
The Helmintholoafcal Society
160
Volume 36 January 1969 /Nurrtbef V PROCEEDINGS The Helmintholoafcal Society -•. "- ' • "'x'"' '""•• ' ' ; ' • ' ' '' : ' : -' ; ' ' on . x x L V ^ semiannual /ouriid/V-of rfesearcli< devoted fo : : ^ / ^vHe/minflio/dgy and dlf branches of Parasifo/ogy ' " " .Supported ir^paft by the \ ''••'•• { -^. n H.,Rahspm MeiiiorJal Try^t Fu,ricl /\.^'-. /'/ :^tt^ Subscription ^7/OQ<ci Volume; Fpr9Jgri / l$7.50l ^ • CONTENTS . - -. r V<'-" ; "-'^\-.-' ^|^f%S ;ABRAM, JAMES B. , Some Gastrointestinal' Helminths of Ondatra zihethicus , . ,; .Linnaeus, tlie Muskrat in Maryland -,...--------:_-..--..--.--.._..-A----->------l-- ---.---.rr -CABLE, ; R. MJ, ANDHADAR ISSEROFF^ VA Protandrotis Uaplopqrid Cercaria, r , Probably Jthe Larva of SaccocQelid.ides sogandanqsi Lumsden, 1963 1..^. COI.GLAZIER, M. L., K. C. KATES/kNb F, D. ENZIE. \. Activity of , ,'V-Tetfainisole, Thiabendazble, and Purified Fin^ Particle Thenothiazine, .-.':Against Experimental/Infections of Haernojichus contorUis--JanA Triclio- stroiigylus Speeies in Sheep ,Liivir— .:-—1^ l-.,r.__...-.i-.J.«__:.: -4^212 CROLL, N./A^ ANDLTD, R. \ViGLjERcmo. Osmpregulatioh arid the Uptilce pf > ^/. Ions in a Marine Nematode ..^^.^.: .... ::..„-.-../ ...... ^ ^.,^...^..' r ' r -l.^ ..... L~^.'.. DORAN, DAVID J., AND JO&N M.^VEiTteRriiNG. Infectiyity of Two Species of ' Poultry Coccidia After Freezing, arid Storage in. Liquid JNfitrogen Vapor\:l._- DORAN;) DAVID ,J.y AND JOHN M. VErrteRirNTG. Influence of Storage Period;on; /' Excystation; and i Development in Cell Culture bf Sporozoites of Eimeria - fneleOfgrimitis Tyzzeir, 1929^. -.^--^—^.. .--l.u--.-Z2 ^_ii.r.—.... ^..^.-:_^lL-. ;/; "';"^ v v ; ; 'K r- - / -'(Continued on BackCover),' - J- '.: .^,;/ \ ... .r -''/:> ,93 68 \; : ; •;"-!' • r J r30 '...-' ••£ 'f /,,;- - X; K '•#• i'l Vx.: -;^: Copyright © 2011, The Helminthological Society of Washington
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of The Helmintholoafcal Society

Volume 36 January 1969 /Nurrtbef V
PROCEEDINGS
The Helmintholoafcal Society- • . " - ' • "'x'"' ' " " • • ' ' ; ' • ' ' '' : ' : - ' ;' '
on.
x x L V ^ semiannual /ouriid/V-of rfesearcli< devoted fo: : ^ / ^vHe/minflio/dgy and dlf branches of Parasifo/ogy' " "
.Supported ir^paft by the \ ' ' • • ' • • { -^.n H.,Rahspm MeiiiorJal Try^t Fu,ricl /\.^'-. /'/
:^tt^Subscription 7/OQ<ci Volume; Fpr9Jgri/l$7.50l ^ •
CONTENTS
. - -. r V<'-" ;"- '^\-.- '
| f%S
;ABRAM, JAMES B. , Some Gastrointestinal' Helminths of Ondatra zihethicus, . ,; .Linnaeus, tlie Muskrat in Maryland -,...--------:_-..--..--.--.._..-A----->------l-- ---.---.rr-CABLE, ;R. MJ, AND HADAR ISSEROFF^ VA Protandrotis Uaplopqrid Cercaria,
r , Probably Jthe Larva of SaccocQelid.ides sogandanqsi Lumsden, 1963 1.. .COI.GLAZIER, M. L., K. C. KATES/kNb F, D. ENZIE. \. Activity of
, ,'V-Tetfainisole, Thiabendazble, and Purified Fin^ Particle Thenothiazine,.-.':Against Experimental/Infections of Haernojichus contorUis--JanA Triclio-
stroiigylus Speeies in Sheep ,Liivir— .:-—1 l-.,r.__...-.i-.J.«__:.: -4 212CROLL, N./A^ ANDLTD, R. \ViGLjERcmo. Osmpregulatioh arid the Uptilce pf
> ^/. Ions in a Marine Nematode ..^^.^.: ....::..„-.-../...... ., ... ..'r'r-l. .....L~ .'..DORAN, DAVID J., AND JO&N M.^VEiTteRriiNG. Infectiyity of Two Species of '
Poultry Coccidia After Freezing, arid Storage in. Liquid JNfitrogen Vapor\:l._-DORAN;) DAVID ,J.y AND JOHN M. VErrteRirNTG. Influence of Storage Period;on;
/' Excystation; and i Development in Cell Culture bf Sporozoites of Eimeria- fneleOfgrimitis Tyzzeir, 1929 . -.^--^—^.. .--l.u--.-Z2 ^_ii.r.—.... .. .-:_ lL-.
;/; "';"^ vv ; ; ' K r- - / -'(Continued on BackCover),' - J- '.: .^,;/ \ ... .r -''/:>
,93
68\;:;•;"-!'
• r Jr30
'...-' ••£' f /,,;-
- X ; K'•#•
i'l Vx.: -;^:
Copyright © 2011, The Helminthological Society of Washington

• i THE HEI INTHOLOGJCAL SOCIETY OF WASHINGTON : ^' ' ".' ''S ' •' ' ' -~ ' ' - J ' '' / ' ' • . ' '~~L.' ' ' "'• -~~' "I •-•' -'""' • ' ''I *'_- .'' - ' "• ^ '- / i
- ; ' . ' . ; " r ' • ••• ;• THE SOCIETY meets once a mpriUi from October dirough May for the presentation, andT ^/ •;' discussion'pf papers in .any \and all branches of parasitology or "related .sciences. All .interested
f /_ r persons are invited to attend. ; J ' //' "/' . ' • . " " ' - " - , ' ; / :\,
x ; ''Persons interested^in inembership in die HelrriintliQlogical Society of.;Was3bin"gton.i may obtainVir vapplicatibn blanks from the Gorresponding gecretalry-Treasurer^ Miss Edna ,/M. Bulirer, Belts--/, N '; ville Parasitplogidal Laboratory, Agricultural Research Center, Beltsville, Maryland, 20705.. Ai ^ year's subscription to,, die Proceedings is^included in die1 annual dues ($6:00).v ;"/ --/
OFFICERS OF7THE SOCIETY FOR
Ptesident:^ALA.N C. PIPKIN : — V .; . .f r , .-;'. *+. • V .. '<£ , . ; . .1.^Vice President: A. JAMES HALEY . *, ), ' • . ; • . ^V*:'-•'•;:M;.;,;,v; ! -yCorresponding Seer•etary-Treamrer: EDNA M; BUHRER -^ ? ^--\1 -. - '/1.Associate Treasurer; LLOYD E. ROZEBOOM ;" ;f i ;" ^/"
:Assistant Correspondin^Seeretanj-Treasiirer:, HALSEY H. VEGQRS {. \ ;r ;
Recording Secretary: E, J>>L;i SOUL'SBY ' - >v ^ ; - >* ^ ; " -V :'Librarian: JUDIT^ M. HUMPHREY .(1962- ) ,' x- "^ \:\4 ^ 1'; :;Afc/iiui^:,;WXLLARD W/BECKLUND (1967- ) ^ ] .V^'--'^/^Representative to the Washington Academy of Sciences: AtlREL O.~F9§TER (1965- , ),Representative to the American Society \of Parasitologists: i ! ^ ' l ; >C , ' :\..^-.;....-,...-' r '; , M, V , GEORGE ^l LUTTERMOSER (1969-Execittive'Gomniittee Members-at-Large: HARLEY^ G. SHEFFIELE^ 1869 ';"A." ', '/!
" \ v C - . < ^ : / s, ',-;"' / .-'. ' "'.' 'r' GILBERT F.-OT^TO, 1970 -'gv. / . ' ^ v ^ ^ - '
"THE PROCEEDINGS <OF THE HELMINTH0LOGICAL OF WASHINGTON. _ • - ^ _ , . .- - : . . s - : - • - - • - . • - .
THE PROCEEDINGS are published semiannually at Lawrence, /.Kansas by die Helmintho-Ipgical 'Society of Washington, "".papers, need not: be '.presented at a meeting to be /published/in'jthe .Proceedirigs. / However, non-members may publish in the Proceedings /only , ifContribute die full cost pf pi\blicatiohv '\-.\ - , ' , « • •
should be sent:te the EjDITOR, /Francis \Gk Tromba.; Beltsville Parasito-logic(al Labordtory-, Agricultural Research Center, Belfsville, Maryland 20705. Manuscriptsmust be typewritten, double i spaced, apd un Nf inished form. Only the, ribbon /copy will /beaccepted for publicatiori; ; it is hce'epted w,ith die understanding that it 'ill be published only
'in the Proceedings, . . \ " ; '-. , • ' - - ' ' / - ' y, . • ; . ' . /^ ;. ',/ ' • ;, '!• \:\ V-: ' • " ' - . / •• i
- REPRlMTS may (be Ordered from the 'PRINTER J at 'die same' time, the corrected proofis retiihiedj'io/die EMTOR./" , ' I ^. ; ^ ^'"'^ - • '\:^^^ "'•'"/• . ! • - ( " ' -- /BACK VOLUMES of the Proceedings are .available; /Inquiries /concerning back volumesand current isubscriptions, .should be directed to: Hehri^thplogical Society, of "Washington,c^p Allen Press, -Inc., .041 Newjilampshire St., Lawrence, 'I|ansas ;66044, U;S.A. . ;
BUSINESS OFFICE. The Sopety^s 'business office is at ; , Lawrence,. Kansas'. 'All/inquiries^concerning subscriptions , or back issues and -all /payments. "for dues, subscriptions, and bapk
, issues should be addressed vto- Hejniindiplogical Society ' of 'Wasliihgton^ c/o Allen-Press, Inc.,/1041 New Hampshire St., tawrencef Kansas 66044, '. tl.'SiA. ',-: ' ^ , /-' : , , - „ /
' ' ' ' ' ' ' '' • • '- ' •' s - '
^PITORIAL! BOARD> ^ I
FRANCIS G. TROMBA/ Editor^
WILBUR L. BULLOCKMAY BELLE ,CHITWOODJACOB H. FISCHTHAL"WILLIAM J. HARGIS, JR.:GLENN L. :HOFFMAN/ l"i
, LOREN R. KRUSBERG VJOHN T.;LUCKER -" )' /
-ALLEN iMcINTOSH ' V,:/XAWILLIAM R. NICKLE i.GILBERT F. OTTO :\Y J. RASKI: s i '
HARLEY G^ SHEFFIELDARISEN .C.^IRJAN-" £.!.PAUL P. WEINSTEIN .-
Copyright © 2011, The Helminthological Society of Washington

PROCEEDINGS OF THEHELMINTHOLOGICAL SOCIETY OF WASHINGTON
VOLUME 36 JANUARY 1969 NUMBER 1
Osmoregulation and the Uptake of Ions in a Marine Nematode
N. A. CROLL1 AND D. R. VlGLIERCHIODepartment of Nematology, University of California, Davis, California
Past studies have demonstrated that if cer-tain animal-parasitic, plant-parasitic and free-living nematodes are placed in hypertonicsolutions they lose water causing a reductionin volume (Stephenson, 1942; Osche, 1952;Lee, 1960; Wallace and Greet, 1964; Anya,1966; and Myers, 1966). In most cases osmo-regulation has then occurred enabling thenematodes to regain their original size. Sim-ilarly when placed in hypotonic media thereis an increase in volume and sometimes a re-turn to normal body size (Stephenson, 1942;Anya, 1966; and Myers, 1966). This com-munication complements these observations bycharacterizing osmoregulatioii and uptake ofions in the marine nematode—Deontostoma(= Thoracostoma) calif ornicum.
Reviewing the data on osmoregulatory mech-anisms in invertebrates, Beadle (1957) wasable to say that "There is no convincing evi-dence as yet that osmotic gradients betweenbody fluid and external medium are due inany degree to the active transport of water."It is, therefore, with the movements of ionsthat osmoregulation is achieved and the abilityto osmoregulate will depend on the ability totake up and lose ions.
Experimental Methods and Materials
Deontostoma calif ornicum was collectedfrom the holdfasts of Laminaria digitata andEgregria laevigata at Dillon Beach, California,at extreme low water spring tide. D. califor-nicum was removed from the holdfasts andstored in seawater on gauze at 5 C as this wasfound to reduce "clumping." The seawater waschanged at weekly intervals, and under these
1 Fulbright Scholar, permanent address: Department ofZoology and Applied Entomology, Imperial College, Uni-versity of London.
conditions the worms remained in an appar-ently good state for up to 6 weeks. All experi-ments were conducted on adult worms fromthis supply at 25 C.
Through water transfer nematodes becomegreatly reduced in size and distended in hyper-tonic and hypotonic solutions respectively. Thenematode response to osmotic solutions andosmoregulation was measurable by changes inits physical dimensions. The distance from theoesophago—intestinal junction to the anteriortip was found to be a convenient sensitiveparameter. In preliminary experimentation itwas found that this measurement was in directproportion to changes in width and overalllength.
Each worm was placed in a perspex groovecontaining seawater, held by a coverglass andmeasured with an ocular micrometer. Thenematodes then were placed in test solutionin 2 cm diameter glass cells in covered Petridishes. A little tap water was placed in eachPetri dish to ensure a high humidity whileallowing free gaseous exchange. The nema-todes became inactive or less active duringthe experiments.
Frequently, exosmosis proceeded until con-tortion and crumpling of the worms occurredand at these extremities of water loss, volumechanges were not truly reflected by lengthmeasurements. These excessive losses wereaccommodated within 15 min and the usualbody shape was recovered. The first reliablemeasurement of water loss (ME) was on therecovery curve R which approximated tolinearity in all but the two cases describedbelow. T was the time taken to regain original(seawater) size (Fig. 1). The rate of recoverywas expressed as ME/T or tan «°.
Copyright © 2011, The Helminthological Society of Washington

PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
PERCENTAGECHANGE INLENGTH
Table 1. Osmotic regulation of Deontostoma cali-fornicum following immersion in various hypertonicmedia.
Figure 1. Schematic diagram of the measure-ments made of the osmoregulatory process inDeontostoma californicum, following immersion inhypertonic media. ME, maximum recorded exos-mosis; T, recovery time; R, recovery line; tan a,rate of recovery.
Osmoregulation in hypotonic andhypertonic solutions of NaCl
Using distilled water, 0.4, 0.6, 0.8, and 1.0 Msolutions of NaCl, the length changes of D.californicum were measured with respect totime. The isotonicity of the worm was thusapproximated as well as its ability to recoverits seawater dimensions.
RESULTS: In both hypertonic and hypotonicsolutions there were immediate changes inlength (Fig. 2) . D. californicum was unableto osmoregulate in hypotonic solutions (Fig.2) even after 24 hr, but recovered its normallength and mobility in seawater. In hypertonicsolutions recovery occurred at an essentiallyconstant rate until the original length had beenreached; R was treated as being linear. Therefrequently occurred an "over-osmoregulation"giving a length larger (usually 5-10%) thanthe original seawater length (Figs. 2, 4) . Hav-
>
Compound
NaCl
NaBr*
Nal
NaNO2*
NaNO.,
Na2SO4
Na^O.,
Na2MoO4
NaWO,
KC1
KBr
Kl
K1O,K3Fe(CN)(.
K1Fe(CN)(.
NH(C1
LiCl
LiNO.,
SrCl2
SrBr2
Srls
Regulation-;- Compound Regulation'!'
+ MgCl2
+ CaCl2
+ KMnO,
-)- Oxamic acid(partial) (COOH-CO-NH2)
+ OxamideNH2-CO-CO-NH2)
— Formamide(CHO-NHg)
Dimethyl oxalate(CH.,-CO-CO-CH3)
— Ribose— Xylose
+ Sorbitol
-)- Inositol
-f- Galactose
O Glucose
— Fructose
— • Trehalose
+ Lactose
+ Maltose
+ Sucrose
+
+
+
+
Ooo
o
o
:—
(slight)
-
—
(slight)
Figure 2. Percentage length changes of D. cali-fornicum with respect to time in NaCl solutions ofvarying molarity.
::: Nonlinear recovery.t +, Recovery to original length; —, No recovery but
normal on return to seawater; Q, No recovery, dead onreturn to seawater.
ing overcompensated, a subsequent return tonormal size was never observed. Followingosmoregulation in a hypertonic solution andtransfer to distilled water, there was an imme-diate and gross increase in size which usuallyled to bursting of the worm. This degree ofincrease did not occur in changes from sea-water to distilled water.
Osmoregulation in other solutions
A comparison was made of the ability ofD. californicum to return to original seawaterlength following immersion in hypertonicsolutions of selected electrolytes and non-electrolytes.
RESULTS: The ability or inability of thenematodes to recover normal length in thehypertonic media, together with the return to
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969
Figure 3. Percentage length changes of D. cali-fornicum with respect to time to hypertonic solu-tions of NaBr and NaNO2, showing nonlinearrecovery.
normal activity in seawater following the treat-ment, is tabulated (Table I). D. californicumreturned to its original size, or increasedslightly in most electrolytes. The recoveryrates (tan «) differed, while the recovery lineR approximated to linearity except in thecases of NaBr and NaNO2 (Fig. 3).
Very slight recovery occurred in sucrose andgalactose and none was observed in any ofthe other sugars.
Oxamic acid, oxamide, formamide, dimethyloxalate, and potassium periodate were used toinvestigate the significance of spatial configura-tion and charge in the regulatory mechanism.Their low solubility, together with their appar-ently high toxicity, made reliable conclusionsimpossible.
In seawater those individuals that had osmo-regulated, expanded in size and frequentlyburst; all of the others (except the oxalic acidderivatives, formamide and potassium perio-date) regained their normal size and mobility.Vigorous activity occurred for up to 30 minfollowing transfer to sodium thiosulphate.
The influence of successive immersionsin different hypertonic solutions onosmoregrilation
After D. californicum had regulated in a 1molar solution of an electrolyte, it was im-mersed in another molar solution of a secondsalt. Salts were selected from those known toallow regulation (Table 1) in order to showany cationic or anionic specificity of the regu-latory mechanism.
Nematodes that had osmoregulated in 1 MNaCl were also transferred to 0.5 M NaCl and
SECOND SALT
, _ .50M NaCl// ,- .75M NaCl
Figure 4. Schematic diagram showing lengthchanges resulting from immersion in a second solu-tion following regulation in the first. A. Transferfrom 1 M solution of first electrolyte to 1 M solu-tion of a second electrolyte. B. Transfer from 1 MNaCl to 0.5 M or 0.75 M NaCl. (Arrow indicatesthe point of transfer.)
0.75 M NaCl and any changes in length weremeasured.
RESULTS: Every change to a molar solutionof the second salt, following regulation in thefirst, resulted in an additional increase in size(Fig. 4A), though'the increase "was subject tosome individual variation. The following pairsof 1 molar successive changes were made:
LiN03 -^ NaCl, NaNO3 -» NaCl,NaCl -* LiNO3, NaCl -> NaNO3.
D. californicum. that had osmoregulated in1 M NaCl also increased further when placedin 0.5 M or 0.75 M NaCl, the increase beinggreater with the more dilute solution (Fig. 4B).
The influence of sulphate on theregulatory mechanism
Since D. californicum was unable to regu-late in sulphate (Fig. 5), thiosulphate, molyb-date, and tungstate (Table 1), recovery wasestimated in various volume to volume hyper-
Copyright © 2011, The Helminthological Society of Washington

PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
v/v N . , S O . / H » C 1
Figure 5. Percentage length changes of D. cali-fornicum with respect to time in hypertonic solu-tions of the sulphate and nitrate salts of lithiumand sodium.
tonic mixtures of 1 M Na2SO4 and 1 M NaCl.The rate of regulation varied with different
v/v mixtures of Cl~ and SO4= (Fig. 6). In100% and 75% sulphate solutions there wasno recovery from ME (« = 0°). At 50:50mixtures there was slight osmoregulation, andthe rate of osmoregulation increased withgreater dilutions of sulphate. From 1% to 50%SO4= the rate of recovery against dilution plotfollowed closely an exponential relation.
The inhibitory effect of high SO.t" concen-trations was temporary and the worms re-covered normal size and motility on beingtransferred to seawater.
Flame photometric determination of ionsTo establish the uptake of ions during osmo-
regulation, groups of 10-12 D. californicumwere weighed (wet wt), dried to constantweight for 3 hr at 100 C and ashed at 600 Cfor 8 hr (ash wt) following treatment. Inaddition to controls taken directly from sea-water, other groups were exosmosed (X) for3 min in 1 M solutions of NaCl, KCl, andMgClo, respectively. The other treatment (R)was exosmosis and recovery in 1 M NaCl, KCl,and MgClo prior to weighing and ashing. Theconcentration of Na, K, and Mg were thenestimated with a flame photometer.
RESULTS: Flame photometric measurementsof ionic concentrations of K1, Na+, and Mg++
following regulation (R) in .1 M KCl, NaCl,and MgClo, respectively, showed increases inall cases. Increases in ionic concentrations also
0/100 55'
Figure 6. Percentage length changes of D. cali-fornicum with respect to time in various volume/volume mixtures of 1 M NaJSO.t and NaCl; showingthe dependence of angle « (Fig. 1) or the Na2SO4/NaCl ratio.
were found for worms exosmosed only ( X ) ,this apparently being due to adsorption or someother form of binding or trapping by the in-tegument. The increases in total cation follow-ing regulation in 1 M NaCl and MgCl2 approxi-mated to the calculated value for a volume of1 M solution equivalent to the volume of solu-tion taken up in the nematodes (Table 2,volume of free water R-X). Insufficient K+
ions entered to account for the total requiredincrement in osmotic pressure needed to attainisotonicity in 1 M KCl.
If the ionic concentration of the water loston drying of D. californicum (wet wt-dry wt100 C) is assumed equivalent to seawater forpurposes of comparison (it is not, of course,because of the hydration of tissues and organicsolutes, etc.), it appears that Mg++ and Na+
are present within the animal in concentra-tions of the same order present in seawater.Potassium ions, however, are present in muchhigher concentrations.
Localization of the regulatory mechanismFollowing regulation in hypertonic solutions,
D. californicum increased greatly in size andoften burst when transferred to distilled water.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969
Table 2. Distribution of ions in Deontostoma calif ornicum, exosmosed only (X), osmoregulated (R),and from seawater controls (C).
s
I1 la
PotassiumK+0 22.9K+x 23.4K>K 23.6
SodiumNa+c 22.9Na+x 23.1Na+jt 23.8
MagnesiumMg++c 22.9Mg++x 22.8Mg++R 23.8
M
1j
Q
37.538.638.5
37,537.938.9
37.538.038.2
c.2 a f
I1'
£^
2.738.62
11.30
44.1060.7078.30
6,3716.5040.40
o C£ to
13
18.918.922.3
18.914.014.7
18.911.014.9
tion
inw
ater
/tg
/ml
OS
9.72
457.00
53.40
M•1
•Sx•^£3 °
ll
1.3
0.4
1.5
0) 2 s *;2 S m^ y.o .« rt O
C bo c 0 C'+3.« -^ '^.2^^
a ff, la « S<^U" O ^ £ g
4.0 50.80
17.0 9.20
22.2 36.48
a c
:ion
tiss
ue w
atre
ase
equi
vale
ieaw
ater
/ig
U.S'o
0.495
4.200
1.950
Ligaturing experiments were conducted to iso-late those regions of the body important inthe osmoregulatory mechanism and in theuptake of ions. Using hypertonic 1 M NaCl,ligatures were tied with 7.0 Ethicon braidedsilk.
RESULTS: When ligatured in the middle ofthe body while still in seawater and placed in1 M NaCl, both anterior and posterior halvesof the body decreased in size. Both halvesbecame grossly distended on being returnedto distilled water. When ligatured behind thestoma and anterior to the anus, the nematodesbecame smaller in hypertonic media and fol-lowing osmoregulation burst when returned todistilled water.
Localization of the area of water uptakeThe ligaturing experiments indicated that
both osmoregulation and the uptake of wateroccurred through the body wall. After osmo-regulation for 30 min in 1 M NaCl (incompleterecovery), D. calif ornicum was placed in dilutevital stains to determine whether the entryof water (and with it the stain) was in anypores or canals. Cotton blue, eosin, congo red,janus green B, crystal violet, and methyleneblue were all used. The reverse process wasalso examined where stained worms wereexosmosed to see any pores or canals fromwhich the dyes diffused out.
RESULTS: Crystal violet (Gentian violet),eosin, and methylene blue entered best andcolored the entire body contents. No canalswere found and no pores were visible for waterpassage in or out of living worms.
The longitudinal movement ofwater in D. californicum
Worms from seawater were placed across avaseline barrier separating distilled water and1 M NaCl. This design contrasted with liga-turing as the environment was separated butthe movement of ions and water in the wormwas free and continuous through its wholelength.
The two halves of the worm behaved inde-pendently with respect to osmotic change.Although the stains entered the portion of theworm in distilled water, there was no indica-tion of the dye moving longitudinally towardthat part of the worm in 1 M NaCl. This mayhave been due to adsorption of the dyes ontomembranes preventing its free passage. Never-theless, an appreciable longitudinal move-ment of water could not be detected in ourtechniques.
DiscussionThe environment of all the animal parasitic
nematodes examined to date are hypertonic to
Copyright © 2011, The Helminthological Society of Washington

PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
the body fluids of the nematodes (Pannikarand Sproston, 1941; Lee, 1960; Rogers, 1962;Anya, 1966). It has been consistently demon-strated that these forms are able to osmoregu-late well in hypertonic media. They are, how-ever, less able or unable to regulate inhypotonic media.
In contrast the free-living Rhabditis terrestmin culture (Stephenson, 1942) and other Rhab-ditis species (Osche, 1952) and Panagrellusredivivus in water (Myers, 1966) inhabithypotonic habitats and regulate well in hypo-tonic media. Isotonicity for these is about0.15 M NaCl. Because of the apparent adap-tation of osmoregulation to habitat, it has beensuggested that marine nematodes would beunable to regulate their osmotic pressure(Rogers, 1962; von Brand, I960; Lee, 1965).Isotonicity of D. californicum. is, however,0.6 M NaCl, or about four times greater thanfor any nematode known. Many marine inver-tebrates have poor powers of osmoregulationbut D. californicum maintains an osmotic equi-librium in hypertonic media—a feature itshares with some animal parasitic forms.
These observations suggest that D. califor-nicum, is partially adapted to intertidal life andcan tolerate exposure and higher salinity. Thisnematode may be poikilosmotic as suggestedby Osche (1952) for free-living soil forms andRogers (1962) for some animal parasiticnematodes.
Beadle (1957) emphasized the importanceof ionic exchange arid uptake in the regulationof osmotic pressure and water content. Hobsonet al. (1952a, 1952b) showed that the bodyfluids of Ascaris lumbricoides changed in ioniccomposition following incubation in dilutedseawater. Myers (1966) reported evidence ofionic absorption in the osmoregulation of Pana-grellus redivivus and Aphelenchus avenae.Regulation of D. californicum relates to thesefindings in other nematodes, taking up ions inhypertonic solutions until osmotic equilibriumis reached with the environment. Bursting ofthe nematode upon re-entry in distilled water,together \vith the flame photometric demon-stration of an increased concentration of ionsfollowing regulation, supported the theory ofionic uptake. There was no suggestion of anoutward movement of ions either in hypotonicmedia or following over-osmoregulation in hy-pertonic solutions (Figs. 2, 4B).
Figure 7. The relationship between molecularweight and the rate of recovery (tan a); — joinscommon cations, joins common anions, = joinsthe chlorides of divalent cations.
It may be seen from Figure 7 that the dis-tribution of salts in a molecular weight againsttan a (rate of osmoregulation) plot is notrandom. When using the chloride, bromide,and iodide salts of Sr++, Na+, and K+, iodidesallowed the fastest rates of recovery, chloridesthe slowest, and both SrBr2 and KBr wereintermediary. The position of NaBr (Figs. 3, 7)cannot be explained, but von Brand (1943)showed it to be toxic to larval Eustrongylidesignotus. In a M wt X tan « plot (Fig. 7) thehalogen salts of each cation had slopes of thesame order. Approximately parallel lines joinrespectively: Nal and KI, NaBr and KBr, andthe chlorides of Li+, NH4+, Na+, and K+.
The anionic and cationic sequence in de-creasing order of preference in the regulatorymechanism is:
Anions I —» Br~ —» Cl~Cations K< -> Na1 -» NH4+ —» Li+
The Stoke's Law radii and ionic mobilities ofSr++ and Ca++ are practically identical andMg++ is very similar (Table 3), correspondingto the rates of regulation in the chlorides ofthese ions (Fig. 7). The ionic mobilities andStoke's Law radii of K+, Na+, and Cl+ also forma series reflecting the rates of regulation of D.californicum (Fig. 7). Having the lowest Mwt, Li+ has the highest charge density, greatesthydration, lowest molecular mobility and,therefore the highest Stoke's Law radius.
Of the anionic halogens the increasing orderof preference for regulation is Ch —> Br~ —> L
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969
Table 3. Ionic mobilities (\°) Stoke's Law Radii,and energy of hydration (~A HK cal mole"1) ofions at 25 C.
Ion
K+
Na+
Li+
Mg++
Ca++Sr++
I-Br-
ci-SO4=Fe(CN)03-Fe(CN),,''-
X°
73.50
50.1038.6053.059.559.476.878.1476.35
Stokes' r
1.251.842.393.473.103.101.201.181.212.302.763.35
-AHK-Kcalmole-1
- 83.37
-103.55
-129.67-473.29
-394.50
-359.22
- 61.8— 72.2- 80.3
Data after Noyes (1962) and Robinson and Stokes (1959).
(Fig. 7). These anions have low hydration,and Stoke's Law radii and their ionic mobilityare essentially the same (Table 3). The seriesCl~ —> Br~ —» I~ does, however, manifest de-creasing charge density and increasing polar-izability. This significance of this polarizabilityappears to be correlated with increasing easeof recovery. A model scheme for a possiblemechanism by which an anion could passthrough a "pore" in the membrane is illustrated(Fig. 8).
The results suggest that in hypertonic solu-tions of single salts, ions pass in by selectivediffusion until an equilibrium is reached forthat salt. The "distention" over normal size ina regulated animal may be explained in partby the osmotic effect of normal residual solublecomponents, over and above that due to theelectrolyte of hypertonicity. This is consistentwith the two-step distention of an animaltreated successively in two different salts. Ina second hypertonic salt, following equilibriumin the first, there is a second equilibrium givinga greater resultant osmotic pressure, and fur-ther increase in size.
The recovery line R (Fig. 1) is approxi-mately linear; the explanation for this maybe found by the interaction of two opposedfactors. The entry of ions by physical meanswould tend to be greatest at the onset of ionic-entry ME (Fig. 1) and gradually become re-duced. As ions enter and the worms dimen-sions increase, its surface area increases as a
Figure 8. Schematic model illustrating a pos-sible mechanism whereby polarizability of an anionfacilitates its passage through a charged membraneof the body integument. A, anion approachingcharged membrane; B and C show7 polar reversibil-ity on passage through the "pore"; D, completionof entry.
function of the square; there is a nonlinearincrease, therefore, in the area for ionic uptake.The resultant of the decreasing rate of diffusionand increasing the surface area tends to givea linear recovery line.
It is suggested that the entry of the cationmay be essentially dependent on differentialionic mobility, reflecting hydrated diametersand modified by the anion depending on itspolarizability. Their entry depends on a livingsystem possibly charged and has the char-acteristics of a one-way selective diffusion,there being no evidence for the outwardmovement of ions.
In attempting to explain the inability ofSO4= and other Group VI anions to enter, itwas not possible to distinguish between themolecular structure or the multiple charge.Monovalent anions of like spatial configura-tion; C1O4~, IO4~, MnO4~ and unlike configura-tion, oxalic acid derivatives, as well as poly-valent anions Fe(CN)G4^, Fe(CN)G3- wereeither toxic or insufficiently soluble for testing.
The penetration of extremely dilute solu-tions of eosin, methylene blue, and gentianviolet, would suggest that there is transfer ofdiverse ions and molecules through the integu-ment, but that it is of an order several timesremoved from that involved in osmotic effects.
The inability to osmoregulate in hypertonicsugar solutions may rest on the very limitedability of nonelectrolytes to cross the integu-ment (Mueller, 1929). The inability of GroupVI anions to enter is consistent with the resultsof Tweedie and Segel (1967) and Yamamotoand Segel (1966) on Penicillium chrysogenumand Aspergillus sp. Krogh (1939) found thatSO4= did not enter marine invertebrates well;and Robertson (1957) found that SO4= in
Copyright © 2011, The Helminthological Society of Washington

PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
many marine invertebrates was usually belowthe equilibrium value for seawater. The inhibi-tory effect is probably of little importance inthe ocean, where SO4~ ion in the Dillon Beachlocality are present in 2.64 parts per thousandat a salinity of 3.48% (Hedgpeth, 1964).
Nematode recovery can be visualized as in-volving several phases: (a) Passage of anionsand cations through a selectively permeablemembrane. The effect between the activity ofthe monovalent cation series and the mono-valent anion series may be in part a reflectionof manner of hydration. Since the water mole-cules around a cation extend their protonsoutward so that ions having a wide range ofsizes may assume configurations more adapt-able to the surrounding water structure whileanions require the hydrogen of water moleculesto be directed inward and as such, the con-figurations of the hydration spheres may bemore dependent on the central ion size, (b)The formation of an osmotically active bodysolution, made up in part by imbibition ofsalts, increased solubilization of organic sub-stances and an increase in bound water fromincreased tissue hydration such as may beinduced by or reflected in cation mobility.
In view of experimental observation it istempting to think that monovalent anions, de-pending upon their polarizabilities, control thepassage of salts across the semipermeablemembrane. The cations, however, are pri-marily involved in solubilization of organiccomponents and enhanced hydration of tissuesmanifested in increased rates of recovery ofbody lengths, according to the series of in-creased ionic mobilities of monovalent cations.
The accumulation of K+ and possibly theother salts suggested in the flame photometricanalyses, may be explained by the observedunidirectional movement inward of salts athigh tide and the partial loss of water throughdrying at low tide over long periods.
If this were the case then it would be neces-sary to postulate an equally slow elimination ofother cations Na+, Mg++ by metabolic meanseg. excretion, loss through the gut, so that thebalance with seawater was maintained.
The ligaturing experiments suggested thatthe uptake of ions occurred through the integu-ment just under the cuticle. It was also foundthat the process did not depend on localizedreceptors, amphids or caudal glands all of
which have at some time been implicated innematode osmoregulation. The gut was shownnot to play an essential role in ionic regulation,although it probably contributes in the naturalhabitat.
There is good evidence that in hookwormlarvae osmoregulation is restricted to the so-called excretory system as it is in flatworms(Weinstein, 1952 and 1960). This differenceis consistent with the broad postulate in biologythat marine organisms, which are usually iso-tonic with their surroundings, undergo ionicexchange and water loss over their entire sur-face. While with migration onto land or fresh-water or another nonisotonic environment,comes a reduction in the surface area of suchmembranes and a localization of osmoticallyactive surfaces.
SummaryDeontostoma californicum was able to re-
cover its osmotic pressure in hypertonic NaClbut not in hypotonic solutions. Osmoregulationwas not possible in hypertonic nonelectrolytesand in SO4=, and other Group VI anions. Inthe other electrolytes used, the rate of recoveryvaried with ionic mobility and energy ofhydration in cationic species with the degreeof polarizability in anionic species. A hypoth-esis is presented that relates the chemical andphysical characteristics of ions and the proper-ties of the integumental membrane to themechanism of ionic and osmotic regulation.
Using ligaturing experiments and aqueousdyes it was concluded that the uptake of ionsand entry of water occurred over the entirebody surface. Finally, our results have beencompared to observations on other free-livingand parasitic nematodes.
Literature CitedAnya, A. O. 1966. Investigation on osmotic
regulation in the parasitic nematode Aspi-culuris tetraptera Schulz. Parasitology 56:583-588.
Beadle, L. C. 1957. Comparative Physiology:Osmotic and ionic regulation in aquatic ani-mals. Ann. Rev. of Physiol. 20: 324-358.
Hedgpeth, J. W. 1964. Introduction to Sea-shore Life of the San Francisco and BayRegion and the Coast of Northern Califor-nia. University California Press, Berkeley.
Hobson, A. D., W. Stephenson, and L. C.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969
Beadle. 1952a. Studies on the Physiologyof Ascaris lumbricoides. I. The relation ofthe total osmotic pressure, conductivity andchloride content of the body fluid to that ofthe total environment. J. Exp. Biol. 29: 1-21.
Hobson, A. D., W. Stephenson, and A. Aden.1952b. Studies on the Physiology of Ascarislumbricoides. II. The inorganic compositionof the body fluid in relation to that of theexternal environment. J. Exp. Biol. 29: 22-29.
Krogh, H. 1939. Osmotic Regulation in Aqua-tic Animals. Cambridge University Press,London.
Lee, D. L. 1960. The effect of changes in theosmotic pressure upon Hammerschmidtielladiesingi (Hammerschmidt, 1938) with ref-erence to the survival of the nematodes duringmoulting of the cockroach. Parasitology 50:241-6.
. 1965. The Physiology of Nematodes.Oliver and Boyd, London.
Mueller, J. F. 1929. Studies on the micro-scopical anatomy and physiology of Ascarislumbricoides and Ascaris megalocephala. Z.Zellforsch 8: 361-403.
Myers, R. F. 1966. Osmoregulation in Pana-grellus redivivus and Aphelenchus avenue.Nematologica 12: 579-586.
Noyes, R. M. 1962. Thermodynamics of ionhydration as a measure of effective dielec-tric properties of water. J. Am. Chem. Soc.84: 513-522.
Osche, G. 1952. Die Bedeutung der Osmoreg-ulation und des Winkverhaltens fur freile-bende Nematoden. Z. Morph. w. Oekol.Tiere 41: 54-77.
Pannikar, N. K., and N. G. Sproston. 1941.Osmotic relations of some metazoan para-sites. Parasitology 33: 214-223.
Robertson, J. D. 1957. Osmotic and ionicregulation in aquatic invertebrates. RecentAdvances in Invertebrate Physiology. Uni-versity Oregon, Eugene.
Robinson, R. A., and R. H. Stokes. 1959.Electrolyte Solutions. The Measurement andInterpretation of Conductance, ChemicalPotential and Diffusion in Solutions of Sim-ple Electrolytes. Second Edition, Butter-worths, London.
Rogers, W. P. 1962. The Nature of Para-sitism. Academic Press, New York.
Stephenson, W. 1942. The effect of variationsin osmotic pressure upon a freeliving soilnematode. Parasitology 34: 253—265.
Tweedie, J. W., and I. H. Segel. 1967. Spec-ificity of Group VI anion transport infilamentous fungi. Bacteriological Proceed-ings, p. 29.
Von Brand, T. 1943. Physiological observa-tions upon a larval Eustrongylides. IV. In-fluence of temperature, pH and inorganicions upon oxygen consumption. Biol. Bull.,84: 148-156.
. 1960. Influence of pli, ions, andosmotic pressure on life processes in Sasserand Jenkins. Nematology. University ofNorth Carolina Press, Chapel Hill.
Wallace, H. R., and D. N. Greet. 1964.Observations on the taxonomy and biologyof Tijlenchorhtjnchus macrurus (Goodey,1932) Filipjev, 1936 and T. icarus sp. nov.Parasitology 54: 129-144.
Weinstein, P. P. 1952. Regulation of waterbalance as a function of the excretory systemof filariform larvae of Nippostrongijlus murisand Ancylostoma caninum. Exptl. Parasit.1: 363-376.
. 1960. Excretory mechanisms and ex-cretory products of nematodes: an appraisal.In: Host Influence on Parasite Physiology.Ed. Stauber. Rutgers University Press, NewJersey.
Yamamoto, L. A., and I. H. Segel. 1966.The Inorganic Sulfate Transport System ofPenicillin chn/sogenum. Arch. Biochem. Bio-phys. 114: 523-538.
Copyright © 2011, The Helminthological Society of Washington

10 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Fine Structure of the Somatic Muscles of the Free-Living MarineNematode Deontostoma californicum Steiner and Albin, 1933(Leptosomatidae)
W. DUANE HOPE1
Department of Invertebrate Zoology, Museum of Natural History, Washington, D. C.
The somatic musculature of nematodes is,in general, comprised of a single layer of longi-tudinally oriented, spindle-shaped muscle fi-bers. Each fiber has a noncontractile region,which gives rise to a "neuromuscular"process. The contractile region of the fiberis applied to the hypodermis of the body-walland encloses an array of numerous ribbonlikebands or "fibers" (Chitwood and Chitwood,1950). Schneider (1860) observed that theseribbons are perpendicular only to that portionof cell membrane adjacent to the hypodermisin some nematodes, whereas in others theymay be perpendicular to the membrane at thebase and sides of the cell as well. The formerhe designated as platymyarian and the lattercoelomyarian muscle. Chitwood and Chitwood(1950) have since described cylindrical musclecells in which the ribbonlike bands are per-pendicular to the cell-membrane all around thecell, enclosing a central core of sarcoplasm, andwhich they termed circomyarian.
As others have indicated (Hanson and Lowy,1960), our understanding of the structure ofthe contractile bands and other features ofnematode muscle has not been advanced sig-nificantly in recent years by light microscopeobservations. But, recent electron microscopestudies have contributed substantially to anunderstanding of the fine structure of nema-tode muscle. In a study of the coelomyarianmuscles of Ascaris himbricoides by Kawagutiand Ikemoto (1958c) and of Parascaris equo-
1 This study was conducted in the Department of Para-sitoloRy, University of Toronto, Toronto, Canada and sup-ported by NRC grant A 3757 from the National ResearchCouncil of Canada.
rum by Hinz (1963) it was demonstrated forthe first time that the "contractile fibers" or"contractile ridges" of light microscope observa-tions, are comprised of myofilaments. Hinzalso mentioned the presence of invaginationsof the sarcolemma which we would now recog-nize to be "T-tubules." But a "T-system,"which is now a well-known structure of verte-brate muscle, was first clearly demonstrated innematode muscles by Rosenbluth (1963;1965a) and Reger (1964) in studies of A.himbricoides. They also showed that cisternaeof the sarcoplasmic reticulum are closely asso-ciated with the T-tubules forming both dyadsand triads, and that both thick and thin myo-filaments are present.
The arrangement of myofilaments in thetype of muscle that occurs in nematodes differsfrom that in vertebrate cross-striated muscle.Our understanding of this arrangement beganwith early studies by Kawaguti and Ikemoto(1958c) on the muscle of Ascaris and on sim-ilar muscle occurring in annelids (1957b;1958a, cl), molluscs (1957a; 1961) and asea squirt (1958b). Their studies revealedthat these muscles have in common an obliquelongitudinal striation and a repeating patternof transverse bands of thick and thin fila-ments. Rosenbluth (1963; 1965a) and Ike-moto (1963) showed that these features weredue to a uniform arrangement of longitudinallystaggered myofilaments in muscle of Ascarisand an oligochaete, respectively. While theseand other studies (Auber-Thomay, 1964;Jamuar, 1966; Lee and Miller, 1967; Watson,1965a; Wright, 1964) have revealed that the
Figure 1. Transverse section of muscle fibers showing noncontractile region (S) bearing the nucleus(N), and contractile regions (C) with horizontal basophilic (A) and nonstained zones (I) and verticalnonstaiiied zones (V). Polyethylene glycol. Haematoxylin. X 2,150.
Figure 2. Transverse section. Dark stained material around the nucleus (N) is glycogen (G); that be-tween cells is the glyco-protein of the sarcolemma (M). Paraffin. PAS. X 2,150.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 11
Copyright © 2011, The Helminthological Society of Washington

12 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
arrangement of myofilaments is basically thesame in most nematode muscle, they have alsodisclosed deviations in the extent of develop-ment and form of the "electron dense material"and sarcoplasmic reticulum, and in the pres-ence or absence of a T-system.
The above studies constitute a significantadvance in our understanding of nematodemuscle, but further studies are desirable tounderstand not only the structural relationshipbetween platymyarian, coelomyarian, and cir-comyarian muscle of nematodes, and betweennematode muscle and obliquely striated anddouble obliquely striated muscle of other in-vertebrates, but the significance of variationsof the sarcoplasmic reticulum and "electrondense material." In view of this, light andelectron microscopy have been employed in astudy of coelomyarian muscle of the free-livingmarine nematode, Deontostoma californicumSteiner and Albin, 1933, and the observationsare compared with those from previous studieson nematodes and higher metazoans.
Materials and MethodsSpecimens were collected from sediment re-
moved from holdfasts of the alga Egregia sp.from intertidal rocks at Dillon Beach, MarinCounty, California. All nematodes were re-tained in sea water at 15 C until fixed; somewere fixed within 24 hours while others werekept in sea water at 15 C for up to one month.
LIGHT MICROSCOPY: Specimens used forgeneral histological examination were fixedwhole for 24 hours in 10% formalin in seawater. To avoid distortions that usually occurduring paraffin embedding, the specimens wereembedded in a 39 to 1 mixture of polyethyleneglycol 1540 and 4000, respectively, by thefollowing modification of the method of Riopeland Spurr (1962). Fixed specimens werewashed for approximately 30 minutes in tap
water and then placed in a Bureau of PlantIndustry Dish filled with a 2.5% aqueous solu-tion of polyethylene glycol 400. The dish wascovered with a cover glass to extend evapora-tion of the water over a period of about oneweek, in much the same way that nematodesare conventionally dehydrated in glycerine.This procedure, in lieu of passing the nema-todes through a graded polyethylene glycol400 series, eliminates collapsing of the bodyand minimizes distortion of tissues. After com-pleting the embedding as recommended byRiopel and Spurr, the nematodes were sec-tioned at 5 /JL on a rotary microtome and stainedfor 5 minutes in 0.1% aqueous hematoxylin atpH 2.4 by the method of Craig and Wilson(1937).
Fasted and nonfasted specimens to be ex-amined for the presence of glycogen were fixedin Rossman's picric acid-alcohol—formalin fixa-tive (Gray, 1954), embedded in paraffin, sec-tioned at 10 fj., and stained by the periodicacid-Schiff method. Control sections weretreated with ptyalin.
ELECTRON MICROSCOPY: Whole specimenswere fixed at room temperature for 20 min-utes in a mixture of 1 volume of Acrolein and9 volumes of M/10 solution of Sorensen's phos-phate buffer adjusted to pH 7.4.
They were then rinsed in the buffer and cutinto pieces approximately 2 mm long in 1%OsO4 in veronal acetate buffer (pH 7.4) con-taining sucrose (Palades sucrose fixative) andpost-fixed in the same solution for 1 hour.
After dehydration in cold ethanol, the tis-sues were embedded by the method of Wrightand Jones (1965) in a mixture of 9 volumesof Maraglas 655, 1 volume of Cardolite Nc513, and 0.25 volumes of DMP-30 (Freemanand Spurlock, 1962).
Sections were cut on LKB and Porter Blumultramicrotomes, mounted on Formvar or
Figure 3. Parasagittal section of a muscle fiber. Basophilic zones (A). Nonstained zones (I). Spindle-shaped profiles (Z). Polyethylene glycol. Haematoxylin. X 2,800.
Figure 4. Electron micrograph of glycogen particles in perinuclear sarcoplasm of nonfasted specimen.Lead hydroxide. X 91,000.
Figure 5. Electron micrograph of cross-sectioned contractile region showing various profiles of thickzone (A and I Bands) material. Note the presence of glycogen particles (G) in the Z planes (ZP). Leadhydroxide. X 33,800.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 13
. - Y -V.'VrCff^v.
Wl
Copyright © 2011, The Helminthological Society of Washington

14 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Formvar and carbon coated grids, stained withWatson's (1958) saturated lead hydroxide orwith permanganate and lead citrate (B. L.Soloff, personal communication), and exam-ined with a RCA EMU-3E (operated at 50 kv)or Zeiss EM 9 electron microscopes.
ObservationsLIGHT MICROSCOPY: The body wall muscu-
lature of D. califo-rnicum consists of a singlelayer of longitudinally oriented, spindle-shapedmuscle fibers. In cross-section, an average of24 fibers appear in each midbody quadrant.The more obvious features of each fiber in-clude the enlarged, noncontractile region,which extends into the pseudocoelom, and arelatively slender contractile region, the baseof which is adjacent to the hypodermis (Fig. 1).
The noncontractile region contains a nucleusand substantial quantities of PAS positive ma-terial (Fig. 2), which is removed by ptyalindigestion. From these observations, and thosemade with the electron microscope (see be-low), it is concluded that this material isglycogen.
The contractile portion of the fiber, whenviewed in cross-section, appears to be com-prised of basophilic zones which usually passuninterrupted from one side of the fiber to theother. These zones are separated from oneanother by nonstained zones that also passfrom one side of the fiber to the other (Fig. 1).Sometimes, however, nonstained zones occurin or near the sagittal2 plane of the musclefibers as well, extending varying distancesfrom the perinuclear sarcoplasm toward thebase of the fiber and separating the stainedzones of this region into right and left halves(Fig. 1). The vertically oriented nonstainedzones resemble the sarcoplasmic core of Ascarismuscle, but differ in being narrower, not ex-tending as far toward the base of the fiber, andin appearing less frequently.
It is evident from parasagittal sections ofthe fiber, that the basophilic and nonstainedzones described above are ribbonlike and ex-tend over much of the length of the fiber (Fig.3). In the same sections, small spindle-shapedprofiles are spaced at rather regular intervals in
2 Sagittal (para-) is used here to indicate longitudinalsections whose plane extends from the base of the fiber tothe apex; frontal sections are also longitudinal with theplane extending between the right and left sides of thefiber.
the nonstained zones, thus resembling similarstructures appearing in longitudinal and ob-lique sections of Ascaris (see Rosenbluth,1965a).
The entire fiber is enclosed in a PAS posi-tive sarcolemma (Fig. 2) which remains reac-tive after ptyalin digestion.
ELECTRON MICROSCOPY: The sarcoplasm ofthe noncontractile region of muscles in non-fasted specimens contains a high density oflead-stained particles 100—230 A in diameter(Fig. 4). These are surmised to be particlesof the beta form of glycogen because of theirsize, form, and staining properties, and alsotheir distribution is coincident with the PASpositive, ptyalin-digestable substance observedin light microscope preparations. Glycogenparticles may also occur in the contractile por-tion of fibers (Fig. 5). Alpha particles oc-curred infrequently in all sections examined,and some specimens maintained in sea waterfor as little as one week without food, con-tained little glycogen in either the noncontrac-tile or contractile regions (Fig. 6).
Mitochondria occur in the noncontractileregion and occasionally were observed subja-cent to the cell membrane in the contractileregion as well. Slender strands of presumablynoncontractile filaments and vesicles bound bysmooth membranes are also evident in non-contractile regions and more readily observedin specimens depleted of glycogen. Golgi zoneswere not observed.
Low magnification electron micrographs ofcross-sectioned muscle fibers reveal, in thecontractile region, an arrangement of widetransverse zones containing small circular pro-files of cross-sectioned filaments and, alternat-ing with the above zones, much narrower zonescontaining bars of electron dense material (Fig.6). This alternating pattern superficially re-sembles the stained and nonstained zones oflight microscope observations (Fig. 1), but thedifference between the widths of the stainedand nonstained zones is not nearly so great asthat between the wide and narrow zones ofelectron microscope observations. Commonly,the wide zones extend uninterrupted from oneside of the fiber to the other, each appearingin cross-section as a broad rectangle with itslonger sides parallel to the base of the fiber.Occasionally, in the apical region of the fiber,each wide zone may be intercepted by vertical
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 15
m. • : ' / • ' • ' . 'it'- ' - ' • .*. - * -^feir'x^i^.t^i^V'-. »'**-. >•*."','" *:^^*'' ' ':-'"'• '"•
Wim!im
Figure 6. Electron micrograph of cross-sectioned muscle showing wide zones (W) and narrow zones(ZP) with bars of electron dense material (Z). Note absence of glycogen in sarcoplasm at upper leftand vertical Z plant at right (V). Permanganate and lead citrate. X 25,500.
Copyright © 2011, The Helminthological Society of Washington

16 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
narrow zones located near the sagittal plane ofthe fiber (Fig. 6) resembling the vertical non-stained zones from light microscope observa-tions. When separated in this manner, thewide zones on one side of the fiber are usuallyat the same level as wide zones of the oppositeside; less frequently are wide zones oppositenarrow zones. Sometimes, several irregularlyoriented narrow zones divide wide zones intoprofiles of various shapes, including circles(Fig. 5) and triangles, immediately basal tothe nuclear region and at the base of the cellas well.
Examination of cross-sections at higher mag-nifications shows that the wide zones are com-posed of two sizes of myofilaments, the thickerwith a maximum diameter of about 200 A, andthe thin about 60 A. These filaments are dis-tributed so as to form five transverse bands ineach thick zone. That is, there are two outerbands of thin filaments (Fig. 7, i ) ; medial tothese are two comprised of both thick and thinfilaments (Fig. 7, a ) ; and in the median planeof each thick zone, a single band of thick fila-ments only (Fig. 7, h) . This pattern is char-acteristic of obliquely striated muscle (Ike-moto, 1963; Rosenbluth, 1963; 1965a) andthe bands are comparable to the I, A, and Hbands, respectively, of vertebrate striated mus-cle. I bands occur on both sides of thin zonematerial regardless of the orientation of thelatter. The filaments of the I bands do notstain with hematoxylin as employed in thisstudy, and so the nonstained zones describedabove correspond to the I bands and thin zonesas well, which they flank. Thus, the nonstainedzones are not wholly comparable to the thinzones of electron microscope observations.
Thick filaments closest to the Z bands areslightly smaller in diameter than those of theH band and, therefore, the thick filaments areassumed to be slightly tapered as in Ascaris(see Rosenbluth, 1965a). Cross-linking be-tween thick and thin filaments in the A bands
was not observed, nor did there appear to bea regular arrangement of thin filaments aroundthe thick. Thin filaments extend to the marginof the narrow zones and here they frequentlyappear to be segregated in small clusters (Fig.7) much as has been found in the case ofAscaris lumbricoides (see Rosenbluth, 1965a).
From parasagittal sections it is evident thatthe wide zones extend uninterrupted for con-siderable distances through the fibers in alongitudinal direction, slightly oblique to thelongitudinal axis of the fiber. The bands ofwide zones are not nearly as distinct in thesesections as in those that are transverse, butit is at least possible to identify the A bands(Fig. 8).
The narrow zones are characterized at lowmagnifications by bars of electron dense ma-terial, the extent of which varies from almostnone in some narrow zones to uninterruptedbars passing from one side of a fiber to theother. Most frequently, however, the densematerial appears one to several times forshort distances in cross-sections of narrowzones and often appears closely applied to, orin some way fused with, the plasma membraneat the sides of the fiber (Fig. 6).
In longitudinal sections, the electron densematerial is apparent as a series of spindle-shaped profiles regularly spaced in a longi-tudinal plane that is slightly oblique to thelongitudinal axis of the fiber. Their periodicityis about 2.0 ^ which agrees very closely withcomparable measurements made from photo-micrographs. A narrow strand, presumably abundle of thin filaments, appears to extendfrom each end of each spindle, and merge withthe filaments of the I band (Fig. 8), as hasbeen observed in the case of Nippostrongyhismuscle (Jamuar, 1966). Consequently, thefilaments from one end of a spindle pass intothe I band below, and those from the oppositeend pass into the I band above the spindle.This apparent continuity between electron
Figure 7. Cross-sectioned muscle showing thick (T) and thin (t) filaments arranged into I, A, andH bands. Note clusters of thin filaments (t') at the margin of the Z plane (ZP). Permanganate andlead citrate. X 81,900.
Figure 8. Electron micrograph of a parasagittal section showing the thick filaments (T) of the widezones (A bands) and what are interpreted to be clusters of thin filaments (t') continuous with cross-sectioned profiles of Z bars (Z). Permanganate and and lead citrate. X 34,400.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 17
¥ jP-TI.'.. -iT" **" ' 2
• . - •-
:
Copyright © 2011, The Helminthological Society of Washington

18 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
dense material and I band filaments suggeststhat at least a portion of the electron densematerial is a Z component comparable to theZ band material of vertebrate striated muscle.For this reason, and for reasons given in thediscussion, the bars of electron dense materialwill be referred to here as Z bars, and theoblique plane bearing them, the Z plane.Frontal sections of muscle fibers disclose thatthe Z planes also bear fibrils oriented at rightangles to the myofilaments. In some instancesit appears that the fibrils are adjacent to, butnot enmeshed with the Z bars (Fig. 9), whilein other cases these components do appear tobe joined with each other (Fig. 10). It is alsopossible, however, that the latter may be anillusion attributable to superposition.
Sarcoplasmic reticulum in the form of nar-row tubules and dilated vesicles also occursconsistently in Z planes. The tubules are usu-ally closely applied to the surface of the Z barsso as to separate the latter from the I bands;in fact, wherever a Z bar is present in cross-sections, at least one and usually both of itssurfaces bear sarcoplasmic reticulum (Figs.11 and 12). Tubules of sarcoplasmic reticulumdo not alternate with Z bars in the Z plane asthey do in Ghjcera (see Rosenbluth, 1968).
Seldom is the lumen of the tubules morethan 400 A wide and often it is so narrow thata lumen is hardly perceptible (Figs. 11, 12).Vesicles, on the other hand, commonly have adiameter approximately equal to the width ofthe narrow zone (Figs. 12, 13). In the Z planethey occur most frequently in those areas de-void of Z bars. Continuity between the tubulesand vesicles is clearly evident in numerousinstances (Fig. 12), and tubules from bothsurfaces of the dense material may have con-tinuity with a common vesicle (Fig. 12). Al-though it was not possible to resolve the unitmembrane structure of the sarcoplasmic retic-ulum, or to measure accurately its membranewidth, membranes enclosing vesicles do notappear to differ from those enclosing tubules.
Furthermore, tubules and vesicles associatedwith the Z plane proper have a similar electrontranslucent medium within them.
Fingerlike projections of the vesicles mayextend into the I band (Figs. 9, 10) where, incross-section, they appear as small circularprofiles (Fig. 12). Therefore, the sarcoplasmicreticulum is not entirely restricted to the Zplane. In no instance was it possible to iden-tify dyads or triads within the Z plane, al-though vesicles (diads) occur subjacent to thelateral sarcolemma (Figs. 13, 14). The longaxis of the vesicles, measured from cross-sec-tions of the fiber, has an average length of0.34 fji and the short axis, 0.12 ^. The averagedistance separating the plasma membrane fromthe membrane of the vesicle is 120 A. In someinstances the lumen of the vesicle is completelyfilled with a rather electron dense, granularmaterial (Fig. 14), while in others it may haverelatively little (Fig. 13). The association be-tween the vesicles and the plasmalemma ishere regarded as a diad and is in accord withsimilar views stated by Rosenbluth (1968)regarding a comparable structural arrangementin Ghjcera muscle.
A T-system was not observed in this muscle,but two types of shallow invaginations of thesarcolemma do occur and both have asym-metrical unit membranes. The first type ofinvagination (Fig. 15) more closely resemblesthe T-tubules of Ascaris (Reger, 1964; Rosen-bluth, 1965a; and Watson, 1965b) in thatthey have been observed only at the level ofthe Z planes and have a diameter comparableto that of Ascaris T-tubules. They differ, how-ever, in being very infrequent, shallow, andsarcoplasmic reticulum has never been ob-served near them as it occurs in diad and triadformations of Ascaris. The second occurs inboth the noncontractile and contractile regionsof the fibers. It is characteristically a verynarrow infolding, the opposing membranesseparated by a uniform distance of 80 A. Theinfoldings may be rather extensive, occasionally
Figures 9 and 10. Electron micrographs of frontal sections demonstrating portions of Z bars (Z) andthin filaments (t) which become continuous (f) with them. Filaments (F) presumed to be of thecyto-skeletal system, traverse the Z plane at nearly right angles to the thin actin filaments. Notetubules and vesicles of sarcoplasmic reticulum (SR) extending into the I bands. Permanganate and leadcitrate. Figure 9, X 80,800. Figure 10, X 81,700.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 19
, . , , .
• • - , : .
H,v/.,;/ I'illMa
Copyright © 2011, The Helminthological Society of Washington

20 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
appear to ramify, and always remain close tothe cell surface (Fig. 16). They have neverbeen, observed to extend between bands offilaments, but may pass between filamentsand subsarcolemmal cisternae, although in thisinstance, the distance between the membranesof the subsarcolemmal cisternae and the in-foldings may be less than that between theformer and the cell surface.
The sarcolemma of Deontostoma muscleconsists of a cell membrane 100 A wide andan extracellular coating 1.5 ^ wide. It is likelythat this coating is responsible for the PASpositive staining both before and after ptyalindigestion, and is a form of glycocalyx whichinvests cells of many types (Fawcett, 1966).Unlike Ascaris, however, it is not comprisedof bands or lamellae.
Wright (1966) reported the occurrence ofcytoplasmic bridges between muscles of Deon-tostoma californicum which were again ob-served in this study.
DiscussionLight and electron microscope studies of
D. californicum reveal that its muscles havethe usual noncontractile portion which is lo-cated along the pseudocoelomic side of thefibers and a more slender contractile regionadjacent to the hypodermis.
NONCONTRACTILE REGION: No attempt hasbeen made to study this region of the musclein detail, but it is worth noting that it con-tains the nucleus, mitochondria, noncontractilefibrils, sarcoplasmic reticulum, and glycogenparticles as has been reported for other species(Wright, 1964; Reger, 1964; Rosenbluth,1965b; Lee and Miller, 1967). The abun-dance of glycogen is particularly impressive asthe intensity of the PAS reaction was muchgreater in somatic muscle than in any of the
other tissues including the gut which gavethe next most intense reaction. Large depositsof glycogen have been found in A. lumbricoidesand Dirofilaria immitis (Rosenbluth, 1965b;Lee and Miller, 1967). Rosenbluth has pointedout that the noncontractile region of the musclemay serve as a glycogen storage depot whichmay sustain the parasite during periods inwhich its host is fasting. This is very likelythe case, but because of comparable depositsof glycogen occurring in D. californicum, afree-living species, it is apparent that storageof glycogen in the muscle belly is not strictlyan adaptation to meet the vicissitudes ofparasitism.
CONTRACTILE REGION: Differences betweenDeontostoma muscle and the muscle of othernematodes are to be found among componentsof the Z plane, namely the electron dense ma-terial or Z bars and the sarcoplasmic reticulum.
This electron dense material may extenduninterrupted for considerable distance acrossthe width of a fiber or may appear intermit-tently in the same Z plane. In photomicro-graphs and electron micrographs of parasag-ittal sections of Deontostoma muscle, it canbe seen that the obliquely regimented pro-files of the dense material occur with regu-larity that suggests they may be an integralpart of the staggered bands of myofila-ments. Indeed, it can be seen at higher mag-nifications of the same sections (Fig. 8, Z)that each profile is tapered and appears tobe continuous with thin filaments of adjacentI bands. For these reasons it is suggested thatat least a part of the electron dense materialof this muscle is Z material; comparable to theconstituents of Z bands in vertebrate muscle.A second component in the Z bars is evidencedby frontal sections in which filaments (Figs.9 and 10, F), presumably of the cyto-skeletal
Figures 11 and 12. Electron micrographs of cross-sectioned muscle cell showing Z bars (Z) and tubulesand vesicles of sarcoplasmic reticulum (SR). Note continuity between tubules and vesicles. Lead hy-droxide. X 76,340.
Figure 13. Electron micrograph of cross-sectioned muscle fibers demonstrating an enlarged vesicle ofsarcoplasmic reticulum (SR) closely applied to the plasma membrane (P) forming a diad at the level ofthe Z plane (ZP). The vesicle is partially filled with granular material. Lead hydroxide. X 97,400.
Figure 14. Same as Figure 13, but the diad is at the level of an A band with a tubule directed intothe Z plane. The vesicle is filled with granular material. Permanganate and lead citrate. X 156,000.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 21
* jf •**.» I** •» ' —- ."£ .. "^ >' <^•9^ «
•'> ,V V.'W."
Copyright © 2011, The Helminthological Society of Washington

22 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
system, are observed traversing the Z planeat right angles to the longitudinal axis of thefiber. These filaments were observed infre-quently which suggests that they either occurinfrequently or that, if they are a commoncomponent of Z bars, they are fused or en-meshed with the Z component and, therefore,difficult to identify.
Z bars, or their counterpart have not beendescribed in detail for ncmatodes except inthe case of Ascaris. According to Rosenbluth(1965a) the dense bands (Z planes) in Ascariscontain "bundles of fibrils" (presumably of thecyto-skeletal system) which appear in cross-section as tightly packed ovoid "dense bodies"and a second component, which he designatesas "Z bundles," comprised of "small, sheaflikeaggregates" of the thin filaments in which thelatter "appear to be linked together." Thesebundles occur not only at the edges of the Izones but within it as well. He further statesthat "thin filaments sometimes seem to join thedense bodies" and later (1967), without fur-ther comment, states that the "thin filamentsinsert into opposite ends of the dense bodies."From these comments it is not entirely clearwhether in Ascaris muscle there are Z bundlesin addition to dense bodies, or whether thedense bodies themselves are the Z juncturesbetween "sarcomeres."
What are interpreted to be aggregates ofthin filaments occur in the Z plane of Deonto-stoma muscle (Fig. 7, t'), as in Ascaris. How-ever, it is suggested that these are not Zbundles, but are aggregates of thin filamentsgrouped together as they converge upon (oremerge from) the Z bars. A similar situationmay exist in Nippostrongylus muscle, since it
appears that there are aggregates of thin fila-ments in the Z plane (Lee, 1966) and thatclearly these are bundles of thin filaments con-tinuous with the Z component (Jamuar, 1966).By this interpretation, the structure in Ascaris,designated by Rosenbluth as "Z bundles" and"ovoid dense bodies," may in reality be aggre-gates of converging or diverging thin fila-ments and ovoid Z bodies, respectively.
From their study on DirofiJaria immitismuscle, Lee and Miller (1967) concludedthat what had been termed "supportingfibers" in the myofibrillar and amyofibrillararea of nematode muscle, are identical withthe Z bundles of Rosenbluth (1965a). Butevidence of fibrils traversing the Z plane atright angles to the myofilaments in Deonto-stoma muscle, supports earlier concepts of asystem of cytoskeletal fibrils separate and dis-tinct from the contractile filaments.
Whereas in Ascaris the Z junctures are inthe form of an ovoid dense body, and barlikein Deontostoma, they apparently have the formof dense thickenings subjacent to the plasmamembrane in Capillaria hepatica muscle stud-ied by Wright (1964). The distribution ofthese dense areas along the base and sides ofthe fibers suggests that it is coelomyarian, buthighly specialized, and possibly degenerate.The fact that the Z juncture is plaquelike andlimited to the periphery of the cell, not extend-ing across the fiber as a series of dense bodiesor a bar, may explain the absence of distincttransverse bands of thick and thin filaments.3
The Z junctures in Nippostrongylus brasil-
3 Wright (1964) reported myofilaments of but onediameter, but disclosed in personal communication that hehas since found thin filaments as well.
Figure 15. Cross-section of a fiber demonstrating one of the few observed examples of relatively broadinvaginations of the sarcolemma located only at the level of the Z plane. Permanganate and lead citrate.X 131,200.
Figure 16. Cross-section of a fiber with examples of the numerous narrow invaginations of the sar-colemma that occurred in contractile and noncontractile regions of the fiber. Permanganate and leadcitrate. X 98,200.
Figure 17. Composite illustration of the Z plane of Deontostoma muscle. Electron dense Z materialpresumably with incorporated filaments of cytoskeletal system (Z); vesicles and tubules of sarcoplasmicreticulum (SR).
Figure 18. Composite illustration of the Z plane of Ascaris muscle based on the authors interpreta-tion of electron micrographs published by Rosenbluth (1965A; 1967). T-tubule (T); electron dense Zmaterial with fibrils of cytoskeletal system (Z); sarcoplasmic reticulum (SR); and filaments of the cyto-skeletal system (F).
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 23
, . f i
•
Copyright © 2011, The Helminthological Society of Washington

24 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
ien-sis muscle are also of interest, not becauseof the shape of the juncture itself, which ap-pears to be rodlike, but in that they areoriented at right angles to the plasma mem-brane at the base of the fiber instead of at thesides and are, therefore, platymyarian. Thesignificance of this is discussed below withregard to oblique striation.
The above variations in the Z component ofnematode muscle, with the possible exceptionof C. hepatica, can also be found among theobliquely striated and double obliquely striatedmuscle of annelids and molluscs. The Z com-ponent appears to have the general shape ofan ovoid structure in the heart muscle (J-gran-ules; Kawaguti, 1963), and mantle muscle ofcuttlefish (J-particles; Kawaguti and Ikemoto,1957a), and is more rocllike in leeches (J-rods,compact sarcotubules and cross filaments;Kawaguti and Ikemoto, 1958a; Pucci andAfzelius, 1962; Rohlich, 1962), polychaetes(J-rods; Z lines; Kawaguti and Ikemoto, 1958d;Rosenbluth, 1968) and oligochaetes (J-rods;Ikemoto, 1963). But in all instances thesestructures are located in what is comparableto the Z plane of Deontostoma muscle, and, inmany instances, there is evidence that thinfilaments are continuous with them (Kawagutiand Ikemoto, 1957a, b; 1958a, b, c; Rohlich,1962; Rosenbluth, 1968; Ikemoto, 1963).
At present it seems that the greatest differ-ence between Z structures of nematode muscle,on the one hand, and those of the obliquelystriated muscles of higher metazoans on the
other, is that the former appears to have acomponent of cytoskeletal fibrils which havenot been observed in the latter. Otherwise, theZ structures seem to be quite comparable inobliquely and double obliquely striated muscle,and it seems they are also comparable to theZ bands of vertebrate cross-striated muscle.
SARCOPLASMIC RETICULUM: The sarcoplas-mic reticulum of the Z plane of Deontostomamuscle is extensively developed while a T-system is lacking or poorly developed. InAscaris, on the other hand, the sarcoplasmicreticulum is not as extensive and a T-systemis well developed. These differences are mani-fest in the basic arrangement of Z plane com-ponents. Transverse sections of Deontostomamuscle fiber disclose that, when all componentsof the Z plane co-exist, the electron densematerial is medial in the Z plane and the nar-row membrane bound cisternae are peripheralto it. Diad or triadlike formations were neverobserved in the Z plane (Fig. 17). In Ascaris,by contrast, the medial region of the plane isoccupied by a relatively broad membrane-bound structure (T-tubule) with the densematerial and narrow cisternae of sarcoplasmicreticulum (which are the lateral elements ofthe diads and triads) displaced to the periph-ery of the Z plane (Fig. 18). Commonly, rela-tively broad vesicles limited by membranes(which are sarcoplasmic reticulum) may oc-cupy the entire width of the Z plane in Deon-tostoma muscle, superficially resembling theT-tubules of Ascaris. However, when this
Figure 19. Cut-away diagram of platymyarian muscle fiber depicting the patterns of oblique I and Abands (H bands not shown) and Z planes. Based on the authors interpretation of electron micrographspublished by Lee (1966) and Jamuar (1966).
Figure 20. Cut-away diagram of a coelomyarian muscle cell showing the pattern of double obliquestriation (partial helix) that would result if the platymyarian arrangement of oblique bands were ex-tended onto opposing sides of the fiber. Based on the authors interpretation of electron micrographspublished by Rosenbluth (1965A; 1967).
Figure 21. Cut-away diagram of circomyarian muscle fiber showing double oblique striation that wouldresult if the platymyarian arrangement of oblique striations were extended around the entire peripheryof the fiber forming complete helices. Inferred from authors interpretation of data by Chitivood (1951),Lee (1966), Jamuar (1966), and Rosenbluth (1965A; 1967).
Figure 22. Enlarged portion of comparable contractile region bracketed on each of the cut-away dia-grams. The same sequence of H, A, and I band and Z planes occurs in all obliquely and doubleobliquely striated muscle.
Figure 23. Cut-away diagram of Deontostoma muscle showing the probable arrangement of bands andZ planes inclined in the same direction and at the same angle in both right and left sides of the fiber.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 25
ZP H A I
'«!••„••«•; f
•
20
23
Copyright © 2011, The Helminthological Society of Washington

26 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
situation exists, electron dense material andmembrane-bound vesicles are not peripheralto it. That these dilated vesicles are com-ponents of the sarcoplasmic reticulum is fur-ther evidenced by their continuity with themuch narrower cisternae. Continuity betweenthe sarcolemma and either the vesicles orcisternae was never observed and, since theplasma membrane of the fiber has asymmet-rical unit membrane structure, while unit mem-brane structure of the sarcoplasmic reticulumcould not be resolved, it seems unlikely thatany of the membrane-bound vesicles withinthe Z plane are derived from the sarcolemma.Rather, it is interpreted that the membrane-bound structures within the Z plane and sub-jacent to the sarcolemma are components ofthe sarcoplasmic reticulum and the vesicleslying adjacent to the sarcolemma, situatedeither above or below Z planes, are associatedwith the sarcolemma so as to form diads.
Sarcoplasmic reticulum in the form describedfor Deontostoma has not been previously de-scribed for other nematodes, but it appearsthat the "membrane units" described for Ca-pillarla by Wright (1964) may also be sub-sarcolemmal cisternae or diads without in-wardly directed tubules. Nippostrongylusbrasiliensis muscle may also be similar to thatof Deontostoma with respect to the absenceof a T-system and greater development ofsarcoplasmic reticulum, while Euchromadoravulgaris and Dirofilaria immitis seem to moreclosely resemble Ascaris muscle in this regard.
The function(s) of sarcoplasmic reticulumis, as yet, not well understood, but one pos-tulate, which has been reviewed by Porter(1961), states that it may be involved inmuscle relaxation, by uptake of calcium, byproduction of a "relaxing factor," or both.With regard to this theory, it is presumed thathypertrophy of the sarcoplasmic reticulum isdirectly related to rapid cycles of contractionand relaxation. If this theory proves to becorrect, it may explain the relatively greaterabundance of sarcoplasmic reticulum in Deon-tostoma, which is a free-living nematode thatpresumably moves about in search of food bymeans of frequent cycles of muscle contractionand relaxation, and less sarcoplasmic reticulumin adult Ascaris, which retains itself in thelumen of the gut of its host by bracing againstthe gut wall (Makidono, 1956), and may,
therefore, move rather infrequently with slowcycles of contraction and relaxation.
A system of sarcoplasmic reticulum similarto that in Deontostoma muscle is more commonin annelid muscle, having been reported in thecase of Glycera muscle (Rosenbluth, 1968),Eisenia muscle (Ikemoto, 1963), Lumbricusmuscle (Heumann and Zebe, 1967), and leechmuscle (Pucci and Afzelius, 1962). However,in these annelids the tubules of sarcoplasmicreticulum are not in direct association with theZ bar as they are in Deontostoma, but occur inthe Z plane midway between each pair of Zbars. This differenece may have some rele-vance to speed of contraction and relaxation.
OBLIQUE AND DOUBLE OBLIQUE STRIATION : Allmuscles with longitudinally staggered bundlesof myofilaments (obliquely striated) appear tohave similar organization and structure whetherfrom representatives of the Nemata, Annel-ida, or Mollusca. However, there are repre-sentatives of the latter two taxa for whichdouble oblique striation has been clearly dem-onstrated, namely the clamworm (Kawagutiand Ikemoto, 1958d), tellinid clam (Kawagutiand Ikemoto, 1961) and the annelid Myxicolasp. (Hanson and Lowy, 1960). In theseexamples, bands of myofilaments occur onopposing sides of the cell and the bands onone side are oblique in a direction opposite tothose of the other side.
Patterns of double oblique striation have notbeen reported for nematode muscle, but in con-sidering the following information from lightand electron microscope studies, it seems quitepossible that patterns of double oblique stria-tion may occur in at least certain types ofnematode muscle. From light microscopy it isknown that "contractile ridges" (bands of myo-filaments) are only at the base in platymyarian(Fig. 19), at the base and sides in coelo-myarian (Fig. 20) or around the entire periph-ery enclosing the medial sarcoplasm in cir-comyarian muscle cells (Fig. 21). Further,platymyarian muscle cells tend to be flattened,that is as wide, if not wider, than tall (Fig.19); those that are coelomyarfan are usuallytaller than wide with an enlarged perinuclearregion at the apex and a narrower base con-taining the myofilaments (Fig. 20); andcircomyarian fibers are cylindrical (Fig. 21).Many nematode muscle cells tend to have anorganization and shape intermediate to those
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 27
assigned to these categories. From the illus-trations in Chitwood and Chitwood (1950)it appears that the somatic muscles of Ethmo-laimiis revaliensis, are very low coelomyarian toplatymyarian while those of Theristus setosusare higher coelomyarian tending toward circo-myarian. Some muscle fibers, such as those ofAscaris, are coelomyarian in the midregion andcircomyarian toward their ends. Furthermore,according to Chitwood and Chitwood (1950),Martini has shown that some nematodes areplatymyarian as first stage juveniles, and laterdevelop coelomyarian muscle. The above ob-servations suggest a similar structural relation-ship between the muscles of these categories.
From electron microscope observations ofcross-sectioned fibers it is known that the samesequence of H, A, and I bands and Z planesoccur in coelomyarian muscle (Rosenbluth,1963, 1965a; Reger, 1964; Watson, 1965a;and Lee and Miller, 1967) as well as platy-myarian muscle of Nippostrongylus brasiliensis(see Jamuar, 1966) and that all bands and Zplanes are at right angles to the cell membrane(Fig. 22). Therefore, the longitudinal stria-tions of platymyarian muscle, if viewed fromthe base or other appropriate angle, wouldappear to have slightly oblique orientationfrom the right side of the fiber to the left orvice-versa (Fig. 19). If exactly the same pat-tern of striations were formed adjacent to acell membrane that was not broad at its base,but curved or folded so as to appear U-shapedin cross section with continuous bands occur-ring on both sides of the fiber as \vell as atthe narrower base (coelomyarian), then thesame pattern of striations would pass fromnear the apex to the base on one side, continueacross the base, and upward toward the apexof the opposite side (Fig. 20). If these as-sumptions are correct, then I, A, and H bandsand Z planes would form partial helices incoelomyarian muscle and if the same patternwere to occur in circomyarian muscle, com-plete helices would be formed (Fig. 21). Iffibers of either of these two types were ob-served in longitudinal optical section withsufficient depth of focus to simultaneously ob-serve the bands and Z plane on both sides ofthe fiber superimposed on one another, itwould have the appearance of double obliquestriation (Figs. 20, 21) comparable to that ofannelids and molluscs.
It is of interest to note further, however, thatbecause the striations are inclined in oppositedirections on opposite sides of the fiber inthe above-suggested situations, each of thebands on one side, in cross section, cannotalways be on the same level with a compa-rable band of the opposite side. In other wordsthe bands of one side will be out of registerwith those of the other. An example of this,and in which double oblique striation has beendemonstrated as well, is to be found in theadductor muscle of the tellinid clam (Kawagutiand Ikemoto, 1961).
From electron micrographs of cross sectionsof Ascaris muscle (Rosenbluth, 1965a) it ap-pears that the bands of one side are out ofregister with those of the other which sug-gests the possibility of double oblique striationin the muscles of this species. However, thisis contrary to the situation in Deontostoma,where in cross section the majority of thebands on one side of the fiber are precisely onthe same level with a comparable band of theother side giving the appearance of continuitythe full width of the fiber (Figs. 1, 2, 23). Asimilar arrangement exists in Glycera muscle(Rosenbluth, 1968). This "in register" con-dition is here interpreted to indicate that thebands are not only inclined at the same angle,but in the same direction and each myofila-ment band would give the appearance of alamella that spans the full width of the fiberand extends obliquely from the apex to thebase of the fiber (Fig. 23). Even whereseparated by a sagittal Z plane, the bands onone side are usually "in register" and parallelwith those of the opposite side. Therefore,even under favorable conditions, doubleoblique striation most likely could not bedemonstrated in Deontostoma muscle.
What significance these considerations havecannot be fully appreciated at this time, butdifferences may be relevant to the function ofnematode muscle and the relationships ofnematode taxa. Perhaps functional implica-tions will not be as readily apparent and maybe appreciated only as more is learned regard-ing the mechanics of nematode muscle con-traction. But, on the other hand, if subsequentinvestigations disclose that somatic musclesnow collectively designated as coelomyarianprove to be a composite of two types, onehelical or double obliquely striated and the
Copyright © 2011, The Helminthological Society of Washington

28 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
other bilaterally parallel, and if all membersof the order Enoplida (to which Deontostomabelongs) have muscle of the former type andthose of Ascarida (to which Ascaris belongs)the latter, then these differences will give fur-ther justification for the wide separation be-tween these two taxa in the taxonomic hier-archy of nematodes and may provide furtherclues to phylogenetic relationship within theNemata.
SummaryThe structure of somatic muscles of Deonto-
stoma californicum is described from light andelectron microscope observations. The non-contractile region bears essentially the sameorganelles as described for previously studiednematodes and contains considerable quanti-ties of glycogen.
The basic organization of the contractileregion is that of obliquely striated muscle withcharacteristic repeated patterns of H, A, and Ibands and Z components and resembles theobliquely striated muscles of other nematodes,annelids, and molluscs. Deontostoma musclediffers from other nematode muscle in that itsZ components appear barlike, and a well-developed T-system is absent. Furthermore, ithas a well-developed system of sarcoplasmicreticulum consisting of tubules and vesicles inthe Z plane which are continuous with diads.Functional implications of the differences inthe extent of the development of sarcoplasmicreticulum are briefly reviewed.
In Deontostoma muscle the continuity of Zcomponents and myofilament bands from oneside of the fiber to the other suggests that eachband forms lamellae which span the full widthof the fiber and extend obliquely from theapex to the base of the fiber. Therefore, theorganization of Deontostoma muscle is suchthat it could not display double oblique stria-tions possibly present in circomyarian andsome coelomyarian nematode muscles. It issuggested that these differences may havefunctional and/or phylogenetic implications.
AcknowledgmentsThe author wishes to express sincere grati-
tude to Dr. A. M. Fallis and Dr. K. A. Wright,Department of Parasitology, University ofToronto, for the hospitality they gave me dur-
ing this study, and especially to Dr. Wrightfor his invaluable guidance. I also wish tothank Dr. Rosenbluth for his helpful review ofthe manuscript and the very capable Mrs.Carolyn Cast for preparing the illustrations.
Literature CitedAuber-Thomay, Michele. 1964. Structure et
innervation cles cellules musculaires de nema-todes. Microscopie 3(1): 105-113.
Chitwood, B. G., and Maybelle B. Chitwood.1950. An Introduction to Nematology. Sec-tion I, Anatomy. Monumental Printing Co.,Baltimore. 213 pp.
Craig, B., and C. Wilson. 1937. The use ofbuffered solutions in staining: theory andpractice. Stain. Technol. 12(3): 99-109.
Fawcett, D. W. 1966. An Atlas of FineStructure. The Cell, Its Organelles and In-clusions. W. B. Saunders, Philadelphia andLondon. 448 pp.
Freeman, J. A., and B. O. Spurlock. 1962.A new epoxy embedment for electron micros-copy. J. Cell Biol. 13: 437-443.
Gray, P. 1954. The Microtomists' Formularyand Guide. Blakiston Co., Inc., New York.794 pp.
Hanson, Jean, and Jean Lowy. 1960. Struc-ture and function of the contractile apparatusin the muscles of invertebrate animals. InBourne, G. H. The Structure and Functionof Muscle I. Structure. Academic Press,New York and London. 472 pp.
Hinz, E. 1963. Elektronenmikroskopische Un-tersuchungen an Parascaris equorum (Integ-ument, Isolations-gewebe, Musculature undNerven). Protoplasma 56(2): 202-241.
Heumann, H. G., and E. Zebe. 1967. tJberFeinbau und Funktionsweise der Fasern ausdem Hautmuskelschlauch cles Regenwurms,Lumbriciis terrestris L. Z. Zellforsch. 78:131-150.
Ikemoto, N. 1963. Further studies in electronmicroscopic structures of the oblique-striatedmuscle of the earthworm, Eisenia foetida.Biol. J. Okayama Univ. 9(3-4): 81-126.
Jarnuar, M. P. 1966. Electron microscopestudies on the body wall of the nematodeNippostrongylus brasiliensis. J. Parasit. 52( 2 ) : 209-232.
Kawaguti, S. 1963. Electron microscopy onthe heart muscle of the cuttlefish. Biol. J.Okayama Univ. 9(1-2): 27-40.
Kawaguti, S., and N. Ikemoto. 1957a. Elec-tron microscopy of the smooth muscle of acuttlefish, Sepia esculenta. Biol. J. OkayamaUniv. 3 (3 ) : 196-208.
. 1957b. Electron microscopy on the
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969
smooth muscle from the body wall of theearthworms, Pheretima communissima andEisenia foetida. Biol. T. Okayama Univ. 3(4 ) : 223-238.
—. 1958a. Electron microscopy on thesmooth muscle of the leech, Hirudo nipponia.Biol. J. Okayama Univ. 4(1-2): 79-91.
. 1958h. Electron microscopy of theheart muscle from the sea squirt, Cionaintestinalis. Biol. J. Okayama Univ. 4(1-2):93-101.
. 1958c. Electron microscopy on thesmooth muscle of the roundworm, Ascarislumbricoides. Biol. J. Okayama Univ. 4(3—4) : 177-190.
. 1958d. Electron microscopy of thesmooth muscle of the clamworm, Nereisjaponica. Biol. J. Okayama Univ. 4(3-4):207-216.
1961. Electron microscopy on the ad-ductor muscle of a tellin, Fabulina nitidula.Biol. J. Okayama Univ. 7(1-2): 17-29.
Lee, Chin-Chiu, and J. H. Miller. 1967.Fine structure of Dirofilaria immitis body-wall musculature. Exper. Parasit. 20: 334-344.
Lee, D. L. 1966. An electron microscope studyof the body-wall of the third stage larva ofNippostrongylus brasiliensis. Parasitology 56:127-135.
Makidono, J. 1956. Observations on Ascarisduring fluoroscopy. Amer. J. Trop. Med.Hyg. 5: 699-702.
Porter, K. R. 1961. The Sarcoplasmic Retic-ulum: Its recent history and present status.J. Biophys. and Biochem. Cytol. 10(4/2):219-226.
Pucci, Ida, and B. A. Afzelius. 1962. Anelectron microscope study of sarcotubulesand related structures in the leech muscle.J. Ultrast. Res. 7(3/4): 210-224.
Reger, J. F. 1964. The fine structure of thefibrillar network and sarcoplasmic reticulumin smooth muscle cells of Ascaris lumbri-coides (var. suum). J. Ultrast. Res. 10:48-57.
Riopel, J. L., and A. R. Spurr. 1962. Carbo-wax for embedding and serial sectioning ofbotanical material. Stain Tech. 37(6): 357-362.
Rohlich, P. 1962. The fine structure of themuscle fiber of the leech Hirudo rnedicinalis.J. Ultrast. Res. 7(5-6): 399-408.
Rosertbluth, J. 1963. Fine structure of bodymuscle cells and neuromuscular junctions inAscaris lumbricoides. J. Cell. Biol. 19(2):82A.
. 1965a. Ultrastructural organization ofobliquely striated muscle fibers of Ascarislumbricoides. J. Cell Biol. 25(3): 495-515.
. 1965b. Ultrastructure of somatic mus-cle cells in Ascaris lumbricoides II. Inter-muscular junctions, neuromuscular junctions,and glycogen stores. J. Cell Biol. 26(2) :579-591.
. 1967. Obiquely striated muscle. III.Contraction mechanism of Ascaris bodymuscle. J. Cell Biol. 34(1): 15-33.
1968. Obliquely striated muscle. IV.Sarcoplasmic reticulum, contractile appara-tus, and endomysium of the body muscle ofa polychaete, Glycera, in relation to its speed.J. Cell Biol. 36(1): 245-259.
Schneider, A. 1860. Ueber die Muskeln undNerven der Nematoden. Arch. Anat, Phys-iol. u. Wiss. Med. 224-242.
Watson, B. D. 1965a. The fine structure ofthe body-wall in a free-living nematode,Euchromadora vulgaris. Quart. J. Microsc.Sci. 106(1): 75-81.
. 1965b. The fine structure of the body-wall and the growth of the cuticle in theadult nematode Ascaris lumbricoides. Quart.J. Microsc. Sci. 106(1): 83-91.
Watson, M. L. 1958. Staining of tissue sec-tions for electron microscopy with heavymetals. II. Applications of solutions con-taining lead and barium. J. Biophys. Bio-chem. Cytol. 4: 727-730.
Wright, K. A. 1964. The fine structure of thesomatic muscle cells of the nematode Capil-laria hepatica (Bancroft, 1893). Can. J.Zool. 42: 483-490.
. 1966. Cytoplasmic bridges and mus-cle systems in some polymyarian nematodes.Can. J. Zool. 44: 329-340.
Wright, K. A., and N. O. Jones. 1965. Sometechniques for the orientation and embeddingof nematodes for electron microscopy. Nema-tologica 11: 125-130.
Copyright © 2011, The Helminthological Society of Washington

30 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Infectivity of Two Species of Poultry Coccidia After Freezing andStorage in Liquid Nitrogen Vapor
DAVID J. DORAN AND JOHN M. VETTERLINGBeltsville Parasitological Laboratory, Animal Disease and Parasite Research Division,Agricultural Research Service, U. S. Department of Agriculture, Beltsville, Maryland
Kouwenhoven (1967) reported that sporo-zoites of Eimeria tenella (Railliet and Lucet,1891) Fantham, 1909, frozen within sporo-cysts released from oocysts, produced infectionin chickens 24 hours after storage at —79 C.He also found that sporozoites within intactoocysts were noninfective after the same inter-val at -79 and -196 C. Doran and Vetterling(1968) stored E. tenella and E. meleagrimitisTyzzer, 1929 sporozoites, excysted and withinreleased sporocysts, in liquid nitrogen vaporfor one month. After either manner in whichsporozoites were stored, they found that ma-ture schizonts developed in cell culture. Theyalso found that frozen excysted sporozoitesinoculated into birds produced oocysts andlesions similar to those produced in birds inoc-ulated with freshly excysted sporozoites,whereas sporozoites within frozen sporocystsproduced less intense infections than in birdsfed freshly released sporocysts.
This report pertains to the infectivity ofexcysted sporozoites, sporozoites within re-leased sporocysts, and sporozoites within in-tact oocysts of E. meleagrimitis and E. tenellaafter storage in liquid nitrogen vapor for inter-vals up to 4 months.
Materials and MethodsOocysts were collected from droppings of
chicks or turkey poults during the first 72 hrof the patent period. The methods for recoveryand sporulation were the same as previouslydescribed (Vetterling, 1969). All oocystsused were from single suspensions that hadbeen made bacteria-free by treatment withundiluted Clorox (Jackson, 1964). They werestored in Ringer's solution at 3 to 6 C and wereeither 35 days or 2.5 months old when groundoocysts (sporocysts) and sporozoites or oocysts,respectively, were frozen.
Sporocysts were released from oocysts bygrinding with a tissue grinder (Doran andVetterling, 1969). They were placed in
sterile distilled water for 24 hours before freez-ing or excysting sporozoites. Sporocysts weretreated with excystation fluid as previouslydescribed (Doran and Vetterling, 1967), untilmore than 85% of the sporozoites had excysted.
Oocysts, sporocysts, and sporozoites of E.tenella were placed in media consisting of 86%Earle's (Earle, 1943) balanced salt solution,5% lactalbumin hydrolysate (2.5% solution inEarle's B.S.S.), 2% calf serum, and 7% di-methyl sulfoxide (DMSO). Those of E.meleagrimiti-s were placed in media containing88% Medium 199 (Morgan et al., 1950) inHank's B.S.S. (Hanks and Wallace, 1949), 5%chicken serum, and 7% DMSO. The mediawere at 37-43 C when parasites were added.
Suspensions were quickly dispensed in 5-mlamounts into 10-ml round-bottom screw-toptubes using a Cornwall automatic pipette.The tubes were then immediately placed in aliquid nitrogen freezer (Canal Industrial Corp.,Bethesda, Md.; Model 2) and frozen to -80 Cat the rate of 1 degree/min. After the tem-perature had dropped to that of the vapor,the tubes were placed in a cylindrical storagetank (Lansdell Cryogenics, Hyattsville, Md.;Type LR-40) about 4 to 5 inches above liquidnitrogen. Each tube contained 2.5 to 3.5million excysted sporozoites, released sporo-cysts, or intact oocysts.
One tube of oocysts, sporocysts, and sporo-zoites of each species was removed from stor-age at 1 week, 1 month, 3 months, and 4months, respectively. After their contents werequickly thawed by vigorous agitation in warmwater at 43 C, the number of oocysts, sporo-cysts, or sporozoites per respective tube wasdetermined. Groups of 3-week-okl chicks andturkey poults, arranged by weight into lots,were inoculated with either the frozen para-sites or fresh oocysts. Uninoculated groupswere included to compare weight gains. Sporo-zoites (200,000) were inoculated by syringeinto either the duodenum (E. meleagrimitis)
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUiME 36, NUMBER 1, JANUARY 1969 31
or cecum (E. tenella); oocysts (25,000) andsporocysts (100,000) were fed by gavage.
Chicks receiving E. tenella were maintaineduntil 8 days after inoculation. The birds werethen weighed and necropsied. After the cecallesions were evaluated, the ceca and contentswere mixed with potassium dichromate andhomogenized in a Waring blender. Droppingsfrom turkeys that received E. meleagrimitiswere collected between 116 and 192 hr afterinoculation. They were placed in potassiumdichromate and stirred for several hours. At192 hr the surviving poults were weighed,necropsied, and their duodenal lesions eval-uated. After the cecal homogenate and fecalsuspension had been diluted with tap water,oocyst counts were made from samples withthe aid of a Levy counting chamber withFuchs-Rosenthal ruling. The total number ofoocysts recovered from each group was cal-culated, and this was divided by the numberof surviving birds in the group to give an esti-mate of the relative oocyst production persurviving bird.
The anticoccidial index (AI) , as proposedby the Merck technical staff (Merck and Co.,1960), was calculated for each group. Thisappeared to be the best available method forcomparing the results obtained with fresh andfrozen parasites. With this method, the growthand survival of the uninfected control (UC)birds are the standards for comparison, andthis group is assigned an index of 200. Lesionsof the infected birds are scored on the basisof 0 = normal; 1 = detectable; 2 = moderate;and 4 = maximal. A lesion value of 2.5 isarbitrarily assigned to each bird that dies ofcoccidiosis during the experiment (McLoughlinand Chute, 1966). Oocyst production is scoredby the following schedule:
Oocysts Oocystrecovered/surviving index
bird X 10° score
0.1-1.02-56-10
01
102040
The AI is calculated according to the equa-tion AI = (% survival + % UC weight gain)- (mean lesion score X 10 + oocyst indexscore).
ResultsOocysts of both species appeared in excel-
lent condition when examined immediatelyafter thawing. They were similar to freshoocysts in all respects. However, they didnot produce infection when fed to their naturalhost.
The membranes of many frozen sporocystswere either slightly indented or collapsed onone side. A few empty sporocysts were alsofound. It was impossible to determine whethersporozoites within the sporocysts were alive.In their dormant condition, they appearedmorphologically similar to those in freshoocysts. When fed to their hosts, both speciesproduced infection but less severe than thatof their controls as judged by the AI (Table 1).
Excysted sporozoites of both species ap-peared morphologically similar to freshly ex-cysted sporozoites; however, because of theirtypically sluggish motility, it was impossibleto tell whether they were alive. Counts indi-cated a loss of less than 5% per vial. Whensporozoites of both species were injected intotheir natural host, infections equal to or greaterthan those of the controls were produced. TheAI's calculated for E. tenella infections werenearly identical, while those of the frozen E.meleagrimitis infected principals were lowerthan those for the control groups.
Discussion
The infections produced in birds with frozensporocysts were entirely due to sporozoiteswithin the sporocysts and not to free sporo-zoites released by grinding. A small number(~3%) were released, but these were killedduring the 24-hour storage before freezing.We previously found that excysted sporozoiteswill not survive 1 hour in water only. Successwith sporocysts and not oocysts was probablydue to differences in permeability of the oocystand sporocyst membranes to the freezingprotectant.
After 4 months storage, frozen sporozoitesof both species produced infections of equalor greater intensity than those produced by asimilar number of sporozoites in the dosagesof fresh oocysts. However, it is doubtfulwhether this really means that the 200,000frozen sporozoites inoculated into the duo-
Copyright © 2011, The Helminthological Society of Washington

32 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Table 1. Infection of turkey poults and chicks with frozen and nonfrozen Eimeria meleagrimitis andE. tenella.
Eimeria meleagrimitisUninoculated controlFresh oocystsFrozen sporocysts
Uninoculated controlFresh oocystsFrozen sporozoites
Eimeria tenellaUninoculated controlFresh oocystsFrozen sporocysts
Uninoculated controlFresh oocystsFrozen sporozoites
Birds surviving/birds started
10/104/109/10
9/95/93/10
10/1010/1010/10
10/1010/1010/10
Mean wt grain/surviving bird
Grams
12885
125
1219041
17393
173
130133153
%UC*
1006698
1007434
10054
100
100100100
Meanlesionscore
02.01.0
01.52.1
02.20.1
01.41.5
Oocysts recovered/surviving bird Anticoccidial
X 10" index
031.031.0
024.538.5
04.50.2
09.97.5
20046
138
20075
4
200122198
200165167
Uninoculated control.
denum or cecum were all alive and just aspotentially infective or viable as 200,000 within50,000 oocysts given orally. When a dosageof oocysts is given orally, a lot of the sporo-zoites are lost—some because of oocysts fail-ing to rupture in the gizzard, some due tofailure to excyst, and others, after excysting,passing out in the feces. Perhaps similar re-sults were obtained with fresh and frozensporozoites because a loss from the controlequalized a loss in the countable frozeninoculum.
There should be no further deterioration orloss of viability during storage for years atthe temperature of liquid nitrogen vapor (Le-vine and Andersen, 1966). Freeze-storage ofcoccidia (excysted sporozoites or releasedsporocysts) by the method described hereinshould save workers a considerable amountof the time, effort, and expense involved inmaintaining cultures by frequent serial passagethrough the host. It also makes possible (1)the retention of pathogenic and drug-resistantstrains without genetic alteration and (2) theestablishment of a "bank" from which workerscould obtain coccidia for experimental pur-poses. Since the morphology of intact oocystswas not altered by freezing (Kouwenhoven,1967; present study), it is possible that a typespecimen collection could also be established.
Summary
Oocysts, sporocysts released from oocysts,and excysted sporozoites of Eimeria melea-grimttis and E. tenella. were placed in mediacontaining 7% dimethyl sulfoxide, frozen to-80 C at the rate of 1 degree/min, and storedabove liquid nitrogen for intervals up to 4months.
Oocysts produced no infection when fed toturkey poults or chicks. After 3 months, sporo-cysts of both species produced infections, butless than those produced by fresh oocysts.However, inoculation of sporozoites that hadbeen stored for 4 months into the duodenumor cecum of their natural host resulted in infec-tions comparable to those caused by freshoocysts.
Literature Cited
Doran, D. J., and J. M. Vetterling. 1967.Cultivation of the turkey coccidium, Eimeriameleagrimitis Tyzzer, 1929 in mammaliankidney cell cultures. Proc. Helm. Soc. Wash.34: 59-65.
, and . 1968. Preservation ofcoccidial sporozoites by freezing. Nature217: 1262.
, and — . 1969. Survival anddevelopment of Eimeria meleagrimitis Tyzzer.1929 in bovine kidney and turkey intestinecell cultures. J. Protozool. (In press.)
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 33
Earle, W. R. 1943. Production of malignancyin vitro. IV. The mouse fibroblast culturesand changes seen in living cells. J. Am.Cancer Institute 4: 165-212.
Hanks, J. H., and R. E. Wallace. 1949. Re-lation of oxygen and temperature in thepreservation of tissues by refrigeration. Proc.Soc. Exp. Biol. Med. 71: 196-200.
Jackson, A. R. B. 1964. The isolation ofviable coccidial sporozoites. Parasitology 54:87-93.
Kouwenhoven, B. 1967. The possibility oflow-temperature freezing of Eimeria tenellasporocysts. Tijdschr. Diergeneesk. 92(24):1639-42.
Levine, N. D., and F. L. Andersen. 1966.
Frozen storage of Tritrichomonas foetus for5.6 years. J. Protozool. 13: 199-202.
McLoughlin, D. K., and M. B. Chute. 1966.Preliminary laboratory trials with novasat inEimeria tenella infections. Avian Diseases10: 410-12.
Merck and Co. 1960. Technical Bulletin, Am-prolium, 56 pp.
Morgan, J. F., H. J. Morton, and R. C.Parker. 1950. Nutrition of animal cellsin tissue. 1. Initial studies on a syntheticmedium. Proc. Soc. Exp. Biol. Med. 73: 1.
Vetterling, J. M. 1969. Continuous-flowdifferential density flotation of coccidiaoocysts with a comparison to other methods.J. Parasit. (In press.)
Influence of Storage Period on Excystation and Development inCell Culture of Sporozoites of Eimeria meleagrimitis Tyzzer, 1929
DAVID J. DOR AN AND JOHN M. VETTERLINGBeltsville Parasitological Laboratory, Animal Disease and Parasite Research Division,Agricultural Research Service, U. S. Department of Agriculture, Beltsville, Maryland 20705
Prolonged storage of oocysts results in lossof infectivity. There are no reports availablethat indicate whether the decrease in infec-tivity is due to one or more of the following:(1) death of sporozoites within sporocystsduring storage; (2) inability of the sporozoiteto excyst because of nonremoval of the sporo-cyst plug or Stieda body; or (3) inability tosurvive and develop through the first asexualgeneration.
In previous work on cultivation (Doran andVetterling, 1967a, b, 1969), oocysts lessthan 18 weeks old were used. In order to havesuch cultures available, it was necessary topropagate oocysts several times. Consequently,many bacteria-free cultures of different ageswere accumulated. In an attempt to determinesome of the reasons for loss of infectivity ordecreased oocyst production in the host, weexcysted sporozoites from these cultures andinoculated them into cell cultures. This re-port concerns Eimeria meleagrimitis culturesthat were between 5 and 60 weeks old.
Materials and MethodsOOCYSTS: All oocysts were of a single strain
isolated from a single oocyst. They were col-lected on the third day of the patent period,and the method for their recovery and sporula-tion was as previously described (Vetterling,1969). After they had been rendered freeof bacteria by treatment with undiluted Clorox(Jackson, 1964), they were stored in Ringer'ssolution at 3 to 6 C until used.
CELL CULTURES: Cell cultures were of bo-vine embryonic kidney (cell line; 20th pas-sage). They were on 10- by 35-mm coverglasses in 16- by 150-mm Leighton tubes. Onlycultures that were confluent and did not con-tain cell aggregates were used. The growthand maintenance media were the same asbefore (Doran and Vetterling, in press).
EXCYSTATION: Five hundred thousand sporo-cysts, released from oocysts of each age bygrinding (Doran and Vetterling, 1969),were treated with excystation fluid (Doran
Copyright © 2011, The Helminthological Society of Washington

34 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Table 1. Survival and development of Eitneria meleagrimitis.
48 hr
_,
no.
1
2
no.
421
4321
.( weeks )
52753
12293460
„zoites
3,2422,900
920
2,9112,8753,146
751
„zoites
2,5252,101
192
2,4112,1202,477
201
72 hrc , . i
I
100644
897692
2
M
26451
4972180
%
4.84.92.5
5.46.54.21.0
c
zoites
1,7351,803
151
1,7211,5232,311
82
c u; „„»
I
22191
192927
0
M
3259
0
397882
2
%
3.04.10.7
3.36.64.52.4
Per centdecrease*
48 hr
18.223.878.6
12.421.117.873.0
72 hr
44.835.183.5
38.943.323.088.8
* Total number of parasites.t I, immature; M, mature (with merozoites).
and Vetterling, 1967a) for 20 minutes. Countswere then made of the number of empty sporo-cysts, sporocysts containing one and two sporo-zoites, and free sporozoites. Percentage ex-cystation (PE) was calculated by two methods:
Method A PE —
Method B PE =
(number of number of sporo-empty sporo- -j- cysts with onecysts X 2) sporozoite
total number of sporocysts X 2
number of free sporozoites
X 100.
total number of sporozoites(free and within sporocysts)
X 100.
INOCULATION AND MAINTENANCE OF CELLCULTURES: Sporozoites were concentrated andresuspended in 20 ml of inoculation mediumconsisting of Medium 199 in Hank's (Hanksand Wallace, 1949) balanced salt solution +3% calf serum. After adjusting the pH to 7.0—7.2 and thoroughly agitating the suspension,1.2 ml was immediately pipetted into each of10 cell cultures.
At 3.5 hr the sporozoite-containing mediumwas removed. The cells were washed oncewith warm inoculation medium and 5 ml ofmaintenance medium (pH 7.2-7.4) was added.Changes of medium were made when neces-sary in order to maintain the pH at 7.0—7.4.Cultures were kept in an incubator adjustedto alternating intervals of approximately 12 hrat 40.6 and 43 C.
FIXING, STAINING, AND COUNTING: At 5, 48,and 72 hr, cover glasses were removed fromfour tubes in each set of inoculated cultures.They were fixed and stained as previouslymentioned (Doran and Vetterling, 1969).Counts were made at 645 X. They represent
the total number of organisms found by exam-ining every 4th "row" across the length ofeach of four glasses. Variation among the fourcultures ranged from 11 to 31 per cent.
ResultsSimilar results were obtained in cell culture
with sporozoites from oocysts 5 to 34 weeksold (Table 1). There was little difference in:(1) numbers of sporozoites found within cellsat 5 hr; (2) percentage of developmentalstages at 48 hrs; and (3) percentage decreasein the total numbers of parasites after 5 hr.However, the results with oocysts 53 and 60weeks old were decidedly different.
The smaller number of sporozoites fromoocysts 53 and 60 weeks old in cells at 5 hris mostly due to death of sporozoites beforeinoculation of cell cultures. There was littledifference between oocysts of all ages whenthe percentage excystation was calculated byMethod A (Fig. 1). When Method B wasused, however, only about 40% excystationwas obtained for the 53- and 60-week oocysts.This finding indicates that many sporozoiteseither died before or shortly after excystation.At least half of the sporozoites inoculated intothese cultures were dead at the time of inocula-tion. Many sporozoites from the 60-week cul-tures were observed to break up while ejnerg-ing from the sporocyst.
DiscussionData indicate that decreased infectivity of
older oocyst cultures is not due to the inabilityof sporozoites to excyst because of nonremoval
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 35
100 -i
80.
60,
op 40
20
Exp. Exp. 2
26 53 12 29 34 60
AGE OF OOCYSTS (WKS.)
Figure 1. Excystation of sporozoites from oocystsof different ages. A - Method A; B = Method B.
of the sporocyst plug. When calculated on thebasis of empty sporozoites (Method A), thepercentage excystation of sporozoites from allcultures was similar.
The low percentage excystation from theolder cultures when calculated on the basis ofsurviving sporozoites (Method B) and thepresence of degenerating sporozoites emergingfrom sporozoites indicate that many sporozoitesin older oocysts are either dead within thesporocyst or die shortly after excysting. Thebreaking up of sporozoites while emerging fromsporocysts suggests that excystation of sporo-zoites depends on external factors (trypsin,bile) and is probably not due to the secretionof any enzyme by the sporozoite as previouslythought (Doran, 1966).
The lower percentages of development andpercentages of survival after 5 hr suggest that
many sporozoites are also unable to survivewithin cells and develop through the firstasexual generation.
SummarySporozoites, excysted from sporocysts re-
leased from Eimeria meleagrimitis oocysts ofdifferent ages, were inoculated into cell cul-tures of bovine embryonic kidney (cell line;20th passage). Data obtained indicate thatdecreased infectivity of older oocyst culturesis due to: (1) death of sporozoites withinsporozoites during storage; and (2) inabilityof sporozoites to survive and develop afterentering cells.
Literature CitedDoran, D. J. 1966. Pancreatic enzymes ini-
tiating excystation of Eimeria acervulinasporozoites. Proc. Helm. Soc. Wash. 33:42-43.
, and J. M. Vetterling. 1967a. Cultiva-tion of the turkey coccidium, Eimeria melea-grimitis Tyzzer, 1929 in mammalian kidneycell cultures. Proc. Helm. Soc. Wash. 34:59-65.
, and . 1967b. Comparative cul-tivation of poultry coccidia in mammaliankidney cell cultures. T- Protozool. 14: 657-662.
, and . 1969. Survival anddevelopment of Eimeria meleagrimitis Tyz-zer, 1929 in turkey intestine and bovinekidney cell cultures. J. Protozool. (In press.)
Hanks, J. H., and R. E. Wallace. 1949. Re-lation of oxygen and temperature in thepreservation of tissues by refrigeration. Proc.Soc. Exp. Biol. & Med. 71: 196-200.
Jackson, A. R. B. 1964. The isolation ofviable coccidial sporozoites. Parasitol. 54:87-93.
Vetterling, J. M. 1969. Continuous-flowdifferential density flotation of coccidiaoocysts with a comparison to other methods.J. Parasit. (In press.)
Copyright © 2011, The Helminthological Society of Washington

36 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Megodontolaimus New Genus (Nematoda: Chromadoridae),with a Description of Two New Species
R. W. TIMMVisiting Professor, Department of Nematology, University of California, Davis
Two new species of a new genus of marinenematodes of the family Chromadoridae, sub-family Hypodontolaiminae, were found in theBay of Bengal. The smaller species has sixlongitudinal alae and a single esophageal bulb,while the larger species has two lateral alae anda double esophageal bulb. Although the twospecies differ in these prominent characters,they are alike in stomatal structure and sohave been kept in the same genus.
Megodontolaimus new genus
DIAGNOSIS: Chromadoridae, Hypodontolaim-inae. Four long and six short cephalic setae.Cuticle with prominent punctation; two or sixalae present, with fine cross bars. Prominentpharyngeal bulb surrounding stoma, asymmet-rical, much larger on dorsal side. Long ven-tral tooth, turning to left side, with tip hookedinwardly and dorsally. Dorsal tooth two-pronged, with cresentic thickening extendingalong dorsal wall of pharyngeal bulb. Trans-verse elliptical amphids. Ovaries amphidel-phic, with flexures.; single outstretched testis.Spicules cephalate; gubemaculum large, par-allel, with lateral sleeves. Preanal midventralsetae present. Tails conical, with pointed tip.
TYPE SPECIES: Megodontolaimus coxbazarisp. n.
REMARKS: Megodontolaimus is most closelyrelated morphologically to that group of Hypo-dontolaimus with a prominent pharyngeal bulbin which the somatic setae are abundant andat least as long as the body diameter. Themost obvious distinction between the twogenera is the long thin crescentic apophysis onthe dorsal side of the pharyngeal bulb in Mego-dontolaimus, together with the two-prongeddorsal tooth. In all Hijpodontolaimus speciesthe large tooth is said to be dorsal (cf. Wieser,1954, p. 75), whereas in Megodantolaimus itis basically ventral (Fig. 1A, B; Fig. 2A). Thepronounced dorsal swelling of the pharyngealbulb may or may not be present in Hijpo-dontolaimus; if present, it is usually tapering
toward the posterior (e.g. H. steineri Wieser,1954), but may occasionally be as prominentas in Megodontolaimus (e.g. H. inaequalis andH. buetschlii in the illustrations of Bresslauand Schuurmans Stekhoven, 1940, plate 9,Figs. 44 and 45). Hypodontolaimus has fourcephalic setae (may have six setose labialpapillae), whereas Megodontolaimus has ten;in the latter the punctation is larger and moredistinct, while the body is paler in color andthe excretory cell is much less distinct.
Megodontolaimus coxbazari sp. n.(Fig. 1 A; Fig. 2 A-B)
FEMALE (n = 20): Length = 0.58-0.78mm; maximum body diameter = 22-30 p.;esophagus = 99-125 p.; esophagus-vulva =0.21-0.30 mm; vulva-anus = 0.19-0.29 mm;tail = 69-80 p..
MALE (n = 13): L = 0.61-0.74 mm; mbd= 18-22 p.; e = 99-118 p, e-a = 0.44-0.56mm; tail = 58-77 p..
HOLOTYPE MALE: L = 0.69 mm; mbd =19 fr e = 109 p.; e-a = 0.51 mm; t = 67 /*.
ALLOTYPE FEMALE: L = 0.75 mm; mbd =26 p.; e = 115 e-v = 0.28 mm; ovi = 0.19mm; ov2 = 0.13 mm; v-a = 0.28 mm; t = 80 p..
DESCRIPTION: Body outstretched in death,with tail loosely curved to ventral side. Cutic-ular punctations elliptical, larger at anterior ofbody and bordering the alae. Annules inclinedforward in cervical region and backward fromopposite tip of anterior ovary. Lateral alaeextending full length of body from amphids totail tip, crossed by thin transverse bands. Headbroadly rounded; head diameter 16—19 p.; 12fine cheilorhabdions surrounding mouth open-ing. Ten fine cephalic setae, the four sub-median longer setae 15—18 p., the six shortersetae 6-7 p. long. Six setiform labial papillae.Somatic setae along full length of body, butnot grouped in sets at same body level; somaticsetae up to 20 p. long. Amphids at extremeanterior, transversely elliptical (appear to beflattened-spiral in one specimen). Large pha-
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 37
o\ o a:o o o o o o o o:goo
300 00 000 0000 00 0=0000000DO oooo oo 6 ; o o o 6 cfc==ooooooo|
Figure 1. A. Megodontolaimus coxbazari, female head. Megodontolaimus sonadiae: B. Male head.C. En face view. D. Optical section at level of ventral tooth tip. E. Optical section just behind ventraltooth tip.
ryngeal bulb, 24-29 ^ long in female, 24-26 pin male, much more expanded dorsally; con-spicuous oblique pharyngeal muscles. Largeventral tooth, appearing obliquely lateral whenviewed from side; tip turned in toward centerof stoma. Dorsal tooth two-pronged at tip, pro-
truding into cheilostome; long thin crescenticdorsal apophysis along outer margin of pharyn-geal bulb. Large double esophageal bulb, 32—42 ju, long in female, 21-32 ^ in male, withexternal indentation into two parts and inter-nal division. Cardia inconspicuous. Intestine
Copyright © 2011, The Helminthological Society of Washington

38 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Figure 2. Megodontolaimus coxbazari. A. Female esophageal region, ventral view. B. Male tail.Megodontolaimus sonadiae. C. Entire female. D. Male tail.
thin-walled, with very scattered pale yellowishglobules. Amphidelphic ovaries, with flexureup to one-half the distance to vulva; vulva at51-56% of body length; spherical sperm inuterus, 2 ^ in diameter, with dense refringentmatter in form of a crescent. Testis conspic-uous, one-half body diameter in width for mostof its length. Spicules slightly arcuate, 21-22 p, long across arc, with narrow pointed tips.Gubemaculum broad, thickly cuticularized,20-24 p. long, with thin lateral sleeve around
spicules. Three fine midventral setae anteriorto anus, equally spaced. Tails in both sexesconical, tapering to pointed tip; tail 4-4.6 analbody diameters long in female, 3.4—4.8 in male;three caudal glands opening by a simple slit.
TYPE HABITAT AND LOCALITY: Fine slitabove sand, tidal pool, Cox's Bazar, EastPakistan.
HOLOTYPE MALE: Collected by R. W. Timm,22 December 1964; Slide 1080, University ofCalifornia Nematode Collection, Davis.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 39
ALLOTYPE FEMALE: Same data as holotype;Slide 1081, UCNC, Davis.
PARATYPES: Slide CB 53 sent to B. E.Hopper; Slides CB 54-61 in collection at NotreDame College, Dacca, East Pakistan.
DIAGNOSIS: This species is distinguishedfrom the following species by its greaterlength, double esophageal bulb and the pres-ence of only two alae.
Megodontolaimus sonadiae sp. n.(Fig. 1 B; Fig. 2 C-D)
FEMALE (n = 20): L = 0.42-0.65 mm;mbd = 22-35 p, e = 80-118 ^ e-v = 0.13-0.23 mm; v-a = 0.14-0.23 mm; t = 48-75 p..
MALE (n = 20): L = 0.41-0.53 mm; mbd= 22-35 p; e = 73-102 p, e-a = 0.27-0.37mm; t = 54-70 /A.
HOLOTYPE MALE: L = 0.5 mm; mbd = 29p; e = 93 p.; e-a = 0.34 mm; t = 68 /*.
ALLOTYPE FEMALE: L = 0.5 mm; mbd =29 p,; e = 96 p.- e-v = 0.17 mm; ov1 = 112 p.;ov2 = 128 p.; v-a = 0.18 mm; t = 51 p.
DESCRIPTION: Body assuming a spiral formor loose curve in death. Cuticle bearing con-spicuous rounded punctations, larger at an-terior and posterior of body and wider apartthan in previous species. Annules inclinedforward in cervical region and backward fromopposite anterior of ovary. One pair lateralalae extending from midesophagus to midtail;two pairs of submedian alae in region fromesophagus to anus; all alae with fine cross bars.Head broadly rounded; head diameter 16-19p.; 12 fine cheilorhabdions surrounding mouthopening (Fig. 1C). Ten fine cephalic setae,the longer 8-13 p. long, the shorter 3 p, (maybe setiform labial papillae). Somatic setaeextending full length of body, up to 13 p, long.Amphids at extreme anterior, transversely
elliptical. Pharyngeal bulb 23-32 p, long infemale, 19-22 p. in male, expanded dorsally.Teeth as in previous species. Single esophagealbulb, 27-33 p. long in female, 16-22 p, in male.Inconspicuous excretory cell a short distanceposterior to esophageal bulb; inconspicuousexcretory pore between bulb and nerve ring.Yellowish refringent circular to elliptical intes-tinal inclusions. Amphidelphic ovaries, withflexure up to one-half or more of distance tovulva; ova 42 by 22 p.; vulva at 52-56% ofbody length. Spicules highly arcuate, 28—31 p.long across arc. Gubernaculum 18—19 p, long,with lateral sleeve around spicules. Eight mid-ventral short, thick, flabby setae anterior toanus, not equally spaced. Tails in both sexesconical, tapering to acute tip; tail 2.2—4.4 analbody diameters long in female, 2.3-3.5 in male;three caudal glands opening by a simple slit.
TYPE HABITAT AND LOCALITY: Mud of inter-tidal zone, north side of Sonadia Island, Bayof Bengal, East Pakistan. Other locality: St.Martin's Island, Bay of Bengal, East Pakistan.
HOLOTYPE MALE: Collected by R. W. Timm,22 December 1964; Slide 1082, UCNC, Davis.
ALLOTYPE FEMALE: Same data as holotype;Slide 1083, UCNC, Davis.
PARATYPES: Slides 1084 and 1085, UCNC,Davis; Slide So 51 sent to B. E. Hopper; SlidesSo 3, 20, 33, 35, 40, 52, 53 in collection atNotre Dame College, Dacca, East Pakistan.
Literature CitedBresslau, E., and J. H. Schuurmans Stekhoven.
1940. Marine freilebende Nematoda aus derNordsee. Mus. Roy. Hist. Nat. Belg., 1-74+ 12 plates.
Wieser, W. 1954. Free-living marine nema-todes. II. Chromacloroidea. Chile Reports17. Lunds Univ. Arsskr., N.F. Avd. 2, 50:1-148.
Copyright © 2011, The Helminthological Society of Washington

40 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Galvanotaxis of Pelodera strongyloides (Nematoda: Rhabditidae)
FRED H.Department of Biology, University of Louisville, Louisville, Kentucky
Numerous experiments concerning the re-sponses of various organisms in an electricalfield have been carried out within the pastsixty years. Some examples listed by Wardenet al. (1940) of the many different organismsthat have exhibited a galvanotactic responseare certain protozoans, bacteria, slime molds,flatworms, annelids, molluscs, insects, crusta-ceans, and echinoderms.
However, as far as it could be determinedfrom the literature, there are only two reportsof previous work of this nature with nematodes(Caveness and Panzer, 1960; Ronald, 1963);hence, the following experiment was under-taken to determine whether a galvanotacticresponse could be elicited from Peloderastrongyloides.
Materials and MethodsAll the stages of P. strongyloides used in
these experiments were cultured in Petri disheson a blood fibrin nutrient agar medium thathad been inoculated with the bacterium Pseu-domonas fluorescens.
The dauer larvae were tested as one group,and the other larval stages with the adultsconstituted the other group. For each seriesof the experiment, approximately 200 washedworms were placed centrally by means of apipette on the bottom of a glass dish measuring12.2 by 3.8 by 3.3 cm that had been filledwith Ringer's solution.
Two calomel electrodes were constructed foruse in the experiments. Each electrode con-sisted of a 10 by 4 cm shell vial in which wereplaced about 15 ml mercury and 50 ml of asaturated solution of HgCl and KC1. Eachvial was fitted with a two-hole cork stopper.Through one hole was inserted a piece of glasstubing in which was fused a piece of platinumwire, one end of which passed into the mer-cury and the other end extended a short dis-tance above the cork. To conduct the current
1 This work represents a portion of a thesis submitted tothe Graduate School, University of Illinois, in partial ful-fillment of the requirements for the degree, Doctor ofPhilosophy.
from the electrode to the Ringer's one arm ofa U-shaped piece of glass tubing filled withKC1 agar was inserted through the other holeinto the HgCl, KC1 solution, and the other armof the agar bridge was immersed in the Ring-er's solution in the dish. The agar bridge alsofunctioned in preventing excessive polarization.
Direct current was produced by a DC powersupply equipped with filters to reduce theripple. An ammeter was placed in the circuitso that total amperage could be read at anytime and current densities calculated. Decaderesistance boxes were also used when smallamperages were desired.
The orientation of the worms was observedwith a dissecting microscope.
Since it was impossible to determine theexact values for the current passing throughthe worms, the responses are thus described interms of the total amperage and/or currentdensity at a particular moment.
For controls, worms were placed in theglass dish filled with Ringer's solution andobserved for 2 hours in the absence of anelectrical current.
ResultsDAUER LARVAE: It was found that at a total
amperage of 3 ^a (current density of 0.00064//,a/mm2) with a resistance of about 2 meg-ohms, all the dauer larvae showed a definitethreshold response by moving toward theanode. In all the experiments the worms firstmoved like one integrated mass toward theobserver (perpendicular to the lines of force)for about 10 minutes and then in mass towardthe anode. Gradual increase of the total am-perage to 100 p.0. had no effect upon the rateof movement of worms. With undulating andthrashing movements, the dauer larvae traveledthe 6 cm in approximately 2 hours eventuallyaggregating near the anode. The experimentwas repeated several times with similar results.
The presence of the electrical field appearedto affect all the worms simultaneously andwith equal intensity. This showed that gal-vanotaxis was not due to electrolytic products,
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 41
in which case those worms nearest one or theother electrode would exhibit galvanotaxis firstand then the other worms in succession.
Passing current through the solution beforeintroducing the worms had no effect uponorientation or movement. There was no differ-ence in the amperage level at which the thresh-old response occurred or in the rate of migra-tion of the worms toward the anode when thedirection of current flow was reversed.
The control worms which were not sub-jected to any current showed random move-ments.
ADULTS AND OTHER LARVAL STAGES: Theadult worms showed no galvanotactic responsewithin a total amperage range of from 1 ^ato 10 ma. Throughout the experiments theseworms remained within the visual field of thedissecting microscope and distinctly showedvery slow random movements, there being noindication of movement toward either of theelectrodes.
In regard to the nondauer larvae, there wasno change in the random movements until atotal amperage of 32 ^a was reached, at whichlevel the young larvae appeared to orientatevery weakly toward the anode and extremelyslow, undulating movements toward thiselectrode then ensued. A slow constant rateof movement continued until a total amperageof 70 p.a. was reached at which point no furthermovement was evident. A current increase to200 p,a. did not induce any further movement;however, after decreasing the amperage below70 fj.a., a resumption of movement occurred,and the worms eventually aggregated near theanode.
The controls for these stages showed randommovements.
DiscussionThe fact that the larval nematodes showed
a constant rate of movement throughout theamperage ranges indicates that the anodalresponse should not be attributed to electro-phoresis. Had the response been electro-phoretic, a greater rate of movement wouldhave resulted with an increase in current.
It cannot be stated with certainty why thedauer larvae first moved perpendicular to thelines of force before orienting and migratingtoward the anode. Such perpendicular move-
ment was noticeably absent in the response ofthe nondauer larvae.
The cessation of movement toward theanode at 70 /j.a by the nondauer larvae mayindicate some inhibitory effect at this amper-age level.
The results of this work differ from thoseof previous investigators. Caveness and Panzer(1960) working with adult Panagrellus redi-viviis and a number of other nematode generain a field of direct current, reported a galvano-tactic movement of these worms to the cathodeon an agar substrate. These investigators re-corded for P. rediviviis a threshold response at0.02 ma below which movement was not sig-nificantly different from the random move-ment of the controls. On a soil substrate, P.rediviviis was reported to have migrated alsoto the cathode, but in numbers less significantthan those which migrated to this electrode onagar.
Using distilled water and dilute saline solu-tions as media, Ronald (1963) was unable toobtain any directional movement with larvaeof Terranova decipiens in continuous directcurrent, in intermittent direct current or byreversing the polarity. He did record, how-ever, a condition of statis with both polarityreversal and alternating current.
Insufficient knowledge of the physiology andbiochemistry of P. strongyloides, especially un-der these experimental conditions, precludesany accurate interpretation of the responsesobtained in these experiments. However, onebroad explanation for such responses as pro-posed by Hyman and Bellamy (1922) en-deavors to relate metabolic gradients to elec-trical gradients. These investigators suggestedthat differences of potential in organisms, par-ticularly those permanent differences thatoccur along the main axis of animals, are dueto differences in metabolic rate at differentregions, the region of highest metabolic ratebeing the most negative in the external circuitand most positive in the internal circuit. Theyfurther suggested that internal potential differ-ences may account for the galvanotaxis ofmany animals so that in a direct current, ani-mals which are electro-positive internally be-come oriented toward the cathode, while thosewhich are electro-negative, become directedtoward the anode. These workers contend thatsuch bioelectric currents are chemical in origin
Copyright © 2011, The Helminthological Society of Washington

42 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
and probably arise through differences in therates of oxidation-reduction processes in differ-ent regions of the animal.
As a matter of speculation based on theaforementioned considerations, the tactic re-sponse of the larvae of P. strongyloides maybe attributed in part to a rate of metabolismgreater than that which occurs in the adult.Oxidation-reduction processes, occurring at agreater rate in the larvae, could be associatedwith, among other things, growth of the orga-nism, tissue differentiation and development ofthe reproductive system and would producelarger amounts of organic salts and other sub-stances. In an electrical field, the cations ofthese organic salts would be attracted to thecathode, and by nature of their structuralsimplicity and small size, would be able topass through membrane barriers fairly readily.On the other hand, most of the correspondingorganic anions, primarily because of theirlarger size, would be unable to pass throughthe organelle and cellular membranes to theanode. Such a condition may therefore resultin the tactic response of the entire organismtoward the anode as occurred in the case ofthe larvae of P. strongyloides.
Further investigations in the area of galvano-taxis are obviously necessary and may even-tually reveal that in some cases such responsesare an active behavioral process, while in othercases, the responses are of a passive nature.
Summary
The galvanotactic response of the larvalstages of the nematode Pelodera strongyloidesis reported. Ringer's solution was employedas the medium in which the larvae migratedto the anode. A threshold response of 3 .awas recorded for the dauer larvae and one of32 ^u,a for the nondauer larvae. The adultworms showed no galvanotactic response withina total amperage range of from 1 ^a to 10 ma.
The results of two previous experiments con-cerning the galvanotaxis of nematodes arebriefly reviewed, and an explanation, entirelyspeculative, is offered as to why the nematodesin the present experiments migrated to theanode.
Literature CitedCaveness, F. E., and J. D. Panzer. 1960.
Nemic galvanotaxis. Proc. Helm. Soc. Wash.27: 73-74.
Hyman, L. H., and A. W. Bellamy. 1922.Studies on the correlation between metabolicgradients, electrical gradients and galvano-taxis. I. Biol. Bull. 43: 313-347.
Ronald, K. 1963. The effects of physicalstimuli on the larval stage of Terranovadecipiens. III. Electromagnetic spectrumand galvanotaxis. Can. J. Zool. 41: 197-217.
Warden, C., T. Jenkins, and L. Warner. 1940.Comparative psychology, Vol. 2: Plants andinvertebrates. 1070 pp. Ronald Press, NewYork.
I N M E M O R I A M
Charles G. Durbin1919-1969
Member since 1949Recording Secretary 1953
Vice President 1960President 1964
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 43
New Species of Tylenchs Associated with Bark Beetles inNew Mexico and Colorado
CALVIN L.
Members of the superfamily Tylenchoideaare among the most important nematode para-sites and associates of bark beetles and otherinsects throughout the United States. In theopinion of the writer and other workers in thefield, many of the free-living or saprophagousforms associated with insects evolve into trueparasites. At times, the sexual free-living stagesof nematode parasites of insects exhibit strikingresemblances to soil-inhabiting or associatedforms. Numerous species of internal parasitesat some period in their taxonomical evolutionhave been placed in genera which are knownto have only a free-living existence. Ruhm1956 placed a parasite of Pissodes pini in thegenus Stictyhis because of the resemblance ofthe sexual forms of the parasite to the sexualforms of that genus. The parasite had beenpreviously included in the genera Tylenchus,Allantoncma, and Parasitijlenchus. Recentlythe same parasite was placed in the genusSpfaaerutariopsis Nickle 1963. The genusStictyhis Thorne 1941 includes species thatare known only to be free-living, althoughtheir habitat lends credence to Ruhm's desig-nation. The writer has been unable to asso-ciate known forms of Stictyhis with a parasitic-counterpart, but closely related genera havebeen collected from insect habitats.
More recently, Bedding (1967) in his studieson nematode parasites of the horntail genusSirex noted the remarkable similarity of thesexual forms of the parasite to the genusDeladenus. The writer, working with the sameparasite at Rotorua, New Zealand, confirmedthe resemblance. The genus Deladenus Thorne1941, at the time of erection, included twospecies, Deladenus diirus and Deladenusobesus. Deladenus durus was originally de-scribed in the genus Tylenchus and was col-
1 Rocky Mountain Forest and Range Experiment Station,Forest Service, U. S. Department of Agriculture, withcentral headquarters maintained at Fort Collins in coopera-tion with Colorado State University; author is located atAlbuquerque in cooperation with the University of NewMexico.
The writer wishes to thank Mr. Gerald Thorne for hissuggestions on the preparation of the manuscript.
lectecl from the galls of chestnut oak, decayingfungus (Pleurota sp.), and under the deadbark of cottonwood. Original specimens ofDeladenus obesus were taken from the frassof bark beetles under the bark of dead whitefir Abies concolor Lindl. It appears that bothspecies were collected from normal insecthabitats.
As research intensifies on the relation ofnematodes to insects and their ecology, it canbe reasonably expected that more and moreexamples of the similarity of parasites to free-living nematodes will arise, and the role ofthe associated forms will assume increasingimportance.
Six new species of tylenchs were collectedfrom trees infested with bark beetles; theywere either associated with the various stagesof the insect or were residing in a habitat thathad been abandoned by the beetles. All typespecimens are in the nematode collection ofthe Rocky Mountain Forest and Range Experi-ment Station at Albuquerque, New Mexico.
Neoditylenchus puniwopus sp. n.(Fig. 1 A-C)
FEMALE (N = 3): L = 0.93-1.17 mm; a =34.0-44.0; b = 6.2-7.4; c = 31.1-35.2; V =90%.
MALE (N = 3): L = 0.59-0.71 mm; a =39.2-51.3; b = 4.5-5.3; c = 23.5-30.8.
FEMALE: Figure IB. Body slender. Cuticlenearly smooth, transverse striae faintly visiblein younger specimens. Head continuous withbody contour. Lip region twice as wide asdeep. Stylet relatively slender with prominentbasal knobs over one-third longer than widthof head. Esophagus as in Figure IB, meta-corpus spindle-shaped with large central val-vular plates and ending in a well-developedterminal bulb. Nerve ring a body width pos-terior to metacorpus. Excretory pore adjacentto nerve ring. Hemizonid posterior to excre-tory pore. Ovary single, outstretched, quadri-columella two body widths in length. Post-uterine branch a body width in length. Lips
Copyright © 2011, The Helminthological Society of Washington

44 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Figure 1. Neoditylenchus puniwopus sp. n. A, Head; B, Female; C, Male tail.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 45
Figure 2. Neoditylenchus yasinskii sp. n. A, Female; B, Male tail.
Copyright © 2011, The Helminthological Society of Washington

46 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
of vulva slightly protruding in some specimens,transverse. Anal opening barely discernibleunder oil immersion objective. Terminusbroadly rounded.
MALE: With anterior body characters offemale. Testes single, outstretched. Spiculespaired tylenchoid, gubernaculum as in Figure1C, one-fourth the length of the spicules.Bursa enveloping terminus which is subacute.
DIAGNOSIS: Closely related to Neoditylen-chus ahieticolus (Ruhm, 1956) Goodey 1963,differs in body length and width, in its longerand more slender stylet and in the nearlysmooth cuticle.
TYPE LOCALITY: Santa Fe National Forestnear Pecos, New Mexico.
TYPE HABITAT: Associated with the Doug-las-fir beetle Dendroctonus pseudotsugae Hop-kins, in Douglas-fir. Pseudotsugae menziesiivar. glauca (Mirb.) Sudw.
Neoditylenchus yasinskii sp. n.(Fig.2A-B)
FEMALE (N = 3): L = 1.25-1.28 mm; a =32.60-38.60; b = 8.54-8.62; c = 20.77-22.82;V = 89%.
MALE (N = 1): L = 0.94 mm; a = 37.16;b = 6.49; c = 18.8.
FEMALE: Figure 2A. Body nearly straightwhen relaxed. Narrow at anterior end, widen-ing rapidly near the vulva. Cuticle with mod-erately fine transverse striations. Lip regiontwice as wide as deep. Head continuous withcontour of body. Stylet with distinct basalknobs, one-third longer than the width of thehead. Esophagus typical of genus. Nerve ringtwo body widths posterior to metacorpus. Ex-cretory pore three-fourths of a body width pos-terior to nerve ring. Hemizonid immediatelyposterior to excretory pore. Ovary single, out-stretched on some specimens, reaching nearlyto basal bulb of esophagus. Postuterine branchone body width in length. Lips of the vulvaprotuberant. Vulva transverse. Anal openingdiscernible only under oil immersion. Terminussubacute.
MALE: Spicules tylenchoid. Gubernaculumas in Figure 2B. Bursa enveloping tail, ter-minus narrowly rounded.
DIAGNOSIS: Closely related to 'Neoditylen-chus pinophilus (Thome) Goodey 1963. Dif-fers from that species in shorter length, shape
of tail, and gubernaculum. Stylet knobs con-siderably more prominent than pinophilus.Also related to Neoditylenchus panurgus(Ruhm, 1956) Meyl 1961—differing from thatspecies in absence of a lateral field.
TYPE LOCALITY: Associated with Dendroc-tonus ohesus (Mannerheim) in Engelmannspruce Picea engelmanni Parry.
Sychnotylenchus scolyti sp. n.(Fig. 3 A-D)
FEMALE (N = 3): L = 0.71-0.87 mm; a =30.6-33.8; b = 5.5-6.2; c = ?; V = 89-90%.
MALE (N = 3): L = 0.70-0.72 mm; a =52-54; b = 5.2-5.4; c = 31-35.
FEMALE: Figure 3A. Body narrowest atanterior end, widest immediately anterior tovulva. Cuticle almost smooth, transverse stria-tions very faint. Head slightly set off. Lipregion more than twice as wide as deep, laterallips narrower than other four and protrudingbeyond body contour. Figure 3B. Stylet stoutwithout basal knobs, slightly longer than widthof head, Figure 3C. Metacorpus of esophaguswith prominent valvular apparatus, spindle-shaped, muscular. Isthmus slender, ending ina prominent elongate terminal bulb. Nervering one-half body width posterior to medianbulb. Excretory pore one body width anteriorto median bulb. Hemizonid about oppositeanterior end of basal bulb. Ovary outstretched,relatively short. Quadricolumella prominent,approximately one-seventh ovary length. Pos-terior uterine branch 1% body widths long.Vulva transverse, lips protuberant. Anal open-ing not discernible. Terminus broadly rounded.
MALE: With head and esophageal char-acters of female. Testes outstretched. Spiculespaired, tylenchoid. Gubernaculum one-thirdlength of spicules, Figure 3D. Bursa envelop-ing tail. Terminus subacute.
DIAGNOSIS: Differs from Sychnotylenchusiilmi Ruhm 1956 in stylet length, absence ofbasal knobs, form of metacorpus, and absenceof a discernible anal opening. Sychnotylenchusscolyti is in general a smaller species thanS. ulmi.
TYPE LOCALITY: Ft. Collins, Colorado.TYPE HABITAT: Associated with Scolytus
multistriatus Marsh, in American elm, Ulmusamericana L.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 47
Figure 3. Sychnotylenchus scolyti sp. n. A, Female; B, Face view; C, Head; D, Male tail.
Copyright © 2011, The Helminthological Society of Washington
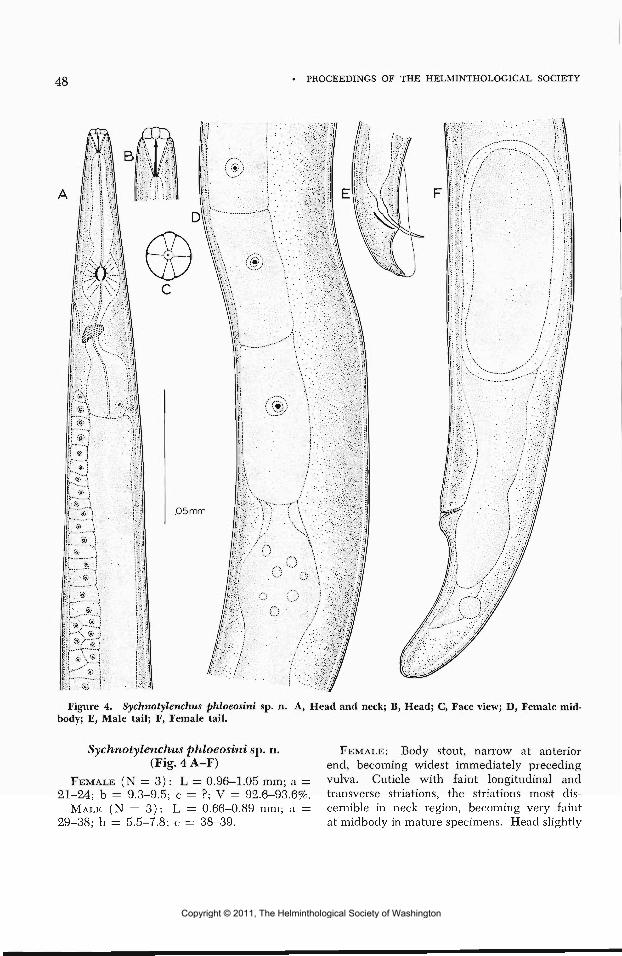
48 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Figure 4. Sychnotylenchus phloeosini sp. n. A, Head and neck; B, Head; C, Face view; D, Female mid-body; E, Male tail; F, Female tail.
Sychnotylenchus phloeosini sp. n. FEMALE: Body stout, narrow at anterior(Fig. 4 A—F) end, becoming widest immediately preceding
FEMALE (N = 3): L = 0.96-1.05 mm; a = vulva. Cuticle with faint longitudinal and21-24; b = 9.3-9.5; c = ?; V = 92.6-93.6%. transverse striations, the striations most dis-
MALE (N = 3): L = 0.66-0.89 mm; a = cernible in neck region, becoming very faint29-38; b — 5.5-7.8; c = 38-39. at midbody in mature specimens. Head slightly
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 49
Figure 5. Aglenchus exiguus sp. n. A, Female; B, Head; C, Male tail; D, Tail showing lateral field;E, Head and neck showing lateral field.
Copyright © 2011, The Helminthological Society of Washington

50 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
set off, flattened, lip region twice as wide asdeep, the lateral lips much narrower than otherfour, protruding beyond body contour in faceview, Figure 4C. Stylet moderately slenderwith basal thickenings, Figure 4B. Procorpusof esophagus stout, narrowing into a prominentovular median bulb in which the valvular ap-paratus is quite distinct. Terminal bulb as inFigure 4A. Nerve ring at the middle of theisthmus. Excretory pore slightly posterior tomedian bulb. Hemizonid not discernible inspecimens examined. Amphidelphic, the ovaryreaching to the median bulb in mature speci-mens, posterior uterine branch as in Figure 4F.Lips of the vulva protuberant. Vulva nearlytransverse, anal opening not discernible. Ter-minus broadly rounded as in Figure 4F.
MALE: Body relatively slender. Head andlips as in female. Testes outstretched, reachingnearly to terminal bulb. Spicules paired, tylen-choid. Gubernaculum one-third to one-halflength of spicules. Figure 4E. Bursa envelop-ing tail. Terminus subacute. Figure 4E.
DIAGNOSIS: Closely related to Sychnotylen-chus intricati Ruhm 1956. Varies from thatspecies in placement of excretory pore andnerve ring, absence of a lateral field, and dis-cernible anal opening.
TYPE LOCALITY: Bandelier National Mon-nument, New Mexico.
TYPE HABITAT: Associated with Phloeo.sinu.ssp. in Juniperus utahensis (Engelm.) Lemm.
Aglenchus exignus sp. n.(Fig. 5 A-E)
FEMALE (N = 2) : L = 0.31 mm; a = 19.7;b = 4.6-4.9; c = ?; V = 64%.
MALE (N = 2): L = 0.31-0.34 mm; a =24.3-25; b = 4.6; c = 3.7.
FEMALE: Figure 5. Body slender ending inan elongate tail. Cuticle coarsely annulated,the annules widest at midbody and interruptedby very prominent lateral fields formed by twobright lines, Figure 5C, D, E. Lateral fieldsextending from two body widths posterior toanterior end to eight body widths anterior toterminus. Head slightly offset, Figure 5B.Lips only faintly discernible in lateral view.Stylet slender with prominent basal knobs,Figure 5B. Esophagus with valvate meta-corpus, the procorpus and metacorpus equalin length to isthmus and terminal bulb. Ex-cretory pore prominent, adjacent to center of
terminal bulb of esophagus. Nerve ring atmiddle of isthmus. Ovary single, outstretched,very short in mature specimens; postuterinebranch short, one-half body width in length.Vulva transverse. Lips covered by vulval flap.Anal opening not discernible. Tail elongate,terminus acute.
MALE: With general body characters offemale. Testis single, outstretched. Spiculestypically tylenchoid. Gubernaculum and bursaas in Figure 5C. Tail elongate, terminus acute.
DIAGNOSIS: Closely related to Aglenchusbryophilus (Steiner, 1914 )Meyl 1961, variesin the shape of the spicules and gubernaculum.Female without discernible anal opening.
TYPE LOCALITY: Mt. Taylor, New Mexico.TYPE HABITAT: Taken from base of Engel-
mann spruce Picea engelmanni Parry infestedwith the Engelmann spruce beetle Dendroc-tonus obesits (Mannerheim).
Neotylenchus nitidus sp. n.-(Fig. 6)
FEMALE (N = 4) : L = 0.79-0.86 mm; a =31.6-37.3; b = 5.7-6.8; c = 11.1-13.3; V =83%.
MALE: Unknown.FEMALE: Figure 6. Body extended when
relaxed. Cuticle with moderately fine trans-verse striations. Lip region hardly set off,continuous with body contour, twice as wideas deep. Stylet moderately coarse with prom-inent basal knobs. Corpus of the esophaguscylindrical, somewhat widened at the centerof its length, then narrowing as it passesthrough the nerve ring, ending in a prominentbasal bulb. Excretory pore immediately pos-terior to hemizonid, both approximately one-half body width anterior to the junction of theesophagus and gut. Ovary single, outstretched,quadricolumella occupying one-eighth of itstotal length. Posterior uterine branch well-developed, approximately one body width inlength, vulva transverse. Anal opening onlyfaintly discernible in some specimens. Ter-minus acute.
DIAGNOSIS: Closely related to Neotylenchusacutus Thorne 1941, varies from that speciesin the prominence of the knobs on the stylet,
2 Nickle (1968) has made Neotylenchus a synonym ofHexattjlus. It is the opinion of the writer that Ncotylenchusis a valid genus. The character of the junction of theesophagus and intestine in members of the group is ofsufficient variation to justify generic rank.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 51
Figure 6. Neotylenchus nitidus sp. n. Female.
Copyright © 2011, The Helminthological Society of Washington

52 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
in the character of the junction of the esoph-agus and gut, and in the presence of a pos-terior uterine branch. It also varies in theabsence of a lateral field.
TYPE LOCALITY: Red Feather Lakes Colo-rado, Roosevelt N.F.
TYPE HABITAT: Abandoned galleries ofDendroctonus obesus (Mannerheim) in Engel-mann spruce, Picea engelmanni Parry.
Literature Cited
Bedding, R. A. 1967. Parasitic and Free-living Cycles in the Entomophagous Nema-todes of the Genus Deladenus. Nature Vol.214: 174-175.
Goodey, T. 1963. Soil and Freshwater Nema-todes. Methuen and Co. Ltd. London, 544pp., illus.
Meyl, A. 1961. Die freilebenden erd und
Siisswassern nematoden (Fadenwiirmer) In.Die Tierwelt Mitteleuropus (Bohmer, Ehr-man and Ulmer) Quelle and Meyer, Leipzig,pp. 1-273, 54 pis., Figs. 1-889.
Nickle, W. R. 1963. Bovienema (Nematoda:Allantonematidae), A New Genus Parasitiz-ing Bark Beetles of the Genus PityogenesBedel with Notes on Other EndoparasiticNematodes of Scolytids. Proc. Helm. Soc.Wash. 30(2): 256-262.
Nickle, W. R. 1968. Observations on Hexa-tijhis viviparus and Neotylenchus abulbosus(Neotylenchidae: Nematoda). Proc. Helm.Soc. Wash. 35: 154-160.
Ruhm, W. 1956. Die Nematoden der Ipiden.Parasitologische Schriftenreihe Jena, No. 6:1-437, Figs. 1-145.
Thorne, G. 1941. Some Nematodes of theFamily Tylenchidae which do not Possess aVulvular Median Esophageal Bulb. GreatBasin Naturalist 2 (2) : 37-85.
The Proposed Synonymy of the Monogenean Genera CleidodiscusMueller, 1934 and Urocleidus Mueller, 1934, with the Proposal ofCleidodiscus bychowskyi sp. n.1
C. E. PRICE2 AND A. MURA3
A total of 43 reports of monogenetic trem-atodes parasitizing North American catfishes(family Ictaluridae) has been recorded. Inall cases, the parasite recovered was eitherCleidodiscus floridanus Mueller, 1936 or C.pricei Mueller, 1936. In light of the numerousreports of these two monogeneans from thegills of several species of Ictaluridae, it ap-pears that close parasite—host relationships existbetween Cleidodiscus parasites and their icta-lurid hosts. The present study includes a de-scription of Cleidodiscus bychowskyi, the thirdspecies of Monogenea to be recovered from amember of this catfish family.
In a comparative study of Cleidodiscus andUrocleidus, Price (1967) expressed the opinionthat these genera were very nearly identicalin morphology. They could be separated from
1 Study supported jointly by the Departments of Biologyof Augusta College and Millersville State College and bya Grant-in-Aid of Research from the Society of Sigma Xi.
'- Department of Biology, Millersville State College,Millersville, Pennsylvania 17551.
3 Department of Biology, Augusta College, Augusta,Georgia 30904.
each other by a single character: the accessorypiece is basally articulated to the cirrus inCleidodiscus whereas these structures existseparately in Urocleidus. Price (op. cit.) de-clined to amalgamate the two genera. Afteradditional consideration, the authors feel thatthis single trait is not of generic magnitude.We consider these genera identical and recom-mend that species presently included underUrocleidus be transferred to Cleidodiscus.
Materials and MethodsHost specimens utilized in this study were
donated by Mr. W. L. McCann of Alexandria,Louisiana, to whom the authors wish to expresstheir thanks.
Hosts were received frozen. After thawing,gills were removed and preserved in 85% iso-propyl alcohol. From this point, gills andrecovered parasites were treated as prescribedby Price (1966). Anatomical terminology em-ployed is that recommeneded by Price andArai (1967) for monopisthocotylean Mono-
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 53
Figures 1-10. Camera lucida illustrations of Cleidodiscus bychowskyi sp. n. 1, whole mount (ventralview); 2, detail of anterior region, showing nature of head organs, cephalic glands, and eyespots with asso-ciated lenses; 3, ventral anchor; 4, dorsal anchor; 5, ventral bar; 6, dorsal bar; 7, 8, hooks, showing twoof the many possible relative positions of the associated domus; 9, cirrus; 10, accessory piece. All figuresexcept Figure 1 drawn to same scale.
Copyright © 2011, The Helminthological Society of Washington

54 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
genea and by Hargis (1958). Measurementswere made according to the recommendationsof Mizelle and Klucka (1953).
Appropriate measurements and illustrationswere made with the aid of a calibrated filarmicrometer ocular and a camera lucida, respec-tively. Average measurements are given first,followed by minimal and maximal values en-closed in parentheses. All measurements areexpressed in microns.
We found that a tincture of Poinceau Sserved as an excellent stain for differentiatinghead organs and cephalic glands.
Cleidodiscus bychowskyi sp. n.HOST AND LOCALITY: Ictalurus punctatus
(Rafinesque), the channel catfish; CalcasieuRiver, near Lake Charles, Louisiana (CalcasieuParish).
NUMBER OF SPECIMENS STUDIED: Ten.TYPES: Holotype USNM Helm. Coll., No.
71238. Paratypes in authors' collections.DESCRIPTION: A dactylogyrid of moderate
size, provided with a thin cuticle. Anteriortwo-thirds of body proper fusiform, taperinguniformly toward peduncle; length 793 (705-878), greatest width of body 162 (144 to209). Cephalic lobes moderately developed.Peduncle and haptor variable in shape, de-pending upon state of contraction, haptorusually well set off from body proper (wholemount: Fig. 1). Two pairs of eyespots, mem-bers of posterior pair larger (Figs. 1, 2).Pharynx prominent, very muscular, and sub-spherical in ventral view. Four pairs of gland-ular head organs, connected to the multilobedcephalic glands by a common duct (Fig. 2).
Two pairs of anchors, one pair in ventralportion of haptor, other pair in dorsal. Allanchors similar in both shape and size (Figs.3, 4 ) . Each composed of: (1) a solid baseprovided with a prominent superficial and ablunt deep root, (2) a solid shaft, and (3) asolid point which meets shaft at a definiteangle. Anchor wings present. Ventral anchorlength 45 (4.1-49), width of base 23 (21-25);dorsal anchor length 47 (45-51), width ofbase 22 (18-25). Bases of each anchor joinedby a simple haptoral bar, the bars similar insize but dissimilar in shape (Figs. 5, 6).
Length of ventral bar 56 (48-63); length ofdorsal bar 60 (53-68).
Hooks 14 in number (7 pairs), all similarin shape and size; hook shape somewhatatypical for Cleidodiscus (Figs. 7, 8). Fivepairs of hooks located on ventral aspect ofhaptor, remaining two pairs on dorsal aspect.Each hook composed of: (1) a solid elongatebase joined to (2) a much narrower shaft, and(3) a sickle-shaped termination provided witha decurved opposable piece. Each hook pro-vided with a domus. All hooks between 14and 17 in length.
Copulatory complex composed of a cirrusand a basally articulated accessory piece (Figs.9, 10). Cirrus equipped with an elongate,heavily sclerotized base. Cirrus tube tapersgradually to a point; length of cirrus in situ80 (73-88), estimated total length 110 (95-129). Accessory piece in form of a slightlycurved sclerotized bar with a flared termina-tion and provided with a deep groove on itsinner surface; length 34 (28-37). Prostaticreservoir single, filled with a yellowish gran-ulated material and emptying into cirrus baseby a fine duct. Vagina, when visible, opensventrally near right body margin; vaginalduct opens into a thin-walled seminal recep-tacle. Ovary somewhat elongate longitu-dinally, larger than the subspherical, post-ovarian testis. Vitellaria composed of darkgranules of essentially uniform size, the vitel-laria forming two broad bands laterally. In-testinal limbs confluent posteriorly.
Discussion
C. bychowskyi does not appear to closelyresemble any known form presently ascribed toeither Cleidodiscus or Urocleidits. It wouldserve no apparent useful purpose to designatespecies possessing bars and/or anchors resem-bling those of the present new species, as agreat degree of homogeneity of these struc-tures exists among most of the included forms.The accessory piece of C. bychowskyi is uniqueamong members of either genus. The hooksof the present form are atypical for bothCleidodiscus and Urocleidus.
DERIVATION OF SPECIES NAME: This parasiteis named in honor of Dr. Boris E. Bychowsky,Director of the Zoological Institute of the
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 55
USSR and author of the most complete accountof the Monogenea thus far published (By-chowsky, 1957). Dr. Bychowsky (personalcommunication) has been a great source ofboth information and encouragement to thesenior author.
Summary
The very similar morphological features ofthe genera Cleidodiscus Mueller, 1934 andUrocleidus Mueller, 1934 have been compara-tively studied. It is proposed that these generaare similar to the extent that they are con-sidered to compose the single genus Cleidodis-cus. It is recommended that species now in-cluded under Urocleidus be transferred toCleidodiscus.
A new species of the North American Mono-genea, Cleidodiscus bychowskyi, recoveredfrom the gills of channel catfishes [letalumspunctatus (Rafinesque) ] captured in Louisi-ana, is described.
Literature CitedBychowsky, B. E. 1957. Monogenetic trema-
todes—their systematics and phylogeny.(Originally in Russian. American translationedited by W. J. Hargis, Jr.) Graphic ArtsPress. Washington, D. C. 506 p.
Hargis, W. J., Jr. 1958. A revised, annotatedlist of terms useful for morphological studiesof monogenetic trematodes. (Mimeographedat Virginia Fisheries Laboratory, GloucesterPoint, Virginia.) 12 p.
Mizelle, J. D., and A. R. Klucka. 1953.Studies on monogenetic trematodes. XIV.Dactylogyridae from Wisconsin fishes. Amer.Midi. Nat. 49(3): 720-733.
Price, C. E. 1966. Urocleidus cavanaughi, anew monogenetic trematode from the gillsof the keyhole cichlid, Aequidens maroni(Steindachner). Bull. Georgia Acad. Sci.24(3): 117-120.
. 1967. Notes on the trematode generaCleidodiscus and Urocleidus. Quart. J. Flor-ida Acad. Sci. 30(1): 61-67.
-, and H. P. Arai. 1967. A proposedsystem of anatomy for freshwater Mono-genea. Canadian J. Zool. 45(6) : 1283-1286.
Studies on Haplobothrium bistrobilae sp. nov.(Cestoda: Pseudophyllidea) from Amia calva L.
G. PREMVATI1
Department of Biology, Florida Agricultural and Mechanical University, Tallahassee, Florida
The genus Haplobothrium Cooper, 1914 ofthe family Haplobothriidae Meggitt, 1924 hasso far only one species, H. globulijormeCooper, 1914, described from only one host,Amia. calva L., 1766. An examination of thesame fish from Florida fresh waters revealedthe presence of a new species of Haploboth-rium. Ten bowfins were examined and allwere found to be infected with this cestode.Four were male fish and had very light infec-tion: each fish harboring not more than threeto four immature worms: one to two with pri-mary scolices and two to three with pseudo-scolices. In the remaining six female fish, thenumber of worms varied from 10-18 with pri-mary scolices and 55 to 65 with pseudoscolices.
1 Present address: Department of Zoology, University ofLucknow, Lucknow, India.
The cestodes were flattened in hot waterand fixed in 10% formalin. The worms werestained for whole mounts with either cochinealor Semichon's carmine or aceto alum carmineor hematoxylin. For sections, worms werefixed in hot Bouin's fluid and stained in hema-toxylin and eosin. Serial frontal and cross sec-tions were prepared from the region of primaryscolex, pseudoscolex, immature strobila, andmature strobila. Osmoregulatory system wasstudied and photographed in living specimens.
All measurements are in microns unlessotherwise indicated.
Haplobothrium bistrobilae sp. nov.(Figs. 1-15)
HOST: Amia calva L., 1766.LOCALITY: Lake Mnnson, Leon County,
Florida.
Copyright © 2011, The Helminthological Society of Washington

56 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
LOCATION: First half of intestine.HOLOTYPE and nine paratypes in USNM
Helm. Coll. Nos. 71209 and 71210.DESCRIPTION: Specimens very active and
show considerable relaxation and contraction,occurring in two phases: primary strobila andsecondary strobila. Primary strobila with truescolex, followed by a growth zone (neckregion), 10—25 mm in length with only ex-ternal segmentation, followed by a long chainof narrow segments. Each segment subdividesanteriorly into three to eight segments of sec-ondary strobila. Anterior segments of sec-ondary strobila, when complete, develop fourauricular lobes: one dorsal, one ventral, andtwo lateral. Original segments with a numberof anterior proglottids break off from primarystrobila and develop into secondary strobila.Thus primary strobila is simply a dividingstrobila which gives rise to secondary strobila.Genital organs never seen to differentiate inproglottids of primary strobila.
Primary strobila has a typical trypanorhyn-chan scolex, with four protrusible and eversibleproboscides. Each proboscis has a proximalspiny base or stump which can also be pro-truded or retracted, and a distal nonspinyelongated tentacle. Proboscides enclosed ininvaginated saclike structures. Cross sectionsshow each proboscis enclosed within a mus-cular sac, and surrounded by a cuticular layer,circular layer and longitudinal layer.
Secondary strobila between 85-100 mm inlength. Anteriormost segment becomes modi-fied into a pseudoscolex, with spines develop-ing on its anterior margin. Segmentation com-plete and begins immediately posterior topseudoscolex. Anterior segments broader thanlong, with distinct four auricular lobes, andthen gradually become longer than broad, andappear beaded to naked eye. Reproductiveorgans appear after about 20—45 segments.With growth of genitalia, internal segmentationstarts to disappear first, and ultimately externalsegmentation also not visible in gravid seg-
ments. Terminal proglottid conical in shape.Testes rounded, medullary in position and
measure 80-100 by 45-50; number of testesin immature proglottids varies from 12—14 oneach side, arranged in two rows, i.e., totalnumber 24-28. This number becomes doubleafter about five or six proglottids, and 24-28testes lie in double alternating rows on eachside. Total number of testes not seen to exceed56 in any proglottid. Vasa efferentia on eitherside join and form vas deferens, submedian inposition and leads to a globular externalseminal vesicle, followed by a cirrus sac en-closing internal seminal vesicle, ejaculatoryduct and cirrus. Cirrus sac measures 250 indiameter; cirrus short, spiny, cylindrical, mus-cular and eversible, opening at level of externalseminal vesicle.
Ovary measures 200—250 by 240, median,and lies at posterior portion of proglottid.Lobes of ovary connected by isthmus; oviductarises from isthmus, runs anteriorly for a shortdistance, and joins common vitelline duct.Vitellaria follicular, medullary, and extendfrom anterior margin to posterior margin ofproglottid on two lateral sides. Follicles mea-sure 40—60 by 40. Two anterior and oneposterior vitelline ducts join in region of ovaryand form a thick short vitelline duct, runningtransversely on either side. Two transversevitelline ducts join in space between twolimbs of ovary and form a very short vitellinereservoir. From this, the common vitellineduct runs posteriorly to join oviduct just beforeootype, which is surrounded by Mehlis' gland.Uterus runs anteriorly as a coiled tube for ashort distance, and then enlarges to form auterine sac, with a permanent uterine pore.Vagina opens on ventral aspect by a circularpore just posterior to cirrus opening. Vaginaenters female genital space and enlarges toform seminal receptacle. Eggs measure 59-60by 25.
Osmoregulatory system may be studied inthree parts: (1) in primary scolex and neck
Figures 1-6. Haplobothrium bistrobilae sp. nov. 1. Camera lucida sketch of primary scolex of livingspecimen to show the proboscides and osmoregulatory system. 2. Pseudoscolex of secondary strobilawith immature proglottids. 3. Two mature proglottids: the first showing vitellaria and the other positionof testes. 4. Enlarged view of female genitalia. 5. Cross section from the region of ovary of a matureproglottid. 6. Cross section from the region of external seminal vesicle of a mature proglottid.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 57
VA
Copyright © 2011, The Helminthological Society of Washington

58 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
region; (2) in immature strobila; and (3) inmature and gravid strobila.
Osmoregulatory canals in primary scolexwith two pairs, a ventral and a dorsal canalon either side. Ventral canals with sinouscourse, run on outside and on reaching basalspiny portion of proboscides, curve inwardly;two dorsal canals also sinuous and run innerto ventral canals, and as they reach anteriorhalf of proboscides, they are connected with atransverse canal. Three small commissures joinventral canal to dorsal canal on either side ofanterior half of scolex. Thereafter dorsal canalruns anteriorly and seen to be confluent withventral canal (Fig. 1). In neck region, twoventral canals run as two separate canals, oneon either side. Dorsal canal single and median,and bifurcates into two canals after formingone or few islets (Figs. 10, 15).
In immature strobila, only three longitudinalcanals run along vertical axis of body, oneventral on either side of body, and a singlemedian dorsal canal (Fig. 13). Dorsal canalslightly thicker at places, otherwise all threecanals practically of uniform width and havesinuous course.
In mature strobila, two ventral canals runlongitudinally in a sinuous course, one oneither side of body. Course of median canalrather unique. It runs both as a single canaland as two bifurcated canals. When presentas two dorsal canals, they run on either side ofgenital organs, and may unite either in sameproglottid or may run as two separate canalsfor a couple of proglottids and then join (Fig.14), thus showing no regularity in bifurcationor union of dorsal canals in mature strobila.
In last proglottid, two ventral canals seen toopen separately by excretory pores a small dis-tance from posterior end. Two dorsal canalsform two excretory vesicles at posteriormostportion of proglottid (Fig. 11). No connectionbetween ventral canal and dorsal canal ofeither side in any proglottid. But when pro-glottids break up and anterior end forms apseudoscolex of secondary strobila, two ven-tral canals connect with single median dorsalcanal without forming any ring canal (Fig. 2).Flame cells numerous in each proglottid, ingroups of four and as many as twenty couldbe counted on either side of one proglottid.
DiscussionGen vis Haplobothrium Cooper, 1914, family
Haplobothriidae Meggitt, 1924, seems to beconfined to only one host, Amia calva L. whichis now found only in North America. H.globuliforme Cooper, 1914, the only species ofthis genus, is so far reported mostly fromnorthern parts of the United States and fromMississippi in the South. This is the first recordof this genus from Florida.
The morphology and systematic position ofH. globuliforme was described in detail byCooper (1914, 1919). Essex (1929), Thomas(1930), and Meinkoth (1947) described itslife cycle and showed that it is similar to thatof pseudophyllideans, involving the develop-ment of procercoid and pleurocercoid, and thatthe characteristic trypanorhynchan scolex de-velops later in the pleurocercoid stage.
Haplobothrium bistrobilae sp. nov. differsfrom H. globuliforme in the following: (1)The total number of testes is only 48-56:24-28
Figures 7-15. Haplobothrium bistrobilae sp. nov. (Photomicrographs). 7. Living primary scolex show-ing completely retracted proboscides. 8. Living primary scolex showing protruded basal spiny portion andretracted tentacles of the proboscides. 9. Frontal section of primary scolex. 10. Osmoregulatory system inneck region of living specimen. Primary strobila, showing bifurcation of median dorsal canal. 11. Pos-teriormost proglottid in living specimen showing excretory pores of ventral canals and two excretoryvesicles of dorsal canals. (Note: Figure retouched.) 12. Cross section of primary scolex showing pro-boscides. 13. Osmoregulatory system in living immature proglottids, showing two ventral canals andsingle median dorsal canal. 14. Osmoregulatory system in living mature proglottids, showing two ventralcanals and union and bifurcation of dorsal canal. 15. A portion of dorsal canal of neck region of livingprimary strobila to show islets before bifurcation into two dorsal canals.
Abbreviations used: B, basal spiny portion of proboscis; C, cirrus sac; DC, dorsal canal; G, male genitalpore; L, auricular lobe; M, Mehlis' gland; O, ovary; P, proboscis; PS, pseudoscolex; R, seminal recep-tacle; S, external seminal vesicle; T, testis; U, uterus; UP, uterine pore; V, vitellaria; VA, vagina; VC,ventral canal; VD, vitelline duct; VP, vaginal pore.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 59
. vr.;;a*«ss..'••, • •v-.-r/ry-i
Copyright © 2011, The Helminthological Society of Washington

60 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
on each side of proglottid arranged in twoalternate rows. In H. globuliforme, the num-ber of testes is 80:40 on each side. (2) Uterinepore persists even in gravid proglottids, butceases to exist in those of H. globuliforme.(3) The proboscides are completely retract-able, while in H. globuliforme the stump orthe base remains permanently protruded. (4)The disposition of vitelline ducts differs fromthat of H. globuliforme. (5) The eggs aresmaller as compared to those of H. globuli-forme. (6) The course of dorsal canal ("me-dian vessel" by Cooper) does not remain singleas in H. globuliforme but becomes double atplaces in mature and gravid proglottids. Theterminal proglottid also shows two distinctvesicles of dorsal canals at posteriormost end.The two openings of ventral canals are seento lie separately on the lateral sides of thelast proglottid.
SummaryHaplobothrium bistrobilae sp. nov., the sec-
ond species of the genus HaplobothriumCooper, 1914, family Haplobothriidae Meggitt,1924, is described from Amia calva L. fromFlorida fresh waters. It is characterized byhaving 48-56 testes (24-28 on each side);permanent uterine pore; completely retract-able proboscides with spiny base and nonspinydistal tentacles, and a unique course of dorsalosmoregulatory canal. Photomicrographs ofthe osmoregulatory system studied in livingworms show that the two ventral canals havea sinuous course throughout the primary andsecondary strobila. The dorsal canal is singlein immature proglottids but bifurcates at placesin mature and gravid proglottids of secondarystrobila. The terminal conical proglottid showstwo distinct vesicles of dorsal canals and twoexcretory pores of ventral canals.
Haplobothrium bistrobilae sp. nov. differsfrom the only other species, H. globuliformeCooper, 1914, in the number of testes, pres-ence of permanent uterine pore, size of eggs,in having completely retractable proboscides,and in not having a single continuous dorsalcanal.
AcknowledgmentsThe author wishes to thank Professor Robert
B. Short of the Florida State University forthe use of his personal library; Dr. N. A.Meinkoth, Department of Biology, SwarthmoreCollege, Swarthmore, Pennsylvania, for thegenerous supply of slides and fixed materialof Haplobothrium globuliforme for purposes ofcomparison, and Dr. W. W. Becklund of theBeltsville Parasitological Labortaory for pro-viding type and paratype slides of H. globuli-forme from USNM collection.
Literature CitedCooper, A. R. 1914. A new cestode from
Amia Calva L. Trans. Roy. Canad. Inst. 10:81-119.
. 1914. On the systematic position ofHaplohothrium globuliforme Cooper. Trans.Roy. Soc. Canad. 8: 1-5.
1919. North American pseudophyllid-ean cestodes from fishes. Illinois Biol.Monogr. 4: 288-541.
Essex, H. E. 1929. The life cycle of Haplo-bothrium globuliforme Cooper, 1914. Science69(1800): 677-678.
Meinkoth, N. A. 1947. Notes on the lifecycle and taxonomic position of Haplo-bothrium globuliforme Cooper, 1914, a tape-worm from Amia calva L. Trans. Amer.Micr. Soc. 65: 256-261.
Thomas, L. J. 1930. Notes on the life cycleof Haplobothrium globuliforme Cooper, atapeworm of Amia calva L. T. Parasit. 16:140-145.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 61
Reesimermis nielseni gen. et sp. n. (Nematoda: Mermithidae)Parasitizing Mosquitoes in Wyoming
YUAN-HWANG TSAI1 AND ALBERT W. GRUNDMANN
Tropical Disease Research Center, St. Clare's Hospital, New York City, New York, andDepartment of Environmental Biology, University of Utah, Salt Lake City, Utah
There has been a renewed interest in theparasites of mosquitoes as the search for orga-nisms that could prove valuable in biologicalcontrol is pursued. As a result of such a studyin northern Utah and southern Wyoming re-gions, a species of mermithid nematode wasfound inhabiting six mosquito species at asite near Lone Tree, Uinta County, Wyoming.Following an extensive study of the life his-tory, pathogenesis in mosquitoes, and mor-phology of this organism, a new genus andspecies name is proposed.
Mermithid nematodes have been reportedfrequently from both larvae and adults of mos-quitoes in North America and elsewhere. Jen-kins (1964) listed four defined species, Lim-nomermis aquatilis Dujardin from larval Anoph-elines in France, Agamomermi-s culicis Stilesfrom adult Aedes sollicitans (Walker) from theUnited States, Paramermis canadensis Sternerfrom Aedes vexans (Meigen) and A. strictusin British Columbia, Canada, and a Mermissp. from larval Aedes aegijpti in Africa(Gendre 1909). Other records of Mermis sp.are: Muspratt (1945) in larval and adultAnopheles, lyengar (1927) in India andWalandouw (1934) in Sumatra. At leastfifteen additional occurrences in mosquitoeshave been reported in North American liter-ature.
Other mosquito-inhabiting species that havebeen described are Hydromermis churchillensisWelch 1960 from larval Aedes communis (DeGeer) from Manitoba, Canada. This specieswas also reported from Canada by Jenkins andWest (1954) from A. communis, A. impiger(Walker), A. nigripes (Zetterstedt), and A.pionips Dyar. Smith (1961) reported what isassumed to be this same species from larvalA. pullatus in Colorado. Welch (1964) de-scribed Romanomermis iyengari from Anoph-eles subpictiis Grassi in India. A species of
1 Present address: Microbiological Department, BristolLaboratories, Syracuse, New York 13201.
Gastromermis was reported by Cox (1966)from Anopheles funestus Giles in west Africa.
Welch (I960) reviewed the status of re-ported mosquito-inhabiting mermithids fromNorth America and proposed referring Para-mermis canadensis to species inqiiirendae be-cause it was indicated that the description hadbeen based on immature specimens. He fur-ther stated that a definite status for Agamo-mermis culicis would be unwise at this timesince the female was unknown and thus thegeneric diagnosis insufficient. Reviewing theworld status of mermithids inhabiting mos-quitoes, three species belonging to three generacurrently are determined valid, while four maybe determined only to genus.
The systematics of the Family MermithidaeBraun 1883 is difficult due to the scatteredliterature and observations on immature forms.Best references are Filipjev (1934), Filipjevand Schuurmans (1941), and the work of anumber of Russian workers that were nicelyreviewed by Welch (1965).
In North America, Johnson (1963) erectedthe genus Octomyomermis from Chironomids.Poinar (1964) established Orthomermis, alsofrom Chironomids. Welch (196()a, 1962b,1963a, 1964, 1965) has described species fromsix genera in Simulid larvae.
General DescriptionGenus Reesimermis, gen. n.
Reesimermis: Mature adult worms elongate,cylindrical, with obtuse ends in both malesand females. Cephalic papillae consisting of acrown of six in a hexagonal position, whichmay represent two lateral and four submedialcephalic nerve endings. Eight longitudinalhypodermal chords in dorsal, ventral, lateral,and submedial positions (Fig. 10). Spiculespaired, slightly curved, medium-sized, fusedposteriorly for more than half of their length(Fig. 8). Vagina straight, of medium length,muscular and pear-shaped. Cuticle smooth, no
Copyright © 2011, The Helminthological Society of Washington

62 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
criss-cross striae on the middle cuticle layer.Mouth opening terminal. Postparasitic juve-niles with tail appendage. Aquatic habitat.
DiagnosisReesimermis is the second genus to be de-
scribed in the family Mermithidae possessingtwo spicules that are fused posteriorly for morethan half their length. Four established generaare found that possess eight longitudinalchords, and Reesimermis differs from each ofthese in several distinct anatomical features.The genus Hydromermis, which contains H.churchillensis and H. contorta, differs fromReesimermis in the shape of the vagina whichis S-shaped in Hydromermis but straight andpear-shaped in Reesimermis, and in the mor-phology of the spicules of which there are twofused for half of their length in the new genusand described as single in Hydromermis. How-ever, H. churchillensis and H. contorta possesstwo spicules fused for more than half theirlength in contradiction to the generic charac-ters but have the S-shaped vaginal characteris-tics of Hydromermis. These two species maybe considered as intermediate evolutionaryforms occurring between the separate, pairedspicules of Isomermis Comaii 1953 and thesingle spicule of Hydromermis. Both generaare similar in other characteristics.
The genus most closely related to the newgenus is Romanomermis which differs pri-marily in having two completely separatespicules. Both genera inhabit mosquitoes. An-other genus appearing in the literature thatshould also be considered is Octomyomermis,but when the published description was com-pared with that of Romanomermis, the twoseemed to be almost identical. Since no speci-mens of either group were examined by the
authors, it is not possible to accurately statethat Octomyomermis should be synonomizeclwith Romanomermis. Other genera possessingpaired spicules, pear-shaped vagina, and eightlongitudinal chords are Allomermis and Ortho-mermis, but these differ from each other andfrom Hydromermis in the arrangement ofcephalic papillae. The following key separatesthese genera from Reesimermis:1. Four cephalic papillae in one plane;
two spicules completely separated;criss-cross striae on the cuticle;mouth opening ventral Allomermis
Different arrangement and number ofcephalic papillae; two spicules sep-arated or fused posteriorly for morethan half of their length; no criss-cross striae on the cuticle; mouthopening terminal 2
2. Two lateral cepalic papillae in oneplane with six papillae in ring un-derneath; two spicules separated ....
OrthomermisSix cephalic papillae in one plane; two
spicules separated or fused morethan half of their length 3
3. Paired spicules separatedRomanomermis (Octomyomermis)
Paired spicules fused for more thanhalf of their length Reesimermis
Reesimermis nielseni sp. nov.GENERAL: All following measurements are
in millimeters unless otherwise noted. Wormlong and slender with smooth cuticle. Criss-cross fibers in middle cuticular layers absent.Cuticle thin, 5.5—7.7 /JL in adults, and 2—3 /j. inpostparasitic larvae. Head convex, roundedwith six cephalic papillae consisting of foursubmedian and two lateral papillae arranged
Figures 1-10. 1. Lateral view of postparasitic male juvenile tail. T, trophosome; CP, caudal appendage(X 100). 2. Lateral view of postparasitic female juvenile tail (X 100). 3. Lateral view of female post-parasitic juvenile vagina. GP, genital primordium (X 150). 4. Lateral view of vagina (mature femaleworm). O, ova; U, uterus; V, vulva; Va, vagina (X 200). 5. Face view of mature worm head. CP, ce-phalic papillae; M, mouth opening (X 400). 6. Dorsal view of mature female head. CP, cephalic papil-lae; A, amphid (X 500). 7. Lateral view of mature female tail. T, trophosome (X 100). 8. Lateral viewof mature male tail. Sp, spicule; Gp, genital papillae (X 100). 9. Ventral view of mature male tail. Sp,spicule; Gp, genital papillae (X 100). 10. Cross section in anterior region of female. D, dorsal hypoder-mal chord; L, lateral hypodermal chord; R, reproductive organ; S, submedial hypodermal chord; T,trophosome; V, ventral hypodermal chord (X 400).
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 63
8
Copyright © 2011, The Helminthological Society of Washington

64 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
hexagonally in one plane and in a slight de-pression of cuticle (Fig. 5). Mouth terminalin slight depression of cuticle. Small neckconstriction visible behind papillar depression.Amphids located behind cephalic papillaepouch-shaped, pores small, situated about2-4 p, behind lateral papillae. Ligament con-necting amphids absent. Oesophagus folded,diameter 4.5-5.5 p. (average 5 /*) near mouthopening, and 4 ^ along main tract; extendsover three-fourths of body length. Nerve ring265-290 p. from mouth. Excretory like glands0.66—0.85 mm from mouth opening. Sticho-cytes beside oesophagus numerous and gland-ular-shaped. Lateral hypodermal chords withthree cells (a few with paired cells). Maleand female worms have obtuse terminus andare slightly curved ventrally. Free-living infec-tive juveniles, postparasitic and parasiticjuveniles retaining needle-shaped caudal ap-pendage.
i. FemaleSeventeen specimens. Body length 15.10
(11.0—21.2), stout, blunt at posterior end,tapering at anterior end. Greatest width ofhead at level of papillae 0.060 (0.055-0.065);at vulva 0.188 (0.160-0.225); and at tailmeasured at the terminus of trophosome 0.141(0.125-0.195). Arnphid 13 by 5 p.. Distancefrom anterior ovary to mouth opening 0.84(0.32-1.33), that of posterior ovary to baseof tail 0,57 (0.43-0.69).
Vulva, 45.7% (41.1-53.1%) of body length,a transverse opening slit (0.049) in ventralview, straightly located in lateral view, lead-ing to muscular pear-shaped vagina that isstraight thick-walled tunnel, 0.112 (0.095-0.130) in length, connected to opposed amphi-delphic musculated uteri, 0.070 in width and0.190 (0.180-0.198) in length.
Rate of anterior ovary to body length 41.8%(37.4—46.2%) and that of posterior ovary tolength is 48.7% (42.3-53.9%), so v =41-845.748-3. Length of posterior end oftrophosome to base of tail 0.150 (0.100-0.170), that of anterior end of trophosome todegenerate mouth opening 0.420; no analopening (Figs. 4, 6, and 7).
ii. MaleEleven specimens. Body length 10.2 (6.2-
14.10). Head width measured at level of
papillae 0.050 (0.048-0.053); width at middlebody 0.144 (0.120-0.160); and width at anus0.116 (0.096-0.130). Tail length 0.157(0.140-0.175). Amphids same as in female.Paired spicules, fused posteriorly for 63-68%of their length, 250 (205-310) p. in length.Individual bases of the separated proximalends 22-26 p.. Width of single separate spic-ule 4.4 p, at midpoint, and that of the middlefused part 6.5 p,; gently curved, tapering tosharp pin-point. Testes two parallel tubesrunning to middle of body possessing someflagellated sperm. Three rows of pregenitalpapillae. Median row with 19—22 papillae,lateral row with 16-19 papillae, postgenitalpapillae irregular, clustered and tiny. The twolateral papillar rows converge close to medianrow at anal area (Figs. 8, 9).
iii. EggsFlattened ellipsoid shaped, 84 by 74 (80-90
by 70-80) p., with cross-section 74 by 68 (70-80 by 63-70) p.. Egg shell thin, transparentand sticky outside, with very thin vitellinemembrane beneath the shell (visible only inpreserved specimens). Eggs unembryonatedwith one egg cell when deposited. Size of eggsin ovaries smaller and irregular in shape,crowded side by side.
iv. Preparasitic juvenilesBody length 0.56-1.20; width at head region
6-7 p., width at the middle of body trunk 15-17 p,. Onchostylet sharp and thin, about 12-14p, in length. Numerous granular cells in body.Head rounded, tail slender, long, whiplike.v. Parasitic juveniles
Body length and width vary with state ofdevelopment. In 28 specimens measured,length 0.88-13.4; greatest width 0.034-0.125.Head rounded, tail long, slender.
vi. Postparasitic juvenilesBody length 7.4-25.5; greatest width 0.103-
0.190. Ratio of head to body width 0.31-0.44.Anterior trophosome arising 0.370 (0.310-0.420) from head end, posterior trophosomearising 0.085—0.200 from base of caudal ap-pendage. Caudal appendages 0.150—0.190long. Sexes are distinguishable (Figs. 1, 2).
SPECIMENS DEPOSITED: Holotype—10 fe-male adults and 10 male adults in the Uni-versity of Utah Parasitological Collection.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 65
Table 1. Seasonal fluctuation in female-male ratio (FMR) for postparasitic juvenile populations ofReesimermis nielseni.
Female
collected
21 May 196612 June 196619 June 196628 June 1966Total worms examined
No ofworms
14261127761
1,091
No.
3232710529
493
Per cent
22.5353.5037.9047.5445.20
Male
No.
11028417232
598
Per cent
77.4746.5062.1052.4655.80
FMR*
0.291.150.610.910.824
* FMR is obtained by dividing number of females by number of males
TYPE HOST: Aedes communis (De Geer).ADDITIONAL HOSTS: Aedes cinereus Meigen,
A. fitchii (Felt and Young), A. increpitus Dyar,A. pullatits (Coquillet), and Culiseta impatiens(Walker).
LOCATION: Haemocoel of host larvae, occa-sionally in the pupae and adults of Aedesincrepitus and A. pullatus.
LOCALITY: Lone Tree, Uinta County, Wyo-ming. Elevation 8,000 feet. Collected May-June 1965-66.
Morphological differences betweenthe male and female
postparasitic juveniles
The chief morphological differences be-tween premale and prefemale postparasiticjuveniles obtained from mosquito larvae havebeen briefly described by Welch (1960) asbeing the larger size of the female and pres-ence of the genital primordium of the vaginain the middle of the female body. Knowledgeconcerning the differential diagnosis of themale and female free-living postparasitic stageis inadequate because in cases of superinfec-tion, male and female worms are often hard toseparate on the basis of size, and in cases ofsingle or double infections, the males are quitelarge. The presence of the genital primordiumof the vagina is difficult to observe in pre-served material and may be very difficult todetect in live specimens. Therefore, an at-tempt was made to distinguish the sexes ofthe postparasitic juveniles obtained followingemergence from the bodies of mosquito larvaeby means of the morphology of the caudal ap-pendages and primordium of the vagina.
The caudal end of the postparasitic larvaewas found to be characteristically different inmales and females, with that of the premalebeing thin and short (150-160 ^ and that of
the prefemale stouter at the base and longer(170-190 /A) . Also the distance from the endof the terminal trophosome to base of thecaudal appendage in the premale is muchgreater (170-200 ^ Fig. 7) than in the pre-female (85-130 ^ Fig. 8). The morphologyof the caudal end is thus a useful characterthat can be observed under the low power of adissecting microscope.
The primordium of the vagina, located nearthe middle of the body (Fig. 3) has beenpreviously used to separate premale and pre-female worms. It is recognizable as a smallslit appearing beneath the juvenile cuticle thatdoes not open to the outside at 46.5% of thedistance from the anterior end. This featureis difficult to recognize in active specimens.
Size differences between premale and pre-female worms can be used for rapid identifica-tions under some conditions, but should beapplied in conjunction with the previousmethods. Postparasitic juvenile females arelonger and stouter, averaging 25 mm in lengthwhile juvenile males are shorter and thinner(average 15 mm in length). Sometimes twoprominent incisions holding big excretorylikegland cells are visible on the ventral side, justunder the stichosome about one-tenth to one-twelfth distance from the anterior end. Theseare far more prominent in premales.
Female—male ratio (FMR) inpostparasitic juveniles
Wide fluctuation appeared in sex ratioswhen postparasitic worms were sexed at dif-ferent times during the season (Table 1).Males made up 77.4% of the population on21 May 1966, dropped to 46.5% on 12 June,increased to 62.1% 19 June, and 52.46% on28 June. The fluctuations correlated well withthe size of worm burdens per host with the
Copyright © 2011, The Helminthological Society of Washington

PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
worms being predominately males when theworm burden was greater than four and almostexclusively female when less than four werepresent. This phenomenon has been describedpreviously by a number of workers (Christie,1929; Johnson, 1955; Couturier, 1963; andParent!, 1965) who pointed out that sex de-termination in mermithids may be environ-mentally controlled with crowding being theprimary factor. Another factor was introducedby Strelkov (1964) who showed that sex ofthe parasite corresponds to that of the host inFilipjevimermis singularis. No attempt wasmade to resolve the latter in this study.
Female-male ratios were obtained both byexamination of free-living postparasitic juve-niles that had emerged spontaneously fromhosts in the laboratory and through dissectionof infected hosts. FMR were similar in bothphases done on the same days, indicating thatit would be possible to predict populationchanges through examination of free-livingindividuals in soil samples from the bottomof ponds, and thus to provide data of valueto potential biological control of mosquitoes.FMR ratios are calculated by dividing thenumber of females by the number of males,with a low FMR being less than one (Table 1).
Life history
The life history of R. nielseni is similar tothat described by Welch (1960a) for Hydro-mermis churchillensis, a parasite of Aedescommunis in Canada.
Unembryonated eggs are deposited in thebottom soil of shallow pools. Observationsshowed embryonation to be evident in 5-10days and the eggs contained fully developedlarvae seven days later. The larvae do nothatch upon completion of development undercold dry field conditions* but remain dormantuntil favorable conditions are present. Theeggs are vulnerable to desiccation so that dry-ing is immediately fatal if the duration is morethan a few seconds, but eggs appear to beresistant to freezing winter temperatures insoil.
In the type locality, snow is present fromOctober to April when melting snow waterforms the pools in which the mosquito hosts
* The eggs, upon completion of development, couldhatch after 1.5-2 months in soil water at 18-20 C in thelaboratory.
develop. Approximately 2 weeks later thelarvae emerge from the eggs and use theirlong, slender tails for swimming in search of ahost. The larvae can exist for about 4 daysunder April and May temperatures and readilyattach by their onchostylets to the mosquitolarvae that appear at this time. Penetrationinto the host body cavity takes several min-utes after which the larvae gradually becomemotionless.
The parasitic juveniles grow rapidly and arelate stage juveniles in 10-15 days. Emergencefrom the host occurs soon after this stage ofdevelopment is reached. The worms emergeby pushing the anterior end through the con-junctivum between the segments of the thorax,abdomen and neck. Complete emergence takesabout 1 minute and is fatal to the host. Thefree-living postparasitic juveniles burrow intothe bottom soil where they become motionlessfor 2-4 weeks. Molting occurs at the end ofthis period with the males preceding the fe-males. Development and sexual differentia-tion are slow and it is usually 1—2 monthsbefore copulation occurs. Another 10-30 dayselapse before egg production occurs and thenumber of eggs produced depends upon thefemale size and the size of the trophosomewhich was determined by the conditions ofparasitism depending mainly on the numberof larvae that inhabit a single host. A largefemale can produce as many as a thousandeggs. After oviposition, the female dies in5—6 days, having exhausted the food storedin the trophosome. Males survive longer thanthe females since they require little storedmaterial for sperm production.
Reesimermis nielseni has one generation peryear, a condition well adapted to the lifecycles of its hosts that are "single brooded"mountain Aedes species confined to the higheraltitudes.
Summary
All life stages of Reesimermis nielseni, gen.et sp. n., a parasite of mosquitoes from LoneTree, Wyoming, are described. A key to theclosely allied genera of Mermithidae is in-cluded. The type host is the larva of Aedescommunis De Geer but the species was alsofound parasitizing Aedes cinereus Meigen, A.fitchii (Felt and Young), A. increpitus Dyar,A. pullatus (Coquillett), and Culiseta im-
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 67
patiens (Walker). A method of sexing post-parasitic juveniles is described as well as theseasonal fluctuation obtained in the female-male sex ratios during May and June of 1966.The life history is included and shows onegeneration per year.
Literature CitedChristie, J. R. 1929. Some observations on sex
in Mermithidae. J. Exp. Zool. 53: 59-76.Couturier, A. 1963. Recherches sur des Mer-
mithidae, nematodes parasites du hennetoncommun (Melolontha melolontha L.). Ann.Epiphities 14: 203-267.
Coz, J. 1966. Contribution a 1'etude du para-sitisme des adultes d' Anopheles funestus parGastromermis sp. (Mermithidae). WHO/EBL/66.76, pp. 1-11.
Filipjev, I. N. 1934. The classification of thefree-living nematodes and their relation tothe parasitic nematodes. Smithsonian Misc.Coll. 89(6): 1-63.
, and J. H. Schuurmans Stekhoven, Jr.1941. A manual of agricultural helminthol-ogy (E. J. Brill, Leiden, Netherlands.) 878 pp.
Gendre, E. 1909. Sur des larves de Mermisparasites des larves du Stegomyia faciata.Bull. Soc. Path. Exct. 2: 106-108.
lyengar, M. O. T. 1927. Parasitic nematodesof Anopheles in Bengal. Far Eastern Assoc.Trop. Med., Trans. 7th Congr. Brit. India.3: 128-133.
Jenkins, D. W. 1964. Pathogens, parasites andpredators of medically important arthropods.Wld. Hlth. Org. Organisation Mondiale dela Sante, 150 pp.
, and A. S. West. 1954. Mermithidnematode parasites in mosquitoes. Mosq.News, 14: 138-143.
Johnson, A. A. 1963. Octomyomermis itascen-sis gen. et sp. nov., a parasite of Chironomusplumosus (L.) . Trans. Am. Micr. Soc. 82:237-241.
. 1965. Hydromermis itascensis sp. n.(Nematoda: Mermithidae), a parasite ofGlyptotendipes labiferus (Say) (Diptera:Chironomidae) and the taxonomic status ofGastromermis. J. Parasit. 51: 53-56.
Muspratt, J. 1945. Observation on the larvaeof tree hole breeding Culicini (Diptera:Culicidae) and two of their parasites. J.Entomol. Soc. S. Africa 8: 13-20.
Parenti, U. 1965. Male and female influenceof adult individuals on undifferentiated larvaeof the parasitic nematode Paramermis con-torta. Nature 207(5001): 1105-1106.
Poinar, G. O. 1964. A new nematode, Ortho-mermis oedobranchus gen. nov. (Mermith-idae), parasitizing Smittia larvae (Chironom-idae) in England. Nematologica 10: 501—506.
Smith, M. E. 1961. Further records of mer-mithid parasites of mosquito larvae. Mosq.News, 21: 344-345.
Strelkov, A. 1964. Biology of Filipjevimermissingularis sp. nov. found in the RybinskiReservoir. Vestnik Leningr. Univ. No. 3,Ser. Biol. 19(1): 55-69.
Walandouw, E. K. 1934. Nematoden als bes-trijders von anopheles larven. Geneesk Tijd-schr. Ned.-Ind. 74(19): 1219-1224.
Welch, H. E. 1960a. Hydromermis churchil-lensis n. sp. (Nematoda: Mermithidae) aparasite of Aedes communis (Deg.) fromChurchill, Manitoba, with observations on itsincidence and bionomics. Can. J. Zool. 38(3 ) : 465-474.
. 1960b. Notes on the identities of mer-mithid parasites of North American mos-quitoes, and a redescription of Agamomermisculicis Stiles, 1903. Proc. Helmin. Soc.Wash. 27(2): 203-206.
. 1962. New species of Gastromermis,Isomermis, and Mesomermis from black flylarvae. Ann. Ent. Soc. Am. 55: 535-542.
1963. Amphimermis bogonae sp. nov.and Hexamermis cavicola sp. nov. from theAustralian bogong moth, Agrotis infusa(Boisd.) with a review of the genus Amphi-mermis Kaburaki and Imamura 1932. (Nema-toda: Mermithidae). Parasitology 53: 55-62.
. 1964. Romanomerrnis iyengari n. sp.,Pilot Register of Zool. Card No. 4.
. 1965. Entomophilic nematodes. Ann.Rev. Ent. 10: 275-302.
Copyright © 2011, The Helminthological Society of Washington

68 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Anthelmintic Activity of Tetramisole, Thiabendazole, andPurified Fine Particle Phenothiazine Against ExperimentalInfections of Haemonchus contortus and TrichostrongylusSpecies in Sheep
M. L. COLGLAZIER, K. C. KATES, AND F. D. ENZIEBeltsville Parasitological Laboratory, Animal Disease and Parasite Research Division,ARS, USDA, Beltsville, Maryland 20705
The efficacy of tetramisole (2,3,5,6-tetra-hydro-6-phenyl-irnidazo [2,1-b] thiazole hy-drochloride), a broad spectrum anthelminticfor man and animals, was first reported byThienpont et al. (1966). Rohrbacher et al.(1967) and Bullock et al. (1968) reportedthat the original optically inactive cW-tetra-misole had been separated into optically actived and / compounds and that anthelmintic ac-tivity was ascribable to /-tetramisole. In pre-liminary trials, /-tetramisole had about the sameanthelmintic activity as the dl compound whengiven at about half the dose rate. Publisheddata on the activity of tetramisole in sheep andgoats pertain to d/-tetramisole; see Corn wellet al. (1967), Fitzsimmons (1966), Forsyth(1966), Gibson (1966), Guilhon (1966),Lyons et al. (1968), Pankhurst and Sutton(1966), Reinecke (1966), Ross (1966), Shoneand Philip (1967), Thomas and Bainbridge(1967), and Walley (1966). Because of themarked promise shown by tetramisole as ananthelmintic for sheep, a trial was conductedto compare the activity of the dl and / com-pounds with that of thiabendazole and pheno-thiazine against experimental infections ofHaemonchus contortus, Trichostrongylus axei,and Trichostrongylus colubriformis in sheep.
Materials and MethodsEXPERIMENTAL ANIMALS: Polled Dorset
lambs (18 wethers and 2 females) used in thistrial were 10-10.5 months old, weighed 21-32kg (avg 27.7 kg), and were raised parasite-free except for insignificant infections ofStrongyloides papillosus and coccidia. Thelambs were allocated to five groups of fourlambs, each group having comparable meanweights (Table 1). Each group was held inseparate, concrete-floored pens which werethoroughly cleaned every 2 days. The lambs
were fed a balanced, pelleted ration in ade-quate quantity to maintain moderate but notmaximum growth.
ORIGIN AND ENUMERATION OF INFECTIVELARVAE: An isolate of H. contortus, designatedAH-2, was established in 1966 from the sheepflock at the Animal Husbandry Division, Agri-cultural Research Service, Beltsville, Maryland.Infective larvae used in this trial were obtainedfrom standard cultures of feces from a stocklamb infected with this isolate.
Infective larvae of Trichostrongylus spp.were of the KH isolate (Kates and Thompson,1967), which contained, on the basis of pre-vious sheep necropsies, about 28 per centT. axei and 72 per cent T. colubriformis.
Prior to larval inoculation of the lambs,infective larvae in the two suspensions (one ofH. contortus and one of Trichostrongylus spp.)were quantitated as follows: After appropriatedilution, the larvae in ten 1-ml samples fromeach suspension were counted in Scott count-ing chambers; undiluted formalin was addedto immobilize the larvae. The per cent stan-dard error (SE) of the larval counts of the H.contortus suspension was 1.87 and of theTrichostrongylus spp. was 1.55.
LARVAL DOSE PREPARATION AND INOCULA-TION OF LAMBS: Each larval dose per lambcontained 5,000 ± 93 SE H. contortus and44,400 ± 688 SE Trichostrongylus spp. (95%confidence interval = SE X 2.26). Each dosewas concentrated by centrifugation and pi-petted onto filter paper. The latter was theninserted into a gelatin capsule and adminis-tered by balling gun.
ANTHELMINTICS EMPLOYED, THEIR ADMIN-ISTRATION, AND DOSAGES: (1) Tetramisole:chemically pure / and dl compounds; Amer-ican Cyanamid Co., Princeton, N. J. (2) Thia-bendazole: Thibenzole; Merck & Co., Railway,
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 69
Table 1. Activity of anthelmintics against H. contortus based upon necropsy worm counts.
DrugZ-Tetramisole
dZ-TetramisoleThiabendazole
Phenothiazinepurified, 2—3 /JL
Controls
No. of lambs1( Avg wt, kg )
4 (28.8)
4 (26.8)4 (27.5)
4 (27.6)
4 (27.7)
Dose ratemg/kg
8
1550
550
—
AdultAvg (range)
0
077
(0-140)<1
(0-3)226
(143-393)
4th-stage larvae Total worms Per cent efficacyAvg (range) Avg (range) 5th-stage 4th-stage-
5(0-20)
01,470
(640-2,380)1,390
(160-3,120)1,520
(920-2,040)
5(0-20)
01,547
(740-2,449)1,391
(163-3,120)1,746
(1,063-2,293)
100
10065
99
99
1003
8
1 All males (wethers), except one female each in dZ-tetramisole and thiabcndazolc groups.2 See text.
N. J. (3) Phenothiazine: 2-3 p. average par-ticle diameter; purified grade (approximatepurity 99.9%); wetting agent 1% lecithin;Atomic Basic Chemical Co., Pittsburgh, Pa.
All the anthelmintics were prepared asaqueous drenches and administered with adose syringe on the 28th day postinfection.The two commercial products were given atcommonly used dose levels, thiabendazole at50 mg/kg and phenothiazine at 550 mg/kg.d£-Tetramisole and /-tetramisole were given at15 and 8 mg/kg, respectively.
EXPERIMENTAL PROCEDURES: Data on weightgains, packed red-cell volume (micro-hemato-crit), and worm eggs per gram of feces wereobtained weekly on all lambs.
All lambs were killed 1 week after treat-ment. The parasites remaining in the treatedand control lambs were recovered and countedby the following procedures: The contents ofeach abomasum and small intestine were re-covered and stored in a large container towhich undiluted formalin was added. Eachorgan was then separately subjected to arti-ficial peptic digestion overnight at 37 C. Thenext day the contents and digests of eachorgan from each lamb were combined, and thefinal preparations made up to 2,000 ml forsampling. From each preparation, five 20-mlsamples were taken for calculating the numbersof Trichostrongylus spp. and larval parasitesin each organ. After the samples were removedfrom the total abomasal material, the remainderwas washed through screens and the adult //.contortus recovered. All adult H. contortuswere counted, but total numbers of the smallnematodes were calculated from the samplecounts.
Efficacy calculations were made by thestandard method for the controlled anthel-mintic test.
SUPPLEMENTAL LAMB INFECTIONS: Becausesubstantial numbers of apparently inhibited4th-stage PI. contortus were recovered 35 dayspostinfection from lambs in the control andtwo treatment groups (Table 1), four morelambs were employed to determine the inhibi-tory influence, if any, of simultaneous infec-tion of Trichostrongylus spp. on the develop-ment of H. contortus. Two lambs wereinoculated with similar numbers of larvae ofH. contortus and Trichostrongylus spp. (5,000and 44,200, respectively) of the same isolatesthat were used in the anthelmintic trial, andeach of two lambs was inoculated with 5,000H. contortus larvae only (Table 4 ) . Subse-quently, these lambs were penned, fed, ne-cropsied, and their parasites recovered andcounted in the same manner as for the lambsin the anthelmintic trial.
ResultsThis trial was planned to obtain compara-
tive data on the activity of certain anthel-mintics against adult populations of three im-portant nematode parasites of sheep. Conse-quently, the lambs were treated 28 days post-infection when practically all worms ordinarilyshould have become adults. Surprisingly, how-ever, the majority of the H. contortus re-covered at necropsy from the control lambsand from two of the four treatment groups(thiabendazole and phenothiazine) were 4th-stage larvae (Table 1). Therefore, for thisand other reasons discussed later, we believethat considerable numbers of inhibited 4th-
Copyright © 2011, The Helminthological Society of Washington

70 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Table 2. Activity of anthelmintics against Trichostrongyhis spp. based upon necropsy worm counts.
Drug
Z-Tetramisole
rfZ-TetramisoleThiabendazolePhenothiazine
purified, 2-3 /J.Controls
No. oflambs1
4
444
4
Dose ratemg/kg
8
1550
550
T. axei-
Avg (range)
65(0-180)
00
30(0-60)3,675
(2,660-4,400)
T. colubriformis
Avg (range)
0
00
655(140-1,260)
10,975(7,060-13,900)
Both speciesAvg (range)
65(0-180)
00
685(200-1,300)
14,470(9,720-18,300)
Per cent efficacy
T. axei T. colubri-formis
98 100
100 100100 10099 94
1 Same lambs inoculated simultaneously with II. contortus (Table 1).2 All Trichostrongijlus spp. recovered were adults.
stage II. contortus were present in lambs ofall 5 groups when the anthelmintics wereadministered. If this assumption is correct,data were obtained on the activity of the drugsagainst 4th-stage //. contortus as well as adults.
The data on the activity of the anthelminticsagainst adult and 4th-stage H. contortus aresummarized in Table 1. Both dl- and Z-tetrami-sole were 100% effective against adult H. con-tortus and almost as effective against 4th-stagelarvae; only one 4th-stage larva was found in20 abomasal samples from the four lambsgiven Z-tetramisole. Phenothiazine was 99%effective against adult H. contortus, but wasineffective against 4th-stage larvae. Thiaben-dazole was only 65% effective against adultH. contortus and almost totally ineffectiveagainst 4th-stage larvae.
Data on activity of the anthelmintics againstT. axei and T. colubriformis are summarizedin Table 2. All were very effective (94-100%) against adults of both species.
All anthelmintics reduced the parasite eggcounts 92-100% (Table 3). Thiabendazolecaused the least reduction because of the re-
tention of adult H. contortus after treatment.No eggs were seen in the posttreatment fecalsamples from lambs treated with tetramisole.The drugs were well tolerated in all trials.
Average pretreatment egg counts in thefive groups of lambs were quite uniform, vary-ing only from 2,445 EPG in the group givenphenothiazine to 3,563 EPG in the controlgroup. This uniformity in the pretreatmentegg counts indicated that reasonably uniformadult infections were established initially inthe lambs. However, an average of 226 adultH. contortus (55% females) were present inthe control lambs at necropsy, whereas thesame lambs had an average of 14,470 adultTrichostrongijlus spp. (54% females) (Tables1, 2). Therefore, substantial numbers of theeggs counted pretreatment were probablythose of Trichostrongylus spp. No attempt wasmade to differentiate the eggs of H. contortusand Trichostrongijlus spp. because quantita-tive differentiation of T. axei and H. contortuseggs in mixed infections is unreliable.
The reduction in average hematocrit valuesover the 4 weeks from infection to treatment
Table 3. Activity of anthelmintics against H. contortus and Trichostrongyhis spp. based upon combinedspecies fecal egg counts.
Avg EPG
Drug
Z-TetramisoledZ-TctramisoleThiabendazolePhenothiazine purified, 2—3 fiControls
No. oflambs1
44444
Dose ratemg/kff
81550
550-
Pretreatment4th week
3,5502,5082,8252,4453,563
Post-treatmentat necropsy
00
20530
2,645
EPGreductionper cent
100100929825
1 Saine lambs for which necropsy worm counts are summarized in Tables 1 and 2.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 71
Table 4. Supplemental lamb infections. Necropsy worm counts 35 days postinfection of nonmedicatedlambs infected (a) simultaneously with H. contortus and Trichostrongylus spp. and (b) with H. contortusonly.
No. oflambs1
(a) 2
(b) 2
Agemo.
,3.5
11
H
Larvae inoculatedper lamb
5,000 H. contortus44,200 Tricliostrongi/his
5,000 H. contortus
. contortus Avg ( range )
Adults 4th-stagelarvae
1,664 20(806-2,522) (0-40)
1,240 0(998-1,482)
T. axei-Avg (range)
4,530(4,300-4,760)
T. colubriformis-Avg (range)
19,850(16,500-23,200)
T. a. + T. c.Avg (range)
24,380(21,260-27,500)
1 All males (wethers) except for one female in ( a ) ; weight range 17—25 kg." All in the adult stage.
varied only from 5-6% per group, which sup-ports our conclusion that average infectionsper group at the time of treatment werecomparable.
Our conclusion that the 4th-stage H. con-tortus larvae recovered at necropsy werederived from the original inocula is supportedby the fact that similar percentages ofH. contortus (34.9%—larvae + adults) andTrichostrongylus spp. (32.6%—adults) of theoriginal inocula were recovered at necropsyfrom the control lambs.
There appears to be no obvious explanationfor the presence of large numbers of 4th-stageH. contortus in the control and some treatmentgroups at the termination of this trial. Theresults of the small scale test (Table 4) con-ducted to determine if simultaneous Tricho-strongylus spp. infections had a reproducibleinhibitory effect on the development of H.contortus were essentially negative. Substan-tial numbers of adult H. contortus (avg 1,664),and very small numbers of 4th-stage larvae(avg 20), were recovered at necropsy fromthe two lambs inoculated with all three speciesof nematodes. Also, substantial numbers ofadult worms (avg 1,240) and no 4th-stagelarvae were recovered from the two lambsinoculated with comparable numbers of H.contortus only. It is also noteworthy that thetwo lambs with mixed infections harboredlarger numbers of adult Trichostrongylus spp.(avg 24,380 = 55% of inocula) than the con-trol lambs in the anthelmintic trial (avg 14,470= 32% of inocula). The results of this supple-mentary test indicate, therefore, that the con-current Trichostrongylus spp. infection was nota major cause of the inhibited development ofH. contortus in the anthelmintic trial.
DiscussionOur results confirm those of previously cited
authors that rf/-tetramisole given orally at 15nig/kg is very effective in removing adultH. contortus, T. axei, and T. colubriformisfrom lambs, and that /-tetramisole at abouthalf the dose rate (8 mg/kg) of the dl com-pound is equally effective. If it is assumedthat at the time of treatment the two groups oflambs treated with tetramisole compounds aswell as the control group had comparablenumbers of inhibited 4th-stage H. contortus,which seems likely, then both tetramisoles werehighly effective against these larvae. Gibson(1966) reported 100% efficacy of 15 mg/kgof rfZ-tetramisole against all larval stages ofH. contortus in lambs, and Lyons et al. (1968)reported that the dl compound at the samedose rate was 97% effective against 4th and87% against early 5th-stages of H. contortus.Also, Shone and Phillip (1967) reported thatat 15 mg/kg d/-tetramisole was more than99% effective against 4th-stage H. contortus,but was only 72.5% effective against parasitic3rd-stage. Furthermore, Gibson (1966) re-ported that, at 15 mg/kg, dZ-tetramisole washighly effective against immature T. colubri-formis in sheep. Data are rapidly accumulat-ing to show, therefore, that the two tetramisolesat appropriate dose levels are highly effectiveagainst both adult and larval stages of H.contortus and Trichostrongylus spp. in sheep.
We obtained relatively poor action withthiabendazole against adult H. contortus (AH-2). These findings were not especially sur-prising, however, because certain unpublishedobservations involving the source flock alsosuggested that the sheep may harbor H. con-tortus that is somewhat resistant to thiabenda-
Copyright © 2011, The Helminthological Society of Washington

72 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
zole. The fact that we obtained with the AH-2isolate an efficacy of only 65% against adultH. contortus, and apparently no activity againstinhibited 4th-stage larvae, tends to support theearlier observations, and indicates that theAH-2 isolate may be resistant to thiabendazoleat the 50 mg/kg level. Further studies are inprogress to compare more precisely the efficacyof thiabendazole against this and other isolatesof H. contortus. Most reports (see Drudgeet al., 1964; Gibson, 1965) indicate that thia-bendazole at 50 mg/kg is at least 90% effec-tive against adult H. contortus in sheep. In-deed, Gibson (1965) recommends this regimenas the treatment of choice. Conway (1964),on the other hand, reported poor results atthis dose level, but improved efficacy at 80mg/kg. At the dose rate of 44 mg/kg, Drudgeet al. (1964) administered six successivedrenches at 4-week intervals to naturally in-fected sheep on pasture with notable lack ofsuccess in controlling H. contortus. Althoughfewer H, contortus were recovered at necropsyfrom the principals than from the controls, theresidual worm burdens in the treated animalswere almost pure infections of H. contortus.They concluded that this treatment programapparently selected, from the original popula-tion of H. contortus, a strain resistant to thedrug at this dose level. Attention should becalled to the reports of Gibson (1964), Gordon(1964), Reinecke et al. (1962), and Southcott(1963) that thiabendazole at 50 mg/kg bodyweight is effective against immature H. con-tortus. However, only Reinecke et al., exam-ined digests for the immature stages.
Purified, 2—3 ^ phenothiazine given at 550mg/kg to a group of four lambs removed 99%of adult H. contortus (AH-2), but apparentlyhad no effect on inhibited 4th-stage larvae.These results may be compared with thosereported by Colglazier et al. (1967) for threeother phenothiazine products given at the samedose rate to lambs experimentally infected withanother isolate (AH) of H. contortus from thesame sheep flock. They reported efficaciesagainst adult parasites of 12% for N.F. 10 ^phenothiazine, 40% for N.F. 7 ^ phenothia-zine, and 75% for purified 6.9 p. phenothiazine.There is a lack of data on the effect of pheno-thiazine against immature H. contortus. How-ever, Gordon (1940) reported that pheno-thiazine in doses of 600 mg/kg of body weight
destroyed immature H. contortus 10 and 15days old. Later, Southcott (1963) reportedthat phenothiazine in similar doses was lesseffective against H. contortus 12 hr to 7 daysold than against parasites 10—28 days old.
A unique finding in this trial was the re-covery of substantial numbers of 4th-stageH. contortus from lambs of the control, pheno-thiazine, and thiabendazole groups. These4th-stage larvae were 1.5—2.5 mm long, andwere equivalent in size and development to4th-stage larvae 4—6 days old, as described byVeglia (1915). For reasons given below, webelieve these larvae developed from the orig-inal inocula, were inhibited in their develop-ment by some factor(s) not now apparent, andwere present in all groups of lambs at the timeof treatment 28 days postinfection. Therefore,efficacy calculations are given in Table 1 alsofor activity of the anthelmintics against 4th-stage H. contortus. On this basis, only the twotetramisoles showed action against these in-hibited larvae.
We cannot be certain the 4th-stage H. con-tortus recovered from the trial lambs camefrom the original inocula and were inhibited intheir development, but the following reasonssupport this thesis: (1) The animal pens werecleaned regularly. (2) The experiment wasconducted in midwinter and the animal quar-ters were too cold for normal development ofinfective larvae in the pens. (3) No parasiticlarvae of Trichostrongylus spp. were recoveredfrom any of the 20 lambs on trial, whichstrongly supports our contention that reinfec-tion did not occur from pen contamination.(4) The numbers of adult H. contortus werelow and the numbers of adult Trichostrongylusspp. were high in the lambs; therefore, ifreinfection had occurred in the pens, substan-tial numbers of immature Trichostrongylusspp. would have been recovered along withimmature H. contortus, which was not thecase. (5) The total numbers of H. contortusand Trichostrongyhis spp. recovered at ne-cropsy from the controls in both instances wereabout one-third of the original inocula.
Summary
In a controlled test with experimental infec-tions of Haemonchus contortus, Trichostron-gylus axei, and Trichostrongylus coluhriformisin lambs, /-tetramisole (8 mg/kg), df-tetrami-
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 73
sole (15 mg/kg), thiabendazole (50 mg/kg),and purified, 2-3 /j, phenothiazine (550 mg/kg) were all highly effective (94-100%)against adult parasites of all species, exceptfor thiabendazole which was only 65% effec-tive against adult //. contortus. The poorresults obtained with thiabendazole suggestthat the H. contortus isolate (AH-2) used inthis work may be resistant to the drug at the50 mg/kg level. Substantial numbers of ap-parently inhibited 4th-stage H. contortus wererecovered from all lambs except those treatedwith tetramisole.
Acknowledgmen t
The authors gratefully acknowledge thetechnical assistance of R. H. Burtner.
Literature Cited
Bullock, M. W., J. J. Hand, and E. Waletzky.1968. Resolution and racemization of dl-tetramisole, dl-6-phenyl-2,3,5,6-tetrahydroim-idazo-[2,l-b] thiazole. J. Med. Chem. 11:169-171.
Colglazier, M. L., F. D. Enzie, and R. P. Leh-mann. 1967. Strain variation in the re-sponse of Haemonchus contortus in sheep andgoats given phenothiazine: Efficacy of N.F.and purified products on pure parasitic infec-tions. Am. J. Vet. Res. 28: 1711-1722.
Conway, D. P. 1964. Variance in the effec-tiveness of thiabendazole against Haemon-chus contortus in sheep. Am. J. Vet. Res.25: 844-845.
Cornwall, R. L., R. M. Jones, J. Berry, T.Jordan, E. A. Mercer, D. A. Pullen, andC. J. Riley. 1967. Field trials in sheepwith the anthelmintic pyrantel tartrate. IV.Comparison of pyrantel tartrate and tetram-isole in the prevention of parasitic gastro-enteritis. Vet. Rec. 80: 676-679.
Drudge, J. H., J. Szanto, Z. N. Wyant, and G.Elam. 1964. Field studies on parasitecontrol in sheep. Comparison of thiabenda-zole, ruelene, and phenothiazine. Am. J. Vet.Res. 25: 1512-1518.
Fitzsimmons, W. M. 1966. The effect of tet-ramisole on the parasitic stages of Tricho-strongylus colubriformis in experimentally in-fected goats. Vet. Rec. 79: 599-600.
Forsyth, B. A. 1966. Tetramisole: A newanthelmintic for sheep. Austral. Vet. 42:412-419.
Gibson, T. E. 1964. The evaluation of anthel-mintics for the removal of gastrointestinalnematodes of sheep—an improved form of
the controlled test. Parasitology 54: 545-550.
. 1965. Veterinary anthelmintic medi-cation. Tech. Com. No. 33, Commonwl.Agric. Rur., Rucks, England, 206 pp.
1966. An evaluation of the anthelmin-tic tetramisole using the improved controltest. Vet. Rec. 79: 601-602.
Gordon, H. McL. 1940. The anthelmintic effi-ciency of phenothiazine against immatureHaemonchus contortus. J. Council Sci. Ind.Research 13: 245-246.
. 1964. Studies of anthelmintics forsheep: Thiabendazole. Austral. Vet. J. 40:9-18.
Guilhon, J. 1966. Action d'un derive de 1'Imi-dazo-thiazole (Tetramisole) sur les Strongy-lides gastro-intestinaux des ovines. Rull.Acad. Vet. 39: 255-264.
Kates, K. C., and D. E. Thompson. 1967.Activity of three anthelmintics against mixedinfections of two Trichostrongylus spp. ingerbils, sheep, and goats. Proc. Helmin. Soc.Wash. 34: 228-236.
Lyons, E. T., J. H. Drudge, and S. C. Tolliver.1968. Activity of tetramisole against gastro-intestinal helminths in a controlled test inlambs. Am. J. Vet. Res. 29: 2157-2162.
Pankhurst, J. W., and D. O. Sutton. 1966.Field trials with a new anthelmintic in lambs.Vet. Rec. 79: 166-170.
Reinecke, R. K. 1966. A larval anthelmintictest. J. So. Afr. Vet. Med. Assn. 37: 29-33.
, A. J. Sniders, and I. G. Horak. 1962.A modification of standard procedures forevaluating the relative efficacy of anthel-mintics. Onclerstepoort J. Vet. Res. 29:241-257.
Rohrbacher, G. H., J. Emro, and E. Walet-sky. 1967. A comparison of the anthel-mintic activity of I and c/Z-tetramisole againstnematodes of cattle and sheep by drench.Prog, and Abst. 42nd Ann. Meet., Am. Soc.Parasitol., p. 44 (also personal communica-tion ).
Ross, D. B. 1966. Critical trials with tetrami-sole given to lambs experimentally infectedwith Haemonchus contortus, Ostertagia cir-cumcincta, Nematodirus battus, and Tricho-strongylus colubriformis. Vet. Rec. 79: 392-395.
Shone, D. K., and J. R. Phillip. 1967. An-thelmintic and toxicity studies tetramisole: I.Anthelmintic efficacy. J. So. Afr. Vet. Med.Assn. 38: 165-176.
Southcott, W. H. 1963. Ovicidal effect ofthiabendazole and its activity against im-mature helminths of sheep. Austral. Vet. J.39: 452-458.
Copyright © 2011, The Helminthological Society of Washington

74 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Thomas, R. J., and M. H. Bainbridge. 1967.The influence of the anthelmintic tetramisoleon the productivity of fattening lambs. Vet.Rec. 80: 266-269.
Thienpont, D., O. F. J. Vanparijs, A. H. M.Raeymaekers, J. Vandenberk, P. J. A.Demoen, F. T. N. Allewijn, R. P. H.Marsboom, C. J. E. Niemegeers, K. H. L.Schellekens, and P. A. J. Janssen. 1966.Tetramisole (R 8299), a new, potent broad
spectrum anthelmintic. Nature. 209: 1084-1086.
Veglia, F. 1915. The anatomy and life-historyof the Haemonchus contortus (Rud.) . Thirdand fourth Rpts. Dir. Vet. Res., Dept. Agric.,Un. So. Afr., pp. 347-500.
Walley, J. K. 1966. Tetramisole (dl 2,3,5,6-tetrahyclro-6-phenyl-imidazo [2,1-b] thiazolehydrochloride—Nilverm) in the treatment ofgastrointestinal worms and lungworms indomestic animals. Vet. Rec. 78: 406-416.
A New Trematode Cephalogonimus sireni sp. nov. (Digenea:Cephalogonimidae) from Florida Mud Eel, Siren lacertina
G. PREMVATI1
Department of Biology, Florida Agricultural and Mechanical University, Tallahassee, Florida 32307
In amphibians, a single genus Cephalogon-imus Poirier, 1886, of the family Cephalo-gonimidae, has been recorded mostly fromfrogs of the genus Rana. From urocleles, onlyone species, C. amphiuma Chandler, 1923, hasbeen reported from Amphiuma means inLouisiana and subsequently Manter (1938)reported the same species from Siren ofFlorida. Two mud eels, Siren lacertina, wereexamined for helminth infection, and one wasfound to be infected with a new trematode ofthe genus Cephalogonimus. The worms werefixed under slight coverglass pressure in hotAFA and stained with Semichon's carmine.
All measurements are in microns, unlessotherwise indicated.
Cephalogonimus sireni sp. nov.(Fig. 1)
HOST: Siren lacertina Linnaeus, 1766.LOCALITY: Lake Munson, Leon County,
Florida.LOCATION: Latter half of intestine.NUMBER OF WORMS: Five from one host.SPECIMENS: Holotype and one paratype in
USNM Helm. Coll. Nos. 71236, 71237.
1 Present address: Department of Zoology, University ofLucknow, Lucknow (U.P.) , India.
Description
Body oval, spinous, small, 1.04—1.44 mmlong, and 760-870 wide. Oral sucker sub-terminal, 170 in diameter; ventral sucker verylarge, 340 in diameter; both suckers have threeto six rows of spines on their margins. Pre-pharynx thick and muscular; pharynx 125-130by 110, very muscular, globular, resemblinga sucker; esophagus absent; intestinal cecaterminate almost at posterior end of body.Distance between ceca and posterior end ofbody 140-150.
Genital pore median and terminal. Testes230-270 by 200-230, ovoid to spherical, sym-metrically opposite, immediately posterior toventral sucker. Vasa efferentia join in acetab-ular region to form the vas deferens. Cirrussac preacetabular, elongated; enclosing elon-gated seminal vesicle, ejaculatory duct, cirrusand prostatic cells; runs toward left of phar-ynx and then joins uterus to form hermaphro-ditic duct that runs dorsal to oral sucker.Ovary submedian, ovoid to lobed, dorsal andto right of ventral sucker, measures 160-170by 100-110. Laurer's canal and seminal re-ceptacle present. Vitellaria follicular, extends
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 75
Figure 1. Cephalogonimus sireni sp. nov. fromFlorida mud eel, Siren lacertina. Holotype: ventral
from intestinal bifurcation to testicular level.Two transverse vitelline ducts dorsal to ven-tral sucker and join to form an elongated vitel-line reservoir. Whole female genital complexlies in acetabular region. Uterus extends pos-teriorly and then runs anteriorly on left ofventral sucker. Excretory bladder Y-shaped,stem of Y being small. Eggs small, numerous,and measure 23—25 by 14-15 (30 measured).
DiscussionThe subfamily Cephalogoniminae Looss
(1899), subsequently raised to family Cepha-logonimidae Nicoll (1914), includes fourgenera: Cephalogonimus Poirier, 1886, Emo-leptalea Looss, 1900 (Syn.: Leptalea Looss,1899), Paracephalogonimits Skrjabin, 1950and Oudhia Dayal and Gupta, 1954.
Oral sucker is larger than ventral sucker inthe three genera, Emoleptalea, Paracepha-logonimus, and Oudhia, and in all speciesexcept five, of the genus Cephalogonimus.Two of these five species, namely C. com-pactus Stunkarcl, 1924 and C. japonicus Ogata,1934, have oral sucker equal to ventral sucker.In the remaining three species, the oral suckeris only slightly smaller than the ventral sucker,the sizes being: 240 and 290 in C. lenoiriPoirier, 1886; 220 and 270 in C. vesicaudusNickerson, 1912; 220-270 and 310-330 inC. manchuricus Oguro, 1941. But in C. sireni,the oral sucker is half the size of ventral suckerand they measure 170 and 340. This ratio of1:2 in the sizes of the two suckers is not seenin any species of the genus Cephalogonimus.
The position of testes in all species of thegenus Cephalogonimus is tandem or diagonal.But in C. sireni, the testes are symmetricallyopposite.
In all species of the genus Cephalogonimus,the shape of the ovary is entire. But ovary islobed in C. sireni.
The position of testes and shape of ovaryare two of the major characteristics used todistinguish the two families Prosthogonimidaeand Cephalogonimidae: the former has sym-metrically opposite testes and lobed ovary,while the latter has tandem or diagonal testesand entire ovary. Cephalogonimus sireni showsprosthogonimid characters in having sym-metrically opposite testes and lobed ovary;but it has typical cephalogonimid shape of thebody, elongated cirrus sac, genital pore atanterior extremity on the dorsal side, and anamphibian host.
Thus C. sireni differs from all the presentspecies of the genus Cephalogonimus in thefollowing: (1) ventral sucker very large,twice the size of oral sucker; (2) symmetricallyopposite testes, and (3) lobed ovary. It is,accordingly, regarded as a new species.
SummaryCephalogonimus sireni sp. nov. (Digenea:
Cephalogonimidae) is described from Floridamud eel, Siren lacertina. It is characterizedby: (1) very large ventral sucker, twice the
Copyright © 2011, The Helminthological Society of Washington

76 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
size of oral sucker; (2) symmetrically oppositetestes, immediately posterior to ventral sucker,and (3) lobed ovary. In the position of testesand shape of ovary, C. sireni shows charactersof the family Prosthogonimiclae; but it hastypical cephalogonimid shape of the body,elongated cirrus sac, genital pore at the anteriorextremity on the dorsal side, and an amphibianhost.
Ack no wledgmen tThanks are extended to Professor Robert B.
Short of the Florida State University, Tallahas-see, for the use of his personal library.
Literature CitedManter, H. W. 1938. A Collection of Trema-
todes from Florida Amphibia. Trans. Amer.Micr. Soc. 57: 26-37.
Intestinal Helminths of the Bullfinch, Pyrrhula pyrrhula (L.), inSouthern England
I. C. WILLIAMS1 AND I. NEWTON2
The bullfinch, Pyrrhula pyrrhula (L.) , is asmall passerine bird which is widely distributedin Europe and northern Asia. Over most of itsrange it favors coniferous or mixed forests, andin Britain thick deciduous woods and hedge-rows. It lives chiefly on seeds and buds, butfeeds its young on a mixture of seeds andsmall invertebrates. By eating the buds offruit trees it has become a serious pest in fruitgrowing areas of southern England, where itis resident throughout the year. The ecologyand general biology of the bullfinch have re-ceived considerable attention (Wright andSummers 1960; Newton 1964, 1966, 1967a, b);on the other hand, knowledge of the helminthparasites of this bird is confined to a few scat-tered records and the brief report on Urotocusrossittensis (Miihling, 1898) by Newton andWilliams (1967). The large numbers of bull-finches killed annually in pest-control programsin fruit growing areas of southern Englandprovided an opportunity to investigate theirintestinal helminths.
Through the kindness of fruit growers, deadbullfinches were received regularly betweenJanuary 1965 and January 1966 from 37 local-ities in 10 English counties. A total of 1,265birds was examined. The gut was removedfrom each bird soon after arrival and that part
1 Department of Zoology, The University, Hull, England.- Edward Grey Institute for Field Ornithology, Botanic
Garden, Oxford, England.
from the gizzard forward was used for ananalysis of the food (Newton 1967a), whilethe rest was stored in 5% formaldehyde andexamined later for helminths. The resultsreported herein thus relate to the duodenum,small and large intestines, rectum, cloaca, andbursa Fabricii. Birds are classed as juvenilefrom the time they left the nest to the end ofthe postjuvenile moult (3-4 months), first-year from then until the first complete moulta year later (age 15-16 months) and adultthereafter. No nestling bullfinches wereexamined.
Results(a) Incidence of infection
Intestinal helminths were found in 71(5.6%) of the bullfinches examined; theycomprised three species of trematodes andtwo species of cyclophyllidean cestodes (Table1). No nematodes or Acanthocephala werefound. The trematodes were members of thefamily Brachylaimidae, namely Brachylaimaarcuata Dujardin, 1845, Leucochloridiummacrostomum (Rudolphi, 1803) and Urotocusrossittensis (Mtihling, 1898), and the cestodeswere Anomotaenia borealis (Krabbe, 1869)and Hymenolepis passeris (Gmelin, 1790).
Not more than one species of helminth wasfound in any infected host and, combining allthe data, there was no significant differencein incidence of infection between male (42/
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 77
Table 1. The occurrence of five helminth species in Bullfinches, Pyrrhula pyrrhula (L.), from differ-ent English counties.
No. oflocal-ities in
County county
BedfordshireBerkshireBuckinghamshireEssexHampshireHerefordshireKentSuffolkSussexWarwickshireTotal
135511
12162
37
Brachylaima LeucocJiloridiiimNo. of arcuata1' " macrostomumbull
finchesexamined
23131119207
6275
32992
21413
1,265
No.inf.
0231104200
13
%inf.
01.52.50.51.601.22001
No.inf.
01000000102
%inf.
00.80000000.500.2
Urotocusrossittensis
No.inf.
123
32
1051420
51
%inf.
417.62.51
1670.34104
Anomotaeniaborealis-
No.inf.
00010000001
%inf.
0000.50000000.1
Hymenolepispasseris
No.inf.
00100210004
%inf.
000.8002.70.30000.3
Total
No. %inf. inf.
1 426 20
7 64 2
11 187 96 26 73 10 0
71 5.6
1 New record in the British Isles.- New host record in the bullfinch.
698, 6.0%) and female (29/567, 5.1%) birds.The most common parasite found was U. rossit-tensis in 51 birds from 13 of the 37 localitiesinvolved and within these areas the propor-tion of infected birds varied greatly. Berk-shire and Hampshire provided the greatestproportion of infected birds (33/193, 17%)and the southeastern counties of Kent, Essex,and Sussex the smallest, with under 1% (5/750) infected. The second most frequenthelminth found was Brachylaima arcuata in13 birds, six of which were from the central-southern counties (Berkshire, Buckingham-shire, and Hampshire, 6/312, 2%) and fivefrom the southeastern counties (Essex, Kent,5/536, 1%); the remaining two infected birdswere from one locality in Suffolk from which92 birds were examined. The distribution ofbullfinches infected with L. macrostomum andthe two species of cestodes was sporadic butit was noticeable that no tapeworms werefound in any birds from Berkshire andHampshire, although these areas containedbullfinches with the highest incidence oftrematodes.
Among birds from Berkshire and Hamp-shire, there was an apparent decline in inci-dence of infection with U. rossittensis withincreasing age (Table 2). Thus, 25% ofjuveniles from these two counties containedthis species compared with 18% of first-yearbirds and 14% adults, but these differenceswere not significant statistically. If the inci-dence of L. macrostomum and Brachylaima
arcuata in birds from Berkshire and Hamp-shire are added to the figures for U. rossittensisthe percentage incidence of infection with agebecomes 25, 19, and 16, respectively. Onlyone group of juveniles (hatched 1965) wasexamined, but two groups of first-year birds(hatched 1964 and 1965) and two groups ofadults (hatched pre-1964 and pre-1965) werereceived. The incidence of trematodes amongthese five groups from Berkshire and Hamp-shire was: adult, hatched pre-1964, 3/20(15%); first-year, hatched 1964, 7/53 (13%);adult, hatched pre-1965, 3/17 (18%); juve-nile, hatched 1965, 6/24 (25%); first-year,hatched 1965, 18/79 (23%). Comparison ofthe two groups of first-year birds suggeststhat, within these localities, there may be dif-ferences in incidence of infection from yearto year.
Among the two species of cestode it wasnoticeable that no infection of adult bull-finches was found.
(b) Intensity of infectionThe number of individual parasites per in-
fected host varied considerably (Table 2), andthere was a tendency for the intensity to begreater in first-year and adult birds than injuveniles, with the exception of U. rossittensisin which the reverse was seen. With U. rossit-tensis there was a marked difference in themean intensity of infected male (9.4) andfemale (2.9) birds; but this was chiefly dueto two of four juvenile males and three of 25
Copyright © 2011, The Helminthological Society of Washington

78 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Table 2. The occurrence of five helminth species in Bullfinches, Pyrrhula pyrrhula (L.), of differentage.
Age class of host
Number examined
Urotocus rossittensisa. All localities1 Number infected
Per cent infectedMean intensity2
Range of intensityb. Berks, and Hamps. Number infected
Per cent infected
Lciicochloridium macrostomum Number infectedPer cent infectedMean intensityRange of intensity
Brachyluima arcuata Number infectedPer cent infectedMean intensityRange of intensity
Anomotaenia borealis Number infectedPer cent infectedIntensity
Hymcnolepis passcris Number infectedPer cent infectedMean intensityRange of intensity
All species Number infectedPer cent infected
Juvenile
301
93
101-426/24
25
00--10.311
10.32
10.311
124
First winter
770
3757
1-7322/132
18
00--
101.31
1-2
00-
30.46.3
1-16
507
Adult Total and means
194
535
3-7
5/3714
21
151-29
211.5
1-3
00-00--95
1,265
5147
1-7333/193
17
20.2
151-29
1311.3
1-3
10.12
40.35
1-10
715.6
1 If data for all areas are combined, as in a, a possible decline in incidence with increase in age of host is maskedby the fact that relatively more juveniles than older birds were received from areas of low infection. The data for Berk-shire and Hampshire are, therefore, given separately.
- Intensity, being the number of parasites per infected host.
first-year males containing exceptionally largenumbers of parasites (42, 28, 39, 73, and 31respectively). If these birds are excluded, themean intensity of infected males becomes 3.2compared with the females 2.9. Correspond-ing adjusted figures for juveniles, first-yearbirds and adults then become 2.9, 2.9, and5.2 respectively, with an overall mean intensityof 3.1 instead of 7 (Table 2). The averagenumber of U. rossittensis in infected hosts fromlocalities with few birds infected was 1.8, com-pared with 9.6 for Berkshire and Hampshire.
(c) Pathology of infectionOf the five helminth species found only
U. rossittensis was seen to have an appreciableaffect on its host. This species is fusiform inshape and may attain 7 mm in length by 0.4mm in diameter; it lives in the ovoicl, saclike,bursa Fabricii where it presses upon the finger-like projections which line this organ and which
are especially rich in lymphoid follicles. Theeffect is to distort and, particularly in a heavyinfection, destroy the functional division oflymphoid follicles into medulla, which pro-duces lymphocytes, and cortex, where thesemature (Figs. 1, 2). At the same time theextensive vascular and connective tissue net-work, which surrounds and supports the fol-licles, is compressed and ultimately destroyed.Within the bursa U, rossittensis feeds on blood,lymphoid cells and connective tissue. Thebreakdown product of hemoglobin, hematin, isespecially noticeable in histological sectionsand can be recognized microspectroscopically.For the most part the worms lie with theirmouth toward the periphery of the bursa asthough browsing on its inner surface. Noinstance of cannibalism by Urotocus was seen.In addition to damage through pressure andfeeding, the cuticular spines of the wormabrade the surface of the lymphoid tissue.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 79
/ Sl>^ , r-^~^;^^ l - : •&•' *< -sffift
¥*$!*> $$^^'r^ . , ; • ' - • • • ' "Figure 1. Sagittal section of the uiiinfected bursa Fabricii of a juvenile male bullfinch showing
numerous lymphoid follicles.
As well as causing damage within the bursaFabricii, heavy infections with U. rossittensisgreatly distend the bursa producing a "cyst"which may attain 1 cm in diameter. As thebursa lies on the dorsal side of the cloaca,it is possible that a heavily infected and dis-tended bursa might, through its presence,interfere with the normal functioning of theorgans of the pelvic region.
DiscussionBullfinches acquire the various helminth
parasites reported above by ingesting the re-spective larval stages, present in certain inver-tebrates, with their food. As animal materialis normally eaten by bullfinches only duringthe first 10 days or so of life, presumably
infection occurs during this early period. Al-though all the nestling's food, consisting mainlyof caterpillars, flies, spiders, slugs, and snails,is collected by their parents the adults, appar-ently, do not eat invertebrates at this time, nordo they predigest it for their young (Newton1967a); they are, therefore, unlikely to becomeinfected. Yet many helminths were found inbirds of postnestling age. This situation thusprovides an unusual opportunity to examinethe longevity of these various parasites innatural infections, and results show thatBrachylaima arcuata can live in bullfinchesfor at least 18 months, L. macrostomum 25months, U. rossittensis 30 months, Anomotae-nia borealift 2 months and Hymenolepis passeris4 months. As many bullfinches older than
Copyright © 2011, The Helminthological Society of Washington

80 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
_ 1 mm*» ;s-k. • V
>^rxt^i>£&r-'V* >,,^*^>i'rr..J£ ^P fiaVI .f;S& :.:A ,(ifc. nrlRcv &*$& i£ -- »i: j H
Figure 2. Sagittal section of the bursa Fabricii of a juvenile male bullfinch heavily infected withUrotocus rossittensis; the peripheral region of the bursa only remains.
those containing the two cestode species wereexamined, it is possible that these particular-parasites can live only a few months in thishost. This agrees with Kintner's (1938) ob-servations on the parasites of 197 house spar-rows, Passer domesticus (L.) , in Indiana, inwhich cestodes were found only in birds ob-tained before November, with none from No-vember to March. Kintner suggested that thecestodes concerned, which included H. pas-seris, either were short-lived, or that the inter-mediate hosts were not available or did nottransmit them in winter. Our own observa-tions on H. passeris in the bullfinch suggestthat Kintner's first alternative, that the ces-todes were short-lived, is very likely, althoughthere is the possibility, in the present study at
least, that infected intermediate hosts werepresent but not eaten.
An interesting comparative study is that ofSokolova (1959) in Russia on the parasites ofthe chaffinch, Fringilla coelebs L., which isclosely related to the bullfinch. Unlike thelatter, however, invertebrates predominate inthe diet of the adult chaffinch during the sum-mer and the young are reared entirely onanimal material (Newton 1967b). From 143birds Sokolova obtained four species of trema-todes; two of these, Prosthogonimus ovatusand P. cunsatus, inhabited the bursa Fabricii,and the other two, Plagiorchis maculosus andLeucochloridium sp., the intestine. The ces-todes found included "Taenia exigua Dujardin1845" and some unidentified specimens. None
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 81
of these parasites was found in bullfinches.Sokolova found that trematodes infectedmainly nestlings but, in contrast to our re-sults, were rare or absent in fully grown birds.
The low mean intensity of infection of bull-finches with Brachylaima arcuata (1.3) is inagreement with that reported for the relatedspecies B. fuscata in the starling, Sturnus vul-garis L., in northern England (3.4) by Owenand Pemberton (1962), and south Wales (4.12)by James and Llewellyn (1967). The latterauthors also reported that the mean numbersof B. fuscata from infected mistle-thrushes,Turdus viscivorus L., at 5, from a song-thrush,T. ericetorum. Turton, at 9 and from 57 in-fected redwings, T. musicus L., at 9.8. It is,perhaps, surprising that the intensities shouldbe fairly low whereas the corresponding inci-dences of infection varied considerably (1%of 1,265 bullfinches; 24 of 358 (6.7%) star-lings (Owen and Pemberton 1962); 25/122(21%) starlings (James and Llewellyn 1967);2/5 (40%) mistle-thrushes, 1/11 (9%) song-thrushes and 57/171 (33%) redwings (Jamesand Llewellyn 1962)).
It is surprising that neither Owen and Pem-berton (1962) nor James and Llewellyn (1967)found L. macrostomum. in the 470 starlings and235 passerines of six other species they exam-ined, as these birds have previously been re-corded as hosts for this parasite. In Poland,however, Joszt (1962) found L. macrostomumin 3 of 190 (1.6%) house sparrows.
Urotocus rossittensis presents an unusualcase owing to the temporary nature of itshabitat which atrophies with the approach ofsexual maturity and is not normally found inthe adult. Yet we found 2.6% of all adultbullfinches infected with this species, whilethe proportion of infected adults from Berk-shire and Hampshire was much higher at 14%.Undoubtedly infection with U. rossittensis musttake place at an age when animal food is eaten,that is as a nestling. Thus the proportion ofanimal material in the diet and, also, the inci-dence of infective larvae in intermediate hostswill influence the incidence and intensity ofinfection in the bird. These two factors mayoperate to produce the differences describedabove, in infection of bullfinches from differ-ent localities. The observations of Newton(1967a), that bullfinch nestlings reared in
woodland areas receive appreciably greaterproportions of animal food in their diet thanbroods raised in farmland areas, may offer apossible explanation of the present observa-tions but we have no direct evidence to thiseffect. Furthermore, infection must occurwhen the canal between the cloaca and thebursa is large enough to admit the youngparasite, which when infective measures 0.2-0.6 mm in length (Timon-David 1957). Aswe were unable to measure the width of thelumen of the canal we do not know at what ageit becomes too narrow to permit entry of theinfective larva. Only then is the bird freefrom the risk of further infection with thisspecies of parasite. Of course, some timeearlier the canal will have become too narrowto permit the adult 17. rossittensis from leavingthe bursa. Thus in adult birds the worms areeffectively encapsulated within the fibro-muscular wall of the atrophied bursa.
As stated above, exceptionally heavy infec-tions were found only in comparatively youngbirds and there was also an apparent declinein incidence with increasing age of the bird.Both these effects might be due either to theparasite dying, and presumably being eatenby their neighbors although no instances ofcannibalism were seen in our sections, or toinfected birds dying in greater proportion thanuninfected. The role of the bursa appears tobe particularly important during the first fewdays of life and in the fowl it is known to pro-duce both lymphocytes (Ackerman and Knouff1964) and circulating antibodies (Click 1964)during this period. Thus, impairment of thehost's defence mechanisms very early in lifemay lead to death, if not directly from worminfections, perhaps from secondary bacterialinfections. In addition, it is possible that aswollen, heavily infected, bursa may pressupon adjacent organs and thereby interferewith their function.
In conclusion, we believe that the aquisitionof helminth parasites by bullfinches is almostcertainly restricted to the nestling stage, butsome species at least remain alive in the hostfor many months and beyond the stage atwhich the bird reaches sexual maturity. Ofthe five helminth species found only one,Urotocus rossittensis which inhabits the bursaFabricii, does any appreciable damage to the
Copyright © 2011, The Helminthological Society of Washington

82 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
host and might influence nestling and juvenilemortality.
Summary
Seventy-one of 1,265 (5.6%) bullfinches,Pyrrhula pyrrhula (L.), received from 37localities in 10 counties in southern Englandbetween January 1965 and January 1966 wereinfected with helminth parasites. These com-prised five species, three being trematodes andtwo cestodes, namely Brachylaima arcuataDujardin, 1845, Leucochloridium macro-stomum (Rudolphi, 1803), Urotocus rossit-ten-sis (Miihling, 1898), Hijmenolepis passeris(Gmelin, 1790) and Anomotaenia borealis(Krabbe, 1869). Urotocus rossittensis was themost common parasite with an incidence of4%, although in birds received from Berkshireand Hampshire the incidence was locally ashigh as 17%; the other four species were com-paratively uncommon. Brachylaima arcuatahas not previously been reported from theBritish Isles, and neither this species norAnomotaenia borealis have been recordedbefore from the bullfinch.
The incidence and intensity of infection ofthe five species with respect to age, sex, andlocality is considered, and the pathology ofinfection of the bursa Fabricii of the bullfinchwith U. rossittensis is described. It is believedthat infection of the bullfinch with helminthparasites takes place during the nestling stagebecause only then is animal food included inthe bird's diet.
Literature Cited
Ackerman, G. A., and R. A. Knouff. 1964.Lymphocytopoietic activity in the Bursa ofFabricius. In The Thymus in Immunobiology(ed. Good, R. A., and A. E. Gabrielson), pp.123-49. London: Harper and Row.
Click, B. 1964. The Bursa of Fabricius andthe development of Immunologic compe-tence. In The Thymus in Immunobiology (ed.Good, R. A., and A. E. Gabrielson), pp.343-58. London: Harper and Row.
James, B. L., and L. C. Llewellyn. 1967. Aquantitative analysis of helminth infestationin some passerine birds found dead on theisland of Skomer. J. Helminth. 41: 19-44.
Joszt, L. 1962. Helminth parasites of sparrow—Passer domesticus (L.) in the environmentof Warszawa. Acta Parasit. Pol. 10: 113-6.
Kintner, K. E. 1938. Notes on the cestodeparasites of English sparrows in Indiana.Parasitology 30: 347-57.
Newton, I. 1964. Bud-eating by Bullfinches inrelation to the natural food-supply. J. Appl.Ecol. 1: 265-79.
. 1966. The moult of the bullfinchPyrrhula pyrrhula. Ibis 108: 41-67.
—-——. 1967a. The feeding ecology of theBullfinch (Pyrrhula pyrrhula L.) in southernEngland. J. Anim. Ecol. 36: 721-44.
. 1967b. The adaptive radiation andfeeding ecology of some British finches. Ibis109: 33-98.
-, and I. C. Williams. 1967. A trema-tode infection of Bullfinches Pyrrhula pyr-rhula. Ibis 109: 110-3.
Owen, R. W., and R. T. Pemberton. 1962.Helminth infection of the Starling (Sturnmvulgaris L.) in northern England. Proc. Zool.Soc~ Lond. 139: 557-87.
Sokolova, G. A. 1959. [Dynamics of the para-site fauna of the Chaffinch (Fringilla coelebsL.).] Vest, leningr. gos. Univ., 3, Ser. Biol.1: 83-90. (In Russian.)
Timon-David, J. 1957. Recherches experimen-tales sur le cycle evolutif d'un trematode dugenre Urotocus Looss (Digenea, Leuco-chloridiidae). Annls Parasit. Hum. Comp.32: 219-42.
Wright, E. N., and D. D. B. Summers. 1960.The biology and economic importance of theBullfinch. Ann. Appl. Biol. 48: 415-8.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 83
Euparadistomum cercopitheci sp. n. (Dicrocoeliidae), a DigeneticTrematode from the Talapoin Monkey from Rio Muni1
JACOB H. FISCHTHAL
The specimens of this report were collectedby Robert W. Cooper, D.V.M.,- Director, NIHPrimate Colony, Institute of Comparative Biol-ogy, Zoological Society of San Diego. Theywere brought to the author's attention by G. E.Cosgrove, M.D., Biology Division, Oak RidgeNational Laboratory. Dr. Cooper (personalcommunication) noted that "The talapoinmonkey from which the five trematodes werecollected was received in our colony from Bata,Rio Muni (West Africa) on 26 April 1968.This animal was an adult female. . . . Talapoinmonkeys are extremely common in Rio Muniand it is unlikely that the specimen was cap-tured more than 25 miles from Bata. . . . In atotal of 20 examinations performed on tala-poins in Rio Muni, in Cameroun, and here inSan Diego, I have found this particular trem-atode on only one other occasion. In this casea single immature specimen was recoveredfrom the gall bladder of a freshly capturedadvanced pregnant female which was pur-chased in Bata from a native on 3 January1968." Specimens were fixed, without pres-sure, in Roundabush solution, stained in Semi-chon's acetocarmine or Mayer's carmalum, andmounted in permount. Measurements are inmicrons.
Euparadistomum cercopitheci sp. n.(Figs. 1-4)
HOST: Cercopithecus (Miopithecus) tala-poin Schreber (Cercopithecidae).
HABITAT: Gall bladder.LOCALITY: Vicinity of Bata, Rio Muni;
Equatorial Guinea.SPECIMENS: USNM Helm. Coll. No. 71288
(holotype); No. 71289 (paratypes).DIAGNOSIS (based on five mature specimens
from one host): Body flat, slightly longer than
1 Contribution from the Department of Biology, StateUniversity of New York at Binghamton, Binghamton, NewYork 13901.
- Under contract No. PH43-63-56 within the SpecialVirus Leukemia Program of the National Cancer Institute,NIH, USPHS, covering studies of the biology and potentialbiomedical research utility of talapoins and other smallprimates.
wide, anterior extremity round, posterior nearlytruncate, tending toward trigonal shape, 3,180-4,385 by 2,630-3,920. Tegument thick, annu-lated; papillae and spines absent. Forebody1,300-1,815 long, hindbody 997-1,450 long;forebody-hindbody length ratio 1:0.70-0.87;preoral space 60-115 long. Suckers trans-versely elongate; oral sucker subterminal ven-tral, 675-835 by 760-974; acetabulum 860-1,166 by 997-1,304, center at level of anterior52-57 per cent of body length; sucker lengthratio 1:1.22-1.39, sucker width ratio 1:1.29-1.45. Prepharynx absent; pharynx usuallylongitudinally elongate, 205-290 by 200-230,entirely dorsal to or considerably overlappingoral sucker; esophagus 70-450 long; cecalbifurcation 335—510 preacetabular; ceca wide,terminating 220-560 from posterior extremity.
Testes two, situated symmetrically at antero-lateral margins of acetabulum in three speci-mens, oblique with left testis anteromedian toacetabulum in two others, usually overlappingacetabulum, margins smooth to slightly lobed,usually transversely elongate but sometimeslongitudinally elongate, right testis 225-335by 225-360; left testis 245-300 by 260-365.Vasa efferentia uniting at cirrus sac. Lattermedian to submedian, straight to curvingslightly, 250-440 by 125-160, lying 75-160preacetabular, containing saccular, undulatingseminal vesicle, short pars prostatica sur-rounded by prostate cells, and muscular cirrus.Genital atrium round, wide, shallow. Genitalpore median, bifurcal to postbifurcal, situated205-520 postpharyngeal, 205-475 preace-tabular.
Ovary smooth to slightly lobed, median tosubmedian (left), overlapping posterior partof acetabulum, longitudinally or transverselyelongate, 265-375 by 290-335. Mehlis' glandwell developed, very compact, round to longi-tudinally elongate, situated dextral to ovaryin three specimens, sinistral in two others,usually overlapping ovary dorsally, 140-205by 120—185. Seminal receptacle situated asfor Mehlis' gland, large, saccular, longitudinally
Copyright © 2011, The Helminthological Society of Washington

84 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Figures 1-4. Euparadistomum cercopithecL 1. Whole mount, holotype, ventral view. 2. Position oftestes in two paratypes, ventral view. 3. Terminal genitalia, paratype, ventral view. 4. Annulated tegu-ment, holotype, anterolateral part of body at level of cecal bifurcation.
Abbreviations: C, cirrus; CS, cirrus sac; GA, genital atrium; GC, gland cells; GP, genital pore; M,metraterm; PC, prostate cells; PP, pars prostatica; SV, seminal vesicle; U, uterus; VE, vas efferens.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 85
elongate, 300-385 by 195-220. Laurer's canalextending to dorsal surface near seminal recep-tacle. Vitelline follicles in lateral extracecalfields, extending from pretesticular, testicularor acetabular level to level anterior or pos-terior to or at cecal ends, anterior and/or pos-terior limits of fields in individual specimenmay be subequal, one or both fields may beinterrupted at ovarian level or both may beuninterrupted. Uterine coils extending to pos-terior extremity, overlapping vitellaria ven-trally, anteriorly extending to sides of oralsucker and lateral body margins; coiling essen-tially as schematically presented by Buckleyand Yeh (1958) in figure 3 of their newspecies Euparadistomum heischi. Metratermshorter than cirrus sac, muscular, anteriormostextent anterior to cirrus sac and genital pore,surrounded by gland cells. Eggs numerous,operculate; shells of younger eggs yellow incolor, becoming yellow-brown, and finallybrown as they progress from proximal to distaluterine coils; 20 older eggs measuring 34-41by 22-26.
Excretory bladder Y-shaped, stem post-ovarian, arms extending to ovarian level; poreterminal.
DiscussionSpecies of Euparadistomum Tubangui, 1931,
have been reported from lizards, birds, andmammals from the Philippine Islands, NorthBorneo, British Solomon Islands, Malaya,Burma, India, Madagascar, Belgian Congo,Kenya, and Brazil. African species are: E.varani var. madagascariensis Capron, Deblockand Brygoo, 1961, from chamaeleonid lizardsfrom Madagascar; E. pipistrelli (Sandground,1937) Travassos, 1944, from a vespertelionidbat from the Belgian Congo; E. heischi Buck-ley and Yeh, 1958, from a domestic cat(Felidae) from Kenya. Species from mam-malian hosts, in addition to the latter twolisted above, are: E. cerivoulae Gogate, 1939,from a vespertelionid bat from Burma; E.
paraense (Jansen, 1941) Travassos, 1944,from a didelphid marsupial from Brazil; E.buckleyi S. N. Singh, 1958, from a fox (Can-idae) from India. E. cercopitheci sp. n. ap-pears closest to E. paraense, E. buckleyi, andE. francolini R. Gupta, 1959 (from a phasianidbird from India). It differs from them in bodyshape, and in possessing an annulated tegu-ment. It differs further from E. paraense inlacking tegumental papillae, and in havingsmaller eggs and sucker ratios; from E. buck-leyi in possessing a well developed Mehlis'gland; and from E. francolini in having smallereggs.
The differences cited above for declaringE. cercopitheci a new species appear to berelatively small in view of experimental studieson host influenced intraspecific morphologicalvariations reported for Opisthioglyphe ranae(Frolich, 1791) Looss, 1907 (Plagiorchiidae)by Grabda-Kazubska (1967), and for Telor-chis bonnerensis Waitz, 1960 (Telorchiidae)by Watertor (1967). However, experimentaldata on morphological variability and on lifecycles are not known for any species ofEuparadistomum. Additionally, E. cercopitheciis from Africa, whereas the most closely relatedspecies are either from Brazil or India. There-fore, the author feels that the new speciesdesignation for the present specimens isjustified.
Literature CitedBuckley, J. J. C., and L. S. Yeh. 1958. On
Euparadistomum heisclii n. sp. from the liverof a domestic cat on Pate Island, Kenya, anda new sub-family Euparadistominae (Dicro-coeliidae). J. Helm. 32: 81-88.
Grabda-Kazubska, B. 1967. Morphological var-iability of adult Opisthioglype ranae (Frolich,1791) (Trematoda: Plagiorchiidae). ActaParasit. Polonica 15: 15-34.
Watertor, J. L. 1967. Intraspecific variationof adult Telorchis bonnerensis (Trematoda:Telorchiidae) in amphibian and reptilianhosts. J. Parasit. 53: 962-968.
Copyright © 2011, The Helminthological Society of Washington

86 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Survival and Egg Laying of Turtle Blood Flukes (Trematoda:Spirorchiidae) on the Chick Chorioallaiitois1
BERNARD FRIED AND RONALD E. TORNWALLBiology Department, Lafayette College, Easton, Pennsylvania 18042
Turtle blood flukes, tentatively identifiedas Spirorchis elegans Stunkard, 1923 from theesophagus, and Spirorchis scripta Stunkard,1923 from the head of naturally infected mid-western painted turtles, Chrysemys picta bellei(Gray), have been transplanted to the chickchorioallantois (Fried and Tornwall, 1965),to determine if this site would be suitable forhandling spirorchiid flukes outside of the host.The details are presented herein.
Materials and Methods
Chrysemys picta bellei obtained from a com-mercial dealer (Fried, 1965) were decapitatedand the heads cut midsagittally to expose thebrain and cranial cavity. Spirorchis scriptawere removed from the surface of the brainand cranial cavity and transferred throughseveral changes of sterile Ringer's solution(Paul, 1960). A turtle's plastron was severedwith bone cutters and the esophagus was re-moved and placed in Ringer's solution. Spiror-chis elegans dissected from the submucosa andpericonnective tissue of the esophagus weretransferred through several changes of sterileRinger's.
Fertile white leghorn hens' eggs were incu-bated at 37-38 C for 9-13 days. Sterilizationprocedures, methods used to operate on eggsand to implant spirorchiids on chorioallantoicmembranes were as previously described foreye flukes (Fried, 1962). Spirorchiids weremaintained on chorioallantoic membranes for1-3 days postimplantation at 30 ± 1 C andeggs were examined as previously described(Fried, 1962).
One hundred and nineteen spirorchiids wereplaced on the chorioallantoic membranes of109 eggs. One to three flukes were used peregg and the membranes were examined 1—5days later. Fifty-nine S. scripta were placed
1 Supported in part by a research grant from the Ad-vanced Studies and Research Fund of Lafayette Collegeand in part by NIH Grants AI-03443 and AI-06835.
on the chorioallantoic membranes of 56 eggs,and 60 S. elegans were placed on the mem-branes of 53 eggs.
Results
The results of the transplant experimentsare presented in Table 1.
Twenty-eight of the 119 chorioallantois-spirorchiids, including the 11 flukes recoveredlive, laid from 1-125 eggs on chorioallantoicmembranes. One live fluke recovered 2 daysafter transplantation laid about 70 eggs on thechorioallantois, and a second worm recovered3 days postimplantation laid about 125 eggson the chorioallantois. Fifty chorioallantois-spirorchiid eggs incubated in pond water atroom temperature for 10 days failed toembryonate.
S. elegans and S. scripta removed from tur-tles and maintained at room temperature inRinger's survived no longer than 6 hr and didnot lay eggs.
Live parasites were sluggish on the chorioal-lantois and did not attach firmly to its surface.However, when removed to Ringer's 1-3days after transplantation, chorioallantois-flukes were as active as in vivo spirorchiidsplaced in Ringer's, and displayed characteristicturtle blood fluke undulations. Dead chorioal-lantoic-flukes were either disintegrated andeasily lifted from the chorioallantois or werewalled off by the chorioallantois and firmlyattached to its surface.
Discussion
The fact that 11 (15%) of the 74 spirorchiidsrecovered on the chorioallantois survived for1—3 days postimplantation at 30 ± 1 C (Table1) indicates that chick eggs can provide ade-quate physicochemical conditions for turtleblood flukes for at least 3 days. Since 28spirorchiids laid eggs on the chorioallantois,including two specimens that laid an averageof 35-40 eggs per day for 2-3 days, turtle
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 87
Table 1. Survival of spirorchiids on chorioallantoic membranes at 30 ± 1 C.
Exper.species
S. scriptaS. elegansTotal
No. ofeggsused1
5653
109
No. ofwormsused2
5960
119
No. of dayson chorio-allantois
1-51-51-5
No. of liveworms
recovered3
56
11
No. of deadworms
recovered1
313263
No. ofcontaminatedor dead eggs
121022
1 Nine to 13 days old.2 One to 2 worms per egg.3 One to 3 days postimplantation.4 On shell or chorioallantois.
blood flukes can produce and lay eggs in thissite. Spirorchiid eggs from the chorioallantoisfailed to embryonate, possibly because intra-specific mating is necessary for normal eggproduction.
A survival rate of 15% indicates that con-ditions on the chorioallantois although ade-quate are suboptimal. Physicochemical con-ditions on the chorioallantois differ from thosein normal sites, esophageal submucosal andpericonnective tissue, and cranial cavity andsurface of the brain. Spirorchiids in theirnormal sites are usually surrounded on all sur-faces by host tissue and fluid, whereas thechorioallantois makes contact only with theventral surface of the worm. Moreover, spiror-chiids have a weak oral sucker, lack an ace-tabulum and pharynx and might not be ascapable of obtaining blood and tissue fromthe chorioallantois as are trematodes that havea pharynx and well-developed suckers andhave been maintained on the chorioallantoicmembrane for periods in excess of 3 days, i.e.,Philophthalmus hegeneri and Polystomoides sp.(Fried, 1962; 1965).
Specimens of Spirorchis scripta obtainedfrom the head and Spirorchis elegans from theesophagus of Chrysemys picta bellei are ref-erable to the descriptions of S. scripta andS. elegans by Stunkard (1923), Byrd (1939),and Schroecler and Ulmer (1959). Goodchildand Kirk (1960) described the life history ofSpirorchis elegans from the heart of Chrysemyspicta picta in Georgia. It is questionablewhether their S. elegans is the same as theS. elegans used in this study. The possibilityalso exists that S. scripta and S. elegans fromChrysemys picta bellei are morphological var-iants of the same species.
Summary
Turtle blood flukes, tentatively identified asSpirorchis elegans from the esophagus, andS. scripta from the head of Chrysemys pictabellei, have survived on the chick chorioal-lantois at 30 ± 1 C for up to 3 days. Spiror-chiids maintained in Ringer's at room tempera-ture survived no longer than 6 hours and didnot lay eggs.
Eleven of 119 spirorchiids transplanted tothe chorioallantoic membranes of 109 9- to 13-day-old chick embryos maintained at 30 rt 1 C,were recovered live and laid eggs up to 3 daysafter transplantation. One chorioallantoic-flukerecovered 2 days after transplantation laid ap-proximately 70 eggs, and a second worm re-covered 3 days postimplantation laid about125 eggs on the chorioallantois. Attempts toembryonate these eggs in pond water wereunsuccessful.
AcknowledgmentsWe wish to thank Mr. Jack Carty for
valuable technical assistance.
Literature CitedByrd, E. E. 1939. Studies on the blood flukes
of the family Spirorchiidae. Part II. Revisionof the family and description of new species.J. Tenn. Acad. Sci. 14: 116-161.
Fried, B. 1962. Growth of Philophthalmus sp.(Trematoda) on the chorioallantois of thechick. J. Parasit. 48: 545-550.
. 1965. Transplantation of a monoge-netic trematode, Polystomoides sp., to thechick chorioallantois. J. Parasit. 51: 983-986.
, and R. E. Tornwall. 1965. Survivaland egg-laying of turtle blood flukes (Trema-toda: Spirorchiidae) on the chick chorioal-lantois. J. Parasit. 51(Suppl.): 46.
Goodchild, C. G., and D. E. Kirk. 1960. The
Copyright © 2011, The Helminthological Society of Washington

88 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
life history of Spirorchis elegans Stunkard,1923 (Trematoda: Spirorchiidae) from thepainted turtle. J. Parasit. 46: 219-229.
Paul, J. 1960. Cell and Tissue Culture. 2nded. Williams & Wilkins Co., Baltimore, Md.
Schroeder, P. J., and M. J. Ulmer. 1959.
Host-parasite relationships of Spirorchis ele-gans Stunkard (Trematoda: Spirorchiidae).Proc. Iowa Acad. Sci. 66: 443-454.
Stunkard, H. W. 1923. Studies on NorthAmerican blood flukes. Bull. Amer. Mus.Nat. Hist. 48: 175-221.
Pathogenicity of the Northern Root-knot Nematode(Meloidogyne hapla) to Potato1
G. D. GRIFFIN AND E. C. JoRGENSON2
The northern root-knot nematode, Meloi-dogyne hapla, is considered a serious pest ofpotatoes in the Western United States. Wehave seen fields where approximately all ofthe tubers are either infected or galled by thisnematode, making them commercially unde-sirable. We have also observed the occurrenceof a deep-seated tuber infection (nematodesnear the center of the potato tuber). We sus-pected that age of plant, soil temperature, andsoil moisture determine the extent of invasionand severity of galling of potato roots andtubers. Gross (1940) found galling of potatotubers by Heterodera marioni (Meloidogynesp.) less in soil irrigated every 3—5 days at asoil temperature of 17-18 C, than under lessfrequent irrigations of 7—9 days where soiltemperature went as high as 26.5 C. Godfrey(1926) reported an increase in root-knotgalling of potato plants up to 27 C. Parris(1948) reported that the higher the soilmoisture, the greater tuber galling by Hetero-dera marioni (Meloidogyne sp.). He foundlarvae near the cambium below swollen len-ticels and concluded that nematodes enteredthrough lenticels of tubers grown in soil witha high moisture content. Godfrey (1926)found that optimum root-knot galling of to-mato roots occurred between 50 and 80%field capacity, and Peacock (1957) obtainedsimilar data. Wallace (1956) observed a
1 Cooperative investigation of Crops Research Division,Agricultural Research Service, United States Departmentof Agriculture and the Utah State Agricultural ExperimentStation. Journal Paper No. 747.
2 Research Nematologists, Crops Research Division, Agri-cultural Research Service, United States Department ofAgriculture, Crops Research Laboratory, Utah State Uni-versity, Logan, Utah.
decline in the mobility of Heterodera schachtiias soil moisture decreased, which may directlyaffect root infection.
The role of host age on root infection byroot-knot nematodes was observed by Christie(1936), who stated that second stage root-knotlarvae usually enter roots near the tip. Thisaccounts for the congregating of nematodesat or near the meristematic root tissue up to 24hr until the root is penetrated (Godfrey andOliveira, 1932; Peacock, 1959; Loewenberg,Sullivan, and Schuster, 1960). The ability ofroot-knot larvae to penetrate only delicate roottissue is evidenced by the galling of only thetip of large tap roots, in some instances infumigated soil.
This study was made to determine (1) therelationship between soil temperature, soilmoisture, and the pathogenicity of Meloi-dogyne hapla on potato under controlled con-ditions, and (2) if size or age of roots andtubers affect penetration and galling by root-knot larvae.
Materials and Methods
To study the effect of temperature on patho-genicity of M. hapla Chitwood on potato(Solanum tuberosum L.), Russet Burbankpotato seed pieces were surface sterilized with1:600 solution of rnethylmercury dicyandi-amicle for 60 sec, suberizecl for 24 hr andplanted in flats of fumigated soil. After 15days, plants, minus seed pieces, were trans-planted in 3.8 liter polyethylene containers ofProvo sand (M.H.C. = 17%) and inoculatedwith 1,000 M. hapla larvae. Plants were grown
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 89
Table 1. Effect of temperature on pathogenicity of Meloidogyne hapla to Russet Burbank potatoes.1
°c20253035
Roots
Avg wt.
4.16b3.48b4.51b0.25a
Gall rating2
2.14d4.00e3.71e0.67c
No. per plant
2.00f3.00f00
Tubers
Avg wt.
6.24g2.85g
Gall rating2
00
—-
Numbers with same letter do not differ significantly at 5% level.1 1,000 M. hapla larvae per plant.2 Gall rating: 0, none; 1, very few; 2, slight; 3, moderate; 4, severe; 5, very severe.
at temperatures of 20, 25, 30, and 35 C. Eachtreatment (temperature) was replicated fourtimes, and grown under a 16 hr day. Theexperiment was concluded after 60 days, andplant growth and iiematode pathogenicitydetermined.
The relationship between galling and plantage was determined by inoculating RussetBurbank seed pieces, and 3- and 6-week-oldplants (minus seed pieces and rhizomes) with1,000 larvae of M. hapla. Plants were grownon a greenhouse bench in 15.2 cm clay potsat 22 ± 4 C; treatments consisted of sixinoculated and six control plants. The experi-ment was ended after 140 days, and tuberand root weights, and nematode pathogenicitydetermined.
The effect of tuber size on nematode infec-tion was studied by inoculating plants fromwhich all but one potato bud or tuber wereremoved. Twenty-five 12-week-old RussetBurbank plants, each labeled as to size of budor tuber, were inoculated as previously statedand transplanted into a 15.2 cm clay pot ofProvo sand. Potato buds and tubers rangedfrom 5—57 mm in diameter. These plants weregrown for 60 days at 22 ± 4 C, and tuberinfection was determined.
Two methods were used to study the effectof soil moisture on infection and galling. The"percent soil moisture method" involved trans-planting 15-day-old plants into 3.8 liter poly-ethylene containers of Provo sand and inoculat-ing each plant with 1,000 larvae of M. hapla.Containers of soil with the moisture levels of25, 50, 75, 100, and 125% of field capacity(F.C.) were replicated 6 times per treatmentin a chamber with relative humidity of 80—100%, and a soil temperature of 22 ± 2 C.Each container was weighed daily, and dis-tilled water added to maintain moisture at the
desired level. To be sure the soil moisture wasmaintained at the desired level, control plantsof comparable size were weighed weekly andcompensation was made for any increase inplant weight. The study was terminated after130 days; root and tuber growth and infectionand galling of roots and tubers by M. haplawas determined.
The other method, the "field capacitymethod," was similar except that the soilmoisture of all containers was initially adjustedto field capacity. Water was not added tothe containers until soil moisture had droppedto or below previously determined soil mois-ture levels of 25, 50, 75, and 100% E.G.; thenenough water was added to restore F.C.(Couch, Purdy, and Henderson, 1967). Repli-cates consisted of six inoculated and six con-trol plants at each soil moisture level. Plantswere grown for 80 days on a greenhouse benchat 22 ± 4 C, and root growth, and root infec-tion by M. hapla was recorded.
Results and DiscussionPotato plant growth was affected as much
by temperature as by the pathogenicity ofM. hapla. The least root growth occurred at35 C, but there were no significant differencesin root growth of plants from 20-30 C (Table1). However, tubers were formed only at20 and 25 C, and root galling -was most severeat 25 and 30 C; root galling, like root growth,was adversely affected at 35 C. It appears thata temperature of approximately 25 C is opti-mum for maximum expression of root gallingon Russet Burbank potato. The slight amountof galling at 20 C agrees with Gross (1940)who found galling of tubers to be less at 17-18C than at temperatures up to 26.5 C. Thisundoubtedly affects the degree of severity oftuber galling from year to year when differ-
Copyright © 2011, The Helminthological Society of Washington

90 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Table 2. Effect of age of plant at time of inoculation on root and tuber galling of Russet Burbankpotatoes by Meloidogyne hapla.
Age ofinoculated
plant
6 weeks — InoculatedControl
3 weeks — InoculatedControl
Seed pieces — InoculatedControl
Avg rootweight
(g )
0.78a5.1 Ob2.25a5.56b4.35b5.33b
Rootgall
rating1
2.00c
2.22c
2.29c
Avgtuber
weight
10.50d21.20e
8.60d20.74e
6.86d7.26d
Per centinfected
85
92
83
Per centtubersgalled
85
92
35
Tubergall
rating1
S.OOf
3.33f
3.57f
Numbers with same letter do not differ significantly at 5% level.1 1,000 M. hapla larvae per plant.Tuber and root gall rating: 0, none; 1, very few; 2, slight: 3, moderate; 4, severe; 5, very severe; temperature, 20-
25 C.
ences in soil temperatures are noted. However,we have no data on soil temperatures in potatofields, and can only speculate.
Tuber infection was not affected by differ-ences in plant age at time of inoculation.Tubers were readily infected on plants grownfrom inoculated seed pieces, and from 3- and6-week-old plants (Table 2). However, fewertubers were galled on plants from inoculatedseed pieces, which apparently was due to thelate tuber set as shown by the small-sizedtubers when compared with those grown from3- and 6-week-old plants. There were signifi-cant differences between root growth of con-trol and inoculated 3- and 6-week-old plants,which showed the pathogenicity of M. haplato potato plants. However, there were nosignificant differences between inoculated andcontrol plants from seed pieces, because morethan one root system developed from a singleseed piece.
Infection was not greatly affected by tubersize. Larvae were able to penetrate large andsmall tubers (Table 3). There were, however,a greater number of larvae infecting tubers
Table 3. Influence of bud and tuber size ofRusset Burbank potato on infection by Meloi-dogyne hapla.
Bud and tuberdiameter
^YniTv)1
5-1516-2526-4141-57
Per cent ofbuds and tubers
infected
100100100100
Degreeof
infection-
2.16 ± 0.422.42 ± 0.282. 65 ±0.363.17 ±0.51
1 Size at time of inoculation.-Degree of infection: 0, none; 1, light; 2, moderate;
3, heavy; 4, very heavy.
from 41—57 mm in diameter, which may beexplained by the greater surface area exposedto larval invasion.
The pathogenicity of M. hapla to RussetBurbank potato roots and tubers was affectedby soil moisture. Roots and tubers were galledat all moisture levels from 25-100% F.C. withthe "percent soil moisture method" (Table 4) .Tubers were not galled at 125% F.C., but 74%of the tubers were infected. Root galling wassignificantly less at 125% of F.C. than at othermoisture levels. This was apparently explainedby Shepherd and Wallace (1959), who foundthat the infection of hosts by HeteroderaschachtU and H. gottingiana was greatest whenmost of the soil pores are free of water. Thismay also be attributable to low oxygen andgaseous exchange in near saturated soil (VanGundy, Stolzy, Szuszkiewicz, and Rackman,1962).
Maximum root growth occurred at 50, 75,and 100% F.C., and minimum root growth wasat 25 and 125% F.C.
The "field capacity method" gave resultssimilar to those obtained with the "percentsoil moisture method," and resulted in root andtuber infection at 25, 50, 75, and 100% F.C.(Table 5). However, tubers were not galled,because of the short duration of the study. Thegreatest root growth occurred at 100%, while25% of F.C. resulted in the smallest root sys-tem. There \vere no differences in root weightsbetween control and inoculated plants at 25%F.C.; roots of these plants were reduced asmuch or more by low soil moisture as by nema-tode pathogenicity. However, at 50, 75, and100% F.C. roots of inoculated plants were sig-nificantly smaller than control plants, although
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 91
Table 4. Effect of soil moisture on root and tuber galling of Russet Burbank potatoes by Meloidogynehapla in controlled moisture chamber of 80-100% relative humidity at 22 — 2 C ("Percent soil moisturemethod").
Soil moisture( per cent field
capacity )
255075
100125
Avgroot wt.
( g )
15.80a26.32b23.66B29.60018.34a
Rootgall
rating1
4.38e4.60eS.OOfS.OOf3.18d
Per centtubers
infected
9596
10010074
Per centtubersgalled
9596
100100
0
Tubergall
rating
4.25g4.60gh4.67ghS.OOh0
Numbers with same letter do not differ significantly at 5% level.1 15-day-old transplants inoculated with 1,000 M. hapla larvae per plant, and grown for 130 days.Root and tuber gall rating: 0, none; 1, very few; 2, slight; 3, moderate; 4, severe; 5, very severe.
galling was as severe at 25 as at 50% F.C.The point of tuber penetration was appar-
ently through the lenticels, because most ofthe nematodes were found immediately belowor near the lenticels at 125% F.C. However,there was no affinity for the area below ornear the lenticels at the other soil moisturelevels, and an epidermal penetration remainsa possibility. Parris (1948) stated that root-knot larvae enter potato tubers through swollenlenticels. We found no significant differencesbetween tubers infected at 50 and 75% F.C.,although tuber lenticels were swollen only at75, 100, and 125% F.C. The low soil moisturesof 25 and 50% F.C. apparently inhibited len-ticel swelling, but did not prevent infectionand galling of tubers by M. hapla. Tubers withswollen lenticels at 100% F.C. were generallythe most heavily galled, which suggests thattubers may be penetrated more readily throughthe swollen lenticels. However, the very severetuber galling at 100% F.C. may be partly dueto the effect of soil moisture on the nematode
mobility (Wallace 1956) and migration of thenematode to the site of infection. Nematodeswere found in the tuber just outside the cam-bial layer at all moisture levels, and less than5% of the nematodes were found beneath thecambium. There was no deep seated infectionobserved in any of the infected tubers. It is,therefore, apparent that deep-seated infectionof tubers by M. hapla is fortuitous or causedby factor(s) not considered in this study.
Comparing the two methods used for con-trolling soil moisture, the "field capacitymethod" is considered the better method sinceit more closely compares to field conditions,and is more easily controlled in the greenhouse.
Although there were significant differencesin root and tuber infection and galling betweenthe different soil moisture levels, it appearsthat soil moisture is not of major importancein the host—parasite relationship. Over 70%of all tubers were infected at all soil moisturelevels; root and tuber galling was severe orvery severe at all moisture levels but 125%
Table 5. Effect of soil moisture on root galling of Russet Burbank potatoes by Meloidogyne hapla ongreenhouse bench at 22 — 4 C ("Field capacity method").
Soil moisture(per cent field
capacity )
25 Inoculated25 Control50 Inoculated50 Control75 Inoculated75 Control
100 Inoculated100 Control
Avgroot wt.
(g)
4.42a4.86a7.86b
10.71c8.09b
12.29c8.31b
16.30d
Rootgall
rating1
4.24e
4.49e.
S.OOf
S.OOf
Per centtuber
infected
71
—100—
100—
100—
Per centtubersgalled
0-0-0-0-
Numbers with same letter do not differ significantly at 5%.1 15-day-old transplants inoculated with 1,000 M. hapla larvae per plant and grown foi-l-loot gall rating: 0, none; 1, very few; 2, slight; 3, moderate; 4, severe; 5, veiy severe.
clays.
Copyright © 2011, The Helminthological Society of Washington

92 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
F.C.; nematode infection occurred outside thecambial layer, and very few (less than 5%)were found immediately below the cambiallayer. However, root weights of infected plantsat all soil moisture levels but 25% F.C. weresignificantly less than those of uninfectedplants. Therefore, we conclude that variationin soil moisture is important only in the degreeof pathogenicity, and while it may contributeto difference in severity of galling, the presenceor absence of root and tuber galling, and deep-seated potato tuber infection are conditionedby other factor(s).
SummaryRoot galling of Russet Burbank potatoes by
the northern root-knot nematode, Meloidogynehapla, increased as temperature increased from20-30 C. Roots were slightly galled at 20 C,and severely galled at 25 and 30 C. Therewas a very light galling at 35 C, which wasassociated with very little root growth. Therewere no differences in root growth from 20-30C, but tubers were formed only at 20 and 25 C.
Tuber infection was not affected by plantage or tuber size at time of inoculation.
M. hapla infected potato tubers at moisturelevels from 25-125% F.C. in Provo sand. Thegreater number of larvae infected tubers at100% F.C., and the least at 125%. Infectionwas mainly through the lenticels.
Literature CitedChristie, J. R. 1936. The development of
root-knot nematode galls. Phytopathology26: 1-22.
Couch, H. B., L. H. Purdy, arid D. W. Hender-son. 1967. Application of soil moistureprinciples to the study of plant disease.
Virginia Polytecnic Inst., Dept. Plant Path.Bull. No. 4. 23 pp.
Godfrey, G. H. 1926. Effect of temperatureand moisture on nematode root knot. J.Agric. Res. 33: 223-254.
, and Juliette Oliveira. 1932. Thedevelopment of the root-knot nematode inrelation to root tissue of pineapple and cow-pea. Phytopathology 22: 325-348.
Gross, A. E. 1940. Controlling root-knot nema-tode in potatoes in the Klamath Basin bymeans of irrigation. Plant Dis. Reptr. Suppl.124: 142.
Loewenberg, J. R., T. Sullivan, and M. L.Schuster. 1960. Gall induction by Meloi-dogyne incognita incognita by surface feed-ing and factors affecting the behavior patternof the second stage larvae. Phytopathology50: 322.
Parris, G. K. 1948. Influence of soil moistureon the growth of the potato plant and itsinfection by the root-knot nematode. Phy-topathology 38: 480-488.
Peacock, F. C. 1959. The development of atechnique for studying the host-parasite rela-tionship of the root-knot nematode Meloi-dogyne incognita under controlled conditions.Nematologica 4: 43-55.
Shepherd, A. M., and H. R. Wallace. 1959.A comparison of the rates of emergence andinvasion of beet eelworm, Heterodera schactiiSchmidt and pea root eelworm Heteroderagottingiana Liebscher. Nematologica 4: 227-235.
Van Gundy, S. D., L. H. Stolzy, T. E. Szuszkie-wicz, and R. L. Rackman. 1962. Influ-ence of oxygen supply on survival of plant-parasitic nematodes in soil. Phytopathology52: 628-632.
Wallace, H. R. 1956. The emergence oflarvae from cysts of the beet eelworm,Heterodera schachtii Schmidt in aqueoussolutions of organic and inorganic substances.Ann. Appl. Biol. 44: 274-282.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 93
Some Gastrointestinal Helminths of Ondatra zibethicusLinnaeus, the Muskrat in Maryland1
JAMES B. ABRAMBiology Department, Maryland State College, Princess Anne, Maryland
The muskrat, Ondatra zibethicus, has beenan important wildlife entity in Maryland, bothfrom the standpoint of adding to the economyand in providing opportunity for scientificinvestigations. The State has long been recog-nized for its large production of pelts, andmuskrat meat is a table delicacy in the area.
Several muskrat studies have been con-ducted in the Dorchester County marshes onthe Eastern Shore. The biology and habitatsof the species have been written of exten-sively, but the aspect of parasitism has beenneglected. Not since the study of helminthsof Maryland muskrats by Price (1931), whodescribed four new trematode species, has asimilar study been reported.
The muskrats of the present study weretrapped near Thurmont in Frederick Countyand in the Dames Quarter marshes in SomersetCounty, from November 1966 through No-vember 1967.
The collecting areas were ecologically dif-ferent in several respects. The Thurmontstudy area consisted of five freshwater ponds,located within a 10-mile radius of one another,near the Catoctin range of the Blue RidgeMountains. The Dames Quarter marshes, in-fluenced by tides from Tangier Sound and theManokin River, are a preferred muskrat habi-tat. Their salinity averaged around 11.91during the study.
Materials and MethodsThe muskrats were captured in a Conibear
trap which kills the animals by drowning. Alltaken in the Thurmont area were Ondatrazibethicus zibethicus; all captured in theDames Quarter area were O. z. macrodon.Organs of the gastrointestinal tract, stomachthrough large intestine, of each muskrat wereseparately examined for helminths. An aliquottechnique or a sieving method was employed
1 A portion of a thesis submitted in partial fulfillment ofthe requirements for the degree of Doctor of Philosophy,Oklahoma State University, Stillwater, Oklahoma.
for helminth recovery. The aliquot techniqueinvolved the use of a dissecting microscope inexamining around 10% of the total volume ofthe contents of each organ. The sievingmethod entailed placing a wide strip of finegauze material over the top of a metallic testtube rack and slowly sieving the organ con-tents through the gauze. The helminths wereretained on the gauze and were removed by adissecting needle. Routine methods for thepreparation of the helminths followed. Thisincluded: staining trematodes with alumcochineal, borax carmine, Semichon's aceto-carmine, and Delafield's hematoxylin. Thenematodes were not stained. After dehydra-tion in ethyl alcohol, the helminths werecleared in beechwood creosote or methyl salicy-late and mounted with Canada balsam.
Results and DiscussionThe gastrointestinal tracts of the muskrats
examined yielded five species of trematodesand one nematode species, as shown in Table I.
Approximately 71% of the Thurmont areamuskrats and about 51% of those of the DamesQuarter area were infected with one or moreof the helminths listed. None of the threetrematode species recovered from the Thur-mont group was found in the Dames Quartergroup; neither trematode collected from thelatter group occurred in the Thurmont group.The nematode, Trichuris opaca Barker andNoyes, 1915, occurred in both localities. TheDames Quarter group had a higher incidenceof infection with it.
A study of the literature indicated that allof the helminths recovered in this study arewidely distributed in muskrats in this country.All prior reports hereinafter cited pertain tooccurrence in this host.
T. opaca reportedly occurs in Michigan(Ameel, 1944), Ohio (Rausch, 1946; Beckettand Gallicchio, 1967), New York (Edwards,1949), Wisconsin (Tiner, 1950), Colorado(Ball, 1952), Illinois (Gilford, 1954), Oregon
Copyright © 2011, The Helminthological Society of Washington

94 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Table 1. Results of survey of the gastrointestinal helminths of 154 Maryland muskrats.
Study area andhelminth
ThurmontEchinostoma rec/olutumQuinqueserialis quinqueserialisWardius zibethicusTrichuris opaca
Dames QuarterNudacotijle novicaEchinochasmus schivartziTrichuris opaca
Locationin host
Small intestineCaecumCaecumCaecum
Small intestineSmall intestineCaecum
No. animalsexamined
79797979
757575
No. animalsinfected
4219267
273
18
Per centinfected
53.224.032.98.9
36.04.0
24.0
(Senger and Neiland, 1955), Alaska (Duna-gan, 1957), and Pennsylvania (Anderson andBeaudoin, 1966).
Echinostoma revolutum (Frolich, 1802),which was recovered only from the Thurmontgroup, had the highest helminth infection rate(53.1%). This helminth also occurs in Massa-chusetts (Rankin, 1946), New York (Edwards,1949), Illinois (Gilford, 1954), Oregon (Sen-ger and Neiland, 1955; Rider and Macy,1947), Pennsylvania (Anderson and Beaudoin,1966), and Ohio (Beckett and Gallicchio,1967).
Wardius zibethicus (Barker and East, 1915),the second most frequently occurring helminthin the Thurmont muskrats, has been reportedin Ohio (Rausch, 1946; Beckett and Gallicchio,1967), Illinois (Gilford, 1954), and Penn-sylvania (Anderson and Beaudoin, 1966).
Quinqueserialis quinqueserialis Barker andLaughlin, 191.1, which also occurred only inThurmont muskrats, occurs in Tennessee (Har-wood, 1939), New York (Edwards, 1949),Maine (Meyer and Reilly, 1950), Illinois(Gilford, 1954), Oregon (Senger and Neiland,1955), Alaska (Dunagan, 1957), Utah (Sen-ger and Bates, 1957), Pennsylvania (Andersonand Beaudoin, 1966), and Ohio (Beckett andGallicchio, 1967).
Nudacotyle novica Barker, 1916, the mostfrequently occurring helminth in the muskratsof the Dames Quarter marshes, occurs in fresh-and brackish-water habitats, and in Texas(Chandler, 1941), Louisiana (Penn, 1942),Michigan (Ameel, 1944), Ohio (Rausch, 1946;Beckett and Gallicchio, 1967), New York (Ed-wards, 1949), Maine (Meyer and Reilly, 1950),and Illinois (Gilford, 1954).
Echinochasmus schwartzi Price, 1931, al-though originally described from muskrats of
the Dorchester County marshes, occurred inonly 4% of the Dames Quarter sample. It alsooccurs in Texas (Chandler, 1941), Louisiana(Penn, 1942), and Tennessee (Byrd andReiber, 1942).
On the basis of the limited studies reported,the helminth fauna of muskrats in Marylandappears to vary from one locality to another.In this particular study, the chief environ-mental factor that appears to be the most in-fluential on these parasite differences is theconsistent salinity of the coastal marshes. Thisfactor limits the ability of the freshwater gas-tropods, preferred intermediate hosts for cer-tain trematodes, to persist in the Dames Quar-ter marshes.
While this article does not dwell on theinfluence that ecology plays in controlling therange of helminths, it is clear that differencesin the biological and physical environment playa key role in parasite distribution.
Literature CitedAmeel, D. J. 1944. A life history of Nuda-
cotyle novica Barker, 1916 (Trematoda,Notocotylidae). J. Parasit. 30(4) : 257-263.
Anderson, D. R., and R. L. Beaudoin. 1966.Host habitat and age as factors in the prev-alence of intestinal parasites of the muskrat.Bull. Wildl. Dis. Assoc. 2: 70-77.
Ball, L. 1952. Notes on the helminth para-sites of muskrats from western Colorado.J. Parasit. 38: 83-84.
Beckett, J. V., and V. Gallicchio. 1967. Asurvey of helminths of the muskrat, Ondatraz. zibethica Miller, 1912 in Portage County,Ohio. J. Parasit. 53: 1169-1172.
Byrd, E. E., and R. J. Reiber. 1942. Mam-malian trematodes II. Three flukes fromsmall mammals. J. Tenn. Acad. Sci. 17:143-148.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 95
Chandler, A. C. 1941. Helminth parasites ofmuskrats in southeast Texas. T- Parasit. 27:175-181.
Dunagan, T. T. 1957. Helminth parasites ofAlaskan muskrats. Trans. Amer. Micr. Soc.76: 318-320.
Edwards, R. L. 1949. Internal parasites of cen-tral New York muskrats (Ondatra zibethicazibethica L.). J. Parasit. 35: 547-548.
Gilford, J. H. 1954. A survey of muskrathelminths in Illinois. J. Parasit. 40: 702-703.
Harvvood, P. D. 1939. Notes on Tennesseehelminths. IV. North American trematodesof the subfamily Notocotylinae. J. Term.Acad. Sci. 14: 332-340; 421-437.
Meyer, M. C., and J. R. Reilly. 1950. Para-sites of muskrats in Maine. Amer. Midi.Natural. 44: 467-477.
Penn, G. H. 1942. Parasitological survey ofLouisiana muskrats. J. Parasit. 28: 348-349.
Price, E. W. 1931. Four new species of trema-tode worms from the muskrat, Ondatrazibethica, with a key to the trematode para-
sites of the muskrat. Proc. U. S. Natl. Mus.79: 1-13.
Rankin, J. S. 1946. Helminth parasites ofbirds and mammals in western Masschusetts.Amer. Midi. Natural. 35: 756-768.
Rausch, R. L. 1946. The parasites of Ohiomuskrats. J. Wildl. Mgmt. 10: 70.
Rider, C. L., and R. W. Macy. 1947. Pre-liminary survey of the helminth parasites ofmuskrats in northwest Oregon, with descrip-tion of Hymenolepis ondatrae n. sp. Trans.Amer. Micr. Soc. 66: 176-181.
Senger, C. M., and J. W. Bates. 1957. Theoccurrence of Hymenolepis evaginata and H.ondatrae in Utah muskrats. Proc. Helm.Soc. Wash. 24: 141-142.
, and K. A. Neiland. 1955. Helminthparasites of some fur-bearers of Oregon. J.Parasit. 41: 637-638.
Tiner, J. D. 1950. Two new species of Trich-uris from North America, with redescrip-tions of Trichuris opaca and Trichuris leporis(Nematoda: Asphasmidia). J. Parasit. 36:350-355.
The Use of Irradiated Third-stage Larvae ofAngiostrongylus cantonensis as Antigen to ImmunizeAlbino Rats Against Homologous Infection1
SHERIDAN H. LEEParasitology Laboratory, Department of Animal Sciences, University of Hawaii, Honolulu, Hawaii 96822
Lim, Ow-Yang and Lie (1965) and Heyne-man and Lim (1965) reported that rats whichreceived one to three sublethal feedings ofnormal third-stage larvae of Angiostrongyluscantonensis were able to develop some degreeof resistance to reinfection as manifested bythe reduced worm burden. Like many otherhelminthic infections, no sterile immunity wasobtained (Sprent, 1963).
Jarret et al. (1959) reported success in ob-taining sterile immunity to Dictyocaulus vivi-parus by using irradiated larvae. This sug-gested that a similar result may also beobtained with A. cantonensis since the latter
1 This investigation was supported by Research GrantNB-04965 from the Institute of Neurological Diseases andBlindness, NIH, USPHS, to Dr. J. E. Alicata. Publishedwith the approval of the Director of the Hawaii Agricul-tural Experiment Station as Technical Paper No. 905.
has a long brain phase 28-day plus, in its lifecycle (Mackerras and Sandars, 1955).
Materials and Methods
Third-stage larvae of A. cantonensis wereharvested from laboratory infected giant Afri-can snails, Achatina fulica, by digesting mincedsnail tissue in \% pepsin and 1% concentratedhydrochloric acid solution. Digested snail tis-sue was sieved through a 40-mesh wire screeninto an Imhoff flask and allowed to settle for40 min. After the supernatant liquid wassiphoned off, the sediment was baermannizedin saline through two 16-ply gauze pads sup-ported by a 40-mesh copperwire sieve. Thelarvae were harvested 30 and 50 min later.Harvested larvae were rebaermannized and
Copyright © 2011, The Helminthological Society of Washington

96 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Table 1. Number of worms recovered from the lungs of immunized and control rats of Experiment I,II, and III following a challenge with 400 normal larvae.
Experiment I
Rat no.
123456
Immunized
948139614649
Control
152234198260173234
Experiment II
Immunized
6102009
Control
14517812272
124132
Experiment III
Immunized1
16 (2)14 (0)34 (6)14 (12)23 (8)19 (0)
Control
254238181134156223
1 Number of young adults on brain surface in parenthesis.
then washed three times in distilled water.About 10,000 washed larvae were placed in2 cc of distilled water in small screw cappedvials and irradiated with the desired dosage.
The radiation source was a 30,000-curiecobalt-60 irradiator located in the departmentof Food Science and Technology of the Uni-versity of Hawaii. When this study began, theoutput was 4.7 Kr/min at 5 inches. All larvaeirradiated were drawn from a common pool.
EXPERIMENT I. Twelve rats were used, eachbeing given one feeding of 200, 40 Kr treatedlarvae. Six of these rats were killed on the30th day after feeding; the other six werechallenged with 400 normal larvae each, 30days after feeding, and killed 42 days afterchallenge.
EXPERIMENT II. Six rats were used, eachwas given two feedings of 200, 40 Kr treatedlarvae with a 30-day interval between eachfeeding. Thirty days after the last feeding,each was challenged with 400 larvae and killed42 days later.
EXPERIMENT III. In this experiment, eachof six rats was given three feedings of 200,20 Kr treated larvae with a 15-day intervalbetween each feeding. Thirty days after thelast feeding, each rat was challenged with 400normal larvae and killed 42 days later.
An equal number of control rats were given400 normal larvae to accompany all the aboveexperiments. All the control rats were of thesame age as the experimental rats.
EXPERIMENT IV. Two rats were given 200,20 Kr treated larvae each. One of these ratswas killed 40 days later and the other waskilled 62 days later.
ResultsEXPERIMENT I. One adult female worm was
recovered from one of the six rats which
received 200, 40 Kr treated larvae each. Thefollowing numbers of worms were recoveredfrom the lungs of the experimental rats whichwere challenged with 400 normal larave: 94,81, 39, 61, 46, and 49. The number of wormsfound in each control rat was 152, 234, 198,260, 173, and 234 (Table I ) .
EXPERIMENT II. The number of wormsrecovered from the lungs of each experimentalrat, which had received two feedings of 200,40 Kr treated larvae before challenge, was 6,10, 2, 0, 0, and 9; whereas, the number ofworms found in each control rat was 145, 178,122, 72, 124, and 132 (Table I). All theworms recovered in both experiments werefertile and none of the rats had any wormsin the brain. None of the rats appeared tohave suffered any ill effects during the entireperiod of this experiment.
EXPERIMENT III. The following resultswere obtained from rats each of which re-ceived three feedings of 200, 20 Kr treatedlarvae before challenge: Rat 1 had two deadyoung adults on its brain surface and in itslungs were 16 adult worms, 7 males and 9females, four of the latter being sterile. Rat 2had in its lungs 14 adult worms, 5 males and9 females, five of the latter being sterile. Afew first-stage larvae were found in the lungtissue. Rat 3 had four live young adults andtwo dead young adults on its brain surface, 7males and 27 females, in its lungs. Five ofthe 27 females were dead with only theircuticle and degenerate digestive tract remain-ing. First-stage larvae were present in thelung tissue. Rat 4 had three live young adultson its brain surface plus the remains of ninedead young adults which appeared as yellow-ish green lines. In its lungs were 3 sterilemales and 11 females, four with eggs and
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 97
two without eggs. Five females were in theprocess of being absorbed. Rat 5 had sevenpartially absorbed young adults plus one liveyoung adult on its brain surface. In its lungswere 13 females, three without eggs, and 10males, seven of which were sterile. A fewfirst-stage larvae were present in the lungparenchyma. Rat 6 had no worm on its brainsurface. There were 8 males and 11 femalesin its lungs, seven of which were sterile. Nofirst-stage larvae were found in the lungparenchyma. The number of worms found inthe control rats for Experiment III was 254,238, 181, 134, 156, and 223 (Table I).
EXPERIMENT IV. Eighteen sterile wormswere found in the lungs of the rat killed onthe 40th day after being fed 200, 20 Kr treatedlarvae. There were no worms in its brain. Thesecond rat which was killed on the 62nd dayafter being fed 200, 20 Kr treated larvae hadsix sterile females in its lungs and six youngadults on the surface of its brain. Two of thesix young adults were still alive whereas theother four were dead.
Discussion
Experiment I indicated that a radiationdosage of 40 Kr was quite detrimental to thefurther development of the third-stage larvaeof A. cantonensis. Yet the antigenecity of these40 Kr treated larvae was not lost as indicatedby the reduced worm burden of the immunizedrats when challenged. This experiment alsoindicated that immunity acquired by the ex-perimental rats did not depend on the existenceof live worms within the rats.
In Experiment II, the complete absence ofworms in two of the experimental rats andthe relative low number of worms in otherrats of this group suggests that two feedingsof 200, 40 Kr treated larvae offers the bestimmunizing procedure.
Experiment IV indicated that most likely,all the sterile worms recovered from the experi-mental rats of Experiment III were developedfrom the 20 Kr treated larvae used for im-munization. It is interesting to note thatExperiment IV not only demonstrated thedetrimental effect of the dosage of 20 Kr tothe germ cells of A. cantonensis but alsorevealed that this dosage will prolong thebrain phase of this worm.
It would seem that the protracted stay ofyoung adults in the subarachnoid space shouldcause the rat to have a stronger immune re-sponse. However, the results of ExperimentsII and III do not support this idea, at leastwhen the quality of immunity is measured interms of worm burden after reinfection. Whythe dosage of 40 Kr makes the third-stage lar-vae of A. cantonensis serve as a better antigenas compared with 20 Kr against homologousinfection remains an unsolved but interestingquestion.
SummaryOf six rats which received two feedings of
200, 40 Kr irradiated third-stage larvae ofAngiostrongyhls cantonensis, two became com-pletely refractive and four showed significantresistance when challenged with normal larvae.Twenty Kr radiation not only destroyed thegerm cells of A. cantonensis, but also causedmost of the young adults to lose their abilityto migrate to the lungs of their definitive host.
AcknowledgmentThe author wishes to acknowledge the tech-
nical assistance of Dr. M. D. Upadhya inirradiating all the larvae used in this paper.
Literature CitedHeyneman, D., and B. L. Lim. 1965. Pro-
longed survival in rats immunized by asmall number of low-level doses of Angio-strongylus cantonensis and challenged with alethal level of infective larvae. Med. J.Malaya 20(2): 162-163.
Jarrett, W. F. H., F. W. Jennings, W. I. M.Mclntyre, W. Mulligan, N. C. C. Sharp,and G M. Urquhart. 1959. Immunolog-ical studies on Dictyocaulus viviparus incalves—double vaccination with irradiatedlarvae. Am. J. Vet. Res. 20: 522-526.
Lim, B. L., C. K. Ow-Yang, and K. J. Lie.1965. Natural infection of Angiostrongyluscantonensis in Malaysian rodents and inter-mediate hosts, and preliminary observationson acquired resistance. Am. J. Trop. Med.14(4): 610-617.
Mackerras, M. J., and D. F. Sandars. 1955.The life history of the rat lungworm, Angio-strongylus cantonensis (Chen) (Nematoda:Metastrongylidae). Austr. J. Zool. 3: 1-25.
Sprent, J. F. A. 1963. Parasitism—an intro-duction to parasitology and immunology forstudents of biology, veterinary science andmedicine. Williams and Wilkins, Baltimore,145 pp.
Copyright © 2011, The Helminthological Society of Washington

PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Hypsoperine ottersoni sp. n. (Nemata, Heteroderidae) InfestingCanary Grass, Phalaris arundinacea (L.) Reed in Wisconsin
GERALD THORNE1
Unusual types of nematode-incited gallswere observed on canary grass in the WindLake area near Kenosha, Wisconsin by Dr.Henry Otterson, Agricultural Chemist and Con-sultant, who submitted specimens for identifi-cation in 1958, which later were recognized asbelonging to the genus Hypsoperine Sledgeand Golden, 1964. Detailed studies were notbegun until 1963 when Arthur Weber under-took the project as a thesis problem in grad-uate work under the direction of Dr. K. R.Barker, University of Wisconsin, Madison.
Validity of the genus Hypsoperine has beenquestioned by Dr. M. W. Allen (personalcommunication). It is true that the elevatedperineal region sometimes is not evident andthe cuticle is not thickened on this species, asreported by Sledge and Golden (1964) forH. graminis. However, the physiological effecton the host, causing an unusual type of gallformation with larvae always lying in the rootwith heads toward the apex, is unlike that ofusual Meloidogyne infestations. The hemizonidof H. graminis is reported anterior to the ex-cretory pore, while that of H. spartinae isposterior, according to Rau and Fassuliotis(1965).
Infestation always takes place just behindthe apical cells and does not stop growth ofthe root which generally forms an arcuate,elongated gall and then proceeds to developa series of similar galls, each containing from1-6 females and in rare instances 15 or 20 mayoccur. Branch roots are produced on the outerperiphery of each gall and these in turn areinfested until a complicated, chainlike massof roots results (Fig. 1A). Occasionally aprogressive multiple infestation occurs in aterminal gall with mature egg-producing fe-males at the gall base, while elongated youngfemales are located near the terminus, resultingin a series of various-aged individuals (Fig.IB).
In all instances observed, the heads of the
nemas were directed toward the root terminus.Eggs usually are deposited in a gelatinousmatrix within the gall tissues, but in someinstances they and the matrix are extrudedoutside the root. Feeding takes place alongthe cells between the xylem tubes which fre-quently are multiplied and broken apart (Fig.IB). Even in extreme instances like this, theroot cap and terminal cells remain intact,enabling continued growth.
Hypsoperine ottersoni sp. n.-Morphology
FEMALE (10): 0.27 by 0.45 mm (0.18-0.32 by 0.39-0.52). Variable in form as indi-cated by these measurements (Fig. 2N—S).Spheroid to elongate pyriform with neck pro-jecting at different angles (Fig. 2N-S). Vulvaand anus usually on a slight elevation, butsometimes this is not present and the posteriorend is similar to that of Meloidogyne. Cuticle3—6 /j. thick on main part of body, leatherywith extremely fine striae except on head andneck where they are easily visible and formannules variable in width. Lip region un-striated, set off by constriction. A face view(Fig. 2L) shows six well-developed lips, thelateral ones being somewhat larger than thesubmedian. Amphid apertures slightly oval,located at apices of lateral lips. Vestibuleslightly sclerotized, forming a spear-guidingapparatus. Spear 10-12 ^ long with smallrounded knobs. Dorsal esophageal gland ori-fice almost adjacent to spear base. Anteriorportion of esophagus set off from median bulbby constriction. Bulb spheroid with refractivevalve and strong radial muscles. Basal bulba flattened, irregular shaped lobe extendingback over intestine, containing the usual threenuclei which generally are difficult to observe.Isthmus very short. Nerve ring obscure. Ex-cretory pore far forward, almost opposite spearbase (Fig. 2K). Intestine and body contentsdensely granular, making observations very
1 Professor Emeritus, Departments of Plant Pathology andZoology, University of Wisconsin.
" First used as a nomen nudum by Weber and Barker(1967).
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 99
Figure 1. A, Root of Phalaris arundinacea infested with Hypsoperine ottersoni. Note how secondaryroots spring from outer contours of the sickle-shaped galls. Upper left and right are females ejecting eggsfrom the galls as occasionally occurs. B, Unusual terminal gall with progressive infestation. The first lar-vae entering it are now adult females producing eggs, while those recently entering near the terminus arestill in early stages of development.
difficult. Ovaries two, elaborately convoluted(Fig. 2N), at maturity completely filling bodycavity. Vulva transverse. Vagina cylindroid,muscular. Perineal pattern with simplerounded striae, usually on a slight terminalelevation (Fig. 2T). Anal opening visiblefrom a lateral view and rarely from ventral(Fig. 3C). Neither spermatheca nor sper-matozoa observed.
Eggs (15) 45 by 105 (40-50 by 95-115),deposited before segmentation by young fe-males. Senile individuals may retain from 5-25 eggs which segment and develop into lar-
vae which probably are not ejected. A gelat-inous matrix surrounds eggs deposited eitherwithin the roots or into the soil.
MALE: (7) 0.9-1.0 mm long, rarely 0.7 mm;a = 34, rarely 40. Body twisted until a truelateral view of head and tail can be seen onlyby cutting specimens after fixation. Striaeabout 2 fj(, apart at midbody, narrower nearhead and sometimes absent on tail. Lateralfields about one-fifth body width, marked byfour incisures, the two median ones sometimesvery obscure. Lip region unstriated; labial discvisible only from a dorsal or Ventral view when
Copyright © 2011, The Helminthological Society of Washington

100 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Figure 2. A, B. Anterior and posterior portions of larva X 1,000. C, D. Male head lateral and dorsalviews. E, F, G. Lateral and dorsal views of posterior portion of males. H. Specimen with two testes. I,J. Variations in median esophageal bulbs. K, L, M. Head, face view and anterior end of female. N-S.Variations in form of females. T. Perineal pattern. U. Duboscquia sp. parasites.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 101
Figure 3. A, B. Lateral and dorsal views of posterior portion of female. C. Ventral view showing anus,vagina, uterus, and attachment of the two ovaries and saccate rectal glands.
the slitlike amphid apertures set it off fromthe lip contour (Fig. 2D). Cephalids obscure,visible only from a lateral view, located wellforward of the spear base as in Meloidogyne(Fig. 2C). Spear 14-16 ^ long with strong,backward-sloping knobs. Nerve ring one bodywidth, and excretory pore twice that distancebehind median bulb. Lumen of esophagusjoining intestine about one body width pos-terior to bulb, the junction being rather abruptas illustrated for the larvae (Fig. 2A). Hemi-zonid about two annules anterior to excretorypore. Basal esophageal lobes average aboutthree times as long as body width. Testes53-65% of body length, usually single, out-stretched, rarely reflexed. Males rare; among14, two were observed with double testes (Fig.2H). Spicula slightly arcuate 19-23 p, longbut difficult to measure because of the twistedbodies. Gubernaculum 3 or 4 ^ long, well-thickened, troughlike. Phasmids almost ter-minal (Fig. 2G). Tail variable in form andlength (Fig. 2E, F). Sperm ducts packedwith spheroid spermatozoa indicating thatmales may be functional although sperms werenot observed in the uteri.
LARVAE (10): 0.43-0.50 mm; a = 23-30,b and c measurements not made because theesophagus base and anal opening could rarelybe accurately determined. Lateral field markedby four fine incisures visible only on favorablespecimens. Framework of the rounded lipregion practically undeveloped. Vestibuleforming a minute sclerotized spear guide.Spear 13—15 p, long with well-developed knobs.Dorsal gland orifice about 4 fjl behind spear.Nerve ring adjacent to bulb, practically en-veloping the unusually short isthmus. Esoph-ageal lumen joining intestine about one body
width behind bulb. Dorsally the esophagusbase is short and rounded while ventrally it isextended in a long lobe 5-7 times the bodywidth. Excretory pore slightly behind nervering. Anal opening very obscure, often un-identifiable. Tail ending in an irregularlyclavate or knobbed terminus.
Holotype female, allotype male and otherspecimens as indexed under Hypsoperine 1,University of Wisconsin nematode collection.
TYPE HOST AND LOCALITY: Canary grass,Phalaris arundianacea (L.) Reed, HomerRanch, Wind Lake area, Wisconsin. Foundwherever canary grass is growing in Racine,Milwaukee, and Waukesha counties and nearWaupun in Green Lake County. Many obser-vations by Dr. Otterson and others have failedto reveal it attacks any crop plants.
Hypsoperine ottersoni is distinguished fromH. graminis, Sledge and Golden, 1964 by themore simple annulation and absence of laterallines in the perineal pattern, smaller males 0.9:1.5 mm, and irregular-clavate terminus of lar-vae. From H. spartinae Rau and Fassuliotis,1965, it is recognized by absence of labialannules (two in H. spartinae}, simple lines ofperineal pattern compared to elaborate ar-rangement, and presence of male hemizonidanterior to excretory pore which is posteriorin H. spartinae.
Perineal sections should be mounted in adrop of hard glycerin jelly to support thecover glass, otherwise the weight will producefolds and wrinkles which destroy the actualpattern of the striae.
Many larvae and some males were attackedwith sporozoans, Duboscquia sp.; in some in-stances these were observed in clusters of
Copyright © 2011, The Helminthological Society of Washington

102 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
eight, unlike any previously recorded (Fig.2U).
Literature CitedRau, G. J., and G. Fassuliotis. 1965. Htjpso-
perine spartinae n. sp., a gall-forming nema-tode on the roots of Smooth Cordgrass. Proc.Helm. Soc. Wash. 32(2): 159-162.
Sledge, E. B., and A. M. Golden. 1964. Hyp-soperine gramini-s (Nematoda: Heteroderi-dae), a new genus and species of plantparasitic nematode. Proc. Helm. Soc. Wash.31(1): 83-88.
Weber, A. J., and K. R. Barker. 1967. Biol-ogy of the pseudo root-knot nematode Hijpso-perine ottersoni. Phytopathology 57(7):723-728.
Studies on Freshwater Larval Trematodes. XXI. Two NewSpecies of Macrocercous Cercariae1
PIR NASIR, MARCOS TULIO DIAZ, Luis J. HAMANA SALAZAR, AND S. GUEVARALaboratorio de Parasitologia, Depto. de Biologia, Escuela de Ciencias,Universidad de Oriente, Cumana, Venezuela
Until now, only bivalve mollusks have beenreported as the first intermediate hosts of theknown macrocercous cercariae. This is thefirst record for the involvement of univalvesas well. The cercariae treated in this paper,as a result of comparative studies discussedlater, proved to be new species and have beennamed, respectively, Cercaria latigazica andC. yacalicola. The latter is readily distin-guished by the presence of a cercarial cham-ber, in the proximal part of its tail, in whichthe cercarial body is characteristically enclosed.The former lacks this structure and its tail isfrequently thrust out in a peculiar whiplikefashion. It is evident from the works of otherauthors that various species of tadpoles andnaiads of insects serve as the second interme-diate hosts for macrocercous cercariae whileurinary bladders of frogs, salamanders, andfishes are the usual sites of adult infections.In certain cases like Phyllodistomiim simile,the precociously encysted cercariae while stillinside sporocysts when experimentally fed totrout resulted in the development of corre-sponding adults (Thomas, 1958). Of course,we are not sure whether these "precociousmetacercariae" were formed by the penetra-tion of cercariae from without. Due partly tothe dearth of material, we have been unable
1 Supported in part by a grant from Comision de Desar-rollo e Investigaciones Cientificas de la Universidad deOriente, Venezuela.
to elucidate life cycles. Even if infected snailshad been more numerous, only a few cercariaeemerged at a given time; however, efforts arein progress. It may be mentioned here thaton one occasion C. yacalicola was escapingfrom the same specimen of the snail, Pomaceaglauca, which was also discharging two addi-tional cercariae, C. cumanensis Nasir, 1965and C. allomacarapanensis Nasir and Diaz,1968.
The recovery of sporocysts presented anintriguing situation. Although several snailswere dissected immediately after the appear-ance of cercariae, sporocysts were never en-countered in the usual sites like gills and hepa-topancreas. A careful examination revealed thepresence of a few, anteroposteriorly elongated,considerably thick-walled, pigmentecl sporo-cysts in the lumen of intestine.
All observations have been made on freshlyemerged cercariae, except measurements whichare based on specimens killed by irrigating withhot 10% formalin.
All measurements are in millimeters.
1. Cercaria latigazica sp. n.(Figs. 1, 2, 7)
HOST: Pomacea glauca (L.) .LOCALITY: La Chorrera, near Caripito; Los
Yacales, La Llanada de San Juan, near Uni-versidad de Oriente, Cumana, Venezuela.
DESCRIPTION: Body without spines, beset
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 103
Figures 1, 2, 7. Cercaria latigasica sp. n. 1. Cercaria, note peculiar structure of tail, stylet not shown.2. Stylet. 7. Sporocyst, note considerably thick wall and pigmented specks.
Figures 3-7. Cercaria yacalicola sp. n. 3. Stylet. 4. Body only, note two groups of penetration glands oneach side. 5. Cercarial chamber and part of tail. 6. Cercarial chamber along with distal, elongated swol-len region. 7. Sporocyst.
Copyright © 2011, The Helminthological Society of Washington

104 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
with 22 rows of setate papillae. Stylet withouta basal bulb (Fig. 2) . Central parts of suckersoccupied by papillae without setae and irreg-ularly distributed bristles. Periphery of bothsuckers with a row of setate papillae. Ventralsucker surrounded by a muscular fold. Pharynxabsent. Esophagus long, not extending to ven-tral sucker. Intestinal ceca extending to pos-terior end of body. Penetration glands small,six on each side along posterior part of esoph-agus, not extending to ventral sucker. Genitalrudiments represented by two cellular masses,one anterior and the other posterior to ventralsucker. Excretory vesicle tubular, with a swol-len posterior region; eight transversely elon-gated glands with granular contents on eitherside of vesicle. Main excretory ducts extendingas far anteriorly as about halfway in preacetab-ular region and after forming a small loop insame region dividing into anterior lateral andposterior lateral collecting excretory tubules.Flame cell formula: 2[(3+3+3) + (3+3+3+3+3+3+3+3)1 = 66. Tail without spines,without cercarial chamber, capable of extend-ing in a peculiar whiplike fashion. Measure-ments of 10 specimens: body 0.210-0.267 by0.057-0.090; tail 0.300-0.375 by 0.078-0.114,when extended more than twice as long as itsnormal length; oral sucker 0.040-0.044 indiam.; esophagus 0.060-0.065 long; ventralsucker 0.043-0.045 in diam.; stylet 0.013-0.016 by 0.003-0.005. Sporocysts 0.300-0.817by 0.206-0.404.
2. Cercaria yacalicola sp. n.(Figs. 3-7)
HOST: Pomacea glauca (L.).LOCALITY: Los Yacales, La Llanada de San
Juan, near Universidad de Oriente, Cumana,Venezuela.
DESCRIPTION: Body without spines, coveredwith setate papillae. Stylet without a basalbulb (Fig. 3). Central parts of suckers withpapillae without setae and irregularly distrib-uted bristles. Periphery of suckers borderedwith rows of setate papillae. Pharynx absent.Esophagus long, extending to ventral sucker.Intestinal ceca extending only slightly pos-terior to ventral sucker. Penetration glandswith uniformly granular contents, in twogroups on each side of body: one group ofthree posterior to oral sucker and another
group of five anterior to ventral sucker; thislatter group may extend laterally around ven-tral sucker. Genital rudiments composed oftwo cellular masses, one anterior and otherposterior to ventral sucker. Excretory vesicletubular, with a swollen posterior region.Twelve to 14 transversely elongated glandswith coarsely granular material on each sideof vesicle. Main excretory tubes dividing intoanterior lateral and posterior lateral collectingtubes at about equatorial level of preacetabularregion. Flame cell formula: 2[ (3+3+3) +(3+3+3+3+3+3+3)] = 60. Tail withoutspines or papillae, divided into two regions:proximal swollen region or cercarial chamberand a distal, narrow, elongated region whichoften presents irregular swellings. Measure-ments of 10 specimens: body of cercaria ex-pressed from cercarial chamber 0.235-0.370 by0.112-0.138; oral sucker 0.060-0.066 in diam.;esophagus 0.060-0.067 long; ventral sucker0.072-0.085 in diam.; stylet 0.027-0.033 by0.007-0.009; cercarial chamber 0.243-0.379 by0.159-0.300; distal part of tail without cer-carial chamber 2.302-2,570 by 0.159-0.300.Sporocysts similar to that of C. latigazica.
DiscussionFor the specific identification only those
characters which are fairly constant or varylittle like the shape and size of stylet, esoph-ageal length, number and arrangement ofpenetration glands, number of "cystogenousglands" around excretory vesicle, type of ex-cretory system mesostomate or stenostomate,number and arrangement of flame cells andfinally presence or absence of cercarial cham-ber have been considered. Unless the relativedifferences in size are of a considerable magni-tude, too much reliance on this character alonecould be misleading. Some "new species"erected on the basis of minor discrepanciesshould be suppressed. Cercaria eriensis Coil,1953 and C. lampsilae Coil, 1954 are indis-tinguishable in having approximately the samesize suckers, esophageal length, intestinallength, shape, and extent of excretory vesicle,similar number of glands around excretoryvesicle and finally both of the species aboundin the same locality, Lake Erie, Ottawa County,USA. Furthermore, the molluscan intermediatehosts are the very closely related species,
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 105
Lampsilis ventricosa and L. siliquoidea. Ac-cording to Coil, 1954, "C. lampsilae differsfrom C. eriensis, its closest relative, by possess-ing (1) a stylet which is slightly shorter andof a different shape, (2) a different dispositionof sensory papillae on the oral sucker, (3) abody length which is shorter." In our experi-ence none of these characters is of substantialtaxonomic importance. The "differences" be-tween the stylets of C. eriensis and C. lamp-silae, as depicted in diagrams (Coil, .1954; fig.5—6), are hardly differences at all, becausea slight pressure of coverglass will distort theshape of one into that of the other. We haveoften encountered this type of distortion whilestudying xiphidiocercariae. Insofar as stylet
I.
size is concerned there is only a difference of2 /x,; the stylet of C. eriensis is 14—17 ^ incontrast with 16-19 of C. lampsilae. The ar-rangement of papillae on oral sucker is alsoa matter of interpretation. Finally, the cri-terion of body size is also questionable. Cer-caria of Gorgodera amplicava Looss, 1899, asdescribed by Krull, 1934, is 4.4-7.5 mm longwhereas the same cercaria reported by Good-child, 1945, is 12.0-17.4 mm long. It is there-fore suggested that C. lampsilae should besynonymized with C. eriensis.
Only those styleted macrocercous cercariaeabout which sufficient structural details areknown are compared and these are separatedby the following key:
A. Tail with a cercarial chamber IIB. Tail without a cercarial chamber III
II. Tail with cercarial chamberA. Penetration glands arranged in two groups on each side of body
1. Groups of 5 and 3 on each side —. C. yacalicola sp. n.2. Groups of 6 and 4 on each side C. raiacauda3. Groups of 8 and 2 on each side ..,. C. steelmani
B. Penetration glands in only one group on each side of body1. Constant number present
a. eleven on each side Phyllodistomum staffordib. four on each side C. coelocercac. six on each side
la. Number of flame cells knownaa. 18 each side C. macrocercabb. 16 each side C. donecercacc. 24 each side Phyllodistomum simile
Ib. Number of flame cells unknowna'. 10 sensory tubercles around oral sucker
aa'. stylet 33—37 p. long Gorgodera amplicavab'. 2 rows of 10 each
bb'. stylet 23-25 ^ long Phyllodistomum solidum — C. conicac'. about 40
cc'. stylet 35 ^ long Gorgoderina attenuata2. Number not constant
a. 3 to 4 pairs on each side C. papillostomab. not arranged in 3 to 4 pairs
la. Excretory system stenostomate C. wabashensis2a. Excretory system not stenostomate C. sphaerocerca
III. Tail without cercarial chamberA. Six penetration glands each side C. latigazica sp. n.B. Four penetration glands each side C. eriensis
Literature Cited
Coil, W. H. 1954. Contributions to the lifecycles of gorgoderid trematodes. Amcr. Midi.Natl. 52: 481-500.
Goodchild, C. G. 1945. Additional observa-
tions on the life history of Gorgodera ampli-cava Looss, 1899. J. Parasit. 31(Suppl.):22-23.
Krull, W. H. 1934. Studies on the life historyof a frog bladder fluke, Gorgodera amplicava
Copyright © 2011, The Helminthological Society of Washington

106 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Looss, 1899. Pap. Mich. Acad. Sc. Arts &Letters 20: 697-710.
Rankin, J. S., Jr. 1939. The life cycle of thefrog bladder fluke, Gorgoderina attenuataStafford, 1902 (Trematoda: Gorgoderidae).Amer. Midi. Natl. 21: 476-488.
Schell, S. C. 1967. The life history of Phtjtto-distomum staffordi Pearse, 1924 (Trematoda:Gorgoderidae Looss, 1901). J. Parasit. 53:569-576.
Thomas, J. D. 1958. Studies on the structure,life history and ecology of the trematodePhtjllodistomum simile Nybelin, 1926 (Gorgo-deridae: Gorgoderinae) from the urinarybladder of brown trout, Salmo trutta L.Proc. Zool. Soc. London 130: 397-435.
Vickers, G. G. 1940. On the anatomy of Cer-caria macrocerca from Sphaerium cornenm.Quart. J. Micr. Sc. 82: 311-326.
On the Morphology and Ultrastructure of the EsophagealRegion of Trichodorus allius Jensen
D. J. RASKI, N. O. JONES, AND D. R. ROGGENUniversity of California, Davis and Laboratoria voor Morfologie en Systematiek, Gent
The search by electron microscopy for plant-pathogenic viruses in nematode vectors beganwith Xiphinema index, the vector of the fan-leaf strain of arabis mosaic virus. No evidenceof the virus was found in X. index but thestudy did yield valuable information on theultrastructure of the nematode vector (Wright,1966; Roggen, Raski and Jones, 1966, 1967).
More recently, the search has been directedto Trichodorus allius, a vector of tobacco rattlevirus, in a similar study of nematode-virus re-lationships. One advantage of this virus is therod-shape of its particles which might be easierto detect in animal tissue than the polyhedralparticles of fanleaf virus.
A study of ultrathin sections of T. allius hasnot been successful in locating virus particlesbut has been fruitful in developing a betterunderstanding of the odontostyle and mecha-nism of feeding and of the ultrastructure ofthe esophagus.
Materials and Methods
Nematodes used in these studies were col-lected originally from soil about the roots ofsugarbeet (Beta vulgaris) and alfalfa (Medicagosativa) near Salinas, California. Populationswere reared at Davis, California, under green-house conditions feeding on the roots of alfalfa,tobacco (Nicotiana tabacum), but principallyon sweetpea (Lathyrus odoratus). No males ofT. allius were seen in any of these populations.
Female and larval nematodes were wet-sievedfrom the soil then hand-picked into tap water.The nematodes then were transferred to 0.9%NaCl and frozen by dry ice in a fume-chamberwith 2% OsO4 in distilled water. After beingallowed to return slowly to room temperaturethey were fumed overnight. This treatment wasfollowed by 2 minutes immersion in 15%DM SO (dimethylsulf oxide) in distilled waterand 20 minutes in 2% OsO4 in distilled water.This procedure was used unless otherwise indi-cated in plate text. The nematodes then werehandled according to Wright and Jones (1965).Embedded worms were cut by diamond kniveson an LKB Ultrotome and examined on anRCA EMU-3G or modified EMU-3E electronmicroscope each having a 50 ^ objective aper-ture and operating mostly at 50 kv, occasionallyat 100 kv.
ObservationsCuticle
The cuticle of the adult female of T. allius(Plate 1, A—D; Fig. 1, A) appears to be com-posed of eight layers as follows:
L1. A triple-layered, thin (0.025 _«,) osmio-philic membrane.
L2. A thick (0.16 /a) layer, uniform in tex-ture. There may be two sub-layers making upL2 because in one section (OsO4 fixative plusPb(OH)2 postsectioning stain for 15 minutes)a fine, outer, whitish band and dark inner band
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 107
were observed. These were not consistently orcertainly set apart in other sections.
Fine cross-striations (cs) (Plate 1, A—C),approximately 3 ^ apart, were present on mostof the body and extended almost to the termi-nus. The striations were formed by an infoldingof L1 and L2 and at most created a slight in-dent in the outer surface of L3.
L3. A layer, varying in thickness, from0.48—1.50 p, with an average of about 0.64 p,.This layer also appeared to have two com-ponents, different in texture and somewhat incolor, but there was no sharp demarcationbetween the two. The outer portion (L3a)comprised 74-82% of the total and was coarser-grained in appearance than the inner portion.In cross-section there was only a vague sug-gestion of structure as concentric lines butwithout regularity. In longitudinal section(Plate I, C) there were many lines runninglongitudinally which appeared to set off sub-layers, 18—20 in number, whitish with darkborders. The inner 14 or 15 were broader, nar-rowing (or doubling) near L2. The inner por-tion (L3b) comprised 18-25% of the total ofL3, was darker in color and of finer grain thanL3a. In longitudinal section there were 11 or12 sublayers similar to those in L3a but darkbands with light borders.
L1. A layer variable in thickness from mini-mal or nonexistent to 0.68 p., spongy or amor-phous in texture.
L"~7. The next three layers were granularwith no apparent structure and about equal inthickness (0.09 p. each). The innermost (L7)had an irregular inner surface projecting inrough, lumpy protuberances.
Ls. Underlying L7 was an irregular and lightgray layer the nature of which is unknown. Itwas intimately connected with the epidermiswhich, on the outer margin, formed an irreg-ular edge with polyp like protuberances inter-mingled with Ls.Oral Aperture
The most anteriad section shows the oralopening to be triradiate (each arm about 0.5—0.7 /i, long). These arms are almost straightbut slightly posteriorly they become irregularwith several short side branches, the distalbranches on two of the arms give an almostforked appearance (Plate II, A ) . Lining theoral opening is the thin, dark osmiophilic layer
(L1) which is continuous with the outermostlayer of the body cuticle.
The opening is set in L2 of the cuticle whichforms a roughly hexagonal shape, each sideabout 0.7-0.9 p. long. The six angles of thehexagon are directed each between two innerlabial papillae (IL) and anteriorly these anglesextend outward joining a similar layer of tissueoutside the papillae. The triradiate oral open-ing also is oriented with each arm directed be-tween two inner labial papillae. Each papillais surrounded by more coarsely granular L3.
About 0.6—0.8 /A posteriad from the anteriorsurface (Plate I, D) is a crease or folding ofthe oral aperture extending parallel with theanterior surface. This fold is about 0.6 ^ longin both directions perpendicular to the longi-tudinal aperture. The osmiophilic layer linesthis fold and is continuous with the lining ofthe oral aperture.Stoma
The first evidence of the stomatal wall (PlateII, B) appears just below the thick anteriorcuticle and is composed of:
L1. The osmiophilic layer forms a very ir-regular outline of the stomatal lumen and isgrossly convoluted or jagged. This lasts only ashort distance and soon becomes more simple,triradiate with a single bulge at the midpointof each side (Plate II, C). Where the lining isconvoluted (Plate II, B) there are some mem-branelike folds inside the lumen of the stoma.
L2. This layer is relatively thick (0.1-0.2 p.),at first uniform in texture, and definitely setoff from the adjacent L3 by a dark line. Latera well-defined membrane appears around L2
(Plate II, C) and soon afterwards tissue ap-pears with the membrane. This tissue probablyis the most anterior projection of pharyngealtissue (ph).
At the same level where the above mem-brane first appears there is also the first evi-dence of the lining of the pharyngeal lumen(pll) inside L2. The innermost lining of thepll seems to be similar to, if not identical with,L1 and continuous with it. As the pll expandsthe L2 diminishes to a thin concentric bandaround it, finally becoming an incomplete band(Plate II, D) and later disappears.
The double-layered L3 surrounds L2. Out-side of L3 is a membrane which broadensslightly to a thin line of epidermal tissue. Sur-
Copyright © 2011, The Helminthological Society of Washington

108 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
rounding the epidermal layer is another whichappears to be Ls and it in turn is surroundedby another layer of epidermis.
L3 retains its double-layered appearance(Plate II, C) through level 1 (Fig. 1, A-B)where the pharyngeal tissue appears betweenit and L2. The pharyngeal tissue continues toexpand through succeeding levels further sep-arating off L3. As this happens L3 becomeshomogeneous in color and evenly granular.Posteriorly L3 disappears (Plate II, E). Thisseems to be the inner limit of the externalcuticle and probably marks the posterior endof the stoma. This is supported by the fact thatmolted cuticles of larvae show only a very shortstomatal cuticular lining which is cast off.
PharynxCoincident with the very first appearance of
the pll there are light colored areas (Plate II,C; level 1, Fig. 1, A—B) which seem to bestrengthening rods or bands (Pr) runninglongitudinally. These may serve as muscle at-tachments and are located centrally near theosmiophilic layer. Similar refractive bands soonappear (Plate II, D) at the ends of the tri-radiate lumen. The 3 inner bands are shorterthan the distal ones and disappear just beforethe stylet tip is seen (Plate II, E). The threedistal bands divide into three each—one at the
tip of the angles of the lumen and one slightlylarger on each side.
The next change in these bands is their dis-appearance in the apical area on all three raysof the lumen accompanied by a lengtheningof the dorsal wall which is gently arched (PlateIV, A) . On the ventral ray of the lumen therefractive bands divide to form two pairs.These refractive longitudinal bands persistsome distance until finally the two pairs onthe ventral ray disappear (Plate II, F) andsoon afterwards the pairs on the two subdorsalrays disappear.
Posteriorly the next development is a smooth,grayish thickening of L1 lining the two sidesof the ventral ray of the lumen (Plate III, G).These thickenings extend posteriorly throughthe folds of the guide ring then disappear. Therest of the pll outside the L1 becomes re-stricted to small narrow areas on both wallsof the ventral ray on each side of the swellingand finally the entire lumen wall becomes auniform, gray texture.
Odontostyle
The odontostyle is first seen (Plate II, E)as a simple circular body about 0.2 ^ across,free in the lumen. It has a uniform gray colorwith a small central darker dot and a fine,slightly darker, outer border. Posteriorly it en-
Plate I. Cuticular layers and markings. A, Longitudinal section, 12,000 X; B, Longitudinal section,38,000 X; C, Cross section, 20,800 X; D, Longitudinal section in stomatal region, (IL) innervations tointerne-labial papillae, 43,700 X.
Plate II. Cross sections of stoma and pharynx including the odontostyle and its extension. A, Nearoral aperture, 14,000 X; B, Stoma, 23,100 X; C, Transition from stoma to pharynx and first evidenceof pharyngeal lumen lining (pll), 19,900 X; D, Transition slightly posterior to D, 18,200 X; E, Pharynxwith first section of odontostyle (Od), 30,400 X; F, Pharynx posterior to E, 20,000 X.
Plate III. Cross sections of stoma and pharynx (cont'd). G, Odontostyle near attachment to dorsalwall, 19,400 X; H, Fusion of odontostyle with dorsal wall and guide ring, 13,400 X; I, Stylet extensionposterior to H, 12,500 X; J, Stylet extension posterior to I, 13,800 X; K, Attachment of odontostyle pro-tractor muscles to stylet extension, 22,000 X; L, Transition of stylet extension with wall of pharyngeallumen, 12,500 X.
Plate IV. Esophageal muscles. A, Cross section at collar of (Ap) amphid, (PPm) perpendicularplvaryngcal muscles, (LPm) longitudinal pharyngeal muscles, 19,800 X; B, Cross section near base ofstylet extension, (OPm) odontostyle protractor muscles, 23,100 X; C, Cross section of esophagus in narrowregion posterior to stylet, (ELm) esophageal lumen muscles, 20,200 X.
Plate V. A-B. Longitudinal sections of amphid (P — amphidial pouch; C = collar; -> indicates basalplates of cilia). A, Near base of amphidial pouch, 9,500 X; B, Junction (S) of cilia and nerve (N), 15,300X; C, Longitudinal section of esophageal gland orifice into lumen of esophagus cut across two rays oflumen (Lu), 30,400 X; D, Longitudinal section between two rays of lumen (Lu), 31,200 X.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 109
cs
Copyright © 2011, The Helminthological Society of Washington

110 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
J
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969111
Uti
-
'
* ' " ' - ' •
Copyright © 2011, The Helminthological Society of Washington

112 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 113
''^m^m," •:>• ' r-. • • !i. V*?*!*!? . '
<**.-.:~ ,; ,-v-- '• f>:*t-\+;.r.
Copyright © 2011, The Helminthological Society of Washington

114 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
larges to about 0.3 ^ across and shows twolayers, a darker band (about 0.05 //, wide) asa concentric ring surrounding the solid innercore with a central black dot (Plate II, F).Later the inner core and the outer band ex-pand and the central dot enlarges, becom-ing more complex and eventually develop-ing an outer membrane surrounding tissuewhich includes endoplasmic reticulum andmitochondria.
The outer portion of the odontostyle reachesa maximum thickness of about 0.1 ^ when thetotal diameter is 0.5 //,. Posteriad this outerportion becomes more narrow as the inter-mediate whitish layer expands to 0.8-1.0 p. indiameter. At the same time the central coreenlarges with a membrane surrounding its tis-sue. When the outer band has disappeared(Plate III, G) the odontostyle consists of acircular, uniformly light, cuticular material sur-rounded by a thin, dark line with a maximumdiameter about 1.1 p, and with an inner tissuecore about 0.3 ^ in diameter. The cuticularlining of the odontostyle is similar in textureto the odontostyle extension and continuouswith it.
Posteriad the odontostyle begins its attach-ment to the dorsal wall and the "guide ring"appears (level 5, Fig. 1, A). The attachmentbegins first toward the left side of the dorsalwall (Plate III, H) and is completed in a shortdistance. Where the odontostyle joins the dor-sal wall (Plate III, H) the pi I folds anteriorlythen again posteriorly, still showing the thick-enings of the L1. In the region of the odonto-style extension posterior to the "guide ring" thelumen of the esophagus (Lu) is small, narrowand immediately ventrad to the extension(Plate III, I, J).
In the odontostyle extension the inner coreof tissue with membrane moves dorsad (PlateIII, I) and finally merges (Plate III, J) withthe nearby tissue (epidermal?) of the pharynxand the odontostyle extension thus forms aU-shape in cross-section with the open end tothe dorsal side. By the time the core of tissueis no longer part of the extension, the latter,in cross-section, is flattened ventrally where itis part of the lumen wall and rounded dorsally.As the extension begins to diminish it becomesirregular in dorsal outline and mottled in tex-ture (Plate III, K). Finally as the last of thewhitish extension tissue disappears it becomes
banded diagonally across the dorsal wall (PlateIII, L). The lumen, as it courses through thenarrow esophagus to the glandular bulb, hasthe familiar osmiophilic membrane lining it
-with simple rounded ends on the three radi-ating arms of the lumen.
MusculatureThe somatic muscles possess two kinds of
elements (myofilaments), fine and large, similarto those in Xiphinema index. These form iden-tifiable areas in the cells where only large onesoccur (near the midline of the cell), or whereboth occur (these areas surround the former).Finally the fine elements seem to be free ofthe larger elements in the outer margins ofeach cell. Neither striae nor any regular re-peating markings were seen in longitudinalsections.
In the esophageal region there appear to besix or seven sets of muscles.
1. LABIAL MUSCLES (LM) (Fig. 1, A). Forwant of a better term these are referred to aslabial because they are the first to appear atthe anterior end. There seem to be four (twoeach subventral and sublateral), quite large,extending from the outer stomatal cuticularlining to the body wall and are almost asbroad as long at their maximum development.They each occur laterad to their respectivedorsal or ventral inner labial papillary inner-vation processes.
It is not certain just where they attach.These are not long muscles and seem to dis-appear before the stylet tip is found in thebuccal cavity. It is possible these represent theanterior extremity of the pharyngeal muscles(PPM).
2. PHARYNGEAL MUSCLES (LPM) LONGITU-DINAL. Four smaller muscles (LPM) appear atabout the same level at which the labial mus-cles disappear. These four are closer in nearthe stomatal region with two each betweenthe subventral and subdorsal inner labial pa-pilla innervations (Plates II, D; IV, A; Fig. 1,A) . The muscles are narrow and longitudinallyparallel with but outside the pharyngeal tissue.The muscles seem to diverge and change loca-tion, one continuing on the left lateral side,the others as a single subdorsal and two sub-ventral in location. The left lateral muscle per-sists longest but not quite to the end of theodontostyle proper.
Copyright © 2011, The Helminthological Society of Washington

L1"3
, lay
ers
of c
utic
leL
m, l
abia
l m
uscl
esP
Pm
, pe
rpen
dicu
lar
phar
ynge
al m
uscl
esP
r, s
tren
gthe
ning
rod
sin
pha
rynx
Epi
, ep
ider
mis
LPm
, lo
ngit
udin
alph
aryn
geal
mus
cles
Sm, s
omat
ic m
uscl
esP
ll, l
inin
g of
pha
-ry
ngea
l lu
men
Od,
odo
ntos
tyle
GR
, gui
de r
ing
OdA
t, at
tach
men
t of
odon
tost
yle
toph
aryn
xO
dEx,
odo
ntos
tyle
exte
nsio
nL
u, lu
men
of
phar
ynx
OPm
, odo
ntos
tyle
prot
ract
or m
uscl
es
A
BFi
gure
1.
Sc
hem
atic
dra
win
g at
he
ad
regi
on a
nd
esop
hagu
s of
T.
al
lius
.
Copyright © 2011, The Helminthological Society of Washington

116 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
3. PHARYNGEAL MUSCLES (PPM) PERPENDIC-ULAR TO LONGITUDINALS (LPM). A set of threemuscles are present in the area near the an-terior tip of the odontostyle when in retractedposition. One is about midventral in positionattaching to the ventral apex of the triradiatestoma (Plate IV, A; Fig. 1, A). The other twoare subdorsal attaching to the two subdorsalapices of the stoma. The muscles extend dis-tally and proceed between somatic muscles tothe body wall, possibly to the innermostlayer of the epidermis. Internally the musclesend at a perpendicular line darkened in colorand adjacent to the basement membrane ofthe epidermal tissue surrounding the pharynx.
4. ODONTOSTYLE PROTRACTORS (OPM). Theseare strongly developed muscles which begin ata level anterior to the base of the odontostyle.Here, there seem to be three, one dorsal andtwo subventral, near the cuticular lining of thepharyngeal lumen. Nearer the end of the odon-tostyle these move to the outer edge of thispharyngeal tissue and appear as four or fivelarge, irregularly shaped muscles. At the begin-ning of the odontostyle extension the musclesdivide into elements (Plate IV, B), 15 or morein number almost lining the entire outer limitof the pharyngeal tissue. Some are dark stain-ing while others are similar to the musclesanterior to this level. These muscles attachventrally, two being slightly subventral leavinga corridor of epidermal tissue directly ventradto that ray of the triradiate lumen. Each ofthese two becomes bifurcate as they approachthe outer edge of the pharyngeal tissue. Wherethe core of the extension emerges, the musclesshift toward the center of the pharynx. Heremitochondria are large and numerous. Muscleattachments to the extension occur where thesclerotized wall begins to reduce in size andthen almost cover the outer dorsal wall of thatorgan (Plate II, K).
5. SOMATIC MUSCULATURE (SM) (Fig. 1, A).These muscles begin at about the level wherethe amphidial pouches enter the body, appear-ing first dorsad and ventrad to the pouches.As the pouches move inward leaving a spacelaterad to them, the muscles appear betweenthe pouches and the outer wall. Compared toXiphinema index, the somatic musculature isnot very high in cross-section.
6. ESOPHAGEAL LUMEN MUSCLES (ELM).Some sections of the narrow part of the esopha-
gus posterior to the odontostyle and also of theenlarged glandular region, show some musclesradiating outward from the angles of thelumen. These appear to attach at the angles ofthe lumen or on the swellings on the arms ofthe lumen (Plate IV, C). Some sections showevidence of attachments of muscles at the endsof the rays of the lumen.
Sensory organs1. INNER LABIAL PAPILLAE. The six inner
papillae when first seen (Plate II, A) are ar-ranged symmetrically about 1 p, from the cen-ter of the oral aperture. The papillae show ascattered arrangement of microtubular ele-ments. Posteriorly these arrange into four cili-ary elements in each papilla (Plate II, B—C).
2. OUTER LABIAL AND CEPHALIC PAPILLAE.Six outer labial and four (two subdorsal andtwo subventral) cephalic papillae are presentwith two or three ciliary elements in each.
3. AMPHIDS. These are quite similar to thosefound in X. index and are formed by invagina-tions of the cuticle. The inner lining is a con-tinuation of the osmiophilic layer. Adjacent tothe osmiophilic layer and surrounding it wascuticular material apparently similar to L3 ofthe outer cuticle but also perhaps some rem-nants of L2.
The sensillar (amphidial) pouch is evenlygranular in texture near the aperture but im-mediately posteriad is provided with some ele-ments. At first the pouch (P) is irregular inoutline then assumes a shape roughly rectangu-lar in outline. The shape then narrows into acircular form with a thick ring of cuticle (C)surrounding it (Plate V, A). In the longitudinalsection the amphid has a goblet-shape with aconcentration of ciliary elements in the narrowcollar (Plate V, A) . Posteriorly the amphidwidens again as numerous ciliary elementswith basal plates (arrows) connect (?) withnerve cells (N) (Plate V, B) . Ciliary struc-tures are similar to those in X. index, dif-fering only in number of elements in individualsensory organs. The cuticular structures of theamphids are molted by larvae and persist asprominent remnants in the shed cuticles.
4. NERVE RING. First seen at the level nearthe posterior end of the odontostyle extensionthe nerve ring forms a U-shaped arc of nerveson the ventral side but not entirely encirclingthe esophagus. In the region of the nerve ring
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 117
Figure 2. Schematic drawings illustrating feeding mechanism of T. allius. a—c, stylet protraction; d—e,stylet retraction; 1, labial muscles (Lm); 2, perpendicular pharyngeal muscles (PPm); 3, longitudinalpharyngeal muscles (LPm); 4, somatic muscles (Sm); 5, odontostyle protractor muscles (OPm).
and posteriorly around the esophagus are manynuclei with dense chromatin arranged aroundthe periphery of and also scattered throughoutthe nucleus.
ESOPHAGEAL GLANDS. There are five nucleivisible in the esophagus of T. allius (Fig. 1,B). One is the large dorsal gland nucleus lo-cated in the dorsal sector of the esophagusabout midway in the posterior bulb or glan-dular region. Two large subventral gland nucleiare more posterior in the glandular region.Slightly anterior to the dorsal gland nucleusare two more subventral nuclei, smaller in sizeand less noticeable.
The dorsal gland extends almost the fulllength of the dorsal sector but has its orificeinto the lumen of the esophagus in the anteriorend of the glandular region. In many specimensthe narrow part of the esophagus between thestylet and the posterior bulb forms a distinctloop or S shape.
The two posterior subventral glands connectwith the lumen of the esophagus in the pos-terior bulbar region not far from the nucleithemselves. However, the orifices of the twosmaller anterior subventral glands were notcertainly established.
The orifices of the dorsal and two largersubventral glands are provided with minute,complex structures which appear to serve ascollection mechanisms. These measure about2.5 n long, 1 fj, in width and height and arelocated next to the lumen and each is betweentwo rays of the lumen. The structure seems tobe a sinus with radiating and branching armsnumbering 20 or more. It is completely sur-rounded by a triple-layered membrane. A sagit-tal section crossing both rays of the lumenshows a kidney- or oval-shaped outline (PlateV, C), becoming a common sinus in the center.Sagittal sections bisecting two rays shows threeor four separated arms of the sinus or 5—10 ormore fingerlike radiating arms becoming fan-shaped and converging to the orifice enteringthe esophageal lumen (Plate V, D).
DiscussionCUTICLE. The report on X. index (Roggen
et al., 1967) concluded that the structure of thecuticle of nematodes is too variable and toofew species have been studied to permit gen-eralizations. It was not possible to homologizethe layers of the cuticle of X. index with thoseof other nematodes.
Copyright © 2011, The Helminthological Society of Washington

118 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Here the same appears to be true and asidefrom the outer osmiophilic layer the cuticularcomponents of T. allius show no certain ho-mology with those of X. index. The secondlayer in T. allius may be the cortical layer butit does not appear to be at all similar to thesecond layer of X. index.
Layer four is interesting in that it varies somuch in thickness and is very spongy in nature.The cuticle of most Trichodorus species isknown to be very loose and tends to fold orwrinkle during movement. In some fixativesit swells, sometimes ballooning out and forwardbeyond the anterior end of the body. The sec-tions seen here indicate most of this changetakes place in L'. This suggests osmotic regula-tion may be active in this layer.
FEEDING PROCESS. The feeding process of T.allius is accomplished by protraction of thestylet to puncture plant cells and to ingest con-tents of the cells through the triradiate lumenof the pharynx and esophagus. The structureof the odontostyle being toothlike with a solidtip (Plate II, E) precludes the ingestion offood through the odontostyle even though itappears to be hollow in sections just prior tothe tip. Anterior to the "guide ring" the styletis located inside the triradiate lumen of theesophagus. Posteriorly the stylet lumen is ven-trad to the extension. The muscle structure ofthe pharynx, as determined by their sections,suggests its function in the protraction and re-traction of the stylet to be as follows (Fig. 2):(a) rest position with all muscles relaxed; (b)by contraction of the somatic muscles, labialmuscles and perpendicular pharyngeal muscles,the stoma is everted with a slight protractionof the stylet; (c) contraction of the odontostyleprotractor muscles results in protraction of thestylet and increased turgor pressure in thepharyngeal wall; (d) upon relaxation of theprotractor muscles, turgor pressure effects sty-let retraction; (e) relaxation of somatic, labial
and perpendicular pharyngeal muscles resultsin reinvagination of the stoma.
The functioning of the pharyngeal lumenappears to be by the classical mechanism foundin most nematodes: opening of the lumen bymuscular action, closing by turgor pressure.
Eversion of the stoma during feeding maypossibly account for the fine folds in the innerstomatal lining or osmiophilic layer (Plate II,B). If eversion does take place the cuticularlining would be stretched and the layers closestto the lumen would be stretched most. A rea-sonable adaptation would be to provide foldsin the cuticle which would disappear on stretch-ing. The folds would have to be most pro-nounced in the innermost layers and less in theouter layers of the stoma wall. Furthermore, itwould be expected that thinner membranescould not withstand as much stress as thethicker ones and should be more folded. Thefolded osmiophilic layer fits this concept.
SummaryObservations are reported here on the ultra-
structure of the cuticle, pharynx, musculature,sensory organs and esophagus of females ofTrichodorus allius. The feeding process is alsodiscussed.
Literature CitedRoggen, D. R., D. J. Raski, and N. O. Jones.
1966. Cilia in nematode sensory organs. Sci-ence 152: 515-516.
. 1967. Further electron microscopicobservations of Xiphinema index. Nemato-logica 13: 1-16.
Wright, K. A. 1965. The histology of theoesophageal region of Xiphinema indexThome and Allen, 1950, as seen with theelectron microscope. Can. J. Zool. 43: 689-700.
, and N. O. Jones. 1965. Some tech-niques for the orientation and embedding ofnematodes for electron microscopy. Nemato-logica 11: 125-130.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 119
Penarchigetes oklensis gen. et sp. n. and Biacetabulum carpiodisp. n. (Cestoidea: Caryophyllaeidae) from Catostomid Fish inNorth America
JOHN S. MACKIEWICZUniversity of Oklahoma Biological Station, and the State University of New York at Albany
During June and July of 1963—1966 approxi-mately 400 catostomid fish (Cypriniformes:Catostomidae), comprising five genera andnine species from southern and eastern Okla-homa, were examined for caryophyllid cestodes.From these collections three new species havebeen described (Calentine and Mackiewicz1966; Mackiewicz 1968b); two other newspecies are described in this paper. Referenceto the new species in the genus BiacetabulumHunter 1927, described below was made in anearlier abstract (Mackiewicz 1964).
Cestodes were removed from freshly killedfish, fixed usually by shaking them vigorouslyin a vial of 5% formalin, stained in Semichon'scarmine or Mayer's paracarmine and mountedin permount (Fisher) or Kleermount (CarolinaBiological). Hematoxalin and eosin sectionswere prepared in the usual manner. Drawingswere made with the aid of a microprojector;measurements are in millimeters unless other-wise indicated.
Comparative material included: Biacetabu-ulm meridianum Hunter 1929, holotype USNMHelm. Coll. No. 51059 (serial longitudinal sec-tions), and 11 of Hunter's original slides ofthis species (his numbers 687.2 to .6; all areserial cross or longitudinal sections) now in theauthor's collection; wholemounts or sections ofArchigetes sieboldi Leuckart 1878, A. brachy-urus Mrazek 1908, A. cryptobothrius Wisniew-ski 1928, A. limnodrili (Yamaguti 1934), andA. iowensis Calentine 1962; and a single whole-mount of Paracaryophyllaeus dubininae Kula-kovskaya 1961.
Since all available examples of B. meridi-anum consisted of serial sections, attempts weremade in August 1966 to recollect this speciesfrom the type locality in North Carolina. Re-grettably, only B. infrequens Hunter 1927 wasfound in two of 14 Moxostoma sp. (total length225—425) and four of five Erimyzon sucetta(Lacepede) (total length 150-225) from theEno River near Hillsboro, North Carolina
(Orange Co.) and University Creek, a tributaryof the Eno.
Order Caryophyllidea Van Beneden(in Olsson 1893)
Family Caryophyllaeidae Leuckart, 1878Penarchigetes gen. n.
(Figs. 1-7)GENERIC DIAGNOSIS: Caryophyllaeidae. Sco-
lex with a pair of median bothria, two pairsof lateral loculi, and a small terminal disc.Cirrus joining uterovaginal canal. Ovary H ordumbbell shaped. Coils of uterus extending toanterior level of cirrus. Preovarian vitellarialateral, sometimes continuous with the post-ovarian vitellaria. External seminal vesicleabsent. Type species: Penarchigetes oklensis.
RemarksIn having a single gonopore, lateral vitellaria,
small terminal disc, and uterus not extendinganteriorly beyond the cirrus in non-gravid in-dividuals, Penarchigetes most closely resemblesthe caryophyllaeid genus Archigetes Leuckart1878. Unlike Archigetes, however, the newgenus lacks all trace of an external seminalvesicle (Figs. 6—7) which in Archigetes is aprominent muscularized structure often ap-proximating the size of the cirrus sac.
Penarchigetes superficially resembles Para-caryophyllaeus Kulakovskaya 1961; however,since the placement of the inner longitudinalmuscles is not described nor the presence orabsence of the external seminal vesicle estab-lished, or the number of gonopores mentioned,adequate comparisons can not be made be-tween the two genera. These important fea-tures could not be discerned with certainty onthe single specimen of Paracaryophyllaeus ob-served. Nevertheless, on the basis of an un-specialized scolex and anterior position of theuterus of Paracaryophyllaeus as well as a greatdifference in the distribution and hosts of thetwo genera—palearctic Cobitidae for Paracary-
Copyright © 2011, The Helminthological Society of Washington

120 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Figures 1-7. Penarchigetcs oklensis gen. et sp. n. from the spotted sucker, unless otherwise specified.1A. Cross section through anterior part of bothria. IB. Cross section through middle part of bothriaillustrating the absence of acetabular musculature. 2. Midsagittal section through gonopore. 3. Immatureindividual. 4. Gravid individual with cirrus everted. 5. Cross section through midpart of body. 6. Fron-tal section through cirrus and external seminal vesicle of Archigetes iowensis Calentine, 1962. 7. Frontalsection through cirrus and vas deferens of P. oklensis illustrating the absence of the external seminalvesicle. Abbreviations: esv, external seminal vesicle; ut, uterus; vd, vas deferens.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 121
ophyllaeus and nearctic Catostomidae for Pen-archigetes, the two genera are regarded asdistinct from each other.
Clearly it is necessary to erect a new genusto receive this form for which the name Pen(pene, L. near) archigetes is proposed. Othergenera in the family Caryophyllaeidae firstdescribed from North American Catostomidae(suckers) are: Glaridacris Cooper 1920, Biace-tabulum Hunter 1927, HijpocanjopliyllaeusHunter 1927, Hunterella Mackiewicz and Mc-Crae 1962, Isoglaridacris Mackiewicz 1965,and Promonobothrium Mackiewicz 1968.
Penarchigetes oklensis sp. n.SPECIFIC DIAGNOSIS (measurements based on
11 gravid and three immature specimens, fivesectioned): Gravid individuals 1.2 (0.9—1.7)long and 0.2 (0.26—0.44) wide at gonopore.Immature individuals from fish 0.9 (0.8—1.0)long. Length 3.3—5.0 times combined lengthof neck and scolex. Scolex 0.34 (0.28-0.39)wide. Neck indistinct. Outer longitudinal mus-cles poorly developed. Inner longitudinal mus-cles consisting of small fascicles, chiefly in adorsal and ventral group that form four largefascicles at base of scolex. Testes number 15(11—21) in two irregular rows beginning 0.29(0.25—0.41) from tip of scolex and extendingto a short distance anterior of cirrus sac. Cirrussac round; cirrus eversible. Preovarian vitellariabeginning 0.33 (0.26-0.41) from tip of scolex,usually continuous with postovarian vitellariaover dorsal part of ovary. Postovarian vitellinefield longer than ovary. Ovary dumbbell-shaped, compact follicular wings 0.10 (0.06—0.15) long. Well developed seminal receptacleabsent. Osmoregulatory canals diffuse, no spe-cific number in midpart of body. Egg withsmooth shell, 56 by 40 ^ (one measured inutero); presence or absence of operculumcould not be definitely established.
The specific name refers to the state otOklahoma.
DEFINITIVE HOST: Spotted sucker, Miny-trema melanops (Raf.) (Cypriniformes, Cato-stomidae). USA, Oklahoma, Cherokee Co.,Northeastern Outing Club Lake, 22 km NEof Tahlequah on Okla. rt. 10. This lake is anox-bow lake of the nearby Illinois River; it isapproximately 10 acres in size, has a meandepth of 1 m and is periodically confluent withthe river.
HABITAT: Intestinal swelling and fore partof intestine; weakly attached between mucosalfolds.
TYPE SPECIMENS: Holotype No. 71262 (Coll.14 July 1965) and Paratypes (4) No. 71263of the USNM Helm. Collection; Paratypes (2)Nos. 1968.7.23.1 and 2 of the British Museum(Natural History) Helm. Collection.
Supplementary material (six slides) consist-ing of immature worms and sections include:USNM Helm. Coll. No. 71263 and BritishMuseum (Natural History) Helm. Coll. Nos.1968.7.23.3 and 4.
Remarks
This species, one of the smallest yet de-scribed, offers some interesting morphologicalcharacteristics. For example, the scolex isexpanded and appears to be intermediatebetween the cuneiform-type of Glaridacriscatostomi Cooper 1920 and the disc-type ofGlaridacris laruei (Lament 1921) or Archigetesioivensis (Figs. 3, 4). In all gravid specimensthe testes contained only a few spermatogonialcells along the testis membrane; this conditionmade it difficult to count the testes, particu-larly the most anterior ones. In immature indi-viduals, or ones with just a few ova in theuterus, the uterus does not extend anterior tothe cirrus sac; in gravid individuals it does(Figs. 2—4). With some vitelline follicles dorsalto the ovary (Fig. 3) Penarchigetes appears toexhibit an intermediate condition between acontinuous and discontinuous vitelline distribu-tion. These vitelline cells have the vacuolatednucleus as recently described by Mackiewicz(1968a).
Of eight M. melanops ranging in total length277 (143-331), only two were infected with13 and 14 cestodes, respectively. Some cestodeswere unquestionably overlooked because oftheir small size. Biacetabulum banghami Mack-iewicz 1968 was the only other helminth as-sociated with P. oklensis. Two Ictiobus bubalus(Raf.) and one I. ct/prinellus (Val.) were notinfected with caryophyllids but one of threeCyprinus carpio L. contained Atractohjtocestushuronensis Anthony 1958 and Khawia iowensisCalentine and Ulmer 1961.
Penarchigetes oklensis is the fourth caryo-phyllaeid to be reported from the monotypicspotted sucker, the others are: Monobothriumulmeri Calentine and Mackiewicz 1966, Bi-
Copyright © 2011, The Helminthological Society of Washington

122 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
14 15Figures 8-15. Biacetabulum carpiodi sp. n. from the river carpsucker. 8. Gravid individual. 9. Three
immature individuals; note that the acetabular sucker is visible even on the smallest specimen (scale thesame as in Fig. 8). 10. Midsagittal sections through the gonopore illustrating an expanding cirrus. 11.Midsagittal section through the scolex showing the basal membrane of the acetabular sucker. 12. Scolexwith the acetabular sucker contracted. 13. Outline of an operculate egg. 14. Cross section through mid-part of body. 15. Detail of the posterior part of body, ventral view.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 123
acetabulum banghami Mackiewicz 1968, Pro-monobothrium minytremi Mackiewicz 1968(Mackiewicz 1968b.) Two others, still understudy appear to be Biacetabulum infrequensHunter 1927 and Glaridacris sp.
Biacetabulum carpiodi sp. n.(Figs. 8-15)
SPECIFIC DIAGNOSIS (measurements based on23 gravid and 11 immature individuals; 11sectioned): Gravid individuals 5.5 (4—7.5)long, 0.47 (0.32-0.70) wide at gonopore. Im-mature cestodes from fish 3.1 (0.9—5) long.Length 3.9 to 7.2 times combined length ofneck and scolex. Scolex with a pair of well-developed acetabular suckers; no accessoryloculi. Neck distinct 0.31 (0.24-0.46) wide.Outer longitudinal muscles poorly developed.Inner longitudinal muscles consisting of largefascicles irregularly arranged about the corticalparenchyma. Testes begin 1.7 (1.2—2.2) fromtip of scolex and extend posteriorly to thecirrus sac. Testes number 69 (59—84) andaverage 130 (90-210) p in diameter (5from each of 5 worms). Vas deferens extendsanteriorly between two rows of testes. Cirrussac oval; male gonoduct joins uterovaginalcanal very close to ventral surface forming anatrium with a large gonopore 50-100 p, indiameter. Preovarian vitellaria in median aswell as lateral position, extending 1.2 (0.8—1.4) from tip of scolex to ovary or sometimescontinuous with postovarian vitellaria. Post-ovarian vitelline field longer than ovary. Ovaryshaped like letters H or M; compact follicularwings 0.3 (0.2-0.5) long. Seminal receptaclepresent. Fourteen osmoregulatory canals atmid-region of body. Eggs operculate, shellsmooth, 53 (51-56) ^ long by 32 (28-34) ^wide (5 measured in utero); undevelopedwhen shed.
The specific name refers to the host.DEFINITIVE HOSTS: River carpsucker, Car-
piodes carpio (Raf.) (Cypriniformes: Cato-stomidae). IOWA: Des Moines River (coll. D.Williams). OKLAHOMA: Marshall Co., LakeTexoma; Payne Co., Lake Carl Blackwell (coll.R. D. Spall). TENNESSEE: Lake Co., MississippiRiver at Powell's Ferry opposite Caruthersville,Missouri, New Madrid Co. TEXAS: GraysonCo., Red River below Denison Dam of LakeTexoma (Type locality). Quillback carpsucker,
C. cypriniis (LeSueur). TENNESSEE: AndersonCo., Oak Ridge, Clinch River at the RullrunSteam Plant. Carpiodes sp. IOWA: Iowa River,South Fork Iowa River (coll. R. Calentine).
HABITAT: Usually in the intestinal swelling"stomach," occasionally in the anterior part ofthe intestine. Securely attached with scolexcompletely imbedded in the mucosa.
TYPE SPECIMENS: Holotype No. 71264 (coll.10 July 1965) and Paratypes (5) No. 71265 ofthe USNM Helm. Collection; Paratypes (2)Nos. 1968.7.23.5 and 6 of the British Museum(Natural History) Helm. Collection.
Supplementary material (8 slides) consist-ing of immature worms and sections include:USNM Helm. Coll. No. 71265 and BritishMuseum (Natural History) Helm. Coll. Nos.1968.7.23.7 to 10.
Remarks
There are several morphological featureswhich distinguish this species from all others inthe genus. The pre- and postovarian vitellarianfields are sometimes continuous dorsally overthe ovary (Fig. 15) similar to that describedfor Penarchigetes. The testes extend in tworows on each side of the vas deferens (Figs.8, 15). And lastly, the cirrus intercepts theuterovaginal canal very close to the ventralsurface and with it forms a common atriumwith a large, single gonopore (Fig. 10). Be-cause of this large gonopore, an expandedcirrus and female gonopore may give the ap-pearance of two gonopores. By its small sizeand reduced number of testes, B. carpiodimost closely resembles B. meridianum and B.macrocephalum McCrae 1962 (Table I ) . How-ever, both of these species have few post-ovarian vitellaria and those present do notreach the ovary as in B. carpiodi. In additionto the gross morphological features mentionedabove, B. carpiodi also lacks the well-developedouter longitudinal muscles of B. meridianum.
Incidence and worm burden varied with hostand locality. Five of 70 river carpsuckers (totallength 232-475) from Lake Texoma, Okla-homa, contained 1—3 worms per fish whereasfour of 16 fish (total length 232-414) fromthe Red River, just below Denison Dam ofLake Texoma contained 3—37 parasites. Fromthe Clinch River, Oak Ridge, Tennessee, fiveof 38 quillback carpsuckers (total length 212—375) contained 1—2 parasites. In all cases,
Copyright © 2011, The Helminthological Society of Washington

124 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
which represented 91 cestodes, there were noconcurrent infections with other caryophyllidsalthough Spartoides wardi Hunter 1929 andanother species of undetermined generic status(from Tenn.) occurred in the same host. Fur-thermore, this species appeared to exhibit amarked host-specificity for Carpiodes sp. notbeing found in approximately 275 other cato-stomid fish representing six genera and 11species.
The only other Biacetabulum reported fromcarpsuckers is that of B. meridianum from 201C. carpio from Lake Texoma (Timmons, 1951;Self and Timmons, 1955); most infected fishwere collected in May. Specimens of theseBiacetabulum were lost in the mail before theycould be re-examined. Except for having testesnumbering 45-53 (N = 6) . Self and Tim-mons reported that all other characters closelycomplied with Hunter's description of B.meridianum. In the absence of the originalslides it is difficult to appraise the validity ofthis record; however, all Biacetabulum fromLake Texoma Carpiodes were first called B.meridianum until critical study indicated thata new species was involved.
Self and Timmons (1955) also reported G.confusa from C. carpio; specimens that couldbe re-examined were redetermined as Sparti-odes wardi Hunter 1929.
Discussion
Both of these species offer variations in cer-tain generic characters. For example, the posi-tion of the uterus with respect to the cirrus sachas long been an important generic criterionin the order Caryophyllidea. In some genera,e.g. Biacetabulum, the uterus clearly twistsand forms loops anterior to the cirrus sac withthe uterine primordia assuming this positionbefore gametogenesis commences (Fig. 9). Inothers, such as Penarchigetes, Archigetes, orGlaridacris (especially G. laruei), in which theuterus normally reaches the anterior level ofthe cirrus sac, its position may vary with thestate of maturity of the cestode. That ova canextend the uterus beyond its normal positionhas recently been forcefully illustrated by Ken-nedy (1965) in the genus Archigetes. Whilethis uterus-cirrus sac relationship may be acontinuously variable character, whatever itsnature it should be judged in immature and
maturing individuals before ova have greatlydistended the uterus.
Whether or not the pre- and postovarianvitelline fields are continuous may be also animportant systematic character although atwhat level is not clear at present. In somegenera, for example in Archigetes (except forA. sieboldi), Atractohjtocestus Anthony 1958(? = Markevitschia Kulakovskaya 1965), Brev-iscolex Kulakovskaya 1962, Capingens Hunter1927, or Wenyonia Woodland 1923, all of thevitellaria clearly appear as one continuousunit. In others, such as Caryophullaeus Gmelin1790, Glaridacris, and Isoglaridacris, to men-tion a few, the vitellaria are definitely confinedto separate pre- and postovarian regions. Pen-archigetes and B. carpiodi as well as Caryo-phyllaeides fennica (Schneider 1902) andKhawia baltica Szidat 1942 appear to exhibitan intermediate condition. In the absence of adistinct continuity it would appear that thisintermediate condition is a variation of the dis-continuous distribution. More precise detailsof the vitelline duct system might aid in theinterpretation of this character.
The genus Biacetabulum has been subjectto various interpretations since described byHunter (1927). Szidat (1937) was the first toconsider some palearctic Archigetes as conge-neric with Biacetabulum. Calentine (1965),reviewing the complex Archigetes-Biacetabu-lum question, clearly demonstrated the validityof the two genera on the basis of life-historvand morphological characters. A further analy-sis of this same problem by Kennedy (1965)resulted in the synonymy of all palearcticBiacetabulum under the genus Archigetes. Ex-cept for the Australian species B. tandaniJohnston and Muirhead 1950, incompletely de-scribed from a single specimen and thereforeof questionable status (it has been omitted inTable I), all other species (six) of Biacetabu-lum are from the nearctic.
All of these species, including B. carpiodi,can be arranged into distinct groups on thebasis of scolex morphology and preovarianvitellaria distribution (Table I) . There can belittle doubt that Group I constitutes a goodspecies group, although some of the species,B. infrequens for example, may represent aspecies complex. The relationship of GroupsII and III to each other and to Group I is
Copyright © 2011, The Helminthological Society of Washington
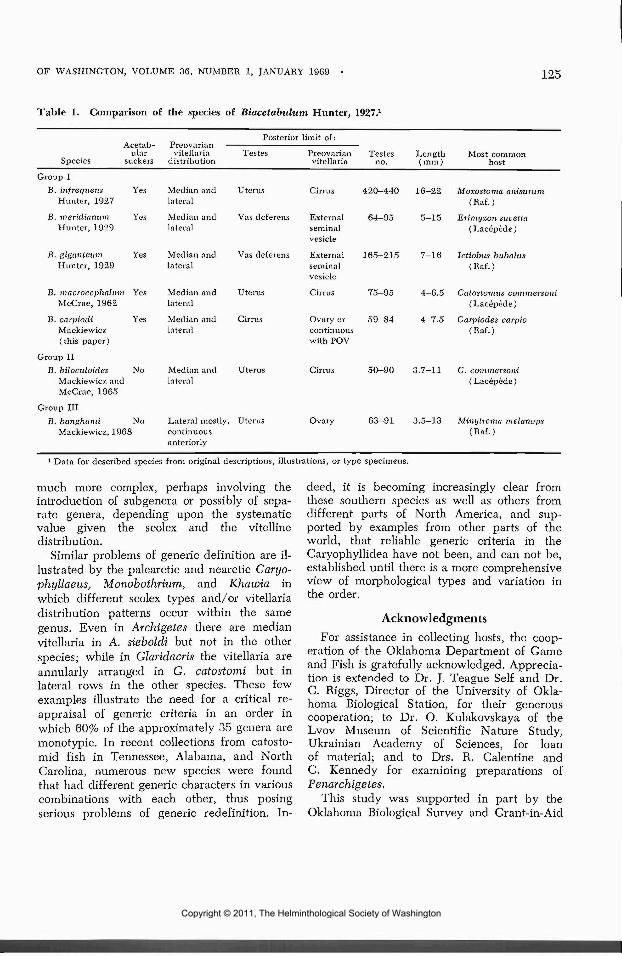
OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 125
Table 1. Comparison of the species of Biacetabulum Hunter, 1927.1
Acetab-ular
Species suckers
Group IB. infrequens Yes
Hunter, 1927
B. meridianum YesHunter, 1929
B. giganteum YesHunter, 1929
B. macrocephalum YesMcCrae, 1962
B. carpiodi YesMackiewicz(this paper)
Group IIB. biloculoides No
Mackiewicz andMcCrae, 1965
Group IIIB. banghami No
Mackiewicz, 1968
vitellariadistribution
Median andlateral
Median andlateral
Median andlateral
Median andlateral
Median andlateral
Median andlateral
Lateral mostly,continuousanteriorly
Posterior limit of:
Testes Preovarian Testesvitellaria no.
Uterus Cirrus 420-440
Vas defereiis External 64-95seminalvesicle
Vas dcferens External 165-215seminalvesicle
Uterus Cirrus 75-95
Cirrus Ovary or 59—84continuouswith POV
Uterus Cirrus 50-90
Uterus Ovary 63-91
Length Most common( mm ) host
16—22 Moxostoma anisurum(Raf . )
5—15 Erirnyzon sucetta( Lacepede )
7-16 Ictiobus bubalus(Raf.)
4—6.5 Catostomits commersoni( Lacepede )
4—7.5 Carpiodes carpio(Raf.)
3.7—11 C. commersoni(Lacepede)
3.5—13 Minytrema melanopft(Raf . )
1 Data for described species from original descriptions, illustrations, or type specimens.
much more complex, perhaps involving theintroduction of subgenera or possibly of sepa-rate genera, depending upon the systematicvalue given the scolex and the vitellinedistribution.
Similar problems of generic definition are il-lustrated by the palearctic and nearctic Caryo-phyllaeus, Monobothrium, and Khawia inwhich different scolex types and/or vitellariadistribution patterns occur within the samegenus. Even in Archigetes there are medianvitellaria in A. sieboldi but not in the otherspecies; while in Glaridacris the vitellaria areannularly arranged in G. catostomi but inlateral rows in the other species. These fewexamples illustrate the need for a critical re-appraisal of generic criteria in an order inwhich 60% of the approximately 35 genera aremonotypic. In recent collections from catosto-mid fish in Tennessee, Alabama, and NorthCarolina, numerous new species were foundthat had different generic characters in variouscombinations with each other, thus posingserious problems of generic redefinition. In-
deed, it is becoming increasingly clear fromthese southern species as well as others fromdifferent parts of North America, and sup-ported by examples from other parts of theworld, that reliable generic criteria in theCaryophyllidea have not been, and can not be,established until there is a more comprehensiveview of morphological types and variation inthe order.
AcknowledgmentsFor assistance in collecting hosts, the coop-
eration of the Oklahoma Department of Gameand Fish is gratefully acknowledged. Apprecia-tion is extended to Dr. J. Teague Self and Dr.C. Riggs, Director of the University of Okla-homa Biological Station, for their generouscooperation; to Dr. O. Kulakovskaya of theLvov Museum of Scientific Nature Study,Ukrainian Academy of Sciences, for loanof material; and to Drs. R. Calentine andC. Kennedy for examining preparations ofPenarchigetes,
This study was supported in part by theOklahoma Biological Survey and Grant-in-Aid
Copyright © 2011, The Helminthological Society of Washington

126 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
No. 20-0160 B from the Research Foundationof the State University of New York.
Literature CitedCalentine, R. L. 1965. The biology and tax-
onomy of Biacetahulum (Cestoda: Caryo-phyllaeidae). J. Parasit. 51: 243-248.
, and J. S. Mackiewicz. 1966. Mono-bothrium ulmeri n. sp. (Cestoda: Caryophyl-laeidae) from North American Catostomidae.Tr. Am. Micr. Soc. 85: 516-520.
Hunter, G. W., III. 1927. Notes on theCaryophyllaeidae of North America. J. Para-sit. 14: 16-26.
Kennedy, C. R. 1965. Taxonomic studies onArchigetes Leuckart, 1878 (Cestoda: Caryo-phyllaeidae). Parasitology 55: 439-451.
Mackiewicz, J. S. 1964. Variations and host-parasite relationships of caryophyllaeids (Ces-todea) from fish of Lake Texoma, MarshallCounty, Oklahoma. J. Parasit. 50 (3, sec. 2 ) :31 (abstract).
. 1968a. Vitellogenesis and eggshell for-mation in Caryophyllaeuf, laticeps (Pallas)and Canjophyllaeides Fennica (Schneider)(Cestodea: Caryophyllaeidea). Z. f. Para-sitenk. 30: 18-32.
. 1968b. Two new caryophyllaeid ces-todes from the spotted sucker, Minytremanielanops (Raf . ) (Catostomidae). J. Parasit.54: 808-813.
Self, J. T., and H. F. Timmons. 1955. Theparasites of the river carpsucker (Carpiodescarpio Raf.) in Lake Texoma. Tr. Am.Micr. Soc. 74: 350-352.
Szidat, L. 1937. Archigetes R. Leuckart, 1878,die progenetische Larve einer fiir Europaneuen Caryophyllaeiden-Gattung Biacetabu-lum Hunter, 1927. Zool. Anz. 119: 166-172.
Timmons, H. F. 1951. A study of the hel-minth parasites of the river carpsucker(Carpiodes carpio) in Lake Texoma. Master'sthesis, University of Oklahoma, 49 p.
The Chromosomes of Hunterella nodulosa Mackiewicz andMcCrae, 1962 (Cestoidea: Caryophyllidea)
JOHN S. MACKiEwicz1 AND ARTHUR W. JONESDepartment of Zoology and Entomology, University of Tennessee, Knoxville, Tennessee
Cytological studies on tapeworms havebeen almost entirely confined to Cyclophyllid-ean cestodes (for reviews see Jones 1945,Walton 1959). Exceptions have been thestudies of four species of the Pseuclophyllideaby Wolcott (1959), and Wikgren and Gus-tafsson (1965), and of Archigetes sp. (orderCaryophyllidea) by Motomura (1929). Wherecomparison between species in the same genuswas made, especially of Hymenolepis by Jones(1945) and Diphyllobothrium by Wikgren andGustafsson (1965), the chromosome numbersand/or karyotypes showed little variation andtherefore could be used little as an aid in thetaxonomic analyses of the polyzoic cestodesstudied.
From the single investigation of Motomura(1929), who was primarily interested in theembryology rather than the cytology of Archi-
1 On academic leave from State University of New Yorkat Albany, New York.
getes, it is impossible to judge the worth ofsimilar cytotaxonomic studies in the monozoictapeworms of the order Caryophyllidea. Onepurpose of the present study, one of a pro-jected series, is to learn if cytological informa-tion on the Caryophyllidea will aid in thepractical problems of taxonomy at the family,genus and species levels; another is to learn ifsuch studies can help in appraising the rela-tionship of these unusual tapeworms to themore common strobilate groups, from whichthey differ greatly in morphology and generalbiology.
Hunterella nodulosa Mackiewicz and Mc-Crae, 1962 (Cestoidea: Caryophyllidea) of thefamily Caryophyllaeidae was chosen for de-tailed cytological studies for two reasons. First,in a preliminary survey of 17 species of caryo-phyllids studied by the squashing technique,squashes of H. nodulosa had large numbers ofgood metaphase plates with relatively large,
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 127
well stained chromosomes in the testes andvitellaria. Second, H. nodulosa is easily identi-fied, widely distributed in North America, andrelatively easy to obtain, occurring as a com-mon intestinal parasite of the white sucker,Catostomus commersoni (Lacepede). It isamong the best known of North AmericanCaryophyllidea with recent studies on syste-matics, morphology, and distribution (Mack-iewicz, I960; McCrae, I960; Mackiewicz andMcCrae, 1962) periodicity (Calentine andFredrickson, 1965) and larval development(Calentine, 1965).
Materials and Methods
Parasites were obtained on 27 November1967 from the white sucker, Catostomus com-mersoni (Lacepede) (Cypriniformes; Cato-stomidae) from Jacks Creek, a tributary of theSmith River (Roanoke drainage), Patrick Co.,Virginia and from the same host from the LittleRiver, Walland, Blount Co., Tennessee on 29March 1968. Living cestodes were droppedinto a vial of Carnoy's fixative (6:1:1, ab-solute ethyl alcohol, chloroform, glacial aceticacid) that was placed in ice water and laterstored in a refrigerator until squashes wereprepared in May, 1968. No differences instaining quality were detected in materialstored for different periods. Carnoy fixationgave good spreads and preservation of thechromosomes, unlike the results of Wikgren(1964) who worked with plerocercoids of D.latum.
Squashes were prepared using the leuco-basic fuchsin (Feulgen) stain prepared ac-cording to Darlington and La Cour (1960).Cestodes were removed from the vial of fixa-tive, hydrated with successive changes (2 hrseach) of 95%, 50% (ethyl alcohol), and tapwater. The whole worm was hydrolyzed in 1N HC1 at 55 C to 60 C for 9-11 min (10 minwas later found to be a good average time)and removed to the Feulgen stain for one hr.Usually the testes and ovary began to show astaining reaction within the first minute. Whenthese organs became deeply stained, (usuallywithin one-half hour), the whole worm wasplaced in 45% acetic acicl for 5-10 min inorder to soften and swell the tissues. A smallpart of the body, approximately one-half mmlong, containing testes and vitellaria was placedon a slide and teased apart in a drop of 45%
acetic acid. A large proportion of the testesand vitelline follicles were teased out and,along with the remaining tissues, dissectedinto small fragments; only large pieces of theintegument were discarded. A 22 mm squarecoverslip (No. 1 or 0) was applied and, afterexcess acetic acid had evaporated, the prepara-tion was squashed by placing the slide withinone fold of paper towel and applying with thethumb the full weight of the body. The pres-ence of Newton rings under the coverslip in-dicated a satisfactory squash. The preparationwas made permanent by using the dry icemethod of Conger and Fairchild (1953), de-hydrating in 95% or 100% ethyl alcohol for 1min and mounting with diaphane.
Dividing cells were most common in thetestes but excellent mitotic figures also oc-curred in the vitelline follicles, particularly incestodes in which spermatogenesis was just be-ginning. Mitoses were rare in the parenchymalcells.
Eighteen squash preparations representinga large number of testes and vitelline folliclesfrom eight cestodes were studied. All illustra-tions and most of the observations were madeon squashes from immature worms collected inVirginia; the Tennessee collections consistedof mostly gravid worms and were used to sup-plement and verify the other observations.Camera lucida drawings of 16 metaphase platesstudied under oil immersion (900 and 1,800powers) were prepared at an enlargement of2,600 X. From 11 of these drawings of meta-phase plates (three from testes and eight fromvitelline cells of two worms) an idiogram wasconstructed by measuring the chromosomes tothe nearest 0.5 mm and expressing each chro-mosome or arm as a percent of the total chro-mosome length in the complement. In this waythere was an adjustment for the differences inchromosomes due to squashing or other factors.In those cases where either the centromere po-sition could not be determined or the chromo-some was folded, no measurements were at-tempted; the sample size for each chromosomemeasured therefore varied. The position of thecentromere of the small rods could not be ac-curately determined but from indirect evidence,as discussed under "observations," these chro-mosomes appeared to be acrocentric and are soindicated on the idiogram. Karyotype analysiswas based on the above eleven plates plus five
Copyright © 2011, The Helminthological Society of Washington

128 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Figures 1-5. Chromosomes of Hunterella nodulosa. Feulgen stained squashes. 1, 2. Metaphase platesfrom vitelline cells. 3. Diakinesis. Note that more than one chiasma is readily visible on most bivalents.4. Metaphase I, showing seven bivalents. 5. Metaphase II, Anaphase II, both dyads and monads arevisible.
others, only parts of which were suitable forpurposes of an idiogram, that included twofrom testes and three from vitelline cells ofthree worms (two from Virginia, one fromTennessee).
The photographs were made with a ZeissPhotomicroscope using phase contrast optics,including an apochromatic oil immersion lensand a green filter.
ObservationsHunterella nodulosa has 14 chromosomes,
diploid (Figs. 1, 2, 7, 8). These include threepairs of metacentrics, ("V's", two long and oneshort pair), three pairs of short acrocentrics("rods"), and one pair of long submetacentrics
("J's") (Figs. 6, 7). At mitotic metaphase inthe spermatogonia and vitelline cells the long-est chromosomes are 6—8 ^, the shortest 2.5-3^ long. As shown by the idiogram (Fig. 6)there are four large "V's," two small "V's," twolarge "J's," and three pairs of small rods. Thelargest chromosome represents approximately12% of the total chromosome complement(TCL). No satellites or fragments were ob-served. The location of the centromere of themitotic metaphase chromosomes could be de-termined for all but the six rods, but fromobservations of metaphase I of spermatogenesisit appears that the centromeres of these rodsare terminal or nearly so.
Meiosis was studied only in spermatogenesis
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969
2N = 14
HUuu
<f
Figures 6-8. Chromosomes of Hunterella nodulosa (cont'd). 6. Idiogram; TCL, total chromosomelength. 7. Karyotype from a single vitelline cell nucleus. 8. Two metaphase plates from vitelline cells.
since no meiotic activity was observed in intra-uterine eggs. Spermatogenesis proceeds asusual with normal sperm being formed aftertwo meiotic divisions. Two meiotic or matura-tion divisions have also been reported for Tri-anophorus lucii (Mull., 1776) (Pseudophyl-lidea) by Rybicka (1962). At diakinesis struc-ture of the chiasmata was readily observable inmost bivalents (Fig. 3). Chiasma frequencywas not worked out but it appears to be 5.0or 6.0 per bivalent, at least in the case of thelongest chromosomes. No attempt was made toascertain the number of primary spermatocytesresulting from a single spermatogonium, detailsbest studied in sections. A single sperm couldeasily be seen adjacent to or wrapped aroundthe ovum in intrauterine eggs.
DiscussionMotomura (1929) was the first to report the
chromosome number of a caryophyllidean;however, there appears to be some questionregarding the identity of the cestode studied.
Motomura designated it as Archigetes appendic-ulatus Ratzel. Unfortunately there have beentwo cestodes with this same binomial, A. ap-pendiculatus Ratzel 1868 and A. appendicula-tus Mrazek, 1897. It has long been recognized,however, that while Ratzel's species was infact the larval stage of Caryophyllaeus laticepsLeuckart, A. appendiculatus Mrazek repre-sented a distinct species (Nybelin, 1922).Since Mrazek's name has been in more com-mon usage with the binomial it is difficult tounderstand why Motomura did not use it. Thecomplex case involving these two binomialshas been thoroughly discussed by Wisniewski(1930). Kennedy (1965b) in his recent re-vision of Archigetes does not list Ratzel's spe-cies (since it refers to Canjophyllaeus); heplaces A. appendiculatus Mrazek in synonymyunder A. sieboldi Leuckart, 1878. A furthercomplicating factor has been the discovery ofanother species of Archigetes, A. limnodridi(Yamaguti, 1934), in Limnodrilus sp. and acobitid fish in Japan (Yarnaguti, 1934; Ken-
Copyright © 2011, The Helminthological Society of Washington

130 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
nedy, 1965a). Motomura was most certainlydealing with Archigetes, because gravid speci-mens came from aquatic oligochetes (Limnod-rilus and Tubifex) yet he did not further de-scribe or illustrate his species. It is thereforeimpossible to know whether he had A. sieboldi(which is not reported from Japan), A. lim-nodrili (originally described from that coun-try), or another, perhaps undescribed, speciesof Archigetes.
Adequate comparisons between the presentwork and that of Motomura (1929) are notpossible since cytological observations were buta minor part of his study. Motomura observeda diploid number of 18 and that the chromo-somes were of unequal size. From his Figure 2of the first maturation division in the ovum, wemeasure the largest chromosome to be 3.3 p.long. H. nodulosa, on the other hand, has adiploid number of 14 and chromosomes ofunequal and larger size (Fig. 8). Accordingto Walton (1959) the only other cestode with14 diploid chromosomes is Liga brasiliensisRansom, 1909 [= Liga punctala (Weinl. 1856)Weinl. 1857 of Yamaguti (1959) and L.brasiliensis Parona, 1901 of Wardle and Mc-Leod (1952)] a cyclophyllidean of birds(Jones, 1945) but distantly related tocaryophyllideans.
More closely related are the Pseudophyllidea,an order under which caryophyllideans haveoften been placed as a family (Hunter, 1930;Joyeux and Baer, 1961). Working with colchi-cine-treated pleroceroids Wikgren and Gus-tafsson (1965) observed that Diphyllobothriumlatum, D. osmeri and D. dendriticum have dip-loid numbers of 18 and that the karyotypes ofsomatic cells of all species are apparently iden-tical. The largest chromosome of D. latum is3.5 //, long with seven pairs of metacentrics andtwo of submetacentrics making up the comple-ment. In contrast, the longest chromosomes ofH. nodulosa are 8 /j, long with only three pairsof metacentrics in the complement. On cytolog-ical grounds the relationship of Diphylloboth-rium and Hunterella does not appear to beclose.
Unlike the experience of Jones (1945) orWikgren and Gustafsson (1965) in which littlevariation in chromosome number or karyotypewas found among the species studied, prelimi-nary studies on the cytology of cestodes of theorder Caryophyllidea indicate that chromo-
some numbers may have systematic value. It istoo early to judge if karyotypes show similarsignificance. Whether differences in chromo-some numbers between Archigetes sp. and H.nodulosa have systematic value can not be de-termined until there are data from more spe-cies. Studies now in progress indicate thatthere is more variation in the chromosome num-bers of caryophyllidean species than any othergroup of cestodes yet studied, that the chromo-somes are the largest yet described for anygroup of cestodes, and that the chromosomescan be demonstrated with few technicaldifficulties.
Acknowledgments
Thanks are extended to the Departments ofFish and Game of Virginia and Tennessee forpermission to collect hosts, to Dr. Etnier forassistance in collecting and to Dr. B. Hochmanfor photographic assistance.
This study was supported in part by Grant-In-Aid 20—0244 A from the Research Founda-tion of the State University of New York.
Literature Cited
Calentine, R. L. 1965. Larval development offour caryophyllaeid cestodes. Iowa Acad.Sci. 72: 418-424.
, and L. H. Frederickson. 1965. Perio-dicity of caryophyllaeid cestodes in thewhite sucker, Catostomus commersoni (Lace-pede). Iowa State J. Sci. 39: 243-250.
Conger, A., and L. Fairchild. 1953. A quick-freeze method for making smear slides per-manent. Stain Tech. 28: 281-283.
Darlington, C. D., and L. F. LaCour. 1960.The Handling of Chromosomes. 3rd Ed.George Allen and Unwin Ltd. London. 247 p.
Hunter, G. W., III. 1930. Studies on theCaryophyllaeidae of North America III. Biol.Monog. 11: 1-186.
Jones, A. W. 1945. Studies in cestode cytol-ogy. J. Parasitol. 31: 213-235.
Joyeux, C. E., and J. G. Baer. 1961. Classedes Cestodaires. In: Traite de Zoologie, T.IV fasc. 1: 327-560.
Kennedy, C. R. 1965a. The life-history ofArchigetes limnodrili (Yamaguti) (Cestoda:Caryophyllaeidae) and its development inthe invertebrate host. Parasitology 55: 427-437.
. 1965b. Taxonomic studies on Archi-getes Leuckart, 1878 (Cestoda: Caryophyl-laeidae). Parasit. 55: 439-451.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 131
Mackiewicz, J. S. 1960. Studies on the Caryo-phyllaeidae (Cestoidea) of Catostomus com-mersoni (Lacepede), with emphasis on thosefrom fish near Ithaca, New York (TompkinsCounty), USA. Ph.D. thesis, Cornell Univ.Microfilm No. 61-1439. Univ. Microfilms,Inc., Ann Arbor, Mich.
, and R. McCrae. 1962. Hunterellanodulosa gen. n., sp. n. (Cestoidea: Caryo-phyllaeidae) from Catostomus commersoni(Lacepede) (Pisces: Catostomidae) inNorth America. J. Parasit. 48: 798-806.
McCrae, R. 1960. Studies on the Caryophyl-laeidae (Cestoda) of the white suckerCatostomus commersoni (Lacepede) in north-ern Colorado. Ph.D. thesis, Colorado StateUniv., Microfilm No. 60-6797, Univ. Micro-films, Inc., Ann Arbor, Mich.
Motomura, I. 1929. On the early develop-ment of monozoic cestode, Archigetes appen-diculatus, including the oogenesis and fer-tilization. Annot. Zool. Japan. 12: 109-129.
Nybelin, O. 1922. Anatomish—SystematischeStudien iiber Pseudophyllideen. GoteburgsKungl. Vetensk. Vitterh. Samh. Handl.Fjarde Puljden 26: 1-228.
Rybicka, Krystina. 1962. Observations sur laspermatogenese d'un cestode pseudophylli-
dien Triaenophorus lucii (Miill., 1776). Bull.Soc. Neuchatel. Sci. Nat. 85: 177-181.
Walton, A. C. 1959. Some parasites and theirchromosomes. J. Parasit. 45: 1-20.
Wardle, R. A., and J. A. McLeod. 1952. TheZoology of Tapeworms. Minneapolis. Univ.Minn. Press. 780 p.
Wikgren, B.-J. 1964. Studies on the mitoticactivity in plerocercoids of Diphyllobothriumlatum L. (Cestoda). Comm. Biol. Soc. Sci.Fenn. 27: 1-33.
, and Margareta Gustafsson. 1965.The chromosomes of somatic cells of threeDiphyllobothrium species, with notes on themode of cell division. Acta Acad. Aboensis,Ser. B. 25: 1-12.
Wisniewski, W. 1930. Das genus ArchigetesR. Leuck. Eine Studie zur Anatomic, His-togenese, Systematik und Biologic. Mem. deL'Acad. Pol. des Sci. et des Lettres., Classdes Sci. Math, et Nat. Ser. B. II: 1-160.
Wolcott, G. B. 1959. The chromosomes ofDipht/llobothrium ursi. J. Parasit. 45: 378.
Yamaguti, S. 1934. Studies on the helminthfauna of Japan. Pt. 4. Cestodes of fishes.Jap. J. Zool. 6: 1-112.
. 1959. The Cestodes of Vertebrates.Systema Helminthum, Vol. II. New York,Interscience Pub. 860 p.
A Protandrous Haploporid Cercaria, Probably the Larva ofSaccocoelioides sogandaresi Lumsden, 1963
R. M. CABLE AND HADAR ISSEROFFDepartment of Biological Sciences, Purdue University, Lafayette, Indiana 47907, andDepartment of Biology, Rice University, Houston, Texas 77005
During the summer of 1967, an unusualcercaria was found among larval trematodesdeveloping in Amnicola comalensis collectedfrom a drainage canal near Galveston Bay,Texas. It not only is the first freshwatercercaria reported for the Haploporidae, a pre-dominantly marine family of trematodes, butalso is sexually precocious with a degree ofprotandry apparently unique among larvaltrematodes. Many cercariae have well devel-oped genital primordia and a few azygiid lar-vae even contain eggs on escape from themollusk. The Texas species lacks eggs but thebody is otherwise equivalent to an adult
trematode. It has a large seminal vesicle filledwith sperms but the testis consists of a largelyempty sac when the larva is shed by the snail.That gonad becomes conspicuously developedin the cercarial embryo, produces sperms, andretrogresses before development in the snail iscomplete. In respects other than sexual devel-opment, the cercaria agrees with the only otherreported haploporid larva, a marine species(Cable, 1962). Given a testis like that of thecercarial embryo, the body would have fea-tures of the genus Saccocoelioides which Szidat(1954) erected for freshwater species inArgentina.
Copyright © 2011, The Helminthological Society of Washington

PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Infected snails were collected near thebrackish ponds where Lumsden (1963) foundsailfin mollies, Mollienesia latipinna, infectedwith a new species of Saccocoelioides, S. so-gandaresi. Several dimensions of the haploporicllarva fall within the ranges Lumsden gave forhis species. Although the two differ in someother respects, they probably are stages of thesame species as Sogandares has suggested in apersonal communication. Evidence to that ef-fect will be presented after describing thestages provided by infected snails.
The entire cercaria was drawn to scale frommeasurements of a representative specimenamong spontaneously emerging larvae killed innear-boiling 2% formalin and measured im-mediately under a freely floating cover glass.Details were added freehand from the study ofliving specimens and wholemounts stained withSemichon's carmine and counterstained withindulin. The metacercaria was drawn from aliving specimen in the same manner, and thecercarial embryo by microprojection of awholemount. All measurements are given inmicrons; lengths and widths of certain cercarialstructures were measured parallel to those di-mensions of the entire larva.
Description of StagesCercaria (Figs. 2-4)
Biocellate, distomate; tail slender, unadorned,attached subventrally. Larva strongly photo-positive, swimming smoothly and almost con-tinuously before encysting in the open. Bodylinguiform to pyriform, 410—436 long; maxi-mum width 175—190, at or somewhat posteriorto midlevel. Tail 485-510 long, 41-45 wide atslight swelling near base. Tegument of bodythick, colorless, uneven; with fine spines fromanterior end almost to eyespot level; par-enchyma yellowish, with much brown pigmentposterior to eyespots, first clumped, then dis-persed except for concentration near caudalattachment; eyespots 23 by 20. Oral sucker61—67 long, 70—76 wide; ventral sucker equa-torial, 69—76 in diameter. Mouth subterminal;
pharynx about midway between suckers, 41—58long, 53—62 wide. Prepharynx almost as longas pharynx, esophagus longer; bifurcation nearposterior edge of ventral sucker; ceca short,divergent, inflated. Testis in emerging larva anelongate, largely empty sac superimposed onexcretory bladder; well developed in embryo,smooth at first but becoming slightly indentedas sperms form. External seminal vesicle filledwith sperms, most conspicuous feature of hind-body; beginning as a wide sac near midlevelof that region, then tapering as it extends an-terolaterally and loops clorsomedially at rightof ventral sucker to turn anteriorly and enterhermaphroditic sac. That sac spherical, over-lapping ventral sucker anterolaterally; contain-ing internal seminal vesicle, prostatic complexand metraterm. Ovary small, ovoid, with fewcells; slightly to left, overlapping externalseminal vesicle posterolaterally. From ovary,oviduct extends anterolaterally to left, joinedfirst by Laurer's canal, then by common vitel-line duct before bending medially to joinootype posterolateral to ventral sucker. Uterusshort, curving to right from ootype, aroundventral sucker to cross neck of external seminalvesicle, enter hermaphroditic sac and join maleduct anterior to pars prostatica. Common gono-duct short; genital pore median, near anterioredge of ventral sucker. Seminal receptacleabsent, amphitypy not observed. Vitelline cellslarge, in massive follicles not in two lateralfields but filling parenchyma of hindbody nototherwise occupied and compressing incon-spicuous cystogenous glands against subtegu-mental muscles. Excretory system stenostomate;primary pores at sides of tail, near base. Blad-der elongate sacciform, extending almost toovary; with distinctly thickened wall contain-ing nuclei. From bladder, main tubules extendto sides of oral sucker, then posteriorly to ven-tral sucker where each receives an anterior andposterior secondary tubule. Flame cells num-erous, apparently three groups served by eachsecondary tubule; five or six flame cells pergroup posterior to ventral sucker. Development
Figures 1-5. 1. Young redia drawn freehand to show excretory system of one side. 2. Cercaria, ven-tral view. 3. Same, showing excretory system on one side of body. 4. Lateral view of cercarial embryoshowing precociously developed testis. 5. Metacercaria.
Abbreviations: ES, external seminal vesicle; EV, excretory vesicle; GP, genital pore; HS, hermaphro-ditic sac; IN, intestine; IS, internal seminal vesicle; MG, Mehlis' gland; OT, ootype; OV, ovary; TE,testis; UT, uterus; VI, vitelline follicles.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 133
4
Copyright © 2011, The Helminthological Society of Washington

134 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
completed in digestive gland of snail afterescape from redia.
HOST: Amnicola comalensis Pilsbry and Fer-riss, 1906.
HABITAT AND,LOCALITY: Drainage canal nearGalveston Bay, Texas.
DEPOSITED SPECIMENS: Nos. 71321-71323USNM Helm. Coll.
Redia (Fig. 1)Elongate, without collar or "feet"; gut short
and wide in older rediae. Pharynx of youngredia 32 in diameter. Excretory pores lateral,just posterior to pharyngeal level. Excretoryformula 2[(3) + (3)] = 12 flame cells.
Metacercaria (Fig. 5)Cyst symmetrically ovoid, not flattened on
one side or adhering to surfaces; with indis-tinct outer layer and well defined inner one,fibrous, 8—11 thick when formed but becomingsomewhat thicker with age and appearance ofspace between worm and cyst wall. Cavity ofcyst 260-293 long, 200-230 wide. Eyespotsbegin to dissociate soon after encystment.
DiscussionAlthough the bladder wall of the cercaria is
appreciably thickened and contains nuclei, itdoes not appear to be of the epithelial type.Instead, the nuclei seem to be those of theprimary excretory tubules and retained afterfusion of the tubules to form the bladder, giv-ing it the appearance of being epithelial. Asimilar bladder has been seen in bucephalidcercariae (unpublished studies).
Features which at first made the cercariaseem unlikely to be the larva of Saccocoelioidessogandaresi were the limited body spination,long prepharynx, extent of the vitellaria, largeexternal seminal vesicle, and reduction of thetestis after its conspicuous development in thecercarial embryo. The last feature suggestedthat the adult would be short-lived, grow littleif any in the vertebrate host, and have avail-able only the sperms provided by the cercaria.In contrast, the adult of S. sogandaresi is de-scribed as having a large testis. However,examination of the paratype of that trematodeand several additional specimens provided byDr. R. D. Lumsden showed that worms com-parable to the cercarial body in size containedeggs but not what could be recognized as atestis. That gonad was distinct only in larger
and presumably older specimens; even then, itusually had the appearance of a young testisbefore the various stages of spermatogenesisare evident as cell clusters. The cercaria inquestion would account for that situation byproviding sperms for early egg production be-fore the testis redevelops and becomes func-tional. Larger adults are about one-fourthlonger and considerably wider than the cer-carial body but dimensions of the suckers andpharynx are essentially the same in both. Dur-ing growth in the vertebrate, utilization ofsperms and vitelline cells provided by thecercaria could account for the less extensiveseminal vesicle and vitellaria in the adult.Meanwhile spination limited to the anteriorend of the cercaria could develop over theentire body.
Morphological differences exclude as theadult of the present cercaria two other hap-loporids known to occur in the region. Nonewas found in a variety of fishes which weseined from the canal where infected snailsoccurred, but Mollienesia latipinna, the de-finitive host of Saccocoelioides sogandaresi wasnot included. Lumsden (personal communica-tion) found in a different snail species ahaploporid cercaria which encysted in theopen. He fed metacercariae to M. latipinna butwith negative results which he considered in-conclusive because he had found anotherhaploporid adult, Saccacoelium beauforti, inmullet from the same region. It now seems thathis cercaria may well be that species and thepresent one the larva of S. sogandaresi.
Although haploporids occur mostly in marinefishes, Szidat (1954) reported several speciesin Argentine freshwater hosts. He regarded theparasites as relicts of the ancient Tethys Seawhich extended from the Mediterranean regionto eastern South America and covered thelower watershed of rivers now in that area. Ina similar manner, former inundations of centralUnited States could account for the far inlandoccurrence of a few trematodes in such pre-dominantly marine families as the Micro-phallidae, Lepocreacliidae, Miero.scapbj.idae,Cryptogonimidae, and Didymozoidae. The lifehistory has been elucidated for at least onefreshwater species in each of those familiesexcept the Microscaphiidae and Didymozoidae,and recent unpublished studies have revealedthe cercariae of Microphallus opacus and two
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 135
lepocreadiids, Homalometron sp. and Micro-creadium sp., developing in Amnicola limosaSay in northwestern Indiana.
It seems significant to the zoogeography ofdigenetic trematodes that all known centraland eastern North American freshwater cer-cariae in the above families as well as theHaploporidae develop in species of Amnicola,and brackish-water larvae in species of Hy-drobia. The fact that malacologists disagree asto whether those genera and Truncatella, agenus of marine snails, belong in the samefamily indicates their close relationship. For-merly, all were placed in the Rissoidae whichnow also includes the marine host of the onlyother described haploporid cercaria (Cable,1962). Knowledge of the life histories of Ar-gentine freshwater haploporids might revealsuch a close affinity of their molluscan hosts tomarine gastropods. Manter (1957) pointed outthat their vertebrate hosts are distantly relatedto those of Mediterranean haploporids, but hesuggested that such euryhaline fishes as mugi-lids may have served as ecological "bridges"for trematodes between marine and freshwaterenvironments.
Present knowledge of life histories stronglyindicates that digenetic trematodes as a groupare less specific for their vertebrate than mol-luscan hosts. Although both are essential tothose parasites in extending their range, in do-ing so they would seem as a rule more adapt-able in acquiring new vertebrate than mollus-can hosts. Such adaptability within the marineenvironment is demonstrated by the crypto-gonimid, Siplwdera vinaledwardsii, which para-sitizes shallow-water fishes from Cape Cod toSouth America. Throughout that range, thecercaria develops in snails of a single genuswhereas the parasite changes its vertebrate hostto fishes of different orders. Similarly, adultsin the Bivesiculidae parasitize a variety offishes in world-wide tropical and subtropicalseas, but all of several known cercariae in that
family develop in snails of the genus Cerithium.It thus is evident that discussions of hostspecificity and zoogeography of digenetictrematodes have unduly neglected molluscanhosts, and not entirely for lack of informationconcerning life histories.
SummaryA cercaria developing in Amnicola comalen-
sis from a freshwater canal near Galveston Bay,Texas, is essentially an adult with a tail exceptfor lacking eggs. A conspicuous testis devel-ops in the embryo, fills a large seminal vesiclewith sperms, and retrogresses to a largelyempty sac before the cercaria emerges fromthe snail. Encystment is in the open. Themorphology and ecology of the cercaria indi-cate it to be the larva of Saccocoelioidessogandaresi. The redia, cercaria and meta-cercaria are described. Representation of theHaploporidae and certain other predominantlymarine families by freshwater species is dis-cussed, suggesting that molluscan hosts havebeen neglected as factors in the spread oftrematodes both within and between marineand freshwater environments.
Literature CitedCable, R. M. 1962. A cercaria of the trema-
tode family Haploporidae. J. Parasit. 48:419-422.
Lumsden, R. D. 1963. Saccocoelioides sogan-daresi sp. n., a new haploporid trematodefrom the sailfin molly, Mollienisia latipinnaLe Sneur in Texas. J. Parasit. 49: 281-284.
Manter, H. W. 1957. Host specificity andhost relationships among the digenetic trema-todes of marine fishes. Premier symposiumsur la specificite parasitaire des parasites devertebres. Inst. Zool. Univ. Neuchatel. 185-198.
Szidat, L. 1954. Trematodos nuevos de pecesde aqua dulce de la Republica Argentina yun intento para aclarar su camcter marine.Rev. Inst. Nac. Invest. 3: 1-85.
Copyright © 2011, The Helminthological Society of Washington

136 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Lissorchis heterorchis sp. n. (Trematoda: Lissorchiidae) fromCatostomus macrocheilus Girard in Oregon
B. BRUCE KRYGIER AND RALPH W. MACYDepartment of Biology, Portland State College, Portland, Oregon 97207
The only previous record of a species otLissorchis in the Pacific Northwest appears tobe that of L. attenuatum which was found byBangham and Adams (1954) in Catostomuscatostomus and C. macrocheilus in BritishColumbia. Magath (1917) established thegenus Lissorchis which, with the new speciesdescribed herein, at present includes elevenspecies.
Materials and Methods
Specimens were removed from the smallintestine of the host, fixed in Gilson's fluid atroom temperature, under slight cover glasspressure, stained in Ehrlich's acid hematoxylin,and counterstained with 0.02% fast green in95% alcohol. Drawings were made with the aidof a microprojector and camera lucida. The de-scription is based upon the six largest speci-mens. Measurements are in microns unlessotherwise stated and the average of each isfollowed by the minimum and maximum inparentheses.
Lissorchis heterorchis sp. n.(Figs. 1-3)
DiagnosisBody elongate, anterior and posterior ends
rounded, 3.1 (2.4-4.0) mm long by 0.87(0.65—1.10) mm wide; cuticular spines ex-tending along body margins from anterior endto a short distance posterior to posterior testis,with few spines on ventral surface. Oral suckersubterminal, aspinose, 322 (283—389) long by365 (313—454) wide, acetabulum in anteriorthird of body, 374 (307-460) long by 401(319—513) wide. No prepharynx, esophagusshort, ceca of moderate width, extending tonear posterior end of body. Testes tandem,lobed, posterior to midbody. Anterior testissmaller, 408 (342-472) long by 330 (260-413) wide; posterior testis 605 (460-779)long by 314 (248-389) wide. Cirrus sacoblique, proximal end just posterior to acetabu-lum and distal end at midlevel of acetabulum
and reaching left body margin, 547 (354—708)long by 121 (83-165) wide; seminal vesiclebipartate with anterior part smaller; prostategland cells present. Genital pore marginal atmidlevel of acetabulum; cirrus protrusible,spined (Fig. 2). Ovary multilobed with vari-able number of lobes immediately anterior toanterior testis, 269 (212-342) long by 324(260-401) wide. Vitelline follicles broadlyinterconnected, fields overlapping ceca anduterine arms, extending from midlevel of ace-tabulum to midlevel of posterior testis. Trans-verse vitelline ducts meeting at posterior mar-gin of ovary at vitelline reservoir; common ductthen proceeds anteriorly over ovarian surfacewhere it is joined by short oviduct leading toootype and beginning of uterus. Laurer's canaland seminal receptacle not seen. Uterus pro-ceeds posteriorly along right side almost to endof body then returns on same side to the levelof ovary, then crosses to the left side where itduplicates the same course on the left side ofthe body, terminating in the metraterm whichlies under the cirrus sac, and reaching thecommon genital pore. Mature uterine eggs,brownish yellow, averaging 26 long by 16wide.
TYPE HOST: Catostomus macrocheilus Girard.HABITAT: Small intestine.TYPE LOCALITY: Multnomah Creek, Mult-
nomah County, Oregon.INCIDENCE: Seven specimens found in one
sucker.SPECIMENS DEPOSITED: USNM Helm. Coll.
Holotype No. 71344 Paratype No. 71345.
Key to the species ofLissorchis Magath, 1917
1. Vitelline follicles 7—12 on each side .... 2Vitelline follicles 16 or more on each
side 32. Ovary deeply trilobed, lobes com-
pletely separated; length of bodyposterior to hind testis approxi-mately equal to combined length of
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 137
Figures 1-3. Lissorchis heterorchis sp. n. 1. Holotype, dorsal view. 2. Detail of genital region. 3. a, b.Outline of testes of two paratypes to show variation.
Copyright © 2011, The Helminthological Society of Washington

138 PROCEEDINGS OF THE IIELMINTHOLOGICAL SOCIETY
the three gonads L. simeri (Van L. attenuatiim (Van CleaveCleave and Mueller, 1932) and Mueller, 1932)
Ovary compact, moderately lobed Vitelline fields reaching posterior thirdbut not appearing as three separate of body; testes in posterior half ofstructures; length of body posterior bodyto hind testis less than length of the L. polylobatum (Haderlie, 1950)three gonads
L. mutabile (Cort, 1918) Results and Discussion3. Cuticular spines extending to posterior The new species is relatively large with a
end of body; postacetabular region broad posterior region. It is the only species ofmuch narrowed and triangular 4 the genus with lobed testes. The posterior
Cuticular spines extending no farther testis is much longer than the anterior one andthan the middle of the posterior the ovary is deeply lobed. The cirrus sac istestis: postacetabular region not nar- relatively short and its proximal end does notrowed, posterior end more broadly extend much posterior to the acetabulum. Therounded 5 cirrus is spined.
4. Cirrus armed Following Smith (1968), we consider Tri-L. gullaris Self and Campbell, 1956 ganodistomum to be synonymous with Lis-
Cirrus not armed sorcliis. Simer (1929) erected a new genusL. fairporti Magath, 1917 Alloplagiorchis for A. garricki but this genus
5. Ovary overlapping both testes, testes also is a synonym of Lissorchis.wider than long, body flask-shaped An examination of paratypes of Lissorchis
L. garricki (Simer, 1929) crassicrurum Haderlie and L. polylobatumOvary not overlapping both testes, Haderlie revealed that the vitelline follicles
testes longer than wide, body more are more numerous than shown in the author'selongate 6 drawings; in the former species the vitelline
6. Anterior limits of vitelline fields much fields extend nearly to the inner margins ofposterior to acetabulum the ceca, in the latter species the vitelline
L. hypentelii (Fishthal, 1942) follicles are overlapping and closely packed.Vitelline fields reaching level of ace- Magath (1917) found that the first and
tabulum 7 second intermediate hosts of Lissorchis fair-7. Acetabulum about twice the size of the porti were the snail Helisoma trivolvis and
oral sucker chironomid larvae, respectively. The cercariaL. transluscens (Simer, 1929) possessed both stylet and tail in contrast to
Acetabulum less than twice the size the cercaria of L. mutabile which, accordingof the oral sucker 8 to Wallace (1941), lacked both stylet and
8. Cuticular spines only on the ventral tail. Cercariae of the latter encyst in the corn-body surface and not on body mar- mensal annelid Chaetogaster limnaei in thegins; combined width of ceca equal respiratory chambers of Helisoma trivolvus andto one-third of body width H. campanulata. The tailless cercariae of L.
L. crassicrurum (Haderlie, 1953) mutabile encyst after being eaten by the an-Cuticular spines on body margins, not nelid. Thus, there is a correlation between
confined to ventral surface; ceca the habitat of the second intermediate hostnarrower 9 and presence or absence of a tail on the cer-
9. Testes distinctly lobed; posterior testis cariae. Smith (1968) found that L. mutabileabout one-third longer than anterior also utilizes freshwater limpets and planarianstestes L. heterorchis n. sp. as intermediate hosts.
Testes entire; posterior testis not one- No Plelisoma snails are known to occur inthird longer than anterior testis 10 the area inhabited by the sucker host of
10. Vitelline fields largely confined to an- Lissorchis heterorchis. More probable snailterior half of body; testes not con- hosts are Flumenicola or Oxytrema, commonfined to posterior half of body stream snails of the region.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 139
SummarySeven specimens of a trematode found in
the intestine of a sucker, Catostomus macro-cheilus Girard, collected in Multnomah Creekbelow Multnomah Falls, Multnomah County,Oregon, were determined to be an imdescribedspecies of Llssorchis Magath (1917). The newspecies herein described is characterized es-pecially by lobed testes unequal in size, aspined cirrus, and numerous, broadly joinedvitelline follicles. The genus Triganodistomumis considered to be a synonym of Lissorchis.A key to the species of Lissorchis is presented.
AcknowledgmentsThanks are expressed to Wanetah D. Bell
for technical assistance and to W. W. Beck-lund for loan of paratypes. This project wassupported by National Science FoundationGrant GB-5384.
Literature CitedBangham, R. V., and J. R. Adams. 1954. A
survey of the parasites of freshwater fishesfrom the mainland of British Columbia. J.Fish Res. Bd. Canada 11: 673-708.
Magath, T. B. 1917. The morphology and lifehistory of a new trematode parasite Lis-sorchis fairporti nov. gen., et nov. spec, fromthe buffalo fish, Ictiobus. J. Parasit. 4: 58-69.
Simer, P. H. 1929. Fish trematodes from thelower Tallahatchie River. Amer. Midi. Nat.11: 563-588.
Smith, R. J. 1968. Ancylid snails as first inter-mediate hosts of Lissorchis mutahile comb. n.(Trematoda: Lissorchiidae). 1. Parasit. 54:283-285.
Wallace, H. E. 1941. Life history and embryol-ogy of Triganostomum mutabile (Cort) Lis-sorchiidae, Trematoda. Trans. Am. Micr. Soc.60: 309-326.
Survival of Eggs and Larvae in Cysts of the Soybean CystNematode, Heterodera glycines, Ingested by Swine
GROVER C. SMART, JR., AND H. R. TuoMAS1
In Virginia, fields infested with the soybeancyst nematode, Heterodera glycines Ichinohe,1952, are planted to peanut, Archis hypogaea,in the rotation system. Although peanut is nota host of the nematode, cysts do adhere to thepods and hay when this crop is grown in in-fested soil (Miller, 1960; Smart and Wright,1962). After peanuts are harvested, swine areoften allowed to roam the fenced fields to eatthe waste and cull peanuts and peanut vinehay. On infested land, they undoubtedly ingestnematode cysts. Swine then may be moved tononinfested fields, and the possibility exists ofspreading the nematode to new locationsthrough the fecal droppings.
Triffit (1929) reported that eggs and larvaein cysts and larvae not in cysts of H. rosto-
1 Respectively, Assoeiate Professor of Nematology, De-partment of Entomology and Nematology, University ofFlorida, Gainesville, and Associate Professor of AnimalScience, Virginia Polytechnic Institute Division of Re-search, Holland, Virginia. The research was conducted in1962 while the senior author was located at Holland, Va.
Florida Agricultural Experiment Stations Journal SeriesNo. 3086.
chiensis Wollenweber, 1923 did not survivepassage through the digestive tract of the 6-week-old pig. Goffart (~1934) fed 3,000 cystsof PI. rostochiensis to a rabbit and mixed theexcrement in sterilized soil planted to potato.Upon examining the potato roots, he found onelarva. Ellenby (1944, 1946) reported thateggs in cysts of H. rostochiensis survived pas-sage through the gut of earthworms, andyielded a greater number of larvae in larvalemergence tests than cysts which did not passthrough earthworms.
Girard (1887) reported that cysts of H.schachtii Schmidt, 1871, which were not dam-aged by the teeth of sheep eating infectedsugarbeets, survived passage through the di-gestive tract of the sheep. He observed larvaeemerging from such cysts and was able to ob-tain reproduction by adding infested sheepexcrement to sand taken from a quarry andseeding sugarbeet. Chatin (1890) was unableto recover viable H. schachtii from sheep fedinfected sugarbeet, some of which were arti-
Copyright © 2011, The Helminthological Society of Washington

140 PROCEEDINGS OF THE HELM1NTHOLOGICAL SOCIETY
ficially infested with large numbers of larvae,males, and females. He did recover eggs andlarvae of parasites of sheep and believed thatearlier workers had misidentified those as H.schachtii. Willot's (1890) results were iden-tical to those of Chatin.
Kofoid and White (1919) reported that eggsof a nematode were recovered from the fecesof military troops. Sandground (1923) showedthat the eggs undoubtedly were those of aroot-knot nematode, Ateloidogyne, and con-ducted experiments showing that root-knotnematode eggs passed through the human di-gestive system and stated, "They were slightlybrown in color and were in all stages of de-velopment, with the metameres so sharplydefined as to indicate their viability." Hefurther stated that the eggs were " . . . appar-ently uninjured." Martin (1968) showed thatMeloidogyne arenaria (Neal, 1889) Chitwood,1949 survived passage through the digestivetract of a cow and subsequently infestedtomato roots.
According to Leukel (1924, p. 944), Mar-cinowski (1909) showed that Anguina tritici(Steinbuch, 1799) Filipjev, 1936 in wheatgalls survived passage through the intestinesof the sparrow, goldfinch, pigeon, and mouse,but did not survive passage through thechicken, gopher, or marmot. However, Da-vaine (1857) found that A. tritici in wheatgalls did not survive passage through the intes-tines of the chicken, pigeon, or sparrow, butdid survive passage through the frog, triton,salamander, and goldfish. Leukel (1924)found that A. tritici did not survive passagethrough the digestive tract of the horse, cow,hog, sheep, or chicken. Wilson (1930) foundthat Ditylenchiis dipsaci (Kiihn, 1857) Filip-jev, 1936 in diseased Phlox did not survivepassage through the intestine of a rabbit.
Research reported in this paper was con-ducted to determine whether eggs and larvaein cysts of Heterodem glycines would survivepassage through the digestive tract of swine.An abstract has been published (Smart, 1963).
Materials and Methods
Six pigs weighing an average of 63 Ib (28.6kg) each (60-65 Ib = 27.5-29.5 kg) and withan average age of 13 weeks (80—96 days) wereused in this study. Three of the pigs werefemales and three were males. At a later date,
rectal temperatures were recorded in six other13-week-old pigs (three of each sex) on threeconsecutive days. Temperatures were re-corded with the pigs unrestrained and in astanding position. The average rectal tem-perature of the six pigs was 104.2 F (40.1 C)ranging from a low of 103.0 F (39.4 C) in afemale to a high of 105.1 F (40.6 C) also ina female.
Six concrete-floored pens were washedthoroughly using a high pressure hose (110psi pressure at the pump). One animal wasplaced in each pen and feed was withheld for40 hr prior to initiating the study; however,drinking water was supplied ad lib. Once-each 24 hr for 3 consecutive days each pigingested 1,000 cysts of H. glycines whichwere mixed with the daily ration of 2 Ib (0.9kg) of a 15.6% crude protein feed. One ofthe pigs (pig 1 in Table 1) ingested cysts fromthe Holland, Virginia, area, and the other fiveingested cysts from the Wilmington, NorthCarolina, area. Control cysts were kept at 4 C.The feed was the standard growing ration usedat the Holland, Va. Station and was of thefollowing mixture: 1,598 Ib (726,3 kg) ofcorn, 252 Ib (114.5 kg) of soybean oil meal(50% crude protein), 100 Ib (45.4 kg) oftankage, 20 Ib (9.1 kg) of limestone, 10 Ib(4.5 kg) of salt (NaCl) , 5 Ib (2,3 kg) ofVitamin Pre-Mix (Charles Pfizer), 0.6 Ib (272g) of Terramycin TM50, 100 g of zinc sul-phate (ZnSO4), and 12 g of copper sulphate(CuSO4).
Feces were collected initially from each penjust before the second feeding of cysts. There-after for 4 additional days, feces were col-lected 3 times a day and composited. Afterthe third collection the feces were examinedby washing through a 60-mesh and a 325-meshscreen. Residues from the 60-mesh screen wereexamined directly for cysts, and residues fromthe 325-mesh screen were placed in Baermannfunnels for recovery of any larvae that mighthave been liberated within the alimentarycanals of the pigs.
On the fourth collection day, 50 cysts re-covered from the feces of each pig were placedin 4-inch (10.2 cm) pots of autoclaved soiland soybean seed, Glycine max Merr. var."Lee," planted. All other cysts on all collectiondays were isolated and placed in sterilized dis-tilled water in 60 mm Petri dishes at the rate
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 141
Table 1. Number and percentage of cysts of Heterodera glycines recovered from the feces of each of6 pigs on 5 consecutive collection days and the number of larvae emerged during incubation of the cystsfor 21 days at 22 C. Each pig ingested 3,000 cysts (1,000 on 3 consecutive days).
Pig number and
Day fecescollected
1234-5
I1-FemaleCysts Larvae
10580
13318528
30100
2-FemaleCysts Larvae
15 039 0
198 0215 0
46 0
sex, number of cysts recovered, and number of larvae emerged
3 -FemaleCysts Larvae
847
278150
27
00000
4 -MaleCysts Larvae
14268
19012778
10000
5-MaleCysts Larvae
53 068 0
300 0189 0104 1
6-MaleCysts Larvae
8713313418090
00000
ControlCysts Larvae
200 3,596200 3,776200 1,767136 65100 59
Total
Percentage of cystsrecovered per pig
531
17.7
513
17.1
510 0
17.0
605
20.2
714 624 0
20.8
836 9,263
1 Pig 1 ingested cysts from the Holland, Va. area, and all other pigs ingested cysts from the Wilmington, N. C. area.- Fifty cysts from each pig collected this day were placed immediately in 4 inch clay pots of soil seeded with "Lee"
soybean.
of 10 cysts per dish and incubated at 22 C.After incubation for 10 days, emerged larvaewere counted and the cysts were transferredto fresh dishes of sterilized distilled water.After 11 more days the emerged larvae wereagain counted. The cysts which had beenincubated and all emerged larvae were placedin 4-inch (10.2 cm) pots of autoclaved soilplanted to "Lee" soybean. After 60 days, thesoybean plants were removed from the soil.The roots were placed on a 60-mesh screenand washed with a spray of water using enoughpressure to remove cysts from the roots. Thesoil was roiled in a container of water andwashed through a 60-mesh screen with theprocess repeated twice. The residue was dis-tributed in counting trays and any cysts presentwere counted with the aid of a dissecting mi-croscope at 15 X.
Results
A total of 3,497 or 19.4% of the 18,000cysts fed to the six pigs were recovered (Table1). Only six larvae emerged from the 3,197cysts subjected to larval emergence tests. Thiswas an average emergence of 0.002 larvae percyst. The control cysts yielded 9,263 larvaefor an average emergence of 11 larvae percyst. The six larvae which emerged from cystsrecovered from the pigs were alive as deter-mined by movement both autonomously andfrom tactile stimulation. The intestines of thelarvae, however, were highly vacuolated. Theintestines of larvae manually released fromeggs in the cysts recovered from pigs also
were vacuolated. These larvae were dead;they remained in a bent position and did notmove either autonomously or from tactilestimulation.
No larvae were recovered from the Baer-mann funnels containing the debris caught onthe 325-mesh screen.
No soybean cyst nematodes developed on"Lee" soybean during a 60-day period fromthe cysts and six emerged larvae used as inoc-ulum. From the control cysts, an average of19 cysts or white females were recovered foreach cyst used as inoculum.
Discussion
Although a few larvae survived passagethrough the digestive tract of the 13-week-oldpig, they did not appear as "normal," basedon the vacuolated appearance of the intestine,as larvae not similarly treated. Balmer andCairns (1963) stated that infectivity droppedsharply in root-knot nematode larvae contain-ing five or more large-sized vacuoles in theintestinal region. The larvae of H. glycineswhich survived passage through the pig, aswell as those which did not survive, had highlyvacuolated intestines. (The number of vac-uoles were not counted since this work wasdone prior to the publication of the work byBalmer and Cairns.)
Triffit (1929), in seeking an explanation forthe death of H. rostochiensis ingested by the6-week-old pig, pointed out that the tempera-ture in young pigs is higher than in older ones,but she did not believe body heat to be the
Copyright © 2011, The Helminthological Society of Washington

142 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
decisive factor in death of the nematode. Werecorded the average rectal temperature of pigs5 weeks old, 13 weeks old, and 24 weeks oldwith the following average temperatures, re-spectively: 104.2 F (40.1 C), 104.2 F (40.1C), and 102.4 F (39.1 C). Since Endo (1962)showed that larvae of H. ghjcines in cysts livedmore than 8 hr at 110 F (43.3 C), we do notbelieve that body heat of the pigs alone wasresponsible for the death of most of the larvaeof H. ghjcines.
Sex of the pigs apparently had no effect onsurvival or death of the larvae, since larvae ofH. ghjcines emerged from cysts recovered fromthe feces of both males and females.
Since a very few larvae in cysts of H.ghjcines did survive passage through the diges-tive tract of swine, it is possible that the nema-tode could be disseminated through swinefeces. However, based upon the vacuolatedappearance of the intestinal region of thoselarvae, and upon the lack of reproduction on asusceptible host plant, we believe that dis-semination of the nematode through the fecesis unlikely.
Summary
Of 18,000 cysts of Heterodera ghjcines in-gested by six pigs, 19.4% were recovered fromthe feces. Six larvae emerged from the cystsincubated for larval emergence. The larvaewere alive, but had highly vacuolated intes-tines. When these larvae and all cysts re-covered from the feces were placed in pots ofsoil containing "Lee" soybean, a known host,no cyst nematodes developed.
Literature Cited
Balmer, E., and E. J. Cairns. 1963. Relation ofphysiological age of root-knot larvae to infec-tivity. Phytopathology 53(6): 621. (Abstr.).
Chatin, Joannes. 1890. Le mouton peut-il pro-pager YHeterodera Schachtii? Compt. Rend.Soc. Biol. 9th Series, 11(2): 17-18.
Davaine, C. 1857. Recherches sur 1'anguilluledu ble niele considered au point de vue de1'histoire naturelle et de 1'agriculture. Compt.Rend. Soc. Biol. 3 (2) : 201-271.
Ellenby, C. 1944. Influence of earthworms onlarval emergence in the potato-root eelworm,
Heterodera rostochiensis Wollenweber. Ann.Appl. Biol. 31(4): 332-339.
1946. Ecology of the eelworm cyst. Na-ture 157(3988): 451-452.
Endo, B. Y. 1962. Responses of Heteroderaglijcines to lethal temperatures. Phytopathol-ogy 52(1): 9. (Abstr.).
Girard, A. 1887. Sur le developpement des ne-matodes de la betterave, pendant les annees1885 et 1886, et sur leurs modes de progaga-tion. Compt. Rend. Acad. Sci. 104, S.
Gof'fart, H. 1934. Ueber die Biologic und Be-kampfung des Kartoffelnematoden (Hetero-dera schachtii Schmidt). Arbeiten Biol. Reichs.Land-u. Forstwirtschaft. 21(1): 73-108.
Kofoid, C. A., and A. W. White. 1919. A newnematode infection of man. J. Amer. MedicalAssoc. 72: 567-569.
Leukel, R. W. 1924. Investigations on the nem-atode disease of cereals caused by Tylenchustritici. J. Agr. Research 27(12): 925-955 plus5 plates.
Marcinowski, K. 1909. Parasitisch und semi-parasitisch an pflanzen lebende nematoden.Arb. k. Biol. Anst. Land. u. Forstw. 7: 1-192.
Martin, G. C. 1968. Survival and infectivity ofeggs and larvae of Meloidogyne arenaria afteringestion by a bovine. Plant Disease Reptr.52(2) : 99-102.
Miller, L. I. 1960. Soybean cyst nematode re-search in Virginia. Virginia-Carolina PeanutNews. 6(1) : 9-11.
Sandground, J. H. 1923. "Oxyuris incognita"or Heterodera radicicola? J. Parasit. 10: 92-94.
Smart, G. C., Jr., and Barbara A. Wright. 1962.Survival of the cysts of Heterodera glijcinesadhering to stored sweet-potato, peanut, andpeanut hay. Virginia J. Sci. (N.S.) 13(4):219-220. (Abstr.).
Smart, G. C., Jr. 1963. Survival of encystedeggs and larvae of the soybean cyst nematode,Heterodera glijcines, ingested by swine. Phyto-pathology 53(8): 889. (Abstr . ) .
Triffit, M. J. 1929. Observations on the inci-dence of Heterodera schachtii at the OrmskirkPotato Testing Station. J. Helminth. 7 ( 2 ) : 93-98.
Willot. 1890. Destruction cle YHeterodera sch-achtii. Compt. Rend. Acad. Sci. 3,S: 801-803.
Wilson, G. F. 1930. Further investigations onthe eelworm disease of Phloxes. J. Royal Hort.Soc. 55(1): 88-100.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 143
The Intrapulmonary Localization of Angiostrongyluscantonensis in the Rat
THOMAS M. SODEMAN,* WILLIAM A. SODEMAN, JR.,2 AND CHARLES S. RICHARDSS
Angiostrongylus cantonensis was describedby Chen (1935). Mackerras and Sandars(1955) in a detailed description of its lifecycle showed that the adult worm migratesfrom the rat's subarachnoid space to its lungsbetween the 28th and 31st day of infection.Fertilization takes place in the pulmonaryarteries and oviposition begins about the 35thday.
The purpose of this investigation was tostudy the localization of adult worms, eggs,and larvae in the lungs of experimentallyinfected rats. Because of the difficulty inmaintaining spatial relationships after re-moval of organs from an animal, particularlywhen making histologic sections from smallblocks of the whole organ, it was elected tosection the whole lung. The Gough-Went-worth (1960) technique for making thick sec-tions (200 ju.) of whole, inflated fixed lungswas used. These sections permitted the local-ization of lesions within the matrix of thepulmonary parenchyma with little loss ofthe spatial relationships between pathologicchanges and normal tissue.
Methods and Materials
Weanling, Sprague-Dawley rats were in-fected with A. cantonensis by feeding of one-half of a Biomphalaria glabrata (= Aiistralorbisglabratus) snail harboring infective third stagelarvae. The rats were divided into six groups.The first group was sacrificed on the 28thday of infection and thereafter a new groupwas sacrificed on the 34th, 37th, 39th, 43rd,and 65th day of infection. The heart andlungs were removed en block, avoiding injuryto the visceral pleural surfaces. The lungs wereinflated with a formalin acetate mixturethrough the trachea by a cannula connectedto a reservoir 4 feet above. The formalinacetate mixture consisted of 500 ml of liquid
1 Section on Primate Malaria, Laboratory of ParasiteChemotherapy, NIAID, P.O. Box 80190, Chamblee, Georgia30341.
2 Liberian Institute of Tropical Medicine, Harbel, Liberia.3 Laboratory of Parasitic Diseases, NIAID, National In-
stitutes of Health, Bethesda, Maryland 20014.
formaldehyde (40%) and 200 gm of sodiumacetate, brought to 5,000 ml with distilledwater. The inflated lungs were fixed in theformalin acetate mixture for a minimum of2 days.
The lungs were separated from the tracheaand heart. Each lung was washed for 72 hrin running water into which 5% nitric acidwas dripped. The entire lung was embeddedin a solution of 250 mg 80—100 bloom gelatin,40 ml ethylene glycol mono-ethyl ether, 5 mlcarpyl alcohol, 5 ml 1% ethylmercurithio-salicylate and 850 ml distilled water. Thespecimen was allowed to stand for 1 hr andwas then transferred to an incubator and keptfor 48 hr at 35 C. The entire block was frozenand kept at —20 C until sectioning. Sectionswere cut at 200 JJL on a Reichert microtome asthawing occurred and they were stored in tapwater. The sections were floated onto glassslides for examination with a dissecting micro-scope. Several sections were stained withhematoxylin and eosin.
Lungs from each group were also embeddedin paraffin and sectioned at 5 p. These sec-tions were stained with Giemsa for comparisonwith the thick sections.
ResultsThe lungs of a rat infected with A. canto-
nensis 65 days prior to sacrifice revealed ma-ture worms only within the vascular system.They were restricted to the larger pulmonaryarteries. The thick sections permitted visualiza-tion of large portions of the adult worms, mostwith ingested blood outlining their digestivetracts, within the mainstem pulmonary arteriesand peripherally in the larger branches (Fig.1). There was no evidence that the matureworms penetrated into the pulmonary paren-chyma or the bronchial system at 65 days.
Eggs surrounded by fluffy white consolida-tion were scattered throughout the lung fields.From serial sections it was apparent that egglesions were associated with blood vessels. Aportion of the left middle lobe was infarcted(Fig. 2) . Serial sections of the lung revealed
Copyright © 2011, The Helminthological Society of Washington

144 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Figures 1—4. Thick sections of the left lung from a rat sacrificed 65 days after infection. 1. Angio-strongylus cantonensis fill the main pulmonary artery. Egg lesions are seen as patchy white areas(X 2.5). 2. Infarcation of a portion of the middle lobe (X 2.5). 3. Thrombosis of the artery leading tothe infarcted lobe in Figure 2. The proximal portion contains eggs, the distal contains developing larvae(X 200). 4. A thick section of the lung demonstrating eggs embolized to the terminal arteries (X 200).
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 145
Figures 5—6. 5. A thin section of the lung from a rat sacrificed at 39 days with an egg in a capillaryof an alveolar wall (X 735). 6. A thin section of the lung from a rat sacrificed at 65 days with eggs inadvanced stages of development surrounded by organizing tissue. A small artery can be traced to anegg lesion. The proximity of the eggs to alveoli and a bronchus is evident (X 580).
thrombosis of the artery to that lobe (Fig. 3).The thrombus was composed of two distinctsections; the older, more distal portion con-tained hatched larvae, the proximal portion,developing eggs. Where eggs were seen out-side of the arterial system, the integrity of thevessel was in question because of surroundinghemorrhage or an intense inflammation withdisruption of normal structures.
In many cases, the plane of the section andits thickness permitted the tracing of arterialvessels leading to egg lesions (Fig. 4) . In thisfigure an egg can be seen occluding a branchof a small artery just distal to its bifurcationfrom the main blood-filled lumen. The patternof egg lesions and their association with arteriessuggested embolization. The bronchi containedno eggs.
Lungs from rats sacrificed on the 28th, 34th,and 37th days were normal. Those sacrificedon the 39th, 43rd, and 65th days postinfectionand processed as 5 /A paraffin sections showedhistologie changes similar to the thick gelatinsections. Adult worms were localized in the
pulmonary arteries. In the early stages of in-fection, eggs were clearly found only in pul-monary vessels (Fig. 5). With advancing agean extensive reaction formed about the eggswith replacement of blood vessel structure byactive fibroblastic proliferation and markedleukocytic and monocytic infiltration. As thelarvae developed, there was an increasedeosinophilic reaction. In many places eggslying in their connective tissue masses wereseparated from alveoli or bronchi only by athin wall of tissue (Fig. 6). After 65 days ofinfection larvae were seen in the alveoli andsurrounded by eosinophils. The patchy whiteconsolidation in the thick sections was seen toconsist of organized tissue, collections ofmacrophages and polymorphonuclear leuko-cytes in the thin sections. Alveolar septa asso-ciated with intracapillary eggs were thickenedand infiltrated with polymorphonuclear leuko-cytes, lymphocytes, and macrophages. In themore advanced stages, the large pulmonaryarteries were dilated and contained masses ofeggs.
Copyright © 2011, The Helminthological Society of Washington

146 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
DiscussionThe results of the thick, as well as thin,
sections indicated that the adult stages wererestricted to the pulmonary arteries. Theirsize limited them to the larger arteries. Noevidence in the stages studied, either in thethick gelatin or thin paraffin sections, indicatedthat the adult worm penetrated into the pul-monary parenchyma. The findings suggestedthat oviposition occurs in the blood stream andthat the wide distribution of pulmonary in-volvement resulted from embolization of eggs.This embolization and impaction in blood ves-sels resulted in thrombosis and infarction. Thatdevelopment may progress within the throm-bus was evident in Figure 3 where hatchedlarvae were seen in the distal portion of athrombus, while in the recently formed prox-imal portion, eggs were found. There wasnothing to suggest a preferential distributionof egg lesions in the lungs. However, becauseof the close association of the bronchial andvascular systems, many of the egg lesions werein peribronchial locations.
When eggs were seen in the pulmonary par-enchyma, they were surrounded by inflamma-tion and organization that obscured their orig-inal intravascular position. Furthermore, it wasevident that eggs outside of vessels may haveresulted from disruption of vessel walls withhemorrhage or from infarction. Both processeswere evident in the sections. The extensiveorganization around the eggs in the arteriesextended into the perivascular and frequentlyinto the peribronchial tissue. The materialsexamined suggest that the primary site of de-velopment is within the arterial system and that
the pulmonary parenchyma acts as a secondarysite for development when eggs escape fromthe vascular system. It is likely that the larvaemigrate from the arteries through the orga-nized tissues directly into the bronchial systemor out into the alveolar sacs.
The use of the thick section technique per-mitted better visualization of eggs within thelung substance without loss of orientation. Theauthors feel this technique deserves considera-tion as an experimental method in studyingpulmonary parasitic lesions.
SummaryThe intrapulmonary localization of Angio-
strongyhis cantonensis in experimentally in-fected rats was studied in lungs embedded ingelatin and sectioned at 200 /.i. Adult wormswere found only intravascularly. Egg deposi-tion and larval development within lungs wereprimarily in blood vessels. Eggs were distrib-uted by embolization. It is suggested that thepreferred site of larval development is intra-vascular rather than in lung parenchyma.
Literature CitedChen, H. T. 1935. Un nouveau nematode pul-
monaire, Pulmonenui cantonensis, N. G., N.SP. Des rats de Canton. Ann. Parasit. Hum.Comp. 13: 312-317.
Mackerras, M. J., and D. F. Sandars. 1955.The life history of the rat lung-worm, Angio-strongylus cantonensis (Chen) (Nematoda:Metastrongylidae). Aust. J. Zool. 3: 1-21.
Gough, J., and J. E. Wentworth. 1960. Thinsections of entire organs mounted on paper.Recent advances in pathology, 7th Ed.(Edited by C. V. Harrison, M.D.), Little,Brown and Company, pp. 80-86.
BUSINESS OFFICE
For the convenience of members and subscribers and to better serve both membersand subscribers, the Society on January 1, 1969 established its business office atAllen Press, Inc., in Lawrence, Kansas. All dues, subscriptions, orders for back issues,and any and all correspondence with reference to these items or the receipt of Pro-ceedings should be addressed to:
HELMINTHOLOGICAL SOCIETY OF WASHINGTONc/o ALLEN PRESS, INC.1041 NEW HAMPSHIRE ST.LAWRENCE, KANSAS 66044U.S.A.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 147
Tylenchus vesiculosus, sp. n. (Nematoda: Tylenchidae)From Soil in Michigan
NATALIE A. KNOBLOCHX AND JOHN A.
Tylenchus vesiculosus sp. n. was recoveredfrom soil samples collected 16 October 1967from a potato field near East Lansing, Mich-igan. The habitat, a field in which potatoeshad been under cultivation for several years,was organic soil (muck) with two out-cropping knolls of sandy mineral soil. T.vesiculosus, while not widely distributedthroughout the field, was found in large num-bers around the root systems of potato plantsgrowing in the sandy soil. Only one male wasrecovered from the original soil samples.
A ratio of one male for about every 50 fe-males was recovered from samples processed inApril 1968 but these males were inadvertentlydestroyed in the laboratory. Since the originalinfestation was to be fumigated prior to springplanting, soil was transferred from the originalhabitat to a microplot at the Entomology Re-search Farm, Michigan State University in anattempt to preserve the species for future biol-ogy investigations.
The following description of the speciesis based on preserved specimens. Measure-ments were made on specimens which hadbeen heat relaxed, fixed in F.A. 4:10, pre-served and mounted according to the glycerol-ethanol method of Seinhorst (1959).
Tylenchus vesiculosus sp. n.(Figs. 1-7)
HOLOTYPE FEMALE: Length = 0.55 mm;a = 29; b = 6.2; c = 4.7; V = ^66; spear =9.75 p..
PARATYPE FEMALES (12): Length = 0.52(0.51-0.55) mm; a = 31.3 (28-34); b = 6.1(6.0-6.6); c = 4.8 (4.7-4.9); V = 66 (66-68); spear = 9-11 p.
ALLOTYPE MALE: Length = 0.48 mm; a =30; b = 5.5; c = 4.5; T = 46; spear = 9.7 p..
1 Research Technician, Entomology Department, Mich-igan State University, East Lansing, Michigan.
- Assistant Professor, Entomology Department, MichiganState University, East Lansing, Michigan.
Published with the approval of the Director of the Mich-igan Agricultural Experiment Station as Journal ArticleNumber 4453.
FEMALE (HOLOTYPE) : Body filiform slightlyarcuate ventrally, tapering more towards pos-terior than anterior end. Cuticle of bodycoarsely striated; annules 1.9 ^ wide at middleof body. Lateral field a plain band borderedby two bright lines one-fourth to one-fifthgreatest body width (Fig. 3).Phasmids notobserved. Lip region conoid, rounded, contin-uous with neck contour; no lip annules ob-served. Spear delicate, 9.75 p long, with well-developed knobs (Fig. 2). Median bulb ofoesophagus ovate with crescentic valve plates;the metacorpus obscure in many of the glyc-erine specimens. Isthmus slender, surroundedby the nerve ring slightly anterior to wherethe isthmus joins the basal oesophageal bulb.Basal bulb elongate-pyriform; cardia conical,well defined. Excretory pore opens at aboutthe middle of the basal bulb (Fig. 1). Vulvaa depressed transverse slit; vagina thick-walled, occupying about three-fifths bodywidth; length of posterior uterine sac aboutfour-fifths vulval body width. Ovary single,outstretched anteriorly, wtih a round to ovalshaped spermatheca with an extended pouch-like extension (Fig. 5). Spermatheca and theextension contained many spermatozoa. Ovarywith oocytes arranged in a single file and ex-tending two-thirds of the distance to oesoph-agus (Fig. 4). Anus distinct; tail length about9 times anal body width, ending in a roundedtip.
MALE (ALLOTYPE) : Length = 0.48 mm;a = 30; b = 55; c = 4.5; T = 46; spear =9.7 p.
Body shape, head and oesophagus same asin female (Figs. 1, 2) .
Testis single, outstretched anteriorly; sper-matocytes arranged in single file. Spiculespaired, curved, of usual tylenchoid type.Gubernaculum thickened, troughlike; bursafaintly crenate, rising at a point a little aboveproximal ends of spicula and extending pastanus a distance equal to about 1% anal bodydiameter (Figs. 6, 7). Tail same shape as infemale; about 10 times anal body width.
Copyright © 2011, The Helminthological Society of Washington

148 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
7
6.
V-'.-V
Figures 1-7. Tylenchus vesiculosus. 1. Anterior portion of female. 2. Female head region. 3. Lateralfield of female. 4. Female reproductive tract showing position of spermatheca. 5. Sketch of spermathecashowing ventral view of attached pouch. 6. Male tail. 7. Posterior portion of male with lateral lines.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 149
DIAGNOSIS: Tylenchus vesiculosus sp. n. canbe differentiated from all known species of thegenus by the spermatheca with an extendedpouch. It is small (0.54 mm) with thick, heav-ily annulated cuticle, plain bandlike lateralfield and tail with a distinctly rounded termi-nus. It differs from Tylenchus davainei Bastian,1865, in the straight tail, bandlike lateral field,unstriated lip region and the spermatheca withthe pouch. Distinguished from Tylenchusditissimus Brzeski (1963) by the coarser bodystriation, no annules in the lip region, bandlikelateral field, position of hemizonid, longerlength, possession of post uterine sac and aspermatheca with a pouch. The extendedpouch of the spermatheca is known for noother species of the genus Tylenchus. How-ever, a similar pouch is described for Dac-ti/lotylenchus crassacuticulus Wu (1968).
HOLOTYPE, FEMALE: Collected 16 October1967 from a potato field in East Lansing,Michigan. Slide T-118t deposited with theUSDA Nematology Collection at Beltsville,Maryland.
ALLOTYPE, MALE: Same data as holotype,slide T-117t.
PARATYPES, FEMALES: Same data as holo-type, 12 slides Tv3-Tvl4 deposited in Depart-
ment of Entomology Collection, MichiganState University, East Lansing, Michigan.
TYPE HABITAT: Organic and mineral soilaround roots of potato, Solanum tuberosum L.
TYPE LOCALITY: A cultivated farm field ofpotatoes grown in organic and mineral soilnear East Lansing (elevation 845 feet),Michigan, USA.
The authors are grateful to Professor GeraldThorne, Department of Plant Pathology, Uni-versity of Wisconsin, Madison, Wisconsin forhis advice during this work and to Richard J.Snider for his patient counseling in drawingtechniques.
Literature CitedBrezeski, M. W. 1963. Tylenchus ditissimus
sp. n., a new nematode from Poland (Nema-toda, Tylenchidae). Bull. Acad. Pol. Sci.(11): 531-535.
Seinhorst, J. W. 1959. A rapid method for thetransfer of nematodes from fixative to anhy-drous glycerin. Nematologica 4: 67-69.
Wu, L. Y. 1968. Dactylotylenchus crassacuti-culus, a new genus and a new species (Tylen-chinae: Nematoda). Can. Jour. Zool. 46(5):831-834.
NEW JOURNALThe Society of Nematologists announces the publication of a new journal, the
JOURNAL OF NEMATOLOGY. Appearing quarterly beginning in January 1969, it isintended as a medium for publication of original research in all phases of nematology.Membership in the Society of Nematologists (annual dues $6.00) includes the Journalof Nematology. Subscriptions are available at $10.00 domestic and $11.50 foreign,per volume. Inquiries should be directed to: Society of Nematologists, Departmentof Botany and Plant Pathology, Auburn University, Auburn, Alabama 36830; or Dr.Seymour D. van Gundy, Editor, Journal of Nematology, Department of Nematology,University of California, Riverside, California 92502.
Copyright © 2011, The Helminthological Society of Washington

150 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Helminths of the Crow, Corvus brachyrhynchosBrehm, 1822, in North Carolina1
LARRY D. HENDRicics,2 REINARD HARKEMA, AND GROVER C. MILLERDepartment of Zoology, North Carolina State University at Raleigh
There are few comprehensive reports onthe helminths of crows in North America.Ward (1935) conducted the first extensivesurvey of crows. This work included dataon the helminths of 50 crows collected inOklahoma. Morgan and Waller (1941) ex-amined 112 crows from Wisconsin and Iowa.Daly (1957a, 1957b) examined 103 southerncrows (Corvus brachyrhynchos paulus Howell)from Virginia. Most references on parasitesfrom the crow are for particular species or onsurveys of hosts which included crows as partof the study. The object of our investigationwas to determine the parasitic fauna and theincidence of parasitism of both external andinternal parasites of the crow in North Caro-lina. The external parasites are reported else-where (Hendricks and Axtell, 1968).
Materials and MethodsOne hundred and fifty-eight crows, 76 fe-
males and 82 males, were collected and ex-amined for parasites between 5 October 1965and 22 April 1967. The majority of thesebirds was taken in the spring of 1966 and1967 in conjunction with the annual hunt ofthe Southeastern Crow Shooters Association inthe vicinity of Henderson, North Carolina.All hosts were data-tagged and sealed in plasticcollecting bags. Whenever possible hosts wereautopsied within a few hours after collection;otherwise they were frozen and stored.
Trematodes were killed and fixed in hotA.F.A.; acanthocephalans and tapeworms werekilled in boiling water and then fixed andstored in A.F.A.; nematodes were killed andfixed in hot 70% ethanol and stored in 10%glycerine in 70% ethanol.
For microscopic study, trematodes, cestodes,and acanthocephalans were stained with Harris'
1 From a thesis submitted to the Graduate Faculty ofNorth Carolina State University at Raleigh in partial ful-fillment of the requirements for the degree of Master ofScience. This project was supported in part by Grant No.AI-05927, National Institutes of Health.
2 Address: 9th Medical Laboratory, APO San Francisco96307.
hematoxylin, Semichon's carmine, or fast green,cleared in methyl salicylate, and mounted inCanada balsam. Nematodes were cleared inlacto-phenol.
Results and DiscussionTwelve species of helminths were recovered
from 158 crows. These included two trema-todes, two cestodes, three acanthocephalans,and five nematodes (Table 1).
Brachijlecithum americamnn Denton, 1945is a common parasite in the liver of birds ofthe families Corvidae and Icteridae and wasdescribed from specimens obtained from mea-dowlarks, grackels, crows, and blue jays fromGeorgia, Tennessee, and Texas. It was foundin the liver of 122 of the 158 crows examined.It was reported from the yellow throat, Geo-thlypis trichas at Durham, N. C. (Wells andHunter, 1960).
Echinostoma revolutum Froelich, 1802 wasfound in the small intestine of seven crows.Daly (1959) reported this form from thesouthern crow in Virginia. This is the firstreport from crows in North Carolina.
Only one specimen of Anomotaenia sp. wasfound in the small intestine. It possibly isA. constricta (Molin, 1858) which was re-ported from the southern crow in Virginia(Daly, 1959). Ransom (1909) listed the fishcrow (C. ossifragus Wilson), the raven (C.corax L.) and the crow as hosts for A.constricta.
Variolepis variabilis (Mayhew, 1925) wasfound in the small intestine of 34 of the hostsexamined and North Carolina is a new localityrecord. It has been reported from the crowin Illinois (Mayhew, 1925), Wisconsin (Mor-gan and Waller, 1941) and Virginia (Daly,1959).
Mediorhynchiis grandit Van Cleave, 1916was found in the small intestine of two crows.The two specimens recovered were immaturemales. This species has been reported fromthe grackle, eastern meadowlark (Van Cleave,
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 151
Table 1. Helminths recovered from 158 crows,Corvus brachyrhynchos Brehm.
Average numberNumber of helminths per
Parasite infeeted infected host
TrematodesBrachijlecithum am ericami mEchinostoma revolutum
CestodesAnomotaenia sp.*Variolepis variabilis*
AcanthocephalaMediorhynchus grandisMacracanthorhynchus sp.fPlagiorhynchns formosum
NematodesCapillaria contorta*Hystrichis sp.*tPhysaloptera sp.*fAcnaria anthuris*Diplotriaena trictispis*
* New locality record.f New host record.
1227
134
221
21122
351
14.52.8
1.02.1
1.01.51.0
1.41.82.02.41.0
1916), and the crow (Van Cleave, 1918, 1947;Daly, 1959).
The infection of two crows with Macra-canthorhynchus sp. probably was accidental.The specimens are immature females and re-semble M. hirudinaceus.
Plagiorhynchus formosum Van Cleave, 1918was found in the small intestine of one crow.It has been reported from the crow in theDistrict of Columbia (Jones, 1928) and theMacgillivray seaside sparrow in North Carolina(Hunter and Quay, 1953).
Capillaria contorta (Creplin, 1839) wasremoved from the mucosa or submucosa ofthe esophagus of 21 hosts. Canavan (1931)reported this species from a crow in thePhiladelphia Zoological Garden and Morganand Waller (1941) reported it from crows inWisconsin and Iowa. This is the first reportof this species from North Carolina.
Hystrichis sp. was recovered from the glandsof the proventriculus of 12 of the 158 crows.The specimens constitute a new species andit will be described separately. This is thefirst report of a representative of this genusin North America and the crow is a new host.
Two immature female specimens of Phy-saloptera sp. were recovered from one host.They resemble F. alata Rudolphi, 1819.
Acuaria anthuris (Rud. 1819) was found inthe crop and gizzard of 35 hosts. This specieswas reported from the crow in Nebraska
(Williams, 1929), Oklahoma (Ward, 1935),New Jersey (Beaudette and Hudson, 1936),Wisconsin and Iowa (Morgan and Waller,1941). This is the first report of this speciesin North Carolina.
Diplotriaena tricuspis (Fedtschenko, 1874)was recovered from an abdominal air sac ofone crow. Probably this is the only adultfilarid species reported from crows in NorthAmerica (Beaudette and Hudson, 1936).Wells and Hunter (1960) reported an imma-ture Diplotriaena sp. from the yellow throat inDurham, N. C.
It is interesting to compare this study withthat of Daly (1959) who examined 103 south-ern crows taken in Albemarle County, Vir-ginia. There is a distance of 100-150 milesfrom the counties in which the majority ofthe crows in this survey was taken, yet onlythree parasites were common to both studies.These were: the trematode Echinostomarevolutum Froelich, 1802, the cestode Hyme-nolepls variabile (Variolepis variabilis of thispaper), and the acanthocephalan Mediorhyn-chus grandis Van Cleave, 1916.
Summary
A total of 158 crows harbored the followinghelminths: Trematoda: Brachylecithum. amer-icanum, Echinostoma revolutum; Cestodes:Anomotaenia sp., Variolepis variabilis; Acan-thocephala: Mediorhynchus grandis, Macra-canthorhynchus sp., Plagiorhynchus formosum;Nematodes: Capillaria contorta, Hystrichis sp.,Physaloptera sp., Acuaria anthuris, Diplotriaenatricuspis. New locality records are noted forAnomotaenia sp., V. variabilis, Hystrichis sp.,A. anthuris, and D. tricuspis, and new hostrecords for Macracanthorhynchus sp., Hys-trichis sp. and Physaloptera sp.
AcknowledgmentsSincere appreciation is extended to Dr.
Helen Ward for her assistance in the spe-cific identification of the acanthocephalans,and to Mrs. MayBelle Chitwood for her opin-ions and suggestions on the new species ofHystrichis.
Literature CitedBeaudette, F. R., and C. B. Hudson. 1936.
Parasitism and tuberculosis in a crow. J. Am.Vet. Med. Assoc. 89: 215-217.
Copyright © 2011, The Helminthological Society of Washington

152 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Canavan, W. P. N. 1931. Nematode parasitesof vertebrates in the Philadelphia ZoologicalGarden and vicinity. Parasitology 23: 196-229.
Daly, E. F. 1959. A study of the intestinal hel-minths of the southern crow (Corvns brachy-rhynchos paulus) in Virginia. Proc. Helm.Soc. Wash. 26: 66.
Hendricks, L. D., and R. Axtell. 1968. Arthro-pods from passerine birds of North Carolina.J. Elisha Mitchell Sci. Soc. 84: 312-314.
Hunter, Wanda S., and T. L. Quay. 1953. Anecological study of the helminth fauna ofMacgillivray's seaside sparrow, Ammospizamaritima macgttlivraii (Audubon). Am. Midi.Nat. 50: 407-413.
Jones, M. F. 1934. An acanthocephalid, Pla-giorhynchus formosum, from the chicken androbin. J. Agr. Res. 36: 773-775.
Morgan, B. B., and E. F. Waller. 1941. Someparasites of the eastern crow (Corvus brachy-
rhynchos brachyrhynchos Brehm). Bird-Band-ing 12: 16-22.
Van Cleave, H. J. 1918. The acanthocepliala ofNorth American birds. Trans. Am. Micros.Soc. 61: 206-210.
. 1947. The acanthocephalan genus, Me-diorhynchus, its history and a review of thespecies occurring in the United States. J.Parasit. 33: 297-315.
Ward, J. W. 1935. Some parasites of crows ofcentral Oklahoma. (Preliminary report) Proc.Okla. Acad. Sci. 15: 6.5-66.
Wells, M. J., and Wanda S. Hunter. 1960.Helminths of the yellow throat, Geothlypistrichae, during migration. J. Parasit. 46: 623.
Williams, O. L. 1929. A critical analysis of thespecific characters of the genus Acuaria, nem-atodes of birds, with descriptions of newAmerican species. Univ. Calif. Publs. Zool.33: 69-107.
SECOND INTERNATIONAL CONGRESS OF PARASITOLOGYMayflower Hotel, Washington, D. C. U.S.A.
September 6-12, 1970The Second International Congress of Para-
sitology was organized by the American Societyof Parasitologists and is co-sponsored by theHelminthological Society of Washington, Na-tional Academy of Science, Society of Nema-tologists, Society of Protozoologists, WildlifeDisease Association, and World Federation forthe Advancement of Veterinary Parasitology.The program will include any and all aspectsof human and animal parasitology and plantnematology except the description of new spe-cies. Parasitology is used in its generally ac-cepted connotation and hence would not in-clude bacteriology, virology, mycology, pestsor pesticides.
Plans for the development of the meetingsare progressing well. The ad hoc OrganizingCommittee and the ad hoc Program Committeehave developed the keynote of informality forfree and informal exchange. Every effort isbeing made to recapture the spontaneity, en-thusiasm, and stimulation that has character-ized some of the few unscheduled and im-promptu discussions that have developedwithin some other international meetings. Ac-cordingly it is proposed to limit formal presen-tations, with invited papers, to the morning
It is proposed to have a series of informaldiscussions in the afternoon sessions. Thusfree contributions (submitted papers) wouldbe grouped in appropriate sessions. It is fur-thermore proposed to prepublish these aslonger than average resumes (perhaps 500words) to serve as the basis for informal dis-cussion. Thus the chairman or any participantmight ask for elaboration, clarification, or dis-cussion of any paper or any aspect therein.Lantern slide projectors will be available.However, it is hoped that free discussion willnot be stifled by the routine reading of aseries of short papers.
This is frankly an experiment in internationalmeetings. The concept has been enthusiasti-cally endorsed in many quarters. We invitecomments and suggestions on the concept andany and all aspects of the program. The adhoc Program Committee welcomes suggestionsfor presiding officers at these sessions whichcould number as many as 60 or more.
Plan to attend the Congress and participate.GILBERT F. OTTOSecretary General
Zoology DepartmentUniversity of MarylandCollege Park, Maryland 20742
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 153
Exposure of Chicks to the Metacercaria ofEchinostoma revolutum (Trematoda)1
BERNARD FRIED AND LINDA J. WEAVERDepartment of Biology, Lafayette College, Easton, Pennsylvania 18042
Although Beaver (1937) has contributed anextensive monograph on the biology, syste-matics and life history of Echinostoma revo-lutum (Froelich, 1802) and Senger (1954)has studied aspects of growth, developmentand survival of this parasite, detailed studieson its infectivity in domestic chicks are lacking.During studies on the growth and developmentof this fluke in ectopic sites (Fried and Von-roth, 1968; Fried, Weaver, and Kramer, 1968)observations on the infectivity of E. revolutummetacercariae in the chick were made and arerecorded herein.
Materials and Methods
Helisoma trivolvis (Say) naturally infectedwith E. revolutum larvae were placed in fingerbowls containing pond water with laboratory-reared Physa sp. snails. Cercariae emergedfrom the H. trivolvis and encysted primarilyin the kidney of the Physa. The number ofcysts varied depending upon the size of thePhysa but ranged from 25—250. Physa werecrushed in pond water from 1-10 days post-infection and cysts were transferred to 3-saltRinger's (Paul, 1960) and teased into clumpsof approximately 25, 50, or 100 cysts. Exactnumbers of cysts were difficult to count soestimates of ± 5, 10, 20 were usually made.Except in experiments F and G (Table II)where cysts were pretreated in Ringer's or dis-tilled water and in the cloacal exposure study,cysts were pretreated in 3% NaHCOH for 30—60 min and then fed to chicks in 1-3 ml ofthe bicarbonate.
Day-old chicks were used in all experimentsexcept the reinfection studies (Table III).Chicks were not given food or water on theday of exposure or autopsy. They were killedby decapitation and either total worm countswere made or parasites found in the rectum,ceca, and ileum were recorded.
1 This work was supported by NIH Grant AI-06835. Theauthors gratefully acknowledge the technical assistance ofMr. Jack Carty.
Results and Discussion
Twenty-nine (93.6%) of 31 chicks eachexposed to 50 ± 10 to 100 ± 20 cysts andautopsied 22—35 days later were infected(Table I ) . Forty (95.2%) of 42 chicks eachexposed to 50 ± 10 cysts in 3% NaHCO:!, 3-salt Ringer's or distilled water and autopsied14 days later were infected (Table II) . Senger(1954) previously demonstrated that thedomestic chick could serve as a host of E.revolutum and reported that 22 (61%) of 36chicks fed metacercariae in water becameinfected.
Bacha (1964) found that an increased wormburden of Zygocotyle lunata (Diesing, 1836)in white rats could be obtained if metacer-cariae were pretreated in NaHCO3, possiblybecause the bicarbonate counteracted theacidity of the stomach. However, in matchedexperiments using NaHCO3 and distilled watersimilar worm burdens of E. revolutum wereobtained (Table II). Moreover, when cystswere pretreated in Ringer's the worm burdenincreased approximately twofold. Many fac-tors influence the rate of infectivity of E.revolutum, in chicks including the strain ofparasite and host, the emptying time of thegut, mechanical and chemical damage of cystsin the crop, proventriculus and gizzard andfurther investigations are needed to elucidatethese factors.
In older infections more worms were foundin the rectum than in the ileum. Thus whenchicks were autopsied in 2 weeks (Table II)272 worms were in the ileum and 241 in therectum, whereas in hosts autopsied 3—5 weeks(Table I) postinfection 105 worms were in theileum and 193 in the rectum. E. revolutum isshortlived in chicks and our records indicatethat rarely do infections persist up to 8 weeks.Senger (1954) reported the life span of E.revolutum in chicks to be about 35 days. Manyworms as they age migrate posteriorly fromtheir original site and either reestablish or passout of the host.
Copyright © 2011, The Helminthological Society of Washington

154 PROCEEDINGS OF THE HELMINTHOLOGICAL SOCIETY
Table 1. Summary o£ exposure experiments using 50±10-100±20 cysts pretreated in 3% NaHCO». Chickswere autopsied 22-35 days postinfectlon.
Exp.
ABCD
Total
No. ofchicks
exposed1
83
173
31
No. ofinfectedchicks
83
153
29
No. ofcysts per
chick
50±10-100±2050±1050±10
100±20
50±10-100±20
Age ofworm atautopsy
22232535
22-35
Number of worms recovered
Ileum
70
5840
105
Ceca
531429
6
102
Rectum
7246750
193
Total
13260
16246
400
Per centrecovery
22.040.021.615.3
22.2
1 All chicks day-old when exposed.
To determine the effects of storage on infec-tivity, cysts maintained for 7 weeks in 3-saltRinger's at 15 C were fed to chicks. Three offour chicks fed 100 ± 20 cysts and autopsied7-14 days later were infected with 9, 10, and23 worms. Senger (1954) reported that cystsstored in 0.4% saline at 5 C for 31 days wereviable. Storage of cysts is important in E.revolutum studies since many Phijsa that be-come heavily infected with metacercariae diewithin 2 weeks postinfection.
To determine if chicks could become in-fected with cysts via the cloaca, cysts wereimplanted into the cloacas of four chicks thatwere autopsied 7—10 days later. Two chickseach received 100 ± 20 cysts pretreated inbicarbonate and two chicks received the samenumber of cysts pretreated in Ringer's. Nonewas infected.
Cysts occasionally formed ectopically on themucus trails of H. trivolvi-s. To determine ifthese cysts were infective, three chicks werefed approximately 20, 60, and 175 mucus cystsand autopsied 7 days later. Only the one chickfed approximately 175 cysts was infected withfour worms suggesting that ectopically formedcysts are not as infective as those obtained fromthe kidney of Physa sp. snails.
To determine the effects of a secondary
infection on a primary one 12, 3-day-old chickswere each exposed to 100 ± 20 cysts and werereexposed to the same number of metacer-cariae 13 days later. Twelve additional chickswere used as controls, half of which receivedonly the primary exposure and the other halfthe secondary exposure. Chicks were autop-sied 9-10 days after the secondary exposure(Table III, H, I, J ) . To determine if chicksolder than 1 month could become reinfected,14 chicks that were 12 days old at the time ofa primary exposure to 50 ± 10 cysts werereexposed each to 50 ± 10 cysts 31 days later.Chicks were autopsied 10—11 days after thesecondary exposure (Table III, K). The re-infection experiments indicate that a primaryinfection of E. revolutum does not protectagainst a secondary infection. Fried (1963)found that a primary infection of Philo-phthalmus hegeneri Penner and Fried, 1963,did not protect chicks against a secondaryinfection and reviewed the literature on re-infection in trematodes. The reinfection ex-periments also indicate that chicks older than2 weeks (Exp. J) can become infected andthat chicks older than 1 month (Exp. K) canbecome reinfected indicating that an ageresistance to this worm was not apparent, atleast within the first month.
Table 2. Summary of exposure experiments using 50±10 cysts and different pretreatment solutions.Chicks were autopsied 14 days postinfection.
Exp.
EFG
Total
No. ofchicks
exposed1
181212
42
No. ofinfectedchicks
161212
40
Pretreatmentsolution
3% NaHCO.,3-salt Ringer'sDistilled H2O
Number of worms recovered
Ileum
10413038
272
Ceca
557335
163
Rectum
6310573
241
Total
222308146
676
Per centrecovery
27.751.324.3
32.2
1 All chicks day-old when exposed.
Copyright © 2011, The Helminthological Society of Washington

OF WASHINGTON, VOLUME 36, NUMBER 1, JANUARY 1969 155
Table 3. Summary of reinfection experiments.
Exp.
HIJK
No. ofchicks
exposed
1266
14
Age ofchick at
1° infection( days )
333
12
Age ofchick at
2° infection(days)
16161643
No. ofpos. chicks
w 1°infection
103-8
Age ofchick attime ofautopsy( days )
25-2625-2625-2653-54
No. ofworms
recoveredfrom 1°infection
8324-
15
No. ofpos. chicks
w 2°infection
9-57
No. ofworms
recoveredfrom 2°
infection
124-
174132
Chicks in Exp. H received 100 ± cysts in 1° and 2° infections.Chicks in Exp. 1 received 100 ± cysts in 1° infection !only.Chicks in Exp. J received 100 ± cysts in 2° infection only.Chicks in Exp. K received 50 ± cysts in 1° and 2° infections.
Summary
Sixty-nine (94.5%) of 73 day-old chicksexposed to 50-100 Echinostoma revolutionmetacercariae and autopsied 2—5 weeks post-infection yielded worm burdens of 15.3—51.3%.Forty (95.2%) of 42 day-old chicks exposed tometacercariae pretreated in distilled water, 3%NaHCO.j, 3-salt Ringer's and autopsied 2weeks later yielded worm recoveries of 24.3%,27.7%, and 51.3%, respectively. Cysts storedin Ringer's at 15 C for 7 weeks were viable.Cysts formed ectopically on the mucus trailsof Helisoma trivolvis snails were not as infec-tive as cysts formed within the kidney ofPhysa sp. snails. Worms were not recoveredfrom cysts implanted into the chick cloaca.A primary infection of E. revolutum did notprotect chicks against a secondary infection.Chicks older than 2 weeks became infectedand chicks older than 1 month became re-infected indicating that an age resistance tothis worm was not apparent, at least withinthe first month.
Literature Cited
Bacha, W. J., Jr. 1964. Effect of salt solutionson the establishment of infections of the trem-atode Zygocotyle lunata in white rats. J.Parasit. 50: 546-548.
Beaver, P. C. 1937. Experimental studies onEchinostoma revolutum (Froelich) a flukefrom birds and mammals. 111. Biol. Monog.15: 3-96.
Fried, B. 1963. Reinfection of chicks with anocular trematode, Philophthalmus hegeneriPenner and Fried, 1963. J. Parasit. 49: 981-982.
, and W. Vonroth, Jr. 1968. Trans-plantation of Echinostoma revolutum (Trem-atoda) into normal and surgically alteredchick intestine. Exper. Parasit. 22: 107-111.
-, L. J. Weaver, and M. D. Kramer. 1968.Cultivation of Echinostoma revolutum (Trem-atoda) on the chick chorioallantois. J. Para-sit. 54: 939-941.
Paul, J. 1960. "Cell and Tissue Culture." 2ndEd., Williams, Baltimore, Md., 312 p.
Senger, C. M. 1954. Notes on the growth, de-velopment and survival of two echinostometrematocles. Exper. Parasit. 3: 491-496.
Copyright © 2011, The Helminthological Society of Washington

156
Academician K. I. Skrjabin — Ninetieth Birthday
On December 7, 1968, Academician K. I.Skrjabin celebrated his ninetieth birthday andthe completion of more than six decades ofscientific work. Regarded as the outstandinghelminthologist of the Soviet Union, he hasgained worldwide recognition for his researchesin this field.
Graduating as a veterinary surgeon in 1905from the Juriev Veterinary Institute, Dr.Skrjabin first became interested in the studyof helminths during an assignment in Turkes-tan. He made extensive collections from bothdomestic and wild animals and later (1912-1914), studied them under the guidance ofBraun, Fuhrmann, Luhe, Railliet, and Henry.In his master's thesis (1916), based on theTurkestan collections, he stressed the impor-tance of establishing helminthology as a sci-ence in Russia. In the following year, he wasnamed to the first chair of parasitology inRussia, established at the Novocherkassk Vet-erinary Institute.
From this vantage point, Dr. Skrjabin gainedincreasing recognition for his concepts of thesignificance of helminthic disease in agricul-ture and public health. In the period 1920-1922 he organized or was instrumental infounding three centers of helminthological re-search: The Helminthological Department ofthe State Experimental Veterinary Institutewhich was later named the Ail-Union K. I.Skrjabin Institute of Helminthology in honorof its founder; the Helminthological Depart-ment of the Moscow Tropical Institute; anda Department of Helminthology at the MoscowState University. In addition to the organiza-tion of helminthological institutions in Moscow,he was the moving force in establishing thestudy of helminthology in research institutionsand universities throughout the Soviet Union.
Despite his administrative activities, Dr.Skrjabin found time to continue his morpho-logical and taxonomic studies. Beginning in1919, he organized and directed surveys ofthe helminth fauna of man and animals in theSoviet Union. Publication of these studies
together with his lectures on helminthologysoon attracted increasing numbers of students.Over 100 candidates have received doctoratesunder his guidance, and many more can becounted as students of his students. Amongthe former are a number of correspondingmembers of the Academies of Sciences of theUSSR.
Dr. Skrjabin's investigations in the field ofmorphology, biology, phylogeny, and syste-matics of helminths, on ecology, epidemiology,and epizootiology of helminthic diseases andtheir control have been reported in over 600publications. Now internationally knownthrough these publications, his contributionsto his country's scientific development as aneducator, mentor, and administrator have beeneven more remarkable. To list only some of thehonors accorded him in recognition of his ac-complishments, he has received honorary de-grees from Humboldt University (Berlin), andthe Institute of Agriculture (Czechoslovakia).He is a member of the Veterinary Academy ofFrance, a member of the Academies of Scienceof Bulgaria, Hungary, Czechoslovakia, andPoland, and an honorary member of the Amer-ican Society of Parasitologists (1932), theIndian Society of Helminthologists, Parasito-logical Society of Poland, and the RumanianParasitological Society. He is a member ofthree academies in the Soviet Union; Academyof Sciences of the USSR, Academy of MedicalSciences, and the All-Union Academy of Agri-cultural Sciences. He is president of the All-Union Society of Helminthologists and founderand Chairman of the Editorial Board of Hel-minthologia. Among other honors bestowedby his government are five Orders of Lenin,and on the occasion of his eightieth birthdaythe name of Hero of Socialist Labor.
The Helminthological Society of Washingtoncongratulates Academician Skrjabin on hisninetieth birthday, a milestone in a career thathas exerted a profound influence on all phasesof helminthology in the Soviet Union.
—FRANCIS G. TROMBA
Copyright © 2011, The Helminthological Society of Washington

MEMBERS OF THE H ELM INTHOLOGICAL SOCIETY OF WASHINGTON;.:'•: ' : ' X"-V> "•: ' / ; . i*\ ' C-' \ V ' •*** '•>*/ ^ "V''^' •" '
(/Alabama through Maryland; remainder of list will appear in July issue) '-•/
'"•\a /-*trv '^' '\\- Cairns, E. J.,- --0 ~
,•", ; -Ford, B.,R. : • ' • ; • ' - ••• : "*. Frandsen,J. "CJ-.-— .... .v
>;.Isensteinr-R. S.v ' •." I Rogers; W. A; r .,-
, - . - V Wellborn.'.T. L., Jr.^ ^Alaska
' ' -iRausch, R. •'-._ "• '; • - • ' •/ Shults, L. M. > f
Arizona V ' '• '• Anderson, G. A.
,""• : Nigh, E.CL., Jr.. . -• .Reynolds; H. W. N \
- '__ \VVilkes S.N. -' Arkansas * /' ft '^ ...
~- v - Slack, D. A, ; y '7,r California ' /
Allen, M. W.,Ash, L>R. - V
• -.. Baker;' N./F.\ .' , . ,.-•'•-,,"''.•. , Brown, R. L. ;L\' v
/ ' • • ) * • • Caveness, F. E. /i:, ,•••' •'.- Dailey,,' M..D. - ( ' - . .
, f • Hart, W. H: ' ,.-. . j•' Hasbrouck, IS. JR. .
Heyneman, ©."{- Holdeman, QJ L.. , '
' Y" : :Kinsella,J.M. ; .' ,:'. Rrassner,""S. M.
, Lamberti, vF.' -r > "1 Lear, B. - / , -.
->-', \, H. W; ->„.• /-."Loos, C. A. - \. .,•'•'•.'• , Maggenti, A. R. • ' :'S
'•• -.Mahkau, R.' • • f ', Martin', W. E. ::. "'-v . ' , ' McBeth, C. W;y , McGuirei W. G. ~ -
< ' ,;. Mizelle, :J,D.' .. '/•' Nahlias, F. M., - -
•"•Noffsiriger, Ella MaePaxman, G. A. <• -f
/' . Poinar,; G. O.^Jr. ^ /
'.-.'- .. •Ros^an^R. 'N. , /^v - Rothman, A. H.
Seh,-A/K.1% ''I,.''- . ' V - ': . \, S. A.
' V SiddiquH. ./A. V • ; "V ' v Steele, A. E.
•\..,;..'Van Gundy, S. D. -, ' • » 'i' . ' . • • . . ; Viglierchio, ;DrR.
Voge, Marietta • ' . . . T• ' '-^xr • / i? Ti ' ."'• •-•" '
- . ' WeiriYnann, ~C. J. ; '.Canal Zone
Walton, B..C.. Colorado ' ,TV
Olsen, O. W. 'tl Schmidt, G. D,'
Stable^, R.,M. . - ,oConnecticut . v \
' . Miller, P. M:; " " •' ' ,.-•^l- Penner, L. R . > r / / ..
Stoffolano, J. G.', Jr.• • • 'Delaware ': • ^ /-^ ,• < , Fielding, M. J. . ' ) (. ^ ' /D i s t r i c t of Colurnbio••- . I . Baisden,(L. A. , r-
1 v Betz, D. O., Jr. • ' - .'•' • r•:- , p ;BriggsrN. T. ' ',.v1 ; x , i ' .^ Buchheit, J. R.'
; - •• V-'\;Bufke, J. C. ' "\v : \.'' '
{ Crawley, Lila R. -'^ Draggan, S. ' ' ,"Duxbury, R. "El . r.Ericksonj D. G. ,•/Gonzalez, Stella M.Habermann, R. T. ''
'Hope, \V- D.Kenworthy, F. T.,. ; -
: Lincicome, D. R. ( 'Martin, L. K.
* Oh'vier, Li J. /Radke, M. G. - .^ledington, B. C.Sadun, E. H. fSlie, RitaB.Wells, Gladys A./: i.
Florida -- ~-< \,Christie, J. R.Esser, R. P. ' . 'Hahnon, C.xL- T '''<-'Kincaid, R. R.
• . , O'Bannon, JVH.> N Overman, Arnegda J.\, V. G.
Short, 'R. B. .Smart, G. C., Jr. ;/Tarjan, A. G. .~~~-/Taylor, A. L.'
GeorgiaBird, G. W,. '
^ '• Ciordia, H. > (J'iCross, Hi F. x ;Denton, J. F.~
;. .Eni, E.-U. r! '£• Goodchild, C. G. "..Heara, R. W., Jr.
Kaganj I. G. i , ' /Marti, O. G., Jr.Minton, N. A- \ '•Ruehle, J.;L:/ /
S, -Stewart, T. B. , -;v xSullivan, J. JV";f Wehunt, E. J.Hawaii '>• f •-. / • v-''r '''
Alicata,/J. E.Apt, w. j. / • rCheng, :T. C.- iGoto, S. cLee,S.H.' " . . » : .Rifkin, E. , " -
Idaho , ." , , , '\' •. •/' •£•'"- vDalliinore,1 G.. E.Illinois - '
Amosuy Ov , , •, Bauman, P. M, •. '•
Duuagan, T,\T; .Edwards, D, I.
v 'Garoiah, G.Huizinga, H- W- _
, Kruidenier.^F. 7-Levine-.-N': D. i
\, R. B.1McCrae, R. C.' , ,Qstdiek, J. L. ; , ,Pillai, J. K. , -Schneider, M. D. :Seitaer, P:iG, •- -" •Singer, I. , ---•' ' . ?•-Taylor, D. PS !Thomas, L. J,Todd, K. S., Jr./K ;
Indiana ' , ' \, R.'J.
' Cable, R. M. . f ; •
::: Ehrenford, F. A.-, -f'-Ferris, Virginia R. . s
X > \. F.Thorson, R. E. ':. . ,
- Weinstein, P. P. , - .,. ,/'' IqWo . J- /'.,'; , ;
' " Blankespoor, H. D.; -1 Ellis, C. J.
.' 'Epstein, A. H. - ' .Goss, R. C. .... '-:'•'
i Greve1, J. H. . ,'>Hsu, H. F. - ^ ^
>. Norton, D. C./" '',':J iu\mei, M. j. -.>•;,:, . , Williams, D.:.D.
-; ', Ziminermann,' W. J. --H , Kansas': ;; / • ..^__',-'
v, : • Ackert, Ji E.,•Ameel, D. Jj
•" ' Ay tan, Songul V ( . - ~" Cml,-W. H7- - ; • ' / " %
. i S . Dickerson, O. J. \ J' . :Haiiseh, M. F. ^ '/•
•• \y " / _ \V'' .Chapman, R: A, ~ •
prudge.J. H. v:v~ /Eversmeyer, H- "EiS
'. > Lyons; E. T. .;•:;. •-.'' .. ; '/Whittaker, F.H..
/ : Louisiana ••' • • -• • ' '.'{Acholomi, A. D.
• • : ''-. ' Bello, T. R. -- ;• ' i.Besch, E. D. . ' ,W
)- Birchfield, W.v .- ' •__"', Hollis; J. P.* Jr. - ; r- ' > Lumsden, R. .D. -?.'
•' Martin', W. J. , 'vNewsom, L. D.
v ., Overstreet, R. M- .; SogandaresrBernal, F;
;. " Warren, ;,L. G. /'" .Jfamaguti, S^ i , . /.
5>v,' Maine ' \ \j-' ' *'-"•{.,' Meyer, M. C." . '•': . . . ,'Maryland -. -, , ' ••fkV\ '-'
, V *. Abram, J. B. : { • v' . . • r Anastos,^G. , ' ' - • -N'. '
/..f -Anderson, R. I. ( \ \ .•Andrews, J. S. . <-. .-
(Bang^F. B. ^_Beaudoin, R. L. -r:^Becklund, W;,W. ->Bergquist, E. J^-_ •• -;•
': - 1 vori Brand, 'T. ' /''- "Buhrer, Edna M.
. s i _ v ;Chitwood,; May B.^-- x'••- 'Gblglazier, M. L.
\, W.-B. -' " . . . . ."< ' . ' . - . • • Diamond, L. S. ' V
'Doran, D. J. '- i-Doss, "Mildred A.' V'
''. ,Douvres,>;F. W. •*- : , Dropkin, V.JH. ,
'•'- Durbin, C::G.'-'• /'Edwards, Shirley J. , /
IS vEndo, B. Y.. r\''i :| Enzie, F, D. A"
' 7,\, 'JMarion M. ,v >' .' , '- - - ; • , l.iFayer.iR. --'';-., ' :..,
•A Feldmesseri, J. •Ferguson, M..S.' '• :'x~
cFoster; A. O. '•* Friedman, W:
• :' Goldberg, A. / :,;i •,. .:
'"-" .-° Golden, AllM.. • . , ' . " • . ., V. ...;i-Good, J.vM.,'Jr. ,
Haley, A. T. y ,Hechler, Helen Carol
^Herlicb.H. V '• ;Herman, C. 'M-^Hernandez, A. - -V •Hill, C. K .Holbrook, 'A. A. 'Huff.C.G.Humphrey, Judith M. ; ~,f
Jachowski, L. A., Jr. ,acobs, L. ;
Tahnes, W. G.Kates, K. C. +
•/• Xriight, R. A. : . ;Krusberg.'L. R. ..... . ' .'
f Lawless, D. K. - 'Leek, R. G. >./Lesser, ,E. ^Lichtenfels, J. R. V JLotze, J.G. ' ''.Lund, E. E. , , ' • / .Lunde, 'M. N: v ' , -Lwtterrnoser,-G. W.
, Malakatis, G. M. SMarchbank, Dorothy 'F.;McIntosh, A. ' -
> ;- McLoughliri, D. K. . 'MorriS, J. A. .:-. .;Murpiy, D. G. "
1 Nickle, ;W. R. ' • - . , ' , • •Osborne, H. SV
; ; OtWi, G. F. . :.-.-1 ; : -Pachecd, Gi . •
i Phillips, B. P. , ^r IPipkin, .A; :C.-" -- ' ' ;
Poiley, Judith A. '- Poole, J. B. -;:Powcrs, K. G. ;
>x ,#rice,:D>L. --' ... : ' ' :.~1 vQuigley, Diane ' •
/ Bichards 'C.NS., ^ . ;. Robinson.'-Marma M.V Roby, T. O. T
Rogers, S.;H.yViRomanowski, R. D. /
V , , - - Rosei J. E:j , Rozeboom, L. E. -^
' Sa%vyer, T. K-.Sayre, R. M. , • : '' •;.Schiller,. E..L. ,,, '"(" '. ';'Schmunis, G. A- " ; • •"• ' \, C.M. t'
) Scott, J. A., ...\r Dorothy B- >
^ x Sheffield, H. G.: Shiroishi, Tsugiye;', . -"
Solomon, G. B.'. N /, ••'/ . Spindler, L. A. ;
Sprague, V. ', '•'iStifewalt, -Margaret A.Stringfellovv, F.Tarshis, L.JB'. . , ' ' . ;
j . Terziaiii 'L. A. '•••'- Traub,^. /•*•£/"'(. TrorribarF.;G. .
7 Turner, J. H. • ' • ' , , .'Undervvood, P. C;Vegprs, Hi H.
'"' )Wehr,E..E-. • \*V.' •, Williamson, ;F.' S. %.^ , : -'-
•Wiseman, ;H.-G. /.
Copyright © 2011, The Helminthological Society of Washington

i r
">')••-,
\VOLUME 36 1969 NUMBER .1
C'/./' •/ •'"^••'^.'••y -' *- .,". -if"' ' • , ; . CONTENTS v . , . . . , T,
'••;^y '.•;•'- ';'.' — •.'/ ' '-• ' '• ' '<pT1'' • • •'" (Continued from Front Coyer} X •1,x-*:^-' '•'••;',/''••:-. ~ : 'v,
FiscHTHAL, JACOB H/ Euparddistomum cercopjthepi sp. n. (Diprbebeliidae), ;af Digenetict Trematode from the Talapoin /Monkey fjrom 'Rio Muni -;.r_-..^i,._.I—;._^i±--r-;-------•>--%
FRIED, BERNARD,-AND RONALD E. TOKSTWALL// Survival arid Egg;Laying( of4T\urtle, >>Blood Flukes (Trernatoda: Spirorphiidae) on $e'jChick Chorioallantois _^.^r-_.:-_..FRIED, BERNARD, AJSTD LINDA J, WEAVER. —Exposure of Chicks to the Metacercafia of
v^ Echinostpma/revolutum'^ Trernatoda) •^^.^.:..^l.:....^:^..^.--.^^..^,......-.^.-..-.^.,..:...GRIFFIN/ G. D., AND: E. C.' JoRdENSON. Pathbgenipity of ;the Northern RoptTknot
Nematbde/ (Meloidogyne hdjila) to Potato _:-^—__--7—7-t.—,-i „—;::. _k 1* —HENDRICKS, LAKRY D., REINARD HABKEMA^ AND GROVER C. MILLEB. jHelminths/of the
Crow, Cbrvus brachyrJiynchos Brehm, 1822, in Nor^i Carbh'na —-I'-^ll. i.;.-...J—..„_..HOPE, W. DUANE. -Fine Structure of the Somatic; Muscles of the Free-Living Marine
cyVj . Nematbde peontostpmq.californicum Steiner and Albin, 1933 '.(Leptosomatidae).,_.....'' Iri .Memoriam .^^.__^= ;i-—--~:-7--"7"f-—---—~l~~~:-l-r:VJ-^_^._L._'Jl'r-.._^—_ --,-..KN6BLpcH",xNATALjE A., AND JOHN A;^^ KNiEHiM. Tylenchus vesjculosus, sp. n/ (Nema-
toda: Tylenchidae) from "Soil, in 'Michigan^-:--l-:-----r_^r----^ —-.::-KRYGIER, B; BRU^E, AND RALPH W. MACY, JZis^orchis'heterbrchissptn.j( Trernatoda:
i: Lissorchiidae^ from Cytbstomus mactocHeiliis Giriard in Oregon „_„,„.^ -„„__;.:_LEE, SHERIDAN-H. .'/The lUse of Irradiated ;Third-stage Larvae- of Angiostrongylus
^( (cantonensis as Antigen tblrnmiinize. Albino'Rats Against Homologous Infection'"^:....MACKIEWICZ, JOHN S. Pjenqrchigeies okhiteis gen. et sp. n. and jBiefeetabulum'carpiodi
:' sp. ii. "(Gestoidea: Caryophylla^eidae) ^frorn Gatostomid Fish, in Islortii America,-,..—MACKIEWICZ, JoHN:qS.,: ANIJi AHTHXIB, W.^ JONES, ^fhe Clirbmosomes -of Hunterella
/ nodulosa Matkiewicz iand McGrae, 1962 (Gestoidea: Carybphyllidea) ........jr. ... ....MASSEYy):CALyiN <L. - New .Species of Tylenchs/Associated witli Bark Beetles in New
Mexico and Colorado :nrf,l. jI-^J.i:-._..J-..—..._... : :_._, :^.....^.7 .„...: u..._:.l.. .pa, MAHGOS 'TiHJO IJiAZ;; Luis J. HAMANA SALAZAR, AI^D S. GUEVARA. Studies
on Freshwater Larval Trernatbdes. XXI. Two -'New Species ,of iMacrocercous
G. ^Studies on Haplobothnuni bistrobilae $p. ;n(Jv. (C^stoda: PseudophylLlidea) <frorn •Amfa. calva L/,-^—::.. -.—..„ .._...__-,—4—v—"---^------^--^--^----"—----'
G/ A New Trematode -Cephjalogoriimus sireni sp. nov. r(Digenea:, Cepha-logbnimidae) from Florida: Mud/Eel, .Siren lacertina ... .4. ..—----- ----—--•,------
"C. E., AND'/A. ,>MURA. The Proposed- Synonymy of !vthb Mohogenean^ Genera'. - Cleidodiscus Mueller, 1934 and tlrocleidus Mueller, -1934, with the Proposal o f ,
^ Cleidoldisci/s .fo^c/ioipsfcVMP^1 n-',.--...^--—-—- .1--...—-,-,-i-.I—^-r-~^-^~-——il-,_-i.RASKj;D.!,,J., N, O. JONES, AND DVTl. ROGGEN. " On the Morphology and Ultrastructure
.x ;of the Esophageal Region of Tric)i()dorus allius Jensen ' '....—....V.^-1——.:....:._.....^-.GROVEPI C., JR.,' AND H. RC TH:O^AS. Survival oL Eggs and Larvae in Cysts of
the 'Soybean* Cys,t Neniatode, Heterodera gli/cines^lngesied by Swine; 1J.........—..ITHOMAS M., ^VILLIAM A. SODEMAN, JR., AND CHARLES S. RICHARDS. The
, - 4 Intrapulmohary Lbcalizatioiivof Angiostrongylus cqntonenisis in the Rat'.... ...—;....THORNE, -GERA-Lp- : Hypsoperine ottersoni sp.^-n. (Nernata, . Heterpderidae) Infesting
— < Can,ary Grass,'J?halaris "arundinacea (j^.~) Reed in Wisconsin ..-:...—:... -. .„.!..„:...........TIMM, R. W. ; Megadohtolaimys New Genus (Nematoda: Chromadoridae), 'witii a
- Description of Two ;New Species'—..:.._-- —..._;—--:------------,-—--'-——-------— -TROMBA:; FRANCIS G; Academician K. I. Sfcrjabin-—r^ihetieth Birthday _. .....;............._.
• TSAI, YuAN-HwxVNG,-AND ALBERT W. GRTJNDMANN; Reesimermis nielseni gen. et sp. n./'--''.-'• , (Nematoda: M.erniitliidae") /Parasitizing Mosquitoes'/in Wyoming /_!:.._„.-__—_.,..——-WHITTAKER, (FRED H. Galvanotaxi§ o'tPelodizra strongyloides (Netnaloda:' Rhabditidae,)/
[. C., AND I. Nj^VTON.l—tntestmal Helminths of tlie ^Bullfinch, fynliulapyrrjiula (L.), in)Southern England/ ..L.iv— I..;—---^------'---- -^- .-.,.-1-.——r 1\: V i . - -.,-• '' - ; ' ' ' ' : "" . . . . ANNQUNPEMENTS \ \_ .^ : ' : '
Businejss Offic^,—-.—.—-.———_.._;^.-.i-._.-_^:_...—-i.-r-.-.L--.-.::^LNew' Journal'- ,C---- —-•- 1T,-^ . ^ -_..ji.—..._.:..l—.-.Second International Gongress of Parasitolqgy. -—_.——_- —......-,\J rSj/V1 '\ •• -•- X; • '' ^ ...-":'*i^'''1* • • .'*" ,'-v' - - . - . -, • ,
' 1 :r/!;H .;," lx>-; Date of Publication^SOJaWary/1968
:'fcX:
183'
V
10
147
136
119
126 -
43
^102
55
74
!52,
106
139
143
98\:36(156
.61\0
76
146149152
•i(:ALLEN PRESS) INC. .LAWRENCE, KANSAS
Copyright © 2011, The Helminthological Society of Washington




















