
The relationship between percentage of singletons and sampling effort: A new approach to reduce the...
35
Elsevier Editorial System(tm) for Ecological Indicators Manuscript Draft Manuscript Number: Title: The relationship between percentage of singletons and sampling effort: a new approach to reduce the bias of richness estimates Article Type: Research Paper Keywords: species richness estimation, sampling intensity, singletons, inventory completeness, chao1, jackknife, bootstrap, chao2, ACE, ICE Corresponding Author: Dr Luiz Carlos Serramo Lopez, Corresponding Author's Institution: Universidade Federal da Paraiba First Author: Luiz Carlos Serramo Lopez Order of Authors: Luiz Carlos Serramo Lopez; Maria P Fracasso; Daniel O Mesquita; Alexandre R Palma; Pablo Riul Abstract: Estimate the richness of a community with accuracy despite differences in sampling effort is a key aspect to monitoring high diverse ecosystems. We compiled a worldwide multitaxa database, comprising 185 communities, in order to study the relationship between the percentage of species represented by one individual (singletons) and the intensity of sampling (number of individuals divided by the number of species sampled). The database was used to empirically adjust a correction factor to improve the performance of non-parametrical estimators under conditions of low sampling effort. The correction factor was tested on seven estimators (Chao1, Chao2, Jack1, Jack2, ACE, ICE and Bootstrap). The correction factor was able to reduce the bias of all estimators tested under conditions of undersampling, while converging to the original uncorrected values at higher intensities. Our findings led us to recommend the threshold of 20 individuals/species, or less than 21% of singletons, as a minimum sampling effort to produce reliable richness estimates of high diverse ecosystems using corrected non-parametric estimators. This threshold rise for 50 individuals/species if non-corrected estimators are used which implies in an economy of 60% of sampling effort if the correction factor is used.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of The relationship between percentage of singletons and sampling effort: A new approach to reduce the...

Elsevier Editorial System(tm) for Ecological Indicators Manuscript Draft Manuscript Number: Title: The relationship between percentage of singletons and sampling effort: a new approach to reduce the bias of richness estimates Article Type: Research Paper Keywords: species richness estimation, sampling intensity, singletons, inventory completeness, chao1, jackknife, bootstrap, chao2, ACE, ICE Corresponding Author: Dr Luiz Carlos Serramo Lopez, Corresponding Author's Institution: Universidade Federal da Paraiba First Author: Luiz Carlos Serramo Lopez Order of Authors: Luiz Carlos Serramo Lopez; Maria P Fracasso; Daniel O Mesquita; Alexandre R Palma; Pablo Riul Abstract: Estimate the richness of a community with accuracy despite differences in sampling effort is a key aspect to monitoring high diverse ecosystems. We compiled a worldwide multitaxa database, comprising 185 communities, in order to study the relationship between the percentage of species represented by one individual (singletons) and the intensity of sampling (number of individuals divided by the number of species sampled). The database was used to empirically adjust a correction factor to improve the performance of non-parametrical estimators under conditions of low sampling effort. The correction factor was tested on seven estimators (Chao1, Chao2, Jack1, Jack2, ACE, ICE and Bootstrap). The correction factor was able to reduce the bias of all estimators tested under conditions of undersampling, while converging to the original uncorrected values at higher intensities. Our findings led us to recommend the threshold of 20 individuals/species, or less than 21% of singletons, as a minimum sampling effort to produce reliable richness estimates of high diverse ecosystems using corrected non-parametric estimators. This threshold rise for 50 individuals/species if non-corrected estimators are used which implies in an economy of 60% of sampling effort if the correction factor is used.

1
The relationship between percentage of singletons and sampling effort: a new approach to reduce 1
the bias of richness estimates. 2
3
Luiz Carlos Serramo Lopez1; Maria Paula de Aguiar Fracasso
2; Daniel Oliveira Mesquita
1; 4
Alexandre Ramlo Torre Palma1; Pablo Riul
3. 5
6
1Departamento de Sistemática e Ecologia, Centro de Ciências Exatas e da Natureza, Universidade 7
Federal da Paraíba, Cidade Universitária, João Pessoa Paraíba, 58059-900, Brazil. 8
[email protected]; [email protected]; [email protected] 9
2Departamento de Biologia, Universidade Estadual da Paraíba, Av. das Baraúnas, 351/Campus 10
Universitário, Bodocongó, 58109-753,Campina Grande, Paraíba, Brazil. [email protected] 11
3Departamento de Engenharia e Meio Ambiente, Centro de Ciências Aplicadas e Educação, 12
Universidade Federal da Paraíba - Campus IV, R: Mangueira s/n, Centro CEP: 58.297-000, Rio 13
Tinto - Paraíba - Brazil. [email protected] 14
Correspondent author: Luiz Carlos Serramo Lopez 15
Address: Departamento de Sistemática e Ecologia, Centro de Ciências Exatas e da Natureza, 16
Universidade Federal da Paraíba, Cidade Universitária, João Pessoa Paraíba, 58059-900, Brazil. 17
Phone: 55 83 9937 6226. Email: [email protected] 18
19
20
ManuscriptClick here to view linked References

2
1. Introduction 21
Species richness is a key indicator for biodiversity and the demand for more accurate 22
richness estimation grows in parallel with the increased human alteration of our biosphere 23
(Clarke et al. 2011, Gotelli and Colwell 2001). However, researches face a trade-off between 24
very complete diversity inventories, which are time and resource consuming, and briefer ones 25
thought to be more imprecise. Longino et al. (2002) and Mao and Colwell (2005) stressed the 26
challenges involved in determining the total richness of a given community, since there is an 27
overwhelming presence of rare species in mega-diverse ecosystems. 28
Using non-parametric richness estimators is a potential tool to evaluate the completeness of 29
an inventory (Chao 1984; Smith and van Belle 1984; Colwell and Coddington 1994). Non-30
parametric estimators are thought to be less dependent on the rate of collection of unseen species 31
discovery or the shape of the assemblage distribution (Chao et al. 2009, Palmer 1990, Palmer 32
1991, Zelmer and Esch 1999). However, they demand a minimum sampling effort to produce 33
reliable estimates (Brose and Martinez 2004, Chao et al. 2009, Chiarucci et al. 2003). 34
Coddington et al. (2009) suggested that many inventories of tropical arthropods suffer from 35
an undersampling bias, strong enough to impair even the use of richness estimators in order to 36
assess the real richness of these assemblages. In a large compilation of tropical arthropod 37
inventories, they also found a significant negative relationship between the percentage of species 38
represented by one individual (singleton) and the sample intensity (abundance divided by 39
richness). Singletons have an intuitive connection with inventory completeness, since we expect 40
that the proportions of singletons should decrease as the sampling effort increases, until we come 41
close to the “real” proportion of singletons present in a community. For instance, Coddington et 42
al. (2009) estimated a true proportion of 4% singletons by lognormal extrapolation from their 43
spider assemblage, which originally presented 29% singletons. 44

3
Solutions to this undersampling bias are either a dramatic increase in sample effort or the 45
development of better richness estimators (Chiarucci et al. 2003, Ulrich and Ollik 2005). Here we 46
proposed that it is possible to correct classical non-parametrical estimators in order to boost their 47
performance under conditions of undersampling. We empirically derived this correction using the 48
relationship between the intensity of sampling and the proportion of singletons found in a large 49
database of communities we obtained from the literature. 50
51
1.1 Deriving the estimator correction 52
53
Our correction to improve a non-parametric estimator under low sampling conditions 54
consists in multiplying the original estimative by 1 plus the proportions of singletons in the 55
sample elevated by a constant: 56
57
SestP= Sest (1+Pz) (1) 58
59
Where SestP is the modified estimate, Sest is the original estimate, P is the proportion of 60
singletons (singletons / observed species richness) and z is a constant higher than one. Since the 61
proportions of singletons (P) falls as the sampling effort increases, this basic formula will 62
improve the performance of the estimator under low sampling effort but will converge to the 63
original estimate at high sampling effort conditions. 64
The constant z in the formula shall mirror the allometric relationship between the 65
proportions of singletons and the intensity of sampling in natural assemblages. The value of the 66
constant z can be empirically derived as 67
z= - (ln I / ln P) (2) 68

4
Where ln I is the natural logarithm of the sampling intensity (the number of individuals 69
divided by the number of species observed in a given sample) and ln P is the natural logarithm of 70
the proportions of singletons in the same sample. To estimate the value of z we made a large 71
compilation of different communities, varying widely in sampling intensity and taxonomical 72
composition. We found the average value of z in this database of 185 assemblages to be close to 73
2 (2.06 ± 0.73 SD, n= 185), leading us to a general transformation to correct non-parametric 74
estimators under low sampling effort 75
SestP= Sest (1+P2) (3) 76
This correction (called P correction) can also be used to species-incidence estimators by 77
substituting the proportion of singletons (P) by the proportions of uniques (Pu). The 78
transformation will increase the estimate (up to 100%) when the proportion of singletons (or 79
uniques) in the sample is high, but decreases exponentially, converging towards the original 80
estimator value when the proportion is low. For example, a sample with a proportion of 0.5 81
singletons (or uniques) will generate a transformed estimate 25% (1+ (0.5 2) = 1.25) higher than 82
the non-transformed estimate, while another sample with a proportion of 0.1 of singletons would 83
be just 1% (1+ (0.1 2) = 1.01) higher. 84
We also used our database of communities to search for trends that could indicate the 85
“real” proportions of singletons in well sampled assemblages and to evaluate the limits that low 86
sample intensities pose to the reliability of non-parametric estimators, with and without the 87
correction we developed. 88
We tested our P correction in the most common used non parametric estimators: Chao1 89
(Chao 1984), Chao2 (Chao 1987), Jackknife1 (Heltshe and Forrester 1983), Jackknife2 (Burnham 90
and Overton 1978), ACE, ICE (Chao and Lee 1992), and Bootstrap (Smith and van Belle 1984). 91
Our goal was to assess whether the P corrected estimators were able to produce better estimations 92

5
at lower sampling intensity, in relation to their uncorrected versions, while converging to their 93
original values at conditions of higher sampling effort. 94
We start by doing test comparisons between the Chao1 estimator and is transformed version 95
(“Chao1P”), because the parameters needed to calculate Chao1 and the transformed “Chao1P” 96
(observed richness, singletons and doubletons) are widely available in the literature allowing us 97
to calculate the difference between both estimations for a large database of communities. After 98
that, we extended our tests to include the other six estimators in order to confirm if the results 99
found with Chao1 correction could be applied to non-parametrical estimators in general. 100
101

6
2. Methods 102
103
2.1 Database of communities 104
We expanded Coddington et al. (2009) original compilation of terrestrial arthropod 105
inventories, adding other taxa (tropical trees, corals and terrestrial vertebrates) to produce a set of 106
185 datasets where the singletons-richness ratio and the intensity of sampling 107
(abundance/richness) were calculated (see Appendix S1 in Supporting Information for details). 108
109
2.2 Testing the efficacy of the P transformation for Chao1 estimator using data from large 110
plots of tropical forests 111
112
We used the data from six inventories produced by research teams belonging to The Center 113
for Tropical Forest Science network of large forests plots around the world ((Condit et al. 2005), 114
(CTFS 2009)). We used data from 3 different continents: Africa (Korup Forest, census 1998 and 115
Edoro Forest, census 2000), Americas (BCI, census 2005 and Luquillo, census 1995) and Asia 116
(Huai Kha Khaeng, census 1999 and Pasoh, census 1995). For each plot, we obtained simulated 117
sets of 100 rarefied sub-samples with increasing average intensities (5, 25, 75 and 100 ind/spp). 118
Simulations were made such that each abundance from the original set was reduced according to 119
the formula: 120
121
Fi = (ai/f)r (3) 122
123
where Fi is the fraction of species i abundance, ai is the original abundance of species i, f is 124
the fraction (varying between 1 and ∞) by which the set is rarefied and r is a random number 125

7
between 0 and 1 (uniform distribution). This procedure generates samples containing fractions of 126
the original set with similar proportions among species to those found in the original one, but 127
with a random noise simulating the effect of random variations due to incomplete sampling. 128
We used the classical Chao1 (Chao 1984) estimator to perform a series of comparisons 129
between the original Chao1 formula and the P corrected “Chao1P”. We calculated the average 130
Chao1 estimates and our corrected Chao1P for each set of rarefaction simulations at different 131
intensities. Using these estimates, we calculated the bias and precision of these two estimators 132
using the scaled mean error (SME) and the coefficient of variation respectively (Walther and 133
Moore 2005). Fitting the average estimates from these rarefactions to power curves, we also 134
inferred the minimum sampling intensity necessary to estimate 100% and 95% of the original 135
richness. 136
137
2.3 Using the database of communities to test the efficiency of Chao1 estimator 138
We calculated the percentage difference between Chao1 and Chao1P estimates for the 185 139
communities in our database. This difference can be derived from the Chao1P formula (eq. 2) as 140
follows: 141
142
Difference Chao1 vs Chao1P= (( f1/ Sobs)2) 100 (4) 143
144
To check the validity of our rarefactions, we calculated the mean difference between Chao1 145
and Chao1P estimates from simulated intensities of 5, 25, 75 and 100 (obtained from the high 146
intensity forest plots) and compared them with the differences obtained from our multi-taxa 147
database with similar non-rarefied intensities. We used the mid-points of intensity intervals 0-10 148

8
(comparing them with simulations with intensity 5), 15-35 (intensity 25), 40-60 (intensity 50), 149
65-85 (intensity 75) and 90-110 (intensity 100). 150
151
2.4 Testing the P correction for Chao 2, Jackknife 1, Jackknife 2, ACE, ICE and 152
Bootstrap estimators. 153
154
To test the performance of the P correction for other non-parametrical estimators, besides 155
the Chao 1, we made simulations using data from BCI tree plot (census 2005) (CTFS, 2009). We 156
created 50 pairs of rarefied samples drawn from BCI data with 5 levels of sampling intensity (5, 157
25, 50, 75 and 100 ind/spp). These subsamples were used as an input data for EstimateS 8.2 158
(Colwell 2009) create the original, uncorrected, estimates using Chao2, Jack1, Jack2, ACE, ICE 159
and Bootstrap estimators (100 simulations per sample). Using the formula (1) we transformed 160
the original estimates in their corrected P versions ( Chao2P, Jack1P etc.) and compare the ability 161
of corrected and uncorrected estimators to estimate 100% of BCI dataset richness (299 species) 162
using sub-sets with reduced intensity of sampling. 163
164
3. Results 165
166
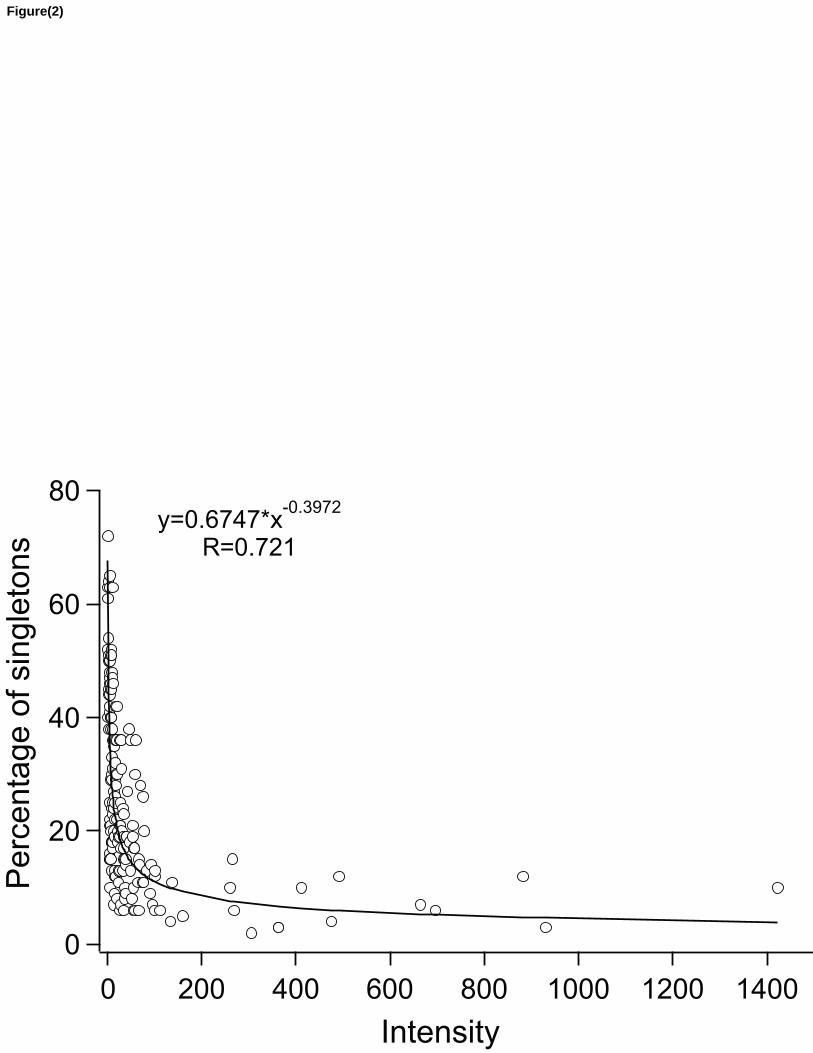
Our expanded database encompasses 185 communities, ranging from 1 to 1,423 ind/spp of 167
intensity and between 2% and 72% singletons. The median intensity was 20.3 and the median 168
percentage of singletons 19.2%. The communities samples belonged to four major groups: 169
terrestrial vertebrates (n=79), terrestrial arthropods (n=72), corals (n=22) and trees (n=12) (see 170
Appendix S1). The correlation between intensity and percentage of singletons was highly 171
significant (log-transformed power curve, r= 0.72; p<0.0001; Fig. 2). The percentage of 172

9
singletons tended to decline as sampling intensity increased, with samples of intensity 5 or less 173
(n= 23) having an average of 46% singletons (±3% SE), while communities with a sampling 174
intensity of 100 or more (n= 20) had an average of 8% of singletons (±1% SE). Due to the shape 175
of the power curve, most of the reduction in singleton percentage occurred at lower intensities, 176
between 0 and 75 species/individuals. 177
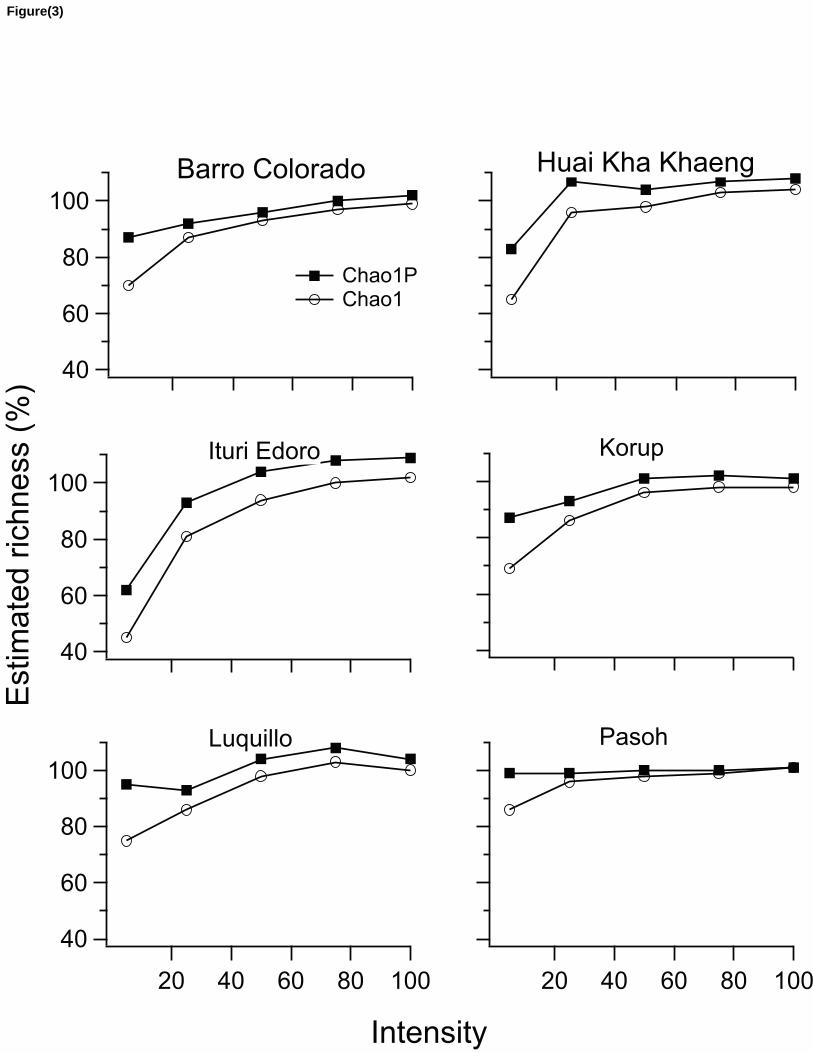
The Chao1P estimator had less bias and precision compared to Chao1, when tested in 178
rarefied sub-samples from six large plots of trees (Table 1). At higher intensities, both estimators 179
yield very similar results (0.8% on average bias difference at original intensities). However, at the 180
low intensity of 5 ind/spp, Chao1P outperforms Chao1 by 17.1% in terms of bias, with an 181
average precision loss of 2.8 % compared to Chao1 (Table 1). 182
The estimated richness increased with rarefaction intensities in a pattern that fits very well 183
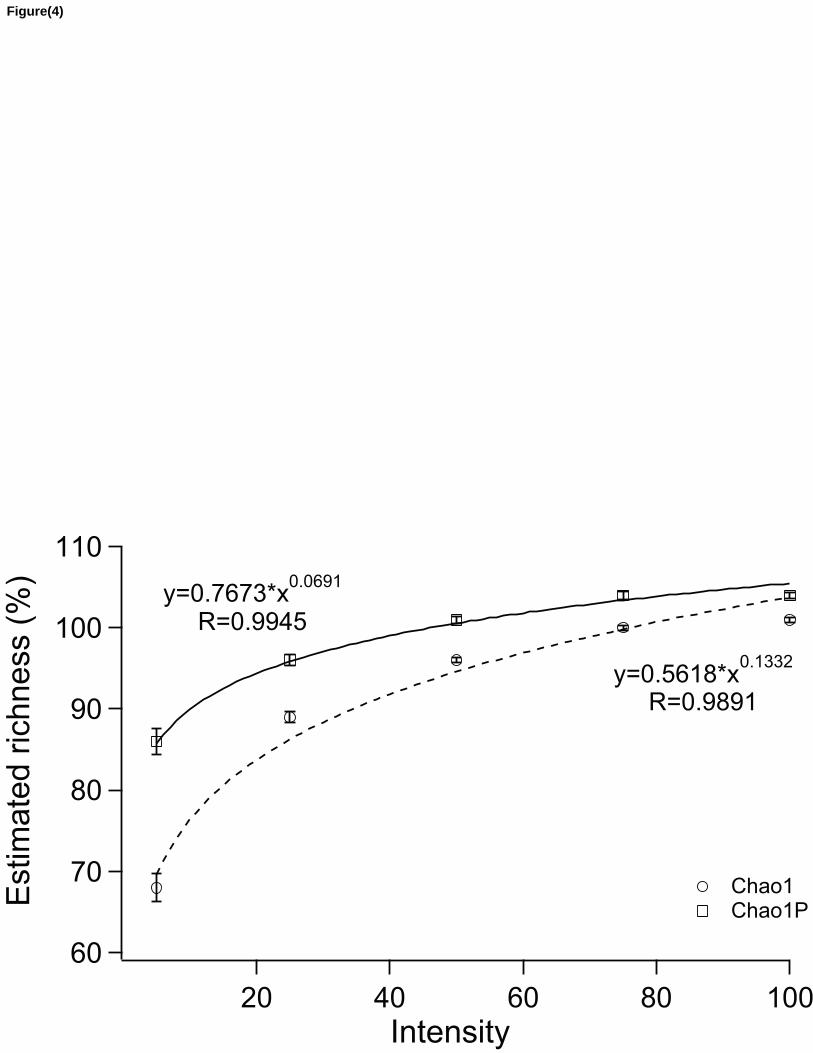
to the power curves for both Chao1P and Chao1 (R2 = 0.99 for Chao1P and 0.98 for Chao1) (Fig 184
4). These curves predict that, on average, Chao1P will estimate 100% of original richness at an 185
intensity of 52.0 ind/spp (± 9.4 SE), while Chao1 will reach 100% at an intensity of 78.7 ind/spp 186
(± 5.5 SE) (Fig. 4). If we use 95% of original richness instead of 100% as a good approximation 187
of original richness (as proposed by Chao et al. (2009)), the thresholds change to 20.7 ind/spp (± 188
5.9 SE) for Chao1P and 50.0 ind/spp (± 9.4 SE) for Chao1. 189
The difference between the two estimators obtained from the rarefaction simulations (from 190
tree plots with intensity 261 or more) showed good agreement with the average difference 191
obtained from samples of taxa that had low intensity (Fig.5). For example, on rarefactions to an 192
intensity of 5 the Chao1P estimates were, on average, 17.1% (± 1% SE) higher than Chao1 while 193
for the 57 communities (29 of arthropods and 28 of vertebrates) in our database with intensities 194
ranging between zero and 10 (midpoint intensity 5) the average difference was 17.2% (± 2% SE) 195
(Fig. 5). 196

10
The P transformation improved the performance of the other 6 non-parametrical estimators 197
(Chao2, Jack1, Jack2, ACE, ICE and Bootstrap) in similar way it did for Chao1. The transformed 198
estimators produced estimates that were more close to BCI real richness compared to their 199
untransformed versions under simulated conditions of low sampling effort (9% less biased, in 200
average, compared to the uncorrected formulas at the intensity of 5 ind/spp) and converge to the 201
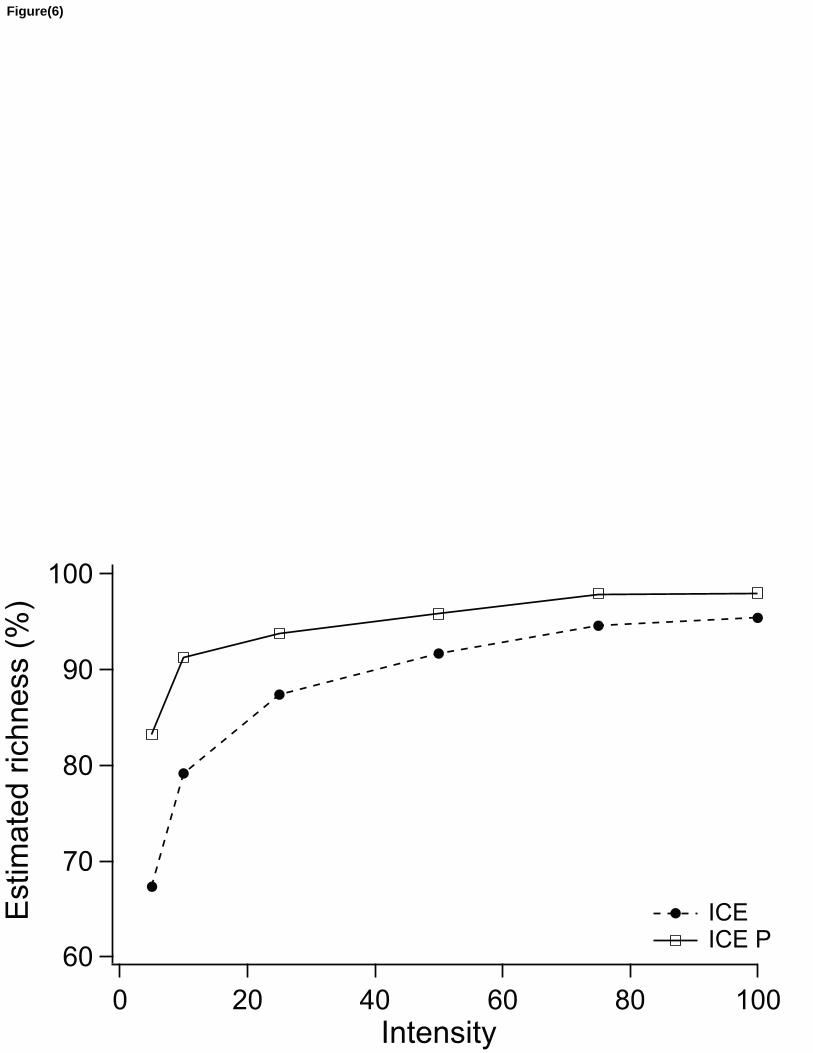
untransformed values as sampling effort increases (Table 2). The ICE corrected estimator 202
(“ICEP”) showed the best overall performance, in these simulations, estimating, in average, 83% 203
of BCI real richness at intensity 5 ind/spp compared to 67% made using its uncorrected version 204
(Fig. 6). 205
206
4. Discussion 207
208
Our expanded dataset confirmed the trend found by Coddington et al. (2009) for their 209
arthropod database: the percentage of singletons tends to decrease with an increase in the 210
sampling intensity in a very consistent way. At lower intensities, one needs to increase the 211
intensity of sampling by five-fold in order to halve the frequency of singletons. However, at 212
higher intensities (roughly, above intensity 100), the frequency of singletons tends to stabilize 213
around 8% (Fig. 2). Given that we have a phylogenetically diverse group of assemblages present 214
in our database (corals, arthropods, vertebrates and trees), we assume that this pattern is a general 215
one among communities. Consequently, the value of 8% (± 4% SD) is probably close to the 216
percentage of singletons expected from most natural communities after severe undersampling 217
bias is removed. 218
Since the proportion of singletons has a robust statistical relationship with the degree of 219
undersampling, it can be used to adjust the results from non-parametrical estimators. Our 220

11
transformation, empirically derived from this relationship, was able to reduce the bias from all 221
the non-parametric estimators tested compared to their untransformed versions at low sampling 222
intensities, while both versions (corrected x uncorrected) converged to very similar values at high 223
intensities. 224
The accuracy of an estimator is a compromise between the variation among estimations 225
(precision) and the distance between the estimated richness and the real richness (bias) (Brose et 226
al. 2003, Walther and Moore 2005). For example, at lower intensities (intensity 5), the corrected 227
Chao1P showed a reasonable trade-off, losing on average 3% precision, but gaining 17% in bias 228
reduction compared to Chao1. A mean of 17% less bias was found in both the rarefaction 229
simulations, drawn from high intensity samples, and from non-rarified lower intensity samples 230
(Fig. 5). This agreement between rarefied sub-samples from large tree plots and other multi-taxa 231
data suggests that the simulations were able to reproduce realistic patterns in low intensity 232
samples of natural situations. 233
Notice that the improvement provided by Chao1P applies not only to the average values but 234
also to the 95% boundary, which can be used to produce less conservative richness estimates. For 235
example, for the BCI dataset rarefied at an intensity of 5 ind/spp, Chao1P improved both the 236
average estimate (24% less bias) and the upper 95% estimate (26% less bias) compared to 237
untransformed Chao1. 238
The rarefactions, using data from six large plots of trees, also allows us to predict the 239
minimum intensity necessary for Chao1 and Chao1P to make estimates close to 100% of original 240
species richness. According to these simulations, it would be necessary to sample, on average, 241
51% more individuals to be able to make an accurate estimation using Chao1 (minimum intensity 242
78.7 ind/spp) compared to Chao1P (minimum intensity 52.0 ind/spp). A difference of this 243

12
magnitude can represent a great economy of time and resources while estimating the total 244
richness of very diverse communities. 245
If one uses 95% of the total richness estimated as a more tenable goal (Chao et al. 2009), 246
the difference in sampling effort between Chao1P and Chao1 becomes even larger, since our 247
simulations predict that one would need to sample, on average, 142% more individuals using 248
Chao1 (minimum intensity 50.0) than for Chao1P (minimum intensity 20.7) to estimate 95% of a 249
total sample richness. Since we found an increase of only 2% on Chao1P precision loss compared 250
to Chao1 at intensity 25 (close to the threshold of 20.7 for Chao1P for 95% estimation), the 251
trade-off between loss of precision and gain in economy of sampling effort in order to estimate 252
95% of total richness appears to be extremely positive. 253
The other non-parametric estimators tested (Chao2, Jack1, Jack2, ACE, ICE and Bootstrap) 254
presented the same pattern found with Chao1 (Fig. 6). The P corrected versions of each estimator 255
tested produced less biased estimates at low sampling intensities compared to their original 256
formulas while the corrected values converge to the original ones as the intensity of sampling 257
increases. The corrected version of ICE (ICEP), for example, was able to estimate 95% of BCI 258
plot using sub-samples with 50 ind/spp of intensity while the uncorrected version of ICE only 259
achieved the same feat at intensity 100 ind/spp (Fig. 6), this difference represents an economy of 260
50% in terms of sampling effort. 261
Consequently, our findings strongly indicate that our correction for non-parametric 262
estimators (equation (3)) produce less biased results and should be used to estimate the richness 263
in ecological studies that are trying to remove the effects of undersampling. An alternative option 264
is to parametric extrapolate the number of species to a given area or number of individuals (Melo 265
et al. 2007, O'Dea et al. 2006, Reichert et al. 2010). However, if such information (the total 266

13
community area, or the final number of individuals expected to be sampled) is not available, a 267
non-parametrical estimation using the correction present in equation (3) is the best option. 268
We also demonstrated that the intensity of sampling (the number of individuals sampled 269
divided by the number of species) and the proportion of singletons (the number of species 270
represented by one individual divided by the total number of species) can be used to indirectly 271
access the accuracy of richness estimates. Since these two parameters can be easily determined at 272
each stage of a real sampling program they can provide useful guidelines for planning and 273
evaluating biodiversity surveys. In our multi-taxa database, for example, 74% of the inventories 274
are below the average intensity threshold necessary to estimate at least 95% of the total richness 275
using Chao1, and 50% did not reach the same kind of threshold for Chao1P. These numbers give 276
support to Coddington et al.’s (2009) arguments that we need greater investment in biodiversity 277
inventories in order to get a realistic picture of the true richness of highly diverse ecosystems. 278
Our results indicate that ecological surveys that present more than 8% of singletons, or less 279
than 100 individuals/species of sampling intensity, probably are suffering from some degree of 280
undersampling and could be improved either by an increase of sampling effort or by using 281
richness estimators. Our simulations and database analysis led us to recommend the threshold of 282
20 individuals/species, or less than 21% of singletons, as a minimum sampling effort to produce 283
reliable richness estimates (at least 95% of richness estimated) using corrected non-parametric 284
estimators. This threshold rise for 50 individuals/species, or less than 14% of singletons, if non-285
corrected estimators are used, which implies in an economy of 60% of sampling effort due to the 286
correction factor. 287
288
289

14
Acknowledgments 290
We thank Nicholas Gotelli, Adriano S. Melo and Carlos Eduardo Grelle for insightful 291
comments on the subject.This work is supported by research fellowship from CNPq to DOM and 292
a post doc fellowship from CNPq/FAPESQ to MPAF. 293
294

15
References 295
296
Brose, U.,Martinez, N. D. 2004. Estimating the richness of species with variable mobility. Oikos 297
105(2), 292-300. 298
Burnham, K. P.,Overton, W. S. 1978. Estimation of the size of a closed population when capture 299
probabilities vary among animals. Biometrika 65, 623–633 300
Chao, A. 1984. Nonparametric-estimation of the number of classes in a population. Scandinavian 301
Journal of Statistics 11(4), 265-270. 302
Chao, A. 1987. Estimating the population size for capture-recapture data with unequal 303
catchability. Biometrics 437, 83-791. 304
Chao, A., Colwell, R. K., Lin, C. W.,Gotelli, N. J. 2009. Sufficient sampling for asymptotic 305
minimum species richness estimators. Ecology 90(4), 1125-1133. 306
Chao, A.,Lee, S. M. 1992. Estimating the number of classes via sample coverage. Journal of the 307
American Statistical Association 87210–217. 308
Chiarucci, A., Enright, N. J., Perry, G. L. W., Miller, B. P.,Lamont, B. B. 2003. Performance of 309
nonparametric species richness estimators in a high diversity plant community. Diversity 310
and Distributions 9(4), 283-295. 311
Clarke, K., Lewis, M.,Ostendorf, B. 2011. Additive partitioning of rarefaction curves: Removing 312
the influence of sampling on species-diversity in vegetation surveys. Ecological 313
Indicators 11(1), 132-139. 314
Coddington, J. A., Agnarsson, I., Miller, J. A., Kuntner, M.,Hormiga, G. 2009. Undersampling 315
bias: the null hypothesis for singleton species in tropical arthropod surveys. Journal of 316
Animal Ecology 78(3), 573-584. 317
Colwell, R. K. 2009. EstimateS: Statistical estimation of species richness and shared species from 318
samples. Version 8.2. User's Guide and application published at: 319
http://purl.oclc.org/estimates. 320
Condit, R. G., M. S. Ashton, H. Balslev, N. V. L. Brokaw, S. Bunyavejchewin, G. B. Chuyong, 321
Co, L., H. S. Dattaraja, S. J. Davies, S. Esufali, C. E. N. Ewango, R. B. Foster, N. 322
Gunatilleke, S. Gunatilleke, C. Hernandez, S. P. Hubbell, R. John, D. Kenfack, S. 323
Kiratiprayoon, P. Hall, T. H. Hart, A. Itoh, J. V. LaFrankie, I. Liengola, D. Lagunzad, S. 324
Lao, E. C. Losos, E. Magard, J. R. Makana, N. Manokaran, H. Navarrete, S. Mohammed 325

16
Nur, T. Okhubo, R. Perez, C. Samper, L. H. Hua Seng, R. Sukumar, J. C. Svenning, S. 326
Tan, D. W. Thomas, J. D. Thompson, M. I. Vallejo, G. Villa Muñoz, R. Valencia, T. 327
Yamakura,Zimmerman., J. K. 2005. Tropical tree α -diversity: Results from a worldwide 328
network of large plots. Biologiske Skrifter 55, 565-582. 329
CTFS 2009. Center for Tropical Forest Science.[WWW document]. URL http://www.ctfs.si.edu/. 330
Gotelli, N. J.,Colwell, R. K. 2001. Quantifying biodiversity: procedures and pitfalls in the 331
measurement and comparison of species richness. Ecology Letters 4(4), 379-391. 332
Heltshe, J. F.,Forrester, N. E. 1983. Estimating species richness using the Jackknife procedure. 333
Biometrics 3, 91–11. 334
Mao, C. X.,Colwell, R. K. 2005. Estimation of species richness: Mixture models, the role of rare 335
species, and inferential challenges. Ecology 86(5), 1143-1153. 336
Melo, A. S., Bini, L. M.,Thomaz, S. M. 2007. Assessment of methods to estimate aquatic 337
macrophyte species richness in extrapolated sample sizes. Aquatic Botany 86(4), 377-338
384. 339
O'Dea, N., Whittaker, R. J.,Ugland, K. I. 2006. Using spatial heterogeneity to extrapolate species 340
richness: a new method tested on Ecuadorian cloud forest birds. Journal of Applied 341
Ecology 43(1), 189-198. 342
Palmer, M. W. 1990. The estimation of species richness by extrapolation. Ecology 711195–1198. 343
Palmer, M. W. 1991. Estimating species richness: the second order jackknife reconsidered. 344
Ecology 72, 1512–1513. 345
Reichert, K., Ugland, K. I., Bartsch, I., Hortal, J., Bremner, J.,Kraberg, A. 2010. Species richness 346
estimation: estimator performance and the influence of rare species. Limnology and 347
Oceanography: Methods 8, 294-303. 348
Smith, E. P.,van Belle, G. 1984. Non-parametric estimation of species richness. Biometrics 40, 349
119–129. 350
Ulrich, W.,Ollik, M. 2005. Limits to the estimation of species richness: The use of relative 351
abundance distributions. Diversity and Distributions 11(3), 265-273. 352
Walther, B. A.,Moore, J. L. 2005. The concepts of bias, precision and accuracy, and their use in 353
testing the performance of species richness estimators, with a literature review of 354
estimator performance. Ecography 28(6), 815-829. 355

17
Zelmer, D. A.,Esch, G. W. 1999. Robust estimation of parasite component community richness. 356
Journal of Parasitology 85, 592–594. 357
18
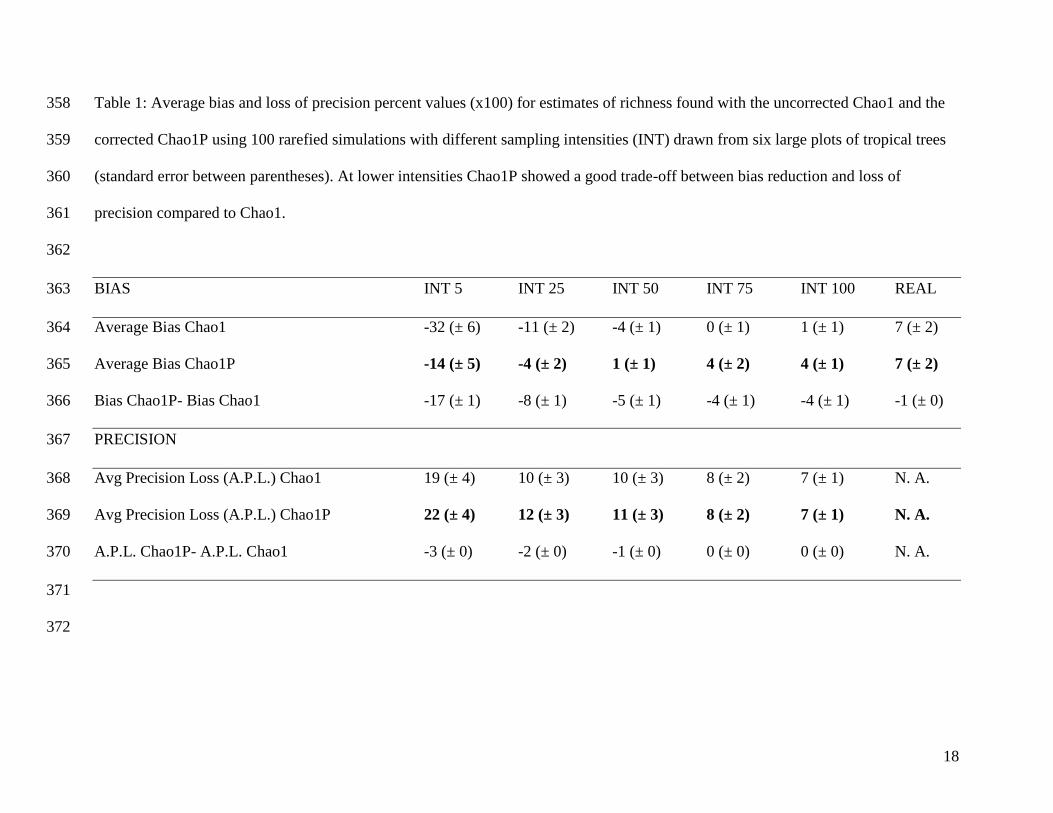
Table 1: Average bias and loss of precision percent values (x100) for estimates of richness found with the uncorrected Chao1 and the 358
corrected Chao1P using 100 rarefied simulations with different sampling intensities (INT) drawn from six large plots of tropical trees 359
(standard error between parentheses). At lower intensities Chao1P showed a good trade-off between bias reduction and loss of 360
precision compared to Chao1. 361
362
BIAS INT 5 INT 25 INT 50 INT 75 INT 100 REAL 363
Average Bias Chao1 -32 (± 6) -11 (± 2) -4 (± 1) 0 (± 1) 1 (± 1) 7 (± 2) 364
Average Bias Chao1P -14 (± 5) -4 (± 2) 1 (± 1) 4 (± 2) 4 (± 1) 7 (± 2) 365
Bias Chao1P- Bias Chao1 -17 (± 1) -8 (± 1) -5 (± 1) -4 (± 1) -4 (± 1) -1 (± 0) 366
PRECISION 367
Avg Precision Loss (A.P.L.) Chao1 19 (± 4) 10 (± 3) 10 (± 3) 8 (± 2) 7 (± 1) N. A. 368
Avg Precision Loss (A.P.L.) Chao1P 22 (± 4) 12 (± 3) 11 (± 3) 8 (± 2) 7 (± 1) N. A. 369
A.P.L. Chao1P- A.P.L. Chao1 -3 (± 0) -2 (± 0) -1 (± 0) 0 (± 0) 0 (± 0) N. A. 370
371
372
19
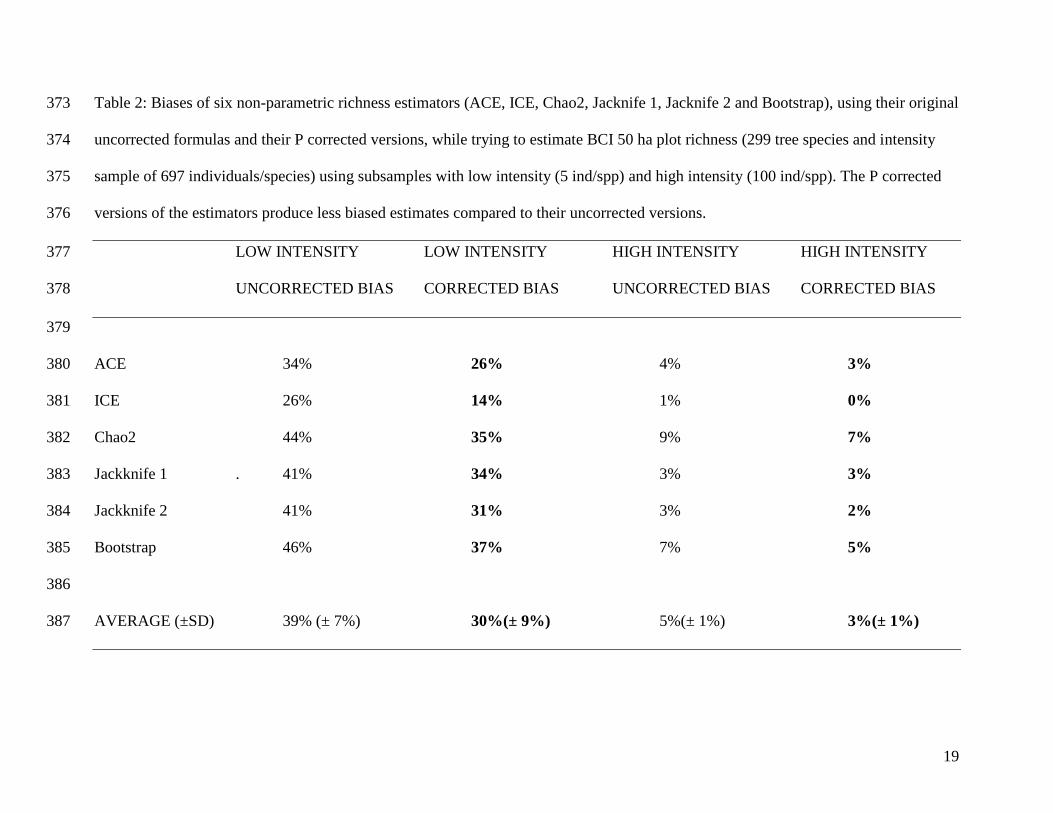
Table 2: Biases of six non-parametric richness estimators (ACE, ICE, Chao2, Jacknife 1, Jacknife 2 and Bootstrap), using their original 373
uncorrected formulas and their P corrected versions, while trying to estimate BCI 50 ha plot richness (299 tree species and intensity 374
sample of 697 individuals/species) using subsamples with low intensity (5 ind/spp) and high intensity (100 ind/spp). The P corrected 375
versions of the estimators produce less biased estimates compared to their uncorrected versions. 376
LOW INTENSITY LOW INTENSITY HIGH INTENSITY HIGH INTENSITY 377
UNCORRECTED BIAS CORRECTED BIAS UNCORRECTED BIAS CORRECTED BIAS 378
379
ACE 34% 26% 4% 3% 380
ICE 26% 14% 1% 0% 381
Chao2 44% 35% 9% 7% 382
Jackknife 1 . 41% 34% 3% 3% 383
Jackknife 2 41% 31% 3% 2% 384
Bootstrap 46% 37% 7% 5% 385
386
AVERAGE (±SD) 39% (± 7%) 30%(± 9%) 5%(± 1%) 3%(± 1%) 387

20
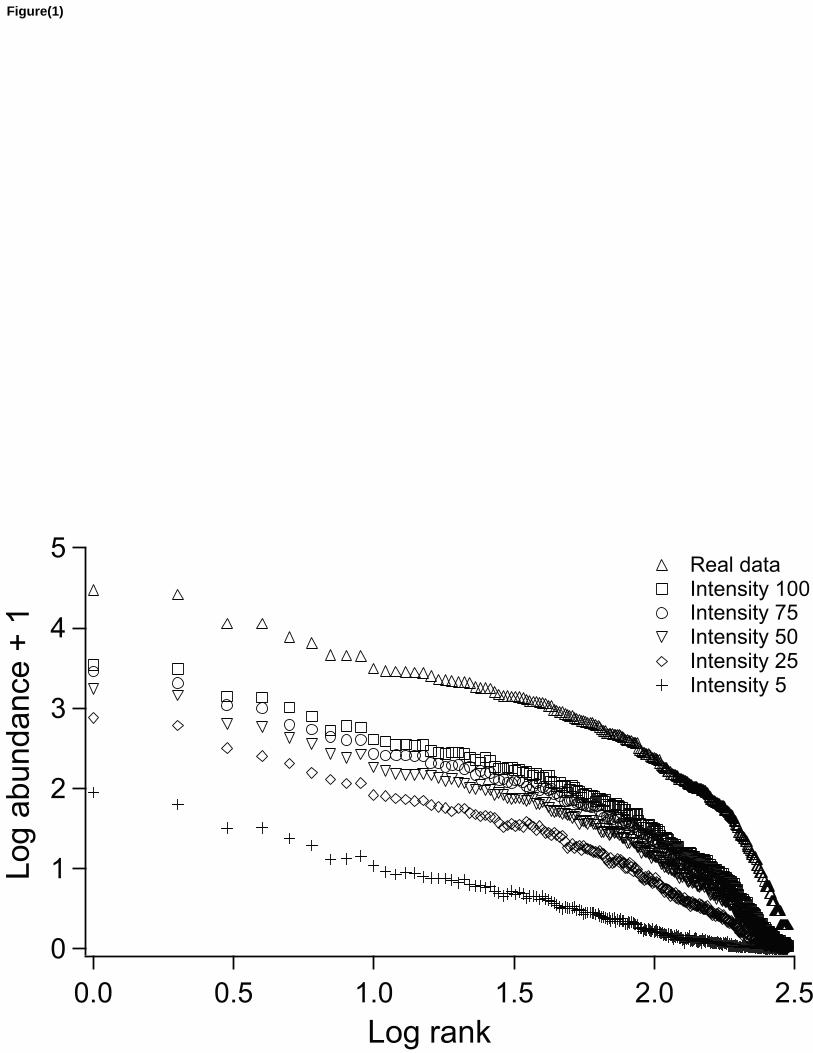
Figure 1: Abundance (log10) of tree species from BCI utilizing real data (census 2005, real 388
intensity 697 ind/spp) and the abundance averages from 100 rarefaction simulations under five 389
sampling intensities (100, 75, 50, 25 and 5 ind/spp). 390
391
Figure 2: Scatterplot between percentage sampling intensity (ind/spp) and percentage of 392
singletons for 185 communities belonging to 4 major taxa (arthropods, corals, trees and 393
vertebrates). The percentage of singletons falls sharply between 0 and 75 ind/spp, but tends to 394
stabilize around 8% singletons above intensity 100. 395
396
Figure 3: Average percentage of original richness estimated with uncorrected Chao1 and 397
corrected Chao1P estimators from rarified simulations with different sampling intensity efforts. 398
At higher intensities, both estimators tend to converge, but Chao1P estimates approaches faster 399
than Chao1 toward 100% of estimation as intensity increased in the six large plots of trees used 400
in the simulations. 401
402
Fig 4- Mean richness (SE bars) estimated by Chao1 and Chao1P estimators obtained from 403
rarefactions of six large plots of tropical trees. The fitted power curves were used to calculate the 404
minimum intensity necessary to estimate 95% and 100% of original richness. Chao1P crosses 405
these thresholds (95% and 100%) with less sampling effort than Chao1. 406
407
Figure 5: Average differences (SE bars) between Chao1 and Chao1P estimates for rarefactions 408
extracted from high intensity samples compared to differences obtained from real samples with 409
original low intensities. The pattern of improved estimation by Chao1P at low intensities 410

21
followed by convergence at higher intensities is very similar between rarefaction simulations and 411
real data. 412
Figure 6 : Comparative performance between corrected (ICEP) and uncorrected (ICE) version of 413
the ICE non-parametric richness estimator trying to predict the richness of BCI 50 ha plot (census 414
2005, richness= 299, intensity= 697 ind/spp). The corrected estimator produced less biased 415
estimates and make better predictions with less sampling effort. 416
5
4
3
2
1
0
2.52.01.51.00.50.0
Real data Intensity 100 Intensity 75 Intensity 50 Intensity 25 Intensity 5
Log rank
Lo
g a
bu
nd
an
ce
+ 1
Figure(1)
80
60
40
20
0
1400120010008006004002000
y=0.6747*x-0.3972
R=0.721
Intensity
Pe
rce
nta
ge
of sin
gle
ton
sFigure(2)
100
80
60
40
Chao1P Chao1
Barro Colorado
Huai Kha Khaeng
100
80
60
40
Ituri Edoro
Korup
10080604020
Pasoh
100
80
60
40
10080604020
Luquillo
Intensity
Estim
ate
d r
ich
ne
ss (
%)
Figure(3)
110
100
90
80
70
60
10080604020
Chao1 Chao1P
y=0.7673*x0.0691
R=0.9945
y=0.5618*x0.1332
R=0.9891
Intensity
Estim
ate
d r
ich
ne
ss (
%)
Figure(4)

20
15
10
5
0
10080604020
Real samples Rarefactions
Intensity
Estim
ate
s d
ife
ren
ce
(%
)Figure(5)
100
90
80
70
60
Estim
ate
d r
ich
ne
ss (
%)
100806040200
Intensity
ICE ICE P
Figure(6)
Table S1: Database utilized in our analyses. We tried to amass a set of data from
literature comprising different taxa with a great variation in sampling intensity.
Inclusive criteria were: have at least one singleton, to allow log transformation of the
singleton percentage, and ten or more species.
Group Taxon Study Site Indiv. Rich. Intensity % Singl. Source
A Araneae Brazil 75 62 1.2 52% (Coddington et al. 2009)
A Araneae Peru 222 123 1.8 63% (Coddington et al. 2009)
A Araneae Mt. Cameroon 573 231 2.5 40% (Coddington et al. 2009)
A Araneae Peru 1821 635 2.9 54% (Coddington et al. 2009)
A Araneae Bolivia 1109 329 3.4 45% (Coddington et al. 2009)
A Araneae Bolivia 654 158 4.1 44% (Coddington et al. 2009)
A Araneae Bolivia 875 191 4.6 47% (Coddington et al. 2009)
A Araneae Peru 5895 1140 5.2 46% (Coddington et al. 2009)
A Araneae Peru 2616 498 5.3 42% (Coddington et al. 2009)
A Araneae Malaysia 6999 578 12.1 25% (Coddington et al. 2009)
A Araneae Guyana 5964 351 17.0 29% (Coddington et al. 2009)
A Araneae Tobago 1777 98 18.1 28% (Coddington et al. 2009) A Araneae Mt. Cameroon 1555 55 28.3 25% (Coddington et al. 2009)
A Araneae Tanzania 5233 149 35.1 23% (Coddington et al. 2009)
A Araneae Tanzania 9096 170 53.5 19% (Coddington et al. 2009) A Araneae Costa Rica 7144 86 83.1 13% (Coddington et al. 2009) A Arthropds Australia 20507 759 27.0 36% (Coddington et al. 2009) A Blattaria Panama 3224 79 40.8 19% (Coddington et al. 2009) V Chiroptera Brazil 99 16 6.2 44% (Luz et al. 2009)
V Chiroptera Brazil 81 13 6.2 15% (Guedes et al. 2000)
V Chiroptera Guinea 107 17 6.3 65% (Fahr et al. 2006)
V Chiroptera Brazil 139 22 6.3 50% (Bordignon 2006)
V Chiroptera Brazil 121 19 6.4 21% (Martins et al. 2006)
V Chiroptera Trinidad and Tobago 143 22 6.5 45% (Clarke and Downie 2001)
V Chiroptera Brazil 268 35 7.7 40% (Martins et al. 2006)
V Chiroptera Brazil 186 21 8.9 52% (Gregorin et al. 2008)
V Chiroptera Ecuador 289 30 9.6 33% (Rex et al. 2008)
V Chiroptera Peru 500 47 10.6 17% (ASCORRA et al. 1996)
V Chiroptera Brazil 178 16 11.1 38% (Tavares et al. 2007)
V Chiroptera Brazil 470 39 12.1 36% (Martins et al. 2006)
V Chiroptera Brazil 231 17 13.6 24% (Gargaglioni et al. 1998)
V Chiroptera Costa Rica 568 40 14.2 35% (Rex et al. 2008)
V Chiroptera Brazil 539 36 15.0 22% Bernard 2001
V Chiroptera Ecuador 895 58 15.4 19% (Rex et al. 2008)
V Chiroptera Honduras 568 35 16.2 26% (Estrada-Villegas et al. 2007)
V Chiroptera Bolivia 396 24 16.5 25% (Loayza and Loiselle 2009)
V Chiroptera Brazil 368 22 16.7 32% (Passos et al. 2003)
V Chiroptera Colombia 244 12 20.3 8% (Sanchez et al. 2007)
V Chiroptera Mexico 338 15 22.5 20% (Chavez and Ceballos 2001)
V Chiroptera Brazil 655 28 23.4 11% (Dias and Peracchi 2008)
V Chiroptera Brazil 659 24 27.5 21% (Cruz et al. 2007)
V Chiroptera Brazil 671 24 28.0 21% (Dias et al. 2002)
V Chiroptera Brazil 390 13 30.0 31% (Camargo et al. 2009)
V Chiroptera Brazil 758 25 30.3 20% (Zortea and Alho 2008)
V Chiroptera Kenya 495 15 33.0 13% (Webala et al. 2004)
V Chiroptera French Guiana 2414 65 37.1 15% (Simmons and Voss 1998)
V Chiroptera Colombia 509 13 39.2 15% (Sanchez et al. 2007)
V Chiroptera Mexico 929 22 42.2 27% (Avila-Cabadilla et al. 2009)
V Chiroptera Brazil 752 14 53.7 21% (Bianconi et al. 2004)
V Chiroptera Mexico 1134 17 66.7 6% (Montiel et al. 2006)
V Chiroptera Bolivia 2548 36 70.8 28% (Espinoza et al. 2008)
Supplementary Material
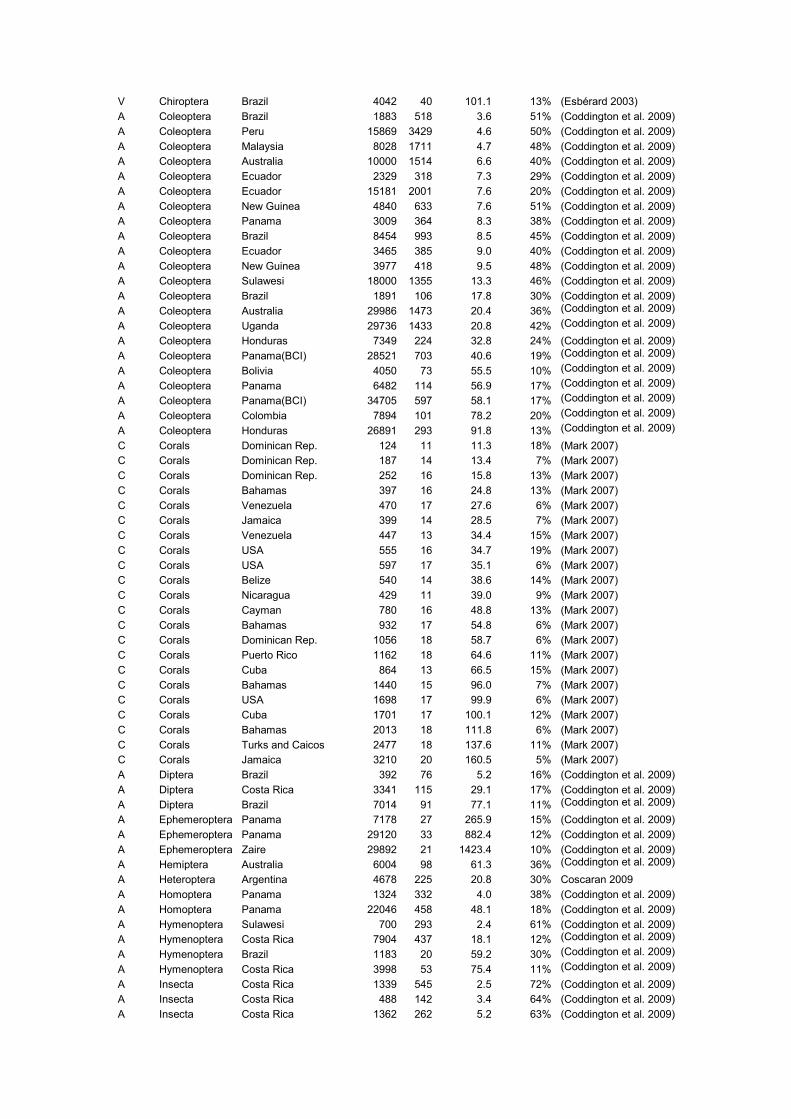
V Chiroptera Brazil 4042 40 101.1 13% (Esbérard 2003)
A Coleoptera Brazil 1883 518 3.6 51% (Coddington et al. 2009)
A Coleoptera Peru 15869 3429 4.6 50% (Coddington et al. 2009)
A Coleoptera Malaysia 8028 1711 4.7 48% (Coddington et al. 2009)
A Coleoptera Australia 10000 1514 6.6 40% (Coddington et al. 2009)
A Coleoptera Ecuador 2329 318 7.3 29% (Coddington et al. 2009)
A Coleoptera Ecuador 15181 2001 7.6 20% (Coddington et al. 2009)
A Coleoptera New Guinea 4840 633 7.6 51% (Coddington et al. 2009)
A Coleoptera Panama 3009 364 8.3 38% (Coddington et al. 2009)
A Coleoptera Brazil 8454 993 8.5 45% (Coddington et al. 2009)
A Coleoptera Ecuador 3465 385 9.0 40% (Coddington et al. 2009)
A Coleoptera New Guinea 3977 418 9.5 48% (Coddington et al. 2009)
A Coleoptera Sulawesi 18000 1355 13.3 46% (Coddington et al. 2009)
A Coleoptera Brazil 1891 106 17.8 30% (Coddington et al. 2009)
A Coleoptera Australia 29986 1473 20.4 36% (Coddington et al. 2009) A Coleoptera Uganda 29736 1433 20.8 42% (Coddington et al. 2009) A Coleoptera Honduras 7349 224 32.8 24% (Coddington et al. 2009)
A Coleoptera Panama(BCI) 28521 703 40.6 19% (Coddington et al. 2009) A Coleoptera Bolivia 4050 73 55.5 10% (Coddington et al. 2009) A Coleoptera Panama 6482 114 56.9 17% (Coddington et al. 2009) A Coleoptera Panama(BCI) 34705 597 58.1 17% (Coddington et al. 2009) A Coleoptera Colombia 7894 101 78.2 20% (Coddington et al. 2009) A Coleoptera Honduras 26891 293 91.8 13% (Coddington et al. 2009) C Corals Dominican Rep. 124 11 11.3 18% (Mark 2007)
C Corals Dominican Rep. 187 14 13.4 7% (Mark 2007)
C Corals Dominican Rep. 252 16 15.8 13% (Mark 2007)
C Corals Bahamas 397 16 24.8 13% (Mark 2007)
C Corals Venezuela 470 17 27.6 6% (Mark 2007)
C Corals Jamaica 399 14 28.5 7% (Mark 2007)
C Corals Venezuela 447 13 34.4 15% (Mark 2007)
C Corals USA 555 16 34.7 19% (Mark 2007)
C Corals USA 597 17 35.1 6% (Mark 2007)
C Corals Belize 540 14 38.6 14% (Mark 2007)
C Corals Nicaragua 429 11 39.0 9% (Mark 2007)
C Corals Cayman 780 16 48.8 13% (Mark 2007)
C Corals Bahamas 932 17 54.8 6% (Mark 2007)
C Corals Dominican Rep. 1056 18 58.7 6% (Mark 2007)
C Corals Puerto Rico 1162 18 64.6 11% (Mark 2007)
C Corals Cuba 864 13 66.5 15% (Mark 2007)
C Corals Bahamas 1440 15 96.0 7% (Mark 2007)
C Corals USA 1698 17 99.9 6% (Mark 2007)
C Corals Cuba 1701 17 100.1 12% (Mark 2007)
C Corals Bahamas 2013 18 111.8 6% (Mark 2007)
C Corals Turks and Caicos 2477 18 137.6 11% (Mark 2007)
C Corals Jamaica 3210 20 160.5 5% (Mark 2007)
A Diptera Brazil 392 76 5.2 16% (Coddington et al. 2009)
A Diptera Costa Rica 3341 115 29.1 17% (Coddington et al. 2009)
A Diptera Brazil 7014 91 77.1 11% (Coddington et al. 2009) A Ephemeroptera Panama 7178 27 265.9 15% (Coddington et al. 2009)
A Ephemeroptera Panama 29120 33 882.4 12% (Coddington et al. 2009)
A Ephemeroptera Zaire 29892 21 1423.4 10% (Coddington et al. 2009)
A Hemiptera Australia 6004 98 61.3 36% (Coddington et al. 2009) A Heteroptera Argentina 4678 225 20.8 30% Coscaran 2009
A Homoptera Panama 1324 332 4.0 38% (Coddington et al. 2009)
A Homoptera Panama 22046 458 48.1 18% (Coddington et al. 2009)
A Hymenoptera Sulawesi 700 293 2.4 61% (Coddington et al. 2009)
A Hymenoptera Costa Rica 7904 437 18.1 12% (Coddington et al. 2009) A Hymenoptera Brazil 1183 20 59.2 30% (Coddington et al. 2009) A Hymenoptera Costa Rica 3998 53 75.4 11% (Coddington et al. 2009) A Insecta Costa Rica 1339 545 2.5 72% (Coddington et al. 2009)
A Insecta Costa Rica 488 142 3.4 64% (Coddington et al. 2009)
A Insecta Costa Rica 1362 262 5.2 63% (Coddington et al. 2009)
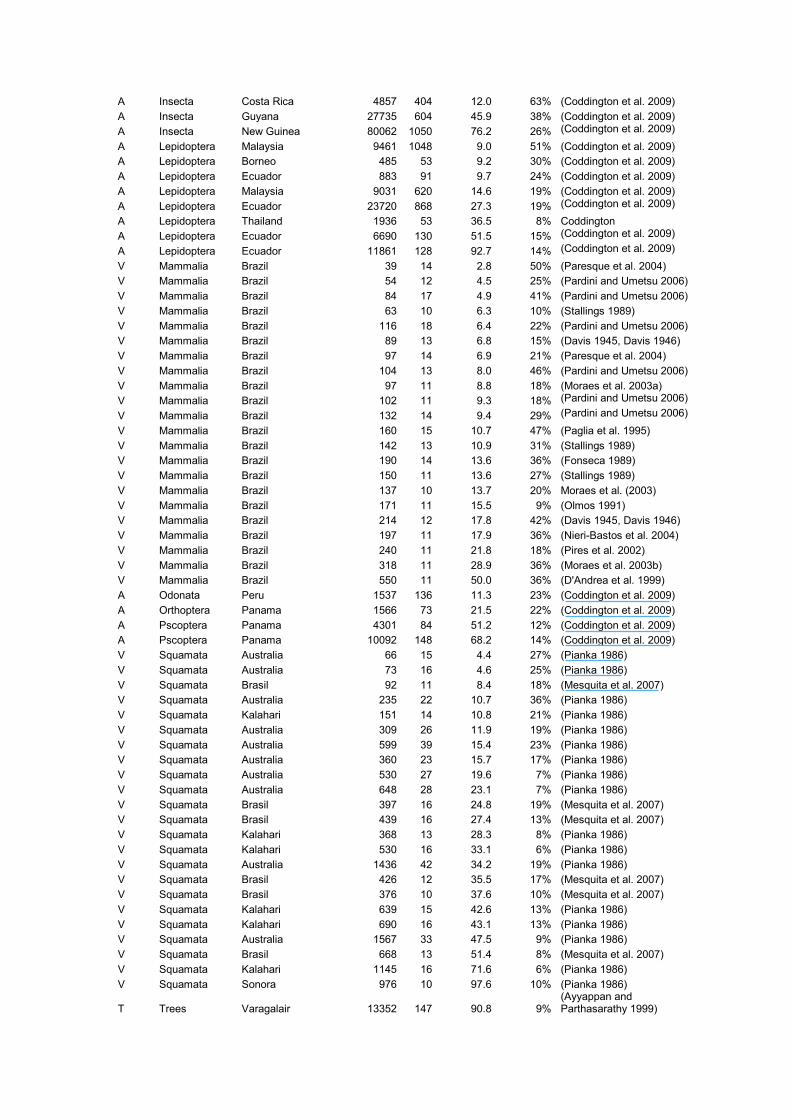
A Insecta Costa Rica 4857 404 12.0 63% (Coddington et al. 2009)
A Insecta Guyana 27735 604 45.9 38% (Coddington et al. 2009)
A Insecta New Guinea 80062 1050 76.2 26% (Coddington et al. 2009) A Lepidoptera Malaysia 9461 1048 9.0 51% (Coddington et al. 2009)
A Lepidoptera Borneo 485 53 9.2 30% (Coddington et al. 2009)
A Lepidoptera Ecuador 883 91 9.7 24% (Coddington et al. 2009)
A Lepidoptera Malaysia 9031 620 14.6 19% (Coddington et al. 2009)
A Lepidoptera Ecuador 23720 868 27.3 19% (Coddington et al. 2009) A Lepidoptera Thailand 1936 53 36.5 8% Coddington
A Lepidoptera Ecuador 6690 130 51.5 15% (Coddington et al. 2009) A Lepidoptera Ecuador 11861 128 92.7 14% (Coddington et al. 2009) V Mammalia Brazil 39 14 2.8 50% (Paresque et al. 2004)
V Mammalia Brazil 54 12 4.5 25% (Pardini and Umetsu 2006)
V Mammalia Brazil 84 17 4.9 41% (Pardini and Umetsu 2006)
V Mammalia Brazil 63 10 6.3 10% (Stallings 1989)
V Mammalia Brazil 116 18 6.4 22% (Pardini and Umetsu 2006)
V Mammalia Brazil 89 13 6.8 15% (Davis 1945, Davis 1946)
V Mammalia Brazil 97 14 6.9 21% (Paresque et al. 2004)
V Mammalia Brazil 104 13 8.0 46% (Pardini and Umetsu 2006)
V Mammalia Brazil 97 11 8.8 18% (Moraes et al. 2003a)
V Mammalia Brazil 102 11 9.3 18% (Pardini and Umetsu 2006) V Mammalia Brazil 132 14 9.4 29% (Pardini and Umetsu 2006) V Mammalia Brazil 160 15 10.7 47% (Paglia et al. 1995)
V Mammalia Brazil 142 13 10.9 31% (Stallings 1989)
V Mammalia Brazil 190 14 13.6 36% (Fonseca 1989)
V Mammalia Brazil 150 11 13.6 27% (Stallings 1989)
V Mammalia Brazil 137 10 13.7 20% Moraes et al. (2003)
V Mammalia Brazil 171 11 15.5 9% (Olmos 1991)
V Mammalia Brazil 214 12 17.8 42% (Davis 1945, Davis 1946)
V Mammalia Brazil 197 11 17.9 36% (Nieri-Bastos et al. 2004)
V Mammalia Brazil 240 11 21.8 18% (Pires et al. 2002)
V Mammalia Brazil 318 11 28.9 36% (Moraes et al. 2003b)
V Mammalia Brazil 550 11 50.0 36% (D'Andrea et al. 1999)
A Odonata Peru 1537 136 11.3 23% (Coddington et al. 2009)
A Orthoptera Panama 1566 73 21.5 22% (Coddington et al. 2009)
A Pscoptera Panama 4301 84 51.2 12% (Coddington et al. 2009)
A Pscoptera Panama 10092 148 68.2 14% (Coddington et al. 2009)
V Squamata Australia 66 15 4.4 27% (Pianka 1986)
V Squamata Australia 73 16 4.6 25% (Pianka 1986)
V Squamata Brasil 92 11 8.4 18% (Mesquita et al. 2007)
V Squamata Australia 235 22 10.7 36% (Pianka 1986)
V Squamata Kalahari 151 14 10.8 21% (Pianka 1986)
V Squamata Australia 309 26 11.9 19% (Pianka 1986)
V Squamata Australia 599 39 15.4 23% (Pianka 1986)
V Squamata Australia 360 23 15.7 17% (Pianka 1986)
V Squamata Australia 530 27 19.6 7% (Pianka 1986)
V Squamata Australia 648 28 23.1 7% (Pianka 1986)
V Squamata Brasil 397 16 24.8 19% (Mesquita et al. 2007)
V Squamata Brasil 439 16 27.4 13% (Mesquita et al. 2007)
V Squamata Kalahari 368 13 28.3 8% (Pianka 1986)
V Squamata Kalahari 530 16 33.1 6% (Pianka 1986)
V Squamata Australia 1436 42 34.2 19% (Pianka 1986)
V Squamata Brasil 426 12 35.5 17% (Mesquita et al. 2007)
V Squamata Brasil 376 10 37.6 10% (Mesquita et al. 2007)
V Squamata Kalahari 639 15 42.6 13% (Pianka 1986)
V Squamata Kalahari 690 16 43.1 13% (Pianka 1986)
V Squamata Australia 1567 33 47.5 9% (Pianka 1986)
V Squamata Brasil 668 13 51.4 8% (Mesquita et al. 2007)
V Squamata Kalahari 1145 16 71.6 6% (Pianka 1986)
V Squamata Sonora 976 10 97.6 10% (Pianka 1986)
T Trees Varagalair 13352 147 90.8 9% (Ayyappan and Parthasarathy 1999)

T Trees Yasuni 2000 145708 1087 134.0 4% (CTFS 2009)
T Trees Huai Kha Khaeng 1999 72604 278 261.2 10% (CTFS 2009)
T Trees Mundamalai 2000 17995 67 268.6 6% (CTFS 2009)
T Trees Lambir 1997 355419 1162 305.9 2% (CTFS 2009)
T Trees Pasoh 2000 296201 816 363.0 3% (CTFS 2009)
T Trees Ituri 2000 144427 350 412.6 10% (CTFS 2009)
T Trees La planada 105163 221 475.9 4% (CTFS 2009)
T Trees Luqillo 1995 67465 137 492.4 12% (CTFS 2009)
T Trees Korup1998 329026 495 664.7 7% (CTFS 2009)
T Trees BCI 2005 208387 299 696.9 6% (CTFS 2009)
T Trees Sinajhara 2001 190707 205 930.3 3% (CTFS 2009)
Ascorra, C. F., Solari, S.,Wilson, D. E. 1996. Diversidad y Ecologia de los Quiropteros
en Pakitza. — In: Wilson, D. E.Sandoval, A. (eds), Manu: The Biodiversity of
Southeastern Peru. Smithsonian Institution Press, pp. 593-612.
Avila-Cabadilla, L. D., Stoner, K. E., Henry, M.,Anorve, M. Y. A. 2009. Composition,
structure and diversity of phyllostomid bat assemblages in different successional
stages of a tropical dry forest. Forest Ecology and Management 258(6), 986-996.
Ayyappan, N.,Parthasarathy, N. 1999. Biodiversity inventory of trees in a large-scale
permanent plot of tropical evergreen forest at Varagalaiar, Anamalais,Western
Ghats, India. Biodiversity and Conservation 81, 533–1554.
Bianconi, G. V., Mikich, S. B.,Pedro, W. A. 2004. Diversidade de morcegos
(Mammalia, Chiroptera) em remanescentes florestais do município de Fênix,
noroeste do Paraná, Brasil. Revista Brasileira de Zoologia 21(4), 943–954.
Bordignon, M. O. 2006. Bat diversity (Mammalia, Chiroptera) from Apore-Sucuriu's
complex, Mato Grosso do Sul, Brazil. Revista Brasileira de Zoologia 23(4),
1002-1009.
Camargo, G., Fischer, E., Gonçalves, F., Fernandes, G.,Ferreira, S. 2009. Morcegos do
Parque NAcional da Serra da Bodoquena, Mato Grosso do Sul, Brasil.
Chiroptera Neotropical 15(1), 417-424.
Chavez, C.,Ceballos, G. 2001. Diversidad y abundancia de murciélagos en selvas secas
de estacionalidad contrastante en el oeste de México. Revista Mexicana de
Mastozoologia 5, 27-44.
Clarke, F. M.,Downie, J. R. 2001. A bat (Chiroptera) survey of mora rainforest in
Trinidad's Victoria-Mayaro Forest Reserve. Biodiversity and Conservation
10(5), 725-736.
Coddington, J. A., Agnarsson, I., Miller, J. A., Kuntner, M.,Hormiga, G. 2009.
Undersampling bias: the null hypothesis for singleton species in tropical
arthropod surveys. Journal of Animal Ecology 78(3), 573-584.
Cruz, L. D., Martínez, C.,Fernandez, F. R. 2007. Comunidades de morcegos em hábitats
de uma Mata Amazônica remanescente na Ilha de São Luís, Maranhão. Acta
Amazonica 37(4), 613 - 620.
CTFS 2009. Center for Tropical Forest Science.(http://www.ctfs.si.edu/).
D'Andrea, P. S., Gentile, R., Cerqueira, R., Grelle, C. E. V., Horta, C.,Rey, L. 1999.
Ecology of small mammals in a Brazilian rural area. Revista Brasileira De
Zoologia 16(3), 611-620.
Davis, D. E. 1945. The home range of some Brazilian mammals. Journal of
Mammalogy. 26(2), 119-127.
Davis, D. E. 1946. The annual cycle of plants, mosquitoes, birds and mammals in two
brazilian forests. Ecological.Monographs. 16, 245-295.

Dias, D.,Peracchi, A. L. 2008. Quirópteros da Reserva Biológica do Tinguá, estado do
Rio de Janeiro, sudeste do Brasil (Mammalia: Chiroptera). Revista Brasileira de
Zoologia 25(2), 333–369.
Dias, D., Peracchi, A. L.,Silva, S. S. P. 2002. Quirópteros do Parque Estadual da Pedra
Branca, Rio de Janeiro, Brasil (Mammalia, Chiroptera). Revista Brasileira de
Zoologia 19(Supl. 2), 113 - 140.
Esbérard, C. E. L. 2003. Diversidade de morcegos em área de Mata Atlântica
regenerada no sudeste do Brasil. Revista Brasileira de Zoociencias 5(2), 189-
204.
Espinoza, A. V., Aguirre, L. F., Galarza, M. I.,Gareca, E. 2008. Ensamble de
murciélagos en sitios con diferente grado de perturbación en un bosque montano
del parque nacional carrasco, Bolivia. Mastozoologia Neotropical 15(2), 297-
308.
Estrada-Villegas, S., Allen, L., García, M., Hoffmann, M.,Munroe, M. L. 2007. Bat
assemblage composition and diversity of the Cusuco National Park, Honduras.
Operation Wallacea, pp. 1-6.
Fahr, J., Djossa, B. A.,Vierhaus, H. 2006. Rapid assessment of bats (Chiroptera) in
Déré, Diécké and Mt. Béro classified forests, southeastern Guinea; including a
review of the distribution of bats in Guinée Forestière. — In: Wright, H.
E.McCullough, J.Alonso, L. E.Diallo, M. S. (eds), A Rapid Biological
Assessment of Three Classified Forests in Southeastern Guinea. Conservation
International, pp. 168-180, 245-47.
Fonseca, G. A. B. 1989. Small mammal species diversity in Brazilian tropical primary
and secundary forests of different sizes. Revista Brasileira.de Zoologia. 6(3),
381-422.
Gargaglioni, L. H., Batalhão, M. E., Lapenta, M. J., Carvalho, M. F., Rossi, R.
V.,Veruli, V. P. 1998. Mamíferos da Estação ecológica de Jataí, Luiz Antônio,
São Paulo. Papeis avulsos de zoologia 40(17), 267-287.
Gregorin, R., Capusso, G. L.,Furtado, V. R. 2008. Geographic distribution and
morphological variation in Mimon bennettii (Chiroptera, Phyllostomidae).
Iheringia Serie Zoologia 98(3), 404-411.
Guedes, P. G., Silva, S. S. P., Camardella, A. R., Abreu, M. F. G., Borges-Nojosa, D.
M., Silva, J. A. G.,Silva, A. A. 2000. Diversidade de mamíferos do parque
nacional de Ubajara (Ceará, Brasil). Mastozoologia Neotropical 7(2), 95-100.
Loayza, A. P.,Loiselle, B. A. 2009. COMPOSITION AND DISTRIBUTION OF A
BAT ASSEMBLAGE DURING THE DRY SEASON IN A NATURALLY
FRAGMENTED LANDSCAPE IN BOLIVIA. Journal of Mammalogy 90(3),
732-742.
Luz, J. L., Costa, L. M., Lourenço, E. C., Gomes, L. A. C.,Esbérard, C. E. L. 2009. Bats
from the Restinga of Praia das Neves, state of Espírito Santo, Southeastern
Brazil. Check List 5(2), 364–369.
Mark, K. W. 2007. AGRRA Database, version (10/2007).
(http://www.agrra.org/Release_2007-10/).
Martins, A. C. M., Bernard, E.,Gregorin, R. 2006. Rapid biological surveys of bats
(Mammalia, Chiroptera) in three conservation units in Amapa, Brazil. Revista
Brasileira de Zoologia 23(4), 1175-1184.
Mesquita, D. O., Colli, G. R.,Vitt, L. J. 2007. Ecological release in lizard assemblages
of neotropical savannas. Oecologia 153(1), 185-195.

Montiel, S., Estrada, A.,Leon, P. 2006. Bat assemblages in a naturally fragmented
ecosystem in the Yucatan Peninsula, Mexico: species richness, diversity and
spatio-temporal dynamics. Journal of Tropical Ecology 22, 267-276.
Moraes, L. B. d., Bossi, D. E. P.,Linhares, A. X. 2003a. Siphonaptera parasites of wild
rodents and marsupials trapped in three mountain ranges of the Atlantic Forest
in Southeastern Brazil. Memórias do Instituto Oswaldo Cruz 98(8), 1071-1076.
Moraes, L. B. d., Bossi, D. E. P.,Linhares, A. X. 2003b. Siphonaptera parasites of wild
rodents and marsupials trapped in three mountain ranges of the Atlantic Forest
in Southeastern Brazil. Memorias Do Instituto Oswaldo Cruz 98(8), 1071-1076.
Nieri-Bastos, F. A., Battesti, D. M. B., Linardi, P. M., Amaku, M., Marcili, A.,
Favorito, S. E.,Rocha, R. P. 2004. Ectoparasites on wild rodents from Parque
Estadual da Cantareira, SÆo Paulo, Brazil. Revista Brasileira de Parasitologia
Veterin ria 13(1), 29-35.
Olmos, F. 1991. Observations on the behavior and population dynamics of some
brazilian Atlantic Forest rodents. Mammalia. 55(4), 555-565.
Paglia, A. P., de Marco, J., Costa, F. M., Pereira, R. F.,Lessa, G. 1995. Heterogeneidade
estrutural e diversidade de pequenos mam¡feros em um fragmento de mata
secund ria de Minas Gerais, Brasil. Revista Brasileira.de Zoologia. 12(1), 67-79.
Pardini, R.,Umetsu, F. 2006. Pequenos mam¡feros nao-voadores da Reserva Florestal
do Morro Grande: distribuicao das especies e da diversidade em uma rea de
Mata Atlantica. Biota Neotropica (Online) 6
Paresque, R., Souza, W. P., Mendes, S. L.,Fagundes, V. 2004. Composicao cariotipica
da fauna de roedores e marsupiais de duas reas de Mata Atlantica do Espirito
Santo, Brasil. Boletim do Museu de Biologia Mello Leitao Nova Serie. 175-33.
Passos, F. C., Silva, W. R., Pedro, W. A.,Bonin, M. R. 2003. Frugivoria em morcegos
(Mammalia, Chiroptera) no Parque Estadual Intervales, sudeste do Brasil.
Revista Brasileira de Zoologia 20(3), 511–517.
Pianka, E. R. 1986. Ecology and Natural History of Desert Lizards: Analyses of the
Ecological Niche and Community Structure. Princeton Univ. Press.
Pires, A. S., Lira, P. K., Fernandez, F. A. S., Schittini, G. M.,Oliveira, L. C. 2002.
Frequency of movements of small mammals among Atlantic Coastal Forest
fragments in Brazil. Biological Conservation 108(2), 229-237.
Rex, K., Kelm, D. H., Wiesner, K., Kunz, T. H.,Voigt, C. C. 2008. Species richness and
structure of three Neotropical bat assemblages. Biological Journal of the
Linnean Society 94(3), 617-629.
Sanchez, F., Alvarez, J., Ariza, C.,Cadena, A. 2007. Bat assemblage structure in two dry
forests of Colombia: Composition, species richness, and relative abundance.
Mammalian Biology 72(2), 82-92.
Simmons, N. B.,Voss, R. S. 1998. The mammals of Paracou, French Guiana: A
neotropical lowland rainforest fauna part - 1. Bats. Bulletin of the American
Museum of Natural History(237), 1-219.
Stallings, J. R. 1989. Small mammal inventories in an eastern Brazilian park.
Bull.Florida State Mus.Biol.Sci. 34(4), 153-200.
Tavares, V. C., Perini, F. A.,Lombardi, J. A. 2007. The bat communities (Chiroptera) of
the Parque Estadual do Rio Doce, a large remnant of Atlantic Forest in
southeastern Brazil. Lundiana 8(1), 35-47.
Webala, P. W., Oguge, N. O.,Bekele, A. 2004. Bat species diversity and distribution in
three vegetation communities of Meru National Park, Kenya. African Journal of
Ecology 42(3), 171-179.

Zortea, M.,Alho, C. J. R. 2008. Bat diversity of a Cerrado habitat in central Brazil.
Biodiversity and Conservation 17(4), 791-805.




















