data-point is based on n = 5±6.
Distinct mechanisms mediate fever and sickness behaviour 4439
ã 2000 Federation of European Neuroscience Societies, European Journal of Neuroscience, 12, 4434±4446
P < 0.05) and a signi®cant interaction between treatment and surgery
(F1,28 = 8.02; P < 0.01). Subdiaphragmatic vagotomy induced Fos
expression in the NTS at the level of the area postrema in saline-treated
animals, as previously observed (Traub et al., 1996). However, the
effect of vagotomy on the number of Fos-immunoreactive cells in the
NTS of saline-treated rats was not signi®cantly different from animals
that underwent sham surgery (P > 0.10). In contrast, LPS induced a
signi®cant increase in the number of Fos-positive cells in the NTS of
sham-operated animals (P < 0.01), and this increase was prevented by
abdominal vagotomy (P < 0.01) (Fig. 9B).
Discussion
The present ®ndings demonstrate that subdiaphragmatic vagotomy
attenuated behavioural depression, but not fever in response to rrIL-
1b or LPS. In parallel, vagotomy blocked LPS-induced Fos
FIG. 4. Subdiaphragmatic vagotomy prevents the increase in the number of Fos-immunoreactive cells in (A) the paraventricular hypothalamus, (B) ventromedialpreoptic area, (C) central amygdala and (D) bed nucleus of the stria terminalis 2 h after intraperitoneal injection of 250 mg/kg LPS. For abbreviations see Fig. 1.*P < 0.05; **P < 0.01, ***P < 0.001; LPS-VGX vs. LPS-Sham. sal-Sham: n = 10; sal-VGX: n = 11; LPS-Sham: n = 10; VGX-LPS: n = 7.
4440 J. P. Konsman et al.
ã 2000 Federation of European Neuroscience Societies, European Journal of Neuroscience, 12, 4434±4446
expression in limbic structures, the PVN and VMPO, but not in the
OVLT. These effects were not due to an attenuation of IL-1b
induction in the brain, but associated with the prevention of Fos
expression in the NTS. Vagotomy did not alter Stat3 translocation in
the OVLT nor induction of the prostaglandin-producing enzyme
COX-2 around blood vessels after LPS injection, suggesting that
circulating IL-6 or prostaglandins acted in the preoptic area to induce
fever.
The observation that vagotomy attenuated rrIL-1b-induced beha-
vioural depression, as measured by reduced social interaction,
con®rms our earlier ®ndings using recombinant human IL-1b and
LPS (Bluthe et al., 1996b). Compared with recombinant human IL-
1b, higher doses of rrIL-1b were needed to induce behavioural
depression. Similar doses of this rrIL-1b preparation were also
needed in naive animals (Anforth et al., 1998), suggesting that this
preparation is less active compared with recombinant human IL-1b.
In the same study it was shown that fever readily occurs at lower
doses of rrIL-1b (Anforth et al., 1998). Similarly, lower doses of the
used serotype of LPS induce fever (Horan et al., 1989), but the dose
of LPS used in the present study was the minimal dose needed to
induce behavioural depression (Bluthe et al., 1992). The behaviou-
rally active doses of rrIL-1b and LPS resulted in long-lasting fevers,
FIG. 5. Fos expression in the ventromedial preoptic area (VMPO; A±D), paraventricular nucleus of the hypothalamus (PVN; E±H) and central amygdala(CEA; I±L) 2 h after intraperitoneal injection of saline or LPS in vagotomized (VGX) and sham-operated rats. The left lane shows sections of a sham-operated animal (Sham 48) injected with 250 mg/kg LPS. The middle left lane displays sections from an animal (VGX 12) injected with LPS, that was judgedto be incompletely vagotomized based on the persistence of isolectin binding in nucleus of the solitary tract (Fig. 1B). The pattern of Fos expression afterLPS in this animal is similar to that seen in a sham-operated animal (left lane). However, LPS-induced Fos expression is attenuated in a rat that wascompletely vagotomized (VGX 32) (middle right lane). Despite a dramatic decrease in LPS-induced Fos expression in the PVN, Fos-positive cells were stillobserved in the ventromedial parvocellular part (G). The right lane shows a completely vagotomized animal injected with saline (VGX 30). Quanti®ed areasare outlined. 3v, third ventricle; opt, optic nerve. Scale bar, 100 mm.
FIG. 6. Vagotomy does not block LPS-induced Fos expression in theorganum vasculosum of the laminae terminalis. Intraperitoneal injection of250 mg/kg LPS results in a robust Fos expression in the organumvasculosum of the laminae terminalis of (A) vagotomized animals 2 h latercompared with (B) saline injection. oc, optic chiasm. Scale bar, 100 mm.
Distinct mechanisms mediate fever and sickness behaviour 4441
ã 2000 Federation of European Neuroscience Societies, European Journal of Neuroscience, 12, 4434±4446
which were not altered by vagotomy. These ®ndings are consistent
with studies showing that vagotomy attenuates short fevers due to low
doses of IL-1b and LPS, but does not affect fever responses to higher
doses (Hansen & Krueger, 1997; Romanovsky et al., 1997; Caldwell
et al., 1999).
The present experiments allow some further conclusions on the
mechanisms behind the effects of vagotomy on LPS-induced
responses. Although reduced induction of IL-1b expression in the
brain has been proposed to underlie the effects of vagotomy (LayeÂ
et al., 1995; Hansen et al., 1998), IL-1b immunoreactivity in
circumventricular organs was not found to be reduced after LPS in
the present study. A similar ®nding was recently published while this
study was under review (Van Dam et al., 2000). RT-PCR studies
might have revealed a vagus-dependent induction of IL-1b mRNA in
the brain that was not detectable with immunocytochemistry, either
due to a differential regulation of IL-1b mRNA and protein
(Dinarello, 1992) or as a consequence of inferior sensitivity. Our
previous study showed that IL-1b-immunoreactive cells in circum-
ventricular organs are phagocytes (Konsman et al., 1999) that are
known to take up circulating molecules (Murabe et al., 1981). Since
subdiaphragmatic vagotomy does not induce marked differences in
circulating lipopolysaccharide after its i.p. injection (Hansen et al.,
2000), it is unlikely that RT-PCR revealed a vagus-dependent
induction of IL-1b mRNA in circumventricular organs. In addition,
vagotomy does not alter IL-1b mRNA induction in the liver or
peritoneal macrophages (Bluthe et al., 1994; Hansen et al., 1998).
This means that IL-1b can still act on IL-1 binding sites in the liver
hilus and associated with vagal paraganglia (Goehler et al., 1997).
Furthermore, IL-1b is known to activate hepatic and gastric vagal
afferent ®bres (Niijima, 1996; Ek et al., 1998). This, together with the
present ®nding that vagotomy prevents LPS-induced Fos expression
in the NTS where vagal afferents terminate, indicates that the effects
of vagotomy are primarily due to interruption of vagal ®bres that are
activated by IL-1b.
The differential effect of vagotomy on behavioural depression and
fever provides evidence for the hypothesis that different cytokine-to-
brain signalling mechanisms are involved. The present study was the
®rst to show that vagotomy prevented LPS-induced Fos expression in
the central amygdala and BNST. This ®nding is consistent with our
hypothesis that the vagus nerve activates limbic structures to produce
behavioural depression. Vagal afferents synapse on catecholaminer-
gic neurons of the NTS (Sumal et al., 1983) that are activated after
peripheral IL-1b or LPS administration (Ericsson et al., 1994; Sagar
et al., 1995). Although the vagus nerve might stimulate catechola-
minergic NTS neurons projecting to the central amygdala (Riche
et al., 1990), local norepinephrine release is not increased after LPS
administration (Lacosta et al., 1999). In fact, the number of central
amygdala projecting neurons in the NTS that are activated after LPS
administration are low compared to the amount of Fos-positive lateral
parabrachial neurons terminating in the central amygdala (Tkacs &
Li, 1999). Since the lateral parabrachial nuclei receive projections
from catecholaminergic neurons in the NTS (Milner et al., 1986), the
vagus nerve is proposed to activate the central amygdala via the
lateral parabrachial nuclei. c-fos mRNA induction in the central
amygdala after IL-1b injection is restricted to encephalinergic
neurons (Day et al., 1999). This, together with the ®nding that
reduction of exploratory behaviour after IL-1b administration in mice
is mediated by opioids (Spadaro & Dunn, 1990), suggests that central
amygdaloid opioid systems play a role in behavioural depression
during sickness.
Vagotomy attenuated Fos induction in the PVN and thus con®rms
earlier studies (Wan et al., 1994; Gaykema et al., 1995). This might
explain inhibition of adrenocorticotropin secretion to i.p. LPS and IL-
FIG. 7. Cyclooxygenase-2 induction in the preoptic area 2 h afterintraperitoneal injection of 250 mg/kg LPS in vagotomized animals. (A)Cyclooxygenase-2 is found in meninges just rostral to the organumvasculosum of the laminae terminalis and (B) around blood vessels in themedial preoptic area after LPS injection. (C) No cyclooxygenase-2 is foundaround blood vessels after injection of saline. (D) Note that some neuronalprocesses were also stained in the preoptic area (B, upper right corner, andC). No staining is found after preabsorbing the antibody with its antigen.3v, third ventricle; oc, optic chiasm. Scale bar, 50 mm.
FIG. 8. Nuclear Stat3 expression indicating translocation in the organumvasculosum of the laminae terminalis of (A) a sham-operated and (B)vagotomized rat 2 h after intraperitoneal injection of 250 mg/kg LPS. No orvery little nuclear labelling is found after saline injection in (C) sham-operated and (D) vagotomized animals. Scale bar, 100 mm.
4442 J. P. Konsman et al.
ã 2000 Federation of European Neuroscience Societies, European Journal of Neuroscience, 12, 4434±4446
1b in vagotomized animals (Gaykema et al., 1995; Kapcala et al.,
1996). The NTS sends noradrenergic ®bres to the PVN (Sawchenko
& Swanson, 1982) and local norepinephrine release after i.p. IL-1b is
prevented by vagotomy (Ishizuka et al., 1997). The observation that
LPS-induced Fos expression in the dorsomedial and ventromedial
parvocellular PVN was less affected by vagotomy is possibly due to a
much weaker innervation from the NTS, compared to other parts of
the PVN (Cunningham & Sawchenko, 1988). Interestingly, a small
population of dorsoparvocellular PVN neurons that project to spinal
cord levels innervating brown adipose tissue are speci®cally activated
after intravenous LPS administration, suggesting that they play a role
in fever (Zhang et al., 2000). However, a more detailed analysis is
needed to determine whether this small population is responsible for
fever in i.p. LPS-injected vagotomized animals. Vagotomy also
inhibited Fos expression in the VMPO after LPS. These results seem
dif®cult to reconcile with ®ndings showing that the cyclooxygenase
inhibitor indomethacin attenuates Fos expression in the VMPO and
PVN in response to LPS (Lacroix & Rivest, 1997). These latter
observations provided support for the hypothesis that prostaglandins
produced along blood vessels activate the VMPO and mediate fever
(Elmquist et al., 1997b). However, the present study showed COX-2
around blood vessels, but no Fos-positive cells in the VMPO of
vagotomized animals demonstrating that Fos expression might occur
independently from prostaglandin synthesis around blood vessels.
Nevertheless, prostaglandin E2 is released within the preoptic area
after LPS, and this release is potentiated by norepinephrine (Sehic
et al., 1996) and blocked by vagotomy (Sehic & Blatteis, 1996).
Since the NTS sends noradrenergic projections to the preoptic region
(Saper et al., 1983) and LPS provokes a rise in preoptic
norepinephrine (Linthorst et al., 1995), Fos expression in the
VMPO might be due to local prostaglandin release after activation
of the NTS. COX-2-immunoreactive neurons are found in the
preoptic hypothalamus (Breder et al., 1995), suggesting that
prostaglandin synthesis by these neurons underlies norepinephrine-
induced prostaglandin release in these structures. Since indomethacin
is highly lipophilic and rapidly found in the brain after peripheral
administration (Bannwarth et al., 1990), it is not unlikely that it
attenuated LPS-induced Fos expression (Lacroix & Rivest, 1997) by
blocking vagus-dependent, norepinephrine-induced prostaglandin
synthesis in hypothalamic neurons.
Although short-lasting fevers in response to low doses of LPS
induce fever via the vagus nerve, higher doses are likely to increase
body temperature via circulating mediators (Romanovsky et al.,
1997). Since humoral routes are slower than neural pathways, it is
important to show that their intervention is compatible in time with
fever onset that occurred 2 h after IL-1b or LPS. COX-2 is essential
in fever, since COX-2 knock-out mice do not develop fever in
response to LPS (Li et al., 1999). Two hours after i.p. LPS injection,
COX-2-immunoreactive cells were found in the meningal strand
penetrating into the OVLT and around blood vessels of the preoptic
area. These cells are probably endothelial cells, as shown by
La¯amme et al. (1999) or juxtaposed perivascular microglia
(Elmquist et al., 1997a). Since LPS is detected in plasma within
15 min after its i.p. injection (Lenczowski et al., 1997), it might act
on CD14 receptors present on brain blood vessels (Lacroix et al.,
1998) to induce COX-2. The prostaglandin EP-3 receptor is necessary
for IL-1b- and LPS-induced fever (Ushikubi et al., 1998), and is
found dorsally and laterally from the VMPO (Nakamura et al., 1999).
However, LPS-induced Fos expression was not observed in the
preoptic area in vagotomized animals. Since the EP-3 receptor signals
FIG. 9. Mechanisms underlying the effects of vagotomy. (A) The relative surface occupied by interleukin-1b-positive cells in the subfornical organ 2 h afterintraperitoneal injection of 250 mg/kg LPS does not differ between sham-operated and vagotomized animals. Abbreviations see Fig. 1. sal-Sham: n = 9; sal-VGX: n = 10; LPS-Sham: n = 8; VGX-LPS: n = 9. (B) Subdiaphragmatic vagotomy prevents the induction of Fos in the nucleus of the solitary tract (NTS)2 h after intraperitoneal injection of 250 mg/kg LPS in vagotomized animals. For abbreviations, see Fig. 1. ** P < 0.01, LPS-VGX vs. LPS-Sham. sal-Sham:n = 9; sal-VGX: n = 11; LPS-Sham: n = 10; VGX-LPS: n = 7.
Distinct mechanisms mediate fever and sickness behaviour 4443
ã 2000 Federation of European Neuroscience Societies, European Journal of Neuroscience, 12, 4434±4446

primarily through the inhibitory G-protein (Audoly et al., 1999), and
Fos induction is never observed after inhibitory responses (KovaÂcs,
1998), this might explain the lack of Fos expression in the preoptic
area. Thus, prostaglandins produced by COX-2 around blood vessels
might act on EP-3 receptors in the preoptic area to induce fever
without Fos.
IL-6 is another essential mediator of fever, as IL-6 knock-out
mice do not develop fever after IL-1b or LPS administration
(Chai et al., 1996). Circulating IL-6 rises 1 h after peripheral LPS
administration (Luheshi et al., 1996; Cartmell et al., 2000) and
both peripheral and central pretreatment with anti-IL-6 serum
abolishes fever (Rothwell et al., 1991; Cartmell et al., 2000). We
studied IL-6 action on the brain after LPS injection by detecting
Stat3 that translocates into the nucleus after activation by the
signal transducer gp130 common to the IL-6 receptor family
(Hirano et al., 1994; Heinrich et al., 1998). Two hours after LPS
injection, nuclear Stat3 was found in the OVLT, suggesting that
circulating IL-6 acted on this area devoid of a BBB. Several
®ndings are compatible with the hypothesis that Stat3 and Fos
induction in the OVLT is related to LPS-induced circulating IL-6
action. The OVLT is known to contain mRNA coding for the IL-
6 receptor and the gp130 signal transducer (VallieÁres & Rivest,
1997) and expresses Fos after peripheral IL-6 administration
(VallieÁres et al., 1997). Furthermore, OVLT lesions block LPS-
induced fever in the presence of circulating IL-6 (Caldwell et al.,
1998), suggesting that IL-6 action on the OVLT plays an
important role in fever. When injected into the brain, IL-6
induces fever but not behavioural depression (Lenczowski et al.,
1999). Altogether, this indicates that IL-6 is part of a speci®c
immune-to-brain pathway underlying fever. Although, the relation
between IL-6 action and COX-2 is still unclear, both are essential
in fever (Chai et al., 1996; Li et al., 1999) and were found to
occur at a time-point consistent with the induction of fever in the
present study.
The present work demonstrated that transmission of an immune
message via the vagus nerve activates limbic structures and
induces behavioural depression. Behavioural depression helps to
reduce heat loss upon infection and thus the maintenance of an
elevated body temperature (Hart, 1988). Since doses of rrIL-1band LPS lower than those needed to provoke behavioural
depression readily result in fever, it can be argued that
thermoregulation by behavioural means occurs only during more
severe infections. Although vagotomy may attenuate fever to low
doses of LPS or IL-1b by preventing activation of the VMPO,
our ®ndings show that fever occurring in parallel with behavioural
depression is not affected by vagotomy. In this case, fever is
likely to result from IL-6 action in the OVLT and prostaglandin
synthesis around blood vessels. Taken together, the present
®ndings can be interpreted to suggest that after infection,
increases in the hypothalamic temperature set-point and regulation
of heat loss by behavioural depression are controlled by different
mechanisms of cytokine-to-brain signalling.
Acknowledgements
The authors would like to thank Dr S. Poole (NIBSC, Potters Bar, UK) for hisgift of recombinant rat IL-1b and the corresponding antiserum. These reagentswere produced within the context of the BIOMED I Concerted Action`Cytokines in the Brain' that was funded by the European Community. Wethank J. Keijser (University of Groningen) for the use of and assistance withthe Quantimet 600 Image Analysis System and Professor Dr J.M. Koolhaasand Dr S. Laye for helpful discussion. Supported by INRA, INSERM and theEuropean Community (CT97-0149 and CT97-2492).
Abbreviations
BBB, blood±brain barrier; BNST, bed nucleus of the stria terminalis; COX-2,cyclooxygenase-2; IL-1b, interleukin-1b; IL-6, interleukin-6; i.p., intraper-itoneal; LPS, lipopolysaccharide; NTS, nucleus of the solitary tract; OVLT,organum vasculosum of the laminae terminalis; PVN, paraventricular nucleusof the hypothalamus; rr, recombinant rat; VMPO, ventromedial preoptic area.
References
Adachi, A. (1984) Thermosensitive and osmoreceptive afferent ®bers in thehepatic branch of the vagus nerve. J. Auton. Nerv. Syst., 10, 269±273.
Anforth, H.R., Bluthe, R.M., Bristow, A., Hopkins, S., Lenczowski, M.J.,Luheshi, G., Lundkvist, J., Michaud, B., Mistry, Y., Van Dam, A.M., Zhen,C., Dantzer, R., Poole, S., Rothwell, N.J., Tilders, F.J. & Wollman, E.E.(1998) Biological activity and brain actions of recombinant rat interleukin-1alpha and interleukin-1beta. Eur. Cytokine Netw., 9, 279±288.
Audoly, L.P., Ma, L., Feoktistov, I., de Foe, S.K., Breyer, M.D. & Breyer,R.M. (1999) Prostaglandin E-prostanoid-3 receptor activation of cyclicAMP response element-mediated gene transcription. J. Pharmacol. Exp.Ther., 289, 140±148.
Bannwarth, B., Netter, P., Lapicque, F., PeÂreÂ, P., Thomas, P. & Gaucher, A.(1990) Plasma and cerebrospinal ¯uid concentrations of indomethacin inhumans. Relationship to analgesic activity. Eur. J. Clin. Pharmacol., 38,343±346.
Blatteis, C.M. (1992) Role of the OVLT in the febrile response to circulatingpyrogens. Prog. Brain Res., 91, 409±412.
BlutheÂ, R.M., Dantzer, R. & Kelley, K.W. (1992) Effects of interleukin-1receptor antagonist on the behavioral effects of lipopolysaccharide in rat.Brain Res., 573, 318±320.
BlutheÂ, R.M., Michaud, B., Kelley, K.W. & Dantzer, R. (1996a) Vagotomyattenuates behavioural effects of interleukin-1 injected peripherally but notcentrally. Neuroreport, 7, 1485±1488.
BlutheÂ, R.M., Michaud, B., Kelley, K.W. & Dantzer, R. (1996b) Vagotomyblocks behavioural effects of interleukin-1 injected via the intraperitonealroute but not via other systemic routes. Neuroreport, 7, 2823±2827.
BlutheÂ, R.M., Walter, V., Parnet, P., LayeÂ, S., Lestage, J., Verrier, D., Poole,S., Stenning, B.E., Kelley, K.W. & Dantzer, R. (1994) Lipopolysaccharideinduces sickness behaviour in rats by a vagal mediated mechanism. C. R.Acad. Sci. III, 317, 499±503.
Boulant, J.A. (1981) Hypothalamic mechanisms in thermoregulation. Fed.Proc., 40, 2843±2850.
Breder, C.D., Dewitt, D. & Kraig, R.P. (1995) Characterization of induciblecyclooxygenase in rat brain. J. Comp. Neurol., 355, 296±315.
Caldwell, F.T., Graves, D.B. & Wallace, B.H. (1998) Studies on themechanism of fever after intravenous administration of endotoxin. J.Trauma, 44, 304±312.
Caldwell, F.T.J., Graves, D.B. & Wallace, B.H. (1999) Humoral versus neuralpathways for fever production in rats after administration oflipopolysaccharide. J. Trauma, 47, 120±129.
Cao, C., Matsumura, K., Yamagata, K. & Watanabe, Y. (1996) Endothelialcells of the rat brain vasculature express cyclooxygenase-2 mRNA inresponse to systemic interleukin-1 beta: a possible site of prostaglandinsynthesis responsible for fever. Brain Res., 733, 263±272.
Cartmell, T., Poole, S., Turnbull, A.V., Rothwell, N.J. & Luheshi, G.N. (2000)Circulating interleukin-6 mediates the febrile response to localisedin¯ammation in rats. J. Physiol. (Lond.), 526, 653±661.
Chai, Z., Gatti, S., Toniatti, C., Poli, V. & Bartfai, T. (1996) Interleukin (IL) -6gene expression in the central nervous system is necessary for feverresponse to lipopolysaccharide or IL-1 beta: a study on IL-6-de®cient mice.J. Exp. Med., 183, 311±316.
Cunningham, E.T.J. & Sawchenko, P.E. (1988) Anatomical speci®city ofnoradrenergic inputs to the paraventricular and supraoptic nuclei of the rathypothalamus. J. Comp. Neurol., 274, 60±76.
Dantzer, R. (1994) How do cytokines say hello to the brain? Neural versushumoral mediation. Eur. Cytokine Netw., 5, 271±273.
Day, H.E., Curran, E.J., Watson, S.J. & Akil, H. (1999) Distinctneurochemical poulations in the rat central nucleus and bed nucleus ofthe stria terminalis: evidence for their selective activation by interleukin-1beta. J. Comp. Neurol., 413, 113±128.
Dinarello, C.A. (1992) Dissociation of transcription from translation of humanIL-1 beta: induction of steady state mRNA by adherence or recombinantC5a in the absence of translation. Proc. Soc. Exp. Biol. Med., 200, 228±232.
Ek, M., Kurosawa, M., Lundeberg, T. & Ericsson, A. (1998) Activation of
4444 J. P. Konsman et al.
ã 2000 Federation of European Neuroscience Societies, European Journal of Neuroscience, 12, 4434±4446

vagal afferents after intravenous injection of interleukin-1beta: role ofendogenous prostaglandins. J. Neurosci., 18, 9471±9479.
Elmquist, J.K., Breder, C.D., Sherin, J.E., Scammell, T.E., Hickey, W.F.,Dewitt, D. & Saper, C.B. (1997a) Intravenous lipopolysaccharide inducescyclooxygenase 2-like immunoreactivity in rat brain perivascular microgliaand meningeal macrophages. J. Comp. Neurol., 381, 119±129.
Elmquist, J.K., Scammell, T.E., Jacobson, C.D. & Saper, C.B. (1996)Distribution of Fos-like immunoreactivity in the rat brain followingintravenous lipopolysaccharide administration. J. Comp. Neurol., 371, 85±103.
Elmquist, J.K., Scammell, T.E. & Saper, C.B. (1997b) Mechanisms of CNSresponse to systemic immune challenge: the febrile response. TrendsNeurosci., 20, 565±570.
Ericsson, A., KovaÂcs, K.J. & Sawchenko, P.E. (1994) A functional anatomicalanalysis of central pathways subserving the effects of interleukin-1 onstress-related neuroendocrine neurons. J. Neurosci., 14, 897±913.
Fleshner, M., Goehler, L.E., Schwartz, B.A., McGorry, M., Martin, D., Maier,S.F. & Watkins, L.R. (1998) Thermogenic and corticosterone responses tointravenous cytokines (IL-1beta and TNF-alpha) are attenuated bysubdiaphragmatic vagotomy. J. Neuroimmunol., 86, 134±141.
Gaykema, R.P., Dijkstra, I. & Tilders, F.J. (1995) Subdiaphragmatic vagotomysuppresses endotoxin-induced activation of hypothalamic corticotropin-releasing hormone neurons and ACTH secretion. Endocrinology, 136,4717±4720.
Goehler, L.E., Relton, J.K., Dripps, D., Kiechle, R., Tartaglia, N., Maier, S.F.& Watkins, L.R. (1997) Vagal paraganglia bind biotinylated interleukin-1receptor antagonist: a possible mechanism for immune-to-braincommunication. Brain Res. Bull., 43, 357±364.
HaÊkansson, M.L. & Meister, B. (1998) Transcription factor STAT3 in leptintarget neurons of the rat hypothalamus. Neuroendocrinology, 68, 420±427.
Hansen, M.K. & Krueger, J.M. (1997) Subdiaphragmatic vagotomy blocks thesleep- and fever-promoting effects of interleukin-1beta. Am. J. Physiol.,273, R1246±R1253.
Hansen, M.K., Nguyen, K.T., Fleshner, M., Goehler, L.E., Gaykema, R.P.,Maier, S.F. & Watkins, L.R. (2000) Effects of vagotomy on serumendotoxin, cytokines, and corticosterone after intraperitoneallipopolysaccharide. Am. J. Physiol., 278, R331±R336.
Hansen, M.K., Taishi, P., Chen, Z. & Krueger, J.M. (1998) Vagotomy blocksthe induction of interleukin-1beta (IL-1beta) mRNA in the brain of rats inresponse to systemic IL-1beta. J. Neurosci., 18, 2247±2253.
Hart, B.L. (1988) Biological basis of the behavior of sick animals. Neurosci.Biobehav. Rev., 12, 123±137.
Heinrich, P.C., Behrmann, I., MuÈller-Newen, G., Schaper, F. & Graeve, L.(1998) Interleukin-6-type cytokine signalling through the gp130/Jak/STATpathway. Biochem. J., 334, 297±314.
Hirano, T., Matsuda, T. & Nakajima, K. (1994) Signal transduction throughgp130 that is shared among the receptors for the interleukin 6 relatedcytokine subfamily. Stem Cells, 12, 262±277.
Horan, M.A., Little, R.A., Rothwell, N.J. & Strijbos, P.J. (1989) Comparisonof the effects of several endotoxin presss on body temperature andmetabolic rate in the rat. Can. J. Physiol. Pharmacol., 67, 1011±1014.
Ishizuka, Y., Ishida, Y., Kunitake, T., Kato, K., Hanamori, T., Mitsuyama, Y.& Kannan, H. (1997) Effects of area postrema lesion and abdominalvagotomy on interleukin-1 beta-induced norepinephrine release in thehypothalamic paraventricular nucleus region in the rat. Neurosci. Lett., 223,57±60.
Kapcala, L.P., He, J.R., Gao, Y., Pieper, J.O. & DeTolla, L.J. (1996)Subdiaphragmatic vagotomy inhibits intra-abdominal interleukin-1 betastimulation of adrenocorticotropin secretion. Brain Res., 728, 247±254.
Kluger, M.J. (1991) Fever: role of pyrogens and cryogens. Physiol. Rev., 71,93±127.
Konsman, J.P., Kelley, K. & Dantzer, R. (1999) Temporal and spatialrelationships between lipopolysaccharide-induced expression of Fos,interleukin-1beta and inducible nitric oxide synthase in rat brain.Neuroscience, 89, 535±548.
KovaÂcs, K.J. (1998) c-Fos as a transcription factor: a stressful (re) view from afunctional map. Neurochem. Int., 33, 287±297.
Kraly, F.S., Jerome, C. & Smith, G.P. (1986) Speci®c postoperativesyndromes after total and selective vagotomies in the rat. Appetite, 7, 1±17.
Lacosta, S., Merali, Z. & Anisman, H. (1999) Behavioral and neurochemicalconsequences of lipopolysaccharide in mice: anxiogenic-like effects. BrainRes., 818, 291±303.
Lacroix, S., Feinstein, D. & Rivest, S. (1998) The bacterial endotoxinlipopolysaccharide has the ability to target the brain in upregulating its
membrane CD14 receptor within speci®c cellular populations. BrainPathol., 8, 625±640.
Lacroix, S. & Rivest, S. (1997) Functional circuitry in the brain of immune-challenged rats: partial involvement of prostaglandins. J. Comp. Neurol.,387, 307±324.
La¯amme, N., Lacroix, S. & Rivest, S. (1999) An essential role of interleukin-1û in mediating NF-kappa-B activity and COX-2 transcription in cells of theblood±brain barrier in response to a systemic and localized in¯ammationbut not during endotoxemia. J. Neurosci., 19, 10923±10930.
LayeÂ, S., BlutheÂ, R.M., Kent, S., Combe, C., MeÂdina, C., Parnet, P., Kelley, K.& Dantzer, R. (1995) Subdiaphragmatic vagotomy blocks induction of IL-1beta mRNA in mice brain in response to peripheral LPS. Am. J. Physiol.,268, R1327±R1331.
Lenczowski, M.J., BlutheÂ, R.M., Roth, J., Rees, G.S., Rushforth, D.A., vanDam, A.M., Tilders, F.J., Dantzer, R., Rothwell, N.J. & Luheshi, G.N.(1999) Central administration of rat IL-6 induces HPA activation and feverbut not sickness behavior in rats. Am. J. Physiol., 276, R652±R658.
Lenczowski, M.J., Van Dam, A.M., Poole, S., Larrick, J.W. & Tilders, F.J.(1997) Role of circulating endotoxin and interleukin-6 in the ACTH andcorticosterone response to intraperitoneal LPS. Am. J. Physiol., 273,R1870±R1877.
Li, H., Nomura, S. & Mizuno, N. (1997) Binding of the isolectin I-B4from Griffonia simplicifolia to the general visceral afferents in thevagus nerve: a light- and electron-microscope study in the rat. Neurosci.Lett., 222, 53±56.
Li, S., Wang, Y., Matsumura, K., Ballou, L.R., Morham, S.G. & Blatteis, C.M.(1999) The febrile response to lipopolysaccharide is blocked incyclooxygenase-2 (±/±), but not in cyclooxygenase-1 (±/±) mice. BrainRes., 825, 86±94.
Lin, M.T. & Chern, Y.F. (1985) Effects of subdiaphragmatic vagotomy onthermoregulatory responses of rats to different ambient temperatures. Exp.Neurol., 88, 467±470.
Linthorst, A.C., Flachskamm, C., Holsboer, F. & Reul, J.M. (1995)Intraperitoneal administration of bacterial endotoxin enhancesnoradrenergic neurotransmission in the rat preoptic area: relationship withbody temperature and hypothalamic ± pituitary ± adrenocortical axisactivity. Eur. J. Neurosci., 7, 2418±2430.
Luheshi, G., Miller, A.J., Brouwer, S., Dascombe, M.J., Rothwell, N.J. &Hopkins, S.J. (1996) Interleukin-1 receptor antagonist inhibits endotoxinfever and systemic interleukin-6 induction in the rat. Am. J. Physiol., 270,E91±E95.
Matsumura, K., Cao, C., Ozaki, M., Morii, H., Nakadate, K. & Watanabe, Y.(1998) Brain endothelial cells express cyclooxygenase-2 duringlipopolysaccharide-induced fever: light and electron microscopicimmunocytochemical studies. J. Neurosci., 18, 6279±6289.
Milner, T.A., Joh, T.H. & Pickel, V.M. (1986) Tyrosine hydroxylase in the ratparabrachial region: ultrastructural localization and extrinsic sources ofimmunoreactivity. J. Neurosci., 6, 2585±2603.
Murabe, Y., Nishida, K. & Sano, Y. (1981) Cells capable of uptake ofhorseradish peroxidase in some circumventricular organs of the cat and rat.Cell Tissue Research, 219, 85±92.
Nakamura, K., Kaneko, T., Yamashita, Y., Katoh, H., Ichikawa, A. & Negishi,M. (1999) Immunocytochemical localization of prostaglandin EP3 receptorin the rat hypothalamus. Neurosci. Lett., 260, 117±120.
Niijima, A. (1996) The afferent discharges from sensors for interleukin 1 betain the hepatoportal system in the anesthetized rat. J. Auton. Nerv. Syst., 61,287±291.
Opp, M.R. & Toth, L.A. (1998) Somnogenic and pyrogenic effects ofinterleukin-1beta and lipopolysaccharide in intact and vagotomized rats.Life Sci., 62, 923±936.
Pomerantz, K.B., Summers, B. & Hajjar, D.P. (1993) Eicosanoid metabolismin cholesterol-enriched arterial smooth muscle cells. Evidence for reducedposttranscriptional processing of cyclooxygenase I and reducedcyclooxygenase II gene expression. Biochemistry, 32, 13624±13635.
Riche, D., De Pommery, J. & Menetrey, D. (1990) Neuropeptides andcatecholamines in efferent projections of the nuclei of the solitary tract inthe rat. J. Comp. Neurol., 293, 399±424.
Romanovsky, A.A., Kulchitsky, V.A., Simons, C.T. & Sugimoto, N. (1998)Methodology of fever research: why are polyphasic fevers often thought tobe biphasic? Am. J. Physiol., 275, R332±R338.
Romanovsky, A.A., Simons, C.T., SzeÂkely, M. & Kulchitsky, V.A. (1997) Thevagus nerve in the thermoregulatory response to systemic in¯ammation.Am. J. Physiol., 273, R407±R413.
Rothwell, N.J., Busbridge, N.J., Lefeuvre, R.A., Hardwick, A.J., Gauldie, J. &
Distinct mechanisms mediate fever and sickness behaviour 4445
ã 2000 Federation of European Neuroscience Societies, European Journal of Neuroscience, 12, 4434±4446

Hopkins, S.J. (1991) Interleukin-6 is a centrally acting endogenous pyrogenin the rat. Can. J. Physiol. Pharmacol., 69, 1465±1469.
Rothwell, N.J. & Hopkins, S.J. (1995) Cytokines and the nervous system II:Actions and mechanisms of action. Trends Neurosci., 18, 130±136.
Sagar, S.M., Price, K.J., Kasting, N.W. & Sharp, F.R. (1995) Anatomicpatterns of Fos immunostaining in rat brain following systemic endotoxinadministration. Brain Res. Bull., 36, 381±392.
Saper, C.B., Reis, D.J. & Joh, T. (1983) Medullary catecholamine inputs to theanteroventral third ventricular cardiovascular regulatory region in the rat.Neurosci. Lett., 42, 285±291.
Sawchenko, P.E. & Swanson, L.W. (1982) The organization of noradrenergicpathways from the brainstem to the paraventricular and supraoptic nuclei inthe rat. Brain Res., 257, 275±325.
Sehic, E. & Blatteis, C.M. (1996) Blockade of lipopolysaccharide-inducedfever by subdiaphragmatic vagotomy in guinea pigs. Brain Res., 726, 160±166.
Sehic, E., Ungar, A.L. & Blatteis, C.M. (1996) Interaction betweennorepinephrine and prostaglandin E2 in the preoptic area of guinea pigs.Am. J. Physiol., 271, R528±R536.
Smith, G.P., Jerome, C., Cushin, B.J., Eterno, R. & Simansky, K.J. (1981)Abdominal vagotomy blocks the satiety effect of cholecystokinin in the rat.Science, 213, 1036±1037.
Spadaro, F. & Dunn, A.J. (1990) Intracerebroventricular administration ofinterleukin-1 to mice alters investigation of stimuli in a novel environment.Brain Behav. Immun., 4, 308±322.
StroÈmberg, H., Svensson, S.P.S. & Hermanson, O. (2000) Distribution of thetranscription factor Signal Transducer and Activator of Transcription 3 inthe rat central nervous system and dorsal root ganglia. Brain Res., 853, 105±114.
Sumal, K.K., Blessing, W.W., Joh, T.H., Reis, D.J. & Pickel, V.M. (1983)Synaptic interaction of vagal afferents and catecholaminergic neurons in therat nucleus tractus solitarius. Brain Res., 277, 31±40.
Tkacs, N. & Li, J. (1999) Immune stimulation induces Fos expression inbrainstem amygdala afferents. Brain Res. Bull., 48, 223±231.
Traub, R.J., Sengupta, J.N. & Gebhart, G.F. (1996) Differential c-fos
expression in the nucleus of the solitary tract and spinal cord followingnoxious gastric distension in the rat. Neuroscience, 74, 873±884.
Ushikubi, F., Segi, E., Sugimoto, Y., Murata, T., Matsuoka, T., Kobayashi, T.,Hizaki, H., Tuboi, K., Katsuyama, M., Ichikawa, A., Tanaka, T., Yoshida,N. & Narumiya, S. (1998) Impaired febrile response in mice lacking theprostaglandin E receptor subtype EP3. Nature, 395, 281±284.
VallieÁres, L., Lacroix, S. & Rivest, S. (1997) In¯uence of interleukin-6 onneural activity and transcription of the gene encoding corticotrophin-releasing factor in the rat brain: an effect depending upon the route ofadministration. Eur. J. Neurosci., 9, 1461±1472.
VallieÁres, L. & Rivest, S. (1997) Regulation of the genes encoding interleukin-6, its receptor, and gp130 in the rat brain in response to the immuneactivator lipopolysaccharide and the proin¯ammatory cytokine interleukin-1beta. J. Neurochem., 69, 1668±1683.
Van Dam, A.M., Bol, J.G., Gaykema, R.P., Goehler, L.E., Maier, S.F.,Watkins, L.R. & Tilders, F.J. (2000) Vagotomy does not inhibit high doselipopolysaccharide-induced interleukin-1beta immunoreactivity in rat brainand pituitary gland. Neurosci. Lett., 285, 169±172.
Wan, W., Wetmore, L., Sorensen, C.M., Greenberg, A.H. & Nance, D.M.(1994) Neural and biochemical mediators of endotoxin and stress-inducedc-fos expression in the rat brain. Brain Res. Bull., 34, 7±14.
Watkins, L.R., Goehler, L.E., Relton, J.K., Tartaglia, N., Silbert, L., Martin, D.& Maier, S.F. (1995) Blockade of interleukin-1 induced hyperthermia bysubdiaphragmatic vagotomy: evidence for vagal mediation of immune-braincommunication. Neurosci. Lett., 183, 27±31.
Yasui, M., Nakao, A., Yuuki, T., Harada, A., Nonami, T. & Takagi, H. (1995)Immunohistochemical detection of endotoxin in endotoxemic rats. Hepato-Gastroenterology, 42, 683±690.
Zhang, Y., Lu, J., Elmquist, J.K. & Saper, C.B. (2000) Lipopolysaccharideactivates speci®c populations of hypothalamic and brainstem neurons thatproject to the spinal cord. J. Neurosci., 20, 6578±6586.
Zhang, J. & Rivest, S. (1999) Distribution, regulation and colocalization of thegenes encoding the EP2- and EP4-PGE2 receptors in the rat brain andneuronal responses to systemic in¯ammation. Eur. J. Neurosci., 11, 2651±2668.
4446 J. P. Konsman et al.
ã 2000 Federation of European Neuroscience Societies, European Journal of Neuroscience, 12, 4434±4446




















