
The molecular classification of the clinical manifestations of Crohn's disease
13
RAPID COMMUNICATIONS The Molecular Classification of the Clinical Manifestations of Crohn’s Disease TARIQ AHMAD,* ALESSANDRO ARMUZZI,* MIKE BUNCE, ‡ KIM MULCAHY–HAWES,* SARA E. MARSHALL, § TIMOTHY R. ORCHARD,* JONATHAN CRAWSHAW,* OLIVER LARGE,* ARJUNA DE SILVA,* JULIA T. COOK, ‡ MARTIN BARNARDO, ‡ SUE CULLEN,* KEN I. WELSH, and DEREK P. JEWELL* *Gastroenterology Unit, University of Oxford, Gibson Laboratories, Radcliffe Infirmary, Oxford; ‡ Department of Transplant Immunology, Oxford Transplant Centre, University of Oxford, Churchill Hospital, Oxford; § Department of Immunology, Wright-Fleming Institute, Imperial College, London; and Clinical Genomics, National Heart and Lung Institute, Imperial College, London, United Kingdom See editorial on page 1161. Background & Aims: Crohn’s disease is a common in- flammatory disorder of the gut characterized by varia- tion in both location and behavior. Chromosome 16 and the HLA region on chromosome 6 have been implicated in susceptibility to disease. Mutations in the NOD2/ CARD15 gene, recently identified on chromosome 16, have been associated with disease overall but are found in only 25% of patients. No data regarding their contri- bution to specific disease subtypes exist. Here we report a detailed genotype-phenotype analysis of 244 accu- rately characterized patients. Methods: A total of 244 white patients with Crohn’s disease recruited from a single center in the United Kingdom were studied. All patients were rigorously phenotyped and followed-up for a median time of 16 years. By using linkage disequilib- rium mapping we studied 340 polymorphisms in 24 HLA genes and 3 NOD2/CARD15 polymorphisms. Results: We show that NOD2/CARD15 mutations determine ileal disease only. We confirm that alleles on specific long- range HLA haplotypes determine overall susceptibility and describe novel genetic associations with suscep- tibility, location, and behavior of Crohn’s disease. Conclusions: The clinical pattern of Crohn’s disease may be defined by specific genotypes. This study may provide the basis for a future molecular classification of disease. C rohn’s disease is a chronic inflammatory disorder of the gastrointestinal tract. Extensive heterogeneity is observed in terms of disease presentation, behavior, and response to treatment. Attempts have been made to define clinical subgroups on the basis of disease location, extent (diffuse or localized), behavior (primary inflam- matory, fistulating, or fibrostenotic), and by surgical history. 1,2 Epidemiologic and genetic data suggest that this disease heterogeneity may be genetically deter- mined: Studies in multiply affected families have shown that affected siblings frequently present at a similar age and concordance rates of up to 80% have been reported for disease site, behavior, and presence of extraintestinal manifestations of disease. 3–6 In addition, case-control association studies have reported genetic associations with the presence of extraintestinal manifestations 7 and perianal fistulating disease. 8 The major focus of recent genetic research in inflam- matory bowel disease (IBD) has been the identification of disease-susceptibility genes. Little emphasis has been placed on the investigation of genes that determine phenotype because of the difficulties of accumulating a large cohort of patients whose disease phenotype and natural history has been accurately determined. Genome- wide scans have identified susceptibility loci on chromo- somes 1, 9 5, 10,11 6, 10,12 12, 13 14, 14,15 16, 13,16 and 19. 10 IBD1, 16 the first and most consistently replicated 17 IBD susceptibility locus identified in the pericentromeric re- gion of chromosome 16, is exclusive to Crohn’s disease. Three independent groups have recently reported associ- ations between the NOD2/CARD15 gene, 18 –20 located within this locus, and susceptibility to Crohn’s disease. NOD2/CARD15, a NOD1/apoptotic protease-activat- Abbreviations used in this paper: Apaf-1, apoptotic protease activat- ing factor-1; BAT1, HLA-B-associated transcript 1; CI, confidence inter- val; HLA, human leucocyte antigen; MICA, major histocompatibility complex class I chain-related gene A; MICB, major histocompatibility complex class I chain-related gene B; NFB, nuclear factor B; PAR, population attributable risk; RR, relative risk; SNP, single nucleotide polymorphism; TNF, tumor necrosis factor. © 2002 by the American Gastroenterological Association 0016-5085/02/$35.00 doi:10.1053/gast.2002.32413 GASTROENTEROLOGY 2002;122:854 – 866
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of The molecular classification of the clinical manifestations of Crohn's disease

RAPID COMMUNICATIONS
The Molecular Classification of the Clinical Manifestationsof Crohn’s Disease
TARIQ AHMAD,* ALESSANDRO ARMUZZI,* MIKE BUNCE,‡ KIM MULCAHY–HAWES,*SARA E. MARSHALL,§ TIMOTHY R. ORCHARD,* JONATHAN CRAWSHAW,* OLIVER LARGE,*ARJUNA DE SILVA,* JULIA T. COOK,‡ MARTIN BARNARDO,‡ SUE CULLEN,* KEN I. WELSH,�
and DEREK P. JEWELL**Gastroenterology Unit, University of Oxford, Gibson Laboratories, Radcliffe Infirmary, Oxford; ‡Department of Transplant Immunology, OxfordTransplant Centre, University of Oxford, Churchill Hospital, Oxford; §Department of Immunology, Wright-Fleming Institute, Imperial College,London; and �Clinical Genomics, National Heart and Lung Institute, Imperial College, London, United Kingdom
See editorial on page 1161.
Background & Aims: Crohn’s disease is a common in-flammatory disorder of the gut characterized by varia-tion in both location and behavior. Chromosome 16 andthe HLA region on chromosome 6 have been implicatedin susceptibility to disease. Mutations in the NOD2/CARD15 gene, recently identified on chromosome 16,have been associated with disease overall but are foundin only 25% of patients. No data regarding their contri-bution to specific disease subtypes exist. Here we reporta detailed genotype-phenotype analysis of 244 accu-rately characterized patients. Methods: A total of 244white patients with Crohn’s disease recruited from asingle center in the United Kingdom were studied. Allpatients were rigorously phenotyped and followed-up fora median time of 16 years. By using linkage disequilib-rium mapping we studied 340 polymorphisms in 24 HLAgenes and 3 NOD2/CARD15 polymorphisms. Results:We show that NOD2/CARD15 mutations determine ilealdisease only. We confirm that alleles on specific long-range HLA haplotypes determine overall susceptibilityand describe novel genetic associations with suscep-tibility, location, and behavior of Crohn’s disease.Conclusions: The clinical pattern of Crohn’s disease maybe defined by specific genotypes. This study may providethe basis for a future molecular classification of disease.
Crohn’s disease is a chronic inflammatory disorder ofthe gastrointestinal tract. Extensive heterogeneity is
observed in terms of disease presentation, behavior, andresponse to treatment. Attempts have been made todefine clinical subgroups on the basis of disease location,extent (diffuse or localized), behavior (primary inflam-matory, fistulating, or fibrostenotic), and by surgical
history.1,2 Epidemiologic and genetic data suggest thatthis disease heterogeneity may be genetically deter-mined: Studies in multiply affected families have shownthat affected siblings frequently present at a similar ageand concordance rates of up to 80% have been reportedfor disease site, behavior, and presence of extraintestinalmanifestations of disease.3–6 In addition, case-controlassociation studies have reported genetic associationswith the presence of extraintestinal manifestations7 andperianal fistulating disease.8
The major focus of recent genetic research in inflam-matory bowel disease (IBD) has been the identification ofdisease-susceptibility genes. Little emphasis has beenplaced on the investigation of genes that determinephenotype because of the difficulties of accumulating alarge cohort of patients whose disease phenotype andnatural history has been accurately determined. Genome-wide scans have identified susceptibility loci on chromo-somes 1,9 5,10,11 6,10,12 12,13 14,14,15 16,13,16 and 19.10
IBD1,16 the first and most consistently replicated17 IBDsusceptibility locus identified in the pericentromeric re-gion of chromosome 16, is exclusive to Crohn’s disease.Three independent groups have recently reported associ-ations between the NOD2/CARD15 gene,18–20 locatedwithin this locus, and susceptibility to Crohn’s disease.NOD2/CARD15, a NOD1/apoptotic protease-activat-
Abbreviations used in this paper: Apaf-1, apoptotic protease activat-ing factor-1; BAT1, HLA-B-associated transcript 1; CI, confidence inter-val; HLA, human leucocyte antigen; MICA, major histocompatibilitycomplex class I chain-related gene A; MICB, major histocompatibilitycomplex class I chain-related gene B; NF�B, nuclear factor �B; PAR,population attributable risk; RR, relative risk; SNP, single nucleotidepolymorphism; TNF, tumor necrosis factor.
© 2002 by the American Gastroenterological Association0016-5085/02/$35.00
doi:10.1053/gast.2002.32413
GASTROENTEROLOGY 2002;122:854–866

ing factor-1 (Apaf-1) family member, is expressed inmonocytes where it is thought to function as a cytosolicreceptor for bacterial lipopolysaccharide.21 Bacteriallipopolysaccharide binds to a leucine-rich repeat re-gion in the NOD2/CARD15 protein, triggering theactivation of nuclear factor–�B (NF-�B). Two nonsyn-onymous, single nucleotide polymorphisms (SNPs;Arg702Trp and Gly908Arg)18 and a frameshift muta-tion (Leu1007fsinsC),18 –20 which leads to a truncationof the protein transcript in the leucine-rich repeatregion, were significantly associated with Crohn’s dis-ease. Functional studies to clarify the precise role ofthese mutations in the pathophysiology of Crohn’sdisease are awaited. These mutations may confer sus-ceptibility to Crohn’s disease itself or may act asphenotype genes, determining the clinical subtypeand course of disease.
The HLA region on chromosome 6 has been impli-cated in determining both disease susceptibility andclinical phenotype by using linkage10,12,22,23 and associ-ation studies.23–26 However, not all genome-wide scanshave shown linkage to this region (IBD3), and associa-tion studies using molecular typing have not producedconsistent results. However, a recent meta-analysis ofmore than 20 studies highlighted associations withHLA-DRB1*0701 (odds ratio, 1.42; confidence interval[CI]: 1.16–1.74) and HLA-DRB3*0301 (odds ratio,2.18; CI: 1.25–3.80).27 Limited conclusions can bedrawn from these association studies for a number ofreasons. Particularly important is the linkage disequilib-rium seen across the HLA region. This results in highlyconserved haplotypes making it difficult to dissect outthe primary association. This problem is compounded bythe fact that the HLA region contains over 200 geneswith similar functions. In addition, studies have focusedalmost exclusively on class II HLA molecules and mark-ers in the tumor necrosis factor (TNF)� complex andlittle is known about associations with genes in the classI and III regions. Furthermore, because IBD3 confers amodest risk, true associations are likely to have beenmissed by studies involving small numbers of patients.Stronger associations have been observed with clinicalsubgroups, but discrepancies in patient profiles haveoften resulted in inconsistency.
The aim of this study was to establish the genetic basisof the clinical manifestations of Crohn’s disease by usingthe 2 strongest susceptibility loci. To avoid the problemsof previous HLA association studies, we fine-mapped andused linkage disequilibrium mapping across 24 genesfrom HLA-A–HLA-DQ by using 340 polymorphisms.
Materials and MethodsParticipants
A total of 244 white patients with Crohn’s disease wererecruited from a single tertiary referral center. Diagnosis ofCrohn’s disease was based on standard clinical, radiologic,endoscopic, and histologic criteria. Phenotypic details wereobtained on 2 occasions by retrospective case-note reviewbetween January and May 2001 by 2 investigators (T.A. andA.d.S). Any interobserver variation was resolved by discussion(Table 1 and Figure 1). Duration of follow-up was defined asthe interval between diagnosis and case-note review. Detailsregarding ethnicity, family history, and smoking history werefurther supplemented by a patient-completed postal question-naire. Fifteen patients reported a family history of IBD notpreviously recorded in the hospital notes and were defined ashaving a family history in this study. Smoking habit refers tousage in January 2001 as reported by patients; for analysispatients were divided into 2 groups only: never vs. current orex-smoker. Ethnicity was defined as Jewish if 3 or more
Table 1. Demographic Characteristics and Clinical Featuresof 244 Patients With Crohn’s Disease
Total number 244Sex (men/women) 102/142Age at diagnosis (yr)
Range 3.0–81.8Mean 28.0Median 24.7Interquartile range 19.9; 31.5
Duration of follow-up (yr)Range 0.4–56.8Mean 17.6Median 16.2Interquartile range 8.6; 25.2
Jewish (%) 3 (1.4)Family history in first- or second-degree relative (%)
Crohn’s disease 48 (19.7)IBD 76 (31.1)
Smoking habit (%)Never 85 (34.8)Ex-smoker 87 (35.7)Current 72 (29.5)
Location of diseaseIleal 201 (82.4)Colonic 150 (61.5)Colonic and normal small bowel 37 (15.2)Perianal 114 (46.7)Perianal and normal small bowel 27 (11.1)
Disease behaviorSurgical stenotic disease 158 (64.8)All stenotic disease 180 (73.8)Fistulating 87 (35.7)Fistulating and normal small bowel 18 (7.4)Severe inflammatory colonic disease 51 (20.9)
Extraintestinal manifestationsType I peripheral arthropathy 32 (13.1)Erythema nodosum 23 (9.4)Uveitis 13 (5.3)Primary sclerosing cholangitis 6 (2.5)
April 2002 MOLECULAR CLASSIFICATION OF CROHN’S DISEASE 855

grandparents were Jewish. Disease phenotype was classified bydisease location and disease behavior.
Disease Location
We defined each individual by 1 or more location sites.The presence of ileal disease was determined by small bowelenema, ileoscopy, or laparotomy findings. To exclude thepresence of more proximal ileal disease, a small bowel enemawas required before positively classifying a patient as havingno ileal involvement. Colonic disease was identified bycolonoscopy or barium enema. Patients with ileal disease whowere shown on colonoscopy to have patchy inflammation in thecaecum adjacent to the ileocaecal valve, but with an otherwisenormal colon were defined as having ileal disease. Perianaldisease was defined by the presence of perianal abscesses,fistulae, or ulcers, but not by the presence of skin tags.
Disease Behavior
Each individual was defined by the presence or absenceof 4 behavioral types. The presence of stenotic ileal disease wasdefined in 2 ways. Group 1 was the surgical stenotic diseasegroup, and included all patients who had undergone surgicalresection or stricturoplasty for obstructive symptoms. Group 2was the all-stenotic disease group, which, in addition, includedpatients with ileal disease who had not undergone resection orstricturoplasty, but yet had strictures or evidence of bariumhold-up on small bowel enema, irrespective of symptoms.Fistulating Crohn’s disease was defined by the presence of anabnormal communication between 2 epithelial surfaces. Thisdefinition included the presence of ischiorectal, intersphinc-teric, and pelvirectal abscesses if they connected with an epi-thelial surface either spontaneously or after surgical interven-tion. However, simple perianal abscesses were excluded fromthis definition because they occur frequently in the healthypopulation, and are frequently treated without involvement ofthe hospital Gastroenterology team. In the absence of Crohn’sactivity index scores, severe inflammatory colonic disease was
defined surgically by the use of colectomy or defunctioningloop ileostomy for refractory disease. Type 1 peripheral ar-thropathy was defined according to previously published cri-teria.28
Control Subjects
The healthy control group was randomly selected fromover 10,000 anonymized blood samples collected from healthscreening clinics in Bedfordshire (OXCHECK).29 Althoughnone of the control subjects had Crohn’s disease, no informa-tion regarding their family history was available. The studywas approved by the central Oxford region ethics committee(COREC 00.083).
Genotyping
All genotyping was performed by using polymerasechain reaction–sequence-specific primers. Genotyping wascomplete except for NOD2/CARD15 typing in 5 controlsubjects (excluded from NOD2/CARD15 and combined HLA/NOD2/CARD15 analysis). To aid accumulation of chromo-somal information, 70% of reactions contained mismatches atthe terminal 3� end of both sense and antisense primers.Medium-resolution classic HLA class I and II typing (HLA A,B, Cw, DRB1, DRB3, DRB5, DQB1) was performed aspreviously described.30 Additional nonsynonymous polymor-phisms in class Ib and III genes were selected from geneticdatabases according to putative gene function, previous asso-ciation with other diseases (including Crohn’s), and an allelefrequency, if known, of greater than 5% (80% power ofshowing a significant association at the 5% level assumingminimum relative risk [RR] of 2.5). Where possible, individ-ual SNPs within a gene were constructed into gene haplotypicblocks for analysis. Haplotypes described in the results arelisted in Table 2. Alleles or SNPs studied included S gene(�C619T, �C1243T), major histocompatibility complexclass I chain-related gene A (MICA; 56 alleles31), major his-tocompatibility complex class I chain-related gene B (MICB;
Figure 1. Anatomic location ofdisease in 244 patients withCrohn’s disease.
856 AHMAD ET AL. GASTROENTEROLOGY Vol. 122, No. 4
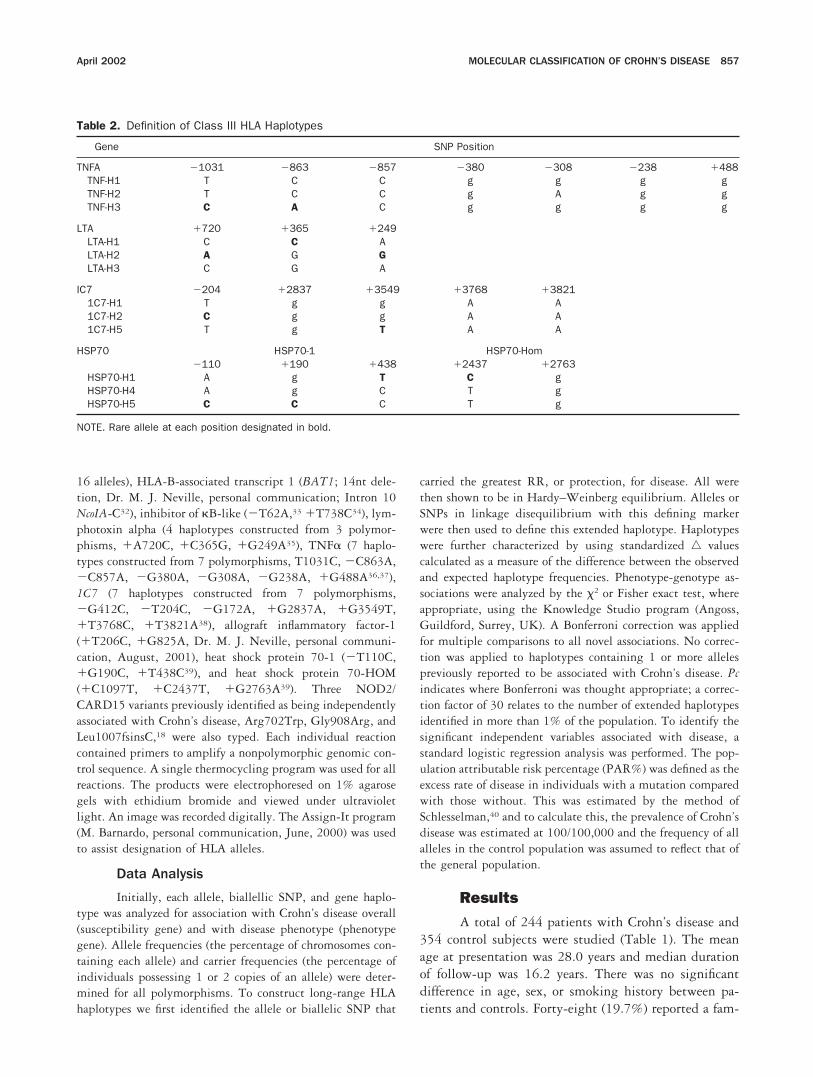
16 alleles), HLA-B-associated transcript 1 (BAT1; 14nt dele-tion, Dr. M. J. Neville, personal communication; Intron 10NcoIA-C32), inhibitor of �B-like (�T62A,33 �T738C34), lym-photoxin alpha (4 haplotypes constructed from 3 polymor-phisms, �A720C, �C365G, �G249A35), TNF� (7 haplo-types constructed from 7 polymorphisms, T1031C, �C863A,�C857A, �G380A, �G308A, �G238A, �G488A36,37),1C7 (7 haplotypes constructed from 7 polymorphisms,�G412C, �T204C, �G172A, �G2837A, �G3549T,�T3768C, �T3821A38), allograft inflammatory factor-1(�T206C, �G825A, Dr. M. J. Neville, personal communi-cation, August, 2001), heat shock protein 70-1 (�T110C,�G190C, �T438C39), and heat shock protein 70-HOM(�C1097T, �C2437T, �G2763A39). Three NOD2/CARD15 variants previously identified as being independentlyassociated with Crohn’s disease, Arg702Trp, Gly908Arg, andLeu1007fsinsC,18 were also typed. Each individual reactioncontained primers to amplify a nonpolymorphic genomic con-trol sequence. A single thermocycling program was used for allreactions. The products were electrophoresed on 1% agarosegels with ethidium bromide and viewed under ultravioletlight. An image was recorded digitally. The Assign-It program(M. Barnardo, personal communication, June, 2000) was usedto assist designation of HLA alleles.
Data Analysis
Initially, each allele, biallellic SNP, and gene haplo-type was analyzed for association with Crohn’s disease overall(susceptibility gene) and with disease phenotype (phenotypegene). Allele frequencies (the percentage of chromosomes con-taining each allele) and carrier frequencies (the percentage ofindividuals possessing 1 or 2 copies of an allele) were deter-mined for all polymorphisms. To construct long-range HLAhaplotypes we first identified the allele or biallelic SNP that
carried the greatest RR, or protection, for disease. All werethen shown to be in Hardy–Weinberg equilibrium. Alleles orSNPs in linkage disequilibrium with this defining markerwere then used to define this extended haplotype. Haplotypeswere further characterized by using standardized ‚ valuescalculated as a measure of the difference between the observedand expected haplotype frequencies. Phenotype-genotype as-sociations were analyzed by the �2 or Fisher exact test, whereappropriate, using the Knowledge Studio program (Angoss,Guildford, Surrey, UK). A Bonferroni correction was appliedfor multiple comparisons to all novel associations. No correc-tion was applied to haplotypes containing 1 or more allelespreviously reported to be associated with Crohn’s disease. Pcindicates where Bonferroni was thought appropriate; a correc-tion factor of 30 relates to the number of extended haplotypesidentified in more than 1% of the population. To identify thesignificant independent variables associated with disease, astandard logistic regression analysis was performed. The pop-ulation attributable risk percentage (PAR%) was defined as theexcess rate of disease in individuals with a mutation comparedwith those without. This was estimated by the method ofSchlesselman,40 and to calculate this, the prevalence of Crohn’sdisease was estimated at 100/100,000 and the frequency of allalleles in the control population was assumed to reflect that ofthe general population.
ResultsA total of 244 patients with Crohn’s disease and
354 control subjects were studied (Table 1). The meanage at presentation was 28.0 years and median durationof follow-up was 16.2 years. There was no significantdifference in age, sex, or smoking history between pa-tients and controls. Forty-eight (19.7%) reported a fam-
Table 2. Definition of Class III HLA Haplotypes
Gene SNP Position
TNFA �1031 �863 �857 �380 �308 �238 �488TNF-H1 T C C g g g gTNF-H2 T C C g A g gTNF-H3 C A C g g g g
LTA �720 �365 �249LTA-H1 C C ALTA-H2 A G GLTA-H3 C G A
IC7 �204 �2837 �3549 �3768 �38211C7-H1 T g g A A1C7-H2 C g g A A1C7-H5 T g T A A
HSP70 HSP70-1 HSP70-Hom�110 �190 �438 �2437 �2763
HSP70-H1 A g T C gHSP70-H4 A g C T gHSP70-H5 C C C T g
NOTE. Rare allele at each position designated in bold.
April 2002 MOLECULAR CLASSIFICATION OF CROHN’S DISEASE 857

ily history of Crohn’s disease in a first- or second-degreerelative. More than 1 site was involved in 68.8% ofpatients (Figure 1). Exclusively ileal, colonic, and peri-anal disease was seen in 54 (22.1%), 16 (6.6%), and 6(2.5%) patients, respectively.
Disease Susceptibility
NOD2/CARD15. The 3 NOD2/CARD15 poly-morphisms were significantly associated with disease aspreviously reported (Table 3).18–20 The RR was greatestfor carriage of the Leu1007fsinsC (RR � 6.5) comparedwith the 908Arg (RR � 2.3) and 702Trp (RR � 2.6)mutations. Although the rare variants of the 3 SNPswere never observed on the same haplotype, the greatestRR was seen in patients possessing a variant allele atmore than 1 of these loci (compound heterozygotes,RR � 29.3) (Table 4). This compared with an RR of 2.4for individuals possessing only a single copy of 1 variantallele (simple heterozygotes) and an RR of 9.8 for ho-mozygotes. The PAR% for Crohn’s disease overall was27.0% for possession of 1 or more variant alleles at anyof these 3 sites.
HLA. In univariate analysis, multiple HLA regionalleles were associated with disease overall. However,when linkage disequilibrium was taken into account,these reduced into 3 extended haplotypes (Figure 2).
Two of these were associated with disease susceptibil-ity whereas 1 was protective: The positive associationwith DRB1*0701 (34% vs. 26%; P � 0.04; RR � 1.5)and the protective association with DRB1*1501 (20%vs. 28%; P � 0.02; RR � 0.6) (Figure 2A ) confirms the
results of previous studies.24,25,41 A negative associationwas seen with other alleles on the DRB1*1501 extendedhaplotype including the TNF� haplotype TNFH1 (com-prising the common alleles at 6 promoter sites) (72% vs.81%; P � 0.01; RR � 0.6), and DQB1*0602 (20% vs.28%; P � 0.03; RR � 0.7). A novel finding was thepositive association with the Cw*0802 haplotype (Figure2B), which comprises Cw* 0802 (12% vs. 4%; P �0.0004; Pc � 0.01; RR � 3.0); B*14 (12% vs. 6%; P �0.004; Pc � NS; RR � 2.3); MICA*011 (8% vs. 4%;P � 0.04; Pc � NS; RR � 2.0); and 1C7�3571*A (8%vs. 4%; P � 0.03; Pc � NS; RR � 2.2). Althoughdetailed analysis revealed a haplotype comprising bothDRB1*0701 and Cw*0802, both alleles were shown tobe independently associated with disease. A further alleleon this haplotype, DRB3*0301, was not associated withdisease overall contrary to a previous report.42 Step-wiselinear regression revealed an additional haplotype basedaround MICA*010 when the DRB1*0701 individualswere removed from both patient and control groups(MICA*010 21% vs. 11%; P � 0.007; Pc � NS; RR �2.10) (Figure 2C ). In common with other HLA-associ-ated diseases, homozygosity for any of these 3 haplotypeswas not associated with additional risk. Furthermore,there was no evidence for synergy between these HLAhaplotypes and the NOD2/CARD15 variants.
Disease Phenotype
Age of presentation. Early age of diagnosis wasassociated with ileal (mean age ileal vs. nonileal 26.7 vs.34.2 yr; P � 0.0004) and fistulating disease (mean agefistulating vs. nonfistulating 25.6 vs. 29.3 yr; P � 0.03)(Table 5). Patients who possessed the Leu1007fsinsCvariant presented earlier when compared with both pa-tients without this variant (mean age, 23.1 vs. 29.0 yr; P� 0.007) and to patients with no NOD2/CARD15variant (mean age, 23.1 vs. 29.0 yr; P � 0.009). NOD2/CARD15 homozygote or compound heterozygote pa-tients presented earlier than patients with no NOD2/
Table 3. NOD2/CARD15 Variant Allele Frequencies inCrohn’s Disease Patients and Controls
Leu1007fsinsC 908Arg 702Trp
Controls 1.6 1.4 5.2Crohn’s disease 9.4 3.3 12.5P value P � 0.0001 P � 0.03 P � 0.0001RR 6.5 (3.3–12.7) 2.3 (1.1–5.2) 2.6 (1.7–4.0)
Table 4. NOD2/CARD15 Genotypes in Crohn’s Disease Patients and Controls
Total
Genotype (%)
No variant Simple heterozygote Homozygote Compound heterozygote Any variant
Controls 349 294 (84.2) 53 (15.2) 1 (0.3) 1 (0.3) 55 (15.8)Crohn’s disease overall 244 150 (61.5) 65 (26.6) 7 (2.9) 22 (9.0) 94 (38.5)P value — P � 0.0001 P � 0.005 P � 0.0001 P � 0.0001RR (95% CI) 1 2.4 (1.6–3.6) 9.8 (1.2–80.3) 29.3 (3.9–) 3.3 (2.3–4.9)PAR % 17.7% 4.1% 12.5% 27.0%
NOTE. Genotypes defined by 3 NOD2/CARD15 variants only (Arg702Trp, Gly908Arg, Leu1007fsinsC), which were never observed on the samehaplotype. Simple heterozygotes defined by possession of a single copy of any 1 of the 3 studied variant alleles; homozygotes defined bypossession of 2 copies of the same variant allele; compound heterozygotes defined by possession of 2 different variant alleles.
858 AHMAD ET AL. GASTROENTEROLOGY Vol. 122, No. 4

CARD15 variant (mean age, 23.5 vs. 29.0 yr; P � 0.04)(Table 6). Age of diagnosis was not associated with anyHLA allele or SNP once Bonferroni correction had beenapplied. The presence of a family history of Crohn’sdisease or IBD was not associated with age of diagnosisor NOD2/CARD15 genotype.
Disease location. A major stable determinant ofphenotype in Crohn’s disease is location.43 A significantassociation was found between ileal disease and posses-sion of 1 or more variant alleles. This was evident whenanalyzed against patients without ileal disease or againstcontrols (Table 7). Eighty-seven of 201 patients withileal disease possessed 1 or more variant alleles comparedwith 55 of 349 controls (P � 0.0001; RR � 4.1). Wheneach NOD2/CARD15 mutation was analyzed indepen-dently, a significant association was found with theLeu1007fsinsC only. All 42 patients who possessed the
Figure 2. Susceptibility, protective, and phenotype-determining long-range HLA haplotypes in Crohn’s disease. (A–D) Construction of HLAhaplotypes described in the Patients and Methods section. RR was calculated for each allele on haplotype. Statistically significant associationsare represented by }. Significant levels are reported in text. The 95% CI, shown for significant associations only, is represented by vertical bars.(A) DRB1*1501 Crohn’s disease protective haplotype, Crohn’s disease patients vs. controls; (B) Cw*0802 Crohn’s disease susceptibilityhaplotype, Crohn’s disease vs. controls; (C ) MICA*010 perianal phenotype haplotype, perianal Crohn’s disease patients vs. controls; (D) BAT1 In 10 Nco1 C colonic phenotype haplotype, colonic Crohn’s disease vs. noncolonic Crohn’s disease.
Table 5. Age of Diagnosis by Disease Phenotype
Diseaselocation
Present Absent
NumberMean age(SD/SEM) Number
Mean age(SD/SEM)
Ileal 201 26.7 (11.2/0.8)a 43 34.2 (17.5/2.7)Colonic 150 28.0 (13.0/1.1) 94 28.1 (12.5/1.3)Perianal 114 26.6 (12.1/1.1) 130 29.2 (13.2/1.2)Disease
behaviorStenotic ileal 190 26.4 (11.3/0.8)b 54 31.9 (16.5/2.2)Surgery for
stenotic 158 27.0 (11.6/0.9) 84 29.8 (14.6/1.6)Fistulating 87 25.6 (19.8/1.2)c 157 29.3 (13.6/26.1)Severe
colonic 51 25.7 (11.7/1.6) 193 28.6 (13.0/0.9)
aDisease location: ileal vs. nonileal; P � 0.0004.bDisease behavior: stenotic ileal vs. all other Crohn’s patients; P �0.01.cFistulating vs. nonfistulating; P � 0.03.
April 2002 MOLECULAR CLASSIFICATION OF CROHN’S DISEASE 859

frameshift mutation had ileal disease (P � 0.001; RR �22.6). A similar trend was observed with the Gly908Argmutation (allele frequencies; ileal disease 3.7%, no ilealdisease 1.2%, controls 1.4%) and the Arg702Trp muta-tion (allele frequencies; ileal disease 13.7%, no ilealdisease 7.0%, controls 5.2%). The RR increased withpossession of more than 1 variant allele; indeed, allcompound heterozygote and homozygote patients hadileal disease (P � 0.0001; RR � 30.3). There was nosignificant difference in NOD2/CARD15 variant allelecarriage between patients without ileal disease and con-trols.
HLA-DRB1*0701 was significantly associated withthe presence of ileal disease when compared with controls(35.8% vs. 25.7%; P � 0.01; RR � 1.61), in addition
to disease overall. However, because 75% of patientswith ileal disease had an overlapping phenotype, we wereunable to dissect the absolute specificity of the associa-tion.
The presence of colonic disease, with or without ilealdisease, was associated with alleles on the classic auto-immune HLA haplotype (A1-B8-DR3) (Figure 2D). Thepeak RR was found at BAT 1 (intron 10 NcoI A, 30.8%vs. 11.8%; P � 0.0007; Pc � 0.02; RR � 3.22).Homozygosity for any of the alleles on this haplotypeconferred no additional risk. Although the numbersstudied were small, involvement of the colon, with orwithout ileal involvement, was associated with the pres-ence of Type I IBD-associated peripheral arthropathy(18.7% vs. 4.2%; P � 0.001; RR � 5.16), and erythemanodosum (12.7% vs. 4.2%; P � 0.03; RR � 3.26). Inthis study, women were more likely than men to havecolonic disease (67% vs. 53%; P � 0.03; RR � 1.78).The 3 NOD2/CARD15 mutations did not confer sus-ceptibility to colonic disease irrespective of the presenceof ileal disease.
The primary genetic association with perianal diseasewas with the haplotype based around MICA*010 (Figure2C ), the same haplotype that was identified by step-wiseanalysis with Crohn’s overall (19.3% vs. 10.5%; P �0.01; RR � 2.1). Two other markers on this haplotypewere also associated with perianal disease when comparedwith controls (B*1501: 21.4% vs. 12.7%, P � 0.03,RR � 1.87; TNFH3: 46.4% vs. 35.3%, P � 0.04, Pc �NS, RR � 1.59). DRB1*0103 was also found to beassociated with perianal disease (10% vs. 4%; P � 0.01;Pc � NS; RR � 2.8). Although the numbers were small,this risk was associated with both the DRB1*0103–
Table 6. Ileal Crohn’s Disease—Age of Onset by NOD2/CARD15 Genotype
NOD2/CARD15 genotypeNumber ofpatients
Mean age at diagnosis(SD/SEM)
Carriage of Leu1007fsinsC 42 23.1 (7.4/1.1)a,b
Absence of Leu1007fsinsC 202 29.0 (13.4/0.9)Simple heterozygote 65 27.7 (11.8/1.5)Homozygote or compound
heterozygote 29 23.5 (7.0/1.3)c
Carriage of any 1 of the 3variants 94 26.4 (10.7/1.1)
None of the 3 variantscarried 150 29.0 (13.9/0.8)
aCarriage of Leu1007fsinsC vs. absence of Leu1007fsinsC, P �0.007.bCarriage of Leu1007fsinsC vs. none of the 3 variants carried, P �0.009.cHomozygote or compound heterozygote vs. none of the 3 variantscarried, P � 0.04.
Table 7. Crohn’s Disease Phenotype and NOD2 Mutations—Disease Location
Totalpatients
Genotype Allele frequency %
Novariant
Minimum 1variant allele
Simpleheterozygote Homozygote
Compoundheterozygote 1007fsinsC 908Arg 702Trp
Ileal disease 201 114 87a,f 58b,g 7c,h 22c,h 11.4d,i 3.7 13.7e
No ileal disease 43 36 7 7 0 0 0 1.2 7.0Colonic disease 150 96 54 38 4 12 9.0 3.0 11.3Colonic and no ileal disease 37 32 5 j 5k 0l 0m 0n 0 6.8No colonic disease 94 54 40 27 3 10 10.1 3.7 14.4Perianal disease 114 73 41 30 2 9 7.9 2.6 12.3No perianal disease 130 77 53 35 5 13 10.8 3.8 12.7Crohn’s disease overall 244 150 94 65 7 22 9.4 3.3 12.5Controls 349 294 55 53 1 1 1.6 1.4 5.2
Ileal disease vs. controls: aP � 0.0001, RR 4.1 (2.7–6.1); bP � 0.0001, RR 2.8 (1.8–4.3); cP � 0.0001, RR 30.3 (7.1–129.3); dP � 0.0001,RR 8.07 (4.1–15.8); eP � 0.0001, RR 2.9 (1.88–4.5).Ileal disease vs. no ileal disease: fP � 0.001, RR 3.7 (1.6–8.8); gP � 0.03, RR 2.5 (1.0–5.9); hP � 0.003, RR 18.8 (1.1–315.7); iP � 0.001,RR 22.6 (1.4–370.1).Colonic and no ileal disease vs. ileal disease: jP � 0.0006, RR 0.2 (0.1–0.6); kP � 0.01, RR 0.3 (0.1–0.9); lP � 0.005, RR 0.1 (0.004–1.00);mP � 0.002, RR 0.05 (0.003–0.8).
860 AHMAD ET AL. GASTROENTEROLOGY Vol. 122, No. 4

DQ*0501 and DRB1*0103–DQB1*0301 haplotypes.Alleles on the Cw*0802 haplotype, associated with over-all susceptibility, appeared to be particularly importantin perianal disease (DRB1*1302: 14% vs. 6%, P � 0.01,Pc � NS, RR � 2.4; DRB3*0301: 14% vs. 6%, P �0.01, Pc � NS, RR � 2.4). In common with colonicdisease, NOD2/CARD15 variants were not associatedwith perianal disease.
The PAR% for the 3 NOD2/CARD15 variants andHLA in ileal disease was estimated at 32% and 17%,respectively. This compared with a PAR% for theHLA region in colonic and/or perianal disease of 40%(Figure 3).
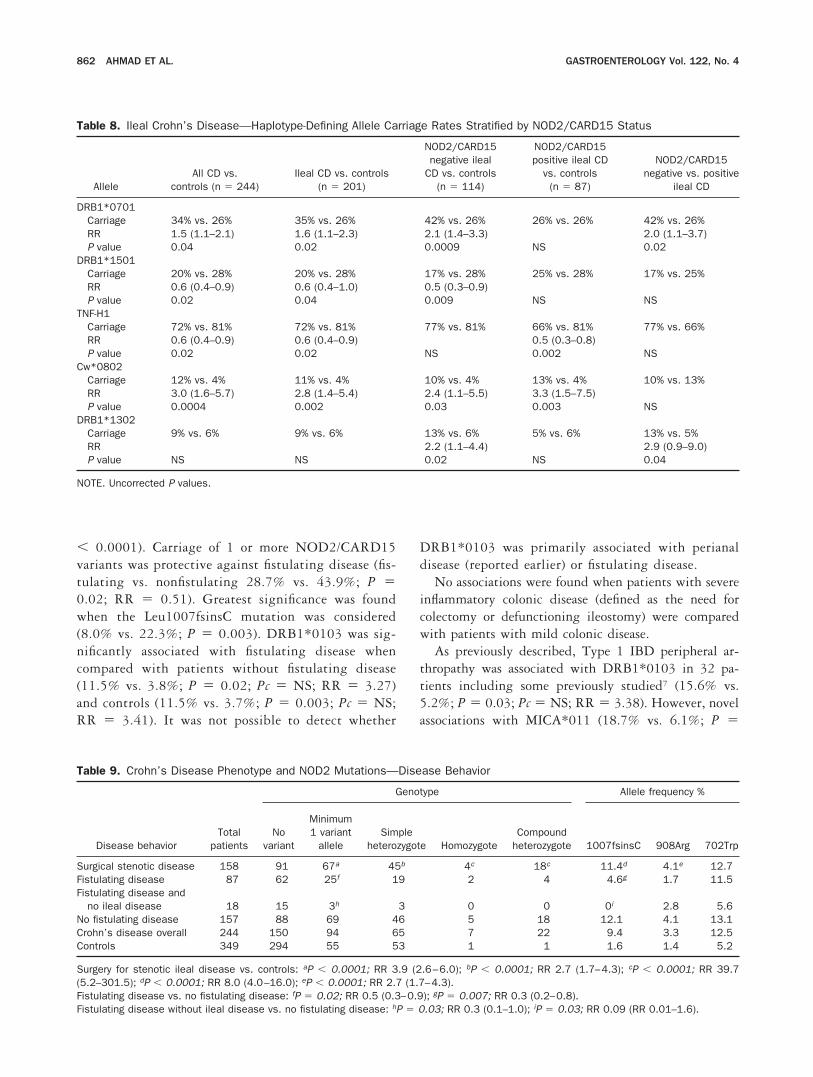
HLA and disease location stratified by NOD2/CARD15 status. HLA associations in patients with ilealdisease were re-analyzed after stratification by NOD2/CARD15 variant allele status. A summary of these datafor the extended haplotype defining alleles is presented inTable 8. DRB1*0701 is associated with ileal disease onlyin NOD2/CARD15-negative patients. A significant as-sociation with this allele was observed when NOD2/CARD15-negative ileal disease patients were comparedwith controls (NOD2/CARD15-negative ileal disease vs.controls 42% vs. 26%; P � 0.0009; RR � 2.1) and toNOD2/CARD15 positive patients (42% vs. 26%; P �0.02; RR � 2.0). The negative association withDRB1*1501 was observed only in ileal patients whowere NOD2/CARD15 negative. (NOD 2-negative pa-
tients vs. controls 16% vs. 28%; P � 0.005; RR � 0.5).The association with Cw*0802 was observed in all pa-tients with ileal disease irrespective of NOD2/CARD15status. Finally, NOD2/CARD15-negative ileal patientswere more likely to be homozygous for the 2 linkedTNF� polymorphisms TNF�857C and TNF�488g(common alleles) than patients with a NOD2/CARD15variant and ileal disease (91.2% vs. 74.8%; P � 0.001;Pc � 0.01; RR � 3.52).
Disease Behavior
Disease behavior in 180 (90%) patients with ilealdisease was defined as stenotic. Surgical resection orstricturoplasty for symptoms of small bowel obstructionwere performed in 158 (88%) of this group. There was asignificant association between both the all-stenotic andsurgical-stenotic ileal group and the NOD2/CARD15mutations (Table 9), but logistic regression showed thisnot to be independent of the association with ileal dis-ease. No genetic variable was shown to predict the needor time to initial surgical resection (data not shown).After a surgical resection, 77 (49%) patients required asecond surgery for obstructive small bowel disease. Nogenetic variable, including NOD2/CARD15, predictedthe rate of relapse as shown by Kaplan–Meier survivalanalysis (Figure 4).
As expected, fistulating disease was significantlyassociated with perianal disease (59.8% vs. 15.1%; P
Figure 3. Crohn’s disease location. PAR% of the NOD2/CARD15 variants and the HLA region. (A) Ileal disease; (B) colonic and/or perianaldisease. PAR% defined as the excess rate of disease in individuals with a NOD2/CARD15 variant or HLA haplotype compared with those withoutand are reported separately for patients with ileal disease and those with colonic and/or perianal disease. To estimate the PAR% of theNOD2/CARD15 variants we calculated the RR associated with carriage of 1 or more of the NOD2/CARD15 variants. To estimate the PAR% ofthe HLA region for ileal disease we calculated the RR associated with possession of either the DRB1*0701 or Cw*0802 haplotypes, and forcolonic and/or perianal disease we calculated the RR associated with possession of either the DRB1*0701, Cw*0802, MICA*010, or BAT 1in 10 Nco1 C haplotypes.
April 2002 MOLECULAR CLASSIFICATION OF CROHN’S DISEASE 861
� 0.0001). Carriage of 1 or more NOD2/CARD15variants was protective against fistulating disease (fis-tulating vs. nonfistulating 28.7% vs. 43.9%; P �0.02; RR � 0.51). Greatest significance was foundwhen the Leu1007fsinsC mutation was considered(8.0% vs. 22.3%; P � 0.003). DRB1*0103 was sig-nificantly associated with fistulating disease whencompared with patients without fistulating disease(11.5% vs. 3.8%; P � 0.02; Pc � NS; RR � 3.27)and controls (11.5% vs. 3.7%; P � 0.003; Pc � NS;RR � 3.41). It was not possible to detect whether
DRB1*0103 was primarily associated with perianaldisease (reported earlier) or fistulating disease.
No associations were found when patients with severeinflammatory colonic disease (defined as the need forcolectomy or defunctioning ileostomy) were comparedwith patients with mild colonic disease.
As previously described, Type 1 IBD peripheral ar-thropathy was associated with DRB1*0103 in 32 pa-tients including some previously studied7 (15.6% vs.5.2%; P � 0.03; Pc � NS; RR � 3.38). However, novelassociations with MICA*011 (18.7% vs. 6.1%; P �
Table 8. Ileal Crohn’s Disease—Haplotype-Defining Allele Carriage Rates Stratified by NOD2/CARD15 Status
AlleleAll CD vs.
controls (n � 244)Ileal CD vs. controls
(n � 201)
NOD2/CARD15negative ileal
CD vs. controls(n � 114)
NOD2/CARD15positive ileal CD
vs. controls(n � 87)
NOD2/CARD15negative vs. positive
ileal CD
DRB1*0701Carriage 34% vs. 26% 35% vs. 26% 42% vs. 26% 26% vs. 26% 42% vs. 26%RR 1.5 (1.1–2.1) 1.6 (1.1–2.3) 2.1 (1.4–3.3) 2.0 (1.1–3.7)P value 0.04 0.02 0.0009 NS 0.02
DRB1*1501Carriage 20% vs. 28% 20% vs. 28% 17% vs. 28% 25% vs. 28% 17% vs. 25%RR 0.6 (0.4–0.9) 0.6 (0.4–1.0) 0.5 (0.3–0.9)P value 0.02 0.04 0.009 NS NS
TNF-H1Carriage 72% vs. 81% 72% vs. 81% 77% vs. 81% 66% vs. 81% 77% vs. 66%RR 0.6 (0.4–0.9) 0.6 (0.4–0.9) 0.5 (0.3–0.8)P value 0.02 0.02 NS 0.002 NS
Cw*0802Carriage 12% vs. 4% 11% vs. 4% 10% vs. 4% 13% vs. 4% 10% vs. 13%RR 3.0 (1.6–5.7) 2.8 (1.4–5.4) 2.4 (1.1–5.5) 3.3 (1.5–7.5)P value 0.0004 0.002 0.03 0.003 NS
DRB1*1302Carriage 9% vs. 6% 9% vs. 6% 13% vs. 6% 5% vs. 6% 13% vs. 5%RR 2.2 (1.1–4.4) 2.9 (0.9–9.0)P value NS NS 0.02 NS 0.04
NOTE. Uncorrected P values.
Table 9. Crohn’s Disease Phenotype and NOD2 Mutations—Disease Behavior
Disease behaviorTotal
patients
Genotype Allele frequency %
Novariant
Minimum1 variant
alleleSimple
heterozygote HomozygoteCompound
heterozygote 1007fsinsC 908Arg 702Trp
Surgical stenotic disease 158 91 67a 45b 4c 18c 11.4d 4.1e 12.7Fistulating disease 87 62 25f 19 2 4 4.6g 1.7 11.5Fistulating disease and
no ileal disease 18 15 3h 3 0 0 0i 2.8 5.6No fistulating disease 157 88 69 46 5 18 12.1 4.1 13.1Crohn’s disease overall 244 150 94 65 7 22 9.4 3.3 12.5Controls 349 294 55 53 1 1 1.6 1.4 5.2
Surgery for stenotic ileal disease vs. controls: aP � 0.0001; RR 3.9 (2.6–6.0); bP � 0.0001; RR 2.7 (1.7–4.3); cP � 0.0001; RR 39.7(5.2–301.5); dP � 0.0001; RR 8.0 (4.0–16.0); eP � 0.0001; RR 2.7 (1.7–4.3).Fistulating disease vs. no fistulating disease: fP � 0.02; RR 0.5 (0.3–0.9); gP � 0.007; RR 0.3 (0.2–0.8).Fistulating disease without ileal disease vs. no fistulating disease: hP � 0.03; RR 0.3 (0.1–1.0); iP � 0.03; RR 0.09 (RR 0.01–1.6).
862 AHMAD ET AL. GASTROENTEROLOGY Vol. 122, No. 4

0.01; Pc � NS; RR � 3.53) and DRB1*1302, bothpresent on the Cw*0802 haplotype, were identified.
DiscussionThis study reports significant genetic associations
with clinical manifestations of Crohn’s disease by using alarge and rigorously phenotyped cohort. Patients wereethnically homogeneous, from a single British referralcenter, and had been followed-up for a median time of 16years. Clinical data collection was complete for all indi-viduals. The most powerful finding was that NOD2/CARD15 polymorphisms are strong predictors of ilealdisease, but are not associated with perianal or colonicdisease. The PAR% of ileal disease, caused by the 3NOD2/CARD15 variants, was estimated at 32%. Ex-tended haplotypes across the HLA were constructed from340 markers. This identified haplotypes that confer over-all susceptibility, and others that are location-specific.The influence of the HLA region was found to be greatestin determining colonic and/or perianal disease (PAR40%) compared with ileal disease (PAR 17%) (Figure 3).As observed in clinical practice, the majority of patientshad disease at more than 1 site in the gastrointestinaltract. Location of disease in these individuals was deter-mined by the possession of more than 1 phenotype-determining allele or haplotype. Thus, both HLA andNOD2/CARD15 associations were identified in the 113patients who had disease located to both the colon andileum.
Overall disease susceptibility. Overall diseasesusceptibility was conferred by alleles on 2 HLAhaplotypes. Our finding of an association with the
DRB1*0701 allele replicates the findings of 2 previousstudies.24,25 This allele conferred an RR of 1.5 (1.1–2.1),comparable with that reported by a recent meta-analy-sis.27 Although this RR is small, this allele is present in34% of Crohn’s patients and, therefore, the overall at-tributable risk is large. The second, Cw*0802-basedhaplotype, was present in 12% of patients, conferring anRR of 3.0 (1.6–5.7). This allele is common in theAshkenazi Jewish population, a group in which Crohn’sdisease is also prevalent. A third HLA haplotype basedon DRB1*1501 was protective against disease with a RRof 0.6 (0.4–0.9), supporting the association with theequivalent serologic type HLA-DR2 identified by themeta-analysis.27
Disease phenotype: age at diagnosis. Patientswho possessed the frameshift mutation, or compoundheterozygotes/homozygotes, presented significantly ear-lier than patients with no NOD2/CARD15 variant. Thisis consistent both with the epidemiologic observationthat younger age of onset is associated with a higherprevalence of ileal disease and stricturing complica-tions3,6 and genetic evidence that younger age of diag-nosis identifies families with greater linkage to the IBD1locus.44
Disease phenotype: location. Possession of aNOD2/CARD15 variant was significantly associatedwith ileal disease (RR � 3.3 for any single variant levelincreasing to 23.1 in the presence of more than 1variant). The RR was greatest for carriage of theLeu1007fsinsC (RR � 6.5). There were, however, 114(57%) patients with ileal disease who did not possess aNOD2/CARD15 variant. It is likely that disease in theseindividuals is associated with loci on other chromosomes,or possibly with other mutations in the IBD1 locus,either within the NOD2/CARD15 gene or another genenearby.18 DRB1*0701 is associated with ileal disease butonly in NOD2/CARD15-negative ileal disease patients.In this study, step-wise analysis identified an additionalassociation of homozygosity for the common TNF-857Cpromoter allele with ileal disease in patients without aNOD2/CARD15 variant allele. Functional experimentsare needed to investigate the possibility that this geno-type confers susceptibility to ileal disease by alteringbinding of the ubiquitous transcription factor OCT-1,45
a potential component of the NF-�B inflammatory cas-cade.
The classic autoimmune HLA haplotype (A1-B8-DR3), linked with other immunopathologic diseases,46
was associated with colonic disease. MICA*00801,MICB*0106, BAT1 intron10 NcoI, and IKBL-63A weremapped to this ancestral haplotype. Peak RR was found
Figure 4. Recurrent stenotic ileal disease—surgery-free survival—NOD2/CARD15 status. Kaplan–Meier survival plot. NOD2/CARD15-negative ileal patients (n � 91) represented by solid line. NOD2/CARD15-positive ileal patients (n � 67) represented by dashed line.
April 2002 MOLECULAR CLASSIFICATION OF CROHN’S DISEASE 863

at BAT1, which encodes a transcription factor associatedwith the down-regulation of cytokine production.47
Finally, the haplotype based on MICA*010, carried by15% of patients, was associated with disease overall instep-wise analysis, and with perianal disease in univariateanalysis. MICA encodes a stress-inducible membrane-bound glycoprotein expressed in intestinal epitheliumthat interacts with V1 �T cells and NK cells.48 Ofnote, MICA*010 encodes a protein that is not ex-pressed,49 the clinical consequences of which are un-known. Also found on this haplotype is the high-pro-ducing TNF-1031C promoter allele, raising thepossibility that previous association of Crohn’s diseasewith this allele50 is owing to linkage disequilibrium withMICA*010.
Disease phenotype: behavior. When disease be-havior was examined, fistulating disease was positivelyassociated with DRB1*0103, an allele previously associ-ated with severe ulcerative colitis.51,52 In addition, thepresence of a NOD2/CARD15 variant, and particularlythe Leu1007fsinsC mutation, was protective against fis-tulating disease (any NOD2/CARD15 variant, P �0.02; Leu1007fsinsC, P � 0.006). Stenotic disease waspositively associated with the presence of a NOD2/CARD15 mutation, but this was not independent of thelink with ileal disease. No other independent associationswere found with disease behavior phenotypes.
The identification that NOD2/CARD15 polymor-phisms are specific for ileal disease is novel. The func-tional effects of these variants and specificity for ilealdisease are at present unexplained. In vitro, theLeu1007fsinsC mutation reduces NF-�B activation, afinding that conflicts with previous work showing anincrease in NF-�B activation in monocytes53 from pa-tients with Crohn’s disease.
Several functional hypotheses have been proposed tolink polymorphisms in the HLA region, bacterial flora,and mucosal inflammation. Cross-reactivity may existbetween a bacterial peptide presented on a specific HLAclass II allele such as DRB1*0701, and a gut autoanti-gen, a mechanism similar to that postulated for reactivearthritis.54 This is supported by identification of murineMHC-restricted CD4� T cells reactive to enteric bacte-rial antigens that are able to induce colitis by adoptivetransfer.55 Alternatively, polymorphism in the genes in-volved in antigen presentation encoded in the HLAregion may alter the binding affinity of bacterial peptideand determine the T-cell repertoire. The genetic associ-ations may, however, simply reflect linkage disequilib-rium with a true disease allele lying hidden in the majorhistocompatibility complex—a region that is rich in
highly polymorphic genes with putative immune func-tion. A further possibility is that changes in the localmicroenvironment, determined either by polymorphiccytokines encoded by the HLA region, or by determi-nants of responsiveness to bacterial lipopolysaccharidesuch as NOD2/CARD15, influence the activation anddifferentiation of regulatory cells within the intestine.56
This study has shown the importance of NOD2/CARD15 and the HLA region in determining clinicalsubgroups of Crohn’s disease and may provide an initialbasis for the construction of a molecular classification ofCrohn’s disease.57 In time, this may provide the frame-work to understand the different mechanisms that un-derlie the disease heterogeneity of Crohn’s disease, andby patient stratification, permit the unraveling of thecomplex interaction between the genetic and environ-mental causes of disease.
References1. Sachar DB, Andrews HA, Farmer RG, Pallone F, Pena A, Prantera
Crutgeerts P. Proposed classification of patient subgroups inCrohn’s disease. Gastro Int 1992;5:141–154.
2. Gasche C, Scholmerich J, Brynskov J, D’Haens G, Hanauer S,Irvine E, Jewell D, Rachmilewitz D, Sachar D, Sandborn W, Suth-erland L. A simple classification of Crohn’s disease: Report of theWorking Party for the World Congress of Gastroenterology, Vienna1998. Inflamm Bowel Dis 2000;6:8–15.
3. Polito JM, Childs B, Mellits D, Tokayer AZ, Harris ML, Bayless TM.Crohn’s disease: influence of age at diagnosis on site and clinicaltype of disease. Gastroenterology 1996;111:580–586.
4. Satsangi J, Grootscholten C, Holt H, Jewell DP. Clinical patternsof familial inflammatory bowel disease. Gut 1996;38:738–741.
5. Bayless TM, Tokayer AZ, Polito JM, Quaskey SA, Mellits ED,Harris ML. Crohn’s disease: concordance for site and clinical typein affected family members—potential hereditary influences.Gastroenterology 1996;111:573–579.
6. Colombel J-F, Grandbastian B, Gower-Rousseau C, Plegat S,Evrard JP, Dupas JL, Gendre JP, Modigliani R, Belaiche J, HosteinJ, Hugot JP, van Kruiningen H, Cortot A. Clinical characteristics ofCrohn’s disease in 72 families. Gastroenterology 1996;111:604–607.
7. Orchard T, Thiyagaraja S, Welsh K, Wordsworth B, Gaston J,Jewell D. Clinical phenotype is related to HLA genotype in theperipheral arthropathies of inflammatory bowel disease. Gastro-enterology 2000;118:274–278.
8. Bouma G, Poen AC, Garcia-Gonzalez MA, Schreuder GMT, Felt-Bersma RJF, Meuwissen SG, Pena AS. HLA-DRB1*03, but not theTNF�-308 promoter gene polymorphism, confers protectionagainst fistulising Crohn’s disease. Immunogenetics 1998;47:451–455.
9. Cho JH, Nicolae DL, Gold LH, Fields CT, LaBuda MC, Rohal PM,Pickles MR, Qin L, Fu Y, Mann JS, Kirschner BS, Jabs EW, WeberJ, Hanauer SB, Bayless TM, Brant SR. Identification of novelsusceptibility loci for inflammatory bowel disease on chromo-somes 1p, 3q, and 4q: evidence for epistasis between 1p andIBD1. Proc Natl Acad Sci U S A 1998;95:7502–7507.
10. Rioux J, Silverberg M, Daly M, Steinhart A, Mcleod R, Griffiths A,Green T, Brettin T, Stone V, Bull S, Bitton A, Williams C, Green-berg G, Cohen Z, Lander E, Hudson T, Siminovitch K. Genome-wide search in Canadian families with inflammatory bowel dis-
864 AHMAD ET AL. GASTROENTEROLOGY Vol. 122, No. 4

ease reveals two novel susceptibility loci. Am J Hum Genet2000;66:1863–1870.
11. Rioux J, Daly M, Silverberg M, Lindblad K, Steinhart H, Cohen Z,Delmonte T, Kocher K, Miller K, Guschwan S, Kulbokas E,O’Leary S, Winchester E, Dewar K, Green T, Stone V, Chow C,Cohen A, Langelier D, Lapointe G, Gaudet D, Faith J, Branco N,Bull S, McLeod R, Griffiths A, Bitton A, Greenberg G, Lander E,Siminovitch K, Hudson T. Genetic variation in the 5q31 cytokinegene cluster confers susceptibility to Crohn disease. Nat Genet2001;29:223–228.
12. Hampe J, Schreiber S, Shaw S, Lau K, Bridger S, MacPherson A,Cardon L, Sakul H, Harris T. A genome-wide analysis providesevidence for novel linkages in inflammatory bowel disease in alarge European cohort. Am J Hum Genet 1999;64:808–816.
13. Satsangi J, Parkes M, Louis E, Hashimoto L, Kato N, Welsh K,Terwilliger JD, Lathrop GM, Bell JI, Jewell DP. Two-stage genome-wide search in inflammatory bowel disease provides evidence forsusceptibility loci on chromosomes 3, 7 and 12. Nat Genet1996;14:199–202.
14. Ma Y, Ohmen J, Li Z, Bentley L, McElree C, Pressman S, TarganS, Fischel-Ghodsain N, Rotter J, Yang H. A genome wide searchidentifies potential new susceptibility loci for Crohn’s disease.Inflamm Bowel Dis 1999;5:271–278.
15. Duerr R, Barmada M, Zhang L, Pfutzer R, Weeks D. High-densitygenome scan in Crohn’s disease shows confirmed linkage toChromosome 14q11-12. Am J Hum Genet 2000;66:1857–1862.
16. Hugot JP, Laurent-Puig P, Gower-Rousseau C, Olson JM, Lee JC,Beaugerie L, Naom I, Dupas JL, Van Gossum A, Orholm M,Bonaiti-Pellie C, Weissenbach J, Mathew CG, Lennard-Jones JE,Cortot A, Colombel JF, Thomas G. Mapping of a susceptibilitylocus for Crohn’s disease on chromosome 16. Nature 1996;379:821–822.
17. The International IBD Genetics Consortium. International collab-oration provides convincing linkage replication in complex dis-ease through analysis of a large pooled data set: Crohn’s dis-ease and chromosome 16. Am J Hum Genet 2001;68:1165–1171.
18. Hugot JP, Chamaillard M, Zouali H, Lesage S, Cezard JP, BelaicheJ, Almer S, Tysk C, O’Morain CA, Gassull M, Binder V, Finkel Y,Cortot A, Modigliani R, Laurent-Puig P, Gower-Rousseau C, MacryJ, Colombel JF, Sahbatou M, Thomas G. Association of NOD2leucine-rich repeat variants with susceptibility to Crohn’s dis-ease. Nature 2001;411:599–603.
19. Ogura Y, Bonen DK, Inohara N, Nicolae DL, Chen FF, Ramos R,Britton H, Moran T, Karaliuskas R, Duerr RH, Achkar JP, Brant SR,Bayless TM, Kirschner BS, Hanauer SB, Nunez G, Cho JH. Aframeshift mutation in NOD2 associated with susceptibility toCrohn’s disease. Nature 2001;411:603–606.
20. Hampe J, Cuthbert A, Croucher PJ, Mirza MM, Mascheretti S,Fisher S, Frenzel H, King K, Hasselmeyer A, MacPherson AJ,Bridger S, van Deventer S, Forbes A, Nikolaus S, Lennard-JonesJE, Foelsch UR, Krawczak M, Lewis C, Schreiber S, Mathew CG.Association between insertion mutation in NOD2 gene andCrohn’s disease in German and British populations. Lancet2001;357:1925–1928.
21. Ogura Y, Inohara N, Benito A, Chen F, Yamaoka S, Nunez G.Nod2, a Nod1/Apaf-1 family member that is restricted to mono-cytes and activates NF-KB. J Biol Chem 2001;276:4812–4818.
22. Yang H, Plevy SE, Taylor K, Tyan D, Fischel-Ghodsian N, McElreeC, Targan SR, Rotter JI. Linkage of Crohn’s disease to the majorhistocompatibility complex region is detected by multiple non-parametric analyses. Gut 1999;44:519–526.
23. Hampe J, Shaw S, Saiz R, Leysens N, Lantermann A, MascherettiS, Lynch N, MacPherson AJ, Bridger S, van Deventer S, StokkersP, Morin P, Mirza MM, Forbes A, Lennard-Jones JE, Mathew CG,Curran ME, Schreiber S. Linkage of inflammatory bowel disease
to human chromosome 6p. Am J Hum Genet 1999;65:1647–1655.
24. Reinshagen M, Loeliger C, Kuehni P. HLA Class II gene frequen-cies in Crohn’s disease: a population based analysis in Germany.Gut 1996;38:538–542.
25. Danze PM, Colombel JF, Jacquot S, Loste MN, Heresbuch D,Ategbo S, Khamassi S, Perichon B, Semana G, Charron D, CezardJP. Association of HLA class II genes with susceptibility toCrohn’s disease. Gut 1996;39:69–72.
26. Trachtenberg E, Yang H, Hayes E, Vinson M, Lin C, Targan S, TyanD, Erlich H, Rotter J. HLA class II haplotype associations withinflammatory bowel disease in Jewish (Ashkenazi) and non-Jew-ish caucasian populations. Hum Immunol 2000;61:326–333.
27. Stokkers PC, Reitsma PH, Tytgat GN, van Deventer SJ. HLA-DRand -DQ phenotypes in inflammatory bowel disease: a meta-analysis. Gut 1999;45:395–401.
28. Orchard TR, Wordsworth BP, Jewell DP. Peripheral arthropathiesin inflammatory bowel disease: their articular distribution andnatural history. Gut 1998;42:387–391.
29. Langham S, Thorogood M, Normand C, Muir J, Jones L, Fowler G.Costs and cost effectiveness of health checks conducted by nursesin primary care: the Oxcheck study. BMJ 1996;312:1265–1268.
30. Welsh K, Bunce M. Molecular typing for the MHC with PCR-SSP.Rev Immunogenet 1999;1:157–176.
31. Robinson J, Perez-Rodriguez M, Waller M, Cuillerier B, Bahram S,Yao Z, Albert E, Madrigal J, Marsh S. MICA sequences 2000.Immunogenetics 2001;53:150–169.
32. Allcock R, Price P, Gaudieri S, Leelayuwat C, Witts C, Dawkins R.Characterization of the human central MHC gene, BAT1: genomicstructure and expression. Exp Clin Immunogenet 1999;16:98–106.
33. Allcock R, Baluchova K, Cheong K, Price P. Haplotypic singlenucleotide polymorphism in the central MHC gene IKBL, a poten-tial regulator of NF-�B function. Immunogenetics 2001;52:289–293.
34. Allcock R, Christiansen F, Price P. The central MHC gene IKBLcarries a structural polymorphism that is associated with HLA-A3,B7, DR15. Immunogenetics 1999;49:660–665.
35. Fanning G, Bunce M, Black C, Welsh K. Polymerase chain reac-tion haplotyping using 3� mismatches in the forward and reverseprimers: application to the biallelic polymorphisms of tumournecrosis factor and lymphotoxin alpha. Tissue Antigens 1997;50:23–31.
36. Wilson AG, di Giovine FS, Blakemore A, Duff G, Putte L, Duff G. Analleleic polymorphism within the human tumour necrosis factoralpha gene detectable by NcoI restriction of PCR product. HumMol Genet 1993;1:353.
37. Higuchi T, Seiki N, Kamizono S. Polymorphism of the 5�-flankingregion of the human tumour necrosis factor (TNF)-alpha gene inJapanese. Tissue Antigens 1998;51:605–612.
38. Ribas G, Neville M, Campbell R. Single-nucleotide polymorphismdetection by denaturating high-performance liquid chromatogra-phy and direct sequencing in genes in the MHC class III regionencoding novel cell surface molecules. Immunogenetics 2001;53:369–381.
39. Milner C, Campbell R. Structure and expression of the threeMHC-linked HSP70 genes. Immunogenetics 1990;32:242–251.
40. Schlesselman J. Case-control studies. Oxford: Oxford UniversityPress, 1982.
41. Purrmann J, Bertrams J, Knapp M, Cleveland S, Hengels KJ,Gemsa R, Strohmeyer G. Gene and haplotype frequencies of HLAantigens in 269 patients with Crohn’s disease. Scand J Gastro-enterol 1990;25:981–985.
42. Forcione D, Sands B, Isselbacher K, Rustgi A, Podolsky D, PillaiS. An increased risk of Crohn’s disease in individuals who inheritthe HLA class II DRB3*0301 allele. Proc Natl Acad Sci U S A1996;93:5094–5098.
April 2002 MOLECULAR CLASSIFICATION OF CROHN’S DISEASE 865

43. Louis E, Collard A, Oger A, Degroote E, Abdoul Nasr El Yafi F,Belaiche J. Behaviour of Crohn’s disease according to the Viennaclassification: changing pattern over the course of the disease.Gut 2001;49:777–782.
44. Brant S, Panhuysen C, Bailey-Wilson J, Rohal P, Lee S, Mann J,Ravenhill G, Kirschner B, Hanauer S, Cho J, Bayless T. Linkageheterogeneity for the IBD1 locus in Crohn’s disease pedigrees bydisease onset and severity. Gastroenterology 2000;119:1483–1490.
45. Hohjoh H, Tokunaga K. Allele-specific binding of the ubiquitoustranscription factor OCT-1 to the functional single nucleotidepolymorphism (SNP) sites in the tumour necrosis factor-alphagene (TNFA) promoter. Genes Immun 2001;2:105–109.
46. Price P, Witt C, Allcock R, Sayer D, Garlepp M, Kok CC, French M,Mallal S, Christiansen F. The genetic basis for the association ofthe B8.1 ancestral haplotype. Immunol Rev 1999;167:257–274.
47. Allcock R, Williams J, Price P. The central MHC gene, BAT1, mayencode a protein that down-regulates cytokine production. GenesCells 2001;6:487–494.
48. Groh V, Steinle A, Bauer S, Spies T. Recognition of stress-induced MHC molecules by intestinal epithelial gamma delta Tcells. Science 1998;279:1737–1740.
49. Li Z, Groh V, Strong RK, Spies T. A single amino acid substitutioncauses loss of expression of a MICA allele. Immunogenetics2000;51:246–248.
50. Negoro K, Kinouchi Y, Hiwatashi N, Takahashi S, Takagi S, SatohJ, Shimosegawa T, Toyota T. Crohn’s disease is associated withnovel polymorphisms in the 5� flanking region of the tumournecrosis gene. Gastroenterology 1999;117:1062–1068.
51. Satsangi J, Welsh KI, Bunce M, Julier C, Farrant JM, Bell JI, JewellDP. Contribution of genes of the major histocompatibility complexto susceptibility and disease phenotype in inflammatory boweldisease. Lancet 1996;347:1212–1217.
52. Roussomoustakaki M, Satsangi J, Welsh K, Louis E, Fanning G,Targan S, Landers C, Jewell DP. Genetic markers may predictdisease behaviour in patients with ulcerative colitis. Gastroenter-ology 1997;112:1845–1853.
53. Schreiber S, Nickolaus S, Hampe J. Activation of nuclear factorkB in inflammatory bowel disease. Gut 1998;42:477–484.
54. Ebringer A, Wilson C. HLA molecules, bacteria and autoimmunity.J Med Microbiol 2000;49:305–311.
55. Cong Y, Brandwein S, McCabe R, Lazenby A, Birkenmeier E,Sundberg J, Elson C. CD4 T cells reactive to enteric bacterialantigens in spontaneously colitic C3H/HeJBir mice: increased Thelper type 1 response and ability to transfer disease. J Exp Med1998;187:855–864.
56. Powrie F, Correa O. Regulatory interactions between CD-45R Bhigh and CD45R Blow CD4� T cells are important for the balancebetween protective and pathogenic cell-mediated immunity. J ExpMed 1994;179:589–600.
57. Bell J. The new genetics in clinical practice. BMJ 1998;316:618–620.
Received October 10, 2001. Accepted October 17, 2001.Address requests for reprints to: Tariq Ahmad, Gastroenterology
Unit, Gibson Laboratories, University of Oxford, Radcliffe Infirmary,Woodstock Road, Oxford OX2 6QX, United Kingdom. e-mail:[email protected]; fax: (44) 0-1865-790792.Supported by a grant from the National Association of Crohn’s and
Colitis (NACC, UK) (to T.A.) and the Institute G. Toniolo di Studi Supe-riori (to A.A.).The authors thank the patients that participated in this study. The
authors acknowledge the contributions of Heather Holt and Sue Gold-thorpe for organization and collection of blood specimens, MatthewNeville for details of novel HLA class III polymorphisms, and RobertWalton for allowing us to use the OXCHECK samples.
866 AHMAD ET AL. GASTROENTEROLOGY Vol. 122, No. 4




















