
The Protector Species Hypothesis: Do Black Skimmers Find Refuge from Predators in Gull-billed Tern...
12
Ethology 104, 273-284 (1998) 0 1998 Blackwell Wissenschafts-Verlag, Berlin ISSN 0179-1613 Department of Biology, Universig of Sotitbwestem Imisiana, 1.ufayette The Protector Species Hypothesis: Do Black Skimmers Find Refuge from Predators in Gull-billed Tern Colonies? SANDRA M. PIUS & PAUL L. LEBERG PIUS, S. M. & LEBERG, P. L. 1998: The protector species hypothesis: do black skimmers find refuge from predators in gull-hilled tern colonies?. Ethology 104, 271-284. Abstract The protector-species hcothesis explains mixed-species coloniality on the basis of benefit.; individuals of a species may receive by nesting with another species, the ‘protector’ species, that responds agressively to potential threats. The reactions of nesting individuals to both natural and model predators wc-rc observed to determine whether black skimmers (R/@Io~J n@r) gain an antipredator advantage by nesting with pll-bilied terns (.Vetemu ndoticu). Observations of natural predators were gathered from three mixed-species and three single- species (black skimmers) subcolonies. Natural predators most commonly encountered by the colonies were herring gulls (IMW u?qentutus), laughing gulls lam^ atncdla), and ruddy turnstones (Annuria intepns). Gull-billed terns responded to the gulls, but not to the turnstones, in higher proporuons than did black skimmers. Two decoys, a mink and a gull, were used to simulate predatory encounters, and a duck decoy was used as a control at two mixed-species and one single-species subcolonies. Gull-billed terns responded in significantly higher proportions than did skimmers to all decoy treatments in the mixed-species subcolonies. Mobbing of both natural and model predators by the terns suggests that skimmers may gain a reproductive advantage 1)) nesting with these terns. However, the response of black skimmers to both natural and simulated predators was independent of the presence of gd-billed terns in the colony, indicating that black skimmers may nut perceive these objects as threats, or may react differently to predators than do gull-billed terns. Corresponding author: Paul L. LEBERG, Department of Biology, University of Southwestern Louisiana, Lafayette, LA 70504, USA. E-mail: [email protected] Introduction Individuals form groups for the purposes of breechng, feeding, migrating, and resting (KUMMER 1968; MECH 1970; SCHALLER 1972; ALEXANDER 1974; HOOGL~ND 1979; PITCHER et al. 1982; WITTENBERGER & HUNT 1985; GOTMARK et al. 1986: HARRISON et al. 1991; CARRASCAL & MORENO 1992; GROVER & O L ~ 1993). Groups may also form because suitable habitat has a clumped distribution (KUMMER 1908). However, in most cases, individuals of a group benefit from being with other individuals and display social attraction and cohesiveness (BURGER & GOCHFELD 1990). Nesting in large groups is thought to be one adaptation to predation pressures (LACK 1968). Although large groups of breeding individuals may act as a beacon for predators, the effect of predation may be buffered through camouflage coloration of nests and eggs FINBF.KGF.N et al. 1967), by synchronous breeding (NISHET 1975), and by antipredator behavior of the group (CAIRNS C. S. Copyright Clearance Center Code Statement: 01 79-161 3/98/1 O4&O273$14.00/0
Transcript of The Protector Species Hypothesis: Do Black Skimmers Find Refuge from Predators in Gull-billed Tern...

Ethology 104, 273-284 (1998) 0 1998 Blackwell Wissenschafts-Verlag, Berlin ISSN 0179-1613
Department of Biology, Universig of Sotitbwestem Imisiana, 1.ufayette
The Protector Species Hypothesis: Do Black Skimmers Find Refuge from Predators in Gull-billed Tern Colonies?
SANDRA M. PIUS & PAUL L. LEBERG
PIUS, S. M. & LEBERG, P. L. 1998: The protector species hypothesis: do black skimmers find refuge from predators in gull-hilled tern colonies?. Ethology 104, 271-284.
Abstract
The protector-species hcothesis explains mixed-species coloniality on the basis of benefit.; individuals of a species may receive by nesting with another species, the ‘protector’ species, that responds agressively to potential threats. The reactions of nesting individuals to both natural and model predators wc-rc observed to determine whether black skimmers (R/@Io~J n@r) gain an antipredator advantage by nesting with pll-bilied terns (.Vetemu ndoticu). Observations of natural predators were gathered from three mixed-species and three single- species (black skimmers) subcolonies. Natural predators most commonly encountered by the colonies were herring gulls (IMW u?qentutus), laughing gulls lam^ atncdla), and ruddy turnstones (Annuria intepns). Gull-billed terns responded to the gulls, but not to the turnstones, in higher proporuons than did black skimmers. Two decoys, a mink and a gull, were used to simulate predatory encounters, and a duck decoy was used as a control at two mixed-species and one single-species subcolonies. Gull-billed terns responded in significantly higher proportions than did skimmers to all decoy treatments in the mixed-species subcolonies. Mobbing of both natural and model predators by the terns suggests that skimmers may gain a reproductive advantage 1)) nesting with these terns. However, the response of black skimmers to both natural and simulated predators was independent of the presence of gd-billed terns in the colony, indicating that black skimmers may nut perceive these objects as threats, or may react differently to predators than do gull-billed terns.
Corresponding author: Paul L. LEBERG, Department of Biology, University of Southwestern Louisiana, Lafayette, LA 70504, USA. E-mail: [email protected]
Introduction
Individuals form groups for the purposes of breechng, feeding, migrating, and resting (KUMMER 1968; MECH 1970; SCHALLER 1972; ALEXANDER 1974; H O O G L ~ N D 1 9 7 9 ;
P ITCHER et al. 1982; WITTENBERGER & HUNT 1985; GOTMARK et al. 1986: HARRISON et al. 1991; CARRASCAL & MORENO 1992; GROVER & O L ~ 1993). Groups may also form because suitable habitat has a clumped distribution (KUMMER 1908). However, in most cases, individuals of a group benefit from being with other individuals and display social attraction and cohesiveness (BURGER & GOCHFELD 1990). Nesting in large groups is thought to be one adaptation to predation pressures (LACK 1968). Although large groups of breeding individuals may act as a beacon for predators, the effect of predation may be buffered through camouflage coloration of nests and eggs FINBF.KGF.N et al. 1967), by synchronous breeding (NISHET 1975), and by antipredator behavior of the group (CAIRNS
C. S. Copyright Clearance Center Code Statement: 01 79-1 61 3/98/1 O4&O273$14.00/0

274 S. M. P1r.s & P. L. LEHBRG
1992). Antipredator behavior, such as increased vigilance (TINBERGFX 1952; ELGAR 1989) and mobbing (GAINS 1992), should become more effective with an increase in group size. Also, as group size increases, it reduces an individual’s risk of predation (HAMILTON 1971).
Costs of predation may be further lowered if individuals of a species join a hetero- specific group in which all species are prone to the same predators but possess different antipredator behavior patterns. Empirical evidence for suggesting the antipredator benefits of heterospecific groups comes from studies of mammals (SINCJAIR 1985; SCHI:EL 1993), fish (MCKAYE & OLIVER 1980), and birds (KRLC‘~< 1964; BGRGER 1984; YC)Uh’G & TIThL4N 1986; BURGER & GOCHFRLD 1990; GROOM 1992). Among birds, more than half of the species that nest in colonies sometimes or always nest in mixed-species groups (BURGER 1981).
If one species of a heterospecific group is more aggressive towards predators than the other species, it is likely that the less aggressive species benefits from the association. This forms the basis of the protector-species hypothesis where members of a species will actively choose to nest in the vicinity of the ‘protector’ species, reducing both their chances of being lulled and their need to respond to predator threats (I(RcuI< 1964; BURGER 1984; YOUNG & TITMAN 1986; BURGER & GOCHFELD 1990). For instance, black skimmers (Rbnchops niger), a colonial-nesting seabird, nest predominantly in colonies of common terns (Sterna hirzlndo) along the beaches of the north-eastern llnited States (ERWIN 1977, 1979; BURGER & GOCHFELD 1990). In the mixed-species colonies, common terns usually respond immelately, aggressively, and in large proportions towards predators. Skimmers in these same colonies are less likely to engage in mobbing behavior and often stay on their nests unless the danger persists. Thus, skimmers not only gain an energetic advantage but also a reproductive one by guarding their nests during a prrdator attack. Furthermore, they lower potential risks of injury and death that might occur during harassment of predators. However, shmmers exhibit some geographic variation in responses to predators; in Peru, both terns and skimmers respond aggressively to avian predators (GROOM 1992). Furthermore, aggressive antipredator behavior m skimmers is dependent on environmental factors, such as the level of threat posed by :I predator and their history of exposure to a predator (BC‘RGER & GOCHFELD 1992).
In coastal Louisiana, where breeding colonies of common terns do not occur, black skimmers nest predominantly with another seabird, the gul!-billed tern, Sterna nzlotica (MARTIN & LESTER 1990). In a state-wide survey, all 16 colonies of gull-hilled terns had black shmmers nesting in them (MARTIN & LESTER 1990); however, unlike the situation in the Northeastern United States (BURGER & GOCHFELD 1990), black skimmers in Louisiana sometimes nested in the absence of terns. The gull-billed terns, like the common terns in the Northeast, arrive at the breeding site and start to nest before the black slummers. This suggests that gull-billed terns show no ‘preference’ for nesting with black skimmers, while it is possible that black skimmers may actively choose to nest with gull- billed terns.
Our objective was to determine if gull-billed terns behave as a ‘protector’ species for skimmers that nest with them and to examine antipredator behavior of skimmers in the presence and absence of gull-billed terns. Evidence for the protector species hypothesis comes largely from studies on ducks nesting with terns or gulls (BOIJRGET 1973;

Response of Seabirds to Predators 275
NUECHTERLEIN 1981; BURGER 1 9 8 4 ; YOUNG & TITMAN 1986), ducks nesting with geese (GIROUX 1981; ROBERTSON 1995), shorebirds and phalaropes nesting with terns (HOHN 1967; ALBERICO et al. 1991), nighthawks and seabirds (GROOM 1992), and seabirds nesting with other species of seabirds (CULLEN 1958; LIED 1963; FUCHS 1977; VEEN 1977; ERVC’IN 1979; GOTMARK & ANDEKSSON 1980; BURGER & G O C H F E L D 1990). If a species dtd confer protection from predators to the heterospecifics nesting with it, then, in the absence of such a ‘protector’ species, either the risk of predation or the anupredator response of the remaining species is expected to increase. A literature survey identified 21 studies of the protector-species hypothesis in birds, but we found that studies comparing both single- and mixed-species groups of birds were r u e (NF,WTON & CAMPELL 1975; GIROUX 1981; BURGER 1984; GOTMARK & AHLUND 1986; Y O U N G & TITMAN 1986; ROBERTSON 1995); the majority of the studies examine only mixed-species colonies. Most of these studies were on ducks or grebes, which typically do not mob predators, nesting with colonial species. None of the seven papers that addressed two or more colonial species of seabirds in the mixed-species groups used single-species colonies of the ‘protected’ species for comparisons (CCLJ~JX 1958; LIND 1963; FCCHS 1977; VEEN 1977; ERWIN 1979; GOTMARK & AXDERSSON 1980; BURGER & GOCHPELD 1990). The availability of single-species subcolonies of black skimmers allowed us to study the antipredator behavior of a species in the presence and absence of its suspected ‘protector’ species, making this study an important contribution to the understanding of not only the protector species hypothesis but also mixed-species coloniality. We also examined whether the association between black skimmers and gull-billed terns is similar to the one proposed by BURGER & GOCHFELD (1990), or if that relationship varies in different regions or with different species associations. The differences in levels of responses of both species to actual and potential threats to the colony are discussed and a lack of aggressiveness toward the only predator observed taking eggs is noted.
Methods
Study Colony and Roost
The Atchafalaya Delta (29’N, 91”Wi) is one of screral areas of suitable nesting habitat for colonial seabirds along Louisiana’s Gulf coast. This delta has many natural and dredge islands. Thc latter are made by the annual dredging of sediments from a shipping canal that runs through the delta. Seabirds ncst o n these dredge islands prior to the establishment of dense vegctation ( L r m w et al. 1997). Approximately 350 gull-billed terns and 1200 black skimmers nest on dredge islands in the Delta, making the breeding populations of thcsc seabirds among the largest along the northern coast of the Gulf of Mexico. Our study was carried out in the summers of 1994 and 1995 on three dredge islands: Skimmer, Donna’s, and Horseshoe Island. Observations were made from approxtmately the second week of May to the first wcek of Aug.
Nesting Characteristics of Study Organisms
In both years of our study, gull-billed terns arrived at the study site about the second week of May and established nests 1-2 wk before the skimmers. Early arriving skimmers occupied nesting sites close to $-billed tern nests and formed the center of the colony from which the later-arriving skimmers spread out in a radial pattern. There were no single-species aggregations within this heterospecific nesting group. About 80-650 breeding pairs of both species nested together during both years in close temporal and spatial association. Typically about 3-5 wk after the start of the main colony, and between 10 m to 40m away from the edgc of the main mixed-species colony, a discrete group of skimmers initiated nesting. We refer to these groups as mixed-

276 S. M. PIUS &. P. L. LEBERG
and single-species subcolonies, the definition o f a ‘subcolony’ being a discrete portion of :I colony separated in space (BURCI-.R & GOcHFI~.r.D 1990).
In 1994, mixed-species subcolonies of black slummers and gull-billed terns formed on both Skimmer Island and Donna’s Island. About 3 4 wk after the mixed-species subcolonies formed, single-species subcolonies of black skimmers appeared, one on each island. The mixed-species subcolony on Skimmer Island and both the single-species subcolonies were used in the 1994 study. In 1995, mixed-species subcolonies formed, one each on Skimmer and Horseshoe Island. When the mixed-species subcolonies were approximately 1 wk old, a single- species subcolony of black skimmers formed on another part of Horseshoe Island. We used all three subcolonies formed in 1995 in this study. Single-species subcolonies in both years were initially spaually isolated by 1011Om from the mixed-species subcolonies on the same island. As both subcolony types recruited more skimmers throughout the breeding season, their respective areas expanded, shortening the distance between them. Although the two types of subcolonies were close in space, they were separated in time; most gull-billed terns had moved with their young away from the colony area towards the edge of the island by the time the single-species subcolonies had completed formation. There were between 50 and 500 black skimmer pairs and 30-1 50 gull- billed tern pairs in the mixed-species subcolonies, and 25-125 pairs of black skimmers i n the single-species subcolonies.
Natural Predators
We observed randomly selected colony types for % 6 h per day (between 07.00 and 20.00 hours) with a spotting scope from an elevated blind about 35 m away. We spent % 245 h observing the subcolonies per breeding season. Every time a predator was observed, we determined its species, the duration of its stay over or in the subcolonies, and the proportions of gull-billed terns and black skimmers that chased and attacked the predator. We used non-parametric statistical procedures because the data did not meet the assumptions of normality and homogeneity o f variances. We tested one-tailed null hypotheses that (1) the proportion of black skimmers mobbing predators was the same o r higher than the proportion of b~ll-billed terns mobbing the same predators, and (2) black skimmers in single-species subcolonies responded to predators in equal or lower proportions than their conspecitics in mixed-species subcolonies. We used contingency tables to test the null hypothesis that there was no association between species or colony type with the proportiun o f predators that elicited mobbing.
Decoy Exposures
Most predators that ehcited a mobbing response from the colony members were observed flying over the colonies. I n an effort to examine other predatory situations, we exposed each colony type t o simulated terrestrial threats using a taxidermy mount of a mink (Mmteh ~ J ~ J O I ? ) and a plastic decoy of a herring ball (Idnns qrniutm), in the 1995 breeding season. The gull decoy was chosen to evaluate how the colony reacted to the presence of this predator on the ground. The mink decoy was chosen to determine the colony response tu a mammalian predator. TI) determine if colony members were instigated to mob because they recogniied the mink and gull decoy as threats, or were just reacting to an object in the colony, we used a plastic duck decoy as a control. As a second control (null), we also monitored colony activity when no decoys o r other thie:its \\-ere present to
determine if the mobbing activity increased when the colony was exposed to a decoy. For each exposure, we randomly picked a decoy along with a colony. Then mc placed the decoy with a
string tied around it under a piece of burlap and positioned it along the edge of the colony. The other end of the string was placed in a clump of bushes or tall grass about 150 m away from the colony. T o minimizc disturbance t o the birds, decoy placement usually took place near sunset. The following day, at a randomly determined daylight time, an investigator crept up to where the end of the string lay. After the birds had stopped mobbing the investigator, the decoy was pulled out slowly from under the burlap s o that it ran ;king the edge o f the colony, and was visible to the nesting birds. The whole episode lasted % 4min and was videotaped from the blind. We divided the breeding srason into three periods, each corresponding to 2-uzk intends. Within each period, we exposed the three subcolonies t o all three decoys and to the null control (a randomly chosen 4-min period, which was videotaped in a fashion identical to that of the decoi- exposures). We later screened each video episode and selected 30 random frames. In each frame, the proportion of each species mobbing the decoys was calculated and for the null episodes, the proportion of each species in the air was calculated. To avoid pseudo- replication (HVRI.HF.RT 1984), we calculated single response variables for each specie\, the statistical means of the proportion of skimmers and proportion of pd-billed terns responding to each decoy q o s u r e .
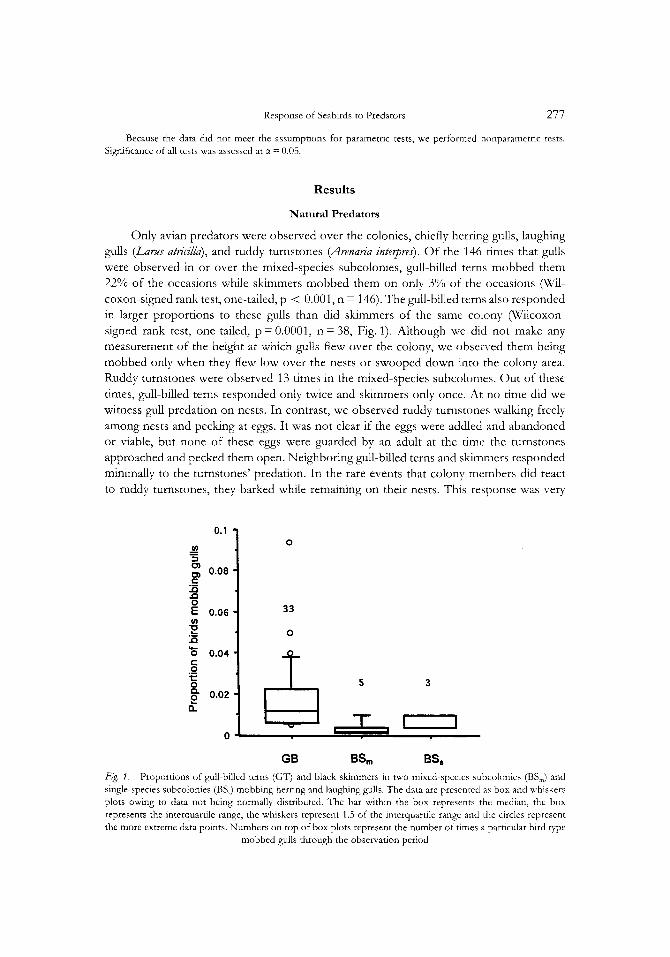
Response o f Seabirds to Predators 277
Because the data did not meet the assumptions for parametric tests, we performed nonparametric tests. Significance of all tests was assessed at LY = 0.05.
Results
Natural Predators
Only avian predators were observed over the colonies, chiefly herring gplls, laughing gulls ( L ~ J atricilla)), and ruddy turnstones (Arenaria zntepm). Of the 146 times that gulls were observed in or over the mixed-species subcolonies, gull-billed terns mobbed them 22% of the occasions while skimmers mobbed them on only 3% of the occasions (Wil- coxon-signed rank test, one-tailed, p < 0.001, n = 146). The gull-billed terns also responded in larger proportions to these gulls than did shmmers of the same colony (Wilcoxon- signed rank test, one-tailed, p=O.O001, n=38, Fig. 1). Although we did not make any measurement of the height at which gulls flew over the colony, we obsened them being mobbed only when they flew low over the nests or swooped down into the colony area. Ruddy turnstones were observed 13 times in the mixed-species subcolonies. Out of these times, gull-billed terns responded only twice and skimmers only once. At no time did we witness gull predation on nests. In contrast, we observed ruddy turnstones walhng freely among nests and pecking at eggs. It was not clear if the eggs were addled and abandoned or viable, but none of these eggs were guarded by an adult at the time the turnstones approached and pecked them open. Neighboring gull-billed terns and skimmers responded minimally to the turnstones’ predation. In the rare events that colony members did react to ruddy turnstones, they barked while remaining on their nests. This response was very
0.1
u)
3 9)
C
0
u)
- - 0.08
z n E 0.06
E a 0 0.04 C 0 iz
Ic
.- 8 0.02 2 n
0
GB BSm BSS @. 1: Proportions of gull-billed terns (GT) and black skimmers in two mixed-species subcolonies (BS,) and single-species subcolonies @S,) mobbing herring and laughing gulls. The data are presented as box and whiskers plots owing to data not being normally distributed. The bar within the box represents the median, the box represents the interquartile range, the whiskers represent 1.5 of the interquarule range and the circles represent the more extreme data points. Numbers on top of box plots represent the number of times a particular bird type
mobbed gulls through the observation period

different from their response to gulls, where they flew up from their nests and chased them.
There was no significant difference between the proportion of times that skimmers in single and mixed-species colony types mobbed gulls (Mann-Whitney-WiIcoxon test, one-tailed, p = 0.137, nsmgIie = 81, nmlncd = 146). Skimmers in single-species subcolonies did not mob in significantly greater proportions when compared with skimmers in mixed- species subcolonies (Mann-Whitney-Wilcoxon test, one-tailed, p = 0.1 72, nrLnglc = 3, nmlxed = 5; Fig. 1). No ruddy turnstones were seen in the single-species subcolonies, mainly because the turnstones had left the study area by the time these subcolonies formed.
0.6 - p 0.5 - 8
P a,
0.4
a n 0.3 - z
v) 2 .- 0.2 ;
g 0 :
'c 0 C .o 0.1 - r 8
Decoys
Shmmers and gull-billed terns in the mixed-species subcolonies were found in greater proportions in the air when decoys were present than when decoys were absent (Mann- Whitney-Wilcoxon test, one-tailed, p 0.0195, nprcscnr = 18, nahScnr = 6). Skimmers in single- species subcolonies were similarly found in larger proportions in the air when in the company of decoys than without (Mann-Whitney-Wilcoxon test, one-tiled, p = 0.01 6, npTcbcnc = 9, nabsent = 3) , indicating that the decoys were viewed as potential threats by the nesting seabirds. Gull-billed terns responded in higher proportions than skimmers of the same colonies to decoy exposures (Wilcoxon-signed rank test, one-tailed, p 0.0001, n = 18, Fig. 2). The response of gull-billed terns differed among the three types of decoys (Friedman two-way analysis of variance, p = 0.050, n = 6). The responses to the mink and gull decoy were similar and much greater than for the duck decoy (Fig.2). In contrast, black skimmers in the single- or mixed-species subcolonies, did not differ in their response to the types of decoys presented (Fig. 2).
.
6
0
T Gull-billed tern
Skimmer-Mixed
Skimmer-Single
6 6 3
2 k L -
Duck Gull Mink Null F>i. 2: Proportions of pd-billed terns and black skimmers in two mixed-species subcolonies, and black skimmers in single-species subcolonies responding to three decoys: mink, duck and p i l l , and to no decoy (Null).
Sample numbers appear above the box plots

Response of Seabirds to Predators 279
The proportion of gull-billed terns responding to decoys dlffered among breeding periods (Friedman two-way analysis of variance, p = 0.002, n = 6, Fig. 3). A multiple com- parison test indicated that the proportions responding to decoys during breeding periods 1 and 2 were similar to each other, and higher and different from breeding period 3 (Fig. 3). The proportion of skimmers responding to decoys did not dlffer between subcolony types (Mann-Whitney-Wilcoxon test, one-tailed, p = 0.1463, nmlxed = 18, nSlllglc = 9, Fig. 2) or among breeding periods (Friedman two-way analysis of variance, p = 0.1 849, nrnlxcLi 6, nslngle = 3, Fig. 3).
Discussion
Many of the studies on mixed-species groups reveal that individuals of one species in the group aggressively protect the entire group from predators. Thus, these ‘protector’ species may be the reason why mixed-species colonies form, attracting other species that are less effective against predators (CULLEN 1958; KRUUK 1964; SMITH 1975; ERWIN 1979; BURGER 1984; ELLIOT 1985; YOUNG & TITMAN 1986). The gull-billed terns a t our study site appear to fit the definition of a ‘protector’ species (KRUUK 1964) when the antipredator behavior of the two species is compared. The gull-billed tern was the predominant species involved in mobbing and chasing both natural predators and predator decoys. As in mixed-species colonies of common terns and skimmers in New York and New Jersey (BURGER & GOCHFELD 1990), in Louisiana, the skimmers participated in lower proportions than the terns, mostly remaining on their nests during a predatory attack. The gull-billed terns successfully chased gulls that visited the colony area; we rarely
en c
0
.- n n E
0 . 6
0.4
0
6
Gull-billed tern
Skimmer-Mixed
Skimmer-Single
6 0 . 3 E .-
n + 0 c 0 .2 0
Q
a
.- 5 2 0.1
0 1 2 3
Breeding Period Fig. 3: Proportions of gull-billed terns and black skimmers in two mlxed-species subcolonies, and black skimmers in single-species subcolonies responding to decoy exposures within each breeding period. Each breeding
period is a 2-wk segment of the breeding season. Sample numbers appear over the bars

280 S. M. Pius & P. L. LEHF.RC;
observed a gull landing in the subcolonies and never observed any nest predation by gulls. Low reproductive success owing to nest predation appears to be common and widespread in seabird colonies frequented by gulls (HULSMAN 1984; BURGER & GOCHFELD 1990; SHEALER & BURGER 1992; EMSL~E et al. 1995). Hence, the reproductive success of the skimmers may be increased by the terns actively warding away gulls.
There was a temporal difference in the response of gull-billed terns to predators. Because the data on natural predators were insufficient, we used only the data from the decoy experiment to examine this temporal variation in response. Tern response towards predators when chicks were present (period 3) was lower compared with when birds were laying and incubating eggs. Although not statistically significant, black skimmers also showed a decline in the proportion of individuals participating in antipredator behavior as the breeding season advanced. Terns and skimmers may be reluctant to leave their nests unguarded with newly hatched chicks. BURGER & GOCHFELD (1990, 1991) report a similar temporal decline in the proportions of terns and skimmers participating in upflights through the breeding season. Because chicks are mobile, and may get lost, hurt or eaten by other colony members or predators, the best strategy for parents may be to stay at their nests for as long as possible. Furthermore, chicks in the latter part of the breeding season find hiding places in nearby vegetation, thereby reducing the importance of mobbing to prevent nest predation.
Ideally, the ‘protector’ species must be able to recognize predators to be an asset to the other species that nest with it. The gull-billed terns appeared to be able to differentiate readily a threat from a non-threat among gulls in the subcolony vicinity and in the experimental decoy treatment. Terns responded more strongly to gulls flying low over the subcolonies or swooping down at nest sites than they did to gulls flying high above the subcolonies, a behavior commonly observed in other tern colonies (BIJKGER & GOCH-
BURGER 1992; EMSLIE et al. 1995) and minks (EMLEN et al. 1966; BUKNESS & MORRIS 1993; BIRKHEAD & NETTLESHIP 1995) are attracted to larid colonies. Terns attacked the mink and gull decoys in significantly larger proportions than they attacked the duck decoy; ducks are not known to pose a threat to terns. However, the skimmers in both subcolony types did not differentiate significantly between the threat and the non-threat decoy; responses to all decoy types were low.
Although gull-billed terns appeared to be protecting the subcolonies as far as gulls and the predator decoys were concerned, neither this species nor the skimmers responded to ruddy turnstones even while this shorebird pecked at nearby eggs and ate the contents that spilled out. To our knowledge, this is the first account of predation by ruddy turnstones on eggs of gukbilled terns. It is possible that ruddy turnstones were pecking at addled e g s abandoned by the breeding pair. However, studies of ruddy turnstones in colonies of other species of the Laridae (larids) indicate that they are capable of being aggressive egg predators, darting and pecking at eggs even while the adult is sitting on the clutch (CROSSIN & HUAER 1970; PARKES et al. 1971; NETTLESHIP 1973; BURGER & GOCHFELD 1991). In one case, turnstones destroyed between 67 and 100%r ofnests in larid colonies (BREAREY & HILDEN 1985), and in another case, they caused an entire colony to abandon the nesting area (LOFTIN 1978). With respect to this nest predator, skimmers may not gain a direct advantage by nesting with gull-billed terns. They may, however, still benefit from having
FELD 1991). Both gulls (HULS~MAN 1984; BURGER & ChCHFE1.D 1c)OO; SHEALER &

Response of Seabirds to Predators 281
their nests interspersed with those of terns; on our study sites, because mixed-species subcolonies tended to be larger than single-species subcolonies, a simple dilution effect could offer protection (HAMILTON 1971). Our data are insufficient to test this hypothesis.
Under the protector-species hypothesis, a species nests with a ‘protector’ t o reduce costs of predator defense or risk of predation. This suggests that a species capable of mobbing would increase antipredator behavior to prevent nest predation if its ‘protector’ is absent. Previous studies of the protector-species hypothesis in seabirds could not examine this prediction because they focused on mixed-species colonies alone (CC12J,EN 1958; LIND 1963; FUCHS 1977; VEEPU’ 1977; ERWIN 1979; GOT MAR^^ & .%NDERSSON 1980; BURGER & GOCHFELD 1990). Because terns actively kept predators from entering the subcolonies, we expected skimmers in the single-species subcolonies to respond more strongly to predators than the skimmers in the mixed-species subcolonies. This hypothesis is not supported by our observations of no difference in skimmer response in the presence or absence of gull-billed terns.
Human activity inside the colonies can reduce skimmer responses to avian predators (BURGER & GOCHFELD 1992). To avoid this confoundmg factor, we made all our observations of the antipredator behavior of both species from blinds about 30 m away from the edge of the subcolonies. The only human present on the entire dredge island was the observer in the blind, except during the decoy trials. During these trials both humans were well hidden. Hence, the apparent low energetic expenditure of mobbing by shmmers did not appear to be a function of nesting with the terns nor the presence of humans, but rather the behavior of the black skimmers.
Gulls and minks are common nest predators PURGER & G o m I m r ) 1990) and thus it is reasonable to assume that skimmers perceive them as threats -yet, they did not mob predators in higher proportions in the absence of gull-billed terns. Thus, predation losses would be expected to increase in single-species groups relative to mixed-species groups. We cannot test this former hypothesis with our data because nest predation was almost nonexistent in both subcolony types.
When faced with predation threats, most skimmers stayed at the nest site instead of mobbing in the air. This is fairly uncommon behavior for a colonial seabird, since the majority of the species defend their group through active group defense (VEEN 1977). The few colonial-nesting species that do not mob predators actively are ones that historically have nested in predator free areas (CUJLEX 1957). Dredge islands are typically free from terrestrial predators (SOOTS & PARKELI. 1975; KARWoWSKl et al. 1995) and this circumstance could have influenced skimmers to respond in low proportions to threats.
Although black skimmers will sometimes actively defend colony sites (GROOM 1992), they often appear to recognize other colonial species that are aggressive towards predators and nest with them. Empirical evidence for this lies along the Atlantic coast of North America, where black skimmers nest predominantly with common terns (ERWIN 1977; BURGER & GOCHFELD 1990), and in coastal Louisiana where they nest predominantly with gull-billed terns (MARTIN & LESTER 1990). If black skimmers do indeed gain an antipredator benefit by choosing to nest with a ‘protector’ species, this benefit may decrease as subcolony size increases. At our study sites, all the single-species subcolonies were formed after mixed-species subcolonies were established. This suggests that late-arriving shmmers may choose to nest by themselves to avoid the costs of nesting at high densities

282 S 'M PII \ & P L LEBFRG
(HUNT et al. 1986) that were common in the mixed-species subcolonies of our study sites. The benefit of nesting in large mixed-species subcolonies is also reduced for the late- nesting birds because gull-billed terns are near the completion of their nesting season and have greatly reduced their own responses to predators.
During our study, skimmers nesting in mixed-species subcolonies tended to have slightly more nests that produced chicks and more chicks per nest than skimmers nesting in monospecific groups ( P I U S - B A L A ~ S H N A N 1996). Because we observed extremely low rates of nest predation we doubt that these small differences in reproductive success are due to variation in the effectiveness of antipredator defenses of the two groups. However, we also feel that the lack of predation on seabird nests in our study does not preclude protection from predators as being one of the reasons that skimmers nest with terns. The creation of dredge islands, which often have relatively low levels of predation compared with more traditional nest sites (SOOTS 81 PAWELL 1975; KARWOWSKI et al. 1995), is a very recent event relative to the evolution of mixed-species coloniality.
No one hypothesis is likely to explain mixed-species coloniality because it is a multifaceted phenomenon. Although an antipredator advantage appears to result from skimmers nesting with terns, it may not be the sole or even the most likely explanation for the association between two species. For example, black skimmers may be using the early arriving gull-billed terns as indicators of good breeding habitat. Slammers require the same general types of breeding area, comprising unvegetated sand-shell substrate common to coastal beaches, barrier islands and dredge islands, as gull-billed terns (LEBFCRG et al. 1997). Other factors such as information transfer of the location of food resources, as well as reduced aggression (PIUS & LEBERG 1997) and cannibalism may also contribute not only to black skimmer preference for nesting with gull-billed terns, but may also help explain other mixed-species associations.
Acknowledgements
We thank M. CARLOSS and the staff of the Atchafalaya Delta Wildlife Management Area for their continuing assistance and support of research in the Atchafalaya Delta, and T. MALLACH for extensive help in the field. Helpful criticisms were provided by J. HATLE, R. JAEGEX, C. Jrism, J. NF.IGEL, R. 'IVlr.1 EY. This work was supported by a University of Southwestern Louisiana University Doctoral Fellowship to S. PIUS and by funds from the Louisiana Department of Wildlife and Fisheries, and National Science Foundation grant DEB-9123943 to P. LEBRRG.
Literature Cited
AI.HERI(:O, J . A. R., RED, J . M. & ORING, L. W. 1991: Nesting near a common tern colony increases and
AI.EXANDFX, R. D. 1974 The evolution of social behavior. Annu. Rev. Ecol. Slst. 5, 325-383 B I R K H ~ A D , T. R. & NETTLFSIIIP, D. N. 1995: Arctic fox influence on a seabird community in Labrador: a
BOURGET, A. A. 1973: Relation of eiders and gulls nesting in mixed colonies in Penohscot bay, Maine. Auk 90,
BREAREY, D. & HILDEN, 0. 1985: Nesting and egg-predation by turnstones Awnuriu zn tqmi in larid colonies.
BL'RGER, J. 1981: A model for the evolution of mixed species colonies in Ciconiiformes. Qu. Rev. Biol. 56,
decreases spotted sandpiper nest predation. Auk 108, 904-910
natural experiment. Wilson Bull. 107, 3 9 7 4 1 2
809-820
Ornis Scand. 16,283-292
143-167

Response of Seabirds to Predators 283
__ 1984 Grebes nesting in gull colonies: Protective associations and early warning. Am. Nat. 123, 327-337 _- & GOCHFRT.D, M. 1990 The Black Skimmer: Social Dynamics o f a Colonial Species. (:olumbia Unir.
__ & -- 1991: The Common Tern: Its Breeding Biology and Social Behavior. Columbia Univ. Press, New
-_ & -- 1992: Experimental evidence for aggressive antipredator behavior in black skiminers (K&rbo).r
BURNESS, G. P. & MORRIS, R. D. 1993: Direct and indirect consequences of mink presence in common tern
CAIRNS, D. K. 1992 Population regulation of seabird colonies. In: Current Ornithology, 9 (P~wF.R, D. M., ed.).
CARRASCAI., L. M. & MORENO, E. 1992: Proximal costs and benefits of heterospecific social foraging in thc
CROSSIN, R. S. & HUBER, H. N. 1970: Sooty tern egg predation by ruddy turnstones. Condor 72, 372-373 CULLEN, E. 1957: Adaptations in the kittiwake to cliff nesting. Ibis 99, 275-302 CULI-EN, J. M. 1958: Some adaptations in the nesting hehaviour o f terns. Proc. Int. Om. Congr. 12, 153-157 EIGAR, M. A. 1989: Predator vigilance and group size in mammals and birds: a critical rewew o f the empirical
Ef.1 fOT, R. D. 1985: The exclusion of avian predators from agqregations of nesting lapwings (I ‘Onelhr z,une//us).
EMEN, J . T., MII.I.ER, D. E., EVANS, R. M. & THOMPSON, D. H. 1966: Predator-induced parental neglect in
EMSLIE, S. D., ICZRNOVSKY, N. & TRI I’IECF:, VV. 1995: Avian predation at penp in colonies on King
ERWIN, R. M. 1977: Black skimmer breeding ecology and behavior. Auk 94, 709-717 _- 1979: Species interactions in a mixed colony of common terns (.Sterna birundo) and black skimmers (Rhynrbops
F u c ~ s , E. 1977: Predation and antipredator behaviour in a mixed colony of terns Stemu sp. and black-headed
GIROGX, J . F. 1981: Ducks nesting in association with Canada geese. J. Wildl. Manage. 45, 778--782 GOTMARK, F. & AHLUND, M. 1986: Nest predation and nest site selection among eiders .\omutetiu moilistma: the
-_ & ANDERSON, M. 1980: Breeding association between common gull Lorus cunws and arctic skua .Stercorutiu.r
_- , WINKLER, D. W. & ANDERSSON, M. 1986: Flock-feeding on fish schools increases indi\itlual success in
GROOM, M. 1992: Sand-colored nighthawks parasitize the antipredator behavior of three nesting bird species.
GROVER, J. J. & OLLA, B. L. 1993: The role of the rhinoceros auklet (C.krorbinru m~nocemtu) in mixed-species
HAMILTON, W. D. 1971: Geometry for the selfish herd. J. Theor. Biol. 31, 295-311 HARRISON, N. M., WHITEHOUSE, M. J., HEINEMANN, D., PRINCE, P. A,, HUNT, G. L. & VHIT, R. R. 1991:
HOHN, E,. D. 1967: Observations on the breeding biology of Wilsons’s phalarope (Stepnopur tricolor) in Central
HOOGLAND, J. L. 1979: The effect of colony size on individual alertness of prairie dogs (Scruridae: CGnoTys
HUISMAN, K. 1984 Selection of prey and success of silver gulls robbing crested terns. Condor 86, 130---138 HUNT, G. L. Jr., EPPIEY, 2. A. & SCHNEIDER, D. C. 1986: Reproductive performance of seabirds: the
HIXLBERT, S. H. 1984: Pseudoreplication and the design o f ecological field experiments. Ecol. Monogr. 54,
I(ARWO\X’SKJ, K., J . E. GATES & I,. H. H A R P E R . 1995: Common terns nesting on navigational aids and natural
KRIJUK, H. 1964: Predators and antipredator behavior of the black-headed gull (Lams ridihundus L). Behaviour,
Press, New York
York
nger). i‘qgr. Behav. 18,241-248
colony. Condor 95,708-71 1
Plenum, New York. pp. 37-61
great tit, Parusmajor. Can. J . 2001. 70, 1947-52
evidence. Biol. Rev. 64, 13-33
Anim. Behav. 33, 308-314
a ring-billed gull colony. Auk 83, 677-679
George Island, Antarctica. Wilson Bull. 107, 31 7-327
nker). h i m . Behav. 27, 1 0 5 k 1 0 6 2
gulls laws ridibwndus with special reference to the sandwich tern .Sterna runduirenrir. C h i s Scand. 8, 17-32
influence of gulls. Ibis 130,111-123
purusitzcws. Ornis Scand. 11, 121-124
gulls. Nature 319, 58‘)-591
Ecology 73,785-793
feeding assemblages of seabirds in the Strait of Juan de Fuca, Washington. Auk 100, 979-982
Observations of multispecies seabird flocks around south Georgia. Auk 108, 801-810
Alberta. Auk 84, 220-244
spp.). Anim. Behav. 27, 37-07
importance of population and colony size. Auk 103, 306-317
187-21 1
islands in the St Lawrence River, New York. Wilson Bull. 107, 4 2 3 4 3 6 .
Suppl. 11.

284 S. M. Pits & P. L. LRBERG
KL'MMER, H. 1968: Two variauons in the social organPzation of baboons. In: Primates: Studies in Adaptation
LACK, D. 1968: Ecological Adaptations for Breeding in Birds. Methuen, London LEBERG, P. L., DESHOTELS, P., PIUS, S. M. & CARI.C)SS, M. 1997: Nest sites of seabirds on spoil islands in
coastal Louisiana. Proc. Southeast. Assoc. Fish Wildl. Agencies 49, 356-366. LIND, H. 1963: Notes on social behaviour in terns. Dansk Ornithologisk 57, 155-175 LOFTIK', R. W. 1979: Ruddy turnstones destroy royal tern colony. Wilson Bull. 91, 133-135 MARTIN, R. P. & LESTER, G. D. 1990: Atlas and Census of Wading Bird and Seabird h-esting Colonies in
Louisiana 1990. Louisiana Dept of Wildl. and Fish., Louisiana Nat. Heritage Progr., Hourque, Baton Rouge MCKAYE, K. R. & OLIVER, M. K. 1980: Geometry of a selfish school: defense of cichlid young 11) bagrid catfish
in Lake Malawi, Africa. Anim. Behav. 28, 1287 Mi+,:ctl, L. D. 1970: The Wolf: the Ecology and Behavior of an Endangered Species. Nat. Hist. Press, New York NETTJ.ESFIIP, D. N. 1973: Breeding ecology of turnstones Arenaria in tepw at Hazen Camp, F.llesmere Island,
NF.WTON, I. & CAMFF.I.I., C. R. G. 1975: Breeding of ducks at Loch Lcvcn, Kinross. Wildfrml 26, 83-103 NISBET, I. C. T. 1975: Selective effects of predation in a tern colony. Condor 77, 221-226 NUECHTERLE~K, G. L. 1981: 'Information parasitism' in mined colonies of western grebes and Furester's terns.
Anim. Behav. 29, 985-989 PARKES, K. C., P<>0I2E, A. & LAPHAM, H. 1971: The ruddy turnstone as an egg predator. Wilson Bull. 83,
306-308 PITCIIER, T. J., WYCEIE, C. J . & MAGUKKAN, A. E. 1982: Evidence for position preferenter in schooling
mackerel. h i m . Behav. 30, 932-934 PI~ 'S-BAI.AKRIS~INAN, S. M. 1996: The maintenance and consequences of mixed species colonies: thc influence
of gull-billed terns (.%mu niLutim) on black skimmers (R&nchops nzgeq'. Phi3 thesis, Univ. of Southwestern Louisiana, Lafayette
_- & LEDERG P. L. 1997: Aggression and nest spacing in single and mixed species groups of seabirds. Oecologica 111, 1 4 6 1 5 0 .
RCIHFJTSCIN, G. J. 1995: Factors affecting nest site selection and nesting success in the common eider .\omatenu mollimma. Ibis 137, 109-1 15
SCtIAI.I.F,R, G. 1972: The Serengeti Lion: h Study of Predator-Prey Relations. Univ. of Chicago Press, Chicago SCHEEL, D. 1993: Watching for lions in the grass: the usefulness of scanning and its effects during hunts. Anim.
stlEAl.f<R, D. A. & BURGER, J . 1992: Differential responses of tropical roseate terns to aerial intruders throughout
SINCJAIR, A. R. E. 1985: Does interspecific competition or predation shape the African ungulate community? J.
SMITt I , A. J . M. 1975: Studies of breeding sandwich terns. Br Birds 68, 142-156 SOOTS, R. F. & PARSELL, J. F. 1975: Ecologcal succession of breeding birds in relation to plant succession o n
TINBERGEN, N. 1952: O n the significance of territoriality in the herring gull. Ibis 94, 158-159 -- , I~VIPF.KOVEN, M. & FRANCK, D. 1967: An experiment on spacing-out as a defense agtainst predation
VEEN, J. 1977 Functional and causal aspects o f nest distribution in colonies of the sandwich tern (.Vema i
W I T T ~ N R ~ R G P B , J. F. & HLINT, G. I>. Jr. 1985: The adapuve significance of coloniality in birds. In: Avian
YOI'NG, A. D. & TITMAN, R. D. 1986: Costs and benefits to red-breasted mergansers nesting in tern and gull
and Variability (JAY, P., ed.). Holt, Rinehart & Winston, New York. pp. 293-312
N.W. T. Ibis 115,202-217
Behav. 46,695-704
the nesting cycle. Condor 94, 712-719
Anim. Ecol. 54, 899-918
dredge islands in North Carolina. Natl. Oceanic and Atmos. Sea Grant Publ. UN(:-S(;-75--27
Behaviour 28,307-321
fandvirenni Lath.). Behaviour, Suppl. 20.
B i o l q q , 8 (PARKF.R, D. S., KING, J. R. & PARKES, K. C., eds). Academic Press, New York. pp. 1-79
colonies. Can. J . 2001. 64, 2339-2343.
Received: Auxust 27, 1996
Accepted: June 30, 1996 (J. Brockman)




















