
The genus Erctella Monterosato, 1894: new molecular evidence (Pulmonata Stylommatophora Helicidae)
12
Biodiversity Journal, 2015, 6 (1): 401–411 The genus Erctella Monterosato, 1894: new molecular evid- ence (Pulmonata Stylommatophora Helicidae) Maria Stella Colomba 1* , Armando Gregorini 1 , Fabio Liberto 2 , Agatino Reitano 3 , Salvatore Giglio 4 & Ignazio Sparacio 5 1 Università di Urbino, Dipartimento di Scienze Biomolecolari (DiSB), via Maggetti 22, loc. Sasso, 61029 Urbino, Pesaro-Urbino, Italy; e-mail: [email protected], [email protected] 2 Strada Provinciale Cefalù-Gibilmanna n° 93, 90015 Cefalù, Palermo, Italy; email: [email protected] 3 Via Gravina 77, 95030 Tremestieri Etneo, Catania, Italy; email: [email protected] 4 Contrada Settefrati, 90015 Cefalù, Palermo, Italy; email: [email protected] 5 Via E. Notarbartolo 54 int. 13, 90145 Palermo, Italy; email: [email protected] * Corresponding author, email: [email protected] ABSTRACT In this paper we report on new molecular data (COI sequences) of different and represent- ative populations of Erctella mazzullii (De Cristofori et Jan, 1832), E. cephalaeditana Gian- nuzzi-Savelli, Oliva et Sparacio, 2012 and E. insolida (Monterosato, 1892) (Pulmonata, Stylommatophora, Helicidae). Present results are compared with those from recent literature and the current knowledge on phylogenetic relationships among Helicidae pulmonate gastropods is reviewed. Obtained results suggest that: i) Cornu Born, 1778 and Cantareus Risso, 1826 are separate and well distinct from Helix Linnaeus, 1758; ii) Erctella Monterosato, 1894 is a valid and independent genus rather than a subgenus of Cornu; iii) Cornu aspersum (O.F. Müller, 1774) is a group of species (i.e. "aspersum" group) whose taxonomic status needs to be defin further studies; iv) Cornu, Cantareus and Erctella might belong to the same tribe that, still, remains to be defined. KEY WORDS Erctella; Helicidae; mitochondrial markers; phylogenetic reconstruction. Received 11.02.2015; accepted 18.03.2015; printed 30.03.2015 Proceedings of the 2nd International Congress “Speciation and Taxonomy”, May 16th-18th 2014, Cefalù-Castelbuono (Italy) INTRODUCTION Colomba et al. (2011) reported on a multidiscip- linary study based on genital morphology, DNA analysis, distribution, ecology and fossil records of Cornu mazzullii (De Cristofori et Jan, 1832), a species endemic to North-Western Sicily. Obtained results supported the hypothesis that C. mazzul- lii should be attributed to the genus Erctella Monterosato, 1894 and that this genus was probably structured in three discrete clades (i.e., the mazzullii group) recognized as species including: (i) the populations living in Monte Pellegrino (Palermo) and nearby mountains, E. mazzullii s. str., (ii) the endemic population of Cefalù, La Rocca, E. ceph- alaeditana Giannuzzi-Savelli, Oliva et Sparacio, 2012, and (iii) the populations living in the mountains of Trapani surroundings, E. insolida (Monterosato, 1892). Based on the phylogenetic reconstruction obtained by the multigenic analysis of nuclear (ITS2) and mitochondrial (16S rDNA, 12S rDNA) molecular markers, Colomba et al. (2011) strongly suggested that the genus Erctella should be kept distinct from the closely related genera Cornu Born, 1778 and Cantareus Risso, 1826. In the
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of The genus Erctella Monterosato, 1894: new molecular evidence (Pulmonata Stylommatophora Helicidae)

Biodiversity Journal, 2015, 6 (1): 401–411
The genus Erctella Monterosato, 1894: new molecular evid-ence (Pulmonata Stylommatophora Helicidae)
Maria Stella Colomba1*, Armando Gregorini1, Fabio Liberto2, Agatino Reitano3, Salvatore Giglio4 & IgnazioSparacio5
1Università di Urbino, Dipartimento di Scienze Biomolecolari (DiSB), via Maggetti 22, loc. Sasso, 61029 Urbino, Pesaro-Urbino, Italy;e-mail: [email protected], [email protected] Provinciale Cefalù-Gibilmanna n° 93, 90015 Cefalù, Palermo, Italy; email: [email protected] Gravina 77, 95030 Tremestieri Etneo, Catania, Italy; email: [email protected] Settefrati, 90015 Cefalù, Palermo, Italy; email: [email protected] E. Notarbartolo 54 int. 13, 90145 Palermo, Italy; email: [email protected]*Corresponding author, email: [email protected]
ABSTRACT In this paper we report on new molecular data (COI sequences) of different and represent-ative populations of Erctella mazzullii (De Cristofori et Jan, 1832), E. cephalaeditana Gian-nuzzi-Savelli, Oliva et Sparacio, 2012 and E. insolida (Monterosato, 1892) (Pulmonata,Stylommatophora, Helicidae). Present results are compared with those from recent literatureand the current knowledge on phylogenetic relationships among Helicidae pulmonategastropods is reviewed. Obtained results suggest that: i) Cornu Born, 1778 and CantareusRisso, 1826 are separate and well distinct from Helix Linnaeus, 1758; ii) ErctellaMonterosato, 1894 is a valid and independent genus rather than a subgenus of Cornu; iii)Cornu aspersum (O.F. Müller, 1774) is a group of species (i.e. "aspersum" group) whosetaxonomic status needs to be defin further studies; iv) Cornu, Cantareus and Erctellamight belong to the same tribe that, still, remains to be defined.
KEY WORDS Erctella; Helicidae; mitochondrial markers; phylogenetic reconstruction.
Received 11.02.2015; accepted 18.03.2015; printed 30.03.2015Proceedings of the 2nd International Congress “Speciation and Taxonomy”, May 16th-18th 2014, Cefalù-Castelbuono (Italy)
INTRODUCTION
Colomba et al. (2011) reported on a multidiscip-linary study based on genital morphology, DNAanalysis, distribution, ecology and fossil records ofCornu mazzullii (De Cristofori et Jan, 1832), aspecies endemic to North-Western Sicily. Obtainedresults supported the hypothesis that C. mazzul-lii should be attributed to the genus ErctellaMonterosato, 1894 and that this genus was probablystructured in three discrete clades (i.e., the mazzulliigroup) recognized as species including: (i) thepopulations living in Monte Pellegrino (Palermo)
and nearby mountains, E. mazzullii s. str., (ii) theendemic population of Cefalù, La Rocca, E. ceph-alaeditana Giannuzzi-Savelli, Oliva et Sparacio,2012, and (iii) the populations living in themountains of Trapani surroundings, E. insolida(Monterosato, 1892).
Based on the phylogenetic reconstructionobtained by the multigenic analysis of nuclear(ITS2) and mitochondrial (16S rDNA, 12S rDNA)molecular markers, Colomba et al. (2011) stronglysuggested that the genus Erctella should be keptdistinct from the closely related genera CornuBorn, 1778 and Cantareus Risso, 1826. In the

M.S. COLOMBA ET ALII402
same paper, this hypothesis was also corroboratedby the analysis of several 16S rDNA partialsequences downloaded from GenBank for othergenera representatives of Western PalaearcticHelicidae taxa; noteworthy, the phylogenetic treetopology clearly showed Cornu and Cantareusdistinct from Helix Linnaeus, 1758 (see Colombaet al., 2011, fig. 42).
Cornu Born, 1778 (type species: Cornu copiaeBorn, 1778) was reintroduced as distinct genus byWaldén (1976) with Cryptomphalus De Charpen-tier, 1837 (type species: Cryptomphalus aspersumO.F. Müller, 1774) as junior synonim; it was some-times considered as subgenus of Helix Linnaeus,1758 (type species: Helix pomatia Linnaeus, 1758)and sometimes as a distinct genus. The descriptionof Cornu copiae was based on a teratologicalspecimen of “Helix” aspersa; due to differentinterpretations of the Article 1.3.2 of the Code, arequest for conservation of the name Cornu is stillpending a ruling of the International Commissionon Zoological Nomenclature.
Cantareus Risso, 1826 (type species: Cantareusapertus Born, 1778) was sometimes considered assubgenus of Helix and sometimes as a distinctgenus.
Schileyko (1978) was the first one who de-scribed the internal structure of male sexual organsof "Helix" aspersa characterized by a penial papillaand a prominent semicircular fold in the distal partof the penis (see also Nordsieck, 2013). Because ofthese anatomical differences, the Author attributedthis species to the genus Cryptomphalus.
Giusti et al. (1995) showed a close similaritybetween genitalia of "Helix" aperta and "Helix"aspersa and, therefore, attributed these twospecies to the same genus, Cantareus, morpholo-gically well distinct from Helix. Moreover, theyreported that Helix has a real penial papilla insidethe penis and, distally, an accessory penial papilla,whereas Cantareus shows a system of a real penialpapilla, a false penial papilla and, distally, an"annular pad".
Neubert & Bank (2006) mainly confirmed thesemorphological differences and concluded in con-sidering Cornu and Cantareus as related but distinctgenera. One year later, similar observations werereported by Alonso & Ibáñez (2007).
At the same time, findings of scientific studiesbased on molecular data were in line with the taxo-
nomic frame showing Helix distinct from Canta-reus and Cornu, the latter two considered the samegenus (Manganelli et al., 2005; Koene & Schu-lenburg, 2005; Wade et al., 2006, 2007).
Nevertheless, despite all these anatomical andmolecular evidence, recently Welter-Schultes et al.(2011) and Welter-Schultes & Audibert (2012) con-sidered Cornu and Cantareus to belong to the genusHelix. Bank (2012) argued that such a systematicposition is wrong, and, above all, it does not takeinto account a number of studies (cited above)suggesting a taxonomic choice closer to the real af-finities among these taxa. Welter-Schultes et al.(2012), however, reaffirmed their beliefs and,besides, Welter-Schultes (2012) reported Erctellaas synonym of Helix.
Nordsieck (2013), reviewing the papers, pub-lished in the last decades, dealing with anatomicaland molecular data, concluded, in summary, that:“According to genital morphology and DNA ana-lysis, “Helix“ aspersa and relatives are not morerelated to Helix than Eobania and other genera ofthe Helicinae […] These species must therefore begenerically separated from Helix. The shell and thegenital differences, especially those of the penis(Giusti et al. 1995, Neubert & Bank 2006,Colomba et al. 2011), are sufficient for the genericseparation of Cantareus and Cornu (or Cryptom-phalus, if the name Cornu is not valid because ofArt. 1.3.2 ICZN, cf. Giusti et al. 1995: 491).Erctella is regarded as a subgenus of Cornu insteadof a genus, because it is more closely related toCornu than to Cantareus”.
More recently, detailed molecular geneticsstudies (Korábek et al., 2014; 2015; Razkin et al.,2015) confirmed Cornu and Cantareus as twodistinct genera forming a group with no sign of aclose relationship with Helix. In addition, ErctellaDNA sequences, when included in such analysis(see Korábek et al., 2015), confirmed this item, inline with Colomba et al. (2011).
At present there seems to be broad agreement inconsidering Cornu and Cantareus distinct genera,while on the position of Erctella opinions are stilldiverging. In order to be able to further test the“genus hypothesis” (Erctella as a distinct genus,Colomba et al., 2011) versus the “subgenus hypo-thesis” (Erctella as a Cornu subgenus, Nordsieck,2013), we performed an additional molecular ana-lysis to characterize and define even better, from a

molecular standpoint, the identity and reliability ofErctella.
In particular, phylogenetic relationships amongtaxa under study were analysed by comparing par-tial sequences of the gene encoding for the cyto-chrome oxidase subunit I (COI) - which is one ofthe most commonly used mitochondrial markers inmolecular evolution and molecular phylogeny.Besides, to provide a little contribute in shedingsome more light on Helicidae systematics, the ana-lysis was extended to hundreds of specimens of thefamily Helicidae whose COI sequences were down-loaded from GenBank database. A similar analysiswas carried out including 16S rDNA partialsequences of the same taxa. Molecular analyseshave been performed either with single (16S orCOI) or combined (16S+COI) molecular datasets.
MATERIAL AND METHODS
Specimens and Collection sites
For each population, 2-5 sicilian Erctella spe-cimens were analysed. Please note that each loc-ality and/or collection site is named in the originallanguage (Italian). Collected samples were iden-tified and [labelled] as follows: Erctella insolida(from Trapani province: Custonaci, Trapani [CU],M.te Cofano, Trapani [COF]; San Vito lo Capo:cala Mancina, Trapani [SV]); Erctella mazzullii(from W-Palermo surroundings: M.te Pellegrino[MP]; Sferracavallo, Palermo [CMS]; Carini:M.te Columbrina, Palermo [COL]; Cinisi: M.tePecoraro, Palermo [PEC]); Erctella cephalaed-itana from Cefalù: la Rocca, Palermo [CM];Cornu aspersum (= H. aspersa) [CA] from Ce-falù, Palermo, Sicily; and Cantareus apertus[CAP] from Cefalù, Palermo, Sicily and Assoro,Enna, Sicily.
DNA extraction, amplification and sequencing
Samples were stored separately at -20 °C in testtubes. Of each individual, a piece of foot tissue wasused for total DNA extraction (by Wizard GenomicDNA Purification Kit, Promega). COI fragments(581-663 bp) were amplified using LCO_1490 (5’-GGTCAACAAATCATAAAGATATTGG-3’) andHCO_2198 (5’-TAAACTTCAGGGTGACCAAAATCA-3’)
(Folmer et al., 1994). PCR cycles were as follows:95°C for 5 min; 95°C for 1 min, 42°C for 1 min,72°C for 1 min (35 cycles); 72°C for 5 min. Toremove primers and unincorporated nucleotides,amplified products were purified with the WizardSV gel and PCR Clean-up kit (Promega). Sequen-cing of purified PCR products was carried out usingautomated DNA sequencers at Eurofins MWGOperon (Germany). All COI sequences generatedin this study were uploaded in GenBank (accessionnumbers: KR921883-KR921914).
Phylogenetic analyses
The analysis was conducted on two partial genesequences: COI and 16S rDNA, integrating our datawith those obtained from GenBank database. Inparticular, in addition to the sequences obtainedfrom specimens tested directly in this study(KR921883-KR921914), to further expand the ana-lysis and refine its resolving power, we included16S rDNA sequences of Erctella mazzullii, E. insol-ida, E. cephalaeditana, Cornu aspersum and Can-tareus apertus previously generated by our researchgroup (GQ402393-GQ402396, GQ402398-GQ402402, GQ402403-GQ402405, GQ402407-GQ402409, GQ402410-GQ402411, GQ402412-GQ402414, GQ402417-GQ402419, GQ402420-GQ402422, GQ402387-GQ402389, GQ402390-GQ402392, see Colomba et al., 2011), joined toboth COI and 16S rDNA sequences downloadedfrom GenBank of the following taxa: Eobaniavermiculata (O.F. Müller, 1774) (KJ458509,KJ458510, KJ458511, JF277395, JF277393,JF277391), Theba geminata Mousson, 1857(KJ458559, HM034468), T. subdentata (Férussac,1821) (KJ458562, HM034496), T. pisana(O.F. Müller, 1774) (KJ458561, JX911311), T. an-dalusica Gittenberger et Ripken, 1987 (KJ458558,KF582631), Murella muralis (O.F. Müller, 1774)(GU391399, JX827154), Helix lucorum Linnaeus,1758 (AF126144, GU784803), Helix pomatia Lin-naeus, 1758 (AF208297, JX911304), Helix secern-enda Rossmässler 1847 (KP072386, KP072387,KP072388, KP072086, KP072087, KP072088),Helix vladika Kobelt, 1898 (KP072303,KF823104), Helix melanostoma Draparnaud 1801(KJ458524, KP072107), Iberus gualtierianus (Lin-naeus, 1758) (AY928605,AY928606, DQ822123,DQ822165, DQ822166, AY546285), Hemicycla
403The genus Erctella Monterosato, 1894: new molecular evidence (Pulmonata Stylommatophora Helicidae)

bidentalis (Lamarck, 1822) (KJ458528, HM147180),Pseudotachea splendida (Draparnaud, 1801)(KJ458552, AY546292), Levantina caesareana(Mousson, 1854) (KP072332, KP072181) Otalalactea (O.F. Müller, 1774) (AY937264, AY937263),O. punctata (O.F. Müller, 1774) (JF717823,JF717824, KJ458545, JF717805, JF717806,JF717807), Helix aspersa (AF126139, AF126135,AF126134, AF126140, AF126136, JN701926,JN701927, GU598217, AY546283, HQ203051,HQ203052, JX911287), Cantareus apertus(KJ458491, JX911286). Finally, Limax maximusLinnaeus, 1758 (Family Limacidae) (KF894386),L. cinereoniger Wolf, 1803 (KF894380), Limacusflavus (Linnaeus, 1758) (FJ896815), Muticariasyracusana (Philippi, 1836) (Family Clausiliidae)(HQ696868, AY425597) and M. neuteboomiBeckmann, 1990 (HQ696866, HQ696867) wereemployed as outgroups.
All sequences were visualized with BioEditSequence Alignment Editor 7 (Hall, 1999), alignedwith the ClustalW option included in this softwareand refined by eye. As far as concerns single (COIor 16S rDNA) molecular data sets, phylogeneticanalyses were conducted in MEGA 5 (Tamura et al.,2011) by Maximum Likelihood algorithm. Substi-tution models, selected according to the “Find BestDNA model” option included in the software, were:HKY+G (COI) and GTR+G (16S rDNA); supportfor the internodes was assessed by bootstrappercentages (BP) (1000 replicates). For the com-bined (COI+16S rDNA) datasets, phylogenetic ana-lyses were conducted in BEAST 1.6.1 (Drummond& Rambaut, 2007) using the *BEAST implement-ation (Heled & Drummond, 2010). A series ofinitial runs were performed to optimize priors andruntime parameter choice to obtain effectivesampling sizes (ESS) above 500 for all estimatedparameters. The best-fit evolution models ofnucleotide substitution were: HKY+G (COI) andGTR+G (16S rDNA) with empirical base com-position; the Yule Process tree prior for mitochon-drial data with piecewise linear population sizemodel was applied with a UPGMA-generated treeas starting point. Trees from all runs were combinedto produce an ultrametric consensus tree usingTreeAnnotator 1.6.1. The first 103 trees were dis-carded as burnin. Support for nodes was expressedas posterior probabilities.
RESULTS AND DISCUSSION
COI and 16S rDNA consensus trees and themulti-genic (COI+16S rDNA) tree included 69molecular sequences, each. Obtained results al-lowed to make a few observations of some interest.In particular, COI consensus tree (Fig. 1), showedthree separate clusters for (A) Erctella (discussedin detail below), (B) Cantareus apertus and (C)Cornu aspersum clearly distinct. Similarly, (D)Eobania vermiculata, (E) Levantina caesareana,(F) Helix spp. (including several species), (G)Otala spp. (O. punctata and O. lactea), (H)Murella muralis, (I) Hemicycla bidentalis,Pseudotachea splendida, Iberus gualtierianus and(L) Theba spp. (T. geminata, T. subdentata, T.pisana, T. andalusica) are separated. With regardto Erctella, the three taxa are clearly distinct andseparated as E. insolida (SV1-SV3, CU4-CU5,COF2-COF4, from Trapani province), E. mazzullii(CMS1-CMS5, COL1-COL3, PEC1-PEC3, MP1-MP3, comprising specimens sampled on M.tePellegrino and the nearby mountains of sur-roundings of Palermo), and E. cephalaeditana(CM1-CM4, from Cefalù, La Rocca).
The 16S rDNA consensus tree topology (Fig. 2)is similar to that shown in figure 1. In fact, also inthis case, Erctella is clearly distinct and well struc-tured in three taxa, Erctella insolida¸ E. cephalaed-itana and E. mazzullii. Once again, it is confirmeda distinction between the (closely related) generaErctella, Cornu and Cantareus; based on 16SrDNA sequences analysis, Erctella appears closerto Cornu, while in the COI tree Cornu is closer toCantareus.
Mean molecular distances among the three taxaof Erctella (assessed by the maximun compositelikelihood method), range from nearly 6 to 10%(16S rDNA) and about 4 to 7.5% (COI). Thesevalues, despite the issues of using mean moleculardistances (see Meier et al., 2008), nevertheless,compared with those reported for other species,including Pulmonata (e.g. Hebert et al., 2003a,2003b; Steinke et al., 2005; Nekola et al., 2009)can, in our opinion, justify the separation ofErctella populations into three species.
Genetic distances between different specieswithin various animal groups, especially inverteb-rates, are variable (see for example Meier et al.,2008 and references therein). This is because they
404 M.S. COLOMBA ET ALII

Figure 1. COI consensus tree. The evolutionary history was inferred by using the Maximum Likelihood method based onHKY model. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1000 re-plicates) are shown next to the branches. A discrete Gamma distribution was used to model evolutionary rate differencesamong sites [5 categories (+G, parameter = 0.6175)].
The genus Erctella Monterosato, 1894: new molecular evidence (Pulmonata Stylommatophora Helicidae) 405
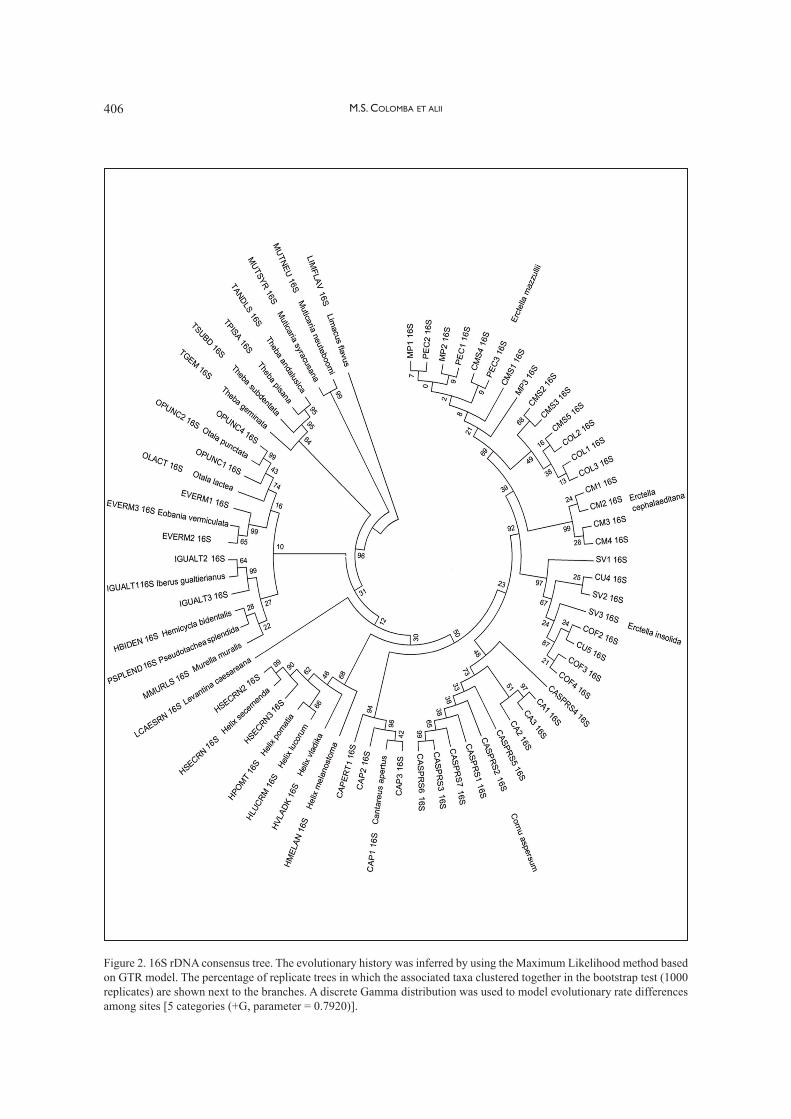
Figure 2. 16S rDNA consensus tree. The evolutionary history was inferred by using the Maximum Likelihood method basedon GTR model. The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (1000replicates) are shown next to the branches. A discrete Gamma distribution was used to model evolutionary rate differencesamong sites [5 categories (+G, parameter = 0.7920)].
M.S. COLOMBA ET ALII406

407
are function of different parameters among which:different rates of nucleotide substitution, differenttypes of environmental pressure, or different typesof mutation which the nucleotide sequences are sub-ject to, which at times (e.g. for retro-mutations)cannot be detected a-posteriori. Moreover, generallyspeaking, genetic distances per se are not sufficientto discriminate between different species and, forpulmonates, a few cases have been documentedwhere distances turned out to be misleading, neces-sitating to be integrated with additional data (seeDavison et al., 2009; Sauer & Hausdorf, 2012).
In Erctella, molecular data combined with othersignificant data such as morphological, biological,ecological and paleontological features allow us toconsider it a genus with three different species,endemic to northwestern Sicily (Liberto et al.,2010; Colomba et al., 2011).
Concatenated-gene analysis was better resolvedthan single-gene analysis and thus represents, prob-ably, more accurately present relationships amongtaxa. It resulted in a tree topology (Fig. 3) which isquite superimposable to that of the ML trees (Figs.1, 2) and, for the most part, in line with a recentreview of the molecular phylogeny of the WesternPalaearctic Helicoidea by Razkin et al. (2015). Inparticular, it is visible the group including Eobaniavermiculata, Otala lactea and O. punctata (tribeOtalini, in pink), the group including Iberus gual-tierianus, Pseudotachea splendida and Hemicyclabidentalis (Allognathini, in red), Theba species(Thebini, in yellow), and several species of Helixand Levantina caesareana (Helicini, in lilac).
In the concatenated-gene analysis, Erctella andCornu, considered two distinct genera, are sistergroups.
Figure 3. Phylogenetic annotated tree based on Bayesian inference analysis of the concatenated data set including 16S rRNAand COI sequences. Numbers correspond to BI posterior probabilities (in %).
The genus Erctella Monterosato, 1894: new molecular evidence (Pulmonata Stylommatophora Helicidae)

Regarding relationships within the groupCantareus-Erctella-Cornu our data differ fromRazkin et al. (2015). In fact, while for Erctella it isnot possible to make a comparison because theAuthors did not include this taxon in their analysis,on the other hand, in our tree, neither Cornu norCantareus can be considered Otalini, rather belong-ing to a distinct cluster (attributable to the “tribe”level) including Erctella.
Therefore, although Cornu and Cantareus showa certain degree of affinity particularly with Eo-bania for genitalia architecture (see Giusti et al.,1995) and share with Otalini similar biogeographic,ecological and evolutionary items typical ofWestern Mediterranean areas where these terrestrialmolluscs differentiated (see Colomba et al., 2011),nevertheless, the consideration of Cornu, Cantareusand Erctella as a separate tribe, which still remainsto be defined, is suggested. Furthermore, Cornu,Cantareus and Erctella share the same chromosomenumber (n = 27) (Vitturi et al., 1982; Vitturi et al.,2005) (see Fig. 3), while Eobania and other Otaliniexamined up to now have n = 26 (Burch, 1965;Thiriot-Quiévreux, 2003). Finally, Otalini show ingenital organs a relatively little dart sac and well-developed digit-like appendiges, Cornu-Cantareus-Erctella, instead, show a massive dart sac and twogroups of digitiform glands with short base andnumerous and short digit-like appendiges.
On the other hand, the separation betweenCornu-Cantareus-Erctella and Helix is supportedby: (i) the different geographical distribution of thegenera: Cornu and Cantareus are widespread inNorth Africa and Southern Europe, with Erctellaendemic to Northwestern Sicily, while Helix ismainly distributed in Central and Eastern Europeand, to a lesser extent, North Africa; (ii) the dif-ferent morphology of genital organs (Schileyko,1978; Giusti et al.,1995; Neubert & Bank, 2006,Alonso & Ibáñez, 2007) showing in Cornu-Can-tareus-Erctella a different form of dart sac and ofdigitiform glands (see above); and (iii) moleculardata (see Korábek et al., 2015 and quotes therein).
Comparing the three phylogenetic trees an inter-esting consideration about Cornu aspersum can bemade. In fact, in line with other studies (Guiller etal., 2001; Guiller & Madec, 2010), in our study aswell, this taxon seems to be not a single species butrather a species group (ie "aspersum" group)
showing a taxonomic situation more complex andheterogeneous than previously hypothesized withinits area of origin and diversification (SouthernItaly, Sicily and NW Africa). This result is furtherconfirmed by personal unpublished morphologicaland molecular data of numerous Italian, Malteseand North African C. aspersum populations.
Finally, the position of Murella muralis remainsto be clarified. In fact, it is not only different in allphylognetic trees but, above all, discordant withwhat reported in other papers. This issue, which isbeyond the aim of the present paper, requires furtherstudy and investigation, possibly increasing thenumber of specimens (joining to sequences down-loaded from the database also sequences obtainedfrom new samples collected directly in the field),increasing the number of genes analyzed and, aboveall, including in the analysis other taxa represent-atives of subfamilies more closely related to Murel-linae, such as Ariantinae.
Overall, present results correspond well toseveral previous molecular studies carried out bynuclear and mitochondrial markers (Koene &Schulenburg, 2005; Colomba et al., 2011; Korábeket al., 2014; Razkin et al., 2015) and confirm thatErctella species lie always outside the clusters ofCornu and Cantareus.
CONCLUSIONS
New molecular evidence provided in this studysuggested also several comments on Erctellaclosely related genera. Hence, on this basis, despitethe difficulties that the argument implies, some con-clusions can be drawn.
The groups comprising Cornu-Cantareus-Erctella on one hand, and Helix on the other hand,appear separate and distinct from each other. In linewith most of the papers reporting on anatomical andmolecular characteristics observed in these animals,there seems to be no evidence that "aperta","aspersa" and / or "mazzullii" may belong to thegenus Helix.
Considering Cornu and Cantareus as Otalini, asassumed by Razkin et al. (2015) is not confirmedin our analysis. However, as mentioned above, theissue certainly needs further study in view of theiraforementioned anatomical and biogeographicalaffinities.
408 M.S. COLOMBA ET ALII

We suggest considering Cornu, Cantareus andErctella as related but distinct genera belonging toindependent lineages; as hypothesized, they mightbe included into a new tribe (between Otalini andHelicini).
Cornu aspersum complex is in need of a thor-ough taxonomic revision in its area of origin.
Finally, it is appropriate to reiterate that our de-cision to consider Erctella a distinct genus includingthree different species (Colomba et al., 2011) was notonly made on the basis of some, although import-ant, molecular evidence, but also by the analysis ofmany other data that allowed us to assign to thevarious Erctella populations morphological, biolo-gical, paleontological and biogeographical peculiarcharacters, amplified by the particular distributionof the taxon, endemic to Northwestern Sicily. In thisregard it is worth remembering that in the charac-terization of a taxon, at different levels, whilegathering as many informations as possible (in-cluding morfological, ecological, molecular data,etc ...) is necessary, taxonomic reconstructions ob-tained with a methodology not always correspondto the ones obtained with another method (seeSchileyko, 2013); for Erctella, instead, all (numer-ous) data are consistent with the hypothesis of dif-ferentiating it from other (similar, closely related)genera.
So that it seems appropriate to conclude with thewords reported by A. Schmidt (1868) who claimedthat, in taxonomy : “Künstliche Systeme entstehendurch consequentes Geltendmachen eines einzelnenPrincips” [“the application of a single criterionproduces artificial classifications”].
In more contemporary terms, we could say withPoins et al. (2014): “Molecular phylogenetics is anirreplaceable tool for taxonomists, but interpreta-tion of the results must be based on clear taxonomicconcepts corroborated by all available resources -that is, the primary reference, the subsequent taxo-nomic literature and the type specimens of theorganisms of interest. Otherwise, molecular phylo-genetics can cause confusion with detrimental con-sequences to follow-up studies (e.g. ecological andevolutionary)”.
ACKNOWLEDGEMENTS
We are grateful to Eike Neubert, Research
Institute Senckenberg, Senckenberganlage, Frank-furt am Main, Germany; Giuseppe Pocaterra, SanPietro in Casale, Bologna, Italy; Fernando Scarlas-sara, Melego di Sarego, Vicenza, Italy.
REFERENCES
Alonso M.R. & Ibáñez M., 2007. Anatomy and functionof the penial twin papillae system of the Helicinae(Gastropoda: Helicoidea: Helicidae) and descriptionof two new, small Hemicycla species from the laurelforest of the Canary Islands. Zootaxa, 1482: 1–23.
Bank R.A., 2012. Comment on Cornu Born, 1778 (Mol-lusca, Gastropoda, Pulmonata, HELICIDAE): re-quest for a ruling on the availability of the genericname (Case 3518). http://iczn.org/node/40276
Burch J.B., 1965. Chromosome numbers and systematicsin euthyneuran snails. Proceedings of the firsteuropean malacological congress, London, 1962,215-241.
Colomba M.S., Gregorini A., Liberto F., Reitano A.,Giglio S. & Sparacio I., 2011. Monographic revisionof the endemic Helix mazzullii De Cristofori & Jan,1832 complex from Sicily and re-introduction of thegenus Erctella Monterosato, 1894 (Pulmonata,Stylommatophora, Helicidae). Zootaxa, 3134: 1–42.
Davison A., Blackie R.L.E. & Scothern G.P., 2009. DNAbarcoding of stylommatophoran land snails: a test ofexisting sequences. Molecular Ecology Resources, 9:1092–1101.
Drummond A.J. & Rambaut A., 2007. BEAST: Bayesianevolutionary analysis by sampling trees. BMC Evol-utionary Biology, 7: 1214–1257.
Hall T.A., 1999. BioEdit: a user-friendly biologicalsequence alignment editor and analysis program forWindows 95/98/NT. Nucleic Acids SymposiumSeries, 41: 95–98.
Hebert P.D.N., Cywinska A., Ball S.L. & deWaard J.R.,2003a. Biological identifications through DNA bar-codes. Proceedings of the Royal Society of LondonB, 270: 313–321. DOI: 10.1098/rspb.2002.2218
Hebert P.D.N., Ratnasingham S. & deWaard J.R., 2003b.Barcoding animal life: cytochrome c oxidase subunit1 divergences among closely related species. Pro-ceedings of the Royal Society of London B (Supple-ment), 270: S96–S99. DOI 10.1098/rsbl.2003.0025
Heled J. & Drummond A.J., 2010. Bayesian inference ofspecies trees from multilocus data. MolecularBiology and Evolution, 27: 570–580.
Folmer O., Black M., Hoeh W., Lutz R. & Vrijenhoek R.,1994. DNA primers for amplification of mitochon-drial cytochrome c oxidase subunit I from diversemetazoan invertebrates. Molecular Marine Biologyand Biotechnology, 3: 294–299.
409The genus Erctella Monterosato, 1894: new molecular evidence (Pulmonata Stylommatophora Helicidae)

Giusti F., Manganelli G. & Schembri P.J., 1995. The non-marine molluscs of the Maltese Islands. MuseoRegionale di Scienze Naturali, Torino, Monografie,15: 1–607.
Guiller A. & Madec L., 2010. Historical biogeography ofthe land snail Cornu aspersum: a new scenario in-ferred from haplotype ditribution in the WestenMedierranean basin. BMC Evolutionay Biology, 10:1–20.
Guiller A., Coutellec-Vreto M.A., Madec L. & Deunff J.,2001. Evolutionary history of the land snail Helixaspersa in the Western Mediterranen: preliminaryresults inferred from mitochondral DNA sequences.Molecular Ecology, 10: 81–87.
Koene J.M. & Schulenburg H., 2005. Shooting darts:co-evolution and counteradaption in hermaphroditicsnails. BMC Evolutionay Biology, 5: 25.
Korábek O., Juřičková L. & Petrusek A., 2014. Resur-recting Helix straminea, a forgotten escargot withtrans-Adriatic distribution: first insights into the gen-eric variation within the genus Helix (Gastropoda:Pulmonata). Zoological Journal of the LinneanSociety, 171: 72–91.
Korábek O., Petrusek A., Neubert E. & Juřičková L.2015. Molecular phylogeny of the genus Helix(Pulmonata: Helicidae). Zoologica Scripta. DOI:10.1111/zsc.12101
Liberto, F., Giglio, S., Reitano, A., Colomba, M.S. &Sparacio I., 2010. Molluschi terrestri e dulciacquicolidi Sicilia della collezione F. Minà Palumbo di Castel-buono. Monografie Naturalistiche, 2. EdizioniDanaus, Palermo, 134 pp.
Manganelli G., Salomone N. & Giusti F., 2005. A mo-lecular approach to the phylogenetic relationships ofthe western Palaearctic Helicoidea (Gastropda:Stylommatophora). Biological Journal of the LinneanSociety, 85: 501–512.
Meier R., Zhang G. & Ali F., 2008. The Use of meanInstead of Smallest Interspecific Distances Ex-aggerates the Size of the “Barcoding Gap” and Leadsto Misidentification. Systematic Biology, 57: 809–813.
Nekola J.C., Coles B.F. & Bergthorsson U., 2009.Evolutionary Pattern and Process within the Vertigogouldii (Mollusca: Pulmonata, Pupillidae) group ofminute North American Land Snails. MolecularPhylogenetics and Evolution, 53: 1010–1024.DOI:10.1016/j. ympev.2009.09.012
Neubert E. & Bank R.A., 2006. Notes on the species ofCaucasotachea C. Boettger 1909 and Lindholmia P.Hesse 1919, with annotations to the Helicidae(Gastropoda: Stylommatophora: Helicidae). Archivfür Molluskenkunde, 135: 101–132, 6 pls.
Nordsieck H., 2013. The systematic position of Helixaspersa Müller 1774. Available at.:
http://www.hnords.de/5356429d6b117f602/535642a15712fcf01/index.html - last access: 10.II.2015
Poins N., Vardinoyannis K., Mylonas M. & PoulakakisN., 2014. Evaluation of the taxonomy of Helix cincta(Müller, 1774) and Helix nucula (Mousson, 1854);insights using mitochondrial DNA sequence data.Journal of Natural History, DOI: 10.1080/00222933.2013.825023
Razkin O., Gómez-Moliner B.J., Prieto C.E., Martínez-Ortí A., Arrébola J. R., Muñoz B., Chueca L.J.,Madeira M.J., 2015. Molecular phylogeny ofthe western Palaearctic Helicoidea (Gastropoda,Stylommatophora). Molecular Phylogenetics andEvolution, 83: 99–117.
Sauer J. & Hausdorf B., 2012. A comparison of DNA-based methods for delimiting species in a Cretan landsnail radiation reveals shortcomings of exclusivelymolecular taxonomy. Cladistics, 28: 300–316. DOI:10.1111/j.1096-0031.2011.00382.x
Schileyko A.A., 1978. Nazemnye molljusk inadse-mejstva Helicoidea. In: Fauna SSSR, Molljuski, III,Leningrad, Nauka, 6: 384 pp.
Schileyko A.A., 2013. Family Helicidae excludingHelicinae (Gastropoda Pulmonata): morphology,taxonomy, and a catalogue of taxa. Ruthenica, 23:127–162.
Schmidt A., 1868. System der europäischenClausilienund ihrernächstenVerwandten. Th Fischer, Cassel, [1-2], 176 pp.
Steinke D., Vences M., Salzburger W. & Meyer A., 2005.TaxI: a software tool for DNA barcoding usingdistance methods. Philosophical Transactions of theRoyal Society of London B, 360: 1975–1980. DOI:10.1098/rstb.2005.1729
Tamura K., Peterson D., Peterson N., Stecher G., Nei M.& Kumar S., 2011. MEGA5: molecular evolutionarygenetics analysis using Maximum Likelihood,Evolutionary Distance, and Maximum Parsimonymethods. Molecular Biology and Evolution, 28:2731–2739.
Thiriot-Quiévreux C., 2003. Advances in chromosomalstudies of gastropod molluscs. Journal of MolluscanStudies, 69: 187–201.
Vitturi R., Rasotto M.B. & Farinella-Ferruzza N.,1982. The chromosomes of 16 molluscan species.Bolletino di zoologia, 49: 1–2, 61–71.DOI:10.1080/11250008209439373
Vitturi R, Libertini A., Sineo L., Sparacio I, Lannino A.,Gregorini A. & Colomba M.S., 2005. Cytogeneticsof the land snails Cantareus aspersus and C. mazzul-lii (Mollusca: Gastropoda: Pulmonata). Micron, 36:351–357
Wade C.M., Mordan P. B. & Naggs F., 2006. Evolutionaryrelationships among the Pulmonate land snailsand slugs (Pulmonata, Stylommatophora). BiologicalJournal of the Linnean Society, 87: 593–610.
410 M.S. COLOMBA ET ALII

411
Wade C.M., Hudelot C., Davison A., Naggs F. & MordanP.B., 2007. Molecular phylogeny of the helicoid landsnails (Pulmonata: Stylommatophora: Helicoidea),with special emphasis on the Camaenidae. Journal ofMolluscan Studies, 73: 411–415.
Waldén H.W., 1976. A nomenclatural list of the landMollusca of the British Isles. Journal of Conchology,29: 21–25 .
Welter-Schultes F., Audibert C. & Bertrand A., 2011.Liste des mollusques terrestres et dulcicoles deFrance continentale (excl. hydrobioïdes). FoliaConchyliologica, 12: 4–44.
Welter-Schultes F., 2012. European non-marine mol-
luscs, a guide for species identification. Planet PosterEditions, Göttingen, 760 pp.
Welter-Schultes F., Altaba C.R. & Audibert C., 2012.Comment on Cornu Born, 1778 (Mollusca,Gastropoda, Pulmonata, HELICIDAE): request fora ruling on the availability of the generic name(Case 3518). Bulletin of Zoological Nomenclature,70: 41–42.
Welter-Schultes F. & Audibert C., 2012. Comment onCornu Born, 1778 (Mollusca, Gastropoda, Pul-monata, Helicidae): request for a ruling on the avail-ability of the generic name. Bulletin of ZoologicalNomenclature, 69: 124–127.
The genus Erctella Monterosato, 1894: new molecular evidence (Pulmonata Stylommatophora Helicidae)

412




















