
The β Subunit Gate Loop Is Required for RNA Polymerase Modification by RfaH and NusG
20
The β subunit gate loop is required for RNA polymerase modification by RfaH and NusG Anastasia Sevostyanova 1 , Georgiy A. Belogurov 1,4 , Rachel A. Mooney 2 , Robert Landick 2,3 , and Irina Artsimovitch 1,* 1 Department of Microbiology and The RNA Group, The Ohio State University, Columbus, OH 43210, USA 2 Department of Biochemistry, University of Wisconsin, Madison, WI 53706, USA 3 Department of Bacteriology, University of Wisconsin, Madison, WI 53706, USA SUMMARY In all organisms, RNA polymerase (RNAP) relies on accessory factors to complete synthesis of long RNAs. These factors increase RNAP processivity by reducing pausing and termination, but their molecular mechanisms remain incompletely understood. We identify the β gate loop as an RNAP element required for antipausing activity of a bacterial virulence factor RfaH, a member of the universally conserved NusG family. Interactions with the gate loop are necessary for suppression of pausing and termination by RfaH, but are dispensable for RfaH binding to RNAP mediated by the β′ clamp helices. We hypothesize that, upon binding to the clamp helices and the gate loop, RfaH bridges the gap across the DNA channel, stabilizing RNAP/nucleic acid contacts and disfavoring isomerization into a paused state. We show that contacts with the gate loop are also required for antipausing by NusG and propose that most NusG homologs use similar mechanisms to increase RNAP processivity. INTRODUCTION The structure, effects on RNA synthesis, and the major binding site on the RNAP of proteins from the NusG family are universally conserved (Belogurov et al., 2009; Hirtreiter et al., 2010; Mooney et al., 2009b). These proteins are also involved in crosstalk between transcription and other key cellular processes, such as RNA processing and translation (Bies-Etheve et al., 2009; Burmann et al., 2010; Peterlin and Price, 2006; Schneider et al., 2006; Wang and Dennis, 2009). The E. coli NusG and RfaH are the best studied members of this family. NusG is a general transcription factor that is essential in the wild-type E. coli and is bound to nearly all transcribed genes (Mooney et al., 2009a). RfaH, a sequence- specific paralog of NusG, preferentially increases distal expression in operons containing promoter-proximal ops DNA elements. The ops sequence mediates RfaH binding to elongating RNAP and thus restricts RfaH action to just a few E. coli operons (Belogurov et al., 2009); consistently, rfaH is dispensable for cell viability. The N-terminal domains of RfaH and NusG are structurally homologous, are predicted to make similar contacts with © 2011 Elsevier Inc. All rights reserved. * To whom correspondence should be addressed: [email protected]. 4 Present address: Department of Biochemistry and Food Chemistry, University of Turku, Turku FIN-20014, Finland. Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain. NIH Public Access Author Manuscript Mol Cell. Author manuscript; available in PMC 2012 July 22. Published in final edited form as: Mol Cell. 2011 July 22; 43(2): 253–262. doi:10.1016/j.molcel.2011.05.026. NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
Transcript of The β Subunit Gate Loop Is Required for RNA Polymerase Modification by RfaH and NusG

The β subunit gate loop is required for RNA polymerasemodification by RfaH and NusG
Anastasia Sevostyanova1, Georgiy A. Belogurov1,4, Rachel A. Mooney2, Robert Landick2,3,and Irina Artsimovitch1,*
1 Department of Microbiology and The RNA Group, The Ohio State University, Columbus, OH43210, USA2 Department of Biochemistry, University of Wisconsin, Madison, WI 53706, USA3 Department of Bacteriology, University of Wisconsin, Madison, WI 53706, USA
SUMMARYIn all organisms, RNA polymerase (RNAP) relies on accessory factors to complete synthesis oflong RNAs. These factors increase RNAP processivity by reducing pausing and termination, buttheir molecular mechanisms remain incompletely understood. We identify the β gate loop as anRNAP element required for antipausing activity of a bacterial virulence factor RfaH, a member ofthe universally conserved NusG family. Interactions with the gate loop are necessary forsuppression of pausing and termination by RfaH, but are dispensable for RfaH binding to RNAPmediated by the β′ clamp helices. We hypothesize that, upon binding to the clamp helices and thegate loop, RfaH bridges the gap across the DNA channel, stabilizing RNAP/nucleic acid contactsand disfavoring isomerization into a paused state. We show that contacts with the gate loop arealso required for antipausing by NusG and propose that most NusG homologs use similarmechanisms to increase RNAP processivity.
INTRODUCTIONThe structure, effects on RNA synthesis, and the major binding site on the RNAP of proteinsfrom the NusG family are universally conserved (Belogurov et al., 2009; Hirtreiter et al.,2010; Mooney et al., 2009b). These proteins are also involved in crosstalk betweentranscription and other key cellular processes, such as RNA processing and translation(Bies-Etheve et al., 2009; Burmann et al., 2010; Peterlin and Price, 2006; Schneider et al.,2006; Wang and Dennis, 2009). The E. coli NusG and RfaH are the best studied members ofthis family. NusG is a general transcription factor that is essential in the wild-type E. coliand is bound to nearly all transcribed genes (Mooney et al., 2009a). RfaH, a sequence-specific paralog of NusG, preferentially increases distal expression in operons containingpromoter-proximal ops DNA elements. The ops sequence mediates RfaH binding toelongating RNAP and thus restricts RfaH action to just a few E. coli operons (Belogurov etal., 2009); consistently, rfaH is dispensable for cell viability. The N-terminal domains ofRfaH and NusG are structurally homologous, are predicted to make similar contacts with
© 2011 Elsevier Inc. All rights reserved.*To whom correspondence should be addressed: [email protected] address: Department of Biochemistry and Food Chemistry, University of Turku, Turku FIN-20014, Finland.Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to ourcustomers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review ofthe resulting proof before it is published in its final citable form. Please note that during the production process errors may bediscovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
NIH Public AccessAuthor ManuscriptMol Cell. Author manuscript; available in PMC 2012 July 22.
Published in final edited form as:Mol Cell. 2011 July 22; 43(2): 253–262. doi:10.1016/j.molcel.2011.05.026.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

RNAP, and are sufficient in vitro for the reduction of transcriptional pausing (Belogurov etal., 2009; Mooney et al., 2009b). However, full-length RfaH and NusG have opposite effectson the expression of poorly translated genes (e.g., foreign DNA) in vivo. NusG acts togetherwith ρ to terminate transcription (Cardinale et al., 2008; Mooney et al., 2009b), whereasRfaH reduces termination via its antipausing (AP) activity and exclusion of NusG(Belogurov et al., 2009; Carter et al., 2004).
RfaH and other antiterminators, such as the phage λN and Q proteins (Roberts et al., 2008),appear to maintain the elongating RNAP complex (EC) in a rapidly-moving mode bypreventing its isomerization into an off-pathway state called an elemental pause (Ederth etal., 2006). From this state, the paused EC is thought to isomerize into long-lived pausedstates (e.g., upon backtracking or formation of a nascent RNA hairpin) or to enter atermination pathway (Artsimovitch and Landick, 2000). Formation of the elemental pauselikely involves fraying of the 3′ RNA nucleotide and rearrangement of an active-site elementcalled the trigger loop, and occurs after nucleotide addition but before RNAP translocation(Sydow et al., 2009; Toulokhonov et al., 2007). Thus, a regulator could suppress pausingeither by promoting translocation or by preventing active-site conformational changes, butthe molecular mechanism of an AP modification remains unknown for any protein.
RfaH is an excellent model to address the question of how a regulator can modify RNAP toresist pause signals. RfaH action can be readily monitored in the cell because it isdispensable and controls just a few operons. In addition, RfaH structure, mechanism ofrecruitment, DNA- and RNAP-binding regions, and binding site on the RNAP have beendetermined. RfaH binds to the β′ clamp helices domain (β′CH) and the non-template (NT)DNA strand at the upstream end of the transcription bubble (Belogurov et al., 2007). Unlikethe phage antiterminators that also stabilize the EC against dissociation (Rees et al., 1997;Yarnell and Roberts, 1999), RfaH does not affect the EC stability or intrinsic termination,except at one unusual signal (Artsimovitch and Landick, 2002; Carter et al., 2004), allowingdirect testing of its AP mechanism. In the simplest scenario, RfaH may favor reannealing ofthe DNA strands and thus push the RNAP forward; indeed, we showed that RfaH favorsforward translocation of the enzyme (Svetlov et al., 2007). However, the location of theRfaH binding site on the EC suggests that the bound RfaH may additionally restrictmovements of the β′ clamp that have been proposed to accompany pausing (Landick, 2001).Here we present the evidence in support of the second mechanism. We show that RfaHaction depends on two RNAP elements, the β′CH and the β subunit gate loop (βGL), whichare located directly across each other on the two sides on the DNA-binding channel. Wepropose that β′CH, βGL, and RfaH jointly form a clamp on the DNA that allows the RNAPto resist pausing and termination signals.
RESULTSRfaH’s HTT motif is required for antipausing but not binding to the EC
To identify the determinants required for AP by RfaH, we have previously carried outmutational analysis of the N-terminal domain, RfaHN, which is sufficient for all in vitroactivities of RfaH. This analysis (Belogurov et al., 2010) revealed three functional regions(Figure 1A): (i) a cluster of residues that likely interact with the NT DNA, (ii) a hydrophobicsurface that is predicted to make van der Waals contacts with the β′CH, and (iii) a cluster ofthree residues (HTT) distant from the NT DNA and the β′CH. Substitutions in the first tworegions are expected to compromise RfaH interactions with the EC; indeed, the defects ofthese substitutions are suppressed at high RfaH concentrations. In contrast, RfaH variantswith substitutions of the HTT residues display dramatically reduced AP activity even whenpresent at high concentrations (Belogurov et al., 2010). These observations are inconsistentwith a model in which RfaH binding to the DNA and the β′CH is sufficient for its AP
Sevostyanova et al. Page 2
Mol Cell. Author manuscript; available in PMC 2012 July 22.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

activity and suggest that an additional interaction between the HTT motif and the EC isrequired. The goal of this study was to characterize this interaction.
First, we wanted to exclude a possibility that, even though the HTT residues are not modeledto interact with either the DNA or the β′CH (Figure 1B), their substitutions compromiseRfaH binding to the EC indirectly, and an altered protein dissociates after recruitment. Toascertain that RfaH variants with HTT residues substituted for Ala remain bound to the EC,we used a σ competition assay that relies on the RfaH ability to sterically block σrecruitment to the EC because, as we showed earlier, both proteins bind to the β′CH domain(Sevostyanova et al., 2008).
To test the ability of altered RfaH variants to compete with σ during elongation, we carriedout a standard single-round transcription assay (Figure 1C) using a linear DNA templatewith a strong T7A1 promoter followed by an initial transcribed region that allows forformation of radiolabeled ECs stalled after incorporation of a G residue at position 37 (G37)when transcription is initiated in the absence of UTP. The template also encodes the opspause signal (opsP), which induces RNAP backtracking and mediates RfaH recruitment tothe EC, the consensus -10 element, and a hairpin-stabilized his pause signal (hisP). Uponaddition of all NTPs, rifapentin (to block re-initiation), and the excess of free σ (1 μM vs 30nM EC), RNAP elongated the nascent RNA, transiently pausing at the opsP (U43) and thehisP sites; σ contacts to bases in a -10-like sequence induced a very long pause at the σP sitefrom which RNAP did not escape under the conditions of our experiments (Figure 1C). Thewild-type (WT) RfaH, when present, delayed RNAP escape from the C45 position (throughinteractions with the ops bases) and accelerated transcription at other sites as evidenced by afaster RNAP arrival at the end of the template (RO). However, on this template, AP effectsof RfaH are difficult to quantify because the hisP signal, which has been shown to respondto RfaH (Artsimovitch and Landick, 2002), is separated from the halted complex position by~100 nt encoding two additional strong and many weak pause sites, which desynchronizethe RNAP population. We have reported the effects of all RfaH variants on pausing at hisPon a less complex template (Belogurov et al., 2010).
Most importantly, the WT RfaH strongly inhibited pausing at the σP site even at a 15-foldmolar excess of σ. We found that H65A and T67A (and T66A, not shown) variants alsocompeted with σ efficiently, indicating that the inability of these variants to accelerateelongation is not due to their failure to remain bound to the EC after recruitment. As acontrol, we used RfaH Y54F variant with a substitution at the RfaH/β′CH interface (Figure1B); this substitution confers a dramatic AP defect that can be rescued at 3 μM RfaH(Belogurov et al., 2010). Y54F was initially recruited at the ops site (as reflected by reducedpausing at U43) but apparently failed to maintain stable post-recruitment interactions: itneither accelerated elongation nor competed with σ (Figure 1C). These results suggest thatthe HTT residues are largely dispensable for RfaH binding to the EC but may be involved ina functional interaction required for the AP modification.
The HTT motif may interact with the β subunit GLThe RfaH/EC model (Belogurov et al., 2007) positions the HTT motif near the βGL (Figure1B). To evaluate a possible role of the E. coli βGL in RfaH function, we substituted the βresidues 368–376 with two glycines. Surprisingly, although the βGL has been proposed toplay a key role in DNA loading during initiation (Vassylyev et al., 2002), we found that theΔβGL variant was active in vitro (Figure 2) and under certain conditions in vivo (Figure S1).
We first tested whether the deletion of the βGL would abolish the RNAP response to RfaHin vitro at the hisP signal. In these experiments, we used the isolated RfaHN which does notrequire an ops element for recruitment (Belogurov et al., 2007), thereby simplifying the
Sevostyanova et al. Page 3
Mol Cell. Author manuscript; available in PMC 2012 July 22.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

kinetic analysis. In a single-round pause assay, RfaHN accelerated RNAP escape from hisPalmost four-fold, reducing the pause half-life from 42 to 11 seconds, but had almost noeffect on the ΔβGL enzyme (Figure 2A). This result shows that the βGL is necessary for theAP activity of RfaH, at least at one model pause site.
To test whether the βGL deletion confers a general AP defect (as opposed to an effectspecific to the his pause), we compared elongation of an rpoB mRNA, which is devoid ofstrong regulatory pauses but punctuated by many elemental pauses that contribute at least65% to the overall rate of elongation (Neuman et al., 2003), by the deletion and the WTenzymes. RfaHN increased the apparent elongation rate by the WT RNAP by 1.8-fold, butslowed down the deletion enzyme ~1.4-fold (Figure 2B); RfaHN has a similar inhibitoryeffect on the WT enzyme at higher NTPs (Svetlov et al., 2007).
One explanation for this phenotype could be that the βGL deletion renders RNAP pause-resistant. We reported previously that RfaH does not accelerate transcription at saturatingsubstrate concentrations or by “fast” RNAP variants with substitutions in β and β′ subunitsthat render the enzyme insensitive to pause and termination signals (Svetlov et al., 2007). Inparticular, an enzyme that is missing a large sequence insertion in the β′ trigger loop, Δβ′ SI3RNAP, fails to respond to RfaH. However, the ΔβGL RNAP was only moderately fasterthan the WT RNAP and displayed a dramatically different response to the hisP signal in thepresence and in the absence of RfaH in a side-by-side comparison with the Δβ′ SI3 enzyme(Figure 2A).
If the βGL deletion removes a contact with the HTT motif, substitutions of the HTTresidues, which abolish RfaH effects on transcription by the WT RNAP, should not haveany additional effects on the ΔβGL enzyme. Indeed, we found that the in vitro transcriptionpattern of the ΔβGL enzyme was the same with the WT, H65A, T66A, and T67A RfaHvariants (Figure S2). Together, these results demonstrate that the βGL is required in vitro forthe AP effects of RfaH and are consistent with a functional βGL/HTT interaction.
The βGL is not required for RfaH binding to the ECTo ascertain that the βGL deletion does not eliminate RfaH binding to the EC, we used σcompetition and electrophoretic mobility shift assays (Figure 3). We found that RfaHN,which does not dissociate from the EC in vitro and thus mimics an in vivo pattern(Belogurov et al., 2007), eliminated σ recruitment to both the WT and the ΔβGL ECs(Figure 3A). As expected, RfaHN drastically altered the pattern of transcription by the WTRNAP, delaying escape from the ops site and reducing pausing at the downstream signals,such as hisP. In contrast, RfaHN had no effect on the ΔβGL RNAP, except at the σP sitewhere it abolished pausing as efficiently as with the WT RNAP. The full-length RfaH hadsimilar effects (AS, data not shown).
We next visualized RfaH binding to the ops-paused ECs by a mobility shift assay (Figure3B). We assembled ECs on nucleic acid scaffolds composed of the template and NT DNAstrands and the RNA (see Experimental Procedures); we used this assay previously toidentify the RfaH target on the EC (Artsimovitch and Landick, 2002). To verify that theassembled complexes were active, we incubated unlabeled ECs with radiolabeledα[32P]GTP, the incoming nucleotide specified by the template DNA. Upon α[32P]GMPincorporation, the position of the ECs could be readily visualized in an agarose gel (Figure3B, left). To monitor RfaH binding to the EC, we radiolabeled RfaH via a kinase recognitionsequence at the N-terminus of the protein. RfaH did not enter the gel when present alone butco-migrated with ECs assembled with the WT and ΔβGL RNAPs (Figure 3B, right). Bycontrast, a β′ I290R substitution at the tip of the β′CH completely abolished RfaH binding
Sevostyanova et al. Page 4
Mol Cell. Author manuscript; available in PMC 2012 July 22.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

(but not GMP incorporation), consistent with a model in which the β′CH is the majoraffinity determinant for RfaH.
Together, these results argue that the βGL is dispensable for RfaH binding. Consistently, wedetected only a very weak (but apparently specific) interaction between RfaHN and the β N-terminal domain in a bacterial two-hybrid assay (Figure S3).
RfaH dramatically reduces ρ-mediated polarity in vivoMany RfaH-controlled genes are horizontally transferred (Belogurov et al., 2009) and arethus expected to be subject to polarity control by ρ (Cardinale et al., 2008). RfaH has beenreported to inhibit the action of ρ in vitro (Belogurov et al., 2009) and in vivo (Stevens et al.,1997). We examined the effect of RfaH on the extent of termination (i.e., polarity) across the11-gene rfb operon (rfbBDACX-glf-rfc-wbbIJKL; Figure 4A). Using ChIP-chip analysis, wehave shown that after recruitment at a promoter-proximal ops site (located 76 bp upstreamfrom the first ORF, rfbB) RfaH remains bound throughout the entire rfb operon and excludesNusG from this and other ops-containing operons (Belogurov et al., 2009). At the sametime, we detected significant association of ρ with the distal part of rfb operon (Mooney etal., 2009a), consistent with a notion that NusG is required for efficient RNA release by ρ butnot for ρ recruitment.
To test the effects of RfaH and ρ on the rfb expression, we compared expression levels ofthe first (rfbB; shown in red in Figure 4) and the eighth (wbbI; blue) genes by qRT-PCR asdescribed in Experimental Procedures. We observed strong polarity (defined here as theratio of rfbB and wbbI messages) in the ΔrfaH strain where wbbI mRNA was barelydetectable (rfbB/wbbI ~800; Figure 4B right). Consistent with our expectations, essentiallyall polarity was caused by ρ, as polarity was eliminated upon the addition of the ρ inhibitorbicyclomycin (BCM; shaded bars in Figure 4B). In the absence of RfaH, addition of BCMincreased the absolute level of wbbI RNA more than 700-fold. In the presence of RfaH,~45% of RNAP molecules reached wbbI gene (Figure 4B; left); this fraction increased to78% when ρ was inhibited by BCM. These results show that even though RfaH does notcompletely abolish ρ-dependent termination (e.g., if not all ECs became modified by RfaH),it eliminates most of the ρ-mediated polarity within rfb operon.
The dramatic effect of RfaH on ρ termination is mediated by two mechanisms. First, RfaHmodifies RNAP into a pause-resistant state (Svetlov et al., 2007), thus making it lesssusceptible to ρ. Second, RfaH excludes the ρ-stimulatory factor NusG through competitionfor binding to the β′ CH. Thus, the levels of NusG associated with the rfb operon are verylow (Figure 4A), despite the fact that NusG concentration in the cell far exceeds that ofRfaH (Belogurov et al., 2009). In the absence of RfaH, RNAP will be terminated morefrequently as a consequence of both (i) increased pausing and (ii) increased binding of NusGwith consequent stimulation of ρ. However, the relative contributions of these twomechanisms to the RfaH action in vivo are not known.
Deletion of the βGL compromises RfaH function in vivoTo assess the contribution of AP to the anti-polar effect of RfaH in vivo, we used the RNAPlacking the βGL. Removal of the GL eliminated AP activity (Figure 2) but not RfaH bindingto the β′CH (Figure 3). Thus, RfaH would be expected to efficiently exclude NusG from thedeletion enzyme and any defect in polarity suppression could be attributed to the loss of AP.We could not construct a chromosomal deletion of the βGL; repeated attempts yielded onlygene duplications instead of a clean deletion allele, suggesting that the βGL is essential forcell growth or viability. Instead, to test the effects of the βGL RNAP, we constructedplasmids that encoded the WT or ΔβGL variant under control of an IPTG-inducible Ptrc
Sevostyanova et al. Page 5
Mol Cell. Author manuscript; available in PMC 2012 July 22.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

promoter. The plasmid copy of rpoB also contained a D516V substitution that confersresistance to rifamycins but does not have any other discernible effects on transcription. Theplasmid-encoded β was incorporated into the RNAP and supported growth in the presence ofrifapentin on plates but not in liquid media (Figure S1). We transformed Ptrc-rpoB plasmidsinto the rfaH+ and ΔrfaH strains, induced the expression of a plasmid-encoded β by IPTG,and added rifapentin to inhibit the chromosomally-encoded WT RNAP. Polarity in rfboperon was analyzed by qRT-PCR as above.
With the plasmid-encoded WT rpoB, we observed essentially the same trend as with thechromosomally-encoded RNAP (Figure 4B); however, the absolute numbers of mRNAsdiffered somewhat (Figure S4). The distal wbbI transcript was present at a ~20% the level ofrfbB in the presence of RfaH and 0.54% in the absence of RfaH, meaning that RfaHdecreased polarity (i.e., increased wbbI mRNA relative to rfbB mRNA) by a factor of ~37(Figure 5A). Deletion of the βGL reduced the effect of RfaH to ~5.5-fold.
Our results show that both RfaH and the βGL affect polarity in the rfb operon (Figure 5B).When both RfaH and the βGL are present, RfaH modifies RNAP and excludes NusG; thisgreatly reduces polarity (5 fold, lane 1). In the absence of RfaH, RNAP both is prone topausing and readily recruits NusG; this causes very strong polarity (185 fold, lane 2).Deletion of the βGL in the rfaH+ background leads to a two-fold increase in polarity(compare lanes 1 and 3), presumably due to the loss of RfaH AP activity in the absence ofthe βGL. However, the βGL role is not limited to its interactions with RfaH: removal of theGL decreased polarity in the rfaH− strain three fold (compare lanes 2 and 4). This defectcould be due to altered kinetic properties of the deletion enzyme or less efficient ρ-mediated,NusG-assisted RNA release. The additional “masking” effect of the βGL prevents preciseevaluation of the contribution of AP to the overall (37-fold) effect of RfaH on polarity: itmay range from a factor of two (if the anti-polar effect of ΔGL is attributed exclusively toaltered interactions with ρ and NusG) to a factor of six (if a somewhat faster elongation rateof the ΔβGL RNAP observed in vitro (Figure 2) accounts for all of its anti-polar effects invivo).
Contacts with the βGL are required for AP activity of E. coli NusGOne would expect that control of processivity is similar among multi-subunit RNAPs whichshare a common structural organization. Indeed, RfaH belongs to the NusG family, the onlygroup of universally conserved transcription elongation factors (Spt5 in Archaea and yeast;DSIF in humans). Recent cryoelectron microscopy (Klein et al., 2011) and X-ray (Martinez-Rucobo et al., 2011) structures of archaeal RNAP bound to Spt4/5 complex are fullyconsistent with our model of RfaH bound to the bacterial EC (Belogurov et al., 2007) andbiochemical analysis of T. thermophilus NusG (Sevostyanova and Artsimovitch, 2010):Spt4/5 binds to the same site on the EC and encircles the DNA, suggesting a similarmechanism of action (Svetlov and Nudler, 2011). We wanted to test whether the inferredcontact with the βGL plays a functional role in AP by NusG. The low resolution of theNusG/EC model and the lack of data on NusG/NT DNA interactions make it difficult topinpoint the residues that could mediate this contact. Therefore, we constructed an E. coliNusG variant in which four residues (79–82) that are structurally homologous to the RfaHHTTT motif (Belogurov et al., 2010) were replaced by Ala.
In contrast to the WT NusG, which reduced RNAP pausing at the scrambled ops signal andincreased an apparent rate of elongation (Figure 6A), the NusG-4A protein did not affecttranscription in vitro. Consistently, deletion of the βGL reduced NusG effect on pausing(Figure S5). However, neither the βGL deletion nor the 4A substitution altered the ability ofNusG to enhance ρ-dependent termination (Figure 6B).
Sevostyanova et al. Page 6
Mol Cell. Author manuscript; available in PMC 2012 July 22.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

These results reveal that interactions with the βGL are essential for AP by NusG but aredispensable for ρ-dependent termination in vitro. However, our observations that the βGLdeletion reduces polarity (Figure 5) suggest that these interactions may play a role in vivo.The loss of putative NusG/GL interactions may facilitate NusG dissociation from the EC orenable NusG to interact more readily with the ribosomal protein S10 (Burmann et al., 2010),thereby recruiting the ribosome to the rfb transcript. Either effect will decrease ρ-dependenttermination. At present, we cannot distinguish between these (and other) scenarios, but apotential role in function of the essential NusG could explain the failure of βGL RNAP tosupport cell viability (Figure S1).
DISCUSSIONTaken together, our results suggest that RfaH increases RNAP processivity by at least twomechanisms. First, RfaH binds to the β′CH and excludes NusG from the transcribing RNAP,thereby inhibiting ρdependent termination. Second, RfaH makes bridging contacts to theclamp domain and the β lobe domain that may restrict movements of the clamp and otherRNAP modules, in turn inhibiting pausing. We propose that the second mechanism is sharedby other proteins from the NusG family, which have been shown to regulate transcription inall living organisms.
RfaH makes a functional contact to the βGLHere, we present evidence that the βGL element, a part of the β lobe, is required for RfaHfunction in vivo and in vitro. The βGL element has been revealed by structural analysis(Vassylyev et al., 2002), and its position in the RNAP holoenzyme suggested a role in“gating” the entry of the promoter DNA during initiation. The role of the βGL intranscription has not been, to our knowledge, studied. We became interested in this elementwhen our molecular modeling suggested that the βGL is positioned next to the newlyidentified HTT cluster in RfaH (Figure 1); substitutions of the HTT residues reduced APactivity but were not predicted to interact with either the NT DNA or the β′CH, the twoknown RfaH targets in the EC (Belogurov et al., 2010). We found that the deletion of theβGL did not confer strong defects in initiation, elongation, termination and RNAP responseto accessory transcription factors in vitro (AS and IA, manuscript in preparation). However,the ΔβGL RNAP failed to respond to AP by RfaH and NusG (Figures 2 and 6A).
Using a two hybrid analysis, we detected an interaction between RfaHN and the N-terminaldomain of β (Figure S3). This interaction was disrupted by substitutions in the GL and theHTT motif, and thus is likely specific. However, it was at least ten-fold weaker than theinteraction between NusGN with a fragment of β′ that includes the β′CH (Nickels, 2009),and we have shown that NusG can t compete with RfaH during elongation (Belogurov et al.,2009). This suggests that the HTT contacts to the βGL do not make a significantcontribution to RfaH s affinity for RNAP. Indeed, we show that the βGL is not required forσ exclusion (Figure 3A) and RfaH binding to the static ECs (Figure 3B). Our data argue thatthe βGL is required for the AP modification of RNAP.
Operon polarity control by RfaH and NusGIn bacteria, if translation is inefficient or interrupted at a stop codon, ρ gains access to thenascent RNA to rapidly terminate transcription (Richardson et al., 1975). ρ establishespolarity along operons: since not all RNAPs reach the end of an operon, promoter-proximalgenes are transcribed at higher levels than the promoter-distal genes. Polarity ischaracteristic for all bacterial operons but is particularly pronounced in poorly translated,e.g., horizontally-transferred operons.
Sevostyanova et al. Page 7
Mol Cell. Author manuscript; available in PMC 2012 July 22.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

E. coli NusG plays opposite roles in polarity control. The N-terminal domain of NusGanchors the factor to the RNAP whereas the C-terminal domain makes contacts to ρ, therebyenhancing ρdependent termination (Mooney et al., 2009b), or to the ribosomal protein S10(Burmann et al., 2010), thereby ensuring that translation and transcription are coupled(Proshkin et al., 2010) and ρ is excluded. NusG also participates in the formation of a largenucleoprotein rrn antitermination complex that substitutes for the ribosome in shielding thenon-coding rRNA transcripts from ρ (Squires et al., 1993).
Enhancement of ρ-mediated RNA release is hypothesized to constitute the essential functionof NusG in E. coli, to limit the expression of foreign DNA (Cardinale et al., 2008).However, some horizontally-transferred genes that play important cellular roles have to beprotected from the defensive action of ρ and NusG. The rfb operon, which encodeslipopolysaccharide biosynthesis and transport genes, lacks a recognizable Shine-Dalgarnoelement in front of the rfbB ORF and has a suboptimal codon distribution, and is thusexpected to be a target for ρ.
Here we show that RfaH, a NusG paralog which has lost the ability to bind ρ, antagonizesthe action of ρ within the rfb operon (Figure 5). We propose that RfaH reduces polarity byseveral independent mechanisms. First, RfaH modifies RNAP into a pause-resistant state(Svetlov et al., 2007), thereby inhibiting ρ that preferentially targets paused RNAPs (Morganet al., 1984). Second, RfaH sterically excludes NusG from the EC (Belogurov et al., 2009).Our results suggest that AP activity suppresses polarity in vivo at least two-fold, in additionto a larger RfaH effect attributed to direct competition with NusG (Figure 5B). Finally,RfaH may promote translation of rfb mRNA. We argued that the stable in vivo associationof RfaH with the EC observed by ChIP-chip (Belogurov et al., 2009) requires sequestrationof the C-terminal domain, likely by the ribosome. Such an interaction has been detecteddirectly (Bailey et al., 2000) but its regulatory significance remains to be determined.
A structural model for AP by RfaHIn this work, we show that RfaH requires contacts to the β′ clamp and the β lobe domains tomediate AP modification of RNAP. We propose that these bridging contacts restrictmobility of RNAP modules, especially of the clamp, which disfavors pausing and, in turn,termination.
The clamp domain has been observed in different positions in various crystal structures(Figure 7) and proposed to undergo conformational changes throughout the transcriptioncycle (Davis et al., 2007; Epshtein et al., 2010; Landick, 2001). The clamp may open topermit entry of promoter DNA during initiation, close to establish the tight grip on DNAduring elongation, and then open again to allow release of DNA during termination (Tagamiet al., 2010). Formation of the elemental pause, a likely target for RfaH, is accompanied bystructural rearrangements near the active site that may include opening of the clamp (Sydowet al., 2009; Toulokhonov et al., 2007). RfaH interactions with the β′CH suggested that RfaHmay modulate RNAP by controlling clamp movements (Svetlov et al., 2007). However, itremained unclear how binding of RfaH (18 kDa) would hinder movements of the largeclamp domain, e.g., as compared to a molecule that binds to the hinge around which theclamp rotates (Mukhopadhyay et al., 2008). Our current genetic data, in combination with invitro and in vivo analyses suggest a direct and intuitively simple mechanism, in which RfaHbinds simultaneously to the β′CH and βGL located on the opposite sides of the main channelto lock the clamp in a closed state (Figure 7).
These bridging contacts would allow RfaH to prevent structural rearrangements that occurduring transcriptional pausing. These rearrangements appear to involve movements of thetrigger loop/trigger helices (Toulokhonov et al., 2007; Zhang et al., 2010), which contact the
Sevostyanova et al. Page 8
Mol Cell. Author manuscript; available in PMC 2012 July 22.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

clamp as part of a three-helix bundle with the bridge helix, mediated in an anchor region byswitch 1. They also involve the β lobe, indirectly, through fork loop 2, one of three flexibleloops whose conformation is interconnected with the bridge helix and trigger loop/helices ina cap region (Hein and Landick, 2010; Seibold et al., 2010; Weinzierl, 2010). Bridgingcontacts of RfaH to the GL and clamp may stabilize the bridge helix-trigger loop/helicesmodule through the cap and anchor regions, respectively, and suppress entry to theelemental pause. Thus, the contribution of the βGL-RfaH contact to AP could be eitherdirect or indirect. Regardless, it is likely that RfaH binding to the clamp and βGL completesa proteinaceous encirclement of the DNA template, thereby increasing RNAP processivity.
Concluding remarksOur results demonstrate that the βGL is essential for AP modification of E. coli RNAP byRfaH and NusG. Together with our previous data, which identified the β′ CH as the bindingsite for RfaH and NusG, and recent structural and functional analyses of archaeal Spt5(Hirtreiter et al., 2010; Klein et al., 2011; Martinez-Rucobo et al., 2011), our observationssuggest a model in which NusG-like proteins form a part of the clamp around the DNA,increasing RNAP processivity. This mechanism is likely ancient and ubiquitous: in allkingdoms, RNAP must bypass numerous barriers, such as sequences that induce pausing,DNA-bound regulators, or DNA-packaging proteins (e.g., nucleoid-associated proteins inBacteria and nucleosomes in Eukarya), to get to the end of the transcribed region. It remainsto be determined whether the NusG-like proteins restrict the clamp and other movements intheir cognate RNAPs and how these effects are coupled to elongation, pausing, andtermination. It is also possible that other, structurally unrelated, proteins inhibit pausing andtermination by locking the two halves of RNAP together through binding to the same ordifferent determinants on RNAP.
EXPERIMENTAL PROCEDURESProteins and reagents
Oligonucleotides were obtained from Integrated DNA Technologies (Coralville, IA) orSigma Aldrich (St. Louis, MO), NTPs and [α32P]-NTPs were from Perkin Elmer (Boston,MA), restriction and modification enzymes – from NEB (Ipswich, MA), PCR reagents –from Roche (Indianapolis, IN), other chemicals - from Sigma (St. Louis, MO) and Fisher(Pittsburgh, PA). Plasmid DNAs and PCR products were purified using spin kits fromQiagen (Valencia, CA) and Promega (Madison, WI). Unless indicated otherwise, enzymesfor RNA manipulations and qRT-PCR were from Epicentre (Madison, WI). ρ was purifiedas described in (Mooney et al., 2009b). The full-length RfaH variants, the RfaHN domain,and RNAP were purified as described in (Belogurov et al., 2007). NusG was purified as in(Artsimovitch and Landick, 2000). Plasmids are listed in Table S1.
Pause assaysHalted ECs were prepared in 50 μl of TGA buffer (20 mM Tris-HCl, 2 mM MgCl2, 20 mMNaCl, 5% glycerol, and 0.1 mM EDTA, pH 7.9) with E. coli RNAP (30 nM), ApU (100μM), and starting NTPs (1 μM CTP, 5 μM ATP and GTP, 10 μCi [α32P]CTP, 3000 Ci/mmol). Transcription was restarted by the addition of CTP, ATP and UTP to 150 μM, GTPto 15 μM, and rifapentin to 25 μg/ml. Samples were removed at times indicated in figurelegends and quenched by the addition of an equal volume of STOP buffer (10 M urea, 50mM EDTA, 45 mM Tris-borate; pH 8.3, 0.1% bromophenol blue, 0.1% xylene cyanol).Samples were analyzed on denaturing urea-acrylamide gels. The RNA products werevisualized and quantified using PhosphorImager and ImageQuant Software (GE Healthcare).
Sevostyanova et al. Page 9
Mol Cell. Author manuscript; available in PMC 2012 July 22.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Sigma competition assayThe assay was performed on a linear DNA template (40nM) amplified from pIA807(Sevostyanova et al., 2008). Halted ECs were prepared in 50 μl of TGA buffer with E. coliRNAP (30 nM), ApU (100 μM), and starting NTPs (1 μM CTP, 5 μM ATP and GTP, 10μCi [α32P]CTP, 3000 Ci/mmol). Elongation factors (1 μM σ, 70 nM RfaH) were addedfollowed by a 3-min incubation at 37°C. Transcription was restarted by the addition of GTPto 15 μM, CTP, ATP and UTP to 150 μM, and rifapentin to 25 μg/ml.
Gel mobility shift assayTEC were reconstituted as described in (Artsimovitch and Landick, 2002). RfaH waslabeled with [α-32P]ATP using the heart muscle kinase catalytic subunit (NEB); theunincorporated label was removed using a size exclusion G50 spin column (GE Healthcare).Reconstituted ECs were mixed with radiolabeled RfaH at 50 nM, incubated for 5 min at37°C, and loaded onto 3% NuSieve agarose gels in 0.5X TBE. To obtain radiolabeled ECs,10 μCi [α-32P]GTP (3000 Ci/mmol) were added instead of RfaH. After electrophoresis atroom temperature at 5 V/cm for 4 hr, the gels were exposed to phosphorimager screens.
qRT-PCRTotal RNA was isolated from the WT or ΔrfaH stains grown under different conditions, andthe amounts of RNA message in rfbB region was compared to that in the wbbI region. Totest the effects of the βGL, overnight cultures of DH5α and IA149 (a ΔrfaH derivative ofDH5α) E. coli strains transformed with pIA898 (ΔGL; D516V rpoB) or pIA183 (D516VrpoB) were diluted 1/100 into LB and grown for 3 h before addition of 1 mM IPTG. After 2h of induction, rifapentin was added to 200 μg/ml, and cells were grown for 1 h. To test theeffects of ρ, overnight cultures were diluted 1/100 into LB, grown for 3 h, and treated withBCM (20 μg/ml) for additional 30 minutes. Total RNA samples were isolated using theNucleic Acid Isolation Kit (Epicentre). qRT-PCR analysis was performed usingMiniOpticon cycler (BioRad; Hercules, CA) and MasterAmp™ GREEN Real-Time RT-PCRkit. For each sample, at least three repeats in two independent experiments (starting fromcell growth and RNA isolation) were performed. To ensure an accurate quantification ofRNA message in rfbB and wbbI regions, qRT-PCR assay was calibrated using dilutions ofan in vitro synthesized RNA (64 pg, 16 pg, 4 pg, 1 pg, 0.25 pg and 0.0625 pg; seeSupplemental Methods for the list of oligonucleotides and additional details). Tc (thresholdcycle) from four independent runs were plotted against the concentration and fitted usingScientist 3.0 software (Micromath). The resulting equations were used to quantify theamount of rfbB and wbbI messages in total RNA samples.
Supplementary MaterialRefer to Web version on PubMed Central for supplementary material.
AcknowledgmentsWe thank Ruth M. Saecker and Dmitri Svetlov for comments on the manuscript. This work was supported by theNational Institutes of Health grants GM67153 (to IA) and GM38660 (to RL).
ReferencesArtsimovitch I, Landick R. Pausing by bacterial RNA polymerase is mediated by mechanistically
distinct classes of signals. Proc Natl Acad Sci U S A. 2000; 97:7090–7095. [PubMed: 10860976]Artsimovitch I, Landick R. The transcriptional regulator RfaH stimulates RNA chain synthesis after
recruitment to elongation complexes by the exposed nontemplate DNA strand. Cell. 2002; 109:193–203. [PubMed: 12007406]
Sevostyanova et al. Page 10
Mol Cell. Author manuscript; available in PMC 2012 July 22.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Bailey MJ, Hughes C, Koronakis V. In vitro recruitment of the RfaH regulatory protein into aspecialised transcription complex, directed by the nucleic acid ops element. Mol Gen Genet. 2000;262:1052–1059. [PubMed: 10660066]
Belogurov GA, Mooney RA, Svetlov V, Landick R, Artsimovitch I. Functional specialization oftranscription elongation factors. EMBO J. 2009; 28:112–122. [PubMed: 19096362]
Belogurov GA, Sevostyanova A, Svetlov V, Artsimovitch I. Functional regions of the N-terminaldomain of the antiterminator RfaH. Mol Microbiol. 2010; 76:286–301. [PubMed: 20132437]
Belogurov GA, Vassylyeva MN, Svetlov V, Klyuyev S, Grishin NV, Vassylyev DG, Artsimovitch I.Structural basis for converting a general transcription factor into an operon-specific virulenceregulator. Mol Cell. 2007; 26:117–129. [PubMed: 17434131]
Bies-Etheve N, Pontier D, Lahmy S, Picart C, Vega D, Cooke R, Lagrange T. RNA-directed DNAmethylation requires an AGO4-interacting member of the SPT5 elongation factor family. EMBORep. 2009; 10:649–654. [PubMed: 19343051]
Burmann BM, Schweimer K, Luo X, Wahl MC, Stitt BL, Gottesman ME, Rosch P. A NusE:NusGcomplex links transcription and translation. Science. 2010; 328:501–504. [PubMed: 20413501]
Cardinale CJ, Washburn RS, Tadigotla VR, Brown LM, Gottesman ME, Nudler E. Termination factorRho and its cofactors NusA and NusG silence foreign DNA in E. coli. Science. 2008; 320:935–938.[PubMed: 18487194]
Carter HD, Svetlov V, Artsimovitch I. Highly divergent RfaH orthologs from pathogenicproteobacteria can substitute for Escherichia coli RfaH both in vivo and in vitro. J Bacteriol. 2004;186:2829–2840. [PubMed: 15090525]
Davis CA, Bingman CA, Landick R, Record MT Jr, Saecker RM. Real-time footprinting of DNA inthe first kinetically significant intermediate in open complex formation by Escherichia coli RNApolymerase. Proc Natl Acad Sci U S A. 2007; 104:7833–7838. [PubMed: 17470797]
Ederth J, Mooney RA, Isaksson LA, Landick R. Functional interplay between the jaw domain ofbacterial RNA polymerase and allele-specific residues in the product RNA-binding pocket. J MolBiol. 2006; 356:1163–1179. [PubMed: 16405998]
Epshtein V, Dutta D, Wade J, Nudler E. An allosteric mechanism of Rho-dependent transcriptiontermination. Nature. 2010; 463:245–249. [PubMed: 20075920]
Hein PP, Landick R. The bridge helix coordinates movements of modules in RNA polymerase. BMCBiology. 2010; 8:144. [PubMed: 21144066]
Hirtreiter A, Damsma GE, Cheung AC, Klose D, Grohmann D, Vojnic E, Martin AC, Cramer P,Werner F. Spt4/5 stimulates transcription elongation through the RNA polymerase clamp coiled-coil motif. Nucleic Acids Res. 2010; 38:4040–4051. [PubMed: 20197319]
Klein BJ, Bose D, Baker KJ, Yusoff ZM, Zhang X, Murakami KS. RNA polymerase and transcriptionelongation factor Spt4/5 complex structure. Proc Natl Acad Sci U S A. 2011; 108:546–550.[PubMed: 21187417]
Landick R. RNA polymerase clamps down. Cell. 2001; 105:567–570. [PubMed: 11389826]Landick R, Wang D, Chan C. Quantitative analysis of transcriptional pausing by RNA polymerase: the
his leader pause site as a paradigm. Meth Enzymol. 1996; 274:334–352. [PubMed: 8902817]Martinez-Rucobo FW, Sainsbury S, Cheung AC, Cramer P. Architecture of the RNA polymerase-
Spt4/5 complex and basis of universal transcription processivity. EMBO J. 2011; 30:1302–1310.[PubMed: 21386817]
Mooney RA, Davis SE, Peters JM, Rowland JL, Ansari AZ, Landick R. Regulator trafficking onbacterial transcription units in vivo. Mol Cell. 2009a; 33:97–108. [PubMed: 19150431]
Mooney RA, Schweimer K, Rosch P, Gottesman M, Landick R. Two structurally independent domainsof E. coli NusG create regulatory plasticity via distinct interactions with RNA polymerase andregulators. J Mol Biol. 2009b; 391:341–358. [PubMed: 19500594]
Morgan WD, Bear DG, von Hippel PH. Specificity of release by Escherichia coli transcriptiontermination factor rho of nascent mRNA transcripts initiated at the lambda PR. J Biol Chem. 1984;259:8664–8671. [PubMed: 6330119]
Mukhopadhyay J, Das K, Ismail S, Koppstein D, Jang M, Hudson B, Sarafianos S, Tuske S, Patel J,Jansen R, et al. The RNA polymerase “switch region” is a target for inhibitors. Cell. 2008;135:295–307. [PubMed: 18957204]
Sevostyanova et al. Page 11
Mol Cell. Author manuscript; available in PMC 2012 July 22.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Neuman KC, Abbondanzieri EA, Landick R, Gelles J, Block SM. Ubiquitous transcriptional pausing isindependent of RNA polymerase backtracking. Cell. 2003; 115:437–447. [PubMed: 14622598]
Nickels BE. Genetic assays to define and characterize protein-protein interactions involved in generegulation. Methods. 2009; 47:53–62. [PubMed: 18952173]
Peterlin BM, Price DH. Controlling the elongation phase of transcription with P-TEFb. Mol Cell.2006; 23:297–305. [PubMed: 16885020]
Peters JM, Mooney RA, Kuan PF, Rowland JL, Keles S, Landick R. Rho directs widespreadtermination of intragenic and stable RNA transcription. Proc Natl Acad Sci U S A. 2009;106:15406–15411. [PubMed: 19706412]
Proshkin S, Rahmouni AR, Mironov A, Nudler E. Cooperation between translating ribosomes andRNA polymerase in transcription elongation. Science. 2010; 328:504–508. [PubMed: 20413502]
Rees WA, Weitzel SE, Das A, von Hippel PH. Regulation of the elongation-termination decision atintrinsic terminators by antitermination protein N of phage lambda. J Mol Biol. 1997; 273:797–813. [PubMed: 9367773]
Richardson JP, Grimley C, Lowery C. Transcription Termination Factor Rho Activity Is Altered inEscherichia coli with suA Gene Mutations. Proc Natl Acad Sci U S A. 1975; 72:1725–1728.[PubMed: 1098042]
Roberts JW, Shankar S, Filter JJ. RNA polymerase elongation factors. Annu Rev Microbiol. 2008;62:211–233. [PubMed: 18729732]
Schneider DA, French SL, Osheim YN, Bailey AO, Vu L, Dodd J, Yates JR, Beyer AL, Nomura M.RNA polymerase II elongation factors Spt4p and Spt5p play roles in transcription elongation byRNA polymerase I and rRNA processing. Proc Natl Acad Sci U S A. 2006; 103:12707–12712.[PubMed: 16908835]
Seibold SA, Singh BN, Zhang C, Kireeva M, Domecq C, Bouchard A, Nazione AM, Feig M, CukierRI, Coulombe B, et al. Conformational coupling, bridge helix dynamics and active sitedehydration in catalysis by RNA polymerase. Biochim Biophys Acta. 2010; 1799:575–587.[PubMed: 20478425]
Sevostyanova A, Artsimovitch I. Functional analysis of Thermus thermophilus transcription factorNusG. Nucleic Acids Res. 2010; 38:7432–45. [PubMed: 20639538]
Sevostyanova A, Svetlov V, Vassylyev DG, Artsimovitch I. The elongation factor RfaH and theinitiation factor sigma bind to the same site on the transcription elongation complex. Proc NatlAcad Sci U S A. 2008; 105:865–870. [PubMed: 18195372]
Squires CL, Greenblatt J, Li J, Condon C. Ribosomal RNA antitermination in vitro: requirement forNus factors and one or more unidentified cellular components. Proc Natl Acad Sci U S A. 1993;90:970–974. [PubMed: 8430111]
Stevens MP, Clarke BR, Roberts IS. Regulation of the Escherichia coli K5 capsule gene cluster bytranscription antitermination. Mol Microbiol. 1997; 24:1001–1012. [PubMed: 9220007]
Svetlov V, Belogurov GA, Shabrova E, Vassylyev DG, Artsimovitch I. Allosteric control of the RNApolymerase by the elongation factor RfaH. Nucleic Acids Res. 2007; 35:5694–5705. [PubMed:17711918]
Svetlov V, Nudler E. Clamping the clamp of RNA polymerase. Embo J. 2011; 30:1190–1191.[PubMed: 21468097]
Sydow JF, Brueckner F, Cheung AC, Damsma GE, Dengl S, Lehmann E, Vassylyev D, Cramer P.Structural basis of transcription: mismatch-specific fidelity mechanisms and paused RNApolymerase II with frayed RNA. Mol Cell. 2009; 34:710–721. [PubMed: 19560423]
Tagami S, Sekine S, Kumarevel T, Hino N, Murayama Y, Kamegamori S, Yamamoto M, Sakamoto K,Yokoyama S. Crystal structure of bacterial RNA polymerase bound with a transcription inhibitorprotein. Nature. 2010; 468:978–982. [PubMed: 21124318]
Toulokhonov I, Zhang J, Palangat M, Landick R. A central role of the RNA polymerase trigger loop inactive-site rearrangement during transcriptional pausing. Mol Cell. 2007; 27:406–419. [PubMed:17679091]
Vassylyev DG, Sekine S, Laptenko O, Lee J, Vassylyeva MN, Borukhov S, Yokoyama S. Crystalstructure of a bacterial RNA polymerase holoenzyme at 2.6 A resolution. Nature. 2002; 417:712–719. [PubMed: 12000971]
Sevostyanova et al. Page 12
Mol Cell. Author manuscript; available in PMC 2012 July 22.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Vassylyev DG, Vassylyeva MN, Zhang J, Palangat M, Artsimovitch I, Landick R. Structural basis forsubstrate loading in bacterial RNA polymerase. Nature. 2007; 448:163–168. [PubMed: 17581591]
Wang MB, Dennis ES. SPT5-like, a new component in plant RdDM. EMBO Rep. 2009; 10:573–575.[PubMed: 19444314]
Weinzierl RO. The nucleotide addition cycle of RNA polymerase is controlled by two molecularhinges in the Bridge Helix domain. BMC Biol. 2010; 8:134. [PubMed: 21034443]
Yarnell WS, Roberts JW. Mechanism of intrinsic transcription termination and antitermination.Science. 1999; 284:611–615. [PubMed: 10213678]
Zhang G, Campbell EA, Minakhin L, Richter C, Severinov K, Darst SA. Crystal structure of Thermusaquaticus core RNA polymerase at 3.3 A resolution. Cell. 1999; 98:811–824. [PubMed:10499798]
Zhang J, Palangat M, Landick R. Role of the RNA polymerase trigger loop in catalysis and pausing.Nat Struct Mol Biol. 2010; 17:99–104. [PubMed: 19966797]
Sevostyanova et al. Page 13
Mol Cell. Author manuscript; available in PMC 2012 July 22.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 1. RfaH interactions with the EC(A) AP-defective substitutions in RfaHN (Belogurov et al., 2010). The affected residues areshown as sticks and are colored according to the EC component with which these residuesare proposed to interact in panel B.(B) A model of RfaHN bound to the EC (Belogurov et al., 2007). The T. thermophilusRNAP is shown as tubes with the bridge helix (β′BH) highlighted in cyan, the templateDNA – in black, the NT DNA – in yellow, the nascent RNA – in red. Position of the RNAPactive site is marked by the catalytic Mg2+ ion (a yellow sphere). RfaHN (blue) is bound tothe NT strand and to β′CH (green); Tyr54 (forest green) interacts with the β′CH. The HTTcluster is positioned next to a mobile βGL (magenta). Figures in A and B were prepared withPyMol (DeLano Scientific).(C) The RfaH retention assay on the linear pIA807 template shown on top with thetranscription start site (+1) and end (224), the ops and the -10 elements, and the hisP signalindicated. Halted ECs were incubated with σ70 and the WT or an altered RfaH whereindicated. Single-round transcription was restarted upon addition of NTPs and stopped at 10,20, 40, 90, 180 and 360 sec. In this and other experiments, some halted ECs become arrestedand do not resume elongation. Positions of the G37, ops, σP, hisP and run-off (RO)transcripts are indicated with arrows. The fraction of RNA at the σP site after a 360-secincubation (as % of total RNA) is presented below each panel; the errors (±SD) werecalculated from four independent experiments.
Sevostyanova et al. Page 14
Mol Cell. Author manuscript; available in PMC 2012 July 22.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 2. The βGL deletion eliminates AP activity of RfaH(A) Single-round pause assays on a pIA171 template that encodes the hisP signal, with (50nM) or without RfaHN. The half-lives of the WT, ΔβGL and Δβ′SI3 RNAPs paused at thehisP site were determined as described in (Landick et al., 1996). Samples were taken at 7,15, 30, 45, 60, 80, 100, 120, 180, 300 and 540 sec.(B) Net elongation rates were determined on a “pause-free” pIA146 template as describedpreviously (Svetlov et al., 2007) at 20 μM NTPs. RfaHN (80 nM) was added whereindicated. Samples were taken at 2, 3, 5, 7, 10, 20 and 40 min. In panels A and B, the errors(±SD) were calculated from four independent experiments.
Sevostyanova et al. Page 15
Mol Cell. Author manuscript; available in PMC 2012 July 22.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 3. RfaH binds to the ΔβGL RNAP(A) The σ competition assay was performed as in Figure 1C.(B) The gel mobility shift assay was performed as in (Artsimovitch and Landick, 2002). ECsassembled from DNA and RNA oligonucleotides and RNAP variants were incubated with[γ32P]ATP-labeled RfaH or [α32P]GTP (the incoming nucleotide specified by the template),and analyzed on an agarose gel.
Sevostyanova et al. Page 16
Mol Cell. Author manuscript; available in PMC 2012 July 22.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
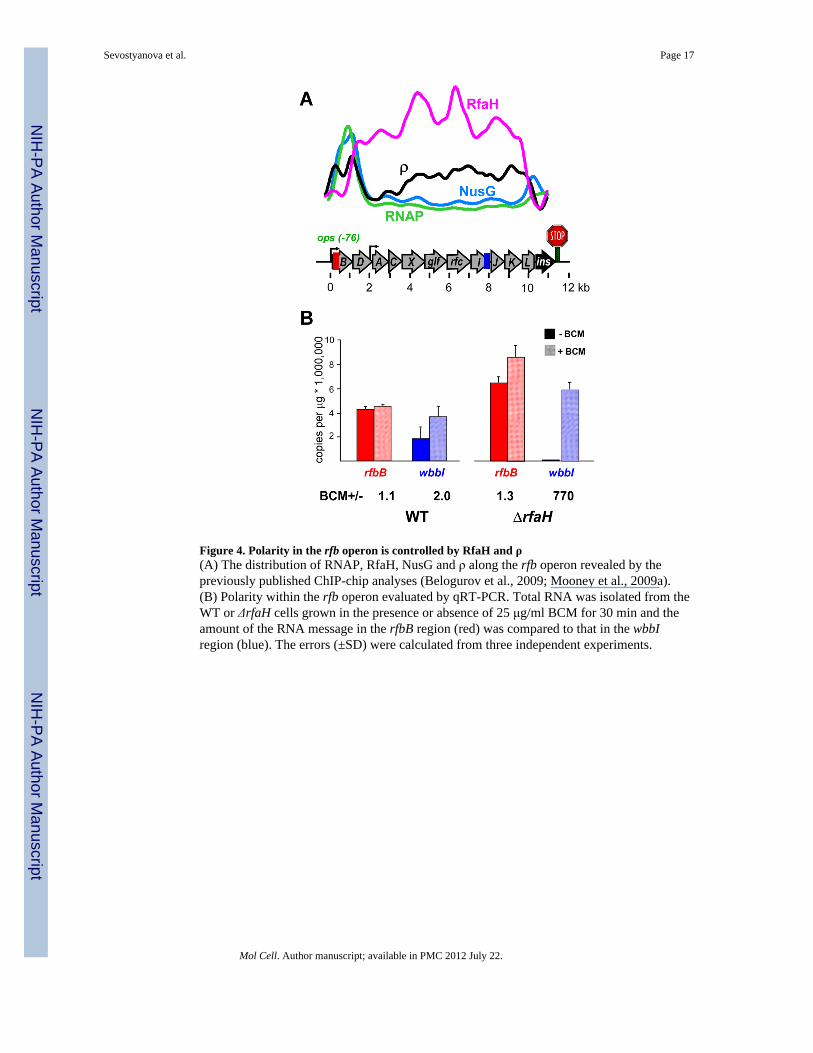
Figure 4. Polarity in the rfb operon is controlled by RfaH and ρ(A) The distribution of RNAP, RfaH, NusG and ρ along the rfb operon revealed by thepreviously published ChIP-chip analyses (Belogurov et al., 2009; Mooney et al., 2009a).(B) Polarity within the rfb operon evaluated by qRT-PCR. Total RNA was isolated from theWT or ΔrfaH cells grown in the presence or absence of 25 μg/ml BCM for 30 min and theamount of the RNA message in the rfbB region (red) was compared to that in the wbbIregion (blue). The errors (±SD) were calculated from three independent experiments.
Sevostyanova et al. Page 17
Mol Cell. Author manuscript; available in PMC 2012 July 22.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 5. Effects of the βGL deletion on RfaH function in vivo(A) Total RNA was isolated from cells expressing the WT or the ΔβGL RNAP in rfaH+ orΔrfaH strain and the amount of wbbI RNA was compared to that in the rfbB region. SeeFigure S4 for the absolute levels of each message.(B) Contributions of AP and NusG exclusion activities of RfaH to polarity. In lane 1, theRNAP is colored green to indicate the AP modification by RfaH. In lane 4, the fading NusG/ρ complex indicates its possible loss from the ΔβGL RNAP. Even though ρ may be bound tothe RNAP throughout transcription and in the absence of NusG (Epshtein et al., 2010; Peterset al., 2009), here we omitted ρ from the RfaH+ lanes since our data show that ρ is largelyinactive under these conditions.
Sevostyanova et al. Page 18
Mol Cell. Author manuscript; available in PMC 2012 July 22.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 6. NusG/GL interactions are required for AP but not for ρ–dependent termination(A) Pause assays on the linear pIA392 template shown on top with the scrambled ops(Artsimovitch and Landick, 2002) and the run-off (RO) indicated. Halted WT A38 ECs wereincubated with the WT or an altered NusG (at 100 nM) where indicated. Transcription wasrestarted upon addition of NTPs and rifapentin. Samples were taken at 5, 10, 20, 40, 90, 180,300 and 600 sec. Positions of the A38 and RO transcripts are indicated with arrows, thepause positions (at 39, 41, 43, and 45) are shown with a bracket. See Figure S5 foradditional details. The fractions of the RO and scrambled ops RNAs are presented as % oftotal RNA on the right; the errors (±SD) were calculated from three independentexperiments.(B) Termination assays were performed on pIA267 template as described (Belogurov et al.,2009). Halted, [α32P]GMP labeled ECs were formed at 40 nM with the WT or the ΔβGLRNAP. ρ (10 nM) and NusG (100 nM) were added where indicated, followed by incubationwith NTP substrates for 15 min at 37 °C. NusG facilitates ρ-mediated RNA release, shiftingthe termination window upstream (marked by a bracket); the fractions of RNA in this regionwere determined from three independent experiments (±SD).
Sevostyanova et al. Page 19
Mol Cell. Author manuscript; available in PMC 2012 July 22.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 7. Model of antipausing by RfaHTwo positions of the clamp domain are shown on a structural outline derived from the NTP-bound T. thermophilus EC (PDB id 2o5j (Vassylyev et al., 2007)) containing the helicalhairpin form of the trigger loop (orange) packed against the bridge helix (cyan). The nucleicacid chains are omitted for clarity. The alternative trigger loop position observed in T.thermophilus holoenzyme (PDB id 1i6h (Vassylyev et al., 2002)) is also shown. Anantitermination protein (AT, blue) can make bridging contacts between the GL (magenta)and clamp helices (green) across the DNA-binding channel. These contacts will preventclamp opening observed in the T. aquaticus core RNAP (darker grey; PDB id 1i6v (Zhang etal., 1999)) to stabilize RNAP interactions with the DNA and increase its processivity.
Sevostyanova et al. Page 20
Mol Cell. Author manuscript; available in PMC 2012 July 22.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript




















