
Maternal or neonatal infection: association with neonatal encephalopathy outcomes

EXPERIMENTAL NEUROLOGY 96,60 l-6 1 1 ( 1987)
The H-Reflex in the Encephalopathy Due to Ammonia Intoxication
Departments ofNeurology of the Veterans Administration Medical Center and The University of Minnesota, Minneapolis, Minnesota 55417
Received October 27, 1986; revision received January 9, I987
Ammonia intoxication has been shown to decrease excitatory synaptic transmis- sion in several regions of the central nervous system. To investigate the relation be- tween an effect of ammonia on excitatory synaptic transmission and the behavioral depression in the encephalopathy due to ammonia intoxication, this study examined in the rat the effects of ammonia intoxication on the H-reflex, the behavioral and neurological signs of the encephalopathy due to ammonia intoxication, and corre- lated the effects on the H-reflex with the signs of encephalopathy. Ammonia intoxica- tion abolished the H-reflex without affecting the M-response. This indicated that am- monia intoxication decreased spinal excitatory synaptic transmission without aflect- ing neuromuscular excitatory synaptic transmission. In the encephalopathy due to ammonia intoxication, the H-reflex disappeared only during a very advanced stage of behavioral depression, i.e.,coma. During early stages of behavioral depression, i.e., during a decrease of reactions to sensory stimuli, the H-reflex was not affected by ammonia intoxication. Therefore, mechanisms other than a decrease of excitatory synaptic transmission in the central nervous system may be responsible for the behav- ioral depression seen in early stages of the encephalopathy due to ammonia intoxication. 0 1987 Academic Press, Inc.
INTRODUCTION
Ammonia2 intoxication produces a potentially reversible encephalopathy. The signs of this encephalopathy include lethargy, coma, and seizures (8). These signs are the result of a dysfunction of the whole central nervous sys-
’ I thank J. Landry for excellent technical assistance. This research was supported by a grant from the Veterans Administration.
* At physiologic pH, ammonia is present as a mixture of NH3 and NH;. The term ammonia is used for the sum of NH, and NH;.
601
0014-4886/87 $3.00 Copyright 0 1987 by Academic Press, Inc. All rights ofreproduction in any form reserved.

602 W. FUABE
tern (CNS) which is caused by the effects of ammonia on the function of individual neurons.
In many neurons, ammonia shifts the equilibrium potential of Cl- to the level of the resting membrane potential (15- 17, 20, 25, 29). This effect dis- turbs inhibitory neuronal interactions mediated by hyperpolarizing Cl--de- pendent postsynaptic inhibition (25). In addition, this effect may also reduce inhibitory effects of Ca*‘- and voltage-dependent hyperpolarizing Cl--cur- rents(l8,21,22).
Recently, it was shown that ammonia also decreases excitatory synaptic transmission in several regions of the CNS, hippocampus ( 1, 3 1, 32) and spinal cord (23,24). At present, it is not known how the effects of ammonia on excitatory synaptic transmission relate to the signs of the encephalopathy due to ammonia intoxication.
In alert and unrestrained animals, excitatory synaptic transmission in the CNS can be monitored by means of the H-reflex. The pathways of the H- reflex include two excitatory synapses, a spinal and a neuromuscular syn- apse. The transmitter at the spinal synapse is presumed to be glutamate (4). The transmitter at the neuromuscular synapse is acetylcholine. Preliminary studies in the cat showed that ammonia intoxication has no effect on neuro- muscular synaptic transmission (W. Raabe, unpublished data). Also, a sys- temic ammonia intoxication affects cerebral cortex and spinal cord simulta- neously to about the same degree (14). Those observations indicated that in the alert and unrestrained animal the H-reflex can be used to correlate the effects of ammonia on excitatory synaptic transmission in the spinal cord with the behavioral and neurologic signs of the encephalopathy due to am- monia intoxication.
This study examined in the rat the effects of acute systemic ammonia in- toxication on the H-reflex and the behavioral and neurologic signs of the encephalopathy due to ammonia intoxication, and correlated effects on the H-reflex with the signs of encephalopathy.
METHODS
Male Sprague-Dawley rats of 264-460 g body weight were prepared in three different ways as follows.
(i) To investigate the effects of ammonia intoxication on the H-reflex and to determine blood NH4+, rats were anesthetized with paraldehyde, 800 mg/ kg i.p. Catheters were placed in the femoral artery and vein for blood pressure monitoring and drug administration, respectively. Ammonium acetate (NH4Ac) was administered i.v. or i.p. In some animals, NH4+ was determined in the arterial blood. At a given time, the abdominal aorta was quickly ex- posed and punctured for blood sampling. Blood plasma was mixed for 5 min with ice-cold 5% perchloric acid and then centrifuged. The supematant was

AMMONIA AND H-REFLEX 603
alkalinized with 10 A4 NaOH to pH 11 to 13. NG was determined as NH3 with a NH3-sensitive electrode (Orion Research, model 95 12).
The H-reflex was recorded as described below. (ii) The effects of ammonia intoxication on the H-reflex were also investi-
gated in unanesthetized animals. On the day before the experiment, the ani- mals were anesthetized with pentobarbital and a catheter was placed in the right jugular vein. Overnight, the distal end of the catheter was hidden in a subcutaneous pocket between the shoulder blades. On the day of the experi- ment, the subcutaneous pocket was infiltrated with a local anesthetic agent, the distal catheter end removed, and connected to an infusion pump. The animal was placed in a soft shoulder harness and held in place by a 20-cm- long semiflexible spring wire attached to the harness. The animal rested com- fortably on its feet and could freely move within a circle of 20 cm diameter. As soon as sedation by NH&c was sufficient, stimulating and recording elec- trodes were placed and the H-reflex recorded.
(iii) The behavioral signs of the encephalopathy due to ammonia intoxica- tion were investigated in unanesthetized animals injected with NH& i.p. In some of these animals, the H-reflex was recorded as soon as sedation by NH4Ac permitted. These animals were used to correlate effects on the H- reflex with the signs of encephalopathy.
The H-reflex was recorded with disk electrodes (9 mm 4) placed on the left planta (active electrode) and the dorsum of the foot (reference electrode) (30). The left posterior tibial nerve was stimulated at 0.5 Hz with needle electrodes inserted posterior to the medial malleolus. A steel needle in the heel grounded the animal. Stimuli of low strength excited only Ia-alTerent fibers from muscle spindles, activated motoneurons monosynaptically, and elicited a muscle twitch, the H-reflex (Fig. 1). Stronger stimuli not only ex- cited Ia-afferents but also the axons of (Y motoneurons. As the result, an M- response, or direct muscle response, appeared before the H-reflex (Fig. 1). Occasionally, the H-reflex could not be elicited without a preceding M-re- sponse. In these instances, stimuli just above the threshold for a-motoneuron axons readily produced an H-reflex. The low stimulus strength used virtually eliminated contamination of the H-reflex by an F-response, a sign of anti- dromic excitation of motoneurons. The F-response is essentially seen only with stimuli that are supramaximal for the M-response ( 19,33). NH4Ac was administered as a 20% solution in HZ0 adjusted to pH 7.4 with NaOH (and HCl, if necessary). At termination of the experiment the animals were killed with an overdose of pentobarbital or thyamylal sodium, i.v. or i.p.
RESULTS
H-Reflex in Ammonia Intoxication. Paraldehyde-anesthetized animals were administered NH&Z i.v. at rates of 0.108 to 0.200 mmol/kg/min until

604 W. RAABE
FIG. 1. A-left tracing: stimulation of the posterior tibia1 nerve elicited an H-reflex. Stimulus strength was low and excited only Ia-alferent fibers. Right tracing: Increase of the stimulus strength elicited an M-response prior to the H-reflex. Stimulus application is indicated by the artifact at the left of the tracings. Tracings are five superimposed responses. B-schematic draw- ing of the pathways of the H-reIlex and M-response. H-reflex: stimulus S excites only Ia-afferent fibers. In the spinal cord, Ia-afferents monosynaptically excite (Y motoneurons. Action potentials travel in cy-motoneurons axons from the spinal cord to the neuromuscular junction and excite muscle fibers. M-response: stimulus S excites Ia-afIerents plus oc-motoneuron axons: an M-re- sponse occurs in addition to the H-reflex. In cy-motoneuron axons action potentials travel from S to the neuromuscular junction and excite muscle fibers to yield the M-response. Because the distance from S to the neuromuscular junction is shorter than from S to the spinal cord and then to the muscle, the M-response has a shorter latency than the H-reflex.
an effect on the H-reflex was seen. The H-reflex disappeared after administra- tion of 3.73 to 12.49 mmol/kg NH& (average: 6.56 mmol/kg, N = 10) (Figs. 2, 3). NH4Ac abolished the H-reflex without affecting the M-response (Figs. 2,3).
Although intravenous administration of NH4Ac was terminated as soon as the H-reflex disappeared, 8 of 10 animals died within 1 to 45 min (average: 23 min) after disappearance of the H-reflex. In those animals the H-reflex remained absent until they died. In the two surviving animals, the H-reflex reappeared 20 and 24 min after termination of NH&c administration. On the day after the experiment, the surviving rats showed no sign of encepha- lopathy.
In another set of experiments, paraldehyde-anesthetized animals received 6.0 to 6.5 mmol/kg NH& or sodium acetate (NaAc) i.p. When the H-reflex disappeared, arterial blood was sampled and NH4f determined. The disap pearance of the H-reflex was accompanied by an increase of blood NH4+ to 1.406 mmol/liter (Table 1).
NH&c i.v. and i.p. had a typical effect on the blood pressure. Initially, the blood pressure decreased. Thereafter, the blood pressure increased beyond the control value. The H-reflex always disappeared during the blood pressure

AMMONIA AND H-REFLEX 605
Control --b-
Recovery - L-1,_-I, mV
FIG. 2. Effects of NH& i.v. on H-reflex and M-response in an unanesthetized rat with a semichronic catheter in the jugular vein. Control: Stimulation of the posterior tibia1 nerve elic- ited a small M-response and a large H-reflex in the plantar muscles. NH&c: 6.45 mmol/kg NH.,Ac i.v. abolished the H-reflex but did not affect the M-response. Recovery: After termina- tion of NH& i.v., the H-reflex reappeared. M-response and H-reflex were slightly changed compared with Control because spontaneous movement of the animal slightly changed the posi- tion of stimulation and recording electrodes.
increase. After the termination of NH&c administration, the blood pressure decreased and the H-reflex recovered (Fig. 3).
The effects of NH&, i.v., on the H-reflex were also studied in five unanes- thetized animals with a catheter in the jugular vein. The H-reflex disappeared after administration of 5.17 to 10.9 1 mmol/kg NH4Ac (average: 6.88 mmol/ kg) at rates of 0.098 to 0.165 mmol/kg/min. All five animals had seizures while the H-reflex was present. Two animals had seizures also after the H- reflex disappeared. Four of these five animals died within 1 to 55 min after disappearance of the H-reflex. One animal recovered fully (Fig. 2).
Encephalopathy Due to Ammonia Intoxication. To investigate the behav- ioral and neurologic signs of the encephalopathy due to ammonia intoxica-
-.‘-
2
-- -~~&-), mV b
FIG. 3. Effects of NH& i.v. on H-reflex and blood pressure in a paraldehyde-anesthetized rat. The H-reflex disappeared when NH.+Ac iv. increased the blood pressure. Upper tracings: l-Control record of H-reflex before NH& i.v.; 2-H-reflex disappeared due to 3.73 mmol/ kg NH& i.v.; 3-Recovery of the H-reflex after termination of NH4Ac iv. Lower tracing: Record of blood pressure during NH& iv. The numbers above the tracing indicate the time when the upper tracings (l-3) were obtained.

606 W. RAABE
TABLE 1
Arterial Blood NH: at the Time of the Disappearance of the H-Reflex after Administration of NH& in Paraldehyde-Anesthetized Rats’
NH+4 (mmol/liter) N
Control (NaAc i.p.), H-reflex present 0.077 2 0.015 5 Nmc i.p., H-reflex disappeared 1.406 + 0.374+ 5
a Values are X + SE. * P < 0.02 vs. control value.
tion, normal rats received various doses of NH& i.p. Only doses greater than 4.6 mmol/kg produced encephalopathy reliably. 5.2 mmol/kg readily produced a reversible encephalopathy. Doses greater than 5.5 mmol/kg fre- quently caused an irreversible encephalopathy, i.e., death of the animal. Therefore, 5.2 mmol/kg NH4Ac i.p. was chosen as the standard dose to pro- duce encephalopathy.
Reliable signs of encephalopathy were the following: (i) hyperventilation (8, 12), (ii) loss of motor activity in response to pull on a limb (motor re- sponse), (iii) loss of the righting reflex, (iv) loss of a whole body jerk in re- sponse to a loud hand clap (startle response), and (v) tonic seizures. If tonic seizures occurred, they were always followed by death of the animal. Partial and clonic seizures were seen in some animals that recovered from NH,& i.p.
The time course and the order of appearance of the signs of encephalopa- thy were examined in 27 animals of 27 1 to 450 g body weight (average: 328 g). The signs of encephalopathy always appeared in the same sequence: hy- perventilation was followed by decreased motor response to pull on a limb, then loss of the righting reflex, and eventually loss of the startle response. The encephalopathy was graded according to the number of abnormal signs present (Table 2). Seizures and death were not included in the grading. For example, hyperventilation plus loss of the motor response to pull on a limb plus loss of the righting reflex plus loss of the startle response yielded a grade 4 encephalopathy. Animals with grade 4 encephalopathy lay motionless on their side and did not respond to sensory stimuli, i.e., were comatose.
The time course of the encephalopathy due to 5.2 mmol/kg NH& i.p. is shown in Fig. 4; 21 animals showed at some time grade 4 encephalopathy. Grade 4 encephalopathy began 18 to 38 min (average: 24 min, N = 2 1) after NI&Ac i.p., and lasted for 8 to 52 min (average: 23 min, N = 21). Seizures were observed only in five animals with grade 4 encephalopathy. Three ani- mals developed partial, clonic seizures and later recovered from the encepha- lopathy. Two animals had tonic seizures and died shortly thereafter.

AMMONIA AND H-REFLEX 607
TABLE 2
Grading System to Stage the Encephalopathy Due to Ammonia Intoxication”
Grade Respiration Motor
response Righting
reflex Startle
response
0 Normal 1 HV 2 HV 3 HV 4 HV
+ + + + + + - + + - - + - - -
’ HV-hyperventilation, + -present, - -absent.
To ascertain if body weight influenced the results, the animals were sepa- rated according to body weight, one group of 27 1 to 308 g, N = 14, and one group of 325 to 450 g, N = 13. Body weight had no effect on the sequence in which encephalopathy signs appeared or on the time course of the encepha- lopathy.
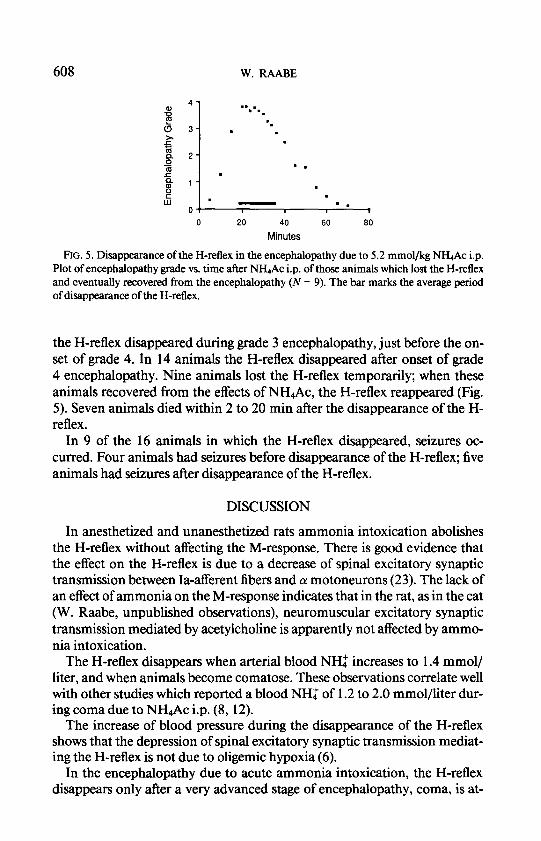
Encephalopathy and H-Reflex. To investigate the disappearance of the H- reflex in relation to the signs and time course of encephalopathy, 30 normal animals (255 to 496 g bodyweight; average: 317 g) received 5.2 mmol/kg NH.& i.p. The H-reflex was recorded as soon as the animals tolerated place- ment of stimulus and recording electrodes. Severity and time course of the encephalopathy were similar to that shown in Fig. 4 (Fig. 5). A total of 20 animals developed grade 4 encephalopathy.
In all animals the H-reflex was present during grade 1, grade 2, and at the onset of grade 3 encephalopathy. The H-reflex disappeared in 16 animals, all of which eventually developed grade 4 encephalopathy. In two animals
60
Minutes
T
120
FIG. 4. Time course of the encephalopathy due to 5.2 mmol/kg NH&c i.p. Plot of the mean encephalopathy grade of all animals alive vs. time. Vertical bars denote *SE. 27 rats were alive at the beginning of the experiment. Downward arrows denote death of one rat each.
608 W. RAABE
4- $ -.-..
L- l . . .
P . ;ij g 2-
3 1 . c . Q 8 1- .
I.5 . . ' 01
.
0 20 40
Minutes
60 80
FIG. 5. Disappearance of the H-reflex in the encephalopathy due to 5.2 mmol/kg NH& i.p. Plot of encephalopathy grade vs. time after NH& i.p. of those animals which lost the H-reflex and eventually recovered from the encephalopathy (N = 9). The bar marks the average period of disappearance of the H-reflex.
the H-reflex disappeared during grade 3 encephalopathy, just before the on- set of grade 4. In 14 animals the H-reflex disappeared after onset of grade 4 encephalopathy. Nine animals lost the H-reflex temporarily; when these animals recovered from the effects of NH4Ac, the H-reflex reappeared (Fig. 5). Seven animals died within 2 to 20 min after the disappearance of the H- reflex.
In 9 of the 16 animals in which the H-reflex disappeared, seizures oc- curred. Four animals had seizures before disappearance of the H-reflex; five animals had seizures after disappearance of the H-reflex.
DISCUSSION
In anesthetized and unanesthetized rats ammonia intoxication abolishes the H-reflex without affecting the M-response. There is good evidence that the effect on the H-reflex is due to a decrease of spinal excitatory synaptic transmission between Ia-afferent fibers and (Y motoneurons (23). The lack of an effect of ammonia on the M-response indicates that in the rat, as in the cat (W. Raabe, unpublished observations), neuromuscular excitatory synaptic transmission mediated by acetylcholine is apparently not affected by ammo- nia intoxication.
The H-reflex disappears when arterial blood NH: increases to 1.4 mmol/ liter, and when animals become comatose. These observations correlate well with other studies which reported a blood NH: of 1.2 to 2.0 mmol/liter dur- ing coma due to NI&Ac i.p. (8, 12).
The increase of blood pressure during the disappearance of the H-reflex shows that the depression of spinal excitatory synaptic transmission mediat- ing the H-reflex is not due to oligemic hypoxia (6).
In the encephalopathy due to acute ammonia intoxication, the H-reflex disappears only after a very advanced stage of encephalopathy, coma, is at-

AMMONIA AND H-REFLEX 609
tamed and death may be pending. The H-reflex is present during the early stages of the encephalopathy. This observation suggests that spinal excitatory synaptic transmission is not depressed when the early signs of depressed be- havior occur, i.e., decreased motor activity in response to pull on a limb and loss of the righting reflex.
Ammonia intoxication affects spinal cord and cerebral cortex simulta- neously to about the same degree ( 14). The transmitter presumed to mediate spinal synaptic excitation, i.e., glutamate, is also presumed to mediate the majority of synaptic excitation in other regions of the CNS including the cerebral cortex (4, 10). Therefore, preservation of the H-reflex during the early stages of encephalopathy provides no support for the hypothesis that a depression of excitatory synaptic transmission in the cerebral cortex or other regions of the CNS initiates the encephalopathy due to ammonia intoxica- tion. A dysfunction of neuronal mechanisms other than (putatively glu- tamatergic) excitatory synaptic transmission, e.g., a disturbance of the equi- librium potential of Cl-, is possibly responsible for the initiation of the en- cephalopathy due to acute ammonia intoxication and the early signs of depressed behavior (26).
Seizures, a sign of neuronal hyperexcitation including excessive excitation of a motoneurons, occur before as well as after disappearance of the H-reflex. The disappearance of the H-reflex does not imply that excitatory synaptic transmission is completely abolished. Rather, the disappearance of the H- reflex suggests that Ia-EPSPs in (Y motoneurons are decreased in amplitude so that they no longer depolarize motoneurons beyond the firing threshold and initiate action potentials. These considerations suggest a potential mech- anism for the occurrence of seizures even after disappearance of the H-reflex. When (putatively glutamatergic) excitatory synaptic transmission is suffi- ciently decreased so that the H-reflex disappears, a massive excitatory synap- tic input to (Y motoneurons can still depolarize beyond the firing threshold and cause excessive discharges of (Y motoneurons.
The observations that behavioral depression and seizures can occur before and after depression of excitatory synaptic transmission, respectively, suggest that one must be cautious in relating specific changes in behavior or specific clinical signs to particular disturbances of neuronal function.
In the rat, selection of the appropriate dose of NH4Ac i.p. can readily pro- duce reversible coma without seizures. This observation contradicts the statement that “Ammonia itself does not reproducibly cause coma in experi- mental animals” (9).
In man, transient neonatal hyperammonemia may acutely increase blood NH4f beyond 1.4 mmol/liter and abolish muscle stretch reflexes (7). Sim- ilarly, some patients in coma due to hepatic (portasystemic) encephalopathy become areflexic (13, 27, 28), although, at blood NE values less than 1 mmol/liter (2, 28). Portasystemic shunting of blood increases the uptake of

610 W. RAABE
NH: from blood into the CNS (3,5,11). Therefore, in patients with portasys- temic encephalopathy, the increase of blood N@ probably need not reach 1.4 mmol/liter to increase NH$ in the CNS to the level that decreases spinal excitatory synaptic transmission and abolishes muscle stretch reflexes.
The clinical observations of absent muscle stretch reflexes in diseases asso- ciated with hyperammonemia correlate well with the findings of this report and suggest that, in man also, ammonia intoxication may decrease spinal excitatory synaptic transmission.
REFERENCES
1. ALGER, B. E., AND R. A. NICOLL. 1983. Ammonia does not selectively block IPSPs in rat hippocampal pyramidal cells. J. Neurophysiol. 49: 138 I- 139 1.
2. CONN, H. O., AND M. M. LIEBERTHAL. 1979. The Hepatic Coma Syndromes andlactulose. Williams & Wilkins, Baltimore.
3. CRUZ, N. F., AND T. E. Dunn. 1983. Local cerebral &rcose metabolism in rats with chronic portacaval shunts. J Cereb. Blood Flow Metab. 3: 3 1 l-320.
4. CURTIS, D. R., AND G. A. R. JOHNSTON. 1974. Amino acid transmitters in the mammalian central nervous system. Annu. Rev. Physiol. 69: 97-188.
5. DUFFY, T. E., F. PLUM, AND A. J. L. COOPER. 1983. Cerebral ammonia metabolism in vivo. Pages 371-388 in L. HERTZ, E. KVAMME, E. G. MCGEER, AND A. SCHOUSBOE, Eds., Glutamine, Glutamate and GABA. Liss, New York.
6. ECCLES, R. M., Y. LOYNING, AND T. OSHIMA. 1966. Effects of hypoxia on the monosynap tic reflex in the cat spinal cord. J. Neurophysiol. 29: 3 15-332.
7. EGGERMONT, E., H. DEVLIEGER, G. MARCHAL, J. JAEKEN, E. VANDENBUSSCHE, E. SMEETS, G. VANACKER, AND L. CORBEEL. 1980. Angiographic evidence of low portal liver perfusion in transient neonatal hyperammonemia. Acta Paediatr. Belg. 33: 1538- 1542.
8. EHRLICH, M., F. PLUM, AND T. E. DUFFY. 1980. Blood and brain ammonia concentrations after portacaval anastomosis. Effects of acute ammonia. J. Neurochem. 34: 1538- 1542.
9. FERENCI, P., S. C. PAPPAS, P. J. MUNSON, AND E. A. JONES. 1984. Changes in glutamate
receptors on synaptic membranes associated with hepatic encephalopathy or hyperam- monemia in the rabbit. Hepatology4: 25-29.
10. FONNUM, F. 1984. Glutamate: a neurotransmitter in mammalian brain. J. Neurochem. 42: l-11.
1 I. GJEDDE, A., A. H. LOCKWOOD, T. E. DUFFY, AND F. PLUM. 1978. Cerebral blood flow and metabolism in chronically hyperammonemic rata. Ann. Neural. 84: 325-330.
12. HINDTFELDT, B., AND B. K. SIESIQ. 197 1. Cerebral effects of acute ammonia intoxication. I. The influence on intracellular and extracellular acid-base parameters. Stand. J. Chn. Invest. 28: 353-364.
13. HOLM, E., H. THIELE, E. M. WOLPERT, M. E. HEIM, AND B. REIMANN-WEHRLE. 1980. Neurologische und psychiatrische Symptome bei akuten und chronischen Leberkrank- heiten. Therapiewoche 30: 4720-4806.
14. LIN, S., AND W. RAABE. 1985. Ammonia intoxication: effects on cerebral cortex and spinal cord. J. Neurochem. 44: 1252-1258.
15. LLIN.~, R., R. BAKER, AND W. PRECHT. 1974. Blockage of inhibition by ammonium ace- tate action on chloride pump in cat trochlear motoneurons. J. Neurophysiol. 37: 522- 533.
16. Lux, H. D. 197 1. Ammonium and chloride extrusion: hyperpolarizing synaptic inhibition in spinal motoneurons. Science 173: 555-557.

AMMONIA AND H-REFLEX 611
17. Lux, H. D., C. LORACHER, AND E. NEHER. 1970. The action of ammonium on postsynap- tic inhibition in spinal motoneurons. Exp. Bruin Res. 11: 43 l-447.
18. MADISON, D. V., R. C. MALENKA, AND R. A. NICOLL. 1986. Phorbol esters block a voltage- sensitive chloride current in hippocampal pyramidal cells. Nature 321: 695-697.
19. MAGLADERY, J. W., A. M. PARK, W. E. PORTER, AND R. D. TEASDALL. 1952. Spinal reflex patterns in man. Res. Publ. Assoc. Nerv. Ment. Dis. 30: 118- 15 1.
20. NICOLL, R. A. 1978. The blockade of GABA mediated responses in the frog spinal cord by ammonium ions and furosemide. J. Physiol. (London) 283: 12 l-1 32.
21. OWEN, D. G., M. SEGAL, AND J. L. BARKER. 1984. A Ca-dependent Cl- conductance in cultured mouse spinal neurones. Nature 311: 567-570.
22. OWEN, D. G., M. SEGAL, AND J. L. BARKER. 1986. Voltage-clamp analysis of a Ca++- and voltage-dependent conductance in cultured mouse spinal neurons. J Neurophysiol. 55: 1115-1135.
23. RAABE, W. 1986. Ammonia decreases excitatory synaptic transmission in cat spinal cord. Sot. Neurosci. Abstr. 12: 825.
24. RAABE, W. 1987. Synaptic transmission in ammonia intoxication. Neurochem. Pathol., in press.
25. RAABE, W., AND R. J. GUMNIT. 1975. Disinhibition in cat motor cortex by ammonia. J. Neurophysiol. 38: 347-355.
26. RAABE, W., AND S. LIN. 1985. Pathophysiology of ammonia intoxication. Exp. Neurol. 87: 5 19-532.
27. SCHENKER, S., K. BREEN, AND A. M. HOYUMPA. 1974. Hepatic encephalopathy: current status. Gastroenterology 66: 12 1- 15 1.
28. SHERLOCK, S., W. H. J. SUMMERSKILL, L. P. WHITE, AND E. A. PHEAR. 1954. Portal- systemic encephalopathy. Neurological complications of liver disease. Luncet 2: 453- 457.
29. SINGER, W., AND 0. CREUTZFELDT. 1974. Unpublished results quoted by Creutzfeldt 0. and J. Houchin. Neuronal basis of EEG waves. Pages 2C3-2C55 in A. REMOND, Ed., Handbook of Electroencephalography and Clinical Neurophysiology, Vol. 2, Part C. Elsevier, Amsterdam.
30. STANLEY, E. F. 198 1. Sensory and motor nerve conduction velocities and the latency of the H-reflex during growth of the rat. Exp. Neurol. 71: 497-506.
3 1. THEORET, Y., M. F. DAVIES, B. ESPLIN, AND R. CAPEK. 1985. Effects of ammonium chlo- ride on synaptic transmission in the rat hippocampal slice. Neuroscience 14: 798-806.
32. THEORET, Y., AND J.-L. BOSSU. 1985. Effects ofammonium salts on synaptic transmission to hippocampal CA1 and CA3 pyramidal cells in viva. Neuroscience 14: 807-82 1.
33. THORNE, J. 1965. Central responses to electrical activation of the peripheral nerves supply- ing the intrinsic hand muscles. J. Neurol. Neurosurg. Psychiatry 28: 482-495.
Copyright © 2022 FDOKUMEN




















