
The Myelopoietic Supportive Capacity of Mesenchymal Stromal Cells Is Uncoupled from Multipotency and...
12
The Myelopoietic Supportive Capacity of Mesenchymal Stromal Cells Is Uncoupled from Multipotency and Is Influenced by Lineage Determination and Interference with Glycosylation VERED MORAD, a MEIRAV PEVSNER-FISCHER, a SIVAN BARNEES, a ALBENA SAMOKOVLISKY, b LIAT ROUSSO-NOORI, a RAKEFET ROSENFELD, b DOV ZIPORI a a Department of Molecular Cell Biology, the Weizmann Institute of Science, Rehovot, Israel; b Procognia (Israel) Ltd., Ashdod, Israel Key Words. Mesenchymal stem cell • Mesenchyme • Hemopoietic support • Multipotency • Differentiation ABSTRACT Cultured bone marrow stromal cells create an in vitro mi- lieu supportive of long-term hemopoiesis and serve as a source for multipotent mesenchymal progenitor cells de- fined by their ability to differentiate into a variety of meso- dermal derivatives. This study aims to examine whether the capacity to support myelopoiesis is coupled with the multi- potency. Our results show that the myelopoietic supportive ability of stromal cells, whether from the bone marrow or from embryo origin, is not linked with multipotency; cell populations that possess multipotent capacity may or may not support myelopoiesis, whereas others, lacking multipo- tency, may possess full myelopoietic supportive ability. How- ever, upon differentiation, the ability of multipotent mesen- chymal progenitors to support myelopoiesis is varied. Osteogenic differentiation did not affect myelopoietic sup- portive capacity, whereas adipogenesis resulted in reduced ability to support the maintenance of myeloid progenitor cells. These differences were accompanied by a divergence in glycosylation patterns, as measured by binding to lectin microarrays; osteogenic differentiation was associated with an increased level of antennarity of N-linked glycans, whereas adipogenic differentiation caused a decrease in an- tennarity. Inhibition of glycosylation prior to seeding the stroma with bone marrow cells resulted in reduced capacity of the stromal cells to support the formation of cobblestone areas. Our data show that myelopoietic support is unrelated to the multipotent phenotype of cultured mesenchymal pro- genitors but is dependent on the choice of differentiation pathway and upon correct glycosylation of the stromal cells. STEM CELLS 2008;26:2275–2286 Disclosure of potential conflicts of interest is found at the end of this article. INTRODUCTION Cultured stromal cells from the bone marrow recreate a hemopoi- etic inductive microenvironment in vitro [1–3]; upon the formation of confluent adherent cell layers the stromal mesenchyme serves as a support for the lodging and long-term proliferation of hemopoi- etic stem cells. Similarly, these cells may promote engraftment of hemopoietic stem cells in vivo [4]. Cells derived from such stromal cultures were further identified as multipotent stromal cells [5, 6], or mesenchymal stem cells (MSCs) [7–9], since they give rise to mesodermal derivatives such as muscle, bone, cartilage, and fat [10, 11]. Distinct mesenchymal cell types from bone marrow cultures have been isolated and propagated in our laboratory as permanent cell lines (MBA series) of osteoblasts, preadipocytes, fibroblasts, as well as cells with endothelial properties [12–14]. In recent years several reports demonstrated the existence of cells, such as multipotent adult progenitor cells (MAPCs) [15, 16], un- restricted somatic stem cells [17], marrow-isolated adult multilin- eage inducible cells [18], amniotic fluid-derived cells [19], skin progenitor cells [20], very small embryonic-like cells [21], and stage-specific embryonic antigen-1 positive cells [22], that give rise either to cells of all embryo germ lines, or in other cases, cells that represent several unexpected lineages. The model that emerges from these studies is of an early mesen- chymal cell that possesses pluripotent stem cell properties. In analogy with the hemopoietic system, it has been suggested that MSCs, as well as the other related cell types, differen- tiate in a hierarchical manner by giving rise to cells of declining potency for self-renewal and decreasing range of cell types to which they would differentiate into [23]. In the hemopoietic system, discrete types of committed progenitors were identified according to cell surface marker expression. These can be isolated to homogeneity and proven to be tripotent, bipotent, or monopotent. By contrast, no restricted progenitors belonging to the MSC lineage have been identi- fied with any certainty (reviewed in [24]). The question whether differentiation of MSCs obeys a hierarchical cascade of decreasing range of differentiation directions therefore remains open. It is further unclear where, within the differ- entiation cascade of MSCs, the hemopoietic supportive ca- pacity is positioned. Author contributions: V.M., M.P.F., and S.B.: design and performance of the biological characterization of mesenchymal cells; L.R.-N.: derivation of MSCs; A.S. and R.R.: performance of the glycoanalysis; D.Z.: study conception. Correspondence: Dov Zipori, Ph.D., Department of Molecular Cell Biology, Weizmann Institute of Science, Rehovot 76,100, Israel. Telephone: 972-8-9342484; Fax: 972-8-9344125; e-mail: [email protected] Received July 9, 2007; accepted for publication June 4, 2008; first published online in STEM CELLS EXPRESS June 12, 2008. ©AlphaMed Press 1066-5099/2008/$30.00/0 doi: 10.1634/ stemcells.2007-0518 THE STEM CELL NICHE S TEM CELLS 2008;26:2275–2286 www.StemCells.com
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of The Myelopoietic Supportive Capacity of Mesenchymal Stromal Cells Is Uncoupled from Multipotency and...

The Myelopoietic Supportive Capacity of Mesenchymal StromalCells Is Uncoupled from Multipotency and Is Influenced by LineageDetermination and Interference with Glycosylation
VERED MORAD,a MEIRAV PEVSNER-FISCHER,a SIVAN BARNEES,a ALBENA SAMOKOVLISKY,b
LIAT ROUSSO-NOORI,a RAKEFET ROSENFELD,b DOV ZIPORIa
aDepartment of Molecular Cell Biology, the Weizmann Institute of Science, Rehovot, Israel; bProcognia (Israel)Ltd., Ashdod, Israel
Key Words. Mesenchymal stem cell • Mesenchyme • Hemopoietic support • Multipotency • Differentiation
ABSTRACT
Cultured bone marrow stromal cells create an in vitro mi-lieu supportive of long-term hemopoiesis and serve as asource for multipotent mesenchymal progenitor cells de-fined by their ability to differentiate into a variety of meso-dermal derivatives. This study aims to examine whether thecapacity to support myelopoiesis is coupled with the multi-potency. Our results show that the myelopoietic supportiveability of stromal cells, whether from the bone marrow orfrom embryo origin, is not linked with multipotency; cellpopulations that possess multipotent capacity may or maynot support myelopoiesis, whereas others, lacking multipo-tency, may possess full myelopoietic supportive ability. How-ever, upon differentiation, the ability of multipotent mesen-chymal progenitors to support myelopoiesis is varied.Osteogenic differentiation did not affect myelopoietic sup-
portive capacity, whereas adipogenesis resulted in reducedability to support the maintenance of myeloid progenitorcells. These differences were accompanied by a divergence inglycosylation patterns, as measured by binding to lectinmicroarrays; osteogenic differentiation was associated withan increased level of antennarity of N-linked glycans,whereas adipogenic differentiation caused a decrease in an-tennarity. Inhibition of glycosylation prior to seeding thestroma with bone marrow cells resulted in reduced capacityof the stromal cells to support the formation of cobblestoneareas. Our data show that myelopoietic support is unrelatedto the multipotent phenotype of cultured mesenchymal pro-genitors but is dependent on the choice of differentiationpathway and upon correct glycosylation of the stromal cells.STEM CELLS 2008;26:2275–2286
Disclosure of potential conflicts of interest is found at the end of this article.
INTRODUCTION
Cultured stromal cells from the bone marrow recreate a hemopoi-etic inductive microenvironment in vitro [1–3]; upon the formationof confluent adherent cell layers the stromal mesenchyme serves asa support for the lodging and long-term proliferation of hemopoi-etic stem cells. Similarly, these cells may promote engraftment ofhemopoietic stem cells in vivo [4]. Cells derived from such stromalcultures were further identified as multipotent stromal cells [5, 6],or mesenchymal stem cells (MSCs) [7–9], since they give rise tomesodermal derivatives such as muscle, bone, cartilage, and fat[10, 11]. Distinct mesenchymal cell types from bone marrowcultures have been isolated and propagated in our laboratory aspermanent cell lines (MBA series) of osteoblasts, preadipocytes,fibroblasts, as well as cells with endothelial properties [12–14]. Inrecent years several reports demonstrated the existence of cells,such as multipotent adult progenitor cells (MAPCs) [15, 16], un-restricted somatic stem cells [17], marrow-isolated adult multilin-eage inducible cells [18], amniotic fluid-derived cells [19], skinprogenitor cells [20], very small embryonic-like cells [21], and
stage-specific embryonic antigen-1 positive cells [22], thatgive rise either to cells of all embryo germ lines, or in othercases, cells that represent several unexpected lineages. Themodel that emerges from these studies is of an early mesen-chymal cell that possesses pluripotent stem cell properties. Inanalogy with the hemopoietic system, it has been suggestedthat MSCs, as well as the other related cell types, differen-tiate in a hierarchical manner by giving rise to cells ofdeclining potency for self-renewal and decreasing range ofcell types to which they would differentiate into [23]. In thehemopoietic system, discrete types of committed progenitorswere identified according to cell surface marker expression.These can be isolated to homogeneity and proven to betripotent, bipotent, or monopotent. By contrast, no restrictedprogenitors belonging to the MSC lineage have been identi-fied with any certainty (reviewed in [24]). The questionwhether differentiation of MSCs obeys a hierarchical cascadeof decreasing range of differentiation directions thereforeremains open. It is further unclear where, within the differ-entiation cascade of MSCs, the hemopoietic supportive ca-pacity is positioned.
Author contributions: V.M., M.P.F., and S.B.: design and performance of the biological characterization of mesenchymal cells; L.R.-N.:derivation of MSCs; A.S. and R.R.: performance of the glycoanalysis; D.Z.: study conception.
Correspondence: Dov Zipori, Ph.D., Department of Molecular Cell Biology, Weizmann Institute of Science, Rehovot 76,100, Israel.Telephone: 972-8-9342484; Fax: 972-8-9344125; e-mail: [email protected] Received July 9, 2007; accepted for publication June4, 2008; first published online in STEM CELLS EXPRESS June 12, 2008. ©AlphaMed Press 1066-5099/2008/$30.00/0 doi: 10.1634/stemcells.2007-0518
THE STEM CELL NICHE
STEM CELLS 2008;26:2275–2286 www.StemCells.com

The ability of bone marrow mesenchyme to support hemo-poiesis is thought to be dependent upon a multitude of factorsincluding components of the extracellular matrix (ECM), cellsurface constituents, and soluble factors (including cytokinesand small molecules) that make up the stem cell niche [3,25–27]. The minimal requirements for hemopoietic supporthave not, thus far, been conclusively defined. We have shownthat the interactions of hemopoietic progenitors with the bonemarrow stroma are affected by free sugars. Some of these effectswere related to the metabolism of these sugars [28]. In contrast,the use of saccharides that do not metabolize indicated thepossible involvement of glycosylated moieties in the interac-tions between mesenchymal cells and hemopoietic progenitors[29].
In the present study, we aimed at examining the relationshipbetween the multipotent phenotype of mesenchymal progenitorsand their corresponding capacity to support myelopoiesis. Weshow that the myelopoietic supportive capacity of the mesen-chyme is not linked to the multipotency of these cells. Further-more, a change in glycosylation patterns of the mesenchymalcells occurred in a specific manner related to their differentia-tion to either adipogenic or osteogenic cells. Inhibition of gly-cosylation is shown to reduce the ability of the stroma to supportthe formation of hemopoietic foci, suggesting a role for glyco-sylated moieties in the control of interactions between thestroma and the hemopoietic progenitors.
MATERIALS AND METHODS
AnimalsMice were maintained under specific pathogen-free conditions.C57BL/6J and Balb/c mice were purchased from Harlan (Rehovot,Israel, http://www.teklad.com). TCR�-deficient mice (C57BL/6J-Tcrbtm1Mom) were obtained from the Jackson Laboratory (Bar Har-bor, ME, http://www.jax.org) and propagated in the WeizmannInstitute’s animal housing facilities. All animal procedures wereapproved by the Weizmann Institute Animal Care Committee.
Cell CultureMBA-13, MBA-15, C3H10T1/2, 14F1.1, NIH-3T3L1 cell lines,and mouse embryo fibroblasts (MEFs) were grown in Dulbecco’smodified Eagle’s medium (DMEM) (Gibco, Grand Island, NY,http://www.invitrogen.com) supplemented with 10% heat-inacti-vated fetal calf serum (FCS) (Biological Industries Ltd., BeitHaemek, Israel, http://www.bioind.com), selected according to itscapacity to support the growth of the ABLS-8 pre-B-lymphoma cellline at the population doubling time of 10 hours, supplemented with60 �g/ml penicillin, 100 �g/ml streptomycin, and 50 �g/ml kana-mycin. MEFs were obtained from day-14 gestation embryo frag-ments treated with trypsin-EDTA and were then propagated inaccordance with procedures previously developed in our laboratoryfor the maintenance of nontumorigenic mesenchyme [12, 13] asfollows: cells were examined frequently, following seeding anduntil reaching confluence. They were then passaged each timeconfluence was regained, rather than at fixed time intervals. Usingthis approach the cells did not undergo crisis and transformation anddid not undergo senescence for more than 15 passages. Antigensmost prominently expressed by MEFs were intercellular adhesionmolecule 1, major histocompatibility complex I, and CXC chemo-kine receptor 4, whereas the hemopoietic marker CD45 was notexpressed at a detectable level. MEF-1, -4, -5, and -8 were derivedfrom C57BL/6J mouse embryos. MEF-2 was derived from hetero-zygote TCR��/� mouse embryos. MEF-3, -6, and -7 were derivedfrom TCR�-deficient mice. MSCs were grown in murine Mesen-Cult Basal Media supplemented with 20% murine mesenchymalsupplement (StemCell Technologies, Vancouver, BC, Canada,http://www.stemcell.com), 60 �g/ml penicillin, and 100 �g/mlstreptomycin. MAPCs were grown in MAPC medium consisting of
60% low-glucose DMEM (Invitrogen Life Technologies, Paisley,Scotland, http://www.invitrogen.com) and 40% MCDB-201 (Sigma-Aldrich, Rehovot, Israel), supplemented with 1 � insulin-trans-ferrin-selenium, 1 � linoleic acid–bovine serum albumin (BSA),10–8 M dexamethasone, 10–4 M ascorbic acid 2-phosphate (all fromSigma-Aldrich), 60 �g/ml penicillin, 100 �g/ml streptomycin alongwith 2% fetal bovine serum (FBS) (HyClone Laboratories, Logan,UT, http://www.hyclone.com), 1,000 units/ml leukemia inhibitoryfactor (LIF) (Chemicon, Temecula, CA, http://www.chemicon.com), 10 ng/ml epidermal growth factor (EGF) (Sigma-Aldrich),and 10 ng/ml platelet-derived growth factor (PDGF)-BB (Pepro-Tech/Cytolab, Rehovot, Israel, http://www.peprotech.com) as de-scribed [15, 30]. All cells were incubated at 37°C in a humidifiedatmosphere of 10% CO2.
MAPC DerivationBone marrow (BM) was collected from the femur and tibia of4-week-old female C57BL/6J mice (n � 7). BM mononuclear cellswere obtained by Ficoll separation and plated on fibronectin (Sigma-Aldrich)-coated plates in MAPC expansion medium containing 2%FBS, EGF, PDGF-BB, and LIF as described [15, 30]. After 6 weeksof expansion in culture, cells were depleted of CD45�/Ter119�
cells using micromagnetic beads (Miltenyi Biotec, Bergisch-Glad-bach, Germany, http://www.miltenyibiotec.com) according to themanufacturer’s instructions. The depleted cells were then replated at10 cells per well in fibronectin-coated 96-well plates and expandedas clones at densities of 2 � 102 cells/cm2. After derivation MAPCswere transferred to MSC medium and these cell strains were des-ignated MAPC-derived MSCs (MD-MSCs).
MSC Isolation and Phenotypic Characterization
BM Cell Extraction. BM cells were obtained from 7- to8-week-old C57BL/6J mice, pelleted, resuspended in phos-phate-buffered saline (PBS) and red blood cells lysis buffer(Sigma-Aldrich) for 5 minutes, and then subjected to an addi-tional centrifugation. The cells were then seeded in 60-mmplates containing MSC medium. Half of the medium was re-placed every 3 days and once a confluent layer was formed, thecells were removed using trypsin (0.05% EDTA, 0.25% trypsin)and reseeded. Cells were grown in culture for 4 weeks until asufficient number of cells was obtained and then subjected tocell sorting.
Cell SortingPrimary BM cells were incubated with antibodies specific toCD45.2 R-phycoerythrin (PE) (Southern Biotechnology Associates,Birmingham, AL, http://www.southernbiotech.com) and CD11b/Mac1 fluorescein isothiocyanate (FITC) (Southern BiotechnologyAssociates) for 1 hour and were then washed and suspended in PBSwith 1% FCS. The cells were sorted using FACSVANTAGE cellsorter (FACSVANTAGE SE; Becton Dickinson ImmunocytometrySystems, San Jose, CA, http://www.bd.com). The double negativecell population was collected and seeded in MSC medium. Pheno-typic characterization was performed as we previously reported[31].
Long-Term Bone Marrow CultureAdherent cells were seeded in 6-well plates (Falcon, FranklinLakes, NJ, http://www.bdbiosciences.com) and allowed to grow toconfluence. BM cells from the femur and tibia of two 6- to 8-week-old Balb/c mice were flushed out and 2 � 105 cells per well wereseeded onto the confluent layers of adherent cells. Cultures weremaintained in � minimum essential medium (Gibco-BRL, Gaith-ersburg, MD, http://www.invitrogen.com) medium supplementedwith 20% horse serum (StemCell Technologies) and 10�6 M hy-drocortisone hemisuccinate (Sigma-Aldrich) at 33°C in a humidi-fied atmosphere of 10% CO2 for 4 weeks unless otherwise speci-fied. Cultures were fed twice a week by replacing half of themedium with fresh medium. After 4 weeks, or on the specified dayin culture, the nonadherent hemopoietic cells were harvested,
2276 MSC Hemopoietic Support Versus Multipotency

counted, and subjected to granulocyte-macrophage colony-formingunit (GM-CFU) assay. Cells were seeded in methylcellulose semi-solid medium supplemented with 10 ng/ml interleukin (IL)-3, 10ng/ml IL-6, 50 ng/ml stem cell factor (SCF) (all from PeproTech/Cytolab), and 3 units/ml erythropoietin (Epoetin alfa; Ortho-BiotechJanssen-Cilag, Baar, Switzerland, http://www.orthobiotech.de).Cultures were maintained at 37°C, 10% CO2 and scored, by mor-phology, on day 8.
Flow Cytometry AnalysisAnti-CD45–phycoerythrin, anti-CD11b–PE, anti-c-Kit-PE, anti–Sca-1–PE, rat IgG2a isotype control–PE, anti-Gr-1–FITC, anti-CD34–FITC, and rat IgG2b isotype control–FITC were purchased fromeBioscience (San Diego, http://www.ebioscience.com). Two weeksafter the seeding of BM cells in long-term bone marrow culture(LTBMC) conditions, hemopoietic cells were harvested and incu-bated with specific antibodies for 1 hour. Next, cells were subjectedto flow cytometry analysis using a FACScan flow cytometer (Bec-ton Dickinson Immunocytometry Systems). Cells were gated ac-cording to their high fluorescence intensity.
Mesodermal Lineage Differentiation, Detection, andQuantificationThe basic medium used in all differentiation experiments wasDMEM � 10% FCS (HyClone Laboratories).
AdipogenesisCells were seeded at concentration to reach subconfluence in a24-well plate. The following day, adipoinductive medium wasadded. Two conditions for adipogenesis were used: medium sup-plemented with 10 �g/ml insulin, 0.5 mM 3-isobutyl-1-methyl-xanthine, and 1 � 10-6 M dexamethasone (all from Sigma-Aldrich)or medium supplemented with 1.5 unit/ml human regular insulin(100 IU/ml Lilly HI0210 [Lilly, Indianapolis, http://www.lilly.com]). Cells were grown for 4 weeks with medium replaced twiceweekly. Both adipoinductive media were used for the comparativestudy of mesenchymal cell populations. In all other experiments,adipogenic differentiation of MSCs and 14F1.1 was carried outusing 1.5 unit/ml human regular insulin only. Adipogenesis wasdetected by oil red O staining. For oil red O quantification, 4%IGEPAL CA 630 (Sigma-Aldrich) in isopropanol was added to eachwell. Light absorbance by the extracted dye was measured in 520nm. Values were normalized to protein concentration.
OsteogenesisCells were seeded at concentration to reach subconfluence in a24-well plate. The next day osteoinductive medium containing 50�g/ml L-ascorbic acid 2-phosphate (Sigma-Aldrich), 10 mM glyc-erol 2-phosphate disodium salt (Sigma-Aldrich), and 1 � 10�8 Mdexamethasone was added. The cells were grown for 2 (for alkalinephosphatase [ALP] staining) or 4 weeks (for alizarin red staining)with medium replaced twice a week. Osteogenic differentiation wasdetected by alizarin red staining. For alizarin red quantification, 0.5N HCl, 5% SDS was added to each well. Light absorbance by theextracted dye was measured in 415 nm. Values were normalized toprotein concentration. ALP activity was detected by BCIP/NBTsubstrate chromogen system (Dakocytomation, Glostrup, Denmark,http://www.dakocytomation.com) according to the manufacturer’sinstructions.
ChondrogenesisCells were grown in micromass culture supplemented with chon-droinductive medium for 4 weeks. Cells at 2 � 105 per tube werecentrifuged for 5 minutes at 1,200g in 15-ml conical polyproylanetubes. Following centrifugation, the supernatant was gently re-moved and 1 ml of chondroinductive medium containing 0.1 mML-ascorbic acid 2-phosphate, 10 ng/ml human TGF�1 (PeproTech/Cytolab), 1 � 10�7 M dexamethasone was added. The tubes wereincubated with the cap slightly loose, with medium replacementtwice a week. After 4 weeks in culture the pellets were fixed with4% paraformaldehyde and embedded in 1.5% low-melting agarose
(Sigma-Aldrich) solution followed by paraffin embedding. Chon-drogenesis was detected by Alcian Blue staining.
Statistical AnalysisThe Wilcoxon rank sum test using Matlab v.7.1 Statistical toolbox(The Mathworks, Inc., Natick, MA, http://www.mathworks.com)was applied to compare the mean of cell counts and GM-CFUcolonies produced in LTBMC. Differences were considered statis-tically significant with p � .05 for the comparison of 14F1.1hemopoietic support after adipogenic differentiation (control-adipo)or with p � .025 when two analyses were performed (MSC control-adipo and control-osteo) to correct for multiple hypotheses with theBonferroni correction. For statistical analysis of cobblestone areaformation, Student’s paired t test was performed.
Semiquantitative Reverse-Transcription–PolymeraseChain ReactionTotal RNA was extracted from MSCs and 14F1.1 using Nucleo-Spin RNA II kit according to manufacturer’s instructions (Mach-erey Nagel, Duren, Germany, http://www.macherey-nagel.com).Two micrograms of total RNA were reverse transcribed usingMoloney murine leukemia virus reverse transcriptase (Promega,Madison, WI, http://www.promega.com). Polymerase chain reac-tion (PCR) amplification of the cDNA was performed with Reddy-Mix PCR Master Mix (ABgene, Surrey, U.K., http://www.abgene.com). glyceraldehyde-3-phosphate dehydrogenase expression wasused as a control for loading and water was used as negative control.Primer sequences used are as follows: Gene:Sense/Antisense
SCF:GCTTGACTACTCTTCTGGACAA/CTGCTGTCATTCCT-AAGGGAG
Flt3-L:CAGTCCCATCTCCTCCAACTT/AGCTGTGTGCAGGT-GTCCTTC
M-CSF:AGTGAGGGATTTTTGACCCAGGAAGCAAA/CTAT-ACTGGCAGTTCCACCTGTCTGT
IL-6:TGCACTTGCAGAAAACAATC/TGGTCTTGGTCCTT-AGCC
GM-CSF:CATTGTGGTCTACAGCCTCTC/GGCAGTATGTCT-GGTAGTAGC
LIF:CATAATGAAGGTCTTGGCCGCA/TGCCATTGAGCTGT-GCCAGTTG
IL-3:CCACCGTTTAACCAGAACGTTG/TCCACGGTTAGGA-GAGACGGA
G-CSF:TGCAGCAGACACAGTCCCTAA/TGGCTGCCACTGT-TTCTTTAGG
GAPDH:ACCACAGTCCATGCCATCAC/TCCACCACCCTGT-TGCTGTA
GlycoprofilingGlycoanalysis of membrane protein extracts was performed usinglectin microarrays.
Sample Preparation. MSCs were grown in 10-cm plates and in-duced to differentiate into adipocytes for 2 weeks and osteoblastsfor 3 weeks with differentiation medium. Control plates were grownin MSC medium and harvested at confluence. NIH-3T3L1 fibro-blasts were grown in 10-cm plates coated with 0.1% gelatin andincubated with or without adipoinductive medium for 10 days.Membrane proteins were extracted from the cells using the Qpro-teome Cell Compartment Kit (Qiagen, Hilden, Germany, http://www1.qiagen.com), as described in the user manual; briefly, ex-traction buffer CE1 was added to cells. This buffer selectivelydisrupts the plasma membrane without solubilizing it, thereby re-sulting in the release of cytosolic proteins. Lysates were centrifugedat 1,000g for 10 minutes at 4°C. The pellet, which contains intactplasma membranes and organelles, such as nuclei, mitochondria, andthe endoplasmic reticulum, was resuspended in extraction bufferCE2, which solubilizes all cellular membranes with the exception ofthe nuclear membrane. The suspension was centrifuged at 6,000gfor 10 minutes at 4°C. The resulting supernatants, which primarilycontain membrane proteins, were biotinylated using N-hydroxysuc-cinimide biotin (Pierce, Rockford, IL, http://www.piercenet.com) ata ratio of 5:1 biotin molecules per protein molecule. Protein con-
2277Morad, Pevsner-Fischer, Barnees et al.
www.StemCells.com

centrations were measured using the BCA Protein Quantification kit(Pierce). The resulting samples were dialyzed in a Slide-A-LyzerMini Dialysis unit with a molecular weight cutoff of 7,000 D(Pierce) for 48 hours.
Glycoanalysis. Lectin microarrays, provided by Procognia Ltd.(Ashdod, Israel, http://www.procognia.com), were blocked using1% BSA (Sigma-Aldrich) and probed with each of the proteinsamples. The arrays were washed with PBS buffer containing 1 mMCaCl2, 1 mM MgCl2, and 0.1 mM MnCl2. Detection of boundsamples was performed using a second step of incubation withcyanin 3-conjugated streptavidin, and arrays were scanned with anAgilent microarray scanner (Palo Alto, CA, http://www.agilent.com). Results are shown for one of three experiments performed foreach differentiation condition. To determine which differences inlectin signals are significant, the two histograms of lectin signals tobe compared are normalized using a robust regression algorithm.The particular algorithm used is a robust regression with maximumlikelihood estimates with high breakdown estimates [32]. This al-gorithm provides both the normalization factor between the twohistograms, and an estimate of the similarity between them, whichcomes from the quality of the fit. This similarity is calculated usingthe weighted root mean square sum of the fit residuals (�), wherethe weights used are the factors assigned to each point by the robustregression calculation. The differences between signals in the twohistograms are then calculated in terms of �, and each differencelarger than 2� is considered significant.
Glycosylation Inhibitor TreatmentMSCs were incubated for 3 days in the absence or presence of 0.4mg/ml deoxymannojirimycin (DMJ; Sigma-Aldrich). Total mem-
brane proteins were extracted from cells and applied to the lectinarrays. Profiles obtained from different lectins specific to complex,terminal sugars of complex glycans and high mannose were de-tected. MSCs were similarly treated with or without DMJ for 2days, washed four times with PBS. Cultures were then seeded withBM cells and subjected to LTBMC conditions. Two and a halfweeks later the number of cobblestones was scored.
RESULTS
Mesenchymal Cell Populations Differ in TheirMyelopoietic Supportive CapacityWe intercompared a series of mesenchymal cell populations fortheir capacity to support myelopoiesis in long-term bone marrowculture. The mesenchymal populations selected for this analysiswere (a) stromal cell lines and clones of bone marrow origin, (b)MSCs, (c) cells obtained according to MAPC derivation procedureand transferred to MSC conditions (MD-MSCs), (d) the MSC-likecell line C3H10T1/2 and, (e) several strains of independentlyderived MEFs. The mesenchymal cells at confluence served as asupportive stroma layer for the seeding of fresh bone marrow cells.Twice a week, the cultures were demidepopulated. The accumula-tion of hemopoietic cells in the liquid phase of the cultures, as wellas the incidence of myeloid progenitors within this population, wasdetermined at the time points indicated. Figure 1A and 1B showone example of bone marrow cultures supported by MEFs. Hemo-poietic cells including myeloid progenitors could be detected until
Figure 1. Myelopoietic supportive capacityof mesenchymal cell populations. (A, B):MEFs support long-term hemopoietic cul-tures starting from total bone marrow. Bargraphs show the total hemopoietic cell count(A) and myeloid progenitor count (granulo-cyte-macrophage colony-forming unit) perculture (B) at different time points. (C, D):Comparison of the ability of different mes-enchymal cell populations to support myelo-poiesis. Hemopoietic cell count (C) and my-eloid progenitors (D) were determined after4 weeks in culture. Abbreviations: E, earlypassages; L, late passages; MEFs, mouseembryo fibroblasts.
2278 MSC Hemopoietic Support Versus Multipotency

Figure 2. Analysis of the ability of different mesenchymal populations to differentiate into mesodermal lineages. (A): MBA-15, MSCs, 14F1.1, multipotent adultprogenitor cell-derived MSC (MD-MSC)-B, MEF-4, and MEF-5 were tested for their ability to differentiate in vitro into adipogenic, osteogenic, and chondrogeniclineages. Adipogenesis was indicated by accumulation of lipid droplets stained with oil red O. Osteogenesis was indicated by the increase of ALP expression ininduced samples compared with control and by calcium mineralization as detected by alizarin red stain. Chondrogenesis was detected by accumulation ofcartilaginous proteoglycans as detected by Alcian Blue staining. Quantification of adipogenic (B) and osteogenic (C) differentiation of mesenchymal cell populations.Cells were cultured in conditions favoring adipogenesis (B): insulin alone (dark gray bars); combination of insulin (Ins), IBMX, and dexamethasone (DEX) (lightgray bars); or without any inducers (black bars); or in conditions favoring osteogenesis (white bars) (C). After 4 weeks of culture cells were stained with oil red O(B) or alizarin red (C). The dye was extracted and measured at 520 nm (B) or 415 nm (C) using spectrophotometer. Values were normalized to protein concentrations.Scale bar � 100 �m. Abbreviations: E, early passages; L, late passages; MEF, mouse embryo fibroblast; MSCs, mesenchymal stem cells; OD, optical density.
2279Morad, Pevsner-Fischer, Barnees et al.
www.StemCells.com

the 43rd day of culture, when the experiment was terminated.Similar experiments were performed with all other mesenchymalpopulations. The data are summarized as number of cells andmyeloid progenitors per culture at 4 weeks of incubation (Fig. 1C,1D). The results represent one of at least three experiments per-formed for each cell population. Whereas the bone marrow–de-rived 14F1.1 preadipogenic stromal cell line supported effectivelylong-term myelopoiesis, other stromal cell lines from bone marroworigin were devoid of this activity. Similarly, some MEF strainswere supportive of myelopoiesis, whereas others, derived using thesame method and under the same conditions, were completelydevoid of this property (Fig. 1C, 1D). Some MEFs retained aconstant phenotype upon repeated passaging, with regard to theireffect of long-term myelopoiesis. Others, such as strain MEF-1 andMEF-3, were unstable, that is, these cells did not support myelo-poiesis at early passages but did perform well at later passages.MSCs showed a stable ability to support long-term myelopoiesis.Conversely, MD-MSCs were ineffective in creating in vitro con-ditions appropriate for myelopoiesis (Fig. 1C, 1D).
Mesenchymal Cell Populations Vary in TheirCapacity to Differentiate into MesodermalDerivativesCells within mesenchymal populations are capable of multilin-eage differentiation and are therefore designated as MSCs. Wetested the multipotent phenotype in the above-studied mesen-chymal populations. As we previously reported, the cells wereinduced toward adipogenesis, osteogenesis, and chondrogenesis[31]. Examples of such induced differentiation are shown inFigure 2A, and quantitative determination of adipogenesis andosteogenesis for the entire cell series is presented in Figure 2B
and 2C, respectively. Clearly, different mesenchymal popula-tions had a divergent capacity to differentiate into mesodermallineages. Although to a different degree, adipogenic potentialwas very prevalent in the different cell populations, whereasosteogenic potential was rare. Adipogenesis was most promi-nently observed in MSCs, C3H10T1/2, and in some of the MEFstrains, particularly MEF-4. MSCs had the highest osteogeniccapacity. The MBA-13 and MBA-15 stromal cell lines differ-entiated effectively into osteogenic cells, as we previously re-ported [33], as did MEF-1L and MEF-4 (Fig. 2B, 2C).
The Myelopoietic Supportive Capacity ofMesenchymal Cells Is Uncoupled from Their MSCMultipotent PhenotypeA schematic summary and comparison between the cell’s abilityto differentiate into the three mesodermal lineages studied aboveand their corresponding capacity to support myelopoiesis isshown in Table 1. Whereas marrow-derived stromal cell linesthat are highly restricted to adipogenesis (14F1.1) had an out-standing capacity to support myelopoiesis, others that differen-tiated into three mesodermal lineages were devoid of suchcapacity (MBA-15). Similarly, MEF-7 exhibited limited differ-entiation (adipogenic potential and some chondrogenic ability)but showed a strong myelopoietic supportive activity. MEF-4supported myelopoiesis as effectively as MEF-7 but in contrastto the latter had a prominent differentiation capacity. Similarly,MSCs that differentiate into all three lineages supported myelo-poiesis effectively. Thus, the stem cell potential of a givenmesenchymal population does not correspond to, and does notpredict, the ability of these cells to create conditions favorablefor myelopoiesis.
Table 1. Summery of the ability of mesenchymal cells to differentiate into mesodermal lineages and to give rise to myelopoietic supportivestroma
Cell type
Fat
Bone
Cartilage Supportive stroma
ALP control ALP inducedAlizarin red
Marrow-derived stromal cell linesMBA-13 �� �� � ���� �� �MBA-15 � �� � ����� � �14F1.1 �� � � � � ���
MD-MSCsA � � � � � �B ��� � � � ��� �
MSCs ����� �� �� ����� ��� ��C3H10T1/2 ����� �� � � � �MEFs
1E �� � � � �� �1L � �� � ��� ��� �2 � � � � � �3E � � � � � �3L �� �� � � � �4 ����� �� � ���� � ���5 �� � �� � � �6 ��� �� �� � � �7 �� �� � � � ���
Scoring key: Fat, Oil red O OD/�g protein: 0–0.5, �; 0.5–1, ��; 1–2, �; 2–3, ��; 3–4, ���; 4–5, ����; � 5, �����. Bone,Alkaline phosphatase (ALP): ��, faint staining; �, regular staining; ��, strong staining. Alizarin red OD/�g protein: 0–0.4, �; 0.4–1, �;1–1.5, ��; 1.5–2, ���; 2–2.5, ����; � 5, �����. Cartilage, Alcian Blue: �, no staining; ��, faint staining; �, regular staining;���, strong staining. Supportive stroma, total colonies (granulocyte-macrophage colony-forming unit) and cell counts: �, � 50 colonies,2 � 105 cells; �, 50–150 colonies, 2–4 � 105 cells; ��, 150–200 colonies, 4–6 � 105 cells; ���, � 200 colonies, � 6 � 105 cells.Abbreviations: MD-MSCs, multipotent adult progenitor cell-derived mesenchymal stem cells; MEFs, mouse embryo fibroblasts; MSCs,mesenchymal stem cells.
2280 MSC Hemopoietic Support Versus Multipotency

The Differentiation of MSCs into Osteogenic LineageDoes Not Hamper Myelopoietic Supportive Capacity,Whereas Adipogenesis Suppresses This FunctionThe stem cell niche in vivo has been shown to include osteo-blasts [25, 34]. We therefore next examined whether the capac-ity of mesenchymal cells to support myelopoiesis would beaffected following induction of differentiation into mesodermallineages. MSCs were grown under osteogenic or adipogenicdifferentiation conditions for 10 days and were then examinedfor their myelopoietic supportive activity. Although osteogenicdifferentiation led to a 44.79% reduction in the hemopoietic cellyield, the incidence of myeloid progenitors recovered remainedunchanged (Fig. 3A, 3B). In contrast adipogenic differentiationreduced both the hemopoietic cell yield and the generation ofmyeloid progenitors by 85.41% and 83.09%, respectively (Fig.3A, 3B). A similar experiment was then performed using the14F1.1 preadipocyte cell clone. These cells are biased towardadipogenesis, as shown above. Upon induction of fat accumu-lation this stromal clone allowed the production of hemopoietic
cells, however, as in the case of adipogenic MSCs, the yield ofmyeloid progenitors was markedly reduced (Fig. 3C, 3D). Fur-ther analysis showed that upon adipogenic differentiation MSCslost their capacity to support myelopoiesis. This was evidencedby complete inhibition of total cells (Fig. 3E), myeloid progen-itors (Fig. 3F), and cobblestone area-forming cells (Fig. 3G). Bycontrast, the control undifferentiated MSCs sustained myeloidprogenitors and supported cobblestone area formation. Flowcytometry analysis of cells maintained in the control MSCsrevealed that most of the cells expressed myeloid markers(91.1% CD11b), whereas a minor fraction was c-Kit� (1.25%)and Sca-1� (1.19%) (Fig. 3H). These findings conform with arecently reported study [35]. Thus, whereas osteogenesis doesnot affect the yield of myeloid progenitors, adipogenesis causesdrastic inhibition of progenitor cell accumulation in long-termbone marrow cultures supported by all mesenchymal cells testedincluding MEFs, preadipogenic cell clone, and MSCs.
The induction of differentiation of MSCs is associated withchanged patterns in gene expression that may account for the
Figure 3. Comparison of the ability of mesenchymal stem cells (MSCs) and 14F1.1 to support hemopoiesis after differentiation. MSCs were grownin normal (Cont), adipoinductive (Adipo), or osteoinductive (Osteo) medium for 10 days and subjected to long-term bone marrow culture (LTBMC)conditions for 4 weeks (A, B) or 2 weeks (E). At the end of this period the cultures were evaluated for total hemopoietic cell counts (A, E), numberof myeloid progenitors (B, F), number of cobblestone-forming units (G), and the distribution of the hemopoietic cell types as measured by flowcytometry (H). 14F1.1 cells were grown in normal (Cont) or adipoinductive (Adipo) medium for 2 weeks and subjected to LTBMC conditions for4 weeks. Hemopoietic cell counts (C) and myeloid colonies (D) were determined. (A) �, p � .017 and .00017 for Control-Osteo and Control-Adipo,respectively; (B) �, p � .003; (D) �, p � .004.
2281Morad, Pevsner-Fischer, Barnees et al.
www.StemCells.com

reduced support of myelopoiesis after adipogenic differentia-tion. A group of cytokine genes was therefore selected based ontheir known contribution to hemopoiesis. IL-6 and granulocyte-macrophage colony-stimulating factor (GM-CSF) expression, atthe mRNA level, were reduced following adipogenic differen-tiation of both MSCs and 14F1.1 cells. Conversely, fms-liketyrosine kinase 3 ligand and SCF mRNA remained unchanged(Fig. 4). Interestingly, within the tested gene group, osteogen-esis did not significantly affect any of the hemopoietic cyto-kines, a finding that corresponds well to the limited effect ofosteogenesis on myelopoietic supportive activity (Fig. 3A, 3B).Whereas granulocyte colony-stimulating factor (G-CSF) expres-sion has been reported for human [36] and rat [37] osteoblastswe did not detect any G-CSF mRNA in the mouse cells studiedhere. We have previously reported that our stromal cells lines,which are derived without the use of strong selective pressuresand without the use of proteolytic enzymes, do not expressseveral cytokine genes including G-CSF [38].
Table 1 summarizes the potential of the various cells studiedto differentiate compared with their ability, in the uninduced,nondifferentiated state, to support myelopoiesis. As can be seen,the mere potential of the cells to differentiate into a particulardirection did not predict their myelopoietic supportive capacity.In contrast the actual differentiation into specific mesodermalpathways seems to determine the ability of the cells to maintainactive myelopoiesis. We have previously reported the wide-spread and constitutive expression of ALP among various stro-mal cell lines from the bone marrow [13]. Here we also detected
ALP expression in uninduced cells. However, upon osteogene-sis induction, an increase of ALP expression in induced sampleswas observed followed by calcium mineralization detected byalizarin red staining.
Differentiated Progeny of MSCs Differ inGlycosylation Patterns That Impact MyelopoieticSupportive CapacityIt has been suggested that epigenetic factors play a role indifferent biological processes [39] including the state of stemcells [40]. We considered the possibility that the glycosylationpatterns of cells, following their lineage-specific differentiation,would change and could contribute to the differences in hemo-poietic support. Glycoanalysis of protein membrane extractsfrom MSCs, prior to and following induction of adipogenic andosteogenic differentiation, was performed using lectin microar-rays. When comparing the binding patterns of the extracts fromundifferentiated MSC population with those of the differentiatedcells, significant differences in signals were observed by a groupof lectins that recognize complex N-linked glycans (Fig. 5A,5B). These lectins recognize branching at either of the two�-mannose residues of the trimannosyl core of N-linked com-plex glycans, and indicate the presence of either triantennary ortetra-antennary structures. Figure 5A depicts the binding ofmesenchymal cell extracts to these lectins and demonstrates thatthe level of antennarity in the osteogenic MSCs is higher thanthat of the undifferentiated MSC population. Figure 5B showsthat following induction of adipogenic differentiation the levelof antennarity of complex glycans in the cells is lower than thatof the undifferentiated MSC population. A similar decrease inthe level of antennarity of complex glycans is shown to accom-pany differentiation of NIH-3T3L1 fibroblasts into adipocytes(Fig. 5C) [41].
One implication of the above study is that glycosylationpatterns may predict the ability of a given mesenchymal popu-lation to support hemopoiesis. Experiments that we performedwith some of the cell types reported here did not support thispossibility (results not shown). This may result from the mask-ing of the glycosylation pattern, relevant for hemopoiesis, bycell type-specific glycosylations related to other cellular func-tions. In addition, some mesenchymal cell types interfere withhemopoiesis by elaborating inhibitory cytokines, a feature un-related to their respective glycosylation patterns.
Adipogenesis is thus associated both with a reduction inthe antennarity of complex N-glycans as well as with adecrease in the capacity of adipogenic MSCs to supportmyelopoiesis. To test the possible contribution of stromaglycosylation to the support of hemopoiesis we chose tointerfere with glycosylation in MSCs and examine the effectson hemopoietic supportive capacity. The initial step in N-glycan synthesis involves synthesis of a precursor oligosac-charide, which is then stepwise processed by several en-zymes, including mannosidase I, to allow synthesis ofcomplex N-linked glycans. DMJ, a known inhibitor of man-nosidase I, blocks the enzyme and therefore inhibits conver-sion of high mannose to complex chains. As a result, treat-ment with DMJ leads to synthesis of glycoproteins withincreased levels of high-mannose glycans and less complexN-linked glycans. MSCs were treated with DMJ (0.4 mg/ml)for 3 days and then analyzed on the lectin arrays. Thistreatment caused a decrease in antennarity (Fig. 5D) that wasalso accompanied by decrease in sugars that are found onantennae termini such as beta galactose and sialic acid (Fig.5E). Moreover, signals from all oligomannose-binding lectinswere significantly increased, suggesting the glycoproteinscontain increased amounts of high-mannose glycans in com-
Figure 4. Cytokine and growth factors expression in mesenchymalstem cells (MSCs) and 14F1.1 cells after differentiation and transfer tolong-term bone marrow culture (LTBMC) conditions. Semiquantitativereverse-transcription–polymerase chain reaction analysis of the expres-sion of stem cell factor (SCF), fms-like tyrosine kinase 3 ligand (Flt3-L),macrophage colony-stimulating factor (M-CSF), interleukin (IL)-6,granulocyte-macrophage CSF (GM-CSF), leukemia inhibitory factor(LIF), IL-3, and granulocyte CSF (G-CSF) in differentiated samples ofMSCs and 14F1.1 cells after 4 weeks of LTBMC. Abbreviations: Adipo,adipoinductive; Cont, normal; GAPDH, glyceraldehyde-3-phosphatedehydrogenase; Osteo, osteoinductive.
2282 MSC Hemopoietic Support Versus Multipotency
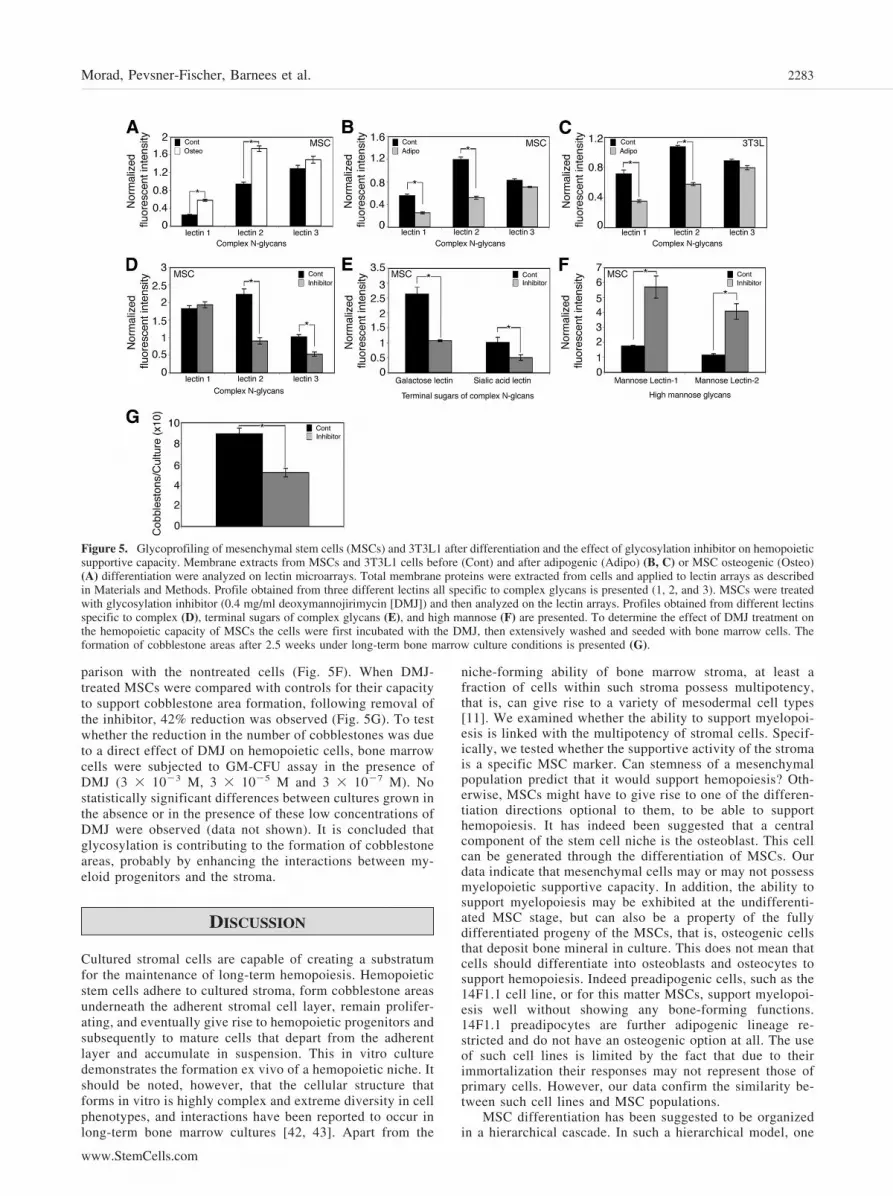
parison with the nontreated cells (Fig. 5F). When DMJ-treated MSCs were compared with controls for their capacityto support cobblestone area formation, following removal ofthe inhibitor, 42% reduction was observed (Fig. 5G). To testwhether the reduction in the number of cobblestones was dueto a direct effect of DMJ on hemopoietic cells, bone marrowcells were subjected to GM-CFU assay in the presence ofDMJ (3 � 10�3 M, 3 � 10�5 M and 3 � 10�7 M). Nostatistically significant differences between cultures grown inthe absence or in the presence of these low concentrations ofDMJ were observed (data not shown). It is concluded thatglycosylation is contributing to the formation of cobblestoneareas, probably by enhancing the interactions between my-eloid progenitors and the stroma.
DISCUSSION
Cultured stromal cells are capable of creating a substratumfor the maintenance of long-term hemopoiesis. Hemopoieticstem cells adhere to cultured stroma, form cobblestone areasunderneath the adherent stromal cell layer, remain prolifer-ating, and eventually give rise to hemopoietic progenitors andsubsequently to mature cells that depart from the adherentlayer and accumulate in suspension. This in vitro culturedemonstrates the formation ex vivo of a hemopoietic niche. Itshould be noted, however, that the cellular structure thatforms in vitro is highly complex and extreme diversity in cellphenotypes, and interactions have been reported to occur inlong-term bone marrow cultures [42, 43]. Apart from the
niche-forming ability of bone marrow stroma, at least afraction of cells within such stroma possess multipotency,that is, can give rise to a variety of mesodermal cell types[11]. We examined whether the ability to support myelopoi-esis is linked with the multipotency of stromal cells. Specif-ically, we tested whether the supportive activity of the stromais a specific MSC marker. Can stemness of a mesenchymalpopulation predict that it would support hemopoiesis? Oth-erwise, MSCs might have to give rise to one of the differen-tiation directions optional to them, to be able to supporthemopoiesis. It has indeed been suggested that a centralcomponent of the stem cell niche is the osteoblast. This cellcan be generated through the differentiation of MSCs. Ourdata indicate that mesenchymal cells may or may not possessmyelopoietic supportive capacity. In addition, the ability tosupport myelopoiesis may be exhibited at the undifferenti-ated MSC stage, but can also be a property of the fullydifferentiated progeny of the MSCs, that is, osteogenic cellsthat deposit bone mineral in culture. This does not mean thatcells should differentiate into osteoblasts and osteocytes tosupport hemopoiesis. Indeed preadipogenic cells, such as the14F1.1 cell line, or for this matter MSCs, support myelopoi-esis well without showing any bone-forming functions.14F1.1 preadipocytes are further adipogenic lineage re-stricted and do not have an osteogenic option at all. The useof such cell lines is limited by the fact that due to theirimmortalization their responses may not represent those ofprimary cells. However, our data confirm the similarity be-tween such cell lines and MSC populations.
MSC differentiation has been suggested to be organizedin a hierarchical cascade. In such a hierarchical model, one
Figure 5. Glycoprofiling of mesenchymal stem cells (MSCs) and 3T3L1 after differentiation and the effect of glycosylation inhibitor on hemopoieticsupportive capacity. Membrane extracts from MSCs and 3T3L1 cells before (Cont) and after adipogenic (Adipo) (B, C) or MSC osteogenic (Osteo)(A) differentiation were analyzed on lectin microarrays. Total membrane proteins were extracted from cells and applied to lectin arrays as describedin Materials and Methods. Profile obtained from three different lectins all specific to complex glycans is presented (1, 2, and 3). MSCs were treatedwith glycosylation inhibitor (0.4 mg/ml deoxymannojirimycin [DMJ]) and then analyzed on the lectin arrays. Profiles obtained from different lectinsspecific to complex (D), terminal sugars of complex glycans (E), and high mannose (F) are presented. To determine the effect of DMJ treatment onthe hemopoietic capacity of MSCs the cells were first incubated with the DMJ, then extensively washed and seeded with bone marrow cells. Theformation of cobblestone areas after 2.5 weeks under long-term bone marrow culture conditions is presented (G).
2283Morad, Pevsner-Fischer, Barnees et al.
www.StemCells.com

would expect the cells to acquire the hemopoietic supportivecapacity upon induction of differentiation and loss of stem-ness. Yet, it appears that the hemopoietic support property iseither associated with the stem cell itself, or alternatively,with a differentiated progeny such as the preadipocyte or theosteocyte, to mention two examples. The property seemstherefore to exist throughout the differentiation cascaderather than emerge at any particular stage. It is therefore notthe case that MSCs differentiate into hemopoietic supportivestroma. Either they may serve this function, as they are (i.e.,while maintaining their undifferentiated stemness), or theymay also fully differentiate and still maintain this function.This latter event occurs during osteogenesis, whereas adipo-genesis seems to interfere, at least partially, with the capacityto support myelopoiesis. Whether this last finding is relatedto the reduced hemopoiesis in adult and old, fat-laden humanbone marrow should be further investigated.
CONCLUSION
It is concluded that the differentiation of MSCs and acquisitionof mature phenotype, in terms of the capacity to support my-
elopoiesis, do not comply with a hierarchical cascade. Ourresults are more compatible with a phase-space model [44] suchas the hypothetical one shown in Figure 6. This model isproposed as a possible interpretation of the results presented inthis study and is also based on our previous observations relatedto the plastic nature of mesenchymal cells [45]. It indicates thatseveral directions of differentiation of the MSC may occur bydirect derivation from the MSC itself, rather then from descen-dents of the MSC that progress through intermediate differen-tiation stages. Furthermore, the model proposes possible re-versibility of differentiated phenotypes. Our past studies onthe derivation of stromal cell lines showed that the MBA-14cell line, which constituted a mixture of fibroblast-like cellsand macrophages, could be separated into these two distinctpopulations, 14F and 14M. The former became strongly fatladen upon separation from the latter [12]. The preadipogeniccell supports hemopoiesis [38] and performs better when inthe preadipogenic rather than in the highly adipose-ladenstate. Upon readdition of the 14M macrophages to the 14Ffat-laden cells, the latter regained the fibroblast appearance[12]. These studies, which indicate possible dedifferentiationof adipogenic cells, are supported by a recent publicationshowing that adipose tissue cells can convert into fibroblastic
Figure 6. Mesenchymal cell differentiationis compatible with a phase-space model.Several directions of differentiation of themesenchymal stem cell (MSC) may occur bydirect derivation from the MSC, itself, ratherthan from descendents of the MSC thatprogress through intermediate differentiationstages. The figure demonstrates that the abil-ity to support hemopoiesis (“stromal cell”property) is shared by a cell biased to adi-pogenesis (14F1.1) and the MSC, itself,which is shown herein to have at list fourpossible differentiation options. Thus, thecapacity to support hemopoiesis is notemerging as a late occurrence in a differen-tiation hierarchy. In addition, this propertymay reversible. Mouse embryo fibroblasts(MEFs) are shown together with MSCs seenfrom the viewpoint of differentiation poten-tial they are shown herein to be similar.Abbreviations: HSC, hemopoietic stem cell;MD-MSC, multipotent adult progenitor cell-derived MSC.
2284 MSC Hemopoietic Support Versus Multipotency

phenotype and, upon this conversion, gain multipotent dif-ferentiation features [46].
One more issue raised by our experiments is that the my-elopoietic supportive function of mesenchymal cell populationsmay be gained by populations that do not possess this functiona priori. Whether this is due to the selection of rare clones,which take over during culture, or to a generalized phenotypicchange, is at the moment unclear. This contention requiresfurther elucidation using single-clone analysis.
The finding that osteogenic differentiation does not interferewith the capacity of MSCs to support myelopoiesis goes alongwith the fact that osteoblasts contribute to the stem cell niche invivo. By contrast adipogenic differentiation of human mesen-chymal cells has been shown to reduce the capacity of thesehuman cells to support the proliferation of cord bloodCD34�CD38� progenitors [35]. Similarly, we show here thatadipocyte differentiation of mouse MSCs results in reducedmaintenance of myeloid progenitors. The mechanism seems toinvolve reduction in expression of IL-6 and GM-CSF. An ad-ditional occurrence associated specifically with adipocyte dif-ferentiation was a change in glycosylation pattern. We havepreviously shown that free saccharides interfere with the inter-actions of hemopoietic progenitor cells and the stroma. Subse-quent studies with bound sugar moieties (neoglycoproteins)substantiated these findings [47, 48]. It is therefore implied thatglycoproteins contribute to the interactions of stromal cells andhemopoietic progenitors and to the maintenance of the hemo-poiesis in long-term culture. It is thus plausible that the modifiedglycosylation pattern that we observed following adipogenesisis associated with the change in myelopoietic support. Indeed, a
use of an inhibitor of glycosylation (DMJ) led to reducedcapacity of treated MSCs to support the formation of cobble-stone areas in cocultures with bone marrow cells. Due to thereversible nature of the DMJ-mediated inhibition [49, 50], it islikely that the effects we observed have to do with the initialimmediate interaction, probably the adhesion, of hemopoieticprogenitor cell with the stroma. Glycosylated moieties present instromal cells seem therefore to contribute to the interactionsbetween the stroma and hemopoietic progenitor cells. Suchinteractions may be of importance for the formation of hemo-poietic stem cell niches in vivo.
ACKNOWLEDGMENTS
D.Z. is an incumbent of the Joe and Celia Weinstein ProfessorialChair at the Weizmann Institute of Science. The authors areindebted to Mrs. Varda Segal for her excellent technical assis-tance. This study was supported by The Helen and MartinKimmel Institute for Stem Cell Research at the WeizmannInstitute of Science, The Gabrielle Rich Center for Transplan-tation Biology, The Minerva Foundation, Germany, and TheLegacy Heritage Foundation.
DISCLOSURE OF POTENTIAL CONFLICTS
OF INTEREST
The authors indicate no potential conflicts of interest.
REFERENCES
1 Friedenstein AJ, Chailakhyan RK, Latsinik NV et al. Stromal cellsresponsible for transferring the microenvironment of the hemopoietictissues. Cloning in vitro and retransplantation in vivo. Transplantation1974;17:331–340.
2 Dexter TM, Wright EG, Krizsa F et al. Regulation of haemopoietic stemcell proliferation in long term bone marrow cultures. Biomedicine 1977;27:344–349.
3 Zipori D. Cell interactions in the bone marrow microenvironment: role ofendogenous colony-stimulating activity. J Supramol Struct Cell Biochem1981;17:347–357.
4 Noort WA, Kruisselbrink AB, in’t Anker PS et al. Mesenchymal stemcells promote engraftment of human umbilical cord blood-derivedCD34(�) cells in NOD/SCID mice. Exp Hematol 2002;30:870–878.
5 Prockop DJ. Marrow stromal cells as stem cells for nonhematopoietictissues. Science 1997;276:71–74.
6 Hung SC, Pochampally RR, Hsu SC et al. Short-term exposure ofmultipotent stromal cells to low oxygen increases their expression ofCX3CR1 and CXCR4 and their engraftment in vivo. Plos One 2007;2:e416:1–11.
7 Caplan AI, Bruder SP. Mesenchymal stem cells: Building blocks formolecular medicine in the 21st century. Trends Mol Med 2001;7:259 –264.
8 Barry FP, Murphy JM. Mesenchymal stem cells: Clinical applicationsand biological characterization. Int J Biochem Cell Biol 2004;36:568 –584.
9 Javazon EH, Beggs KJ, Flake AW. Mesenchymal stem cells: Paradoxesof passaging. Exp Hematol 2004;32:414–425.
10 Bruder SP, Fink DJ, Caplan AI. Mesenchymal stem cells in bone devel-opment, bone repair, and skeletal regeneration therapy. J Cell Biochem1994;56:283–294.
11 Pittenger MF, Mackay AM, Beck SC et al. Multilineage potential ofadult human mesenchymal stem cells. Science 1999;284:143–147.
12 Zipori D, Friedman A, Tamir M et al. Cultured mouse marrow cell lines:Interactions between fibroblastoid cells and monocytes. J Cell Physiol1984;118:143–152.
13 Zipori D, Duksin D, Tamir M et al. Cultured mouse marrow stromal celllines. II. Distinct subtypes differing in morphology, collagen types,myelopoietic factors, and leukemic cell growth modulating activities.J Cell Physiol 1985;122:81–90.
14 Benayahu D, Kletter Y, Zipori D et al. Bone marrow-derived stromal cellline expressing osteoblastic phenotype in vitro and osteogenic capacity invivo. J Cell Physiol 1989;140:1–7.
15 Jiang Y, Jahagirdar BN, Reinhardt RL et al. Pluripotency of mesenchy-mal stem cells derived from adult marrow. Nature 2002;418:41–49.
16 Serafini M, Dylla SJ, Oki M et al. Hematopoietic reconstitution bymultipotent adult progenitor cells: precursors to long-term hematopoieticstem cells. J Exp Med 2007;204:129–139.
17 Kogler G, Sensken S, Airey JA et al. A new human somatic stem cellfrom placental cord blood with intrinsic pluripotent differentiation po-tential. J Exp Med 2004;200:123–135.
18 D’Ippolito G, Diabira S, Howard GA et al. Marrow-isolated adult mul-tilineage inducible (MIAMI) cells, a unique population of postnatalyoung and old human cells with extensive expansion and differentiationpotential. J Cell Sci 2004;117:2971–2981.
19 De Coppi P, Bartsch G, Jr., Siddiqui MM et al. Isolation of amniotic stemcell lines with potential for therapy. Nat Biotechnol 2007;25:100–106.
20 Toma JG, Akhavan M, Fernandes KJ et al. Isolation of multipotent adultstem cells from the dermis of mammalian skin. Nat Cell Biol 2001;3:778–784.
21 Kucia M, Wysoczynski M, Ratajczak J et al. Identification of very smallembryonic like (VSEL) stem cells in bone marrow. Cell Tissue Res2008;331:125–134.
22 Anjos-Afonso F, Bonnet D. Nonhematopoietic/endothelial SSEA-1�cells define the most primitive progenitors in the adult murine bonemarrow mesenchymal compartment. Blood 2007;109:1298–1306.
23 Muraglia A, Cancedda R, Quarto R. Clonal mesenchymal progenitorsfrom human bone marrow differentiate in vitro according to a hierarchi-cal model. J Cell Sci 2000;113(pt 7):1161–1166.
24 Zipori D. The stem state: Plasticity is essential, whereas self-renewal andhierarchy are optional. STEM CELLS 2005;23:719–726.
25 Zhang J, Niu C, Ye L et al. Identification of the haematopoietic stem cellniche and control of the niche size. Nature 2003;425:836–841.
26 Scadden DT. The stem-cell niche as an entity of action. Nature 2006;441:1075–1079.
27 Taichman RS. Blood and bone: Two tissues whose fates are intertwinedto create the hematopoietic stem-cell niche. Blood 2005;105:2631–2639.
28 Zipori D, Sasson T. Myelopoiesis in the presence of stromal cells frommouse bone marrow: II. Mechanism of glucose dependent colony for-mation. Exp Hematol 1981;9:663–674.
29 Zipori D, Sasson T, Frenkel A. Myelopoiesis in the presence of stromalcells from mouse bone marrow: I. Monosaccharides regulate colonyformation. Exp Hematol 1981;9:656–663.
2285Morad, Pevsner-Fischer, Barnees et al.
www.StemCells.com

30 Jiang Y, Vaessen B, Lenvik T et al. Multipotent progenitor cells can beisolated from postnatal murine bone marrow, muscle, and brain. ExpHematol 2002;30:896–904.
31 Pevsner-Fischer M, Morad V, Cohen-Sfady M et al. Toll-like receptorsand their ligands control mesenchymal stem cell functions. Blood. 2007;109:1422–1432.
32 Marazzi A. Algonithms, Routines and S Functions of Robust Statistics.Belmont, CA: Wadsworth and Brooks/Cole, 1993.
33 Benayahu D, Horowitz M, Zipori D et al. Hemopoietic functions ofmarrow-derived osteogenic cells. Calcif Tissue Int 1992;51:195–201.
34 Calvi LM, Adams GB, Weibrecht KW et al. Osteoblastic cells regulatethe haematopoietic stem cell niche. Nature 2003;425:841–846.
35 Ookura N, Fujimori Y, Nishioka K et al. Adipocyte differentiation ofhuman marrow mesenchymal stem cells reduces the supporting ca-pacity for hematopoietic progenitors but not for severe combinedimmunodeficiency repopulating cells. Int J Mol Med 2007;19:387–392.
36 Taichman RS, Emerson SG. Human osteoblasts support hematopoiesisthrough the production of granulocyte colony-stimulating factor. J ExpMed 1994;179:1677–1682.
37 Felix R, Cecchini MG, Hofstetter W et al. Production of granulocyte-macrophage (GM-CSF) and granulocyte colony-stimulating factor (G-CSF) by rat clonal osteoblastic cell population CRP 10/30 and theimmortalized cell line IRC10/30-myc1 stimulated by tumor necrosisfactor alpha. Endocrinology 1991;128:661–667.
38 Zipori D, Lee F. Introduction of interleukin-3 gene into stromal cellsfrom the bone marrow alters hemopoietic differentiation but does notmodify stem cell renewal. Blood 1988;71:586–596.
39 Feinberg AP. Phenotypic plasticity and the epigenetics of human disease.Nature 2007;447:433–440.
40 Zipori D. The nature of stem cells: State rather than entity. Nat RevGenet 2004;5:873–878.
41 Ben-Yakar Maya R, Amor Y, Rosenberg R et al. Glycoanalysis on alectin array: Applications to the development of biopharmaceuticals andlife science research. In: Sansom C, Markman O, eds. Glycobiology. 1sted. Oxfordshire, U.K.: Scion Publishing Ltd, 2007:340–351.
42 Allen TD, Dexter TM. Ultrastructural aspects of erythropoietic dif-ferentiation in long-term bone marrow culture. Differentiation 1982;21:86 –94.
43 Allen TD, Dexter TM. Long term bone marrow cultures: An ultrastruc-tural review. Scan Electron Microsc 1983;(pt 4):1851–1866.
44 Kirkland MA. A phase space model of hemopoiesis and the concept ofstem cell renewal. Exp Hematol 2004;32:511–519.
45 Sternberg D, Peled A, Shezen E et al. Control of stroma-dependenthematopoiesis by basic fibroblast growth factor: stromal phenotypicplasticity and modified myelopoietic functions. Cytokines Mol Ther1996;2:29–38.
46 Matsumoto T, Kano K, Kondo D et al. Mature adipocyte-derived dedi-fferentiated fat cells exhibit multilineage potential. J Cell Physiol 2008;215:210–222.
47 Aizawa S, Tavassoli M. In vitro homing of hemopoietic stem cells ismediated by a recognition system with galactosyl and mannosyl speci-ficities. Proc Natl Acad Sci U S A 1987;84:4485–4489.
48 Aizawa S, Tavassoli M. Interaction of murine granulocyte-macrophageprogenitors and supporting stroma involves a recognition mechanismwith galactosyl and mannosyl specificities J Clin Invest 1987;80:1698–1705.
49 Takahashi H, Parsons PG. Rapid and reversible inhibition of tyrosinaseactivity by glucosidase inhibitors in human melanoma cells. J InvestDermatol 1992;98:481–487.
50 Ogier-Denis E, Trugnan G, Sapin C et al. Dual effect of 1-deoxy-mannojirimycin on the mannose uptake and on the N-glycan process-ing of the human colon cancer cell line HT-29. J Biol Chem 1990;265:5366 –5369.
2286 MSC Hemopoietic Support Versus Multipotency




















