
The Unsustainable Flow of Himalayan Fir Timber
17
The Unsustainable Flow of Himalayan Fir Timber Author(s): Laurent Umans Source: Mountain Research and Development, Vol. 13, No. 1 (Feb., 1993), pp. 73-88 Published by: International Mountain Society Stable URL: http://www.jstor.org/stable/3673645 . Accessed: 13/03/2014 03:53 Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at . http://www.jstor.org/page/info/about/policies/terms.jsp . JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range of content in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new forms of scholarship. For more information about JSTOR, please contact [email protected]. . International Mountain Society is collaborating with JSTOR to digitize, preserve and extend access to Mountain Research and Development. http://www.jstor.org This content downloaded from 137.224.252.10 on Thu, 13 Mar 2014 03:53:25 AM All use subject to JSTOR Terms and Conditions
Transcript of The Unsustainable Flow of Himalayan Fir Timber

The Unsustainable Flow of Himalayan Fir TimberAuthor(s): Laurent UmansSource: Mountain Research and Development, Vol. 13, No. 1 (Feb., 1993), pp. 73-88Published by: International Mountain SocietyStable URL: http://www.jstor.org/stable/3673645 .
Accessed: 13/03/2014 03:53
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
International Mountain Society is collaborating with JSTOR to digitize, preserve and extend access toMountain Research and Development.
http://www.jstor.org
This content downloaded from 137.224.252.10 on Thu, 13 Mar 2014 03:53:25 AMAll use subject to JSTOR Terms and Conditions

MOUNTAIN RESEARCH AND DEVELOPMENT, VOL. 13, No. 1, 1993, PP. 73-88
THE UNSUSTAINABLE FLOW OF HIMALAYAN FIR TIMBER
LAURENT UMANS
Department of Forestry Agricultural University Wageningen
The Netherlands
ABSTRACT The sustainability of Himalayan fir (Abies spectabilis) timber production in Nepalese village forests was analyzed by focusing on the availability of fir timber at village level. The availability was determined by the natural processes of change and by current utilization and management practices. Analysis of the natural processes of change started with an assessment of the internal dynamics of the fir forest. It revealed that regeneration was inhibited by the fir litter and this resulted in even-aged patches. Furthermore, large-scale rare destructive events were shown to have an important role in synchronizing regeneration. A combination of these natural processes results in periodic changes whereby a phase dominated by mature forest without regeneration is followed by complete destruction and fir regeneration. Thus, under natural conditions the flow of fir timber is not sustainable during all phases of this cycle.
This natural unsustainability is increased by several forest utilization practices: the oldest firs are selectively logged for shingle production; the young firs are harvested for pole production; fir regeneration is prevented by the accumulation of the litter layer, by grazing, and/or by the lopping of rhododendrons for fuelwood. All these practices, by their impact on fir regeneration, contribute to a decrease of the already naturally restricted possibilities for a sustainable flow of timber from the forest to the village society. To improve this sustainability alternative management options (based on ecological aspects only) have been proposed.
RgSUMI Le manque de stabiliti de la production de bois de sapin himalayien. La stabilit6 de la production de bois de sapin himalayien
(Abies spectabilis) dans les forets nepalaises a ete 6tudi6e en examinant la disponibilit6 au niveau des villages, d6termin6e par les processus naturels de changement et par les pratiques actuelles d'exploitation et de gestion. L'analyse des processus naturels de changement a commenc6 par une 6tude de la dynamique interne de la foret de sapin. Celle-ci a montr6 que la r6g6neration 6tait inhib6e par les d6bris et produisait des parcelles dont I'&ge '6tait uniforme. De plus, de rares ev6nements destructifs a grande 6chelle ontjou6 un r61e important dans la synchronisation de la r6g6n6ration. La combinaison des processus naturels a engendr6 des changements p6riodiques par lesquels une phase domin6e par la forkt de haute futaie sans r6g6neration est suivie d'une destruction complete et d'une r6g6neration de la fort de sapins. Donc, dans les conditions normales, le niveau de production de bois de sapin n'est pas stable pendant toutes les phases du cycle.
Ce manque naturel de stabilit6 est renforc6 par certains modes d'exploitation de la forkt: les vieux sapins sont abattus d'une maniere s6lective pour la production de bardeaux et les jeunes sapins pour la production de poteaux; la reg6n6ration est inhib6e par l'accumulation de d6bris, par le paturage et/ou par la coupe de rhododendrons pour le bois de chauffage. En vertu de leur impact sur la r6g6neration de la foret de sapins, toutes ces pratiques contribuent au d6clin du potentiel de production soutenue de bois de sapin, d6j'a limit6 par les facteurs naturels. D'autres options de gestion, uniquements bas6es sur les aspects &cologiques, ont ete propos6es de maniere ' augmenter la stabilite de la production.
ZUSAMMENFASSUNG Der nicht nachhdltige Fichtenholzertrag aus dem Himalaya. Die himalayische Fichtenholzproduktion (Abies spectabilis) auf nepalesischen Dorfwaldflaichen wird untersucht, wobei die Verffigbarkeit des Fichtenholzes ffir die Dorfbewohner im Mittelpunkt steht. Sie wird durch natfirliche Verinderungsprozesse und derzeitige Nutzungs- und Managementmethoden bestimmt. Eine Analyse der natfirlichen Verfinderungen begann mit der Beurteilung der internen Dynamik des Fichtenwaldes. Es ergab sich, daB die Regeneration durch umherliegende Fichtenspreu gehindert wurde, was wiederum zu Stellen mit gleichaltrigem Baumwuchs flhrte. AuBerdem zeigte sich, daB groBflaichige, destruktive Ereignisse eine wichtige Rolle im Regenerationszyklus spielen. Eine Kombination dieser natfirlichen Vorgfinge hat periodische Verfinderungen zur Folge, wobei auf eine Phase alten Baumbestandes ohne Regeneration eine vollkommene Destruktion und erst danach eine Regeneration des Baumbestandes folgt. Daraus ergibt sich, daB der Fichtenholzertrag unter natiirlichen Bedingungen nicht wihrend aller Phasen dieses Kreislaufs aufrechterhalten werden kann.
Diese natfirliche Ertragsbegrenzung wird noch durch verschiedene Waldnutzungsmethoden verstirkt: Die ailtesten Biume werden ffir die Dachschindelproduktion ausgesucht, und die jungen Biiume werden ffir die Pfahlherstellung gefiillt. Die Baumregeneration wird auBerdem durch eine Spreuschicht, durch das Weiden und/oder durch das Abschlagen des Rhododendrons zwecks Feuerholzgewinnung verhindert. Alle diese Tatsachen tragen durch ihre Einwirkungen auf die Fichtenregeneration dazu bei, die ffir die Dorfgemeinschaft bereits natfirlich begrenzten M6glichkeiten der Holzbeschaffung weiterhin zu schmilern. Alternative Managementm6glichkeiten unter Beriicksichtigung rein 6kologischer Aspekte, die den Holzertrag verbessern k6nnten, werden vorgeschlagen.
? International Mountain Society and United Nations University
This content downloaded from 137.224.252.10 on Thu, 13 Mar 2014 03:53:25 AMAll use subject to JSTOR Terms and Conditions

74 / MOUNTAIN RESEARCH AND DEVELOPMENT
INTRODUCTION
The vegetation of the lower subalpine zone in the Nepalese Himalaya is characterized by dense fir forests. These remote fir forests are normally not threatened by large-scale deforestation. Utilization is limited to the se- lective logging of fir trees for timber, the coppicing of Rhododendron trees for fuelwood, and grazing. Tree utiliza- tion practices are confined to small areas and have a direct impact at the micro level (specific sites) rather than at the macro level (the whole forest area). Grazing occurs virtually throughout the forest area.
In this paper the impact of utilization on the sustain- ability of fir timber production is evaluated. The sustained yield analysis refers to the future availability of forest products involving a large variety of factors. The focus is on fir demography and vegetation dynamics, and as-
pects such as soil nutrient depletion are not discussed. Data on vegetational processes in the fir forest are
derived from empirical research conducted by the author who thus gained an understanding of the ecology of the fir forest. The human impact will be evaluated in the light of this knowledge on ecosystem dynamics. Additional data were gathered on the successional changes linking the fir forest ecosystem to other vegetation types (particularly pastures). To thoroughly understand these relationships, a literature survey (particularly focusing on pure Rhododen- dron forest) was undertaken. To complete the overview the effects of pastoralism have also been taken into account, contributing to the aim of the paper: integration of the findings of the author's empirical research into a broader review of literature and research.
RESEARCH AIMS, SITES, AND METHODS
The data in this paper were derived from 15 months of fieldwork in central Nepal between 1985 and 1989. The objective of the research was threefold. First, to analyse the internal dynamics in the fir forest ecosystem. Second, to get a better understanding of the conditions underlying the various vegetation types present in the fir forest: are these types related to each other through succession? Third, to assess the effects of fir forest utilization on the sustainability of fir timber production to analyse possible fir timber depletion.
RESEARCH SITES The research area is situated in the lower subalpine
belt between 2,900 and 3,500 m above sea level, south of the Himalayan range in central Nepal, dominated by Abies spectabilis and Rhododendron trees (Rh. arboreum, Rh. barbatum, and Rh. campanulatum), commonly in a mixed forest with an Abies overstorey and Rhododendron under- storey. Pure forests of either Abies or Rhododendron species do exist. The term "fir forest" is used for either pure Abies forest or mixed Abies-Rhododendron forest. On some sites, sometimes due to particular human interventions, there were other types of vegetation, such as pastures, shrubs, bamboo thickets, and Juniperus forest.
From within this vast fir forested area the forests of two villages (Salme in Nuwakot District and Lapilang in Dolakha District) were studied in detail (Figure 1). Both villages had a similar population pressure (the surface of accessible fir forest divided by the number of inhabitants) and exposure of their fir forest (southeast). A general impression of a large part of the research area was gained by extensive trekking.
METHODS
Ecological research in the fir forest was conducted to gather information referring to the first two objectives of the research. A total of 97 homogeneous plots of 20 x 20 m (horizontal projection) were surveyed. The
number of plots was based on the time available. The choice of a plot was limited to a range in altitude (2,900-3,500 m), exposure (90'-170' or east to south), and angle of inclination (30%-90%). Plots were chosen in such a way to maximize a representative picture of the total village fir forest and to represent the various phases of fir forest development. Within the plots, girth measure- ments of all of the trees (at 1.3 m) were taken including the logged trees (where possible at 1.3 m), provided the girth was larger than 5 cm. Of fir seedlings and saplings (with a girth smaller than 5 cm) only the tree height was measured. Firs with a scar (made to check the suitability for logging) were recorded separately. Besides detailed research within plots, observations were made on the vegetation dynamics during extensive trekking. Informa- tion for the first two aims of the research was completed by reviewing the literature.
The third objective (the effects of utilization on the sustainability of fir timber production) required the analysis of fir forest utilization combined with the assess- ment of fir product availability. For this purpose 83 inhabitants of Salme and Lapilang were interviewed using formal open-ended interviews. Only men were inter- viewed because women do not log in the fir forest. Forest utilization was studied through participant observation. To indicate possible changes in the quality of fir timber, measurements of 20 old and 20 new shingles (the most important fir product) on each of 11 houses in Salme and 15 houses in Lapilang were carried out. The surfaces of various fir forest stands were estimated to relate fir population characteristics to fir product availability. These estimates were based on observations, photographs, and maps (Frits Thyssen Stiftung: Lapchi Kang Nepal, 1:50,000, Wenen; HMG Survey Department: Land Utiliza- tion Map, 71H/4 and 721/1, 1:50,000, Kathmandu; Wiart (1983), Unit6s &cologiques du versant de Salme, 1:25,000, Grenoble). Additional information was derived from the literature.
This content downloaded from 137.224.252.10 on Thu, 13 Mar 2014 03:53:25 AMAll use subject to JSTOR Terms and Conditions

L. UMANS / 75
TIBET (CHINA)
NEPAL Lapilang Salme
Kathmandu S main roads
to India FIGURE 1. Map of the research sites within Nepal.
RESULTS AND DISCUSSION
In the structural analysis of the fir forests in the research villages three relevant levels were distinguished: micro, meso, and macro level (Figure 2). The macro level encompasses the fir forest of one village. Several forest stands were distinguished within this village fir forest (e.g., mature fir forest, regenerating fir forest, pure Rhododendron forest) as well as pastures and shrub vegeta- tion. These forest stands and other ecological land units comprised the meso level. Focusing on the structural characterisics of the fir forest stands revealed that these areas were made up of more or less even-aged fir patches,
which represented the micro level. The three levels (patches, stands, and village forest) are presented schemat- ically in Figure 2. They provided a framework within which analysis of the vegetation dynamics and fir timber production at the appropriate scales (micro, meso, and macro) was accomplished. Thus, the first objective of the research refers to the dynamics within the meso level of fir forest stands. The second aim refers to the dynamics in other ecological land units, and the third to the overall availability of fir timber in the village (macro level).
FIR FOREST AND ITS INTERNAL DYNAMICS
The assessment of future availability of fir products at village level started with a fir population analysis revealing the internal dynamics of the fir forest. The first part of this analysis was an investigation on the conditions underlying regeneration of fir trees and fir forest forma- tion. The second part was a study on the transformation of mature fir forest into other types of vegetation.
FIR REGENERATION WITHIN THE MATURE FIR FOREST
Analysis of fir seedling establishment revealed that seedlings are often found on fallen logs without litter. The near absence of seedlings on the ground and their presence on fallen logs were observed both in and outside gaps, indicating that germination is not restricted by light availability. The regeneration of firs on shady sites such as under Tsuga dumosa trees, in Juniperus squamata forest,
and in some pure Rhododendron arboreum patches also supported this view. The pattern of seedling establish- ment probably reflects soil or litter properties. The presence of fir litter seems to be an important restrictive factor in the regeneration of firs and affects future availability of fir products. Either physical properties, such as moisture content, or chemical properties of the litter might cause this inhibition of regeneration. The latter case, called allelopathy, is mentioned in publications on fir forests in Europe (Westhoff, 1965). It has been suggested that the needles of the fir contain a chemical substance, leaching into the soil and suppressing the germination of fir seeds. Apart from the above mentioned factors the lack of seedlings might be a result of grazing.
If seedling establishment is successful, the sapling
This content downloaded from 137.224.252.10 on Thu, 13 Mar 2014 03:53:25 AMAll use subject to JSTOR Terms and Conditions

76 / MOUNTAIN RESEARCH AND DEVELOPMENT
village boundary - -
S\ 3500 meter I pure Rhododendron stand pasture
S young fir stand
pasture with patches
mature fir stand A pasture with micro level patches / \
2900 meter
bamboo thicket
shrub
regenerating fir stand
with patches
Legend: . . indicates macro level (village fir forest) indicates meso level (fir stand) indicates micro level (fir patch)
FIGURE 2. Schematic representation of various scale levels.
phase is reached at first branching. The absence of saplings within the plots representing the mature fir forest was striking (Richard, 1980). The survival of saplings in mature fir forests is severely limited, probably due to the lack of sufficient light or lack of the appropriate sub- statum in the case of establishment on fallen logs. It could be concluded that there is no regeneration (seedling and sapling establishment and growth) within the mature fir forest.
Micro level: even-aged patches The lack of successful regeneration in an established
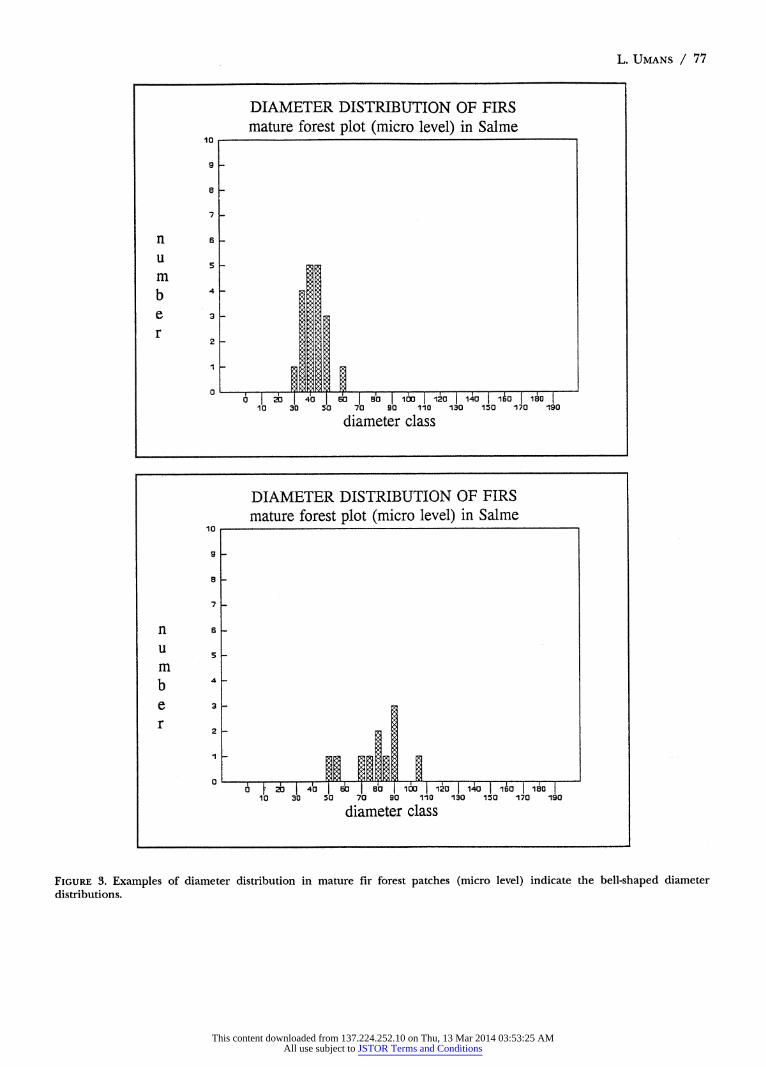
fir patch is reflected in the diameter distribution of older patches. Analysis at micro level (400 m2 plots) reveals a bell-shaped diameter distribution as shown in Figure 3.
Similar bell-shaped diagrams were produced by re- searchers when analysing the age distribution of firs (Sueron, 1985; Schmidt-Vogt, 1990a). Their results in- dicate that at micro level regeneration occurs within a period of 40 years (Sueron, 1985). Thereafter, the homo- geneous fir patch ages without further regeneration. The bell-shaped distribution pattern remains more or less constant and shifts along the horizontal axis.
Meso level: fir forest stands Because regeneration does not occur in an established
fir forest patch, it is important to determine the gap-size required for successful regeneration. Observations re- vealed that abundant fir regeneration can occur in openings of 300 m2 and larger. In a mature fir forest
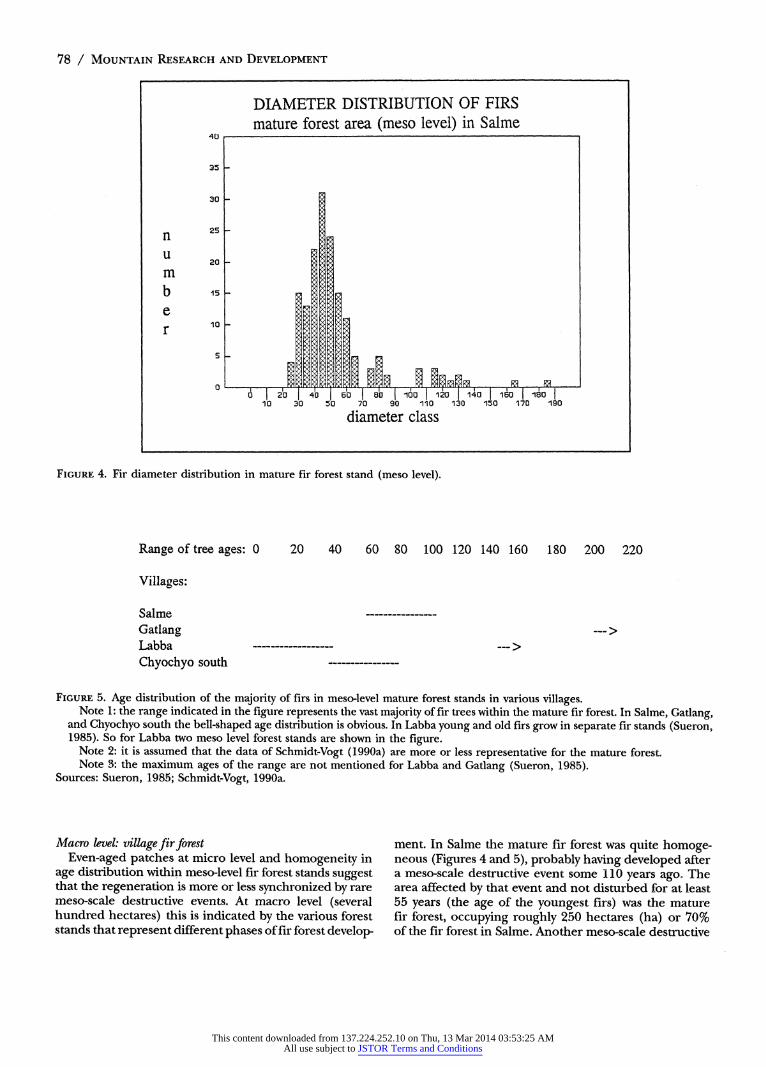
there are considerable changes in the density and/or surface area of the firs every 500 to 1,000 m2 (Sueron, 1985). This is an indication of the size of regeneration units. In general, the fir forest stands can be charac- terized as a patchy pattern of more or less homogeneous fir patches. Although the diameter distribution of firs within the various patches differed from one patch to another, overall differences were limited. Aggregating representative plots within a larger stand of several hectares (meso level) revealed once again a bell-shaped diameter distribution of firs (Figure 4). In the literature similar results were obtained for age analysis within meso- level mature forest stands (Figure 5). Focusing on age differences between regeneration patches along a tran- sect led to the conclusion that the rate of change in density and/or surface area was high (indicating the small patch sizes) but the amplitude of change was limited (Sueron, 1985).
Up to now the focus has been mainly on the mature fir forest characterized by firs of 15-35 m height with a diameter of 25 cm or more. In some stands the fir forest formation at meso level had started comparatively re- cently so the fir forest is not yet mature, for example in Labba (Figure 5). The meso-level young fir forest stand in Lapilang is another example (Figure 6), where the majority of firs were 2-10 m high, with a diameter of 5-25 cm. In Salme the regenerating fir "forest" sampled was characterized by fir seedlings and saplings that do not yet attain a height of over 2 m (Figure 7).
This content downloaded from 137.224.252.10 on Thu, 13 Mar 2014 03:53:25 AMAll use subject to JSTOR Terms and Conditions
L. UMANS / 77
DIAMETER DISTRIBUTION OF FIRS mature forest plot (micro level) in Salme
10
9-
n B u
b 4
e r
2
01
4 0 80 100 120 140 160 180 10 30 50 70 90 110 130 150 170 10
diameter class
DIAMETER DISTRIBUTION OF FIRS mature forest plot (micro level) in Saline
10
7-
n s- U m b 4 e 3- r 2-
0 Y1lol Icu ~ 4 ~ a 2b 40 5 1 810 160 12a .0 1 16,0 18'a 10 30 50 70 90 110 130 150 170 190
diameter class
FIGURE 3. Examples of diameter distribution in mature fir forest patches (micro level) indicate the bell-shaped diameter distributions.
This content downloaded from 137.224.252.10 on Thu, 13 Mar 2014 03:53:25 AMAll use subject to JSTOR Terms and Conditions
78 / MOUNTAIN RESEARCH AND DEVELOPMENT
DIAMETER DISTRIBUTION OF FIRS mature forest area (meso level) in Saline
4U
30-
S 25-
U 20 - m b e r 10 2
SX
0K (3.... 1)t q o B b so 6c ;o I1i4o I
i & o 10 30 5 70 90 110 130 150 170 190
diameter class
FIGURE 4. Fir diameter distribution in mature fir forest stand (meso level).
Range of tree ages: 0 20 40 60 80 100 120 140 160 180 200 220
Villages:
Salinme ---------------- Gatlang --- > Labba ------------------- Chyochyo south------------
FIGURE 5. Age distribution of the majority of firs in meso-level mature forest stands in various villages. Note 1: the range indicated in the figure represents the vast majority of fir trees within the mature fir forest. In Salme, Gatlang,
and Chyochyo south the bell-shaped age distribution is obvious. In Labba young and old firs grow in separate fir stands (Sueron, 1985). So for Labba two meso level forest stands are shown in the figure.
Note 2: it is assumed that the data of Schmidt-Vogt (1990a) are more or less representative for the mature forest. Note 3: the maximum ages of the range are not mentioned for Labba and Gatlang (Sueron, 1985).
Sources: Sueron, 1985; Schmidt-Vogt, 1990a.
Macro level: village fir forest Even-aged patches at micro level and homogeneity in
age distribution within meso-level fir forest stands suggest that the regeneration is more or less synchronized by rare meso-scale destructive events. At macro level (several hundred hectares) this is indicated by the various forest stands that represent different phases of fir forest develop-
ment. In Salme the mature fir forest was quite homoge- neous (Figures 4 and 5), probably having developed after a meso-scale destructive event some 110 years ago. The area affected by that event and not disturbed for at least 55 years (the age of the youngest firs) was the mature fir forest, occupying roughly 250 hectares (ha) or 70% of the fir forest in Salme. Another meso-scale destructive
This content downloaded from 137.224.252.10 on Thu, 13 Mar 2014 03:53:25 AMAll use subject to JSTOR Terms and Conditions

L. UMANS / 79
DIAMETER DISTRIBUTION OF FIRS young forest area (meso level) in Lapilang
80
70-
sO -
50o n u
40-
m b 30 o e
20 r
10
20 40 a0 0 1l 120 140 160 150 10 30 50 70 90 110 130 150 1L0 10
diameter class
FIGURE 6. Fir diameter distribution in young fir forest stand (meso level).
HEIGHT DISTRIBUTION OF FIR SEEDLINGS regenerating plot (micro level) in Salme
30
28 - 2S - 24-
22
20 n 19 fl is U 15
m 14
b 12-
e 10 e
r 6
4
2
0 20140 60 80 100 120 140 160 180 2 0
10 30 50 70 90 110 130 150 170 190 210
height class
FIGURE 7. Height distribution of fir seedlings and saplings in regenerating fir forest (micro level) representing the burned meso- scale stand.
This content downloaded from 137.224.252.10 on Thu, 13 Mar 2014 03:53:25 AMAll use subject to JSTOR Terms and Conditions
80 / MOUNTAIN RESEARCH AND DEVELOPMENT
DIAMETER DISTRIBUTION OF FIRS Salme (macro level)
150 180 -
170 -
160 -
150 -
140 -
130 -
120 -
m so- 60 -
50 - r 40 -
30- 20 - D 10
0
10 30 50 70 90 110 130 150 170 190
diameter class
DIAMETER DISTRIBUTION OF FIRS Lapilang (macro level)
150
180 -
170 -
ISO -
150I-
140 -
130 -
120 -
110 -
U 100 -
m 90- m so b 70 -
e 0 - so r
40- 30 -
20
10
0 20 40 so0 80 1W 2 40 15 10 10 30 50 70 90 110 130 150 170 190
diameter class
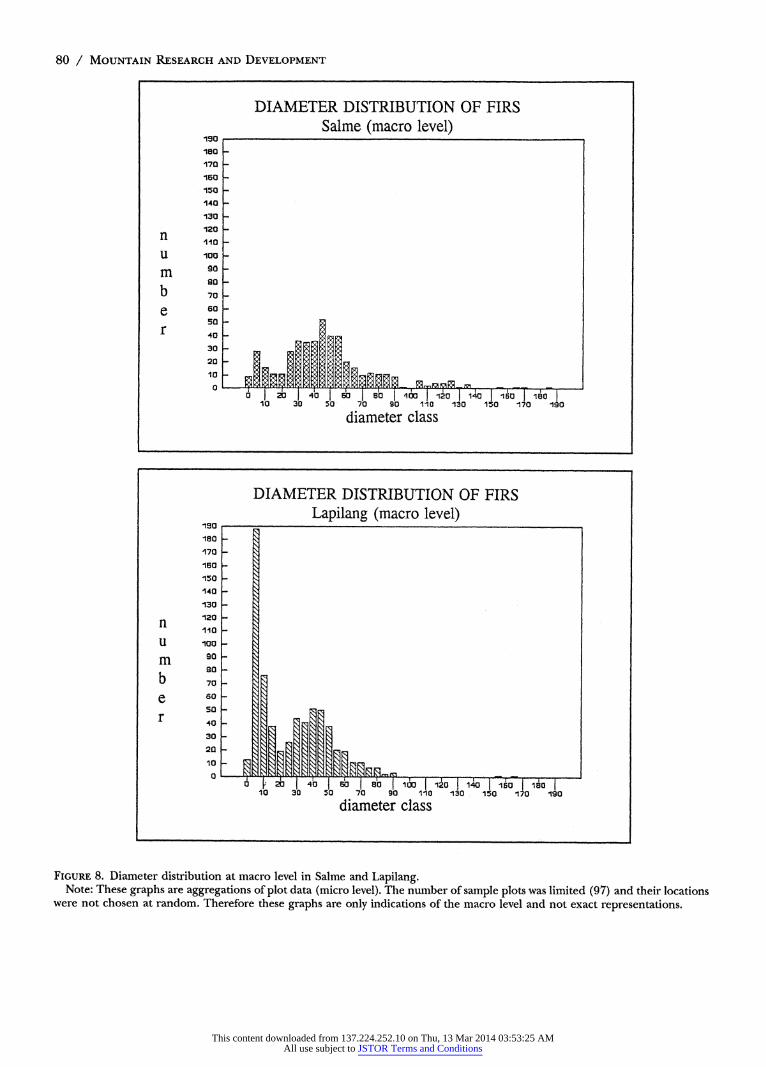
FIGURE 8. Diameter distribution at macro level in Salme and Lapilang. Note: These graphs are aggregations of plot data (micro level). The number of sample plots was limited (97) and their locations
were not chosen at random. Therefore these graphs are only indications of the macro level and not exact representations.
This content downloaded from 137.224.252.10 on Thu, 13 Mar 2014 03:53:25 AMAll use subject to JSTOR Terms and Conditions

L. UMANS / 81
event occurred about 15 years ago when a site of approx- imately 20 ha (6% of the fir forest in Salme) was burned. Abundant regeneration of firs observed on this site (Figure 7) showed that fir forest development had just begun. Thus, at macro level two meso-level forest stands could be distinguished: the mature forest and the regen- erating burned area. These meso-level stands reflected the history of meso-scale destructive events. However, due to the small size of the area and the small size of the regenerating firs, the diameter distribution at macro level (Figure 8) was similar to that of the mature fir forest at meso level (Figure 4).
In Lapilang the young fir forest stands (Figure 6) were approximately 45 ha (12% of the fir forest in Lapilang). The mature fir forest was 160 ha (45%). The relative contribution of the young fir forest was large enough to show up in the diagram at macro level (see Figure 8).
In most Nepalese villages visited on the trek, the village fir forest consisted of one large and a few small meso- level forest stands (the Salme situation). The diameter distribution at macro level is roughly similar to that of the dominant meso-level forest stand.
At macro level, the relative proportion of the various more or less even-aged meso-level forest stands is of great significance for the future availability of fir timber. In Salme, this is not only restricted by the lack of regenera- tion in the mature fir forest at meso level but, more importantly, by the fact that there is a lack of meso-level regenerating forest stands at macro level. Thus the frequency and size of these meso-scale destructive events are extremely important.
Rare large-scale destructive events Rare meso-scale destructive events seem to play an
important role in triggering the regeneration of firs. However, the nature of these events is not yet clear. The first possibility is the meso-scale collapse of the even-aged forest due to natural mortality of old trees or pest outbreaks. In mixed Abies-Rhododendron forests this leads to pure Rhododendron forest because Rhododendron outlives Abies (Schmidt-Vogt, 1990b). In these pure Rhododendron forests there is no regeneration of firs, so the re- establishment of a fir forest via a phase of pure Rhododen- dron forest depends on the internal dynamics of the latter (see below).
The second possibility is large-scale fire. Recently this factor has received much attention due to catastrophes such as that in Yellowstone National Park (USA) in 1988. Studies on fire histories have revealed that large, con- tinuous, even-aged forests occur on sites where conditions allow fuel accumulation and consequently, intense fires during droughts (Tande, 1979). The accumulating bio- mass in aging trees in even-aged patches fuels the fire. In young forests the accumulated biomass is limited and the fires are small. However, at a certain point in time the accumulated biomass in even-aged forest is so large that a fire can affect vast areas of up to thousands of hectares. The 1988 fires in the approximately 250-300-year-old lodgepole pine (Pinus contorta var. lat- ifolia) forest of Yellowstone National Park are an example
(Sprugel, 1991). The extensive, even-aged pure fir and spruce fir stands in Canada are similarly unquestionably due to fire (as well as spruce bud worm). The Picea-Abies forests burned less frequently than fire adjusted climax forests of pines but these fires that did occur were more commonly severe, stand-replacing crown fires (Barbour and Billings, 1988). Although there are no accounts of such meso-scale fires in Himalayan fir forests, its occur- rence must be considered as highly probable. The lack of accounts may be because of the dominance of relatively young forests. Gatlang (Figure 5) is an exception. The rarity of such events may also be an explanation.
These results lead to my hypothesis that the fire regulating mechanism is an ecological equivalent of the chain reactions in nuclear physics after the so-called critical mass is surpassed. The "critical biomass" can therefore be defined as the minimal accumulated biomass necessary to enable potential disturbances to develop into a self-expanding process through chain reactions. As long as the critical biomass is not yet reached, the occuring fires affect small areas and are exhausted rapidly. They probably do not develop into crown fires. When the critical biomass is reached and conditions are appro- priate, ground fires develop into large-scale intense crown fires. These self-expanding fires are very difficult to control by man and his machines and are limited by exhaustion of fuel, natural firebreaks, or a change in the weather (Wright and Heinselman, 1973).
This hypothesis on critical biomass is in line with the observations on the vegetation in the Himalayan fir forest: even-age fir forest units are either small patches within a large stand or considerable areas (large meso- scale units). Small meso-scale even-aged fir forests are virtually absent. Small meso-scale vegetation units are mostly pastures and sometimes logged areas. Natural fires seem to play no role at this level. However, these observa- tions and the concept of "critical biomass" need further investigation to test the hypothesis. This might pose problems when the natural fire regime is influenced by man. This seems the case in northern North American forests, where Indians have influenced the natural fire regime prior to European settlement (Barrett and Arno, 1982). In these cases the hypothesis of "critical biomass" cannot be tested. The same situation might apply to the fir forests on southern-exposed slopes in the Himalaya, where herders have a long history of burning. However, the northern-exposed slopes seem unaffected by man and thus suitable for investigating the critical biomass.
If fruitfull, the assessment of the "critical biomass" is an important parameter to be studied in Himalayan fir forest and in American fire-adjusted forests. It can refer to several types of biomass: inflammable litter, standing timber stock accumulating over time in the even-aged patches, and possibly the biomass of insects causing extensive mortality and thus increasing the susceptibity to fire (e.g., the interaction between bud worm and fire).
Reviewing the internal dynamics of the fir forest the following statements can be made:
This content downloaded from 137.224.252.10 on Thu, 13 Mar 2014 03:53:25 AMAll use subject to JSTOR Terms and Conditions

82 / MOUNTAIN RESEARCH AND DEVELOPMENT
* Rare meso-scale destructive events synchronize regeneration.
* The fir as a pioneer tree gradually colonizes the affected site. A patchy pattern characteristic for large- scale pioneer vegetation emerges.
* After forest establishment on a particular micro-level site further fir regeneration is prevented due to the fir litter properties (especially allelopathy) and in- sufficient light. The age distribution of firs on a micro site is bell-shaped.
* Under natural conditions the disturbance of the fir
forest is rare. Gaps required for regeneration hardly appear. The fir tree population ages without sig- nificant changes and the patchy pattern remains intact.
* Eventually the fir forest gives way to pure Rhododen- dron forest or is destroyed by fire.
* At macro level, a period or phase of mature fir forest without regeneration is eventually followed by a period or phase of abundant regeneration without mature fir forest. The new "generation" emerges only after the death of the old "generation."
MANAGEMENT AND UTILIZATION OF FIR TIMBER
The internal dynamics as outlined above have serious consequences for the sustainable flow of fir products from the forest to society. Under natural conditions both the regeneration of firs and the fir timber availability at macro level are discontinuous. Within the period of this natural cycle there is no sustainability at macro level. It is obvious that utilization and management should be directed towards manipulating the natural cycle in such a way that the sustainability is enhanced. In short this might be done in the following ways:
* preventing wasteful large-scale destructive fires in old- growth fir forest,
* ensuring regeneration in stands dominated by mature fir forest by initiating small-scale clearcutting, litter removal, or controlled burns.
* ensuring regeneration on burned sites by restricting grazing.
* restricting the utilization of mature firs left after a large-scale destructive event (resource scarcity, seed trees).
The information on internal dynamics and manage- ment options is used as a reference, and the present forest utilization practices are described together with an assess- ment of their impact on sustainability.
SELECTIVE LOGGING FOR SHINGLE PRODUCTION In the field, the present use of the fir forest was studied
by means of interviews and participant observation. Additional information was derived from literature re- search (Wiart, 1983; Schmidt-Vogt, 1990a). The results indicate that fir timber is the main product harvested by villagers. Fir timber falls into two distinct categories: mature trees logged for processing into shingles and young firs logged for poles.
Shingle production requires large firs with straight wood fibers and a high first branch so that the trunk segments can be split into shingles by using wedges. Before logging a tree the peasants determine the grain by cutting off a piece of wood so that unsuitable trees are not logged. In depleted areas of the mature fir forest unsuitable trees make up 35 to 50% of the original fir biomass. Thus, overexploitation does not lead to defor- estation (in the sense of a decrease in forest area) but to product depletion and minor structural and possibly
genetic changes of the forest. Once trees are logged they are processed into shingles at the logging site. Processing and further transportation to the village is done by man power. The effects on the soil are therefore limited to a few small paths. At first sight intensive exploitation does not cause the common environmental problems such as deforestation, soil compaction, erosion, and hydrological disturbance. Instead economic problems due to increas- ing resource scarcity (increasing time investment, ex- pensive substitution) arise.
When the fir regeneration conditions are taken into account, it would appear that selective fir logging is silviculturally inappropriate. This method of logging does not cause enough disturbance of the litter layer to trigger abundant fir seedling establishment; nor does it create gaps of at least 300 m2 to enable sapling growth or gradual decomposition of the fir litter. Because logging does not trigger regeneration, the mature fir forest is gradually exhausted. An indication of the speed of this process is the sharp decline in shingle size (Figure 9), where shingles made in 1985 are compared with shingles of previous years on the same roofs. In fact the old shingles may even have been wider when they were produced because the sides are sometimes chopped off as part of the maintenance process.
Resource depletion is very obvious in a relatively old fir forest that has no regeneration patches. Logging trees with the largest diameters initially has little effect because every next-largest class of diameters has more individuals (Figure 10). However, if all trees larger than the median diameter are logged or are unsuitable, product depletion is accelerated because every suitable next-largest class has fewer individuals. Furthermore, by decreasing tree size, the number of trees needed increases. In Salme, the average stump of a tree logged for shingle production was 78 cm and the average number of shingles harvested per tree was 294. In Lapilang, these numbers were 57 and 268 respectively. In Salme, the village fir forest can be characterized as "user-friendly" due to the present resource abundance. Nevertheless, accelerated product depletion is a serious threat to a sustainable flow of fir shingles. In Lapilang, the present scarcity of big firs poses problems for the users. Nevertheless, conditions for the sustainable production of fir timber production seem better than in Salme.
This content downloaded from 137.224.252.10 on Thu, 13 Mar 2014 03:53:25 AMAll use subject to JSTOR Terms and Conditions

L. UMANS / 83
WIDTH DISTRIBUTION OF SHINGLES old shingles
32
30
29-
26k
24-
22-
20-
19 U 16 -
m 14. b 12-" e 10" r 8a- 6-
4
2
3 7 5 11 13 15 17 19 21 23 25 27 25 31 33 33 37 35
width class
WIDTH DISTRIBUTION OF SHINGLES new shingles
32
30 -
29 -
26 -
24 -
22
20
18
u 16 m 14 b 12
e 10 r 8-
6
4
2 C
0 5 97 11 13 15 17 19 21 23 25 27 25 31 33 35 37 39
width class
FIGURE 9. Width of old and new (made in 1985) shingles in Salme indicating resource depletion.
This content downloaded from 137.224.252.10 on Thu, 13 Mar 2014 03:53:25 AMAll use subject to JSTOR Terms and Conditions

84 / MOUNTAIN RESEARCH AND DEVELOPMENT
GRADUAL RESOURCE DEPLETION
54-
n U
3 m b
2
e r
0 0 10 20 30 40 50 50 70 so80 90 100 110
diameter class Ssuitable present harvest unsuitable
ACCELERATED RESOURCE DEPLETION 5
5
4
n u
3 m" b
2 e cc r Da
'1a
0 - - -
0. 10 20 30 40 50 60 70 80 s0 100 110
diameter class Ssuitable s present harvest unsuitable
FIGURE 10. Gradual and accelerated resource depletion assuming preferential logging of the largest diameters.
This content downloaded from 137.224.252.10 on Thu, 13 Mar 2014 03:53:25 AMAll use subject to JSTOR Terms and Conditions
L. UMANS / 85
REVERSED J-SHAPED DISTRIBUTION an example
loo
90o-
8 0-
70 -
n 60so -
U m b 40
e 30
r 20
10
kq
5 10 15 20 25 30 35 40 45 50 class
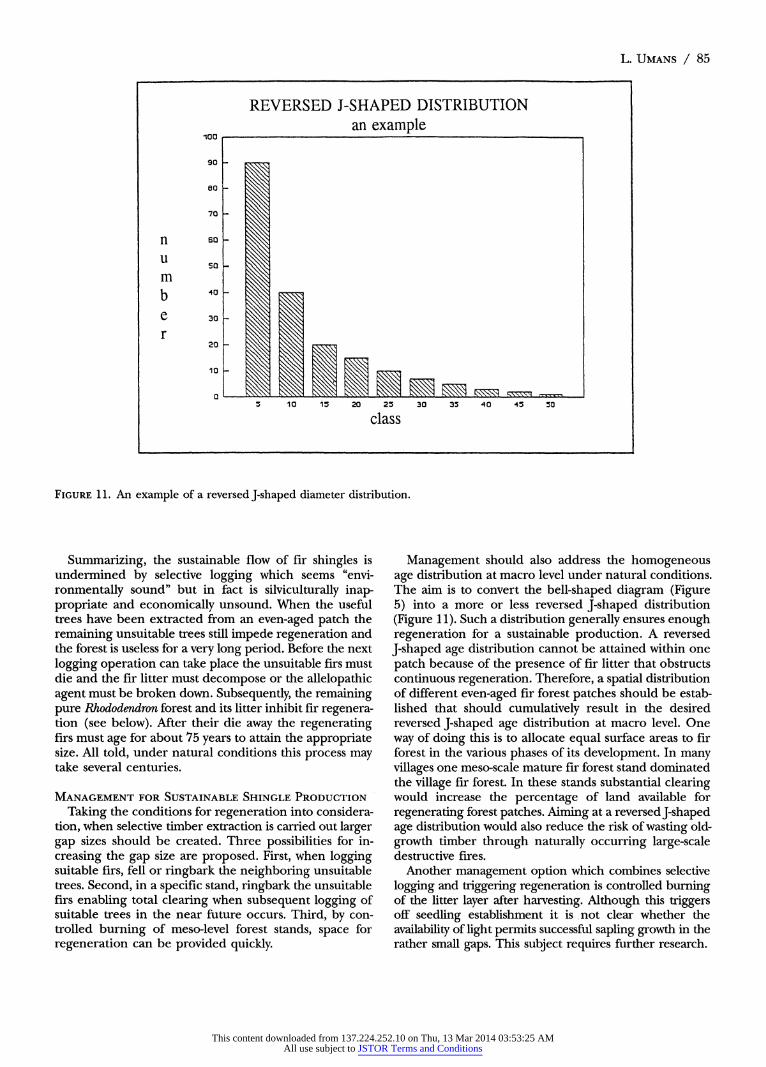
FIGURE 11. An example of a reversed J-shaped diameter distribution.
Summarizing, the sustainable flow of fir shingles is undermined by selective logging which seems "envi- ronmentally sound" but in fact is silviculturally inap- propriate and economically unsound. When the useful trees have been extracted from an even-aged patch the remaining unsuitable trees still impede regeneration and the forest is useless for a very long period. Before the next logging operation can take place the unsuitable firs must die and the fir litter must decompose or the allelopathic agent must be broken down. Subsequently, the remaining pure Rhododendron forest and its litter inhibit fir regenera- tion (see below). After their die away the regenerating firs must age for about 75 years to attain the appropriate size. All told, under natural conditions this process may take several centuries.
MANAGEMENT FOR SUSTAINABLE SHINGLE PRODUCTION
Taking the conditions for regeneration into considera- tion, when selective timber extraction is carried out larger gap sizes should be created. Three possibilities for in- creasing the gap size are proposed. First, when logging suitable firs, fell or ringbark the neighboring unsuitable trees. Second, in a specific stand, ringbark the unsuitable firs enabling total clearing when subsequent logging of suitable trees in the near future occurs. Third, by con- trolled burning of meso-level forest stands, space for regeneration can be provided quickly.
Management should also address the homogeneous age distribution at macro level under natural conditions. The aim is to convert the bell-shaped diagram (Figure 5) into a more or less reversed J-shaped distribution (Figure 11). Such a distribution generally ensures enough regeneration for a sustainable production. A reversed J-shaped age distribution cannot be attained within one patch because of the presence of fir litter that obstructs continuous regeneration. Therefore, a spatial distribution of different even-aged fir forest patches should be estab- lished that should cumulatively result in the desired reversed J-shaped age distribution at macro level. One way of doing this is to allocate equal surface areas to fir forest in the various phases of its development. In many villages one meso-scale mature fir forest stand dominated the village fir forest. In these stands substantial clearing would increase the percentage of land available for regenerating forest patches. Aiming at a reversed J-shaped age distribution would also reduce the risk of wasting old- growth timber through naturally occurring large-scale destructive fires.
Another management option which combines selective logging and triggering regeneration is controlled burning of the litter layer after harvesting. Although this triggers off seedling establishment it is not clear whether the availability of light permits successful sapling growth in the rather small gaps. This subject requires further research.
This content downloaded from 137.224.252.10 on Thu, 13 Mar 2014 03:53:25 AMAll use subject to JSTOR Terms and Conditions

86 / MOUNTAIN RESEARCH AND DEVELOPMENT
UTILIZATION AND MANAGEMENT OF YOUNG FIRS
Young firs processed into poles are used for construc- tion of houses and cattle sheds as well as for religious purposes (Wiart, 1983). Currently, the harvesting of poles is mainly practiced in accessible mature fir forests (espe- cially in Salme and Gatlang, where young forests are absent). This practice of harvesting poles diminishes the already limited future availability of suitable trees.
A more appropriate management option is the thin- ning of young densely-stocked fir forests. With proper management, high turn-over pole production sites can be established. In one plot (Schmidt-Vogt, 1990a) a 53-year-old fir dominated a new generation of 16-26-year-
old trees. Harvesting of the old tree and all but one of the 24 young trees might reproduce a similar cycle every 35 years. Pole production can be sustained by continued well-planned disturbance. Pole production on specific sites thus contributes to the conversion of a bell-shaped into a reversed J-shaped age distribution. Pole production and shingle production should not always be spatially separated. As part of sustaining the future shingle availa- bility by the creation of meso-level regenerating forest patches, thinning operations in these stands can contrib- ute to pole production. Moreover, it diminishes competi- tion among the young trees.
REVIEW OF RELATED LITERATURE
PURE RHODODENDRON FOREST AND ITS INTERNAL DYNAMICS
The presence of pure Rhododendron forest was men- tioned in the introduction. In the section on the fir forest it was said that a pure Rhododendron forest can result from the death of fir trees. It can also occur as a result of intensive fir logging (Schmidt-Vogt, 1990a). Some authors point out that natural causes can underlie the presence of a Rhododendron forest (Dobremez, 1976). The useful- ness of these pure Rhododendron forests for society is limited (Wiart, 1983) and the conversion of these forests into fir forest is desired. By evaluating the literature on the internal dynamics of pure Rhododendron forest an assessment is made of the conditions under which this conversion can be accomplished.
Fir regeneration in Rhododendron forest In pure Rhododendron stands fir seedlings as well as
Rhododendron seedlings are rare (Schmidt-Vogt, 1990b). Their presence seems restricted to sites where the litter layer is absent (Schmidt-Vogt, 1990a). This situation applies to young Rhododendron forest stands, where there has not been sufficient time to build up the litter layer. In mature forests a similar situation can be created by removing the accumulated litter.
It has been acknowledged in the literature that a young Rhododendron forest is established in meso-level openings. Rhododendron is a light-demanding pioneer. Observations and age analysis in mature fir forests confirm that successful regeneration of Abies occurs among these young Rhododendrons. In many mature fir forests the Rhododendron understorey is 10-20 years older than the Abies overstorey (Schmidt-Vogt, 1990a). Once the Rhodo- dendrons are older than 20 years the possibilities for fir regeneration decrease which is probably due to the build- up of a litter layer of Rhododendron leaves. Thus, if conditions for fir regeneration are met on time a mixed Abies-Rhododendron forest will be established, otherwise a pure Rhododendron forest might develop.
How can an older pure Rhododendron forest be trans- formed into a mixed Abies-Rhododendron forest? There are two options: removal of the Rhododendron litter and
removal of the Rhododendron forest. Removal of the litter layer in a pure Rhododendron forest can occur in two ways. First, by a combination of surface runoff and erosion, as observed in forest stands at the lower fringe of overgrazed pastures. Second, the litter can be removed by fire. In young and middle-aged Rhododendron forests the impact of fire seems limited to litter-burns, not killing the allegedly fire-resistant Rhododendrons. After the fire, fir seedling establishment has been observed in several cases (Schmidt-Vogt, 1990a). In other cases a build-up of the Rhododendron litter layer has recommenced.
Fire is not always restricted to litter-burns. In old Rhododendron forests with a thick and highly inflammable litter layer the fire is large-scale and destroys all trees. "The destructiveness of fire in Rhododendron forests is related to the thickness of the litter layer" (Schmidt-Vogt, 1990b). The accumulating litter layer is a kind of ecolog- ical time bomb for the Rhododendron forest. Initially it effectively obstructs any invasion of young trees, thus ensuring the status quo and a steady build-up of the litter. Ultimately this causes the ecosystem's own destruction by fire.
Fire is an important agent for vegetation dynamics enabling a fir forest to develop from a pure Rhododendron forest. Fir seedlings may become established after litter- burns. After large-scale burning that destroys the forest structure, various vegetation types could develop: pas- tures, bamboo thickets, and regeneration of mixed Abies- Rhododendron forest. The role of management in favor of vegetation in which fir regeneration occurs, is significant. Restricting grazing is one aspect (Alirol, 1979); repeated burning of the bamboo thickets is another (Mahat et al., 1987).
The insights gained on vegetation dynamics are com- bined in Figure 12.
SILVO-PASTORALISM AND PASTURE MANAGEMENT Fir forests are not only used for timber extraction, they
also contribute to the fodder required for sustaining animal husbandry. Migrating pastoralists and their herds regularly stay in the fir forest. Herds of mixed livestock (cows, buffalo) move to pastures created within the fir
This content downloaded from 137.224.252.10 on Thu, 13 Mar 2014 03:53:25 AMAll use subject to JSTOR Terms and Conditions

L. UMANS / 87
fir forest: simultaneous young mature regeneration forest forest
activity: restricted
selective grazing N
shIfglogging
for shingles zero-events:
grazing clearing natural
fire
for poles collapse
activity: overgrazing "
other pasture pure bamboo vegetation: Rhododendron thickets
forest
FIGURE 12. Vegetational dynamics and human intervention in the fir forest.
forest for longer periods in summer. Herds of Chauri (local cross-breeds) or sheep and goats, visiting these pastures briefly as part of their transhumance movement, stay for longer periods on the alpine pastures above timberline. Pressure on most pastures is severe and fodder is quickly depleted, and the animals browse in the adjoining forest. Abies seedlings are grazed or trampled except those growing under Viburnum shrubs (Richard, 1980) and Rhododendron shrubs (Schmidt-Vogt, 1990a). The fir forest is overgrazed especially in the upper reaches. On these sites several factors induce the accelera- tion of soil erosion on a massive scale: browsing, tram- pling, steep slope gradient, needle ice formation, and monsoonal rainfall (Schmidt-Vogt, 1990a).
The seasonal movement of herds is part of a manage- ment system for overall resource utilization (Alirol, 1979). Environmental concerns do not always underlie these systems. Agricultural needs (Berthet-Bondet, 1983; Metz, 1990) and economic interests (Brower, 1990) can result in "fallow" periods that are too short to ensure rehabilita- tion. Thus grazing can be seen as a large-scale continuous destruction of regenerating firs. In this way it is neither in line with the natural disturbance regime (large-scale rare destruction of mature forest) nor with the manage- ment options for a sustainable flow of timber products. Present grazing practices with detrimental effect on regeneration threaten the sustainable flow of fir products.
Alternative management options should be directed towards the temporary closure of those grazing areas
where conditions are right for regeneration. This can be achieved by "biological fencing" (e.g. establishment or protection of Rhododendron shrubs) and by "social fenc- ing" (e.g. temporary restrictions on rights of access).
Fuelwood and additional demands of pastoralists The high-altitude fir forests are usually too far away
from the village to serve as its resource for fuelwood. Fuelwood is however gathered by pastoralists for boiling milk, for cooking, and for heating. Due to the wet conditions a continuous fire is required to ensure a continuous supply of dry fuelwood (Schmidt-Vogt, 1990a) and to prevent decay of the roof (Metz, 1990). Rhododen- dron species are preferred and are intensely lopped in the fir forests surrounding the pastures. Regeneration of firs is impossible as the removal of the Rhododendron shrubs leaves the seedlings vulnerable to grazing and trampling (Schmidt-Vogt, 1990a).
In addition to fuelwood there are other demands on the forest resources by pastoralists. They need poles and shingles for their temporary shelters (prone to rapid decay under the wet conditions); they strip the bark off mature firs to cover floors; they lop fir for bedding material and utensils, severely reduce the fir population, particularly in the vicinity of pastures. As a result, the size of the pastures increases as does the area prone to accelerated soil erosion. Although economically profitable, the present practices associated with animal husbandry are ecologically unsound (causing deforestation and erosion) and silviculturally in- appropriate (restricting fir regeneration).
This content downloaded from 137.224.252.10 on Thu, 13 Mar 2014 03:53:25 AMAll use subject to JSTOR Terms and Conditions

88 / MOUNTAIN RESEARCH AND DEVELOPMENT
CONCLUSIONS
The ecological mechanisms underlying the internal dynamics of the fir forest result in a cyclic and syn- chronized process of change. Within the period of this cycle (up to several centuries) and within the spatial limits of the synchronizing process (several hundreds of hectares) the flow of fir timber from the forest to society is not sustainable. This naturally restricted sustainability can be improved by management practices that could manipulate the natural processes. First, the preventing of macro-level destructive events is of great importance. Second, the enabling of successful regeneration on the proper scale is essential. This might imply destruction of parts of the mature fir forest at meso level in combination with gap management to assure fir regeneration.
In respect to the growing interest in environmentally sound selective timber extraction, it should be noted that the sustainability of such a management system also depends on the ecology and population dynamics of the timber species. Selective timber logging seems inap- propriate if regeneration in small gaps is suppressed (e.g., through allelopathy or insufficient light) and/or if condi- tions are promoted for a large-scale rare destructive event. In a natural disturbance regime characterized by such large-scale rare destructive events, different management decisions serve different purposes:
* Sustainable timber availability is enhanced by meso- scale controlled destructive events. This management includes frequent removal of biomass accumulations on small-scale patches and clearing once conditions
for a catastrophe have developed. For this purpose the concept of "critical biomass" could be introduced and needs further investigation. In such a manage- ment system zoning and land-use planning are im- portant elements needed to avoid potentially negative impacts (such as erosion, land squatting, loss of aesthetical values, and loss of game.).
* Forest conversion is enhanced by continuous destruc- tive events such as grazing or annual burning in fir forest.
* The natural situation (not the natural vegetation) is preserved or enhanced by large-scale rare destructive events.
The manager thus has the choice between various fire regimes, ranging from "leaving it up to nature" to maximal fire suppression or maximal management to enhance the sustainable flow of forest products. If rural development is the primary goal, such as in the Nepalese context, then the sustainable flow of timber seems the best management objective.
ACKNOWLEDGEMENTS
The research on which this paper is based was made possible through the cooperative efforts of Drs. B. Arts, H. E Linskens, and J. E Dobremez, and Nepalese assist- ants. Dr. E Bongers, Dr. T. Brock, and Ir. K. E Wiersum provided useful comments on the drafts of this paper.
REFERENCES Alirol, P., 1979: Transhuming animal husbandry systems in the
Kalingchowk regions (Central Nepal). SATA. Kathmandu. Barbour, M. G. and Billings, W. D. (eds.), 1988: North American
Terrestrial Vegetation. Cambridge University Press, 434 pp. Barrett, S. W and Arno, S. E, 1982: Indian Fires As an Ecological
Influence In the Northern Rockies.Journal ofForestry, 80(10): 647-651.
Berthet-Bondet, J., 1983: Analyse du systime d'levage dans les collines prehimalayennes. Paris.
Brower, B., 1990: Range conservation and Sherpa livestock management in Khumbu, Nepal. Mountain Research and Devel- opment, 10(1): 34-42.
Dobremez, J. E, 1976: Le Nopal: Ecologie et Biogdographie. CNRS, Paris, pp. 356.
Mahat, T. B. S., Griffin, D. M., and Shepherd, K. R., 1987: Human impact on some forests in the Middle Hills of Nepal. Part 3. Forests in the subsistence economy of Sindhu Palchok and Kahbre Palanchok. Mountain Research and Development, 7(1): 53-70.
Metz, J. J., 1990: Conservation practices at an upper-elevation village of West Nepal. Mountain Research and Development, 10(1): 7-15.
Richard, D., 1980: Variations de la Structure, de l'Architecture et de la Biomass de Forets du Centre N6pal. Th6se, Universit6 de Grenoble, Grenoble, 162 pp.
Schmidt-Vogt, D., 1990a: High Altitude Forests in the Jugal Himal (Eastern Central Nepal). Forest types and human impact. Geoeco- logical research Vol. 6. Franz Steiner Verlag, Stuttgart.
, 1990b: Fire in High Altitude Forest of the Nepal Himalayas. In Goldammer, J. G. and Jenkins, M.J. (eds.), Fire in Ecosystem Dynamics. SPB Academic Publishing by., The Hague, pp. 191-199.
Sprugel, D. G., 1991: Disturbance, Equilibrium, and Environ- mental Variability: What is 'Natural' Vegetation in a Changing Environment? Biological Conservation, 58:1-18.
Sueron, C., 1985: Regeneration, Structures et Production d'une For&t "naturelle": la Sapiniere a Abies spectabilis (D.Don) Mirb. du N6pal Central Th6se, Universit6 de Grenoble, Grenoble. 62 pp.
Tande, G.E, 1979: Fire history and vegetation pattern of coniferous forests in Jasper National Park, Alberta. Canadian Journal of Botany 57: 1912-1931.
Westhoff, V., 1965: Plantengemeenschappen. In Uit de Planten- wereld. Palladium-reeks nr. 15, W de Haan, Zeist en Van Loghum Slaterus, Arnhem.
Wiart,J., 1983: Ecosysthme Villageois Traditionnel en Himalayas N6palais: la production forestiere suffit-elle aux besoins de la population? These, Universit6 de Grenoble, Grenoble, 267 pp.
Wright, H. E. and Heinselman, M. L., 1973: Introduction. Quaternary Research, 3: 319-328.
This content downloaded from 137.224.252.10 on Thu, 13 Mar 2014 03:53:25 AMAll use subject to JSTOR Terms and Conditions




















