
The locust's use of motion parallax to measure distance
10
J Comp Physiol A (1990) 167:579-588 Journal of Comparative Springer-Verlag 1990 The locust's use of motion parallax to measure distance Erik C. Sobel* Institute of Neurological Sciences, University of Pennsylvania, Philadelphia, PA 19104, USA Accepted July 30, 1990 Summary. 1. Locusts perform a visual behavior known as peering before jumping to targets. Peering consists of a side to side translational movement of the head. Wallace (1959) first proposed that this behavior is per- formed in order to obtain depth information through motion parallax. The present study analyzes more quan- titatively the role of peering in the locust's (Schistoeerca americana) estimation of distance in an attempt to un- derstand the neural mechanisms involved. 2. Jump velocity was found to be related monotoni- cally to target distance and was used as a measure of the locust's judgement of target distance. 3. By presenting locusts with a target which moved laterally while they peered (artificial parallax), it was possible to simulate the motion parallax of a target at any distance. Jump velocities elicited by means of artifi- cial parallax were the same as jump velocities elicited by the corresponding real distances, demonstrating that locusts use motion parallax as a cue to distance. 4. By moving the target in the same direction but further than the locust's head it was possible to simulate targets whose motion parallax specified a position be- hind the animal. Locusts jump forward to such paradox- ical stimuli with the jump velocity appropriate for targets located at the absolute value of the simulated distance. This suggests that locusts are insensitive to the relation between the direction of head and image motion. 5. Measurement of jump velocity in locusts with one eye occluded revealed an interaction between the paral- lax signals from the two eyes - locusts appear to sum (or average) the motion perceived in each eye. 6. Jump velocity is predicted more accurately by tar- get distance than by either image displacement or image velocity during peering. This implies that the locust's computation of target distance involves signals concern- ing its own head motion. * Present address: Laboratory for Sensorimotor Research, Nation- al Institutes of Health, Building 10, Room 10-C-101, Bethesda, Md 20892, USA Key words: Motion parallax - Distance - Locust - Peer- ing - Depth perception Introduction Motion parallax is the phenomenon in which an observ- er's translational motion through a stationary environ- ment results in the perception of apparent motion of objects in the environment. The apparent motion of a particular object depends on its distance from the ob- server and the extent of the observer's motion. Closer objects appear to move faster and farther than more distant objects. If the translational motion of the observ- er is known, the distance between the observer and any object in the scene can be determined. Consequently, motion parallax can be a cue to distance. It has been demonstrated that many insects perceive the distances to food, mates and landmarks (Maldonado and Rodriguez 1972; Wehner and Horn 1975; Collett and Land 1975, 1978; Cartwright and Collett 1979; Wehner 1981; Collett and Harkness 1982; Cheng et al. 1987; Collett 1988; Lehrer et al. 1988). Due to the char- acteristics of compound eyes motion parallax is widely believed to be the dominant visual cue for distance among arthropods. Accommodation cannot provide a cue. The focal length of an ommatidial lens is short and fixed so images are in focus when object distances exceed a few millimeters (Wehner 1981). Stereopsis has limited range due to small interocular distance and low acuity (although mantids at least do use stereopsis for short range distance perception (Rossel 1983, 1986)). The apparent size of an object (number of ommatidia stimulated) can only be used as a cue for distance if the viewer knows the actual size of the object. This cue is exploited by hoverflies in their pursuit of conspecifics (Collett and Land 1975, 1978). The use of motion paral- lax by insects as a distance cue has been previously dem- onstrated in locusts (Wallace 1959; Collett 1978), crick- ets (Goulet et al. 1981), and bees (Cartwright and Collett
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of The locust's use of motion parallax to measure distance

J Comp Physiol A (1990) 167:579-588 Journal of Comparative
�9 Springer-Verlag 1990
The locust's use of motion parallax to measure distance
Erik C. Sobel*
Institute of Neurological Sciences, University of Pennsylvania, Philadelphia, PA 19104, USA
Accepted July 30, 1990
Summary. 1. Locusts perform a visual behavior known as peering before jumping to targets. Peering consists of a side to side translational movement of the head. Wallace (1959) first proposed that this behavior is per- formed in order to obtain depth information through motion parallax. The present study analyzes more quan- titatively the role of peering in the locust's (Schistoeerca americana) estimation of distance in an attempt to un- derstand the neural mechanisms involved.
2. Jump velocity was found to be related monotoni- cally to target distance and was used as a measure of the locust's judgement of target distance.
3. By presenting locusts with a target which moved laterally while they peered (artificial parallax), it was possible to simulate the motion parallax of a target at any distance. Jump velocities elicited by means of artifi- cial parallax were the same as jump velocities elicited by the corresponding real distances, demonstrating that locusts use motion parallax as a cue to distance.
4. By moving the target in the same direction but further than the locust's head it was possible to simulate targets whose motion parallax specified a position be- hind the animal. Locusts jump forward to such paradox- ical stimuli with the jump velocity appropriate for targets located at the absolute value of the simulated distance. This suggests that locusts are insensitive to the relation between the direction of head and image motion.
5. Measurement of jump velocity in locusts with one eye occluded revealed an interaction between the paral- lax signals from the two eyes - locusts appear to sum (or average) the motion perceived in each eye.
6. Jump velocity is predicted more accurately by tar- get distance than by either image displacement or image velocity during peering. This implies that the locust's computation of target distance involves signals concern- ing its own head motion.
* Present address: Laboratory for Sensorimotor Research, Nation- al Institutes of Health, Building 10, Room 10-C-101, Bethesda, Md 20892, USA
Key words: Motion parallax - Distance - Locust - Peer- ing - Depth perception
Introduction
Motion parallax is the phenomenon in which an observ- er's translational motion through a stationary environ- ment results in the perception of apparent motion of objects in the environment. The apparent motion of a particular object depends on its distance from the ob- server and the extent of the observer's motion. Closer objects appear to move faster and farther than more distant objects. If the translational motion of the observ- er is known, the distance between the observer and any object in the scene can be determined. Consequently, motion parallax can be a cue to distance.
It has been demonstrated that many insects perceive the distances to food, mates and landmarks (Maldonado and Rodriguez 1972; Wehner and Horn 1975; Collett and Land 1975, 1978; Cartwright and Collett 1979; Wehner 1981; Collett and Harkness 1982; Cheng et al. 1987; Collett 1988; Lehrer et al. 1988). Due to the char- acteristics of compound eyes motion parallax is widely believed to be the dominant visual cue for distance among arthropods. Accommodation cannot provide a cue. The focal length of an ommatidial lens is short and fixed so images are in focus when object distances exceed a few millimeters (Wehner 1981). Stereopsis has limited range due to small interocular distance and low acuity (although mantids at least do use stereopsis for short range distance perception (Rossel 1983, 1986)). The apparent size of an object (number of ommatidia stimulated) can only be used as a cue for distance if the viewer knows the actual size of the object. This cue is exploited by hoverflies in their pursuit of conspecifics (Collett and Land 1975, 1978). The use of motion paral- lax by insects as a distance cue has been previously dem- onstrated in locusts (Wallace 1959; Collett 1978), crick- ets (Goulet et al. 1981), and bees (Cartwright and Collett

580 E.C. Sobeh Motion parallax in locusts
1979; Lehrer et al. 1988; Srinivasan et al. 1989; Kirchner and Srinivasan 1989).
The most readily observed and most frequently cited example of a scanning movement used to obtain motion parallax information is the peering behavior of the locust (Wallace 1959). When placed on a stick, the locust will walk to its end. It will then jump to the nearest surface even when given the choice between surfaces of the same angular size at different distances (Wallace 1959). Before jumping the locust performs a loosely stereotyped be- havior in which it sways its head and body from side to side while pivoting about the tip of the abdomen. This behavior is known as peering or scanning (Wallace 1959). Locusts typically perform several peers at a poten- tial target and may change body orientation toward an- other target and begin peering again. Peering becomes especially intense immediately prior to jumping and pre- cedes all directed jumps (it may be absent prior to escape jumps).
Collett (1978) found that in a typical peer, the tip of the abdomen remains stationary while the body moves from side to side in an arc. While the body is moving, the head counter-rotates against the body movement so that the net movement of the head is strictly translation- al, in a plane perpendicular to the line of sight. Counter- rotation of the head effectively prevents a confounding component of image motion due to observer rotation. As a result, the movement of the target's image across the locust's frontal retina is strictly a function of the distance between the locust and the target and the extent of the locust's head movement. The counter-rotation be- havior which Collett studied is consistent with the hy- pothesis that locusts use motion parallax to estimate distance.
Wallace (1959) investigated whether peering was in- volved in the visual estimation of distance and found that the distance the locust jumped was related to the motion of the target's image across the retina. Locusts were placed on a platform facing a single target (tall black rectangle). The target was mounted on a lever which was invisible to the locust. As the locust peered to the left the target was moved to the right, and as the locust peered to the right the target was moved to the left. Wallace moved the target against the locust's peering movement creating an exaggerated change in visual angle and simulating the motion parallax of an apparent target which lay closer to the locust than the actual target. Locusts in this experiment jumped short of the target strongly suggesting that the locust measured the motion of the target image and computed jumping distance through motion parallax.
One limitation of Wallace's experiment was that he could only record whether the locust hit or missed the target. The locust's jump trajectory ensures that it hits the target even if it jumps too hard. Consequently, over- jumping could not be assessed reliably. The experiments reported in the present paper exploit modern technology. Jumps were recorded on videotape allowing quantitative analysis. The linkage of the locust's peering to stimulus motion was controlled automatically. This improvement allowed the experimental simulation of a wide range of
different target distances including distances both short- er and longer than the actual target distance.
Materials and methods
All experiments were performed on male 4th and 5th instar juvenile Sehistocerca americana (Forsk~l). The locusts were obtained from a laboratory colony at the University of Pennsylvania. Five or 6 individuals from the colony were housed in a small cage with a 16/8 h day/night cycle at 30-33 ~ The locusts were given ad lib access to young wheat plants, supplemented by a 3:1 mixture of wheat bran/wheat germ, and access to water. Only locusts which showed jumping behavior in the test paradigm were used.
Locusts are attracted to vertical contrasting edges, such as a tall black rectangle against a white background (Wallace 1958a, b, 1959). If placed on a platform of the type used by Wallace (1959) at short distance from such a target, the locust would some- times jump directly at the target, but more often the locust walked down the side of the platform and over to the target. To induce jumping, a different type of platform (1 inch diam disc covered with cotton fabric) was placed in a small pan of water (9 inch diam, 1 inch deep aluminum pie pan) just above the surface of the water. A locust placed on such a platform would orient toward a vertical black rectangle or any contrasting vertical edge and peer at it repeatedly. Occasionally the animal would jump at the edge, more often it did not. To coax the locust to jump a heat lamp was placed approximately 30 cm above the animal. As its tempera- ture rose, the animal appeared to search for a contrasting edge to jump at. When the entire apparatus was surrounded with plain, white paper so that the black target was more salient, the animal primarily directed its attention to the target. After performing sev- eral peers with intermittent pauses the animal jumped at the target. When the animal landed on the target the heat lamp and an addi- tional flood lamp were both immediately extinguished (see Fig. 1).
Locusts were placed on the jumping platform in front of one target placed at one of several possible distances. The sizes of the targets were fixed so that from the center of the platform all subtended the same visual angle (18 ~ horizontally, 57 ~ vertically). The jumps were recorded on videotape (shutter speed: 1/2000 s)
Video Camera~D . . . . . . . . . . _/L = = ~ ~ = ~ ~
Fig. 1. Experimental setup with a locust jumping from the jumping platform, a small disc in the center of a pie pan filled with water. Floor and walls were covered with white paper. The target was a tall vertical rectangle covered with black cotton fabric. The ani- mal was videotaped from above by means of a mirror placed above the animal

E.C. Sobel: Motion parallax in locusts 581
and analyzed frame by frame using a Panasonic AG1950 video cassette recorder.
The position of the locust's head in successive frames was measured by superimposing the video image from a 12 inch NEC color monitor on a 14 inch EGA computer monitor with a beam splitter. The video monitor and computer monitor were placed at right angles with the beam splitter placed at a 45 ~ angle between them. A cross-hair was moved across the computer screen by means of a mouse pointing device. The mouse was then clicked to store the x-y coordinates of the head.
By subtracting the positions of the locust between successive video frames of its jump (30/s) a measure of average jump velocity was obtained. Since a typical jump lasted-3-5 frames, 2-4 such subtraction measurements were acquired, and the largest of these was defined as the velocity of that jump. The alternative procedure of using the average rather than the maximum of the 2-4 velocity measurements gave similar values.
Experiments concerning the peering mo-~ements and those re- quiring automated tracking of the head position were performed as follows. A second video camera (shutter speed 1/60 s) viewed the animal from above. A small white spot was placed on the top of the animal's head (typewriter correction fluid sandwiched between two coats of cyanoacrylate glue). The animals's head was prepared by swabbing it with a mild detergent solution, rinsed and air-dried. The animal was wrapped in paper towel during this procedure and supported by an alligator clip.
The output of the head tracking video camera was sent to a real-time video digitizing board (Datacube IVG128) in an AT- class micro-computer. The digitizing board digitized the complete video frame and performed image thresholding at video frame rates (30/s). The video image was thresholded so that only the white spot on the animal's head was visible. A small window (20 • 20 pix- els) was placed by the experimenter over the white spot in the video image by means of a mouse. Once the computer had detected the spot in the window, it automatically tracked and stored x-y coordinates of the centroid of the moving white spot. A beep alerted the experimenter if the spot escaped the window. This sys- tem provided 30 pairs of x-y coordinates per second which pro- vided sufficient resolution to capture the rather slow peering move- ment (0.5 cm/s on average). The last 33.3 s of x-y data before each jump were stored to disk.
Animals were presented with targets at various distances as described above, and their peering movement was recorded and analyzed to extract peer amplitude and peer velocity. Peer ampli- tude was defined as the distance the head travelled between succes- sive reversals of direction. Peer velocity was defined as the peer amplitude divided by the time between direction reversals. All peer- ing measurements were filtered to remove pauses. Pauses were de- fined as a series of at least 6 successive frames with no detectable movement between frames.
To alter the parallax during peering, head position was tracked by computer and the resulting signal used to move the target. The target was mounted 12 cm or 16 cm from the locust on the pen assembly of an x-y plotter and its lateral position relative to the locust controlled by the computer using a MetraByte PIOll output card and a digital to analog converter, AD557, from Analog Devices. This setup made it possible to place the target in any desired position relative to the head. The dynamic response of the x-y plotter was sufficient to keep pace with the locust's peering.
Results and discussion
Jump velocity as a measure o f perceived distance
In order to investigate whether the m o t i o n paral lax gen- erated dur ing peering is used to est imate target distance, one needs a behaviora l measure of the locust ' s distance j u d g m e n t as exhibited in its j u m p i n g behavior . Because
A to
to
i . -
5 O --I gJ > a.. :E
300
200
100
300
200
J 1 3
. i L
10
J l l
300
200
' ' ' 100 2 0 3 0 4 0
300
200
J 1 9
!
10
J 9
t
i | i
2 0 3 0 4 0
9 100 ' ' ' ' 100 ' ' ' '
0 10 2 0 3 0 4 0 0 10 2 0 3 0 4 0
TARGET DISTANCE (CM)
Fig. 2. Jump velocity in direction of the target (horizontal) as a function of target distance for 4 animals. Each data point is the average of 3-4 jumps (_ 1 SEM) with target at the specified dis- tance. Target distances varied randomly between jumps. Target size at each distance was selected so that the targets always sub- tended the same visual angle (18 ~ horizontally and 57 ~ vertically) at each distance when viewed from the center of the jumping plat- form
j u m p i n g is ballistic (Bennet -Clark 1975; Heit ler and Bur- rows 1977a, b; Benne t -Cla rk and Alder 1979) take-off velocity should provide a conven ien t an d reliable index of the dis tance the locust in tends to j u m p . Juvenile lo- custs are wingless, a nd once a i rborne have little means of in-f l ight j u m p correct ion, except perhaps t h rough pos tura l changes (Cooter 1973; Arbas 1983). However , due to the shor t du ra t i on of the j u m p (100-200 ms) it is unl ikely tha t posture exerts a s ignif icant effect on the locust ' s m o m e n t u m . Consequen t ly , the locust mus t de- cide how hard to j u m p at a target before it jumps .
Based on these cons idera t ions I first examined the dependence of j u m p velocity on target distance. The re- sults show that j u m p velocity increases m o n o t o n i ca l l y with target dis tance (Fig. 2). Thus , j u m p velocity is a reliable measure of the locust ' s in t ended j u m p i n g dis- tance, a nd therefore, of its est imate of target distance. In addi t ion , this measure enables one to discern in tended j u m p i n g distances which are no t only nearer bu t also far ther t han the actual target distance, which was no t possible in the earlier studies (see In t roduc t ion) .
The distance at which the locust makes con tac t with the target will vary with j u m p velocity, j u m p angle and the posture assumed dur ing the j um p . Because typical targets (stalks) are extended and the locust is itself quite large there can p resumab ly be considerable la t i tude in these parameters . It is perhaps surpr is ing that j u m p ve- locity should be so closely control led. One reason might be to avoid b o u n c i n g off the target or d a m a g i n g the l imbs or head as a result o f l and ing too hard.

582 E.C. Sobel: Motion parallax in locusts
Do locusts use mot ion p a r a l l a x?
To determine whether locusts use mot ion parallax to compute distance I employed an updated version of Wal- lace's (1959) experiment in which he moved the target while the animal peered at it. Here the locust's head position was tracked automatically (30 times/s) and the target movement controlled by computer (see Methods). A locust was placed on the jumping platform facing a target as in the previous experiment. The target was mounted 12 cm or 16 cm f rom the locust on the pen of an x-y plotter and was moved laterally as a function of the lateral position of the locust 's head. The locust peered at this stimulus as it did at stat ionary targets and jumped. Jump velocity was recorded.
I f the locust computes distance f rom motion paral- lax, then moving the target against the peering move- ment creates an exaggerated change in visual angle, mak- ing the target appear closer to the animal than it actually is. Moving the target with the locust 's peering creates a diminished change in visual angle, and so simulates a more distant target. By varying the sign and magnitude of the coupling between the lateral mot ion of the head and the lateral mot ion of the target it is possible to simulate the paral lax corresponding to a target located at any experimenter-specified distance. The geometric relationship below was calculated by the computer which moved the target, based on the locust 's head position, to stimulate the parallax produced by a target located at the specified distance f rom the locust.
Eq. (1)
T = H • [ ( D ' - D)/D']
T = target posit ion; H = locust head position; D ' = simu- lated target distance; D = real target distance.
For example, a target located 16 cm from the animal moving horizontally in the opposite direction from the locust's peering movement and with equal amplitude simulates the parallax produced by a stationary target 8 cm away. To simulate the parallax of a stationary tar- get 32 cm away the target on the x-y plotter 16 cm away must move in phase with the locust with half the ampli- tude of the locust's movement . I f the locust uses mot ion parallax, the moving target should create the proximal stimulus of a target at the simulated distance (Fig. 3). Jump velocity corresponding to a simulated distance could then be compared with jump velocity elicited by a stationary target at that distance.
Animals were first tested with stationary targets at a range of distances, and then tested with the same dis- tances as simulated by a moving target. The results are shown in Fig. 4. The jump velocities are the same for stationary targets at a particular distance as for targets at that simulated distance. A three-way repeated mea- sures A N O V A was applied in the comparison of 7 ani- mals, with 3 trials at each of 4 distances under both con- ditions (stationary targets at real distances and moving target simulating parallax). Bonferroni t-tests performed for each target distance revealed no significant difference (P>0 .01) between jump velocity in the two conditions, for distances greater than 8 cm. At the 8 cm target dis- tance there was a significant difference between the two conditions ( P < 0.01), this is reflected in Fig. 4. In some animals, the closest simulated distances (8 cm) elicited higher jump velocities than their stationary target coun- terparts. This may be due to confounding distance cues other than motion parallax, such as stereopsis. More likely, this discrepancy arises because of limitations of the stimulus presentation equipment. Simulating an 8 cm target distance requires a large amount of target move- ment for which the inertial mass of the plotter arm, and the discrete nature of the computer specified motion
A. STATIONARY TARGET B. MOVING TARGET (Target Appears Closer)
D = h/tan(a) D' = h/tan(b) b>a
C. MOVING TARGET (Target Appears More Distant)
/ D' = h/tan(c) /
g.~'o # ] o.
g ~
Fig. 3A-C. Geometry of motion parallax and artificial parallax. A A locust shown peering h cm to one side. Target is located D cm away. As the locust moves through distance h, angular position of the target changes from initially 0 ~ (straight ahead) to its final angular position b. Distance to the target may be computed by formula D = h/ tan(a). B A moving target creates artificial parallax of a closer target. As the locust moves a distance h the target is moved in opposite direction. Perceived change in angular position is b degrees and so a locust using the formula, D = h/tan(b) will estimate the target distance as the smaller distance, D', rather than the actual distance, D (because b > a). C A moving target creates the artificial parallax of a farther target. As the locust moves a distance h the target is moved t in same direction but not quite as far as the locust's movement (t<h). Perceived change in angular position is c degrees and so a locust using the formula, D = h/tan(c) will estimate the target distance as the greater distance, D', rather than the actual distance, D (because c < a)

E.C. Sobel: Motion parallax in locusts 583
,.=.,..,
O I,LI
=E t . )
>, I--
o ,.I LB >
O. =E
300
200
100
300 J 5 1
a 0 200
is 100
0 i i = I I 0
10 20 3O 4O 0
300 300 J 3 8
200
100
200
100
0 I i i i 0
10 2O 3O 4O 0
300 300 J 3 9
200
II 100
11 t It 200
J 3 2
6
6
I i i
10 20 30
J 3 4
100
! ! i
10 20 3O
J 3 6
&
i
4O
i
4O
0 i i | I I i I i I I
10 20 30 40 10 20 30 40
T A R G E T D I S T A N C E (CM) Fig. 4. Jump velocity vs target distance with a stationary target (filled circles) and an artificial parallax target located at a fixed distance from the locust (open triangles). All targets have same apparent size (subtend same visual angle). Symbols: average jump velocity from 3 jumps at each distance (except data for J51 which shows average of 7jumps at each distance). Error bars: + 1 SEM. Artificial parallax generated by moving target laterally based on monitored head position of the locust. All moving targets for data on the left side of the figure (J51, J38, J39) were located 16 crn from the animal, so the 'moving target' condition in which target motion simulated the parallax of a target 16 cm away required no target movement. Moving targets for data on right side (J32, J34, J36) were located 12 cm from the locust and required no target movement to ' simulate' that distance
may become more salient, leading to a breakdown of the illusion. The reason this discrepancy is seen in some animals and not others may be because these locusts used a larger peering movement which further increased the amount of target motion required to simulate a near target.
Further evidence that close distances can be success- fully simulated was obtained by moving the target so as to simulate the parallax appropriate for distances of 2-5 cm. The animal peering at such a stimulus reaches with its forelegs and turns and flails its hindlegs in the direction of the target although it is 16 cm away, provid- ing striking evidence that the moving target creates the illusion of a closer target. This behavior is also observed when the locust is presented with a truly nearby target (in which case it is very effective at acquiring a purchase
on the target). This reaching behavior is rarely seen with more distant targets. Thus if other distance cues are available they must have little weight in controlling jumping.
Accordingly, when the target is made to move exactly in phase with the locust with the same amplitude as the peering movement (simulating a very distant target) the locust loses interest in the target and turns repeatedly around the platform searching in vain for some other surface at which to jump.
The close correspondence between jump velocity for real and simulated distances shows that motion parallax is the major factor in the locusts estimate of target dis- tance and confirms Wallace's (1959) conclusion that peering locusts exploit motion parallax for distance per- ception.
Paradoxical parallax
Because the movement of the target is electronically linked to the movement of the locust the normal rela- tionship between head and image movement may be al- tered. For example the target can be moved in the same direction as the locust's head but farther. When this is done the resulting image motion is in the same direc- tion as head movement. This is contrary to the normal situation in which any motion parallax is always oppo- site to the direction of head movement. The term para- doxical parallax has been coined to describe this abnor- mal relation between head and image motion. The alge- braic consequence of reversing the sign of motion paral- lax is a reversed sign of the computed distance. For con- venience this will be referred to as a situation in which the target is located at negative distance (i.e. behind the animal) (Fig. 5).
To examine how locusts reacted to such a stimulus, I compared jump velocity in locusts presented 1) with stationary targets (baseline condition) at different dis- tances in front of the animal, 2) moving targets simulat- ing the same positive distances, 3) with moving targets simulating negative distances (paradoxical parallax con- dition).
Quite surprisingly, the locusts did not appear to find this stimulus unusual and jumped forward with the same velocities they used for targets located at the absolute value of the specified distances (Fig. 6). A three-way re- peated measures ANOVA was applied to data from 4 animals which performed 3 trials at each of 4 distances under the two conditions (stationary targets at different distances and a moving target presenting paradoxical parallax). There was no significant difference between jump velocities under the two conditions (P--0.279). This result suggests that either locusts measure the mag- nitude of the apparent motion but, rectify (or are insensi- tive to) direction of motion, and/or locusts are insensi- tive to the direction of head movement. Thus in comput- ing target distance locusts seem to ignore the relation between the directions of head and image movement.

584 E.C. Sobel: Motion parallax in locusts
D, E
PARADOXICAL PARALLAX
D' = h/tan(b)~ t ~ I
Fig. 5. Geometry of paradoxical parallax. A locust peering h cm to one side. Target is located D cm away. As the locust moves distance h the target is moved a distance t in same direction. Conse- quently, while angular position of the target is initially a, final angular position of the target is b. Because b is on left of midline tan (b)= - t an (a ) and distance computed by formula D = h(tan(b)) yields negative distance D'. In this case Ibl = lal so D'= -(D). Situa- tion is paradoxical because negative distance implies that target is behind the locust, but the target's angular position clearly speci- fies a position in front of the locust
A
O w
O
O O .J lu >
275
a g 225
175
125
75 -40
275
225
175
125
75 -40
A
| | I | -
-20
-20
J51
�9 . | . . |
0 20 40
�9 , ! . . !
20 40
275
225
175 II ~
125
7 5 " " m " "
-40 -20 275
225
125
75 . . i . . -40 -20
J39
i �9 . | .
20
J40
i � 9
mll
�9 i
40
�9 i , , i
20 40
T A R G E T D I S T A N C E (CM)
Fig. 6. Jump velocity vs target distance for locusts jumping at sta- tionary targets (filled circles) and locusts jumping at moving targets specifying paradoxical parallax (open triangles). Symbols: average of 3 jumps per target distance condition (except J51, 7 jumps/each distance). Target distance varied randomly. Error bars: _+ 1 SEM
Monocular locusts
In theory, one eye is sufficient for the computation of distance from motion parallax�9 Locusts have two eyes and so it is natural to ask how motion parallax signals from the two eyes combine in the determination of the distance estimate�9 Wallace (1959) tested animals with all
290
W-~ 190
0
90 o
o .A 290 W >
--~ 190
90 0
6 J 5 2
2.!, | ! !
10 20 30
10 20 30
TARGET
290 J29
iQ0 I/~ ii
90 ' ' 40 10 20
~ 290 J 2 7
&
190 'A
�9 �9 ! 90 40 10 20
DISTANCE (CM)
i i
30 40
i i
30 40
Fig. 7. Jump velocity as function of target distance in monocular locusts. Animal J52 had the front of its left eye occluded, while animals J20, J27, J29 had the front of their right eyes occluded. Filled circles: baseline data for intact (binocular) locusts. Open triangles: data from monocular conditions. All symbols represent mean of 3jumps at each target distance, error bars: ___1 SEM. Solid line: jump velocity expected from the hypothesis that locusts sum or average information from both eyes. These curves were generated by shifting data points from the intact locusts (filled circles) to the left by one half of the target distance, and then interpolating a line between the shifted points
or part of one eye covered. Like intact locusts these animals successfully chose the nearer of two targets (of the same apparent size), suggesting that monocular lo- custs have distance perception�9
To investigate the performance of monocular locusts more quantitatively, locusts were tested with both eyes to establish baseline data as in Fig. 4, and then were tested with one eye occluded. Locusts were prepared with either black opaque wax (beeswax, rosin and black enamel paint) or black opaque epoxy covering the ante- rior half of one eye. To avoid affecting counterrotation, the lateral retina was left unobstructed. Collett (1978) demonstrated that during peering the lateral retina is involved in maintaining accurate counter-rotation of the head against the arc through which the body moves�9 Counter-rotation ensures that the line of sight is always perpendicular to translational head movement�9 This be- havior prevents the additional image motion which re- sults from rotation of the line of sight and which would confound the measurement of motion parallax�9
The results are shown in Fig. 7. Although jump ve- locity still increased with target distance, locusts jumped with a greater velocity to targets at a given distance when one eye was occluded. Because greater jump veloci- ty implies a greater estimate of distance, and greater distance gives rise to less motion parallax, this observa- tion is consistent with the hypothesis that locusts per- ceive less motion parallax with only one eye, and that ordinarily the motion parallax observed in each eye is

E.C. Sobeh Motion parallax in locusts
combined (perhaps summed or averaged). The jump ve- locities expected under the hypothesis that locusts sum or average the mot ion paral lax seen by each eye is de- picted by the solid lines in Fig. 7. These curves were generated by re-plotting the data f rom intact locusts, as if the target had been presented at half of the actual target distance. The data f rom the monocular condition fall near these curves suggesting that monocular locusts estimate targets to be twice as distant as they appear with both eyes.
The possibility that increased j ump velocity under the monocular condition was caused by increased arous- al, due to the procedure of occluding one eye, was exam- ined by allowing the animal additional time (1 day) to habituate to the monocular condition. The additional time did not abolish the effect. The possibility remains, however, that the monocular condition leads to in- creased arousal during testing. To address these issues and to more quantitatively examine the interaction be- tween the eyes it is necessary to present different mot ion parallax stimuli to each eye simultaneously perhaps by using polarizing filters.
Binocular interactions of mot ion parallax signals have also been suggested by Cloarec (1986) in the water stick and Eriksson (1985) in the Australian bulldog ant. The summat ion or averaging of parallax signals f rom the two eyes may increase the locust 's sensitivity to the small amounts of mot ion it normally encounters and extend the range of distance perception.
585
The first strategy only requires measurement of im- age motion. A locust could equate a specific amount of image mot ion with a particular distance, making pre- cise knowledge of head position (or velocity) unneces- sary. I f the locust used this strategy one would expect the locust to peer with the same ampli tude (or velocity) independent of target distance. In addition j ump velocity should depend on the amount of image motion.
The second strategy only requires measurement of the amount of head motion. This greatly simplifies the computa t ion of distance f rom mot ion parallax because the locust need only be sensitive to a threshold amount of image mot ion on its retina - eliminating the need to quantitatively measure the image displacement (or velocity). I f the locust used this strategy one would ex- pect peer amplitude (or velocity) to increase linearly with distance and the locust 's jump velocity to be relatively independent of the amount of image mot ion it sees.
Alternatively, using the third strategy the locust would need to measure the amount of mot ion on its retina and combine this with information concerning its own mot ion (by efference copy or re-afference). One can formalize this relationship as follows
O • " 1.4 P 3 6 1.4
u,I a
Does the locust combine both head and image motion when ~ o.g computing distance ? <
a , .
t u
The amount of mot ion parallax seen by an observer ,."' is a function of target distance and the extent o f observer movement . I f the extent of observer movement is not = 0.4
0 specified it is impossible to determine the distance which gave rise to the perceived mot ion parallax. To use mo- o tion parallax for the estimation of distance an observer ~ o.o must either, take into account the extent o f his move- __.
,.I ment, or assume a constant extent of movement . ,~ 0.7
Motion paral lax decreases linearly with increasing iv,
target distance. Consequently, if a locust peers the same ~ 0.5 amount for targets at all distances the amount of mot ion *- paral lax it sees will decrease linearly with distance. It follows that if a locust increases its peering linearly with ~ ~ target distance it will see the same amount of mot ion parallax for all target distances.
These considerations suggest that there are 3 broad strategies a locust might use to compute distance f rom mot ion parallax, 1) it could always move its head a stan- dard amount and only measure image displacement (or image velocity), 2) it could move so that the target image is always displaced by a s tandard amount and then mea- sure the amount of head mot ion required to achieve that displacement (or a similar strategy using image ve- locity instead of image displacement), or 3) it could use neither a standard amount of head motion, nor a stan- dard amount of image motion, but instead measure both quantities and scale image mot ion by the extent o f head motion.
�9 ~ �9 o.g
~ = = 0.4 10 2 0 3 0 4 0 0
P 3 6
0.g
i ~ i 0.7
0.5
, I , I ' ' 0.3 10 2 0 3 0 4 0 0
P3g
! I I I m I
10 2 0 3 0 4 0
P 3 9
| i ! i
1 0 2 0 3 0 4 0
TARGET DISTANCE (CM)
Fig. 8. Average and maximum peer amplitude of a bout of peering as a function of target distance. Peer amplitude measured from locusts presented with targets at different distances. Target distance varied randomly between trials. Peer amplitude measured by track- ing the locust's head position with a video tracking system as de- scribed in Methods. The average or maximum amplitude of a bout of peers (3-12 peers per bout) was recorded as the peer amplitude for that trial A trial was defined as the bout of peering immediately preceding a jump. Symbols represent average of 3 trials (i.e. mean of maxima of 3 trials or the mean of means of 3 trials). Error bars, + 1 SEM. Both average and maximum data are provided because the locust increases peer amplitude and velocity over a bout of peers. Consequently, it is possible that the locust uses information from all peers in a bout of peering or just information from the largest/fastest peer
586 E.C. Sobel: Motion parallax in locusts
9 ir
,.'5,
o.9. >-
o 9 r
O
1.4 P36 t} P39 n O J46
J "~ '"t " " 1 2 5 [ �9 �9 125 [ I
. . , . . ,�9 20 30 40 ~ ; 3 5 ' ; ; ' ; ' ; ' ;
0.8 ~ P39 >'~'1 J48 J47
: [ ] _ 4 P o �9
0.4 lira ~ � 9
0.2 ~ ' ' ' 0.2 = , , , 75 ' I , ! 75 ' I , ! 0 10 20 30 40 10 20 30 40 3 5 7 3 5 7
AVG IMAGE DISPLACEMENT (DEGREES) TARGET DISTANCE (CM)
Fig. 9. Same as in Fig. 8, but using average and maximum peer velocity rather than peer amplitude
Eq. (2)
D = h/tan(a);
B 325
275
"-" 225 o u.i 0~ 175
,~ 125 >- I-- 75
0 325 --I g.I ~> 275 O.
225 --i
175
125
325 J49
275 [ ]
[ ] D I ~ I ~ 225
175
D = distance estimate; h = head position; a = angle to the target.
Collett 0978) found the amplitude of peering to in- crease slightly with target distance. The automated tracking system used here allowed straightforward mea- surement of the effect of target distance on peer velocity as well as peer amplitude. The results are shown in Figs. 8 and 9. In agreement with Collett (1978) peer am- plitude was observed to increase slightly with distance. In addition, peer velocity was also observed to increase slightly with distance. Both increased over the course of several peers within a bout of peering. Since the mo- tion parallax created by a fixed amount of lateral move- ment decreases with distance and peer amplitude and peer velocity increase with distance (Figs. 8 and 9) there are likely to be situations in which identical mot ion on the locust 's retina arises f rom targets at different dis- tances. I f the locust judges distance purely on the basis of image mot ion and assumes that it always peers a s tandard amount it should be unable to disambiguate these situations. Jump velocity should be predicted more accurately by image displacement or image velocity, than by target distance. On the other hand, if locusts use the amount of head mot ion in their computat ion of dis- tance, they should jump according to the target distance.
To address this question I investigated the relation- ship between the locust 's j ump velocity (a measure of its distance estimate) and image motion on its retina. These experiments involved up to 20 jumps to targets at each of two similar distances (8 cm and 10 cm or 8 cm and 12 cm). Image displacement and image velocity were obtained f rom peer amplitude (or peer velocity) using the following expression:
�9 ,25
I I , I 75 3 5 7
J47 325
~ 225
y �9 [ ] 175
125
! 75 7
[ ] [ ] J46
�9 - ee
�9 Q
i I i I i I
3 5 7
J48
275
o[] t~
"'4: 75 I I = I I ,
3 5 3 5
AVG IMAGE VELOCITY (DEGREES/SEC)
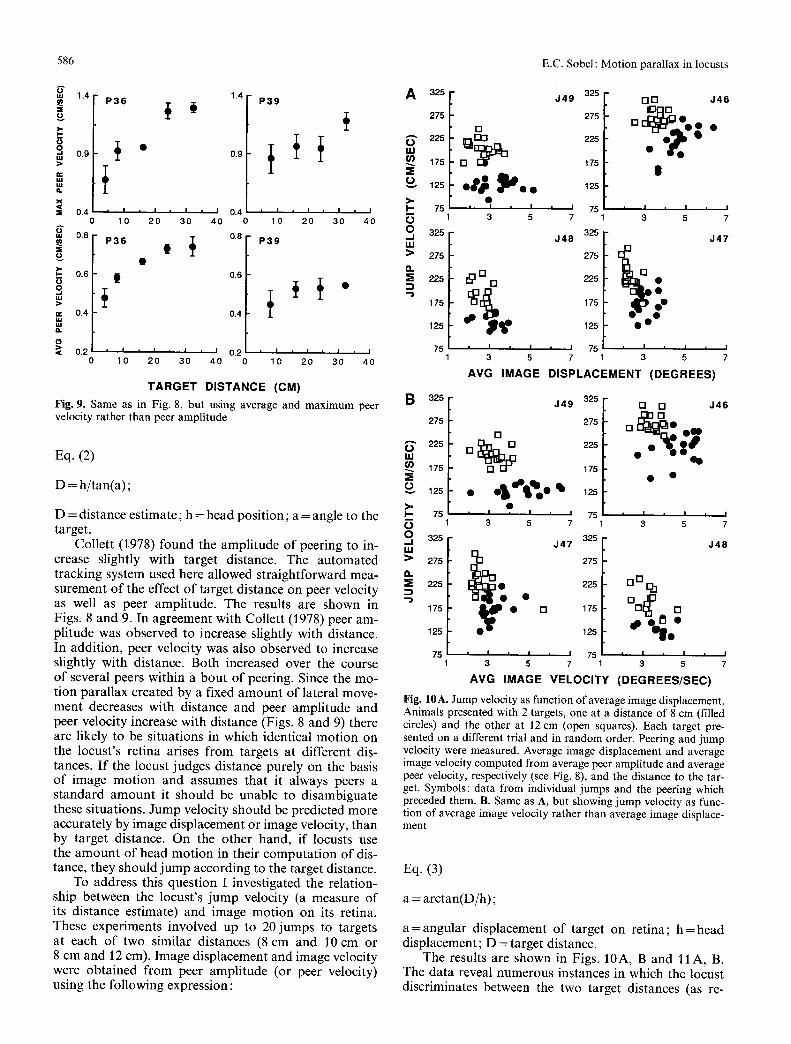
Fig. 10A. Jump velocity as function of average image displacement. Animals presented with 2 targets, one at a distance of 8 cm (filled circles) and the other at 12 cm (open squares). Each target pre- sented on a different trial and in random order. Peering and jump velocity were measured. Average image displacement and average image velocity computed from average peer amplitude and average peer velocity, respectively (see Fig. 8), and the distance to the tar- get. Symbols: data from individual jumps and the peering which preceded them. B. Same as A, but showing jump velocity as func- tion of average image velocity rather than average image displace- ment
Eq. (3)
a = arctan(D/h);
a = angular displacement of target on retina; h = head displacement; D = target distance.
The results are shown in Figs. 10A, B and 11A, B. The data reveal numerous instances in which the locust discriminates between the two target distances (as re-

E.C. Sobel : Motion parallax in locusts 587
A 325 J 4 9 325 ~ J 4 6
275 275 i~~ D - -
2a5 ~ 225 O . l ~ " " , ~ ca eP � 9 �9
175 I-II~ 175 �9
12s N I P ~ o �9 12s
~1 ~ 75 0 ' 0 ; �9 10 " 1=5 " 2 0 75 0 ; " 110 " 15 ' 210
325[ 325[ . ,8 = > 2,51 [] [] 2,5
q~dJe
,25 1 � 9149 ~25 1 e l i ~ 75 l . = , t . t . , 75 = ' t , J , ~ . ,
0 5 10 15 2 0 0 5 10 15 2 0
MAX IMAGE DISPLACEMENT (DEGREES)
a 325 r J , 9 325 ' [ ] J , 6
27s I- 275 ~ _ _ eg r [] . ~ �9
�9
175 l- Jill-- 175 ' �9
,,-I, 325 F . , , 7 325 J , 8
~ ' 275 ~ [ ] [ ] 275 I 1 " 1
2 6 10 14 2 6 10 14
MAX IMAGE VELOCITY (DEGREES/SEC)
Fig. l l A. Jump velocity as function of maximum image displace- ment. Data from same experiments as data in Fig. 10A, B. Here jump velocity is plotted against maximum image displacement rath- er than average image displacement (see Fig. 9). Maximum image displacement computed by using only the largest peer in a bout of peering and the distance to the target. It is necessary to look at both average and maximum values because locusts increase am- plitude of their peers over a bout of peering and thus it is uncertain whether average image displacement or maximum image displace- ment is used by the locust to estimate distance. However, this alternative method of analysis yields similar results to using aver- aged measures of these same quantities as in Fig. 10A, B. B. Same as A, but showing jump velocity as function of maximum image velocity rather than maximum image displacement
vea led by d i spa ra t e j u m p veloci t ies to the two targets) desp i te s imi la r image d i sp lacemen t s or image velocit ies . A n analys is o f cova r i ance app l i ed to these d a t a revea led a s igni f icant effect ( P < 0 . 0 1 ) o f t a rge t d i s tance on j u m p veloci ty , despi te a lack o f co r r e l a t i on be tween ta rge t dis- t ance a n d the a m o u n t o f image d i sp l ace me n t or veloci ty.
Consequen t ly , the locus t d i sc r imina tes be tween dif- ferent t a rge t d i s tances even when they give rise to ident i - cal a m o u n t s o f image d i sp l a c e me n t (or veloci ty) . To ac- compl i sh this the locus t m u s t inc lude i n f o r m a t i o n con- cern ing its own m o v e m e n t in its c o m p u t a t i o n o f d i s t ance f rom m o t i o n pa ra l l ax . W h e t h e r the locus t combines b o t h se l f -mot ion a n d image m o t i o n ( th i rd s t ra tegy above) o r s imply uses its se l f -mot ion (second s t ra tegy above) re- m a i n s to be resolved.
In conclus ion , locusts c lear ly do use m o t i o n p a r a l l a x to c o m p u t e d i s tance and do so in an in t r igu ing ly s imple m a n n e r - ignor ing the re la t ive d i rec t ions o f image a n d head mot ion . They a p p e a r to c o m b i n e i n f o r m a t i o n f rom each eye and to inc lude in thei r c o m p u t a t i o n o f d i s tance the extent o f la te ra l head mot ion . To pu r sue the phys io - logical basis o f the c o m p u t a t i o n o f d i s tance f rom m o t i o n pa ra l l ax , fu r ther be ha v io r a l a n d e l ec t rophys io log ica l ex- pe r imen t s are necessary . D i s t ance pe rcep t ion by m o t i o n pa ra l l ax in the locus t is an a t t r ac t ive p a r a d i g m in which to pu r sue neura l c o m p u t a t i o n , due to the fact t ha t the c o m p u t a t i o n is well def ined and the b e h a v i o r is easi ly measured .
Acknowledgements. I am greatly indebted to Dr. C.R. Gallistel for his support and guidance. I also thank Dr. T. Abrams for providing locusts and invaluable advice. Special thanks to Dr. T.S. Collett for advice and comments regarding these experiments and the preparation of this manuscript. During this work I was sup- ported by a predoctoral training grant from the National Institutes of Health (MH17168). Support for equipment and materials was provided through the Bernard L. and Ida E. Grossman Term Pro- fessorship of C.R. Gallistel.
References
Arbas EA (1983) Aerial manoeuvring reflexes in flightless grass- hoppers. J Exp Biol 107:509-513
Bennet-Clark HC (1975) The energetics of the jump the locust Schistocerca gregaria. J Exp Biol 82:105-121
Bennet-Clark HC, Alder GM (1979) The effect of air resistance on jumping performance in insects. J Exp Biol 63 : 53-83
Cartwright BA, Collett TS (1979) How honey-bees know their dis- tance from a near-by visual landmark. J Exp Biol 82 : 367-372
Cheng K, Collett TS, Pickhard A, Wehner R (1987) The use of visual landmarks by honeybees: bees weight landmarks accord- ing to their distance from the goal. J Comp Physiol A 161:469- 475
Cloarec A (1986) Distance and size discrimination in a water stick insect, Ranatra linearis (Heteroptera). J Exp Biol 120:59-77
Collett TS (1978) Peering - a locust behaviour pattern for obtaining motion parallax information. J Exp Biol 76:237-241
Collett TS (1988) How ladybirds approach nearby stalks: a study of visual selectivity and attention. J Comp Physiol A 163:355- 363
Collett TS, Harkness LIK (1982) Depth vision in animals. In: Ingle D J, Goodale MA, Mansfield RJW (eds) Analysis of Visual Behavior. Cambridge, MIT Press, pp 111-176
Collett TS, Land MF (1975) Visual control of flight behaviour in the hoverfly Syritta pipiens L. J Comp Physiol 99 : 1-66
Collett TS, Land MF (1978) How hoverflies compute interception courses. J Comp Physiol 125:191-204
Cooter RJ (1973) Flight and landing posture in hoppers of Schisto- cerca gregaria (Forsk). Acrida 2:307-317
Eriksson ES (1985) Attack behaviour and distance perception in the australian bulldog ant Myrmecia nigriceps. J Exp Biol 119:115-131

588 E.C. Sobel: Motion parallax in locusts
Goulet M, Campan R, Lambin M (1981) The visual perception of relative distances in the wood cricket, Nemobius sylvestris. Physiological Entomology 6:357-367
Heitler WJ, Burrows M (1977a) The locust jump. I. The motor programme. J Exp Biol 66:203-219
Heitler W J, Burrows M (1977 b) The locust jump. II. Neural circuits of the motor programme. J Exp Biol 66:221-2241
Kirchner WH, Srinivasan MV (1989) Freely flying honeybees use image motion to estimate object distance. Naturwissenschaften 76: 281-282
Lehrer M, Srinivasan MV, Zhang SW, Horridge GA (1988) Motion cues provide the bee's visual world with a third dimension. Nature 332:356-357
Maldonado H, Rodriguez E (1972) Depth perception in the praying mantis. Physiol Behav 8:751-759
Srinivasan MV, Lehrer M, Zhang SW, Horridge GA (1989) How honeybees measure their distance from objects of unknown size. J Comp Physiol A 165:605-613
Rossel S (1983) Binocular stereopsis in an insect. Nature 302:821- 822
Rossel S (1986) Binocular spatial localization in the praying mantis. J Exp Biol 120:265-281
Wallace GK (1958a) Visual perception and behavior in the desert locust. Anim Behav 6:242-243
Wallace GK (1958b) Some experiments on form perception in the nymphs of the desert locust Schistocerca gregaria Forsk~l. J Exp Biol 35:765-775
Wallace GK (1959) Visual scanning in the desert locust Schistocerca gregaria Forskgd. J Exp Biol 36:512-525
Wehner R (1981) Spatial vision in arthropods. In: Autrum H (ed) Comparative physiology and evolution of vision in inverte- brates. (Handbook of Sensory Physiology, vol VII/6c). Sprin- ger, Berlin Heidelberg New York, pp 287-616
Wehner R, Horn E (1975) The effect of object distance on pattern preferences in the walking fly, Drosophila melanogaster. Exper- ientia 31:641-643




















