
Chironomus esai n. sp. (Diptera: Chironomidae) in lakes and ...
Upload
independentCategory
view
1download
0
Freshwater Biology (1995) 34,1-12
The non-lethal effects of predators and the influenceof food availability on life history of adultChironomus tentans (Diptera: Chironomidae)
S. L. BALL* AND R. L. BAKERDepartment of Zoology, Erindale College, University of Toronto, Mississauga, Ontario L5L JC6, Canada
*Present address: Division of Biological Sciences, University of Missouri - Columbia, Columbia, MO 65211, U.S.A.
SUMMARY
1. We used a laboratory experiment to determine effects of a predator (other thanstraightforward prey consumption) and food availabiUty, during the larval stage, onadult size, age at emergence and fecundity of Chironomus tentans.
2. Predator presence and decreased food availability resulted in reduced adultemergence.3. Predator presence and food availabihty resulted in smaller size and greater age atemergence of male and female chironomids. There were no significant interactionsbetween effects of predator presence and food availability.4. Predator presence had no significant effect on fecundity.5. Smaller size and greater age at emergence can have important iniplications for adultsurvival, size and age at first reproduction and, thus, intrinsic rate of population growth.
Introduction
Organisms often respond to environmental stress (i.e.some environmental condition that impairs Darwinianfitness; Sibly & Calow, 1989) through changes in theirlife history. For aquatic invertebrates changes in bodysize, development time and fecundity may be inter-preted as phenotypically plastic responses to temper-ature (Giberson & Rosenberg, 1992; Rempel & Garter,1987), food availability (Anderson & Gummins, 1979;Gollins, 1980; Colbo & Porter, 1979) and food quality(Rosillon, 1988; Vanni & Lampert, 1992). Life-historychanges may also occur as a result of predation.Predators may shape prey life history directly, throughmortality, or indirectly through various non-lethaleffects. Responses to local fluctuations in predationpressure may involve changes in prey morphology(Havel, 1987; Harvell, 1990) and/or behaviour (Dill,1987; Sih, 1987; Lima & Dill, 1990). Predators havebeen shown to induce growth of enlarged spines,helmets and crests in several zooplankton species(Grant & Bayly, 1981; Herbert & Grewe, 1985; Stenson,1987; Parejko, 1991), as well as changes in shell mor-
© 1995 Blackwell Science Ltd
phology in barnacles (Lively, 1986 a,b), and the growthof abdonoinal hairs in chironomids (Hershey &Dodson, 1987), all of which function to inhibit inges-tion of prey by predators. In addition, it has been welldocumented that many species of aquatic animalsalter their activity (Semlitsch, 1987a; Blois-Heulin et al,1990; Wemer, 1991; Macchiusi & Baker, 1992; Stauffer& Semlitsch, 1993), movement patterns (Ramcharan &Sprules, 1991), or habitat use (Wemer et al, 1983;Harvey, 1991; Fraser & Gilham, 1992; Persson, 1993)in response to predator presence. Numerous studieshave shown that, under the risk of predation, preyreduce their foraging activity (damselflies, Dixon &Baker, 1988; waterstriders, Sih, Krupa & Travers, 1990;notonectids, Sih, 1982; stoneflies, Feltmate & Williams1989; isopods. Short & Holomuzki, 1992; anuranlarvae, Lawler, 1989; fish, Prejs, 1987). Althoughdecreased activity may reduce the immediate threatof predation, there may also be costs of reducedforaging. Organisms that reduce their activity inresponse to predators, therefore, may grow at a slower

2 S. L Ball and R. L. Baker
rate and suffer reduced body size, prolonged develop-ment and decreased fecundity.
While several studies have documented predator-induced reductions in body size and developmenttime for the larvae of aquatic insects (Heads, 1986;Dixon & Baker, 1988; Feltmate & Williams, 1991;Peckarsky et al, 1993) and anurans (Skelly & Werner,1990) few have focused on the effects of predatorpresence, during the larval stage, on the adult stage (forexceptions see Semlitsch, 1987b; Feltmate & Williams,1991; Peckarsky et al, 1993). For animals with complexHfe cycles, growth during the larval stage can havesignificant effects on adult body size, age at maturity(Wilbur & Collins, 1973; Wilbur, 1980; Semhtsch, Scott& Pechman, 1988) and fecundity (Rowe & Ludwig,1991; Fraser & Gilliam, 1992). This is particularlyimportant for insects with non-feeding adults, wherelarvae must channel much of their energy into produ-cing reproductive tissues. Thus, predator-mediatedeffects on larval growth and development could trans-late into smaller adult body size and changes in ageat maturity. This, in tum, can affect survival, fecundityand mating success, all of which can influence indi-vidual fitness and ultimately, population growth.
The objective of our study was to examine the effectsof non-Ietha! presence of predatory pumpkinseed sun-fish {Lepomis gibbosis L.) and food availability, in thelarval environment, on the life-history traits of adultChironomus tentans Fab. The larvae are benthic, residingin loosely constructed tubes composed of particlesfrom the substratum held together with a silk-likesalivary secretion (Oliver, 1971). Tubes, when coupledwith reduced larval activity, provide refuge for larvaeby reducing encounters with predators (Bohanan,1988; Macchiusi & Baker, 1991). A chironomid larvafeeds by extending out of its tube while its abdomenis anchored within the tube; when the larva detects apredator it rapidly pulls back into its tube. In choiceexperiments, Hershey (1987) found predators select-ively preyed upon chironomid spedes which spentmore time outside their tubes. Recent laboratory stud-ies have also shown that C. tentans larvae, in thepresence of pumpkinseeds, reduced the amount oftime spent outside their tubes (Macchiusi & Baker,1992). By reducing foraging activity in the presence offish, larvae may suffer reduced growth rates whichconsequently result in changes in adult life-historytraits, such as smaller body size and longer develop-ment time.
Materials and methods
Study animals
A culture of Chironomus tentans was established inspring 1992 from egg masses supplied by DonnaBedard, Ministry of the Environment, Ontario, Canada.Larvae were reared in 3.0-1 opaque polypropylenetanks (26 X 15 x 10-cm deep) containing a 3-cm layerof white silica sand (particle diameter 250-500 |xm)which provided a substratum for larvae to build tubes.Tanks were filled with dechlorinated water and weremaintained at 22 °C under a 16L : 8D photoperiod.Larvae were fed ad libitum with a suspension of finelyground 'Tetra Conditioning Food, Vegetable Diet forTropical Fish' (Melle, Germany) in distilled water,every second day.
Pumpkinseed sunfish {Lepomis gibbosus) were keptin 68-1 glass aquaria in the laboratory at 22 "C, undera 16L : 8D photoperiod, and were fed ad libitum with'Tetra Bits Bite Size Food Bits for Discus, mid waterand bottom feeding tropical and marine fish' (Melle,Germany), Daphnia, C. tentans larvae, chopped frozenfish, and odonate larvae. Fish were collected from apond on the Erindale College campus. University ofToronto (43°33' N, 79°40' W) and from Lake Opiniconat the Queen's University Biological Station (44^34' N,79°15' W), Ontario, Canada.
Ejfects of predator presence and food level on size andage at emergence
We used a laboratory experiment with two levels ofpredator presence ('non-lethal' presence or absence ofa pumpkinseed) crossed with three levels of foodavailability for chironomids (low, medium and high)to determine the effects of predators and food availabil-ity on size and age at emergence of adult chironomids.We included food level in our experiment because theresponse of larval chironomids to fish predators maydiffer with food availability. At low food level larvaemay already be near starvation, and predators cannotfurther reduce larval food intake. Similarly, at highfood level predators may not substantially affect devel-opment.
Sixty (ten blocks of the six treatment combinations)3.0-1 opaque polypropylene tanks (26 X 15 X 10 cmdeep) were each filled with 3-cm-deep layer of finesilica sand (grain size = 200-500 \xm). Each tank was
© 1995 Blackwell Science Lid, Freshwater Biohgy, 34, 1-12

filled with 2.6 1 of dechlorinated water and equippedwith a 2-mm-diameter plastic tubing taped to theinside wall of the tank and connected to common airtubing to provide aeration. Each tank was fitted witha removable screen cover to prevent adults fromescaping. Newly emerged adults flew up to theunderside of the screen where we could easily col-lect them.
The experiment was initiated over 12 days with oneblock set up each day, except for blocks 2 and 9 whichwere set up 2 days after the preceding block. All tankswere kept at the same height on metal shelves in anenvironmental chamber. The location of each blockwithin the chamber was assigned randomly. The cham-ber was kept at 22 °C under a 16L : 8D photoperiod.
Fifty final-instar larvae were placed in each tankand treatments were assigned randomly. We usedrecently moulted final instar larvae which were distin-guishable by their bright orange-pink appearanceand a dark dorsal stripe on the head capsule. Thereare four larval instars, with the final instar comprisingapproximately half of the larval duration. Formationof reproductive structures occurs during the finalinstar (Butler, 1984). Thus, larvae were exposed topredators during the longest and potentially mostimportant stage of their development.
We placed larvae in treatment tanks 18 h beforeadding fish to allow larvae sufficient time to buildtubes. Larvae were fed a suspension of 5 g of finelyground 'Tetra Conditioning Food, Vegetable Diet forTropical Fish' in 25 ml of distilled water, every secondday. Low, medium and high food levels consisted ofone, two and four drops, respectively, of the foodsuspension. In an effort to distribute food evenlywithin each tank, we gently swirled the water with aglass pipette while adding drops of food.
Predator treatments consisted of the presence orabsence of a small (5.0-8.0 cm standard length) pump-kinseed sunfish. We prevented the fish from eatinglarvae by sewing their mouths shut with white cottonthread. This was done in accordance with the Univer-sity of Toronto Animal Care Committee (protocolnumber 1504) and the Canadian Council on AnimalCare. We used fish on a rotational basis so that eachfish was deprived of food for only 2 consecutive daysand, while not being used in the experiment, spent atleast 2 consecutive days in an aquarium where it waswell fed before being used again. Fish were removedfrom treatment tanks using a small net; they were
© 1995 Blackwell Science Ltd, Freshwater Biology, 34, 1-12
Life history of adult Chironomus tentans 3
caught easily, with little disturbance to chironomidlarvae. To control for any possible disturbance causedby addition or removal of fish in predator treatmenttanks, we gently moved a net through the water ofno predator treatment tanks at the time of fish changes.Fish faeces and dead chironomid larvae and pupaewere removed twice daily, because larvae would eatanything left in the tank. This allowed us to controlfood level and to document larval and pupal mortality.We removed 1.0 1 of water from each tank every 12-15 days and replaced it with fresh dechlorinated waterto prevent ammonia from accumulating. Changes wereperformed on days when food was not added so thatwe would not be removing food inadvertently. Weadded distilled water to all tanks weekly to replacethat lost through evaporation. This was done byslowly pouring water on to a 12.0 X 14.5-cm Plexiglasrectangle equipped with a wooden handle held at thewater surface. This enabled us to add water to tankswith minimal disturbance to the substratum and with-out destropng larval tubes.
Each day we noted the presence of dead larvae orpupae and coUected adults that had emerged. Foreach adult, we recorded; treatment, emergence dateand sex. Adults were individually frozen in smallplastic vials until we could measure them. Size meas-urements were taken on specimens that were thawed at22 °C for at least 20 min. We then measured abdomenlength (from the junction of the thorax and firstabdominal segment to the tip of the abdomen), winglength (from the axillary sclerites to the tip of the rightVkdng), and head width (at the widest point). Specimenswere dried at 55 °C for at 3 days and weighed toobtain dry mass. We used these three different mor-phometrics, as well as dry mass, to estimate adultbody size because we did not have prior knowledgeof how predators would affect the size of chironomids.In a study of the effects of food limitation on conditionand size of damselflies. Baker (1989) found that therewas no difference in head width of Ischnura verticalis(Say) larvae on various diets, although body 'condi-tion' (relaHve mass/unit head width) did vary withdiet. Thus, we measured severed aspects of body sizeso that we could detect any effects of predators onsize of adult chironomids.
Because larval mortality differed between treat-ments we used two sets of criteria to determine whento end the treatment in each experimental tank. Highand medium food treatments were ended if no larvae

4 S. L. Ball and R. I. Baker
or pupae were visible, if at least thirty of the fiftylarvae could be accounted for (either as adults or asdead larvae or pupae), and if no adults had emergedfor at least 14 days. Larval mortality at low food levelwas extremely high and, in many cases, fewer thanthirty larvae could be accounted for through emer-gence or mortality. In these cases, treatments wereended if no larvae or pupae were visible and if noadults had emerged for at least 28 days. We carefullyexamined each tank when we ended the treatmentand noted whether any living or dead larvae or pupaewere found. Of the 3000 larvae used in the experiment,1318 adults emerged and only two live larvae wererecovered from the sixty tanks at the end of theexperiment. Therefore it is unlikely that our resultswould be affected by time at which a treatmentwas ended.
Effects of predator presence and food level on fecundity
To determine treatment effects on fecundity, newlyemerged females from the experiment described abovewere mated with males from the stock culture. Pairswere placed in 70-ml plastic snap-cap vials containing10 ml of dechlorinated water. Females usually ovi-posited a single large C-shaped egg mass within 24 h,although pairs were left undisturbed for an additional24 h because females sometimes laid an additionalsmall fragment of an egg mass. We noted whetherfemales had oviposited and removed them to indi-vidual vials where they were frozen for later morpho-metric measurement. We measured fecundity as thenumber of larvae hatched from egg masses and surviv-ing to day 5. We chose number hatched, and notsimply the number of eggs per egg mass becausechironomid egg masses consist of approximately 2000tiny eggs spirally arranged in a clear gelatinous matrix.The small size of eggs and their spiral arrangementmade counting extremely difficult. Egg masses weremaintained at 22 "C for 3 days. We then added onedrop of food suspension to each vial for newly hatchedlarvae. Two days later we transferred the contents ofeach vial to a 20-HI1 glass screw-cap vial and added10 ml of 10% formaldehyde to each vial to preserveits contents. We counted the number of larvae fromeach vial under a dissecting microscope at 25 X magni-fication.
Due to a shortage of adult males in the stock culture,not all females were given the opportunity to lay eggs.
Oc
CD
QJCD
100-,
80-
60-
• No Predator
• Predator
Low Medium
Food Level
High
Fig. 1 Effects of predator presence and food availability on themean percentage of adult chironomids emerging. Valuespresented here are untransformed means and standarddeviations. Different letters indicate significant differences(P < 0.05).
Of those given the opportunity to lay eggs, not allfemales did so. We treated each of these three groupsof females separately (not mated, mated but no eggslaid, mated and eggs laid) and performed statisticalanalyses only on those females that laid eggs (FLE)and those that did not lay eggs when given theopportunity (FNLE). Analysis was not performed onthe few females that were not paired with males.
Results
Effects of predators and food on adult emergence
Mean percentage emergence of adults from each ofthe treatments was compared using a three-way ran-domized block ANOVA. The blocking factor was theday on which treatments were initiated. Angulartransformed mean percentages (Zar, 1984) were norm-ally distributed and homoscedastic. There was a signi-ficant effect of predator presence (fi,45 = 11.13, P =0.002) and food availabUity (F2,45 = 86.00, P < 0.0001)on emergence (Fig. 1). There was a significant effectof the blocking factor (F9.45 = 2.56, P = 0.02) but nosigruficant interaction between predator presence andfood leve! (f2,45 = 1.75, P = 0.19). Tukey's HSD mul-tiple comparison showed emergence was highestunder conditions of no predator and high food avail-ability (HNF), but was similar for the 'highfood-predator' (HP), 'medium food-no predator'(MNP), and 'medium food-predator' (MP) treat-
© 1995 Blackwell Science Ltd, Freshwater Biology. 34, 1-12
Life history of adult Chironomus tentans 5
4.60
Low Medium Low Medium High
c)
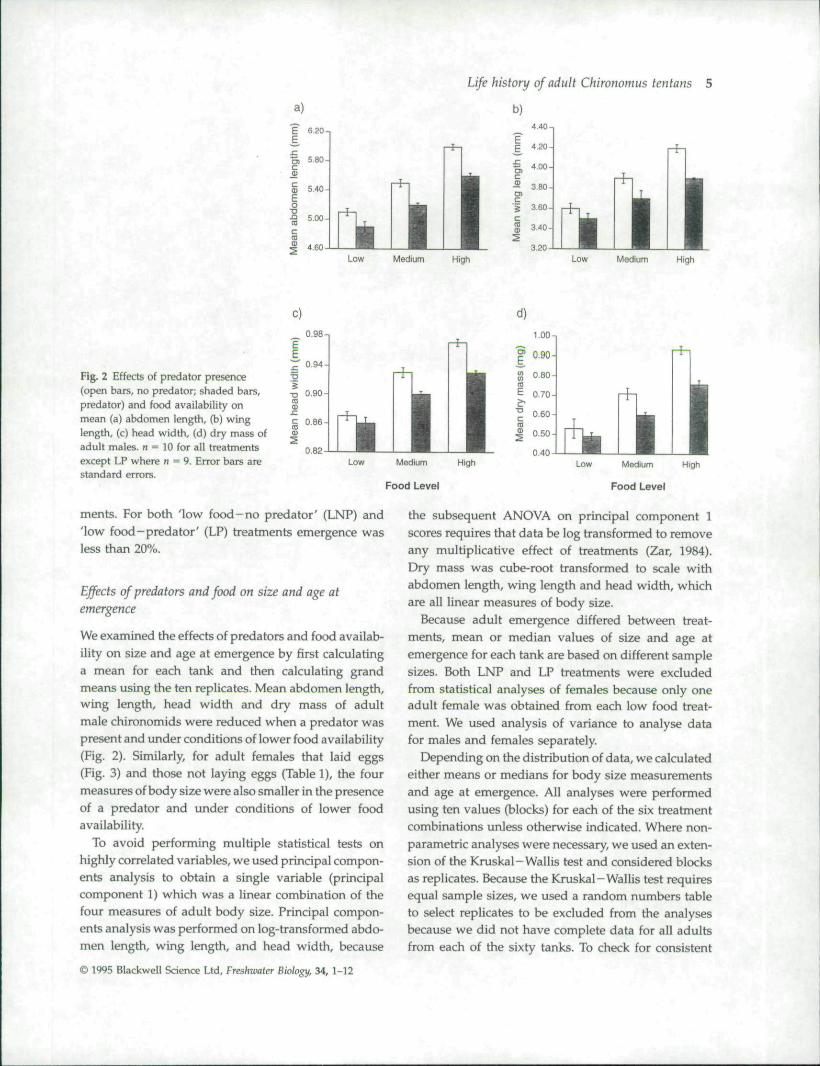
Fig. 2 Effects of predator presence(open bars, no predator; shaded bars,predator) and food availability onmean (a) abdomen length, (b) winglength, (c) head width, (d) dry mass ofadult males, n = 10 for all treatmentsexcept LP where n = 9. Error bars areStandard eirors.
0.98 n
0,94-
1,00-,
0.90-
0.86-
0.62Low Medium High
Food Level
Low Medium High
Food Level
merits. For both 'low food-no predator' (LNP) and'low food—predator' (LP) treatments emergence wasless than 20%.
of predators and food on size and age atemergence
We examined the effects of predators and food availab-ility on size and age at emergence by first calculatinga mean for each tank and then calculating grandmeans using the ten replicates. Mean abdomen length,wing length, head width and dry mass of adultmale chironomids were reduced when a predator waspresent and Linder conditions of lower food availability(Fig. 2). Similarly, for adult females that laid eggs(Fig. 3) and those not laying eggs (Table 1), the fourmeasures of body size were also smaller in the presenceof a predator and under conditions of lower foodavailability.
To avoid performing multiple statistical tests onhighly correlated variables, we used principal compon-ents analysis to obtain a single variable (principalcomponent 1) which was a linear combination of thefour measures of adult body size. Principal compon-ents analysis was performed on log-transformed abdo-men length, wing length, and head width, because
© 1995 Biackwell Science Ltd, Freshvmter Biohgy, 34, 1-12
the subsequent ANOVA on principal component 1scores requires that data be log transformed to removeany multiplicative effect of treatments (Zar, 1984).Dry mass was cube-root transformed to scale withabdomen length, wing length and head width, whichare all linear measures of body size.
Because adult emergence differed between treat-ments, mean or median values of size and age atemergence for each tank are based on different samplesizes. Both LNP and LP treatments were excludedfrom statistical analyses of females because only oneadult female was obtained from each low food treat-ment. We used analysis of variance to analyse datafor males and females separately.
Depending on the distribution of data, we calculatedeither means or medians for body size measurementsand age at emergence. All analyses were performedusing ten values (blocks) for each of the six treatmentcombinations unless otherwise indicated. Where non-parametric analyses were necessary, we used an exten-sion of the Kruskal-Wallis test and considered blocksas replicates. Because the Kruskal-Wallis test requiresequal sample sizes, we used a random numbers tableto select replicates to be excluded from the analysesbecause we did not have complete data for all adultsfrom each of the sixty tanks. To check for consistent

6 S. L Ball and R. L. Baker
4,00Low Medium High Low Medium High
0.82 0.20-
L.OW Medium High
Food Level
Low Medium High
Food Level
Fig. 3 Effects of predator presence(open bars, no predator; shaded bars,predator) and food availability onmean (a) abdomen length, (b) winglength, (c) head width, (d) dry mass ofadult females that laid eggs, n = 10 forall treatments except MNP, M = 9; LNP,n = 1; LP, « = 1. Ermr bars arestandard errors.
Table 1 Effects of predator presence and food level on body size of adult females that did not lay eggs when given theopportunity. Values are means ± standard errors. H, high food level; M, medium food level; NP, no predators; P, predators
Variable Treatment
HNP {n = 9) HP {n = 8) MNP (fi = 9) MP (« = 9)
Abdomen length (mm)Wing length (mm)Head width (mm)Dry mass (mg)
5.60 ± 0.164.83 ± 0.080.96 ± 0.0091.23 ± 0.09*
4.85 ± 0.144.41 ± 0.090.91 ± 0.0130,97 ± 0.09
4,79 ± 0.134.49 ± 0.110.91 ± 0.0141.03 ± 0.08
4.50 ± 0.104.30 ± 0.080.88 ± 0.0070.80 ± 0.05
results, we performed the test three times, each timeexcluding randomly selected replicates. If the sameresults were obtained for each of the three tests thenwe report statistics and graph results from only thefirst test.
For adult males, a principal component 1 (PC 1)explained 94.5% of the total variance while PC 2, 3and 4 explained only 3.8, 0.9 and 0.7% of the totalvariance, respectively. All four measures of body sizeloaded heavily on PC 1 (Table 2) indicating that abdo-men length, wing length, head width and dry masswere equally good measures of adult male body size.We used an exter\sion of the Kniskal—Wallis testto determine effects of predator presence and food
Table 2 Component loading for prindpal component 1 scoresfor log transformed abdomen length, wing length, head width,and cube root transformed dry mass of adult males (M), adultfemales that laid eggs (FLE), and adult females that did notlay eggs when given the opportunity (FNLE)
Variable M FLE FNLE
Abdomen lengthWing lengthHead widthDry mass
0.97850.98840.97740.9439
0.93300.95580.96020.8930
0,92250.93060,46460.7792
availability on male body size at emergence becausePC 1 scores were heteroscedastic. Data for only ninereplicates were obtained from the LP treatment. For
1995 Blackwell Science Ltd, Freshwater Biology, 34, 1-12

Ufe history of adult Chironomus tentans 7
all three repetitions of the analysis, male size atemergence was smaller in the presence of a predator(H = 5.06, d.f. = 1, P < 0.03) and under conditions oflower food availability (H = 39.61, d.f. = 2, P < 0.001).There was no significant interaction (H = 0.43, d.f. =2, P > 0.50) between effects of predator presence andfood level (Fig. 4), although it appears that the effectof predators on male size at emergence was reducedat low food level.
Principal components analysis performed on thefour transformed measures of size at emergence forfemales that laid eggs, yielded a principal component1 which explained 87.6% of the total variance. Percent-age total variance explained by principal components2, 3 and 4 was small (PC 2, 6.7%; PC 3, 3.7%; PC 4,2.0%). All four measures of body size loaded heavilyon PC 1 (Table 2). Principal component 1 scores werenormally distributed and homoscedastjc so we useda three-way randomized block ANOVA for analysis.Females were significantly smaller in the presenceof a predator {F^ g = 24.42, P < 0.0001) and underconditions of lower food availability (F] 26 " 33.78, P< 0.0001); there was no significant effect of the blockingfactor (^9^6 = 0.30, P = 0.97) and no interaction(f 1,26 ~ 0.38, P = 0.54) betvk-een effects of predatorpresence and food level (Fig. 4).
For adult female chironomids that did not lay eggs,principal component 1 explained 63.5% of the totalvariance. Principal components 2, 3 and 4 accountedfor only 23.8, 9.5 and 3.2% of the total explainedvariance, respectively. All variables loaded heavily onthe first principal component except for head width(Table 2). Principal component 1 scores were normallydistributed and homoscedastic so we used a three-way randomized block ANOVA for analysis. Adultfemales not laying eggs were significantly smaller inthe presence of a predator (F, 22 = 21.17, P = 0.0001)and under conditions of less food (F j^ = 16.43, P =0.0005); there was no significant block effect ( 8,22 -0,63, P = 0.74) and no interaction (F,,22 = 1 41, P =0.25) between predator presence and food level (Fig. 4).
To determine whether predator presence or foodlevel influenced age at emergence, we recorded thenumber of days from the beginning of the experimentuntil emergence for each adult. We used an extensionof the Kruskal-Wallis test for analysis of median ageat emergence for males because data were heterosced-astic and could not be corrected through transforma-tion. Median age at emergence increased in the
a)
c)
2 i
£oo
CO
oCL OHcCO
^ -n
b)
O(JCO
0)
oo
CO
-2
MALES
n c
i b
1-
O 0-1
- 1 -
-2
Low Medium High
FLE
1 -
^ 0
-1 -
-2
Medium High
FNLE
I
Medium High
Food LevelFig. 4 Effects of predator presence (open circles, no predator;filled circles, predator) and food availability on body size atemergence of (a) males, (b) females that laid eggs (FLE),(c) females that did not lay eggs (FNLE). Values presentedhere are medians and interquartile ranges or back transformedmeans and standard errors. Different letters indicate significantdifferences (P < 0,05).
1995 Blackwell Science Ltd, Freshwater Biology, 34, 1-12
8 S. L Ball and R. L Baker
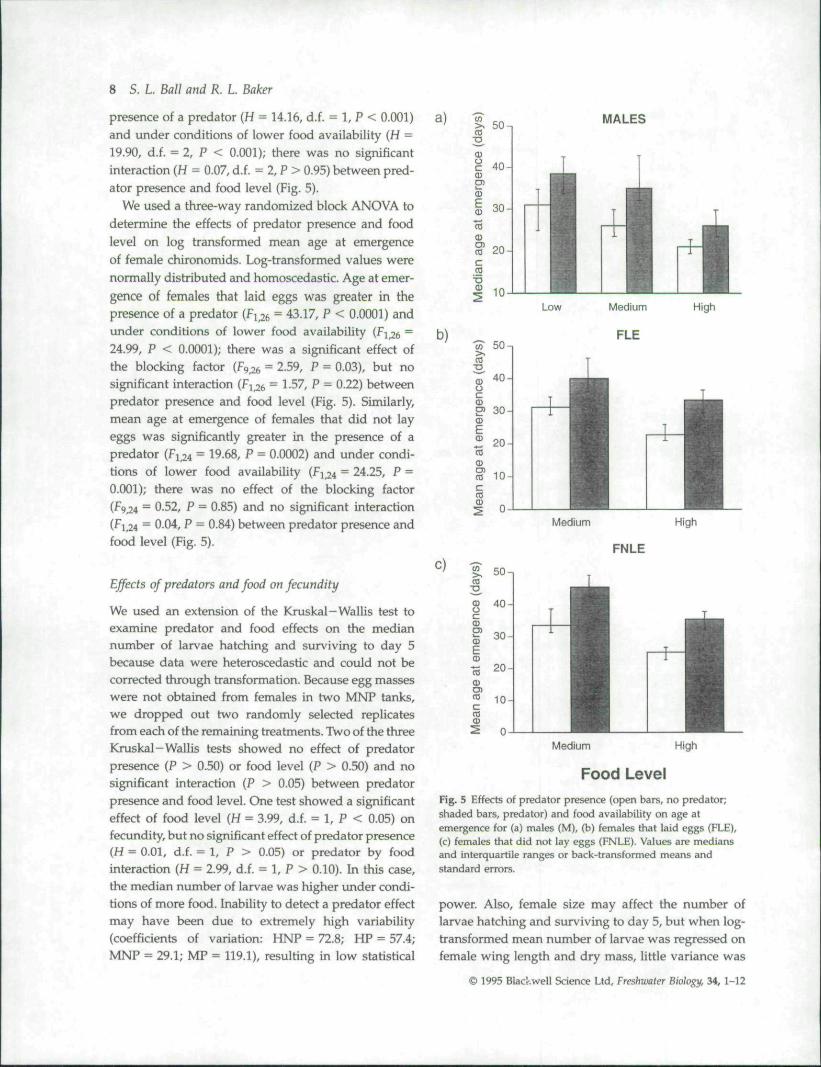
presence of a predator (H = 14.16, di. - 1, P < 0.001)and under conditions of lower food availability (H =19.90, di. = 2, P < 0.001); there was no significantinteraction (H = 0.07, di. = 2, P > 0.95) between pred-ator presence and food level (Fig. 5).
We used a three-way randomized block ANOVA todetermine the effects of predator presence and foodlevel on log transformed mean age at emergenceof female chironomids. Log-transformed values werenormally distributed and homoscedastic. Age at emer-gence of females that laid eggs was greater in thepresence of a predator (F^ ^ = 43.17, P < 0.0001) andunder conditions of lower food availability (F] 26 ~24.99, P < 0.0001); ther^ was a significant effect ofthe blocking factor (F^^e = 2.59, P = 0.03), but nosignificant interaction (Fi_26 = 1-57, P = 0.22) betweenpredator presence and food level (Fig. 5). Similarly,mean age at emergence of females that did not layeggs was significantly greater in the presence of apredator (Fi,24 = 19.68, P = 0.0002) and under condi-tions of lower food availability (Fi 4 = 24.25, P =0.001); there was no effect of the blocking factor(F9_24 = 0.52, P = 0.85) and no significant interaction(F; 24 ~ 004' P ~ 0S4) between predator presence andfood level (Fig. 5).
Effects of predators and food on fecundity
We used an extension of the Kruskal—Wallis test toexamine predator and food effects on the mediannumber of larvae hatching and surviving to day 5because data were heteroscedastic and could not becorrected through transformation. Because egg masseswere not obtained from females in two MNP tanks,we dropped out two randomly selected replicatesfrom each of the remaining treatments. Two of the threeKruskal-Wallis tests showed no effect of predatorpresence (P > 0.50) or food level (P > 0.50) and nosignificant interaction (P > 0.05) between predatorpresence and food level. One test showed a significanteffect of food level (H = 3.99, di. = 1, P < 0.05) onfecundity, but no significant effect of predator presence(H = 0.01, d.f. = 1, P > 0.05) or predator by foodinteraction (H = 2.99, d.f. = 1, P > 0.10). In this case,the median number of larvae was higher under condi-tions of more food. Inability to detect a predator effectmay have been due to extremely high variability(coefficients of variation: HNP = 72.8; HP = 57.4;MNP = 29.1; MP = 119.1), resulting in low statistical
MALES
b)50-,
40-
CT 3 0 -
::; 20 -
1 0 -
O
H
Low Medium
FLE
High
Medium High
FNLE
Medium - High
Food LevelFig. 5 Effects of predator presence (open bars, no predator;shaded bars, predator) and food availability on age atemergence for (a) males (M), (b) females that laid eggs (FLE),(c) females that did not lay eggs (FNLE). Values are mediansand interquartile ranges or back-transformed means andstandard errors.
power. Also, female size may affect the number oflarvae hatching and surviving to day 5, but when log-transformed mean number of larvae was regressed onfemale wing length and dry mass, little variance was
1995 BlacV.well Science Ltd, Freshwater Biology, 34, 1-12

explained by either wing length (r = 0.04, n = 38,P =0.23) or dry mass (r^ - 0.009, n = 38, P = 0.57).
Although most adtilt females were given the oppor-tunity to mate and lay eggs not all females did so. Todetermine if predator presence or food level affectedwhether a female laid eggs, we compared the propor-tion of females laying eggs from each treatment. Wecalculated the proportion for each tank and comparedmean proportions for each treatment using a three-way randomized block ANOVA. Proportions wereangular transformed (Zar, 1984); data were normallydistributed and homoscedastic. There was no effectof predator presence (Fi_27 = 0.32, P = 0.56) or foodavailability (Fi,27 = 2.43, P = 0.13) on the mean propor-tion of females laying eggs (proportion laying: HNP =0.63 ± 0.17; HP - 0.70 ± 0.22; MNP = 0.58 ± 0.24;MP = 0.52 ± 0.22, mean ± standard deviation; n = 10for all treatments). Neither the block effect (Fq_27 =0.97, P = 0.49) nor the predator by food interaction
= 0 41, P = 0.53) were significant.
Discussion
Both presence of a fish predator and decreased foodavailability significantly reduced emergence of adultC. tentans. At low food level emergence was less than20%, suggesting the majority of larvae were so foodlimited, regardless of fish presence, they could notcomplete development. There was no significant inter-action between effects of predator presence and foodlevel. These non-lethal effects of predators on adultemergence should be interpreted with some caution,however, because larval mortality may have resultedfrom direct fish predation. Although fishes mouthswere sutured we found four fish, very early in theexperiment, with sutures undone or loosened. How-ever, the effects of direct predation were probablynegligible; there was no evidence of fewer adultsemerging from those tanks with unsutured fish. Inaddition, we checked tanks every day and thus theamount of time unsutured fish were in tanks comparedwith the total time the experiment was run was small.
Both predator presence and decreased food availab-ility significantly reduced body size at emergence andresulted in greater age at emergence of male andfemale chironomids. We predicted predator effectswould be reduced at high and low food levels andgreatest at medium food level. Studies with zooplank-ton have shown that predators resulted in smaller
© 1995 Blackwell Sdence Ltd, Freshwater Biohgy, 34, 1-12
Life history of adult Chironomus tentans 9
body size (Dodson & Havel, 1988) as well as reducedsurvivorship, fecundity and increased helmet size(Hanazato, 1991), however, these effects were reducedor absent under low food conditions. We found nosignificant interaction between predator presence andfood level, although Fig. 4(a) suggests predator effectson adult male size were absent at low food level.
Under conditions of decreased food availability,there was a trend towards reduced fecundity, but noeffect of predator presence. Inability to detect a pred-ator effect was likely due to extremely high variabilityin the data which may have resulted, in part, fromlarval cannibalism. Our observations of newly hatchedlarvae suggest larvae feed on the gelatinous matrix ofthe egg mass. However, after 2-3 days the matrixcompletely disappears, leaving larvae without food.Our observations also suggest that larger larvae mayconsume moribund larvae. We tried to minimize anycannibalism by providing larvae with a small amountof food, although we do not know whether the foodaddition affected cannibalism.
To date, most studies of predator- and food-inducedchanges in prey life history have focused on larval orimmature stages of development. Few have examinedhow larval environments affect adult fitness. Environ-mental conditions which result in decreased larvalgrowth can have profound effects on adult fitness.Semlitsch (1987b) showed pond-drying regime, foodlevel and initial larval density reduced larval growthand had a marked impact on fitness of adult sala-manders, Ambystoma talpoideum (Holbrook), in termsof smaller body size and decreased fecundity. Feltmate& Willianns (1991) showed that stonefly nymphs,reared in sections of a stream containing rainbow trout{Oncorhynchus mykiss Walbaum), emerged at a smallersize and suffered reduced fecundity due to non-lethaleffects of trout presence. Similarly, Peckarsky etal.(1993) showed that mayflies suffered reduced growthrates, smaller adult body size, and decreased fecunditywhen reared in the non-tethal presence of a predatorystonefly. Changes in these parameters which are oftencorrelated with adult fitness may have importantconsequences for population growth.
For C. tentans fecundity is highly correlated withadult female wing length (Macchiusi, 1991). Thus,snrwller females may pay a cost of predator avoidancein terms of fewer eggs. The potential costs of smallbody size in adult males are not so clear Because veryHttle is known about the mating system of C. tentans.

10 S. L. Ball and R. L. Baker
it is extremely difficult to predict the fitness implica-tions of male size. In insects, male size is oftencorrelated with mating success (Thomhill & Alcock,1983). In Drosophila meknogaster Meigen, larger maleshad an increased probability of acquiring a mate,although they also paid a cost in terms of feweroffspring per mating (Pitnick, 1991). Thus, large malesize may not always be advantageous. In chironomidspecies which form mating swarms, small male bodysize is thought to confer a mating advantage{McLachlan, 1986; McLachlan & Allen, 1987; Neems,McLachlan & Chambers, 1990). Mating takes place inflight and small males, which are more aerobatic mayhave greater access to females that enter into theswarm. There does not appear to be any obviousmating advantage for small C. tentans males becausethis species does not form mating swarms (Downes,1969). Instead, mating takes place on the ground andon vegetation at the waters edge (D. Oliver, personalcommunication). Also, larger adults may have a fitnessadvantage through greater longevity, although it isnot known to what extent life span increases relativeto body size for C. tentans.
It is well known that changes in life-history traits,such as size, age at maturity and fecundity, can leadto changes in the intrinsic rate of population growth(Caswell & Hastings, 1980; Charlesworth, 1973; Cole,1954). Thus, such indirect effects of predators, as wehave shown here for Chironomus tentans, may exertpotentially important influences on population growthas well as community structure. To determine theimpact of such non-lethal effects, however, we needaccurate measurements of these effects in the field.
Acknowledgements
We thank Donna Bedard (Ministry of NaturalResources, Ontario) for providing us with chironomidsto culture and Hala Ali for her help in the laboratory.Thanks also to Nick Collins and Jean Richardson fortheir constructive comments on earlier versions of themanuscript. Research was supported by an NSERCCanada operating grant to R.L.B. and University ofToronto Open Fellowships to S.L.B.
References
Anderson N.H. & Cummins K.W. (1979) Influences ofdiet on the life histories of aquatic insects, journal ofthe fisheries Research Board of Canada, 36, 335-342.
Baker R.L. (1989) Condition and size of damselflies: afield study of food limitation. Oecologia, 81,111-119.
Blois-Heulin C, Crowley RH., Arrington M. & JohnsonD.M. (1990) Direct and indirect effects of predators onthe dominant invertebrates of two freshwater littoralcommunities. Oecologia, 84, 295-306.
Bohanan R.E. (1988) The efficacy of case construction as ananiipredatory behaviour for some lotic chironomidae: costsand benefits. PhD thesis. University of Wisconsin,Madison, WI.
Butler M.G. (1984) Life histories of aquatic insects. TheEcology of Aquatic Insects (eds V.H. Resh and D.M.Rosenberg). Praeger Publishers, KY, U.S.A.
Caswell H. & Hastings A. (1980) Fecundity,developmental time, and population growth rate: ananalytical solution. Theoretical Population Biology, 17,71-79.
Charlesworth B. (1973) Selection in populations withoverlapping generations. V. Natural selection and lifehistories. American Naturalist, 107, 303-311.
Colbo M.H. & Porter G.N. (1979) Effects of the foodsupply on the life history of Simuliidae (Diptera).Canadian Journal of Zoology, 57, 301-306.
Cole L.C. (1954) The population consequences of lifehistory phenomena. Quarterly Review of Biology, 29,103-137.
Collins N.C. (1980) Developmental responses to foodlimitation as indicators of environmental conditionsfor Ephydra cinerea Jones (Diptera). Ecology. 61, 650-661.
Dill L.M. (1987) Animal decision making and its ecologicalconsequences: the future of aquatic ecology andbehaviour. Canadian journal of Zoology, 65, 803-811.
Dixon S.M. & Baker R.L. (1988) Effects of size on predationrisk, behavioural response to fish, and costs of reducedfeeding in larval Ischnura verticalis (Ceonagrionidae:Odonata). Oecologia, 76, 200-205.
Dodson S.I. & Havel J.E. (1988) Indirect prey effects:Some morphological and life history responses ofDaphnia pulex exposed to Notonecta undulata. Limnologyand Oceanography, 33, 1274-1285.
Downes J.A. (1969) The swarming and mating flight ofDiptera. Annual Review of Entomology, 14, 271-298.
Feltmate B.W. & Williams D.D. (1989) Influence ofrainbow trout {Oncorhynchus mykiss) on density andfeeding behaviour of a perlid stonefly. Canadian Journalof Fisheries and Aquatic Sciences, 46, 1575-1580.
Feltmate B.W. & Williams D.D. (1991) Evaluation ofpredator-induced stress on field populations ofstoneflies (Plecoptera). Ecology, 72, 1800-1806.
Fraser D.F. & Cilliam J.F. (1992) Nonlethal impacts ofpredator invasion: facultative suppression of growthand reproduction. Ecology, 73, 959-970.
Ciberson D.J. & Rosenberg D.M. (1992) Effects of
1995 Blackwell Science Ltd, Freshwater Biology, 34, 1-12

temperature, food quantity, and nymphal rearingdensity on life-history traits of a northern populationof Hexagenia (Ephemeroptera: Ephemeridae). Journal ofthe North American Benthological Society, 11, 181-193.
Grant J.W.G. & Bayly l.A.E. (1981) Predator induction ofcrests in morphs of the Daphnia carinata King complex.Limnology and Oceanography, 26, 210-218.
Hanazato T. (1991) Influence of food density on the effectsof a C/ifloborus-released chemical on Daphnia ambigua.Freshwater Biology, 25, 477-483.
Harvel CD. (1990) The ecology and evolution of inducibledefenses. Quarterly Review of Biology, 65, 323-340.
Harvey B.C. (1991) Interactions among stream fishes:predator-induced habitat shifts and larval survival.Oecologia, 87, 29-36.
Havel J.E. (1987) Predator-induced defenses: a review.Predation: Direct and Indirect Impacts on AquaticCommunities (eds W.C. Kerfoot and A. Sih). UniversityPress of New England, Hanover, NH.
Heads P.A. (1986) The costs of reduced feeding due topredator avoidance: potential effects on growth andfitness in Ischnura elegans larvae (Odonata: Zygoptera).Ecological Entomology, 11, 369-377.
Herbert PD.N. & Grewe PM. (1985) ChaoborusAnducedshifts in the morphology of Daphnia ambigua. Limnologyand Oceanography, 30, 1291-1297.
Hershey A.E. (1987) Tubes and foraging behaviour inlarval Chironomidae: implications for predatoravoidance. Oecologia, 73, 236-241.
Hershey A.E. & Dodson S.I. (1987) Predator avoidanceby Cricotopus: cyclomorphosis and the importance ofbeing big and hairy. Ecology, 68, 913-920.
Lawler S.P. (1989) Behavioural responses to predatorsand predation risk in four species of larval anurans.Animal Behaviour, 38, 1039-1047.
Lima S.L. & Dill L.M. (1990) Behavioural decisions madeunder the risk of predaHon: a review and prospectus.Canadian Journal of Zoology, 68, 619-640.
Lively CM. (1986a) Predator-induced shell dimorphismin the acom bamacle Chthamalus anisopoma. Evolution,40, 232-242.
Lively CM. (1986b) Competition, comparative lifehistories, and maintenance of shell dimorphism in abamacle. Ecology, 67, 858-864.
Macchiusi F. (1991) Effects of predators and food availabilityon activity and growth of Chironomus tentans{Chironomidae: Diptera). MSc thesis. University ofToronto.
Macchiusi F. & Baker R.L. (1991) Prey behaviour andsize-selective predation by fish. Freshwater Biology, 25,533-538.
Macchiusi F. & Baker R.L. (1992) Effects of predators andfood availability on activity and growth of Chironomus
© 1995 Blackwell Science Ltd, Freshwater Biology. 34, 1-12
Life history of adult Chironomus tentans 11
tentans (Chironomidae: Diptera). Freshwater Biology, 28,207-216.
McLachlan A.J. (1986) Sexual dimorphism in midges:strategies for flight in the rain-pool dweller Chironomusimicola (Diptera: Chironomidae). Journal of AnimalEcology. 55, 261-267.
McLachlan A.J. & Allen D.F. (1987) Male mating successin Diptera: advantages of small size. Oikos, 48, 11-14.
Neems R.M., McLachlan A.J. & Chambers R. (1990)Body size and lifetime mating success of male midges(Diptera: Chironomidae). Animal Behaviour. 40,648-652.
Oliver D.R. (1971) Life history of Chironomidae. AnnualReview of Entomology, 16, 211-230.
Parejko K. (1991) Predation by chaoborids on typical andspined Daphnia pulex. Freshwater Biology, 25, 211-217.
Peckarsky B.L., Cowan CA., Penton M.A. & AndersonC (1993) Sublethal consequences of stream-dwellingpredatory stoneflies on mayfly growth and fecundity.Ecology, 74, 1836-1846.
Persson L. (1983) Predator-mediated competition in preyrefuges: the importance of habitat dependent preyresources. Oikos, 68, 12-22.
Pitnick S. (1991) Male size influences mate fecundity andremating interval in Drosophila mclanogasler. AnimalBehaviour, 41, 735-745.
Prejs A. (1987) Risk of predation and feeding rate intropical freshwater fishes: field evidence. Oecotogia, 72,259-262.
Ramcharan CW. & Sprules W.G. (1991) Predator-inducedbehavioural defense and its ecological consequencesfor two calanoid copepods. Oecologia, 86, 275-286.
Rempel R.S. & Carter J.C.H. (1987) Temperatureinfluences on adult size, development and reproductivepotential of aquatic diptera. Canadian Journal of Fisheriesand Aquatic Sciences, 44, 1743-1752.
Rosillon D. (1988) Food preference and relative influenceof temperature and food quality on life historycharacteristics of a grazing mayfly, Ephemerella ignita(Poda). Canadian Journal of Zoology, 66, 1474-1481.
Rowe L. & Ludwig D. (1991) Size and timing ofmetamorphosis in complex life cycles: time constraintsand variation. Ecology, 72, 413-427.
Semlitsch R.D. (1987a) Interaction between fish andsalamander larvae: costs of predator avoidance orcompetifion? Oecologia. 72, 481-186.
Semlitsch R.D. (1987b) Density-dependent growth andfecundity in the paedomorphic salamander Ambystomatalpoideum. Ecology, 68, 1003-1008.
Semlitsch R.D., Scott D.E. & Pechman H.K. (1988) Tuneand size at metamorphosis related to adult fitness inAmbystoma talpoideum. Ecology, 69, 184-192.
Short T.M. & Holomuzki J.R. (1992) Indirect effects of

12 S. L Ball and R. I. Baker
fish on foraging behaviour and leaf processing by theisopod Lirceus fontinalis. Freshwater Biology, 27, 91-97.
Sibly R.M. & Calow P. (1989) A life-cycle theory ofresponses to stress. Biological Journal of the LinneanSociety, 37, 101-116.
Sih A. 1987. Predators and prey lifestyles: an evolufionaryand ecological overview. Predation: Direct and IndirectImpacts on Aquatic Communities (eds W.C. Kerfoot andA. Sih). University Press of New England, Hanover,NH, U.S.A.
Sih A., Krupa J. &. Travers S. (1990) An experimentalstudy on the effects of predation risk and feedingregime on the mating behaviour of the water strider.American Naturalist, 135, 284-290.
Sih A. (1992) Integrafive approaches to the study ofpredafion: general thoughts and a case study on sunfishand salamander larvae. Anrmles Zoologid Fennid. 29,183-198.
Skelly D.K. & Werner E.E. (1990) Behavioural and Ufe-historical responses of larval American toads to anodonate predator. Ecology, 7, 2313-2322.
Stauffer H.P, & Semlitsch R.D. (1993) Effects of visual.
chemical and tactile cues of fish on the behaviouralresponses of tadpoles. Animal Behaviour, 46, 355-364.
Stenson J.A.E. (1987) Variafion in capsule size ofHolopedium gibberum (Zaddach): a response toinvertebrate predafion. Ecology. 68, 928-934.
Thornhill R, & Alcock J. (1983) The Evolution of InsectMating Systems. Harvard University Press, Cambridge.
Vanni M.J. & Lampert W. (1992) Food quality effects onlife history traits and fitness in the generalist herbivoreDaphnia. Oecologia, 92, 48-57.
Wemer E.E. (1991) Nonlethal effects of a predator oncompetitive interacfions between two anuran larvae.Ecology, 72, 1709-1720.
Wemer E.E., GilUam J.R, Hall D.J. & Mittelbach G.G.(1983) An experimental test of the effects of predationrisk on habitat use in fish. Ecology, 64, 1540-1548.
Wilbur H.M. (1980) Complex life cycles. Annual Reviewof Ecology and Systematics, 11, 67-93.
WUbur H.M. & CoUins J.P (1973) Ecological aspects ofamphibian metamorphosis. Science, 182, 1305-1314.
Zar J.H. (1984) Biostatistical Analysis. 2nd edition. Prentice-Hall, NJ.
{Manuscript accepted 5 January 1995)
1995 Biackwell Science Ltd, Freshwater Biology, 34,1-12

Copyright © 2022 FDOKUMEN




















