
The evolution of kin inclusivity levels
8
The Evolution of Kin Inclusivity Levels [Artificial Life Track] Anya Elaine Johnson BEACON Center for the Study of Evolution in Action Michigan State University, U.S.A. [email protected] Heather J. Goldsby BEACON Center for the Study of Evolution in Action University of Washington, U.S.A. [email protected] Sherri Goings Carleton College, U.S.A [email protected] Charles Ofria BEACON Center for the Study of Evolution in Action Michigan State University, U.S.A. [email protected] ABSTRACT Altruism is a ubiquitous strategy among organisms ranging from microbes to mammals. Inclusive fitness theory indi- cates that altruistic strategies can be beneficial when an altruist acts to benefit organisms that share its genes. It is common for such altruistic strategies to be negatively af- fected by cheaters that do not act altruistically. A more sub- tle form of cheating involves altruists that are more selective. For example, a selective organism may benefit from a distant kin’s altruistic actions without reciprocating. We consider an organism’s kin inclusivity level to be the maximum num- ber of mutational differences where the other organism will be considered kin. We use evolving computer programs (dig- ital organisms) to explore competitions among organisms with different kin inclusivity levels. Using competition as- says that vary environmental parameters, we find that high mutation rates favor more inclusive colonies. When we com- peted colonies with a wide range of kin inclusivity levels, we found that moderate mutation rates and populations sizes led to intermediate inclusivity levels winning the competi- tions, indicating that extreme inclusivity levels were not al- ways optimal. However, when organisms could set their own kin inclusivity level, we found that high mutation rates se- lected for highly inclusive organisms. Categories and Subject Descriptors I.2.8 [Artificial Intelligence]: Problem Solving, Control Methods, and Search Permission to make digital or hard copies of all or part of this work for personal or classroom use is granted without fee provided that copies are not made or distributed for profit or commercial advantage and that copies bear this notice and the full citation on the first page. Copyrights for components of this work owned by others than the author(s) must be honored. Abstracting with credit is permitted. To copy otherwise, or republish, to post on servers or to redistribute to lists, requires prior specific permission and/or a fee. Request permissions from [email protected]. GECCO’14, July 12–16, 2014, Vancouver, BC, Canada. Copyright is held by the owner/author(s). Publication rights licensed to ACM. ACM 978-1-4503-2662-9/14/07 ...$15.00. http://dx.doi.org/10.1145/2576768.2598283 . Keywords Digital evolution; Avida; kin altruism 1. INTRODUCTION The term altruism is used to describe any act where an organism lowers its own fitness in order to raise another or- ganism’s fitness [24]. Altruistic behaviors are observed fre- quently within nature in organisms ranging from E. coli [1] to termites [20] to human beings [5]. For example, one vari- ant of E. coli engages in suicidal behavior to release toxins (colicins) into the environment [1]. Specifically, when an E. coli colony encounters a rival group, some of the members will produce toxins and then explode. This explosion kills the colony member and releases toxins into the surrounding area. Close kin are resistant to the toxin, however mem- bers of the rival colonies are killed. Thus, the toxin reduces resource competition for the rest of the suicidal organism’s colony. Altruism in general, and suicidal altruism in partic- ular, is puzzling from the evolutionary perspective since an individual is harming or killing itself and therefore decreas- ing its ability to propagate its own genes. An explanation for this behavior is kin-selection (or inclusive fitness ), where the sacrificial act makes it more likely that many of the or- ganism’s kin, which share some of its genes, will survive and flourish [12, 13, 3, 7, 15]. One challenge to the evolution of altruistic behaviors is cheating, where an organism performs the altruistic act less frequently or forgoes altruistic actions altogether [24, 4, 23, 6, 22, 14, 8]. A subtle form of cheating involves organisms that behave altruistically to close kin, but not to more dis- tant kin that behave altruistically to them. We consider an organism’s kin inclusivity level to be the maximum number of mutational differences where the other organism will be considered kin. If an organism has a high kin inclusivity level, then they will be altruistic to a wide range of related organisms (clan altruists ). An organism with a low kin inclu- sivity level may only be altruistic toward their exact clones (clone altruists ). In the case that a clan altruist is related to a clone altruist, the clan altruist will behave altruistically to the clone altruist, but the clone altruist will not recip-
-
Upload
michiganstate -
Category
Documents
-
view
1 -
download
0
Transcript of The evolution of kin inclusivity levels

The Evolution of Kin Inclusivity Levels
[Artificial Life Track]
Anya Elaine JohnsonBEACON Center for the Study
of Evolution in ActionMichigan State University,
Heather J. GoldsbyBEACON Center for the Study
of Evolution in ActionUniversity of Washington,
Sherri GoingsCarleton College, U.S.A
Charles OfriaBEACON Center for the Study
of Evolution in ActionMichigan State University,
ABSTRACTAltruism is a ubiquitous strategy among organisms rangingfrom microbes to mammals. Inclusive fitness theory indi-cates that altruistic strategies can be beneficial when analtruist acts to benefit organisms that share its genes. Itis common for such altruistic strategies to be negatively af-fected by cheaters that do not act altruistically. A more sub-tle form of cheating involves altruists that are more selective.For example, a selective organism may benefit from a distantkin’s altruistic actions without reciprocating. We consideran organism’s kin inclusivity level to be the maximum num-ber of mutational differences where the other organism willbe considered kin. We use evolving computer programs (dig-ital organisms) to explore competitions among organismswith different kin inclusivity levels. Using competition as-says that vary environmental parameters, we find that highmutation rates favor more inclusive colonies. When we com-peted colonies with a wide range of kin inclusivity levels, wefound that moderate mutation rates and populations sizesled to intermediate inclusivity levels winning the competi-tions, indicating that extreme inclusivity levels were not al-ways optimal. However, when organisms could set their ownkin inclusivity level, we found that high mutation rates se-lected for highly inclusive organisms.
Categories and Subject DescriptorsI.2.8 [Artificial Intelligence]: Problem Solving, ControlMethods, and Search
Permission to make digital or hard copies of all or part of this work for personal orclassroom use is granted without fee provided that copies are not made or distributedfor profit or commercial advantage and that copies bear this notice and the full citationon the first page. Copyrights for components of this work owned by others than theauthor(s) must be honored. Abstracting with credit is permitted. To copy otherwise, orrepublish, to post on servers or to redistribute to lists, requires prior specific permissionand/or a fee. Request permissions from [email protected]’14, July 12–16, 2014, Vancouver, BC, Canada.Copyright is held by the owner/author(s). Publication rights licensed to ACM.ACM 978-1-4503-2662-9/14/07 ...$15.00.http://dx.doi.org/10.1145/2576768.2598283 .
KeywordsDigital evolution; Avida; kin altruism
1. INTRODUCTIONThe term altruism is used to describe any act where an
organism lowers its own fitness in order to raise another or-ganism’s fitness [24]. Altruistic behaviors are observed fre-quently within nature in organisms ranging from E. coli [1]to termites [20] to human beings [5]. For example, one vari-ant of E. coli engages in suicidal behavior to release toxins(colicins) into the environment [1]. Specifically, when an E.coli colony encounters a rival group, some of the memberswill produce toxins and then explode. This explosion killsthe colony member and releases toxins into the surroundingarea. Close kin are resistant to the toxin, however mem-bers of the rival colonies are killed. Thus, the toxin reducesresource competition for the rest of the suicidal organism’scolony. Altruism in general, and suicidal altruism in partic-ular, is puzzling from the evolutionary perspective since anindividual is harming or killing itself and therefore decreas-ing its ability to propagate its own genes. An explanationfor this behavior is kin-selection (or inclusive fitness), wherethe sacrificial act makes it more likely that many of the or-ganism’s kin, which share some of its genes, will survive andflourish [12, 13, 3, 7, 15].
One challenge to the evolution of altruistic behaviors ischeating, where an organism performs the altruistic act lessfrequently or forgoes altruistic actions altogether [24, 4, 23,6, 22, 14, 8]. A subtle form of cheating involves organismsthat behave altruistically to close kin, but not to more dis-tant kin that behave altruistically to them. We consider anorganism’s kin inclusivity level to be the maximum numberof mutational differences where the other organism will beconsidered kin. If an organism has a high kin inclusivitylevel, then they will be altruistic to a wide range of relatedorganisms (clan altruists). An organism with a low kin inclu-sivity level may only be altruistic toward their exact clones(clone altruists). In the case that a clan altruist is relatedto a clone altruist, the clan altruist will behave altruisticallyto the clone altruist, but the clone altruist will not recip-

rocate. In this case, the clone altruist would “cheat” theirclan altruist relatives, demonstrating kin discrimination [18].Without mechanisms in place for organisms to detect andprevent cheating behaviors based on kin inclusivity levels,clone altruists are likely to take over in highly-related pop-ulations. However, minimal kin inclusivity levels that allowonly clonal offspring will prevent further adaptive evolutionby killing mutants and thus limiting further adaptation. Inthis study, we explore the environmental conditions underwhich higher and lower kin inclusivity levels are adaptive.
To explore questions surrounding the evolution of kin in-clusivity levels, we use the Avida digital evolution plat-form [17], which has been used previously to study the prop-erties of evolving systems (e.g., [10] and [16]). Within Avida,digital organisms, which are computer programs that self-replicate and potentially incur mutations, compete with neigh-boring organisms for resources, including space. Using or-ganic experimental systems to study the evolution of kininclusivity levels is a time consuming and imperfect process.Even the fastest evolving natural organisms typically takemonths or years to undergo the thousands of generationsnecessary for evolution, whereas digital organisms take merehours. Moreover, controlling factors such as kin inclusivitylevels in natural organisms can be challenging, if not impos-sible. Because digital organisms evolve within a computer,we can rapidly perform experiments spanning thousands ofgenerations with complete experimental control and datacollection.
In previous work, we competed clone altruists (called ‘kin-cheaters’) against clan altruists [9]. We found that within ge-netically similar groups, the clone altruists frequently droveclan altruists out of the population. We also discovered thatin larger populations clan altruists were occasionally ableto outcompete initially genetically similar clone altruists.Here, we extend that work by exploring how a number ofenvironmental factors influence whether higher or lower kininclusivity levels are favored by evolution.
2. METHODSFor this study, we used the Avida digital evolution plat-
form [17]. Figure 1 depicts a population of digital organ-isms that resides in a toroidal grid of cells within the Avidaframework. Each digital organism has a genome, which is acircular list of instructions from a simple, assembly-like com-puter language. The Turing-complete instruction set fromwhich this list is derived includes instructions for basic com-putation (e.g., addition and increment), control of genomeinstruction execution (e.g., jumps), and the instructions re-quired for self-replication. An organism’s hardware consistsof a virtual CPU, several registers, and two stacks. An or-ganism must execute instructions to replicate itself. Whenan organism replicates, its offspring is placed in a randomcell adjacent to the parent organism. If the selected adjacentcell was occupied, the previous occupant is killed.
Our experiments use an instruction set that includes thedefault Avida instructions and several additional altruistic(explode) instructions. These instructions are modeled af-ter the suicidal behavior of colicinogenic E. coli. When anexplode instruction is executed by an organism, the organ-ism explodes with a 5% probability. An explosion will killthe organism itself, as well as any non-kin within two gridcells. An organism considers a neighbor kin if the Hammingdistance between its genome and the neighbor’s genome is
Figure 1: An Avida population, where half-circlesrepresent organisms and boxes represent the cellsthat they live in. A single organism’s hardware andgenome are superimposed. Different colors repre-sent different genomes.
less than its kin inclusivity level (χ). For example, if anorganism has a kin inclusivity level of χ = 3, it considersall neighboring organisms whose genomes differ by less thanthree instructions kin. All organisms’ genomes were fixedat 100 instructions and insertion and deletion mutations arenot permitted, therefore only substitution mutations couldoccur. The use of Hamming distance provides the organ-isms with a perfect measurement of genetic similarity in-stead of an approximation, such as kinship [2]. When anorganism explodes, it kills all organisms with a Hammingdistance greater than or equal to its kin inclusivity level.Thus, continuing with our example, an organism with a kininclusivity level of 3 would kill all neighboring organismswhose genomes differed in three or more instructions. Forsome experiments, we fixed the kin inclusivity level of theorganisms; for others we let the kin inclusivity level evolve.When the kin inclusivity level was fixed, it was statically setin a configuration file for a specific instruction. Organismswith a low kin inclusivity level recognize fewer organisms askin. For example, clone altruists have a minimal kin inclu-sivity level (χ = 1), and therefore kill any organism with oneor more genomic differences upon exploding (Figure 2A). Incontrast, organisms with higher kin inclusivity levels recog-nize more organisms as kin. For example, clan altruists haveχ = 6, so they kill organisms with six or more mutations inan explosion (Figure 2B).
Within our competition experiments, we required each or-ganism to maintain one and only one copy of an explode in-struction in its genome, whereas the rest of the genome stillundergoes mutation based on the set mutation rate. Thismeans an organism was not able to mutate its explode in-struction to a different Hamming distance, as that wouldrequire that fixed instruction to mutate to a different in-struction. Additionally, an organism was required to exe-cute the explode instruction at least once (with a 5% chanceof actually exploding) before the organism was allowed toreproduce. These constraints prevent the confounding ef-

Figure 2: A depiction of the organisms killed when aclone altruist (A) or clan altruist (B) explodes. Eachsquare represents an organism. The number in thesquare represents the Hamming distance betweenthat organism and the organism depicted in the mid-dle square. When that middle organism explodes asa clone altruist (χ = 1), it kills all organisms withintwo cells of it (the red-shaded area) that have anedit distance of greater than or equal to one. Whenthe middle organism is a clan altruist (χ = 6) andexplodes, it kills all organisms with an edit distanceof greater than or equal to six within two squares ofit. Organisms that were killed are overlaid with redX’s.
Figure 3: A pair of genetically similar (A) and apair of genetically dissimilar (B) organisms. For thegenetically similar case, there is a one-instructiondifference between the two genomes. For the genet-ically dissimilar case, there is an eleven-instructiondifference.
fects of organisms mutating to either avoid exploding or toexplode multiple times with different kin inclusivity levels,therefore allowing us to determine which genotype is favoredby evolution.
We performed competition assays in which we varied boththe genetic relationship among the colonies and also thenumber of distinct colonies present within the population,where a colony is a group of organisms descended from thesame ancestor organism and sharing the same explode in-struction. Specifically, we started each competition assaywith colonies that were either genetically similar or geneti-cally dissimilar to each other. Figure 3 depicts sample clanand clone altruist genomes for both cases. In the genetically-similar treatments, the only instruction that varied betweenthe colonies was the explosion instruction. Thus, in a two-colony experiment, the clan altruist considered the clone al-truist kin, but the clone altruist did not consider the clanaltruist kin. We created genetically dissimilar, but func-tionally equivalent, altruist genomes by inserting ten “no-operation” instructions into each colony’s genome. Becausethe clan altruists (the most inclusive genotype tested) wereset to have a kin inclusivity level below ten for our experi-ments (χ = 6), this genomic change meant that no colonyconsidered another colony to be kin. Thus, if a memberof any colony exploded, it would kill members of the othercolonies. In experiments with more than two competingcolonies, the no-operation instructions were placed in differ-ent locations to make all colonies genetically dissimilar fromall other colonies.
The goal of our competition experiments was to ascer-tain if a particular genotype was favored by evolution. Assuch, in the two-colony competition assays, we started bothcolonies at 50% of the population in two equal-sized patchesfilling the environment. In the multi-colony competitionassays, all colonies also started at equal frequencies in sixequal-sized patches made as close to perfectly square as pos-sible.

Number of Fixation Events in Genetically Similar Populations Over Multiple Population Sizes and Mutation Rates With Two Phenotypes
Mut
atio
n R
ate
Population Size Kin Inclusivity Level
Num
ber o
f Fix
atio
n E
vent
s
µ=0.
02
µ=0.
2
µ=2.
0
N=1000 N=5000 N=10000
Clone
Clan Clan Clan
Clan Clan Clan
Clan Clan Clan
Clone
Clone
Clone
Clone
Clone
Clone
Clone
Clone
Figure 4: The number of fixation events by clonealtruists (green) and clan altruists (purple) startingfrom equal proportions of genetically-similar clan al-truists and clone altruists at three different muta-tion rates and three different population sizes. Atthe low mutation rate, the clone altruists fix in allbut one replicate. At the medium mutation rate,however, the clan altruists win the majority of repli-cates. Finally, at the high mutation rate, the clanaltruists fix in every replicate.
Replicates were run until one colony reached 100% of thepopulation. We determined this by assigning a label to eachcolony at the beginning of the experiment and that label isinherited when an organism reproduces.
We used population sizes of 1000, 5000, and 10,000 forboth the two-colony and six-colony competitions. We usedgenomic mutation rates of 2.0, 0.2, and 0.02. All treatmentswere replicated 30 times.
3. RESULTSFor this study, we first performed competition assays to
examine the behavior of organisms that exhibit two extremekin inclusivity levels (e.g., clone altruists with χ = 1 and clanaltruists with χ = 6). We then expand our study to includeorganisms of intermediate kin inclusivity levels. Lastly, weallow organisms to set their own kin inclusivity level.
3.1 Clan and Clone Altruist Competition As-says
In our first experiment, we performed competition assaysthat started with one colony of clone altruists (χ = 1) andone colony of clan altruists (χ = 6). We varied whether ornot the two groups started genetically similar to each other,the population size (N = 1000, 5000, or 10,000 organisms),and the genomic mutation rate (µ = 0.02, 0.2, or 2.0 ex-pected mutations per organism reproduction).
Figures 4 and 5 present the results of our two-colony ex-periments with starting colonies that were genetically similarand dissimilar, respectively. When the colonies were genet-
Number of Fixation Events in Genetically Dissimilar Populations Over Multiple Population Sizes and Mutation Rates With Two Phenotypes
Mut
atio
n R
ate
Population Size Kin Inclusivity Level
Num
ber o
f Fix
atio
n E
vent
s
µ=0.
02
µ=0.
2
µ=2.
0
N=1000 N=5000 N=10000
Clone
Clan Clan Clan
Clan Clan Clan
Clan Clan Clan
Clone
Clone
Clone
Clone
Clone
Clone
Clone
Clone
Figure 5: The number of fixation events by clonealtruists (green) and clan altruists (purple) start-ing from equal proportions of genetically-dissimilarclone altruists and clan altruists at three differentmutation rates and three different population sizes.The clan altruists were able to outcompete the clonealtruists in all but one replicate across all treat-ments.
ically similar (Figure 4), at a high mutation rate (µ=2.0),the clan altruists outcompeted the clone altruists in all repli-cates at every population size. However, additional analysisrevealed that the clone altruists were often unable to sur-vive at the high mutation rate (µ=2.0), even in isolation,which likely confounded our results. Thus, we also testeda slightly lower mutation rate of µ=1.0. At this lower mu-tation rate, the clone altruists could consistently survive inisolation, but the clan altruists still fixed in all runs at ev-ery population size. When examined at a low mutation rate(µ=0.02), the clone altruists outcompeted the clan altru-ists in all replicates at the small and medium populationsize and 29/30 at the large population size. At the moder-ate mutation rate (µ=0.2), the clan altruists were able towin the majority of the replicates at each population size(16/30, 29/30, and 30/30 at the small, medium, and largepopulation sizes, respectively). These results indicate thatpopulations with more genetic diversity, due to a higher mu-tation rate or larger population size, permit individuals witha high kin inclusivity level (i.e., the clan altruists with χ = 6)to drive colonies with a lower kin inclusivity level (i.e., theclone altruists with χ = 1) out of the population.
Next, we performed the same series of competition assays,but used genetically dissimilar colonies as the initial popu-lations. Our results are depicted in Figure 5. Under theseconditions, the clan altruists won all but one of the compe-titions regardless of mutation rate and population size. Toensure these results were not driven by the clone altruists’problem with surviving at the high mutation rate (µ=2.0),we again tested a slightly lower mutation rate of µ=1.0. Theclan altruists still won all 30 replicates even at this reducedmutation rate. These results, when compared to the previ-

Number of Fixation Events in Genetically Similar Populations Over Multiple Population Sizes and Mutation Rates With Six Phenotypes
Mut
atio
n R
ate
Population Size Kin Inclusivity Level
Num
ber o
f Fix
atio
n E
vent
s
µ=0.
02
µ=0.
2
µ=1.
0
N=1000 N=5000 N=10000
1 2 3 4 5 6 1 2 3 4 5 6 1 2 3 4 5 6
1 2 3 4 5 6 1 2 3 4 5 6 1 2 3 4 5 6
1 2 3 4 5 6 1 2 3 4 5 6 1 2 3 4 5 6
Figure 6: The number of fixation events by eachof six genetically similar colonies with differing kininclusivity levels (χ = 1 through χ = 6). We per-formed these competition assays across three differ-ent mutation rates and three different populationsizes. Each panel represents the results of one com-petition assay. As mutation rate increases, colonieswith larger kin inclusivity levels fix more frequently.At intermediate mutations rates, larger populationsizes enable colonies with higher kin inclusivity lev-els to fix. At the highest mutation rate, larger popu-lation sizes enable a wider range of high-inclusivity-level colonies to reach fixation.
ous genetically similar results, demonstrate that the geneticsimilarity (or dissimilarity) among colonies within the ini-tial population strongly influenced which colony was able toreach fixation.
3.2 Intermediate Kin Inclusivity Levels Com-petition Assays
In nature, it is likely that organisms exhibit intermedi-ate phenotypes (i.e., kin inclusivity levels that range be-tween those selected for our clan and clone altruists), ratherthan constraining themselves to extreme phenotypes. Thus,in our second set of experiments, we performed competi-tion assays among six different colonies of organisms, whereeach colony had a different kin inclusivity level (i.e., χ = 1through χ = 6). In general, we examined the effects ofthese intermediate phenotypes under the same set of mu-tation rate and population size parameters we used for ourtwo-colony experiments. However, because clone altruists(χ = 1) are not able to survive in isolation at the highestmutation rate µ=2.0, we used a mutation rate of µ=1.0 asour highest mutation rate for these competition assays.
Figures 6 and 7 depict the results of our experiments whenall six colonies started genetically similar or dissimilar toeach other, respectively. When the colonies start geneticallysimilar, mutation rate strongly influences the results of thecompetition (Figure 6). At a low mutation rate, (µ=0.02)colonies with χ = 1 outcompeted all other colonies in all runs
Number of Fixation Events in Genetically Dissimilar Populations Over Multiple Population Sizes and Mutation Rates With Six Phenotypes
Mut
atio
n R
ate
Population Size Kin Inclusivity Level
Num
ber o
f Fix
atio
n E
vent
s
µ=0.
02
µ=0.
2
µ=1.
0
N=1000 N=5000 N=10000
1 2 3 4 5 6 1 2 3 4 5 6 1 2 3 4 5 6
1 2 3 4 5 6 1 2 3 4 5 6 1 2 3 4 5 6
1 2 3 4 5 6 1 2 3 4 5 6 1 2 3 4 5 6
Figure 7: The number of fixation events by eachof six genetically dissimilar colonies with differingkin inclusivity levels (χ = 1 through χ = 6). Panelsrepresent three mutation rates and population sizes.As the population size and mutation rate increase,colonies with higher kin inclusivity levels fix moreoften, except at the highest mutation rate. At thehighest mutation rate, larger population sizes en-able a wider range of kin inclusivity levels to reachfixation.
at all population sizes. At a high mutation rate (µ=1.0),colonies with χ = 6 tended to fix. In a few cases, other highkin inclusivity level colonies (i.e., χ = 5 or even χ = 4) wonthe competitions. However, within the moderate mutationrate (µ=0.2) treatments, the intermediate phenotypes withχ = 2 through χ = 5 were able to win the majority ofcompetitions.
Figure 7 depicts the results of our genetically dissimilartreatments with six colonies. Similar to the results pre-sented for the two-colony experiments, the higher kin in-clusivity level colonies generally outcompete the lower kininclusivity level colonies at the moderate (µ=0.2) and high(µ=1.0) mutation rates. However, at the low mutation rate(µ=0.02), the organisms with intermediate inclusivity levelsare favored by selection. At the smallest population size, amix of intermediate and high kin inclusivity level colonieswin the competitions. As the population size increases, theχ = 6 colony (the clan altruist) wins more of the competi-tions.
The geographical starting positions of the six colonies inthe initial population could potentially influence the compe-tition results. To assess this dynamic, we performed five dif-ferent sets of experiments that permuted the initial positionsof the various colonies. The starting location of each colonyis defined by which colonies directly neighbor the colony,so for the permutations we sought to test the majority ofvariations of direct neighbors for each colony. The resultsof these experiments were qualitatively identical, suggest-ing that the initial geography has little influence over thecompetition outcome.
0
10
20
30
40
50
0
10
20
30
40
50
0
10
20
30
40
50
Population Size
Ave
rage
Evo
lved
Ham
min
g D
ista
nce
Mut
atio
n R
ate
Median Hamming Distance Evolved Over Multiple Population Sizes and Mutation Rates
Mut
atio
n R
ate
Population Size
Med
ian
Ham
min
g D
ista
nce
Use
d
µ=0.
02
µ=0.
2
µ=1.
0
N=1000 N=5000 N=10000
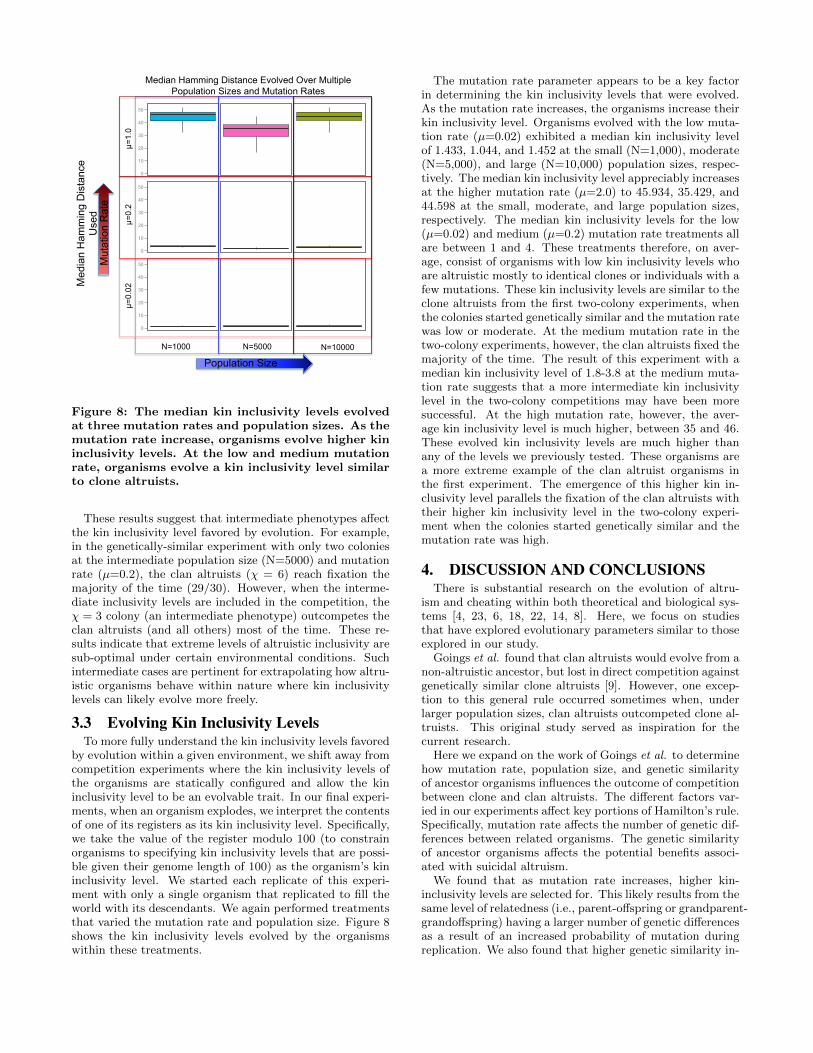
Figure 8: The median kin inclusivity levels evolvedat three mutation rates and population sizes. As themutation rate increase, organisms evolve higher kininclusivity levels. At the low and medium mutationrate, organisms evolve a kin inclusivity level similarto clone altruists.
These results suggest that intermediate phenotypes affectthe kin inclusivity level favored by evolution. For example,in the genetically-similar experiment with only two coloniesat the intermediate population size (N=5000) and mutationrate (µ=0.2), the clan altruists (χ = 6) reach fixation themajority of the time (29/30). However, when the interme-diate inclusivity levels are included in the competition, theχ = 3 colony (an intermediate phenotype) outcompetes theclan altruists (and all others) most of the time. These re-sults indicate that extreme levels of altruistic inclusivity aresub-optimal under certain environmental conditions. Suchintermediate cases are pertinent for extrapolating how altru-istic organisms behave within nature where kin inclusivitylevels can likely evolve more freely.
3.3 Evolving Kin Inclusivity LevelsTo more fully understand the kin inclusivity levels favored
by evolution within a given environment, we shift away fromcompetition experiments where the kin inclusivity levels ofthe organisms are statically configured and allow the kininclusivity level to be an evolvable trait. In our final experi-ments, when an organism explodes, we interpret the contentsof one of its registers as its kin inclusivity level. Specifically,we take the value of the register modulo 100 (to constrainorganisms to specifying kin inclusivity levels that are possi-ble given their genome length of 100) as the organism’s kininclusivity level. We started each replicate of this experi-ment with only a single organism that replicated to fill theworld with its descendants. We again performed treatmentsthat varied the mutation rate and population size. Figure 8shows the kin inclusivity levels evolved by the organismswithin these treatments.
The mutation rate parameter appears to be a key factorin determining the kin inclusivity levels that were evolved.As the mutation rate increases, the organisms increase theirkin inclusivity level. Organisms evolved with the low muta-tion rate (µ=0.02) exhibited a median kin inclusivity levelof 1.433, 1.044, and 1.452 at the small (N=1,000), moderate(N=5,000), and large (N=10,000) population sizes, respec-tively. The median kin inclusivity level appreciably increasesat the higher mutation rate (µ=2.0) to 45.934, 35.429, and44.598 at the small, moderate, and large population sizes,respectively. The median kin inclusivity levels for the low(µ=0.02) and medium (µ=0.2) mutation rate treatments allare between 1 and 4. These treatments therefore, on aver-age, consist of organisms with low kin inclusivity levels whoare altruistic mostly to identical clones or individuals with afew mutations. These kin inclusivity levels are similar to theclone altruists from the first two-colony experiments, whenthe colonies started genetically similar and the mutation ratewas low or moderate. At the medium mutation rate in thetwo-colony experiments, however, the clan altruists fixed themajority of the time. The result of this experiment with amedian kin inclusivity level of 1.8-3.8 at the medium muta-tion rate suggests that a more intermediate kin inclusivitylevel in the two-colony competitions may have been moresuccessful. At the high mutation rate, however, the aver-age kin inclusivity level is much higher, between 35 and 46.These evolved kin inclusivity levels are much higher thanany of the levels we previously tested. These organisms area more extreme example of the clan altruist organisms inthe first experiment. The emergence of this higher kin in-clusivity level parallels the fixation of the clan altruists withtheir higher kin inclusivity level in the two-colony experi-ment when the colonies started genetically similar and themutation rate was high.
4. DISCUSSION AND CONCLUSIONSThere is substantial research on the evolution of altru-
ism and cheating within both theoretical and biological sys-tems [4, 23, 6, 18, 22, 14, 8]. Here, we focus on studiesthat have explored evolutionary parameters similar to thoseexplored in our study.
Goings et al. found that clan altruists would evolve from anon-altruistic ancestor, but lost in direct competition againstgenetically similar clone altruists [9]. However, one excep-tion to this general rule occurred sometimes when, underlarger population sizes, clan altruists outcompeted clone al-truists. This original study served as inspiration for thecurrent research.
Here we expand on the work of Goings et al. to determinehow mutation rate, population size, and genetic similarityof ancestor organisms influences the outcome of competitionbetween clone and clan altruists. The different factors var-ied in our experiments affect key portions of Hamilton’s rule.Specifically, mutation rate affects the number of genetic dif-ferences between related organisms. The genetic similarityof ancestor organisms affects the potential benefits associ-ated with suicidal altruism.
We found that as mutation rate increases, higher kin-inclusivity levels are selected for. This likely results from thesame level of relatedness (i.e., parent-offspring or grandparent-grandoffspring) having a larger number of genetic differencesas a result of an increased probability of mutation duringreplication. We also found that higher genetic similarity in-

creases the competitive advantage of colonies with low kin-inclusivity levels. This effect most likely occurs as the resultof organisms with low kin-inclusivity levels (i.e., clone altru-ists) receiving a benefit from related organisms with higherkin-inclusivity (i.e., clan altruists) without reciprocating. Incontrast, genetic dissimilarity enables clan altruists to dif-ferentiate themselves from clone altruists. In addition, wefound that larger population sizes confer an advantage tocolonies with higher kin-inclusivity levels (i.e., clan altru-ists). This effect could result from larger population sizesenabling colonies to be isolated from competing colonies formore evolutionary time, thus allowing larger amounts of ge-netic dissimilarity to occur as a result of drift. This enablescolonies with higher kin inclusivity levels to successfully be-have altruistically to colony members, while killing compet-ing colonies with lower inclusivity levels. Finally, we alsofound that organisms exhibiting intermediate kin inclusiv-ity levels (i.e., levels 2-5) frequently were able to outcompetemore extreme organisms.
Greig and Travisano explored the effects of populationdensity in single-celled yeast that are known to cooperatein the wild [11]. They found that the cheating colony hadhigher fitness as the density of the population was increasedacross treatments. While this is not identical to our factor ofpopulation size, treatments with small world sizes fill morequickly than treatments with large world sizes. Therefore,small population sizes are likely to have higher populationdensities when averaged over the course of the treatmentthan larger population size treatments. When consideringthis, our study exhibits similar results across population size.Smaller population size treatments favor the clone altruistswhen the colonies are genetically similar and the clone altru-ists can be considered to be “cheating” on the clan altruists.
Two notable studies suggest that lower mutation rates fa-vor altruism. First, Spector and Klein [21] explored howmutation rate and territorial structure affected competitionbetween groups of kin altruists. The territorial structuredetermined how many neighbors along a ring an individualorganism had the chance of being paired with. An organ-ism’s tag, which was inherited at birth, indicated the kingroup of the organism. Organisms donated to other neigh-bors whose tags were within a tolerance amount of their own.Given these parameters, they found that tag-mediated al-truism was more likely to evolve with lower mutation ratesand smaller interaction radii. However, the salient differ-ence from our work is that when the organisms’ tags weremutated they were changed to a random value. Thus, thedistance between tags (used to establish whether or not anorganism would donate) did not measure genetic similarityunless the organisms had identical tags. In contrast, withinour study, the edit distance between two genomes is a perfectmeasure of similarity.
Paolucci and Conte studied the effect of roost size and mu-tation rate on the evolution of altruism in simulated vampirebats [19]. Within this study, the mutation rate determinedthe probability that an offspring organism would be an al-truist or a cheater. They found that high mutation ratesincreased the invasion of cheaters and that smaller roostsizes decreased the invasion rates. The apparent differencebetween these conclusions and those put forth by our studyresults from the implementation of the mutation operator.Our mutation operator can increase genetic differences be-
tween a parent and offspring without affecting whether ornot the organism behaves altruistically.
There are many possible future directions for this study.First, to get a better understanding of the evolution of kin in-clusivity levels in spatially-structured populations, we couldexplore how the distribution of kin inclusivity levels variesover space. Second, most organisms in nature have someform of communication. By allowing our digital organismsto communicate, it is likely that they will evolve to exhibit amore sophisticated strategy for determining when to behavealtruistically. Within our current experiments, often organ-isms will explode when they are completely surrounded bytheir kin, due to the stochastic nature of the instruction. Ifwe provide them with the ability to sense related (or un-related) organisms around them, then new and interestingstrategies that make use of the explosion instructions maydevelop, such as facultative cheating [6]. Finally, currently,the organisms are not able to move. If able to move in the en-vironment, the organisms may strategically place themselvesnear unrelated organisms before executing the explode in-structions, similar to various species of eusocial insects thatstrategically explode when a predator draws near [20].
5. ACKNOWLEDGEMENTSWe thank the DevoLab and BEACON Center for the
Study of Evolution in Action for advice and support. Thiswork was funded in part by the Clare Boothe Luce Founda-tion and the NSF. This material is based in part upon worksupported by the National Science Foundation under Coop-erative Agreement No. DBI-0939454 and No. OCI-1122620.Any opinions, findings, and conclusions or recommendationsexpressed in this material are those of the author(s) anddo not necessarily reflect the views of the National ScienceFoundation.
6. REFERENCES[1] L. Chao and B. R. Levin. Structured habitats and the
evolution of anticompetitor toxins in bacteria.Proceedings of the National Academy of Sciences,78(10):6324–6328, Oct. 1981.
[2] J. Clune, H. J. Goldsby, C. Ofria, and R. T. Pennock.Selective pressures for accurate altruism targeting:evidence from digital evolution for difficult-to-testaspects of inclusive fitness theory. Proceedings of theRoyal Society B: Biological Sciences,278(1706):666–674, Jan. 2011.
[3] R. Dawkins. The Selfish Gene. 2006.
[4] S. P. Diggle, A. S. Griffin, G. S. Campbell, and S. A.West. Cooperation and conflict in quorum-sensingbacterial populations. Nature, 450(7168):411–414,2007.
[5] E. Fehr and U. Fischbacher. The nature of humanaltruism. Nature, 425(6960):785–791, Oct. 2003.
[6] F. Fiegna and G. J. Velicer. Competitive fates ofbacterial social parasites: persistence and self-inducedextinction of Myxococcus xanthus cheaters.Proceedings of the Royal Society B: BiologicalSciences, 270(1523):1527–1534, July 2003.
[7] K. R. Foster, T. Wenseleers, and F. L. W. Ratnieks.Kin selection is the key to altruism. Trends in Ecology& Evolution, 2006.

[8] O. M. Gilbert, K. R. Foster, N. J. Mehdiabadi, J. E.Strassmann, and D. C. Queller. High relatednessmaintains multicellular cooperation in a social amoebaby controlling cheater mutants. Proceedings of theNational Academy of Sciences, 104(21):8913–8917,May 2007.
[9] S. Goings, J. Clune, C. Ofria, and R. T. Pennock. Kinselection: The rise and fall of kin-cheaters. InProceedings of the Ninth International Conference onArtificial Life, pages 303–308, 2004.
[10] H. J. Goldsby, A. Dornhaus, B. Kerr, and C. Ofria.Task-switching costs promote the evolution of divisionof labor and shifts in individuality. Proceedings of theNational Academy of Sciences, 109(34):13686–13691,Aug. 2012.
[11] D. Greig and M. Travisano. The Prisoner’s Dilemmaand polymorphism in yeast SUC genes. Proceedings ofthe Royal Society B: Biological Sciences, 271(Suppl3):S25–S26, Feb. 2004.
[12] W. D. Hamilton. The evolution of altruistic behavior.The American Naturalist, 1963.
[13] W. D. Hamilton. The genetical evolution of socialbehaviour. II. Journal of theoretical biology, 1964.
[14] A. Khare, L. A. Santorelli, J. E. Strassmann, D. C.Queller, A. Kuspa, and G. Shaulsky.Cheater-resistance is not futile. Nature,461(7266):980–982, Sept. 2009.
[15] L. Lehmann and L. Keller. The evolution ofcooperation and altruism – a general framework and aclassification of models. Journal of EvolutionaryBiology, 19(5):1365–1376, Sept. 2006.
[16] R. E. Lenski, C. Ofria, R. T. Pennock, and C. Adami.The evolutionary origin of complex features. Nature,423:139–144, May 2003.
[17] C. Ofria and C. O. Wilke. Avida: A software platformfor research in computational evolutionary biology.Artificial Life, 10(2):191–229, Mar. 2004.
[18] E. A. Ostrowski, M. Katoh, G. Shaulsky, and D. C.Queller. Kin discrimination increases with geneticdistance in a social amoeba. PLOS Biology, 2008.
[19] M. Paolucci and R. Conte. Roost size for multilevelselection of altruism among vampire bats. InMulti-Agent-Based Simulation VII, pages 69–79.Springer Berlin Heidelberg, Berlin, Heidelberg, 2007.
[20] J. R. Shorter and O. Rueppell. A review onself-destructive defense behaviors in social insects.Insectes Sociaux, 59(1):1–10, Dec. 2011.
[21] L. Spector and J. Klein. Genetic stability andterritorial structure facilitate the evolution oftag-mediated altruism. Artificial Life, 12(4), Oct.2006.
[22] J. E. Strassmann, O. M. Gilbert, and D. C. Queller.Kin discrimination and cooperation in microbes.Annual Review of Microbiology, 65(1):349–367, Oct.2011.
[23] G. J. Velicer, L. Kroos, and R. E. Lenski.Developmental cheating in the social bacteriumMyxococcus xanthus. Nature, 404(6778):598–601,2000.
[24] S. A. West, A. S. Griffin, and A. Gardner. Socialsemantics: altruism, cooperation, mutualism, strongreciprocity and group selection. Journal ofEvolutionary Biology, 20(2):415–432, Mar. 2007.




















