
The Truthful Messenger: Visual Methods and Representation in Qualitative Research in Education
Upload
independentCategory
view
3download
0
www.elsevier.com/locate/ynimg
NeuroImage 39 (2008) 469–482The self in conflict: The role of executive processes during truthfuland deceptive responses about attitudes
Ray Johnson Jr.,a,b,⁎ Heather Henkell,b Elizabeth Simon,b and John Zhua
aDepartment of Psychology, Queens College/CUNY Flushing, NY 11367, USAbDepartment of Psychology, The Graduate Center/CUNY NY, NY 10016, USA
Received 3 April 2007; revised 5 July 2007; accepted 20 August 2007Available online 31 August 2007
This study sought to extend previous results regarding deceptionsabout specific memories by investigating the role of executive processesin deceptions about evaluative judgments. In addition, given thatprevious studies of deception have not included valence manipulations,we also wanted to determine whether the goodness/badness aspect ofthe items would affect the processes used during deception. Thus, wecompared behavioral and event-related potential (ERP) activity whileparticipants made truthful and directed lie (i.e., press opposite of thetruth) responses about attitude items with which they either stronglyagreed or disagreed. Consistent with previous results, deceptiveresponses required greater cognitive control as indicated by slowerRTs, larger medial frontal negativities (MFN) and smaller late positivecomponents than truthful responses. Furthermore, the magnitude ofthese deception-related effects was dependent on the valence thatparticipants assigned to the items (i.e., agree/disagree). Directed lieresponses about attitudes also resulted in greatly reduced pre-responsepositivities, an indication that participants strategically monitoredtheir responses even in the absence of explicit task demands. Itemvalence also differentially affected the amplitude of three ERPcomponents in a 650 ms pre-response interval, independently ofwhether truthful or deceptive responses were made. Analyses usingdipole locations based on results from fMRI studies of evaluativejudgments and deception indicated a high degree of overlap betweenthe ERP and fMRI results and revealed the possible temporalcharacteristics of the hemodynamic activations.© 2007 Elsevier Inc. All rights reserved.
Keywords: Deception; Executive processes; Attitudes; ERPs
Although the factors behind being deceptive may vary acrossdeceptions, from person to person and even from day to day, theneed to consciously select and execute a response that isincompatible with the truth is a necessary component of alldeceptions. Hence, by virtue of their role in controlling and
⁎ Corresponding author. Department of Psychology, Queens College/CUNY, 65-30 Kissena Blvd., Flushing, NY 11367, USA. Fax: +1 718 9973257.
E-mail address: [email protected] (R. Johnson).Available online on ScienceDirect (www.sciencedirect.com).
1053-8119/$ - see front matter © 2007 Elsevier Inc. All rights reserved.doi:10.1016/j.neuroimage.2007.08.032
coordinating the selection and execution of willed actions, theexecutive processes used to implement cognitive control (see Bushet al., 2000; van Veen and Carter, 2006 for reviews) play anessential role in deceptive responding. Such processes includethose that automatically and continuously monitor actions forconflicting response tendencies and errors (i.e., tactical monitoring)(e.g., Carter et al., 1998; Botvinick et al., 1999, 2004). Still othersare responsible for adjudicating the conflicting response tendenciesin favor of the deceptive response and inhibiting the pre-potenttruthful response and/or other unwanted responses. In addition, wehypothesized that at least some deceptions would also require theuse of longer-term (i.e., strategic) monitoring processes to ensurethat selected responses conform to one’s plans and goals (Johnsonet al., 2003, 2004). In this scheme, there is a hierarchy ofmonitoring processes that are affected by internal as well asexternal factors. In this way, a personTs goals affect the operation ofthe higher-order, strategic monitoring processes which, in turn,affect the operating context of tactical monitoring processes. Forexample, the goal of being deceptive alters the circumstances suchthat stimuli that may not normally create conflict will now do so.Also, unlike tactical monitoring processes, strategic monitoringprocesses are posited to operate at a conscious level and requireprocessing resources. Thus, the degree of cognitive control neededto compensate for this increased response conflict is greater fordeceptive responses than for truthful responses.
To characterize better the cognitive processes and brainmechanisms underlying deception, we conducted a series ofevent-related brain potential (ERP) experiments to assess therelative roles of tactical and strategic monitoring processes indeceptive responding (Johnson et al., 2003, 2004, 2005). Therefore,we included different conditions in which participants made either“directed” or “self-generated” lies. Directed lies, which wereintended to increase the need for tactical monitoring processes,required participants to press the button opposite of the truthfulresponse for each stimulus. To prevent participants from eliminatingthe response conflict for deceptive responses by surreptitiouslyreversing the stimulus–response assignments, we included catchstimuli randomly on 20% of the trials that required participants torespond truthfully. In support of the idea that deceptive responsesrequire greater cognitive control, the behavioral results showed that

470 R. Johnson Jr. et al. / NeuroImage 39 (2008) 469–482
these reaction times (RT) were significantly slower and less accuratethan those for truthful responses. In contrast self-generated lies wereintended to invoke the use of strategic monitoring processes. In thiscondition, although participants were free to choose whether tomake a truthful or deceptive response on any given trial, they wereinstructed to make their selections conform to the long-term goal ofmaking roughly equal numbers of truthful and deceptive responsesabout both the memorized (i.e., old) and non-memorized (i.e., new)items. The RTs for self-generated lies were much slower and morevariable, even when compared to the RTs for directed lies.
The ERP results revealed that deceptive responding produced acomplex pattern of altered brain activity with different spatio-temporal characteristics. For example, compared to truthfulresponses, deceptive responses elicited significantly larger medialfrontal negativities (MFN), with much larger increases for self-generated lies than for directed lies. This ERP component, whichpeaks at about 70 ms after a correct response, has been linked toresponse monitoring and the degree of response conflict created bya stimulus (Gehring and Knight, 2000; Vidal et al., 2000; Gehringand Willoughby, 2002; Mathalon et al., 2003; Johnson et al., 2004,2005). Based on results from a number of studies, investigatorshave concluded that the MFN is generated in or near the anteriorcingulate cortex (ACC) (Ullsperger and Von Cramon, 2001;Gehring and Willoughby, 2002; Johnson et al., 2004), a structureknown to play an important role in response monitoring andconflict detection (e.g., Bush et al., 2000; van Veen and Carter,2006). The MFNs elicited by the directed and self-generated lieswere each due to different patterns of brain activity, which wereboth different from that for truthful MFNs (Johnson et al., 2004).This finding is consistent with the reported multi-functional natureof the ACC (cf., Turken and Swick, 1999). The results also showedthat, in the 250–300 ms prior to the response, another medialfrontal maximal ERP component was elicited. In contrast to onlysmall reductions in the amplitude of this pre-response positivity(PRP) for directed lies, the PRP was virtually eliminated for self-generated lies, which required strategic monitoring to be successful(Johnson et al., 2004, 2005). The pre-response timing of the PRPalterations is important because it is compatible with the fact that,to be effective, strategic monitoring processes should precederesponse execution. Based on this pattern of results, wehypothesized that the PRP amplitude reductions result from thetemporally overlapping strategic monitoring processes.
The use of strategic monitoring processes and the upregulationof cognitive control required to deal with the increased conflictcreated by deceptive responses requires effort and thus additionalprocessing resources. In accord with findings showing that theamplitude of the parietal late positive component (LPC) of the ERP(also known as the P300) decreases in proportion to the amount ofresources allocated to secondary tasks (e.g., responding decep-tively) (see Johnson, 1986, 1988 for reviews), smaller LPCs wereelicited for deceptive responses than for truthful responses.Furthermore, the LPC amplitude decreases for the two types ofdeceptive responses mirrored the magnitude of the increases innegativity over medial frontal scalp (i.e., greater LPC decreases forself-generated lies than for directed lies) (Johnson et al., 2003,2005). Taken together, the results indicate that deception relies ongeneral purpose cognitive processes and thus they supportconclusions based on autonomic measures that there is no “specificlie response” (e.g., Ben-Shakhar and Furedy, 1990).
Consistent with the idea that deceptive responses require greateruse of executive processes, recent studies using functional
magnetic resonance imaging (fMRI) have shown that, along withtheir slower RTs (Spence et al., 2001; Ganis et al., 2003; Nunez etal., 2005; Abe et al., 2006), deceptive responses elicit greater bloodflow in the ACC than truthful responses (Langleben et al., 2002;Spence et al., 2001; Ganis et al., 2003; Kozel et al., 2004; Lee etal., 2002; Abe et al., 2006; Nunez et al., 2005). Recently, Nunezand colleagues (2005) showed that the magnitude of the behavioraland hemodynamic changes could be increased by increasing theimportance of the stimuli to the person. The deceptive responsesused in all these fMRI studies are roughly equivalent to ourdirected lies about episodic memories (i.e., participants maderesponses that were opposite of the truth) and thus, despite theirapparent simplicity, even directed lies are sufficient to producesignificant changes in behavioral and brain measures. In summary,despite differences in the nature and circumstances of thedeceptions studied to date, the similarity of the results, both acrossexperiments and brain imaging techniques, suggests that they arecapturing an essential aspect of many, if not all, deceptions.
The fact that the stimuli used in deception studies are inherentlyself-referential raises the possibility that there may be some overlapbetween the cognitive processes used during deception and thoseused to provide the basis of the “self” (e.g., self knowledge, self-referential decisions). Recent hemodynamic studies in whichparticipants judged whether particular trait adjectives describethem found activation in midline brain areas, including the anteriormedial frontal cortex, ACC and precuneus cortex (e.g., Gusnard etal., 2001; Kelley et al., 2002; Fossati et al., 2003). Similar midlinefrontal and posterior brain activations have been reported whenparticipants made evaluative judgments about food preferences(e.g., Seger et al., 2004), socially relevant concepts (e.g.,Cunningham et al., 2004) and during moral judgments (see Mollet al., 2005 for a review). Other studies have shown that some ofthese same brain areas are also activated during retrieval ofautobiographical memories (e.g., Craik et al., 1999) and in theoryof mind tasks (e.g., Schmitz et al., 2004). Such results have ledinvestigators to posit that these brain areas are part of a largernetwork underlying the cognitive and affective processes used toprovide the basis of self awareness and self concept (e.g., Northoffand Bermpohl, 2004; Amodio and Frith, 2006). Nevertheless,many aspects of the functioning of this network remain unspecified(cf., Gillihan and Farah, 2005).
The focus of the aforementioned deception studies was onspecifying the executive processes used to make deceptiveresponses about items in episodic memory. Similar results havebeen found for both autobiographical (Johnson et al., 2002a;Nunez et al., 2005) and personal semantic (i.e., factual) memories(Johnson et al., 2002b). However, although all these stimuli werebased on specific memory representations, we hypothesized thatthe same results would obtain if participants made deceptiveresponses about their attitudes. Unlike episodic memory retrieval, acentral aspect of attitude evaluations is the requirement to make anaffective judgment about the goodness/badness of the attitudeobject based on internal scales reflecting the individual’s personalvalue system. Such judgments are an important component ofeveryday life and are made frequently in order to determinewhether one approaches or avoids particular people, situations orobjects (cf., Cunningham et al., 2003, 2004). Given their centralrole in guiding decisions, choices and actions, we expected thatattitudes are likely to be perceived as being more important and/ormore central to the self than most individual episodic orautobiographical memories.

471R. Johnson Jr. et al. / NeuroImage 39 (2008) 469–482
One recent study investigated how the brain activity evokedduring attitude judgments differed from that evoked by other typesof evaluations. Zysset and colleagues (2002) compared fMRI scanswhile participants made agree/disagree judgments about a series ofshort sentences (e.g., I like Leipzig) with those obtained duringautobiographical (e.g., I have been to Leipzig) and semanticevaluations (e.g., Leipzig is a large city). Their results revealed thatthe same midline frontal and precuneus brain areas implicated inprocessing of the self were differentially active during these threedecisions. For example, whereas evaluative and autobiographicaldecisions activated anterior medial frontal cortex, semanticdecisions did not. Moreover, evaluative judgments showed greateranterior medial frontal activity than retrieval of autobiographicalmemories, with the opposite pattern evident for precuneusactivations. The authors interpreted the anterior medial frontalactivations as reflecting self-referential processing and theprecuneus activations as being more related to memory retrieval.Zysset and colleagues also found that left inferior prefrontal cortexwas active for semantic and evaluative judgments, a result assumedto reflect the use of selection processes (cf., Thompson-Schill et al.,1997).
Investigators have also used ERPs to study evaluativejudgments. In a series of studies, Cacioppo and colleagues workedtoward developing a direct indicator of a person’s attitudes thatwould bypass the potential confounds inherent in havingparticipants make overt like/dislike responses (Cacioppo andBernston, 1994; Cacioppo and Gardner, 1996; Crites andCacioppo, 1996; Ito et al., 1998). Hence, they used a paradigmin which positive and negative trait adjectives were presented incongruent or incongruent contexts. These contexts were created bypresenting the trait words in a series of words of the same oropposite valence. The results showed that both negative andpositive trait words elicited a larger LPC when presented withwords of the opposite valence than when they were embedded withwords of the same valence. Moreover, the size of the LPCamplitude increase was directly related to the magnitude of thediscrepancy between the participant’s view of the stimulus andtheir view of the contextual stimuli (Cacioppo and Bernston, 1994;Cacioppo and Gardner, 1996). The authors also reported that theLPC results were unaffected when participants made directed liesabout their attitudes (Crites et al., 1995; Cacioppo and Gardner,1996). However, in view of the fact that they did not report anyresults regarding truthful–deceptive comparisons, for either RT orLPC amplitude, it is impossible to compare their results with ours.More recently, Cunningham and colleagues (2005) studiedparticipants while they made good–bad judgments about sociallyrelevant concepts presented in a neutral context (i.e., a randomseries of positive and negative items). Measuring ERP activity overfrontal scalp, they found that good and bad stimuli elicited greateramounts of activity at left and right frontal scalp, respectively.They interpreted the lateness of their ERP effects (i.e., 475–1200 ms) as indicating that they reflected controlled processes usedduring valence evaluation. Given that none of these studiesmeasured ERP activity related to executive processes, their role inevaluative judgments remains unknown.
The present study sought to extend our results on deceptionsabout specific episodic and autobiographical memories to decep-tions about one’s attitudes. For the reasons mentioned above, wehypothesized that the evaluative nature of attitudes would producesimilar but more exaggerated changes in measures of behavioraland brain activity than deceptions about specific personal
memories. In addition, although previous deception studies havenot included valence manipulations, we expected that the perceivedgoodness/badness of items would not qualitatively affect theprocesses used during deception. Thus, we compared behavioraland brain activity when participants made truthful and directed lieresponses about attitude objects that they had rated as being eitherhighly positive or negative.
Methods
Participants
Seventeen participants (13 females) were recruited from theQueens College campus and paid US $10 per hour forparticipation. Data from one female were dropped because hercatch trial results indicated that she had reversed the stimulus–response assignments. The remaining 16 participants had a meanage of 22.6 years (S.D.=4.0; range=18–30) and a mean of15.9 years (S.D.=1.6) of education. This sample was ethnicallymixed, consisting of seven Caucasians, five African-Americans,two Asians and two Hispanics. All participants were right-handed,native English speakers, with normal or corrected-to-normalvision. Participants were pre-screened by asking them a series ofquestions, including if they had any history of neurologicaldisorders (e.g., head injury with loss of consciousness; history offainting or headaches), psychiatric and/or emotional problems(e.g., panic disorder, mood disorders) or drug abuse. Finally, theywere asked if they were currently taking any psychoactivemedications. Positive answers regarding any of these led to theirbeing excluded from the study. Participants were then briefed aboutthe nature of the experiment and informed consent was obtainedfrom each in accord with Queens College Institutional ReviewBoard procedures.
Stimuli
Approximately 1 week prior to the experimental session(mean=6.7 days, S.D.=2.4), participants were interviewed togather information about their attitudes by having them makeevaluative judgments, via a computerized questionnaire, about atotal of 118 items. These covered a wide variety of social, political,religious and moral issues (e.g., interracial marriage, birth control,censorship, abortion, murderers), as well as widely knownindividuals (e.g., Bill Clinton, Bin Laden, Rudy Giuliani) andpreferences (e.g., rap music, tattoos). As indicated by theseexamples, the concepts varied considerably in terms of theircomplexity, goodness/badness and concreteness. For each item,participants rated their attitudes on a scale of 1 (extremelyunfavorable/disagree) to 7 (extremely favorable/agree). The sameitems were also rated on both their importance and strength using ascale of 1 (not at all important/strongly) to 7 (extremely important/strongly) (Olson et al., 2001). Strength was defined as howstrongly the attitude is held and/or how much work it would take tochange your attitude. Importance was defined as how much thisattitude influences your life and/or your actions. Using theseratings, individualized stimulus files were created for eachparticipant. Because we sought to use strongly held attitudes,items with the highest disagree and agree ratings (1 and 7,respectively) and the highest strength ratings were used first. Wethen proceeded down the strength scale until we had 26–30 itemseach in the agree and disagree categories.

472 R. Johnson Jr. et al. / NeuroImage 39 (2008) 469–482
Procedure
After electrode placement, participants were seated in a dimlylit room. All stimuli were presented on a computer monitor for400 ms in all uppercase white letters on a black background, with arandomized inter-stimulus interval ranging from 2450 to 2850 ms.Stimuli with fewer than 12 letters were displayed on a single linewhile those with more than 10–12 letters were split into two linesin the center of the screen (i.e., one above the other). Prior to theexperimental blocks, participants were given a practice seriesinvolving a simple orthographic decision to familiarize them withthe procedures and reduce their RT.
Although the experiment included three types of stimuli(episodic, autobiographical, attitudes), only the attitude resultsare discussed here. Each stimulus type was presented under threeinstructional conditions: Truthful, Directed Lie and Semantic(presented elsewhere). In the Truthful condition, participants wereinstructed to press one button for items with which they agreed/feltfavorable toward and the other button for items with which theydisagreed/felt unfavorable toward. For neutral items, participantswere instructed to press according to how they were leaning. In theDirected Lie condition, although participants were instructedinitially to make the same responses as in the Truthful condition,they were also told to lie or try to hide their true attitudes bypressing opposite of the instructions on all trials except the catchtrials (see below). All conditions were given in a balanced order sothat half the participants performed the truthful condition beforethe deceptive condition and half performed the deceptive conditionfirst. The stimulus–response button assignments were counter-balanced across participants.
Catch trials
Because participants were instructed to press opposite of thetruth for every stimulus in the Directed Lie condition, there wouldbe nothing to prevent them from simply reversing the stimulus–response assignments in order to eliminate the conflicting responseinformation. To prevent the use of this strategy, catch trialsconsisting of the words “AGREE” and “DISAGREE” werepresented randomly on 17% of the trials in both the truthful anddeceptive conditions (90 attitude items, 18 catch). Participantswere instructed that these trials were included “to keep you honest”and “that it is important that you try hard always to press correctlyon the catch trials so it will not be obvious that you are pressing theopposite buttons on the other trials.” Note that the use of catchtrials meant that participants needed to make truthful responses on17% of the trials in the Directed Lie condition.
ERP recordings and quantification
ERP activity was recorded from 32 scalp sites, all referred to aleft pre-auricular electrode, using tin electrodes embedded in anelasticized cap. The sites (Fp1, Fp2, F7, F3, Fz, F4, F8, FC5, FC1,FC2, FC6, T7, C3, Cz, C4, T8, CP5, CP1, CP2, CP6, P7, P3, Pz,P4, P8, O1, O2, Cb1, Cb2, right pre-auricular, E1, E2) were locatedin accord with the American Electroencephalographic Society(1991) guidelines. To reduce the presence of possible electomyo-graphic artifacts, the Cb1 and Cb2 electrodes were placed 1.2 cmabove their standard International 10–20 System locations.Participants were grounded with a forehead electrode (FPz). TheEEG was amplified 10,000 times with a bandpass of 0.01–35 Hz
(−3 dB/octave) and sampled at 100 Hz. Eye movements (EOG)were recorded from above (FP1) and 2 cm below the outer canthusof the left eye (E1) and trials contaminated with EOG artifacts(signals greater than 50 μV during any 6 sampling points) wereexcluded from the averages. During averaging, all scalp-recordedactivity was digitally re-referenced to an average of the left andright pre-auricular sites. Response-locked ERPs were calculated foran 1450 ms epoch, extending from 1150 ms before the responseuntil 300 ms after the response. The use of an extended pre-response interval (i.e., one that roughly equaled the mean RTsfound here) was intended to ensure that the baseline interval(−1150 to −1000 ms) was, on average, around the time of stimulusonset when there is essentially no stimulus evoked brain activity.Therefore, these response-locked averages provide a neutralbaseline with which to measure the amplitudes of the ERPcomponents.
The behavioral and ERP component amplitudes were analyzedin a series of repeated-measures ANOVAs using the factorsResponse (Truthful, Deceptive), Valence (Agree, Disagree) andElectrode, separately for each ERP component. ANOVAs onbehavioral results had the same designs without the electrodefactor. Because using data from multiple electrode sites may lead toa violation of the sphericity assumption, all ANOVA results werecorrected using the Greenhouse–Geisser procedure. The uncor-rected degrees of freedom are presented along with the corrected pvalues and epsilons (ε). Given that we hypothesized that alldeceptive responses require increased use of executive processes,regardless of item valence, the ANOVAs were followed by plannedcomparisons designed to determine if there were amplitudedifferences between truthful and deceptive responses about bothagree and disagree items for each component. Partial eta-squared(ηp2) is reported as an index of effect size.
Dipole analyses
To determine whether the scalp-recorded activity might berelated to the brain activations reported in fMRI studies dipolesources were seeded using the coordinates of activated brain areasin hemodynamic studies (see Results for specific details). Thesedipoles were fixed within the brain space and the orientation andstrength of the dipoles were allowed to vary using the BrainElectrical Source Analysis program (BESA 5.08; Megis GmbH,Munich, Germany; Scherg, 1990). The program uses a four-shell,ellipsoid head model with the relative conductivities set to 0.330for brain, 0.330 for scalp, 0.042 for skull and 1.00 for cerebrospinalfluid. Scalp and skull thickness were set to 6 and 7 mm,respectively. To evaluate the solutions, the residual variance (RV),which provides an estimate of the amount of ERP power notexplained by the seeded dipoles, was calculated by comparing thesquared total error to the squared data (data power). These analyseswere performed on the grand averages after they were algebraicallyre-referenced to an average reference.
Results
The across-participant mean evaluation, strength and impor-tance ratings for the agree, disagree and neutral stimulus categoriesare shown in Table 1. These data show that the procedure used tocreate the stimulus files was successful in creating lists of agreeand disagree items that were rated significantly differently fromone another [t(15)=40.45, pb .0001]. Importantly, mean impor-

Table 1Mean (S.D.) of evaluation, strength and importance ratings for the agree,disagree and neutral stimulus categories
Evaluation Strength Importance
Agree 6.50 (0.35) 5.63 (1.06) 5.37 (1.09)Disagree 1.23 (0.24) 6.03 (0.68) 5.68 (0.90)Neutral 4.18 (0.60) 2.82 (0.95) 2.59 (0.94)
All ratings were collected at the same time using a 7-point scale(1=disagree, not at all important/strongly to 7=agree, extremely impor-tant/strongly). See text for further details on how the agree, disagree andneutral stimuli were created.
473R. Johnson Jr. et al. / NeuroImage 39 (2008) 469–482
tance ratings for the agree and disagree categories were comparable(p=.15) and, although the difference in strength ratings for theagree and disagree items was borderline significant [t(15)=2.07,p=.06], it is evident that the overall mean difference of 0.40 wasquite small. Finally, of the remaining items, the 30 rated in themiddle of the scale were considered neutral. As can be seen inTable 1, these neutral items were also rated much lower on strengthand importance. Comparing neutral items with agree and disagreeitems is problematic because they can elicit either no affectiveresponse or an ambiguous response containing both positive andnegative feelings. Therefore, in the absence of a method todetermine the true nature of the processing engendered by anygiven neutral item, these data were not included in the analyses.
Table 2Mean (SD) reaction time as a function of condition for the agree and disagreestimuli
Truthful Directed lie
Agree 948 (250) 1162 (268)Disagree 987 (232) 1152 (243)
Behavioral results
Overall, deceptive responses were less accurate than truthfulresponses (86.2% vs. 93.1%, respectively) [F(1,15)=4.3, p=.06,ηp2= .22]. Accuracy was also affected by valence, with slightlyhigher accuracy rates for agree (91.4%) than disagree items(88.0%) [F(1,15)=8.0, pb .02, ηp2= .35]. This difference meantthat the Response×Valence interaction approached significance[F(1,15)=3.5, p=.08, ηp2= .19].
The RT results are shown in Table 2. As expected, the increasedresponse conflict for deceptive responses led to their being almost200 ms slower than truthful responses (1157 ms vs. 968 ms) [F(1,15)=21.4, pb .0005, ηp2= .59]. Although valence did not affectprocessing time (1055 vs. 1070 ms for agree and disagree items,respectively) (Fb1), there was a trend in which the cost of makinga deceptive response (deceptive RT− truthful RT) was greater foragree than disagree items (214 ms vs. 165 ms, respectively)(Response×Valence: F(1,15)=3.2, p=.10, ηp2= .18). Taken to-gether, the accuracy and RT data are consistent with the presenceof greater response conflict for deceptive compared to truthfulresponses, with slightly more conflict for deceptive responsesabout agree items.
ERP results
Medial frontal measures of executive functionThe ERPs recorded over medial frontal scalp by truthful and
deceptive responses, shown in Fig. 1, indicate that response typeand valence had different effects on brain activity in the intervalaround the response (i.e., from −400 ms to +100 ms). The activityof two ERP components in this interval (i.e., pre-responsepositivity, medial frontal negativity) will be discussed, beginningwith the better characterized MFN potential.
An ANOVA on MFN areas between 10 and 80 ms post-response (electrodes: Fz, FC1, FC2, Cz) confirmed that largerMFNs were elicited by deceptive responses than by truthfulresponses [F(1,15)=27.8, pb .0001, ηp2= .65] (Fig. 1). There wasalso a significant effect of valence because agree items elicitedlarger MFNs than disagree items [F(1,15)=4.6, pb .05, ηp2= .24],although the Response×Valence interaction was not significant[F(1,15)=2.0, p=.18, ηp2= .12]. Potential maps (Fig. 2) show thatthe MFN was maximal over left medial frontal scalp (i.e., FC1 site)[Electrode: F(5,75)=6.6, ε=.653, pb .001, ηp2= .31]. Plannedcomparisons confirmed that deceptive responses produced largerMFNs than truthful responses for both agree [F(1,15)=11.8,pb .005, ηp2= .44] and disagree items [F(1,15)=7.9, pb .02,ηp2= .35]. Post hoc tests confirmed that agree items elicited largerMFNs than disagree items for deceptive responses [F(1,15)=9.6,pb .01, ηp2= .39] and that there were no differences for truthfulresponses (Fb1).
Both response type and valence also affected the PRPcomponent elicited in the 400 ms preceding the response (Fig.1). As can be seen in the maps in Fig. 2, the PRP was also maximalover medial frontal scalp, although the focus of this distributionwas slightly more left lateralized than the MFN distribution. ThePRP was significantly less positive between −100 and 0 ms fordeceptive responses compared to truthful responses, whichappeared to be due to the presence of an overlapping negativity[F(1,15)=28.4, pb .0001, ηp2= .66]. Valence affected PRP ampli-tude because agree items elicited smaller positivities than disagreeitems [F(1,15)=5.7, pb .05, ηp2= .28], to the same extent fortruthful and deceptive responses (Response×Valence: Fb1). Theseresults suggest that the response and valence variables each hadindependent and additive effects on PRP amplitude. Plannedcomparisons confirmed that deceptive responses were associatedwith smaller PRPs than truthful responses for both agree [F(1,15)=10.3, pb .01, ηp2 = .41] and disagree stimuli [F(1,15)=18.2,pb .001, ηp2= .55].
LPC measure of workload
As reviewed above, previous studies have shown that LPCamplitude decreases as processing resources are allocated tosecondary or concomitant tasks. As evident from the waveforms(Fig. 3) and potential maps (Fig. 2), this relation was upheld herebecause LPC amplitude varied inversely with MFN amplitude. AnANOVA on the waveform areas (−100 ms to +100 ms) over parietalscalp (electrodes: P3, Pz, P4, CP1, CP2) confirmed that deceptiveresponses elicited smaller LPCs than truthful responses [F(1,15)=25.7, pb .0002, ηp2= .63]. Also in accord with the MFN findings,valence affected LPC amplitude, with smaller LPCs elicited by agreeitems than by disagree items [F(1,15)=7.1, pb .02, ηp2= .32]. Ascan be seen in Fig. 3, although the Response×Valence interactionwas not significant (Fb1), smaller LPCs were elicited by agreeitems than by disagree items when deceptive responses were made
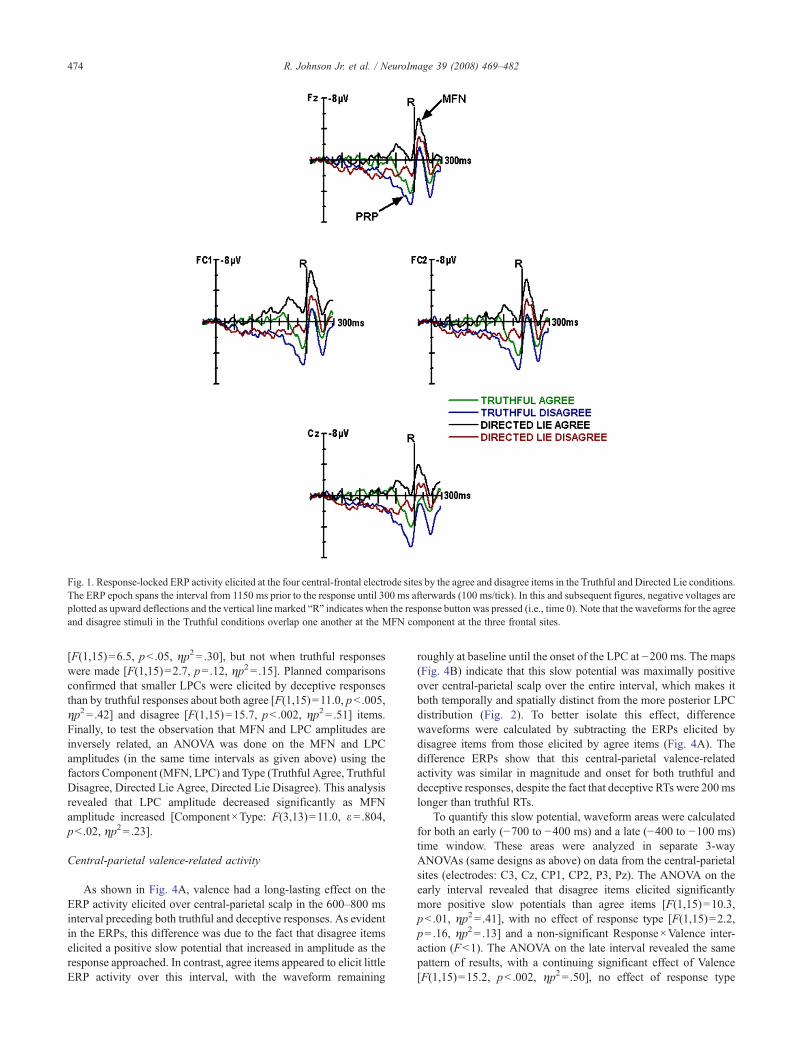
Fig. 1. Response-locked ERP activity elicited at the four central-frontal electrode sites by the agree and disagree items in the Truthful and Directed Lie conditions.The ERP epoch spans the interval from 1150 ms prior to the response until 300 ms afterwards (100 ms/tick). In this and subsequent figures, negative voltages areplotted as upward deflections and the vertical line marked “R” indicates when the response button was pressed (i.e., time 0). Note that the waveforms for the agreeand disagree stimuli in the Truthful conditions overlap one another at the MFN component at the three frontal sites.
474 R. Johnson Jr. et al. / NeuroImage 39 (2008) 469–482
[F(1,15)=6.5, pb .05, ηp2= .30], but not when truthful responseswere made [F(1,15)=2.7, p=.12, ηp2= .15]. Planned comparisonsconfirmed that smaller LPCs were elicited by deceptive responsesthan by truthful responses about both agree [F(1,15)=11.0, pb .005,ηp2= .42] and disagree [F(1,15)=15.7, pb .002, ηp2= .51] items.Finally, to test the observation that MFN and LPC amplitudes areinversely related, an ANOVA was done on the MFN and LPCamplitudes (in the same time intervals as given above) using thefactors Component (MFN, LPC) and Type (Truthful Agree, TruthfulDisagree, Directed Lie Agree, Directed Lie Disagree). This analysisrevealed that LPC amplitude decreased significantly as MFNamplitude increased [Component×Type: F(3,13)=11.0, ε=.804,pb .02, ηp2= .23].
Central-parietal valence-related activity
As shown in Fig. 4A, valence had a long-lasting effect on theERP activity elicited over central-parietal scalp in the 600–800 msinterval preceding both truthful and deceptive responses. As evidentin the ERPs, this difference was due to the fact that disagree itemselicited a positive slow potential that increased in amplitude as theresponse approached. In contrast, agree items appeared to elicit littleERP activity over this interval, with the waveform remaining
roughly at baseline until the onset of the LPC at −200 ms. The maps(Fig. 4B) indicate that this slow potential was maximally positiveover central-parietal scalp over the entire interval, which makes itboth temporally and spatially distinct from the more posterior LPCdistribution (Fig. 2). To better isolate this effect, differencewaveforms were calculated by subtracting the ERPs elicited bydisagree items from those elicited by agree items (Fig. 4A). Thedifference ERPs show that this central-parietal valence-relatedactivity was similar in magnitude and onset for both truthful anddeceptive responses, despite the fact that deceptive RTs were 200mslonger than truthful RTs.
To quantify this slow potential, waveform areas were calculatedfor both an early (−700 to −400 ms) and a late (−400 to −100 ms)time window. These areas were analyzed in separate 3-wayANOVAs (same designs as above) on data from the central-parietalsites (electrodes: C3, Cz, CP1, CP2, P3, Pz). The ANOVA on theearly interval revealed that disagree items elicited significantlymore positive slow potentials than agree items [F(1,15)=10.3,pb .01, ηp2= .41], with no effect of response type [F(1,15)=2.2,p=.16, ηp2= .13] and a non-significant Response×Valence inter-action (Fb1). The ANOVA on the late interval revealed the samepattern of results, with a continuing significant effect of Valence[F(1,15)=15.2, pb .002, ηp2= .50], no effect of response type

Fig. 2. Voltage maps showing the whole brain activity for Truthful (top row) and Directed Lie (bottom row) responses about agree items. The maps show brainactivity from 490 ms before to +90 ms after participant's responses (110° projections; front of head at top). Contour lines are spaced at intervals of 0.6 μV. Themaps were calculated on the basis of the across-participant averages, after re-referencing to an average reference.
475R. Johnson Jr. et al. / NeuroImage 39 (2008) 469–482
(Fb1) and a non-significant Response×Valence interaction (Fb1).Hence, this valence effect occurred relatively late after stimulusonset (i.e., at least 350 ms) and it was unaffected by whetherparticipants responded truthfully or deceptively about theirattitudes.
To determine if this slow potential activity might be due tosome factor other than valence, the single trials were alsoaveraged according to the strength and importance of theparticipant’s attitudes (Table 1). Separate averages were createdfor items rated low (1 to 3) and high (5 to 7) on these two scales.However, due to the fact that the attitude items were originallychosen to be the most strongly held and most important items(see Methods), some participants had too few low-strength and/orlow-importance trials (i.e., fewer than 12) to calculate ERPs.Thus, the strength and importance grand averages were based on
Fig. 3. Response-locked averages showing the LPCs elicited at the midlineparietal site (Pz) by the agree and disagree items in the Truthful and DirectedLie conditions.
12 and 14 participants, respectively. ANOVAs were donecomparing the ERP activity elicited in the low- and high-ratingaverages using the same temporal intervals and electrode sites asin the above analyses. These tests revealed no significant effectsof either attitude strength (psN .30) or importance (psN .56) ineither temporal window. Thus, the central-parietal valence effectcannot be attributed to differences in attitude strength or impor-tance. We also note that, when the same analyses were done onthe MFN, the tests showed no differences in MFN amplitude as afunction of either attitude strength (p= .48) or importance(p=.88).
Occipital response- and valence-related activity
As shown in Fig. 5, another slow potential, located over striateand extrastriate brain areas, was present from −600 to −100 msbefore the response. This slow potential had a negative polarity andfor most of its duration its amplitude was affected by the nature ofthe subsequent response, but not valence. An ANOVA on thewaveform areas in this interval at four occipital sites (O1, O2, CB1,CB2) revealed that truthful responses elicited larger negativitiesthan deceptive responses [F(1,15)=14.0, pb .002, ηp2 = .48],whereas valence had no effect [F(1,15)=1.3, p=.28, ηp2= .08].Electrode produced a significant effect due to the fact that thesepotentials were larger over the right hemisphere [F(3,45)=5.4,ε=.654, pb .05, ηp2= .27].
Close inspection of the ERPs in Fig. 5 indicates that valencehad an effect over primary occipital scalp (O1, O2) in a brief timewindow because agree items elicited larger negativities thandisagree items between −400 and −100 ms before all responses.An ANOVA on the amplitudes in this interval, using only the O1and O2 sites, revealed that disagree items elicited significantlysmaller negativities than agree items [F(1,15)=4.7, pb .05,ηp2= .24] and that this valence effect did not differ as a functionof response (Fb1).
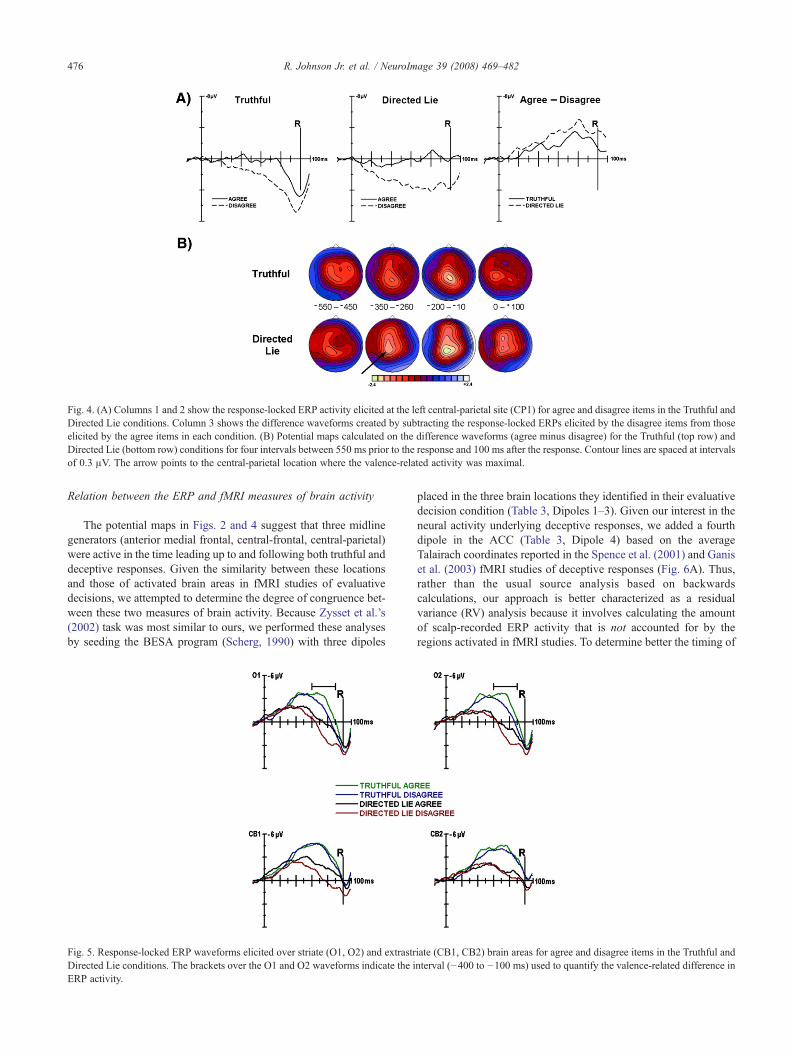
Fig. 4. (A) Columns 1 and 2 show the response-locked ERP activity elicited at the left central-parietal site (CP1) for agree and disagree items in the Truthful andDirected Lie conditions. Column 3 shows the difference waveforms created by subtracting the response-locked ERPs elicited by the disagree items from thoseelicited by the agree items in each condition. (B) Potential maps calculated on the difference waveforms (agree minus disagree) for the Truthful (top row) andDirected Lie (bottom row) conditions for four intervals between 550 ms prior to the response and 100 ms after the response. Contour lines are spaced at intervalsof 0.3 μV. The arrow points to the central-parietal location where the valence-related activity was maximal.
476 R. Johnson Jr. et al. / NeuroImage 39 (2008) 469–482
Relation between the ERP and fMRI measures of brain activity
The potential maps in Figs. 2 and 4 suggest that three midlinegenerators (anterior medial frontal, central-frontal, central-parietal)were active in the time leading up to and following both truthful anddeceptive responses. Given the similarity between these locationsand those of activated brain areas in fMRI studies of evaluativedecisions, we attempted to determine the degree of congruence bet-ween these two measures of brain activity. Because Zysset et al.’s(2002) task was most similar to ours, we performed these analysesby seeding the BESA program (Scherg, 1990) with three dipoles
Fig. 5. Response-locked ERP waveforms elicited over striate (O1, O2) and extrastrDirected Lie conditions. The brackets over the O1 and O2 waveforms indicate the iERP activity.
placed in the three brain locations they identified in their evaluativedecision condition (Table 3, Dipoles 1–3). Given our interest in theneural activity underlying deceptive responses, we added a fourthdipole in the ACC (Table 3, Dipole 4) based on the averageTalairach coordinates reported in the Spence et al. (2001) and Ganiset al. (2003) fMRI studies of deceptive responses (Fig. 6A). Thus,rather than the usual source analysis based on backwardscalculations, our approach is better characterized as a residualvariance (RV) analysis because it involves calculating the amountof scalp-recorded ERP activity that is not accounted for by theregions activated in fMRI studies. To determine better the timing of
iate (CB1, CB2) brain areas for agree and disagree items in the Truthful andnterval (−400 to −100 ms) used to quantify the valence-related difference in
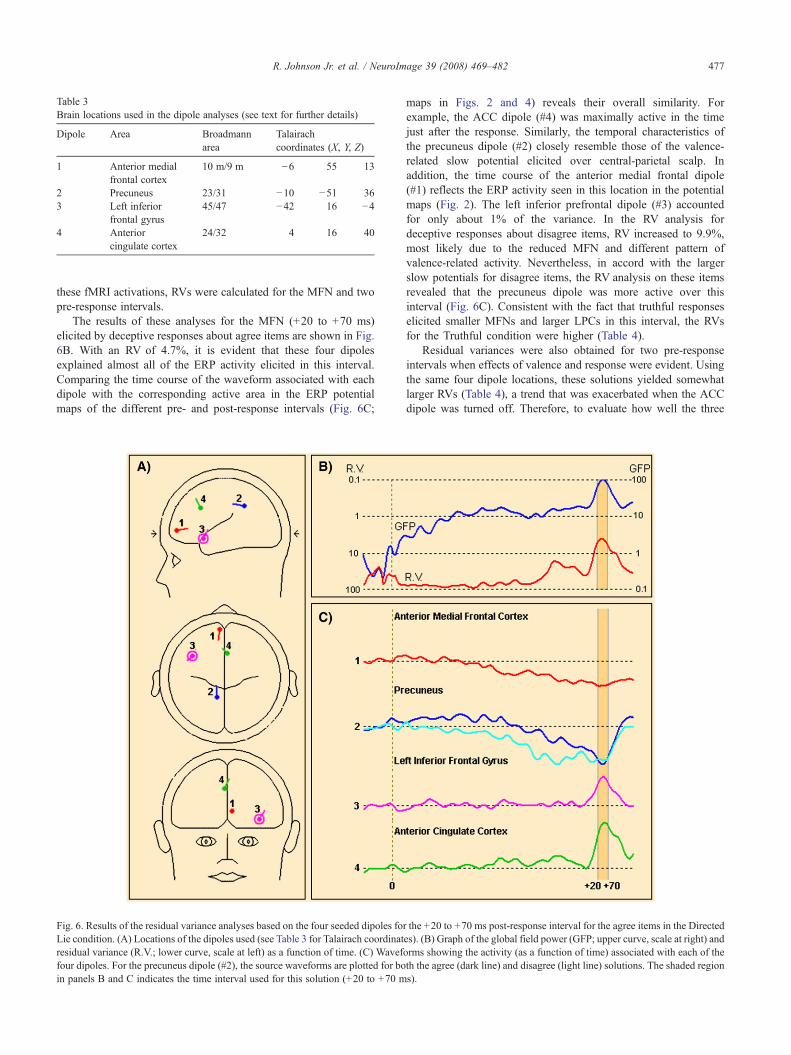
Table 3Brain locations used in the dipole analyses (see text for further details)
Dipole Area Broadmannarea
Talairachcoordinates (X, Y, Z)
1 Anterior medialfrontal cortex
10 m/9 m −6 55 13
2 Precuneus 23/31 −10 −51 363 Left inferior
frontal gyrus45/47 −42 16 −4
4 Anteriorcingulate cortex
24/32 4 16 40
477R. Johnson Jr. et al. / NeuroImage 39 (2008) 469–482
these fMRI activations, RVs were calculated for the MFN and twopre-response intervals.
The results of these analyses for the MFN (+20 to +70 ms)elicited by deceptive responses about agree items are shown in Fig.6B. With an RV of 4.7%, it is evident that these four dipolesexplained almost all of the ERP activity elicited in this interval.Comparing the time course of the waveform associated with eachdipole with the corresponding active area in the ERP potentialmaps of the different pre- and post-response intervals (Fig. 6C;
Fig. 6. Results of the residual variance analyses based on the four seeded dipoles forLie condition. (A) Locations of the dipoles used (see Table 3 for Talairach coordinatresidual variance (R.V.; lower curve, scale at left) as a function of time. (C) Waveffour dipoles. For the precuneus dipole (#2), the source waveforms are plotted for boin panels B and C indicates the time interval used for this solution (+20 to +70 m
maps in Figs. 2 and 4) reveals their overall similarity. Forexample, the ACC dipole (#4) was maximally active in the timejust after the response. Similarly, the temporal characteristics ofthe precuneus dipole (#2) closely resemble those of the valence-related slow potential elicited over central-parietal scalp. Inaddition, the time course of the anterior medial frontal dipole(#1) reflects the ERP activity seen in this location in the potentialmaps (Fig. 2). The left inferior prefrontal dipole (#3) accountedfor only about 1% of the variance. In the RV analysis fordeceptive responses about disagree items, RV increased to 9.9%,most likely due to the reduced MFN and different pattern ofvalence-related activity. Nevertheless, in accord with the largerslow potentials for disagree items, the RV analysis on these itemsrevealed that the precuneus dipole was more active over thisinterval (Fig. 6C). Consistent with the fact that truthful responseselicited smaller MFNs and larger LPCs in this interval, the RVsfor the Truthful condition were higher (Table 4).
Residual variances were also obtained for two pre-responseintervals when effects of valence and response were evident. Usingthe same four dipole locations, these solutions yielded somewhatlarger RVs (Table 4), a trend that was exacerbated when the ACCdipole was turned off. Therefore, to evaluate how well the three
the +20 to +70 ms post-response interval for the agree items in the Directedes). (B) Graph of the global field power (GFP; upper curve, scale at right) andorms showing the activity (as a function of time) associated with each of theth the agree (dark line) and disagree (light line) solutions. The shaded regions).
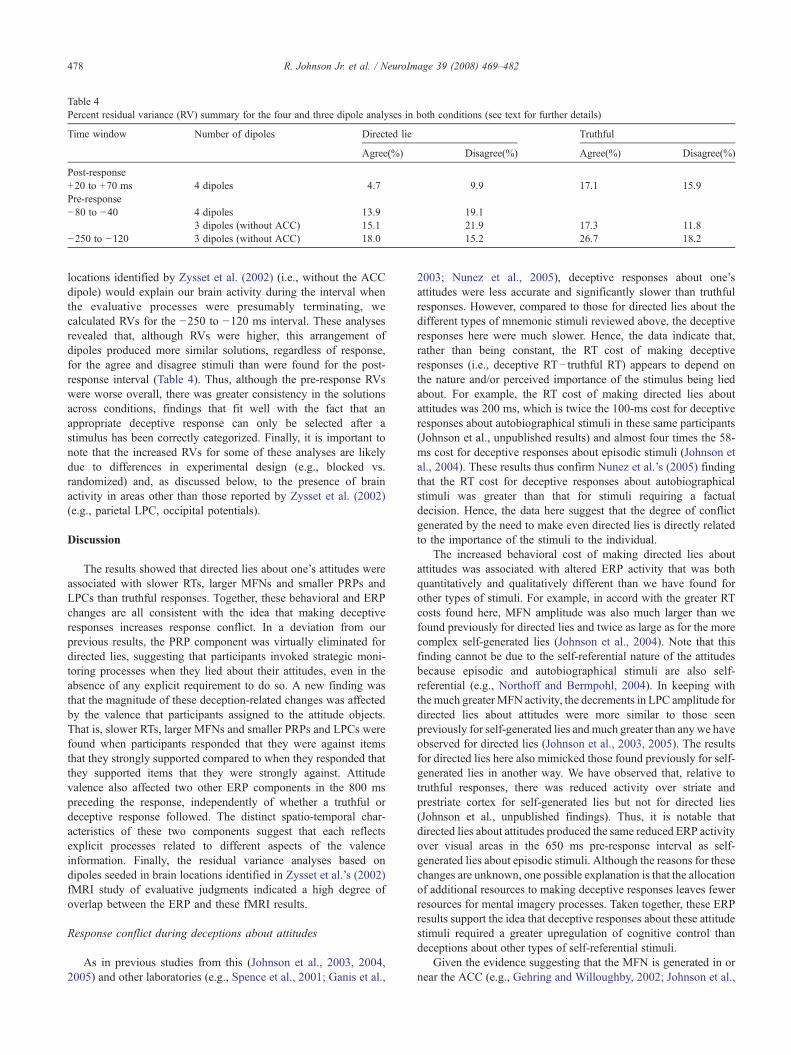
Table 4Percent residual variance (RV) summary for the four and three dipole analyses in both conditions (see text for further details)
Time window Number of dipoles Directed lie Truthful
Agree(%) Disagree(%) Agree(%) Disagree(%)
Post-response+20 to +70 ms 4 dipoles 4.7 9.9 17.1 15.9Pre-response−80 to −40 4 dipoles 13.9 19.1
3 dipoles (without ACC) 15.1 21.9 17.3 11.8−250 to −120 3 dipoles (without ACC) 18.0 15.2 26.7 18.2
478 R. Johnson Jr. et al. / NeuroImage 39 (2008) 469–482
locations identified by Zysset et al. (2002) (i.e., without the ACCdipole) would explain our brain activity during the interval whenthe evaluative processes were presumably terminating, wecalculated RVs for the −250 to −120 ms interval. These analysesrevealed that, although RVs were higher, this arrangement ofdipoles produced more similar solutions, regardless of response,for the agree and disagree stimuli than were found for the post-response interval (Table 4). Thus, although the pre-response RVswere worse overall, there was greater consistency in the solutionsacross conditions, findings that fit well with the fact that anappropriate deceptive response can only be selected after astimulus has been correctly categorized. Finally, it is important tonote that the increased RVs for some of these analyses are likelydue to differences in experimental design (e.g., blocked vs.randomized) and, as discussed below, to the presence of brainactivity in areas other than those reported by Zysset et al. (2002)(e.g., parietal LPC, occipital potentials).
Discussion
The results showed that directed lies about one’s attitudes wereassociated with slower RTs, larger MFNs and smaller PRPs andLPCs than truthful responses. Together, these behavioral and ERPchanges are all consistent with the idea that making deceptiveresponses increases response conflict. In a deviation from ourprevious results, the PRP component was virtually eliminated fordirected lies, suggesting that participants invoked strategic moni-toring processes when they lied about their attitudes, even in theabsence of any explicit requirement to do so. A new finding wasthat the magnitude of these deception-related changes was affectedby the valence that participants assigned to the attitude objects.That is, slower RTs, larger MFNs and smaller PRPs and LPCs werefound when participants responded that they were against itemsthat they strongly supported compared to when they responded thatthey supported items that they were strongly against. Attitudevalence also affected two other ERP components in the 800 mspreceding the response, independently of whether a truthful ordeceptive response followed. The distinct spatio-temporal char-acteristics of these two components suggest that each reflectsexplicit processes related to different aspects of the valenceinformation. Finally, the residual variance analyses based ondipoles seeded in brain locations identified in Zysset et al.’s (2002)fMRI study of evaluative judgments indicated a high degree ofoverlap between the ERP and these fMRI results.
Response conflict during deceptions about attitudes
As in previous studies from this (Johnson et al., 2003, 2004,2005) and other laboratories (e.g., Spence et al., 2001; Ganis et al.,
2003; Nunez et al., 2005), deceptive responses about one’sattitudes were less accurate and significantly slower than truthfulresponses. However, compared to those for directed lies about thedifferent types of mnemonic stimuli reviewed above, the deceptiveresponses here were much slower. Hence, the data indicate that,rather than being constant, the RT cost of making deceptiveresponses (i.e., deceptive RT− truthful RT) appears to depend onthe nature and/or perceived importance of the stimulus being liedabout. For example, the RT cost of making directed lies aboutattitudes was 200 ms, which is twice the 100-ms cost for deceptiveresponses about autobiographical stimuli in these same participants(Johnson et al., unpublished results) and almost four times the 58-ms cost for deceptive responses about episodic stimuli (Johnson etal., 2004). These results thus confirm Nunez et al.’s (2005) findingthat the RT cost for deceptive responses about autobiographicalstimuli was greater than that for stimuli requiring a factualdecision. Hence, the data here suggest that the degree of conflictgenerated by the need to make even directed lies is directly relatedto the importance of the stimuli to the individual.
The increased behavioral cost of making directed lies aboutattitudes was associated with altered ERP activity that was bothquantitatively and qualitatively different than we have found forother types of stimuli. For example, in accord with the greater RTcosts found here, MFN amplitude was also much larger than wefound previously for directed lies and twice as large as for the morecomplex self-generated lies (Johnson et al., 2004). Note that thisfinding cannot be due to the self-referential nature of the attitudesbecause episodic and autobiographical stimuli are also self-referential (e.g., Northoff and Bermpohl, 2004). In keeping withthe much greaterMFN activity, the decrements in LPC amplitude fordirected lies about attitudes were more similar to those seenpreviously for self-generated lies and much greater than any we haveobserved for directed lies (Johnson et al., 2003, 2005). The resultsfor directed lies here also mimicked those found previously for self-generated lies in another way. We have observed that, relative totruthful responses, there was reduced activity over striate andprestriate cortex for self-generated lies but not for directed lies(Johnson et al., unpublished findings). Thus, it is notable thatdirected lies about attitudes produced the same reduced ERP activityover visual areas in the 650 ms pre-response interval as self-generated lies about episodic stimuli. Although the reasons for thesechanges are unknown, one possible explanation is that the allocationof additional resources to making deceptive responses leaves fewerresources for mental imagery processes. Taken together, these ERPresults support the idea that deceptive responses about these attitudestimuli required a greater upregulation of cognitive control thandeceptions about other types of self-referential stimuli.
Given the evidence suggesting that the MFN is generated in ornear the ACC (e.g., Gehring and Willoughby, 2002; Johnson et al.,

479R. Johnson Jr. et al. / NeuroImage 39 (2008) 469–482
2004; Ullsperger and Von Cramon, 2001), the present results fitwell with those of fMRI studies showing that the ACC is involvedin conflict detection and resolution (e.g., Barch et al., 2000;Botvinick et al., 1999; Bush et al., 2000; Carter et al., 1998, 2000).Similarly, the results here are consistent with those from fMRIstudies of deceptive responses showing increased ACC activation(Langleben et al., 2002; Spence et al., 2001; Ganis et al., 2003;Kozel et al., 2004; Lee et al., 2002; Abe et al., 2006; Nunez et al.,2005). The evidence indicating that the MFN is generated in theACC was bolstered here by our RV analysis using an ACC dipolewhose location was derived from the Spence et al. (2001) andGanis et al. (2003) fMRI studies. This dipole produced a waveformwith a time course very much like that of the MFN, which supportsthe studies showing that the MFN is generated in the ACC andsuggests that the ACC activations observed in hemodynamicstudies of deception may be occurring within a several hundredmillisecond interval surrounding the response.
Valence affects on response conflict
An unexpected finding here was that, unlike the case for the pre-potent truthful responses, attitude valence affected the degree ofconflict created by deceptive responses. Although valence has notbeen manipulated previously, the behavioral and ERP data here allsupport the idea that deceptive responses about agree items producedmore conflict than those about disagree items. That is, larger MFNsand smaller LPCs were elicited when participants responded thatthey disagreed with items with which they actually agreed comparedto when they responded that they agreed with items with which theydisagreed. In addition, there was a trend towards greater RTcosts fordeceptive responses about agree items than for disagree items. Thisvalence effect does not appear to be attributable to any fundamentaldifferences between the items used in the agree and disagreecategories, which were closely matched on the basis of eachparticipants’s strength and importance ratings. Indeed, the lack ofvalence-related MFN or LPC amplitude differences for truthfulresponses about the same items argues strongly against the idea thatvalence inherently affects the levels of response conflict duringevaluative judgments. Thus, all three behavioral and ERP measurespoint to there being a differential need to increase cognitive controlas a function of attitude valence, but only for deceptive responses.
The greater conflict created by having to make directed liesabout positively viewed items raises the possibility that makingthis type of overt response may represent a kind of “denial of self”reaction. Conversely, the lesser degree of conflict created bydeceptive responses about items that one is against may representan instance of compliance. Compliance is a well-studiedphenomenon in which people alter their responses to reducepotential interpersonal conflict in a variety of every day socialsituations (e.g., Cialdini and Goldstein, 2004). Given thatcompliance is a relatively common and adaptive aspect of socialinteractions, it may reduce the conflict associated with makingdeceptive responses of this type, as well as the ease with witchsuch responses can be made. Nevertheless, this explanation mustremain tentative until additional supporting data become available.
Strategic monitoring and attitude evaluation
Another result suggesting that deceptions about the attitudestimuli are fundamentally different from those about episodic/autobiographical stimuli was the extent of the reductions in PRP
amplitude. The magnitude of these changes was unexpectedbecause, across a variety of self-referential stimuli, directed lies inour previous studies only produced small decrements in PRPamplitude. In contrast, the PRP reductions for directed lies aboutattitudes were large enough to nearly eliminate the PRP to the pointwhere they closely resembled the PRP changes found previouslyonly when participants made self-generated lies about episodic(Johnson et al., 2004, 2005), autobiographical (Johnson et al.,2002a) or personal semantic memories (Johnson et al., 2002b). Themain difference between self-generated lies and directed lies is thatthe former require participants to strategically monitor theirresponses while the latter do not. Nevertheless, the present resultssuggest that participants invoked strategic monitoring processeswhen making deceptive responses about their attitudes, despite thefact that the task did not require them.
Given that much of our everyday behavior is driven by ourattitudes and preferences, it is reasonable to posit that invokingstrategic monitoring processes could be seen as necessary toensuring that, over time, these evaluative judgments both conformto one another and to one’s self image. Similarly, strategicmonitoring is presumably necessary for maintaining a consistentpresentation of the self when interacting with others. In manycircumstances, failure to maintain a consistent pattern of responsescan have adverse social consequences (e.g., being judged asundependable or inconsistent). In support of this interpretation, theamplitudes of the PRPs elicited by truthful responses about agreeitems were smaller than those elicited when the same participantsmade decisions about their autobiographical memories (Johnson etal., unpublished data). Attitude valence also affected PRPamplitude, with greater reductions for agree items than for disagreeitems. The fact that this valence-related difference in PRP activitywas present to the same extent regardless of whether it wasfollowed by a truthful or deceptive response suggests that itemsviewed as being good and bad are automatically processeddifferently. Hence, the current PRP results suggest that makingresponses about one’s moral, social and religious views auto-matically invokes strategic monitoring, to a different extentdepending on stimulus valence and the intended response.Alternatively, it may be that attitudinal stimuli are viewed asbeing more important and/or central to the self than typicalepisodic or autobiographical stimuli and thus strategic monitoringis automatically invoked and/or required during their processing. Ineither case, future studies may find that the presence and extent ofstrategic monitoring for attitude evaluation also depend on whetherparticipants are at the high or low end of a self-monitoring scale.
Although different in polarity and timing from the MFN, thescalp distribution of the PRP was similar to that of the MFN,suggesting that it is also generated in or near the ACC. Despite thisdistributional similarity, however, the results indicate that these twoERP components reflect different underlying processes. That is, fortruthful responses, whereas valence affected PRP amplitude, it didnot affect MFN amplitude. Nevertheless, the results for both thesecomponents fit with the idea that the ACC is involved inmonitoring the processing of self-referential and moral judgments(e.g., Northoff and Bermpohl, 2004; Moll et al., 2005).
Other valence effects
In addition to their cognitive components, the affectivecomponent of goodness/badness judgments can evoke perceptual/implicit and/or conceptual/explicit processes (e.g., Cunningham et

480 R. Johnson Jr. et al. / NeuroImage 39 (2008) 469–482
al., 2003, 2004). By definition, both implicit and explicitprocessing of affective information should be independent of thenature of the subsequent response and three of our response-lockedERP components met this criterion: an early and long-lastingcentral-parietal slow potential, a late brief potential difference overprimary visual areas and the PRP (discussed above).
The major valence-related brain activity here was a central-parietal positive slow wave elicited by negative items in the 800-ms interval preceding the response. The relatively late onset of thisslow potential (i.e., 350–500 ms after stimulus onset), as well asthe fact that this activity reached its maximum immediately prior tothe response, suggests that it represents an explicit process. Forunknown reasons, this slow potential was only loosely coupled tostimulus onset, as indicated by the finding that its duration was thesame time for both truthful and deceptive responses, despite thefact that deceptive RTs were about 200 ms longer than truthful RTs.An interesting aspect of this valence effect is that it was due to thenegative (i.e., disagree) items because the ERPs for positive itemsremained essentially at baseline throughout this time interval. Asimilar dissociation in which greater activation was found fornegative compared to positive evaluations has also been reported inboth fMRI (e.g., Cunningham et al., 2003, 2004) and ERP studies(e.g., Ito et al., 1998).
The residual variance analyses here suggest that the neuralgenerators of this valence-related activity lie in the precuneus areabecause the dipole seeded there produced a source waveform thatclosely matched the time course of this slow potential. In additionto the Zysset et al. (2002) results, others have found precuneusactivations in paradigms involving the processing and/or evalua-tion of emotional salience (e.g., Maddock, 1999; Maddock et al.,2003), as well as during moral judgments (e.g., Greene et al.,2001). Based on these and similar data, Northoff and Bermpohl’s(2004) model of the neural basis of the self posits that theprecuneus is responsible for integrating stimuli into the emotionaland autobiographical context of the self. Given that the duration ofsuch integrative processes is likely to depend on the complexity ofthe decisions that are required about the stimuli, the extendedduration of this slow potential for attitude evaluations accords wellwith their model. Finally, because our stimulus sets only includedstrongly held attitudes, it remains unknown if this, or any of ourother valence effects, can be graded as a function of attitudestrength.
The next valence-related ERP activity spanned a 300-msinterval that ended about 100 ms before the response and wasspatially limited to the two electrodes located over primary visualcortex. This difference, consisting of a decreased negativity fornegative items, was superimposed on a truthful–deceptive responsedifference that was recorded over a larger scalp region encom-passing both occipital and extrastriate areas, which began severalhundred milliseconds earlier. Although the processes associatedwith both these ERP effects remain unknown, the finding ofsignificantly less amplitude for negatively valenced items fits withsome hemodynamic results. For example, Fossati et al. (2003)found significantly decreased blood flow in left and right occipitalcortex when participants evaluated whether negative, but notpositive, trait words described themselves.
All three of the valence-related ERP effects found here wereonly apparent in response-locked ERP averages. Hence, the factthat none has, to the best of our knowledge, been reportedpreviously is most likely due to the fact that previous studies onlyquantified stimulus-locked averages (e.g., Cacioppo and Bernston,
1994; Cacioppo and Gardner, 1996; Cunningham et al., 2005).However, given that our valence-related potentials were indepen-dent of the response, they all represent the kind of brain responsesbeing sought by previous investigators as veridical indications ofcovert attitudes. Thus, the present results add to the armamentar-ium of ERP components that could be used to reveal anindividual’s true attitudes, without the need for an overt response.
Spatio-temporal analyses of brain activity during attitudejudgments
Although fMRI data provide detailed information on thelocations of activated brain areas, they do not reveal temporaldetails such as the sequencing or duration of these activations. Incontrast, ERPs reveal the timing of brain activity between stimulusand response with a millisecond resolution. To combine thesuperior temporal information of ERPs with the superior spatialinformation of fMRI, we calculated the extent to which fourdipoles, located at Talairach coordinates taken from related fMRIstudies, would explain the ERP activity in different temporalintervals leading up to and following truthful and deceptiveresponses.
Overall, the residual variance analyses indicated a very highdegree of congruence between the ERP activity recorded here andthe results of fMRI studies using comparable paradigms. Followingdeceptive responses, in a time window containing peak MFNactivity, four dipoles explained over 95% of the ERP activityelicited by agree items and a bit less for the disagree items. Inkeeping with the lower levels of response conflict and lesser MFNactivity, these dipoles explained less of the ERP activity elicited bytruthful responses. Our results suggest, therefore, that the threemidline brain areas (e.g., anterior medial frontal cortex, precuneus,ACC) found in fMRI studies of self-referential judgments areactive simultaneously in the interval immediately preceding andfollowing the response. Furthermore, the anterior medial frontalcortex and precuneus also appear to be simultaneously active forseveral hundred milliseconds prior to the response, with increasingactivation as the response approached. Hence, there is thepossibility that these two brain areas are functionally linked duringthe evaluation process. Nevertheless, even if these brain areas arelinked, other results here indicate that they process different aspectsof the stimulus. That is, the self-referential focus of the ERPactivity produced by the anterior medial frontal generator wasdemonstrated by the fact that the same activity was elicited forjudgments about autobiographical stimuli but not for semanticjudgments (Johnson et al., in preparation). In contrast, we foundvalence effects over posterior scalp but not over anterior medialfrontal scalp, a finding that fits well with Fossati et al.’s (2003)results for judgments about positive and negative trait adjectives.
The advantage of using forward calculations to determine theamount of scalp-recorded activity not explained by the fMRI-identified brain activations (i.e., residual variance) is that it doesnot involve any backward calculations or assumptions about eitherthe numbers or locations of the active brain areas. However, onedisadvantage is that differences in experimental design and stimuliwill affect the final goodness of fit for the dipole solutions. Forexample, because we found significant response- and valence-related differences in ERP activity at additional scalp locations,which could not be modeled by these four dipoles, our solutionswere adversely affected by the fact that the RV represents allunexplained brain activity in any given temporal interval. That is,

481R. Johnson Jr. et al. / NeuroImage 39 (2008) 469–482
none of the seeded dipoles could account for the LPC activity,which may explain why RVs increased along with LPC amplitudefor the truthful and disagree deceptive response solutions.Similarly, the RVs were worse in the pre-response interval whenlarge amounts of ERP activity were elicited over visual areas,activity that was not modeled by any of the dipoles seeded here.Hence, the cost of our not making assumptions about generatorlocations was that the RVs were likely larger than would have beenobtained had we seeded extra dipoles to account for the additionalbrain activity elicited in our paradigm.
Conclusions
The present results demonstrated that making even simple,directed lies about the goodness/badness of attitude items resultedin quantitatively and qualitatively different patterns of behavioraland brain activity. Quantitatively, deceptions about attitudesappeared to produce much greater levels of conflict signalingthat such responses required greater cognitive control than isnecessary for deceptive responses about other types of stimuli.Qualitatively, the attitude stimuli were different in that theyappeared to invoke strategic monitoring prior to both truthful anddeceptive responses, even in the absence of any task requirementto do so. Thus, one new finding here is that evaluations aboutsome types of self-referential stimuli appear to automaticallyengage strategic monitoring processes, possibly to ensure that theself is presented in a consistent manner to others. Furthermore, thevalence assigned to each item by the participants affected both theextent to which strategic and tactical monitoring processes wereengaged, with greater amounts of both being evident for thepositive items. Although ethnically mixed, the participants wereall healthy young individuals and thus the generalizability of ourresults to people of other ages awaits future studies. Finally, usingthe forward-calculation option, the RV analyses suggest that thebrain activity evoked by truthful and deceptive responses aboutone’s attitudes arises, at least in part, from brain circuits positedon the basis of results in fMRI studies to provide the neural basisof the self.
Acknowledgments
We thank Doreen Nessler for performing the BESA analysesand Sarit Golub for suggesting the “denial of self” explanation forour MFN results in the deception condition. We also want to thankMaria Dayanayeva and Elisheva Diamond for their assistanceduring data collection and Yuriy Lehman for his invaluabletechnical assistance. This research was supported by the ResearchFoundation of the City University of New York.
References
Abe, N., Suzuki, M., Tsukiura, T., Mori, E., Yamaguchi, K., Itoh, M., et al.,2006. Dissociable roles of prefrontal and anterior cingulate cortices indeception. Cereb. Cortex 16, 192–199.
American Electroencephalographic Society, 1991. Guidelines for standardelectrode position nomenclature. J. Clin. Neurophysiol. 8, 200–202.
Amodio, D.M., Frith, C.D., 2006. Meeting of minds: the medial frontalcortex and social cognition. Nat. Rev., Neurosci. 7, 268–277.
Barch, D.M., Braver, T.S., Sabb, F.W., Noll, D.C., 2000. Anterior cingulateand the monitoring of response conflict: evidence from an fMRI study ofovert verb generation. J. Cogn. Neurosci. 12, 298–309.
Ben-Shakhar, G., Furedy, J.J., 1990. Theories and Applications in theDetection of Deception: A Psychophysiological and InternationalPerspective. Springer-Verlag, New York.
Botvinick, M., Nystrom, L.E., Fissell, K., Carter, C.S., Cohen, J.D., 1999.Conflict monitoring versus selection-for-action in anterior cingulatecortex. Nature 402, 179–181.
Botvinick, M.M., Cohen, J.D., Carter, C.S., 2004. Conflict monitoring andanterior cingulate cortex: an update. Trends Cogn. Sci. 8, 539–546.
Bush, G., Luu, P., Posner, M.I., 2000. Cognitive and emotional influences inanterior cingulate cortex. Trends Cogn. Sci. 4, 215–222.
Cacioppo, J.T., Bernston, G.G., 1994. Relationship between attitudes andevaluative space: a critical review, with emphasis on the separability ofpositive and negative substrates. Psychol. Bull. 115, 401–423.
Cacioppo, J.T., Crites Jr., S.L., Gardner, W.L., 1996. Attitudes to the right:evaluative processing is associated with lateralized late positive event-related brain potentials. Pers. Soc. Psychol. Bull. 2, 1205–1219.
Carter, C.S., Braver, T.S., Barch, D.M., Botvinick, M.M., Noll, D., Cohen,J.D., 1998. Anterior cingulate cortex, error detection, and the onlinemonitoring of performance. Science 280, 747B749.
Carter, C.S., Macdonald, A.M., Botvinick, M., Ross, L.L., Stenger, V.A.,Noll, D., et al., 2000. Parsing executive processes: strategic vs.evaluative functions of the anterior cingulate cortex. Proc. Natl. Acad.Sci. U. S. A. 97, 1944–1948.
Cialdini, R.B., Goldstein, N.J., 2004. Social influence: compliance andconformity. Annu. Rev. Psychol. 55, 591–621.
Craik, F.I.M., Moroz, T., Moscovitch, M., Stuss, M., Winocur, D.T.,Tulving, G., 1999. In search of the self, a positron emission tomographystudy. Psychol. Sci. 10, 26B34.
Crites Jr., S.L., Cacioppo, J.T., 1996. Electocortical differentiation ofevaluative and nonevaluative categorizations. Psychol. Sci. 7,318–321.
Crites Jr., S.L., Cacioppo, J.T., Gardner, W.L., Bernston, G.G., 1995.Bioelectrical echoes from evaluative categorization: II. A late positivebrain potential that varies as a function of attitude registration rather thanattitude report. J. Pers. Soc. Psychol. 68, 997–1013.
Cunningham,W.A., Johnson, M.K., Gatenby, J.C., Gore, J.C., Banaji, M.R.,2003. Neural components of social evaluation. J. Pers. Soc. Psychol. 85,639–649.
Cunningham, W.A., Raye, C.L., Johnson, M.K., 2004. Implicit and explicitevaluation: fMRI correlates of valence, emotional intensity, and controlin the processing of attitudes. J. Cogn. Neurosci. 16, 1717–1729.
Cunningham, W.A., Espinet, S.D., Deyoung, C.G., Zelazo, P.D., 2005.Attitudes to the right- and left-frontal ERP asymmetries associated withstimulus valence and processing goals. NeuroImage 28, 827–834.
Fossati, P., Hevenor, S.J., Graham, S.J., Grady, C., Keightley,M.L., Craik, F.,et al., 2003. In search of the emotional self: an fMRI study using positiveand negative emotional words. Am. J. Psychiatry 160, 1938–1945.
Ganis, G., Kosslyn, S.M., Stose, S., Thompson, W.L., Yurgelun-Todd, D.A.,2003. Neural correlates of different types of deception: an fMRIinvestigation. Cereb. Cortex 13, 830–836.
Gehring, W.J., Knight, R.T., 2000. Prefrontal–cingulate interactions inaction monitoring. Nat. Neurosci. 3, 516–520.
Gehring, W.J., Willoughby, A.R., 2002. The medial frontal cortex andthe rapid processing of monetary gains and losses. Science 295,2279–2282.
Gillihan, S.J., Farah, M.J., 2005. Is self special? A critical review ofevidence from experimental psychology and cognitive neuroscience.Psychol. Bull. 131, 76–97.
Greene, J.D., Sommerville, R.B., Nystrom, L.E., Darley, J.M., Cohen, J.D.,2001. An fMRI investigation of emotional engagement in moraljudgment. Science 293, 2105–2108.
Gusnard, D.A., Akbudak, E., Shulman, G.L., Raichle, M.E., 2001. Medialprefrontal cortex and self-referential mental activity: relation to a defaultmode of brain function. Proc. Natl. Acad. Sci. U. S. A. 98, 4259–4264.
Ito, T.A., Larsen, J.T., Smith, N.K., Cacioppo, J.T., 1998. Negativeinformation weighs more heavily on the brain: the negativity bias inevaluative categorizations. J. Pers. Soc. Psychol. 75, 887–900.

482 R. Johnson Jr. et al. / NeuroImage 39 (2008) 469–482
Johnson Jr., R., 1986. A triarchic model of P300 amplitude. Psychophysiol-ogy 23, 367B384.
Johnson Jr., R., 1988. The amplitude of the P300 component of the event-related potential: review and synthesis. In: Ackles, P.K., Jennings, J.R.,Coles, M.G.H. (Eds.), Advances in Psychophysiology, vol. III. JAIPress, CT, pp. 69–138.
Johnson Jr., R., Barnhardt, J., Adler, N., Simon, E., Zhu, J., 2002a. An ERPstudy of deception: lying about personal experiences. Invited PaperPresentation at the 42nd Annual Meeting of the Society forPsychophysiological Research. Washington, D.C. Psychophysiology,vol. 39, p. S11.
Johnson Jr., R., Barnhardt, J., Adler, N., Simon, E., Zhu, J., 2002b. An ERPstudy of deception: lying about personal facts. Paper Presented at the42nd Annual Meeting of the Society for Psychophysiological Research.Washington, D.C. Psychophysiology, vol. 39, p. S44.
Johnson Jr., R., Barnhardt, J., Zhu, J., 2003. The deceptive response: effectsof response conflict and strategic monitoring on the late positivecomponent and episodic memory-related brain activity. Biol. Psychol.64, 217–253.
Johnson Jr., R., Barnhardt, J., Zhu, J., 2004. The contribution of executiveprocesses to deceptive responding. Neuropsychologia 42, 878–901.
Johnson Jr., R., Barnhardt, J., Zhu, J., 2005. Differential effects of practiceon the executive processes used for truthful and deceptive responses: anevent-related brain potential study. Brain Res. Cogn. Brain Res. 24,386–404.
Kelley, W.M., Macrae, C.N., Wyland, C.L., Caglar, S., Inati, S., Heatherton,T.F., 2002. Finding the self? An event-related fMRI study. J. Cogn.Neurosci. 14, 785–794.
Kozel, F.A., Padgett, T.M., George, M.S., 2004. A replication study of theneural correlates of deception. Behav. Neurosci. 118, 852–856.
Langleben, D.D., Schroeder, L., Maldigian, J.A., Gur, R.C., McDonald, S.,Ragland, J.D., et al., 2002. Brain activity during simulated deception: anevent-related functional magnetic resonance study. NeuroImage 15,727–732.
Lee, T.M.C., Liu, H., Tan, L., Chan, C.C.H., Mahankali, S., Feng, C., et al.,2002. Lie detection by functional magnetic resonance imaging. Hum.Brain Mapp. 15, 157–164.
Maddock, R.J., 1999. The retrosplenial cortex and emotion: new insightsfrom functional neuroimaging of the human brain. Trends Neurosci. 22,310–316.
Maddock, R.J., Garrett, A.S., Buonocore, M.H., 2003. Posterior cingulatecortex activation by emotional words: fMRI evidence from a valencedecision task. Hum. Brain Mapp. 18, 30–41.
Mathalon, D.H., Bennett, A., Askari, N., Gray, E.M., Rosenbloom, M.J.,Ford, J.M., 2003. Response-monitoring dysfunction in aging andAlzheimer’s disease: an event-related potential study. Neurobiol.Aging 24, 675–685.
Moll, J., de Oliveira-Souza, R., Moll, F.T., Ignacio, F.A., Bramati, I.E.,Caparelli-Daquer, E.M., et al., 2005. The moral affiliations of disgust: afunctional MRI study. Cogn. Behav. Neurol. 18, 68–78.
Northoff, G., Bermpohl, F., 2004. Cortical midline structures and the self.Trends. Cogn. Sci. 8, 102–107.
Nunez, J.M., Casey, B.J., Egner, T., Hare, T., Hirsch, J., 2005. Intentionalfalse responding shares neural substrates with response conflict andcognitive control. NeuroImage 25, 267–277.
Olson, J.M., Vernon, P.A., Harris, J.A., Jang, K.L., 2001. The heritability ofattitudes: a study of twins. J. Pers. Soc. Psychol. 80, 845–860.
Scherg, M., 1990. Fundamentals of dipole source potential analysis. In:Grandori, F., Hoke, M., Romani, G.L. (Eds.), Auditory EvokedMagnetic Fields and Electric Potentials. Advances in Audiology,vol. 5. Karger, Basel, p. 40B69.
Schmitz, T.W., Kawahara-Baccus, T.N., Johnson, S.C., 2004. Metacognitiveevaluation, self-relevance, and the right prefrontal cortex. NeuroImage22, 941–947.
Seger, C.A., Stone, M., Keenan, J.P., 2004. Cortical activations duringjudgments about the self and an other person. Neuropsychologia 42,1168–1177.
Spence, S.A., Farrow, T.F.D., Herford, A.E., Wilkinson, I.D., Zheng, Y.,Woodruff, P.W.R., 2001. Behavioural and functional anatomicalcorrelates of deception in humans. NeuroReport 12, 2849–2853.
Thompson-Schill, S.L., D’Esposito, M., Aguirre, G.K., Farah, M.J., 1997.Role of left inferior prefrontal cortex in retrieval of semantic knowledge:a reevaluation. Proc. Natl. Acad. Sci. U. S. A. 94, 14792–14797.
Turken, A.U., Swick, D., 1999. Response selection in the human anteriorcingulate cortex. Nat. Neurosci. 2, 920B924.
Ullsperger, M., Von Cramon, D.Y., 2001. Subprocesses of performancemonitoring: a dissociation of error processing and response competi-tion revealed by event-related fMRI and ERPs. NeuroImage 14,1387–1401.
van Veen, V., Carter, C.S., 2006. Conflict and cognitive control in the brain.Psychol. Sci. 15, 237–240.
Vidal, F., Hasbroucq, T., Grapperon, J., Bonnet, M., 2000. Is the ‘errornegativity’ specific to errors? Biol. Psychol. 51, 109–128.
Zysset, S., Huber, O., Ferstl, E., von Cramon, D.Y., 2002. The anteriorfrontomedian cortex and evaluative judgment: an fMRI study. Neuro-Image 15, 983–991.
Copyright © 2022 FDOKUMEN




















