A Review on Pressure Ulcer: Aetiology, Cost, Detection and Prevention Systems

FEMS Immunology and Medical Microbiology 41 (2004) 161–167
www.fems-microbiology.org
The Helicobacter pylori plasticity region locus jhp0947–jhp0949is associated with duodenal ulcer disease andinterleukin-12 production in monocyte cells
Ramon de Jonge a,b, Ernst J. Kuipers a, Sabine C.L. Langeveld a, Ruud J.L.F. Loffeld c,Jeroen Stoof a, Arnoud H.M. van Vliet a, Johannes G. Kusters a,*
a Department of Gastroenterology and Hepatology, Room L-459, Erasmus MC-University Medical Center Rotterdam,
Dr. Molewaterplein 40, 3015 GD Rotterdam, The Netherlandsb Department of Gastroenterology, VU University Medical Center, Amsterdam, The Netherlandsc Department of Internal Medicine, de Heel Zaans Medisch Centrum, Zaandam, The Netherlands
Received 23 December 2003; received in revised form 12 February 2004; accepted 2 March 2004
First published online 9 April 2004
Abstract
Colonization with Helicobacter pylori always results in chronic gastritis, which is controlled by infiltration of mononuclear cells
and the subsequent release of cytokines like interleukin (IL)-12. To identify H. pylori factors involved in inducing cytokine pro-
duction in mononuclear cells, a random H. pylori mutant library was screened for the inability to induce IL-12 production in
monocyte THP-1 cells. Of the 231 random mutants screened, one mutant (M1) showed a consistent twofold decrease in the amount
of IL-12 induction compared to the parental strain 1061 (P < 0:01). Further characterization of mutant M1 revealed that the
kanamycin resistance cassette had integrated in the jhp0945 gene, which is situated in an H. pylori strain-specific plasticity region.
Three reference strains possessing this plasticity region induced significantly higher amounts of IL-12 when compared to the
H. pylori 26695 reference strain, which does not possess this plasticity region. The role in disease outcome of jhp0945 as well as the
neighbouring plasticity region genes jhp0947 and jhp049 was assessed in a Dutch population cohort. Firstly, the presence of jhp0947
was completely linked with that of jhp0949 and was roughly associated with jhp0945 (P ¼ 0:072), but not with the cag pathogenicity
island (PAI) (P ¼ 0:464). The presence of the jhp0947 and jhp0949 genes, but not of jhp0945, was significantly associated with
duodenal ulcer disease when compared to gastritis (P ¼ 0:027). Therefore, the jhp0947–jhp0949 locus may be a novel putative
H. pylori marker for disease outcome independent of the cag PAI.
� 2004 Federation of European Microbiological Societies. Published by Elsevier B.V. All rights reserved.
Keywords: Helicobacter pylori pathogenesis; Cytokine; Virulence factors; Plasticity region; IL-12
1. Introduction
Colonization of the human stomach with Helicobac-
ter pylori leads to chronic gastritis, which may further
complicate into peptic ulceration, atrophic gastritis and
gastric adenocarcinoma [1]. Colonization with H. pylori
is also characterized by massive infiltration of poly-
morphonuclears (PMNs) and mononuclear cells, which
* Corresponding author. Tel.: +31-10-463-2982; fax: +31-10-463-
2793.
E-mail address: [email protected] (J.G. Kusters).
0928-8244/$22.00 � 2004 Federation of European Microbiological Societies
doi:10.1016/j.femsim.2004.03.003
are hallmarks for inflammation. In addition, increase of
the local production of pro-inflammatory cytokines likeinterleukin (IL)-8, IL-12, IL-1b, TNFa and IFN-c has
been reported in vivo in H. pylori-positive patients, as
well as in vitro using H. pylori stimulated epithelial and
mononuclear cells [2–6].
The cytotoxin-associated gene (cag) pathogenicity is-
land (PAI) was the first H. pylori factor for which a role
in host cell cytokine production was demonstrated [7].
The cag PAI is present in 50–70% of H. pylori isolatesfrom Western populations, and in 90% or more of the
strains in Eastern populations [1]. It plays an important
. Published by Elsevier B.V. All rights reserved.

162 R. de Jonge et al. / FEMS Immunology and Medical Microbiology 41 (2004) 161–167
role in the induction of IL-8 production in gastric epi-
thelial cells [7]. Although IL-8 is an important factor of
the innate host immune response during initial H. pylori
colonization, it is not required for development of the
pro-inflammatory T helper 1 (Th1) immune response.Proliferation of naive Th lymphocytes towards the
inflammatory Th1 cells, which are abundantly present
during H. pylori colonization [8], is regulated by the re-
lease of IL-12 by mononuclear cells [9]. Although IL-12
has recently been shown to be essential for protection
againstH. pylori [10], prolonged production of IL-12 and
thus activation of the pro-inflammatory Th1 immune
response is associated with the development of pepticulcer disease [11]. Although we have previously demon-
strated that H. pylori mediated IL-12 production in
monocyte cells is not dependent on the cag PAI [5], many
of the H. pylori genes that play an essential role in IL-12
production by mononuclear cells are still unknown.
In a previous study we described a technique for
random direct insertion mutagenesis in H. pylori [12]. In
the present study, a set of such H. pylori random mu-tants has been evaluated for their ability to induce IL-12
production in monocyte cells. Subsequent analysis
revealed that integration of the kanamycin resistance
cassette in the H. pylori strain-specific plasticity region
jhp0947–jhp0949 gene resulted in a significantly de-
creased production of IL-12. Subsequently, the presence
of jhp0945, jhp0947 and jhp0949 in a set of H. pylori
isolates obtained from a well-characterized Dutch pa-tient population cohort was assessed. This showed that
the presence of jhp0947 and jhp0949 is significantly
associated with development of duodenal ulcer disease.
2. Materials and methods
2.1. Culture conditions
The H. pylori cag-positive strains J99, 26695, G27
[13], the cag-negative strain 1061 [14], and a set of 231
random mutants derived from a random H. pylori 1061
mutant library [12] were routinely cultured on Columbia
agar plates supplemented with 7% lysed horse blood and
Dent selective supplement (Oxoid, Basingstoke, UK),
further referred to as Dent plates. When appropriate,Dent plates were supplemented with 20 mg l�1 kana-
mycin. Broth cultures were grown in Brucella broth
supplemented with 3% Newborn Calf Serum (Invitro-
gen-Life technologies, Paisley, Scotland) to yield
approximately 108colony forming units (cfu) ml�1.
H. pylori was routinely grown for 24–48 h at 37 �C in an
atmosphere of 5% O2, 10% CO2, and 85% N2.
Human monocytic THP-1 cells were cultured inRPMI 1640 medium containing GlutamaxI supple-
mented with 10% fetal calf serum (Gibco BRL, Paisley,
Scotland) (RPMIþ). Cell suspensions were seeded in 24-
wells microtiter plates (Costar, Cambridge, MA) and
incubated at 37 �C in a 5% CO2 incubator until a cell
density of 106 cells ml�1 was obtained. Subsequently, the
cells were washed with phosphate buffered saline and
harvested by centrifugation at 370g for 10 min. Forsubculturing, cells were diluted in RPMIþ to a final
concentration of 105 cells ml�1.
2.2. H. pylori mediated IL-12 production in THP1
monocyte cells
Wild-type H. pylori 1061 and the 231 random
H. pylori mutants were screened for their capacity toinduce IL-12 production in monocyte THP-1 cells, using
an assay as described previously [5]. THP-1 cells (4� 105
cells ml�1) were incubated for 24 h after which the cells
were stimulated with H. pylori (bacteria: cell ratio 100:1)
for 24 h. As a negative control, THP-1 cells were stim-
ulated with H. pylori for less than 5 min. The amount of
the IL-12 p40 protein in cell culture supernatants was
determined by sandwich ELISA (BioSource Interna-tional, Nivelles, Belgium). The lower limit of detection
for IL-12 was 50 ng l�1. Only those mutants which
showed a decrease in H. pylori mediated IL-12 produc-
tion of >30% when compared to the wild-type strain
1061 in at least three independent induction experiments
were included in subsequent stimulation assays.
2.3. Characterization of the integration site of the
kanamycin resistance cassette
Determination of genes affected by the integration of
the kanamycin resistance cassette was essentially per-
formed as described previously [15,16]. Genomic H. py-
lori mutant and wild-type DNA was digested with CfoI
(0.5 U ll�1) and specific CfoI linkers (2 pmol ll�1; Table
1) were ligated with 0.15 U ll�1 T4 ligase (Promega,Leiden, The Netherlands) with addition of 0.1 M NaCl
and 100 ng ll�1 BSA at 37 �C for at least 2 h. Subse-
quently, PCR experiments using a combination of CfoI
adapter primers (0.5 pmol ll�1) and primers specific for
the kanamycin resistance (kanR) cassette (0.2 pmol ll�1)
were performed to selectively amplify the genome frag-
ment containing the kanR cassette. Fragments were am-
plified in PCR buffer containing 3 mM MgCl2 and 0.02U ll�1 Taq (Promega) in 40 cycles at 95 �C for 30 s, 65 �Cfor 30 s with a decrease of 1 �C per cycle to a minimum of
55 and 72 �C for 45 s, using the primers listed in Table 1.
The resultedPCR fragmentswere cloned into pGEMTeasy
(Promega) and subjected to sequence analysis.
2.4. Identification of the plasticity region genes and the
cag PAI in H. pylori isolates
Chromosomal DNA was purified using standard
protocols from a total of 45 H. pylori clinical isolates
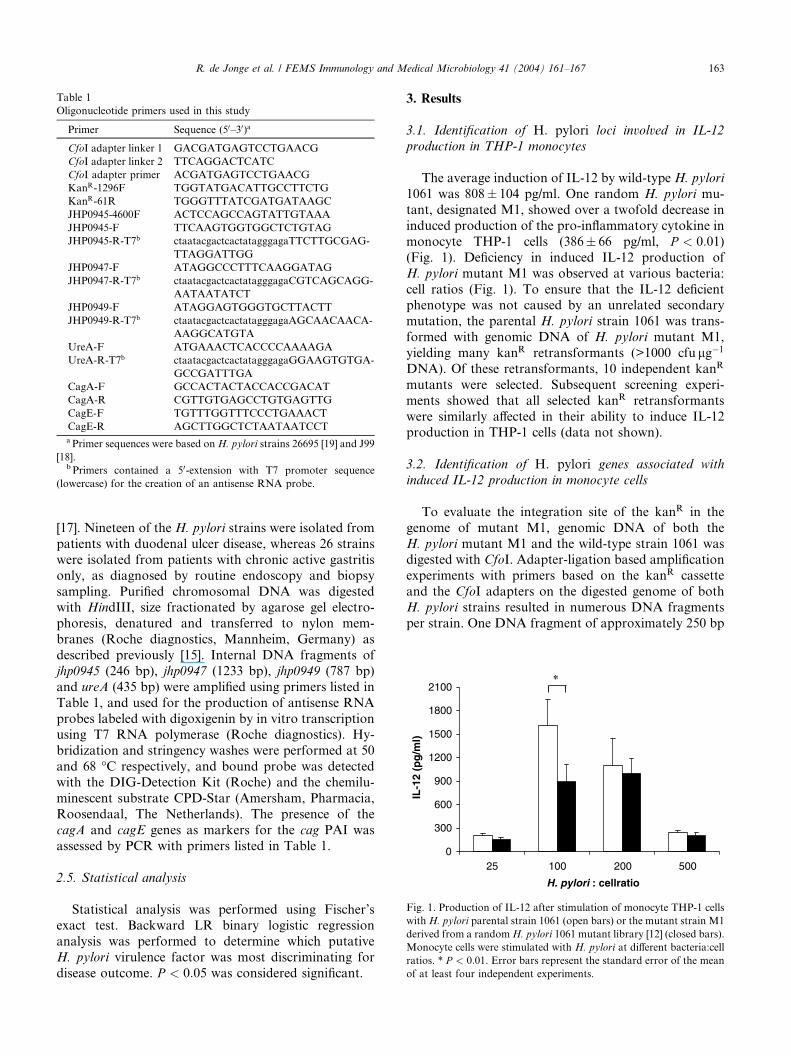
Table 1
Oligonucleotide primers used in this study
Primer Sequence (50–30)a
CfoI adapter linker 1 GACGATGAGTCCTGAACG
CfoI adapter linker 2 TTCAGGACTCATC
CfoI adapter primer ACGATGAGTCCTGAACG
KanR-1296F TGGTATGACATTGCCTTCTG
KanR-61R TGGGTTTATCGATGATAAGC
JHP0945-4600F ACTCCAGCCAGTATTGTAAA
JHP0945-F TTCAAGTGGTGGCTCTGTAG
JHP0945-R-T7b ctaatacgactcactatagggagaTTCTTGCGAG-
TTAGGATTGG
JHP0947-F ATAGGCCCTTTCAAGGATAG
JHP0947-R-T7b ctaatacgactcactatagggagaCGTCAGCAGG-
AATAATATCT
JHP0949-F ATAGGAGTGGGTGCTTACTT
JHP0949-R-T7b ctaatacgactcactatagggagaAGCAACAACA-
AAGGCATGTA
UreA-F ATGAAACTCACCCCAAAAGA
UreA-R-T7b ctaatacgactcactatagggagaGGAAGTGTGA-
GCCGATTTGA
CagA-F GCCACTACTACCACCGACAT
CagA-R CGTTGTGAGCCTGTGAGTTG
CagE-F TGTTTGGTTTCCCTGAAACT
CagE-R AGCTTGGCTCTAATAATCCTaPrimer sequences were based onH. pylori strains 26695 [19] and J99
[18].b Primers contained a 50-extension with T7 promoter sequence
(lowercase) for the creation of an antisense RNA probe.
0
300
600
900
1200
1500
1800
2100
25 100 200 500
H. pylori : cellratio
IL-1
2 (p
g/m
l)
*
Fig. 1. Production of IL-12 after stimulation of monocyte THP-1 cells
withH. pylori parental strain 1061 (open bars) or the mutant strain M1
derived from a randomH. pylori 1061 mutant library [12] (closed bars).
Monocyte cells were stimulated with H. pylori at different bacteria:cell
ratios. * P < 0:01. Error bars represent the standard error of the mean
of at least four independent experiments.
R. de Jonge et al. / FEMS Immunology and Medical Microbiology 41 (2004) 161–167 163
[17]. Nineteen of the H. pylori strains were isolated from
patients with duodenal ulcer disease, whereas 26 strains
were isolated from patients with chronic active gastritis
only, as diagnosed by routine endoscopy and biopsy
sampling. Purified chromosomal DNA was digestedwith HindIII, size fractionated by agarose gel electro-
phoresis, denatured and transferred to nylon mem-
branes (Roche diagnostics, Mannheim, Germany) as
described previously [15]. Internal DNA fragments of
jhp0945 (246 bp), jhp0947 (1233 bp), jhp0949 (787 bp)
and ureA (435 bp) were amplified using primers listed in
Table 1, and used for the production of antisense RNA
probes labeled with digoxigenin by in vitro transcriptionusing T7 RNA polymerase (Roche diagnostics). Hy-
bridization and stringency washes were performed at 50
and 68 �C respectively, and bound probe was detected
with the DIG-Detection Kit (Roche) and the chemilu-
minescent substrate CPD-Star (Amersham, Pharmacia,
Roosendaal, The Netherlands). The presence of the
cagA and cagE genes as markers for the cag PAI was
assessed by PCR with primers listed in Table 1.
2.5. Statistical analysis
Statistical analysis was performed using Fischer’s
exact test. Backward LR binary logistic regression
analysis was performed to determine which putative
H. pylori virulence factor was most discriminating for
disease outcome. P < 0:05 was considered significant.
3. Results
3.1. Identification of H. pylori loci involved in IL-12
production in THP-1 monocytes
The average induction of IL-12 by wild-type H. pylori
1061 was 808� 104 pg/ml. One random H. pylori mu-
tant, designated M1, showed over a twofold decrease in
induced production of the pro-inflammatory cytokine in
monocyte THP-1 cells (386� 66 pg/ml, P < 0:01)(Fig. 1). Deficiency in induced IL-12 production of
H. pylori mutant M1 was observed at various bacteria:
cell ratios (Fig. 1). To ensure that the IL-12 deficientphenotype was not caused by an unrelated secondary
mutation, the parental H. pylori strain 1061 was trans-
formed with genomic DNA of H. pylori mutant M1,
yielding many kanR retransformants (>1000 cfu lg�1
DNA). Of these retransformants, 10 independent kanR
mutants were selected. Subsequent screening experi-
ments showed that all selected kanR retransformants
were similarly affected in their ability to induce IL-12production in THP-1 cells (data not shown).
3.2. Identification of H. pylori genes associated with
induced IL-12 production in monocyte cells
To evaluate the integration site of the kanR in the
genome of mutant M1, genomic DNA of both the
H. pylori mutant M1 and the wild-type strain 1061 wasdigested with CfoI. Adapter-ligation based amplification
experiments with primers based on the kanR cassette
and the CfoI adapters on the digested genome of both
H. pylori strains resulted in numerous DNA fragments
per strain. One DNA fragment of approximately 250 bp

164 R. de Jonge et al. / FEMS Immunology and Medical Microbiology 41 (2004) 161–167
was markedly present in the digested genome fragment
pool of mutant M1, but was absent in that of the wild-
type strain. This DNA fragment contained the 50 end of
the kanR cassette as well as a region with no obvious
differences in amino acid sequence to the jhp0945 gene ofH. pylori strain J99 [18]. Disruption of jhp0945 by the
kanR cassette in H. pylori mutant M1 was confirmed by
additional PCR experiments on genomic DNA of H.
pylori mutant M1 using primers specific for jhp0945 and
the kanR cassette.
The H. pylori strain specific jhp0947–jhp0949 locus
is part of a region of 38 open reading frames
Fig. 2. Genetic variability of the jhp0945–0947–0949 genes in H. pylori clinic
the jhp0947 gene and flanking regions of the H. pylori J99 strain. The vertica
(ORFs) of strain J99 are indicated by arrows and numbered sequentially, be
pylori strain 26695 is indicated below the number of the J99 ORF. The shaded
RNA probes for Southern blot hybridization. (B) The genetic variability of th
isolates as determined by Southern hybridization. Hybridization with the ur
experiment is shown here. The positions of relevant markers are shown on t
(Fig. 2A), which has been termed a plasticity region
[18,19] and is present in H. pylori strain J99 [18], but
not in strain 26695 [19]. To further substantiate a link
between the plasticity region and IL-12 production in
THP-1 cells, we determined the ability to induce IL-12of the plasticity-region-positive H. pylori reference
strains J99 and G27 and the plasticity-region negative
strain 26695. H. pylori J99 and G27 induced high
levels of IL-12 production (954� 124 pg/ml and
773� 21 pg/ml IL-12, respectively), whereas H. pylori
26695 was only a poor IL-12 inducer (299� 16 pg/ml
IL-12).
al isolates. (A) Genetic organization of the plasticity region containing
l line indicate the end of the plasticity region. The open reading frames
low the arrows. The corresponding homologous gene, if present in H.
fragments show the DNA fragments used for development of antisense
eH. pylori genes jhp0945, jhp0947 and jhp0949 of eight clinicalH. pylori
eA gene is included as positive control. A representative hybridization
he right, probes used are indicated on the left.

R. de Jonge et al. / FEMS Immunology and Medical Microbiology 41 (2004) 161–167 165
3.3. The presence of the H. pylori jhp0947 and jhp0949
genes is associated with duodenal ulcer disease
The presence of the plasticity region thus correlated
with IL-12 induction. Previous studies have suggestedthat such an induction in vivo correlates with more se-
vere disease outcome [11]. Therefore, we studied the
association of the H. pylori plasticity region genes
jhp0945, jhp0947 and jhp0949 with disease outcome in a
Dutch population cohort [17], using Southern hybrid-
ization with jhp0945–0947–0949 specific probes (Fig. 2).
Hybridization with the ureA gene was used as positive
control, while a cag PAI-specific PCR used to confirmthe association between the cag PAI and disease out-
come, and to determine the association between the
plasticity region and the cag PAI. Fig. 2B shows a rep-
resentative example of the Southern hybridizations used
in our analyses.
All H. pylori strains tested positive for ureA. The cag
PAI was present in 95% of the duodenal ulcers strains,
while only 68% of the H. pylori strains isolated fromgastritis patients were positive for the cag PAI
(P ¼ 0:056). The plasticity region genes jhp0945,
jhp0947, and jhp0949 were present in 40%, 33%, and
33% of all H. pylori strains tested respectively. The
presence of the jhp0947 gene was completely linked with
that of jhp0949 and was roughly associated with jhp0945
(P ¼ 0:072), but not with the cag PAI (P ¼ 0:464). Theplasticity region gene jhp0947 was present in 19% and53% of H. pylori strains isolated from patients with
gastritis and duodenal ulcer respectively (P ¼ 0:027),while jhp0945 was present in 37% and 42% of duodenal
ulcer and gastritis strains respectively (P ¼ 0:767) (Table2). Both jhp0947 0949 and the cag PAI were present in a
single strain in 25% of the H. pylori population, and this
was highly associated with the presence of duodenal
ulcer (P ¼ 0:0045). Binary logistic regression analysis,which was performed to determine the most discrimi-
nating H. pylori factor in disease outcome, showed that
both the presence of jhp0947 and jhp0949 (P ¼ 0:014) aswell as the cag PAI (P ¼ 0:023) were significantH. pylori
Table 2
Association of the presence of H. pylori plasticity region genes with
disease outcome
Disease
outcome
No. of
patients
jhp0945
N (%)
jhp0947–jhp0949a
N (%)
Duodenal ulcer 19 7(37) 10(53)
Gastritis 26 11(42) 5(19)
Total 45 18(40) 15(33)
P-valueb 0.767 0.027a The presence of jhp0947 and jhp0949 is completely linked in all
H. pylori isolates tested.b P value indicating the relation between the presence of the re-
spective gene and duodenal ulcer disease, as determined by Fisher’s
exact test. P < 0:05 were considered significant (bold).
predictive factors for duodenal ulcer when compared to
gastritis.
4. Discussion
Colonization with H. pylori induces an influx of im-
mune cells into the gastric mucosa, and subsequent
production of IL-12 and other cytokines. IL-12 polar-
izes naive T-cells to the pro-inflammatory Th1 type, and
prolonged activation of the Th1 immune response may
result in severe tissue damage such as peptic ulcer dis-
eases [9,11]. Colonization of H. pylori is strongly asso-ciated with increased IL-12 production by mononuclear
cells. Indeed, we have previously demonstrated that
many H. pylori strains are able to induce IL-12 pro-
duction in monocyte cells and that this occurs inde-
pendently of the H. pylori cag PAI [5]. However,
H. pylori factors that play a role in H. pylori mediated
IL-12 production in monocyte cells are still largely
unknown.In this study we show that disruption of the jhp0945–
0947–0949 locus in H. pylori strain 1061 significantly
decreased its ability to induce IL-12 production in
monocyte THP-1 cells. While we cannot exclude the
possibility of polar effects, we have characterized the
presence of three large genes within the jhp0944–jhp0950
locus in such a way that almost the entire locus was
covered (Fig. 2A). In addition, the ability to induceH. pylori mediated IL-12 production was completely
associated with the presence of the jhp0945, jhp0947, and
jhp0949 genes in the high IL-12 inducing H. pylori
strains J99, G27, 1061 and conversely the absence of
these genes in the low IL-12 inducing H. pylori strain
26695.
The jhp0947 gene is homologous to jhp0938 (hp0990)
and jhp253 (hp1333), which all encode for hypotheticalproteins. Interestingly, the N-terminal site of JHP0947 is
also homologous to that of JHP0477 (HP0528), which is
part of the cag PAI and has been identified as an im-
portant building block for the type IV secretion system
encoded by the cag PAI [20]. Similar to jhp0477, jhp0947
is part of a region of numerous ORFs, which has been
identified as a plasticity region (jhp0914–jhp0951) [18,19].
Plasticity regions are often associated with increasedvirulence, and the jhp0947 gene has already been pro-
posed to be a novel virulence factor of H. pylori, albeit
not independent of the cag PAI [22]. Therefore the
presence of the H. pylori plasticity region genes jhp0945,
jhp0947 and jhp0949 was analyzed in a well defined
Dutch study population. Since the jhp0946 and jhp0948
have already been designated pseudo-genes [21], they
were not included in our analyses. Southern hybridiza-tion analysis of 45 clinical H. pylori isolates confirmed
the heterogeneity in the presence of the plasticity region
genes jhp0945, jhp0947, and jhp0949 in the tested

166 R. de Jonge et al. / FEMS Immunology and Medical Microbiology 41 (2004) 161–167
H. pylori strains. The presence of jhp0947 and jhp0949,
but not jhp0945 is significantly associated with duodenal
ulcer compared to gastritis. This observation is in
agreement with a previous study [22], although in that
report, the jhp0947 and jhp0949 genes were not com-pletely linked, and were associated with the presence of
the cag PAI. These differences may well reflect the
geographic variation of the H. pylori isolates used in
both studies. Indeed, geographic differences in the
presence and expression of H. pylori factors have been
reported for several H. pylori virulence factors, includ-
ing the cag PAI [23,24]. The presence of both jhp0947–
0949 and the cag PAI is highly associated with duodenalulcer disease when compared to gastritis, which is in the
agreement with the concept that expression of multiple
H. pylori virulence factors increases the pathogenicity of
the bacterium.
In conclusion, the jhp0947 and jhp0949 genes proba-
bly play an important role in H. pylori-mediated IL-12
production in monocyte cells in vitro, and the presence
of jhp0947 and jhp0949 are significantly associated withdevelopment of duodenal ulcer disease in vivo. It is
tempting to speculate that proteins encoded by jhp0947
or jhp0949 interact with monocyte cells to stimulate IL-
12 production leading to a pro-inflammatory Th1 re-
sponse, which on its turn results in increased tissue
damage and disease outcome. The presence of jhp0947
and jhp0949 was not linked with the presence of the cag
PAI in our study, and therefore, the jhp0947–0949 locusmay act as a novel H. pylori marker for disease outcome
in Western populations independent of the cag PAI.
Acknowledgements
We thank R.G.J. Pot for excellent technical assis-
tance. This study was supported by a grant from theUniversitair Stimulerings Fonds (USF) of the Vrije
Universiteit, Amsterdam, The Netherlands to E.J.K.
References
[1] Kuipers, E.J. (1998) Review article: relationship between Helico-
bacter pylori, atrophic gastritis and gastric cancer. Aliment.
Pharmacol. Ther. 12, 25–36.
[2] Meyer, F., Wilson, K.T. and James, S.P. (2000) Modulation of
innate cytokine responses by products of Helicobacter pylori.
Infect. Immun. 68, 6265–6272.
[3] Mohammadi, M., Czinn, S., Redline, R. and Nedrud, J. (1996)
Helicobacter-specific cell-mediated immune responses display a
predominant Th1 phenotype and promote a delayed-type hyper-
sensitivity response in the stomachs of mice. J. Immunol. 156,
4729–4738.
[4] Karttunen, R.A., Karttunen, T.J., Yousfi, M.M., el-Zimaity,
H.M., Graham, D.Y. and el-Zaatari, F.A. (1997) Expression of
mRNA for interferon-gamma, interleukin-10, and interleukin-12
(p40) in normal gastric mucosa and in mucosa infected with
Helicobacter pylori. Scand. J. Gastroenterol. 32, 22–27.
[5] de Jonge, R., Kusters, J.G., Timmer, M.S., Gimmel, V.,
Appelmelk, B.J., Bereswill, S., van Vliet, A.H.M., Meuwissen,
S.G., Kist, M., Vandenbroucke-Grauls, C.M.J.E. and Kuipers,
E.J. (2001) The role of Helicobacter pylori virulence factors in
interleukin production by monocytic cells. FEMS Microbiol. Lett.
196, 235–238.
[6] Guiney, D.G., Hasegawa, P. and Cole, S.P. (2003) Helicobacter
pylori preferentially induces interleukin 12 (IL-12) rather than
IL-6 or IL-10 in human dendritic cells. Infect. Immun. 71, 4163–
4166.
[7] Sharma, S.A., Tummuru, M.K., Miller, G.G. and Blaser, M.J.
(1995) Interleukin-8 response of gastric epithelial cell lines to
Helicobacter pylori stimulation in vitro. Infect. Immun. 63, 1681–
1687.
[8] D’Elios, M.M., Manghetti, M., De Carli, M., Costa, F., Baldari,
C.T., Burroni, D., Telford, J.L., Romagnani, S. and Del Prete, G.
(1997) T helper 1 effector cells specific for Helicobacter pylori in
the gastric antrum of patients with peptic ulcer disease. J.
Immunol. 158, 962–967.
[9] Trinchieri, G. (1995) Interleukin-12: a proinflammatory cytokine
with immunoregulatory functions that bridge innate resistance
and antigen-specific adaptive immunity. Annu. Rev. Immunol. 13,
251–276.
[10] Hoffman, P.S., Vats, N., Hutchison, D., Butler, J., Chisholm, K.,
Sisson, G., Raudonikiene, A., Marshall, J.S. and Veldhuyzen van
Zanten, S.J. (2003) Development of an interleukin-12-deficient
mouse model that is permissive for colonization by a motile
KE26695 strain of Helicobacter pylori. Infect. Immun. 71, 2534–
2541.
[11] Bauditz, J., Ortner, M., Bierbaum, M., Niedobitek, G., Lochs, H.
and Schreiber, S. (1999) Production of IL-12 in gastritis relates to
infection with Helicobacter pylori. Clin. Exp. Immunol. 117, 316–
323.
[12] deJonge, R., Bakker, D., van Vliet, A.H.M., Kuipers, E.J., Van
denbroucke-Grauls, C.M.J.E. and Kusters, J.G. (2003) Direct
random insertion mutagenesis ofHelicobacter pylori. J. Microbiol.
Meth. 52, 93–100.
[13] Covacci, A., Censini, S., Bugnoli, M., Petracca, R., Burroni, D.,
Macchia, G., Massone, A., Papini, E., Xiang, Z., Figura, N. and
Rappuoli, R. (1993) Molecular characterization of the 128-kDa
immunodominant antigen of Helicobacter pylori associated with
cytotoxicity and duodenal ulcer. Proc. Natl. Acad. Sci. USA 90,
5791–5795.
[14] Goodwin, A., Kersulyte, D., Sisson, G., VeldhuyzenvanZanten,
S.J., Berg, D.E. and Hoffman, P.S. (1998) Metronidazole resis-
tance in Helicobacter pylori is due to null mutations in a gene
(rdxA) that encodes an oxygen-insensitive NADPH nitroreduc-
tase. Mol. Microbiol. 28, 383–393.
[15] Sambrook, J., Fritsch, E.F. and Maniatis, T. (1989) Molecular
cloning, a laboratory manual, 2nd ed. Cold Spring Harbor
Laboratory, Cold Spring Harbor, NY.
[16] Kwon, Y.M. and Ricke, S.C. (2000) Efficient amplification of
multiple transposon-flanking sequences. J. Microbiol. Meth. 41,
195–199.
[17] Loffeld, R.J.L.F., Werdmuller, B.F., Kusters, J.G. and Kuipers,
E.J. (2000) IgG antibody titer against Helicobacter pylori
correlates with presence of cytotoxin associated gene A-positive
H. pylori strains. FEMS Immunol. Med. Microbiol. 28, 139–
141.
[18] Alm, R.A., Ling, L.S., Moir, D.T., King, B.L., Brown, E.D.,
Doig, P.C., Smith, D.R., Noonan, B., Guild, B.C., deJonge, B.L.,
Carmel, G., Tummino, P.J., Caruso, A., Uria-Nickelsen, M.,
Mills, D.M., Ives, C., Gibson, R., Merberg, D., Mills, S.D.,
Jiang, Q., Taylor, D.E., Vovis, G.F. and Trust, T.J. (1999)
Genomic-sequence comparison of two unrelated isolates of
the human gastric pathogen Helicobacter pylori. Nature 397,
176–180.

R. de Jonge et al. / FEMS Immunology and Medical Microbiology 41 (2004) 161–167 167
[19] Tomb, J.F., White, O., Kerlavage, A.R., Clayton, R.A., Sutton,
G.G., Fleischmann, R.D., Ketchum, K.A., Klenk, H.P., Gill, S.,
Dougherty, B.A., Nelson, K., Quackenbush, J., Zhou, L.,
Kirkness, E.F., Peterson, S., Loftus, B., Richardson, D., Dodson,
R., Khalak, H.G., Glodek, A., McKenney, K., Fitzegerald, L.M.,
Lee, N., Adams, M.D., Hickey, E.K., Berg, D.E., Gocayne, J.D.,
Utterback, T.R., Peterson, J.D., Kelley, J.M., Cotton, M.D.,
Weidman, J.M., Fujii, C., Bowman, C., Watthey, L., Wallin, E.,
Hayes, W.S., Borodovsky, M., Karpk, P.D., Smith, H.O., Fraser,
C.M. and Venter, J.C. (1997) The complete genome sequence of
the gastric pathogen Helicobacter pylori. Nature 388, 539–547.
[20] Tanaka, J., Suzuki, T., Mimuro, H. and Sasakawa, C. (2003)
Structural definition on the surface of Helicobacter pylori type IV
secretion apparatus. Cell Microbiol. 5, 395–404.
[21] Occhialini, A., Marais, A., Alm, R., Garcia, F., Sierra, R. and
Megraud, F. (2000) Distribution of open reading frames of
plasticity region of strain J99 inHelicobacter pylori strains isolated
from gastric carcinoma and gastritis patients in Costa Rica. Infect.
Immun. 68, 6240–6249.
[22] Santos, A., Queiroz, D.M., Menard, A., Marais, A., Rocha, G.A.,
Oliveira, C.A., Nogueira, A.M., Uzeda, M. and Megraud, F.
(2003) New pathogenicity marker found in the plasticity region
of the Helicobacter pylori genome. J. Clin. Microbiol. 41, 1651–
1655.
[23] Yamaoka, Y., Kodama, T., Gutierrez, O., Kim, J.G., Kashima,
K. and Graham, D.Y. (1999) Relationship between Helicobacter
pylori iceA, cagA, and vacA status and clinical outcome: studies in
four different countries. J. Clin. Microbiol. 37, 2274–2279.
[24] de Jonge, R., Pot, R.G.J., Loffeld, R.J.L.F., van Vliet, A.H.M.,
Kuipers, E.J. and Kusters, J.G. (2004) The functional status of the
Helicobacter pylori sabB adhesin gene as a putative marker for
disease outcome. Helicobacter 9, 158–164.
Copyright © 2022 FDOKUMEN




















