
Foraging ecology of Nilgiri Langur (Trachypithecus Johnii) in ...

OIKOS 90: 399-407. Copenhagen 2000
The effects of gerbil foraging on the natural seedbank and consequences on the annual plant community
Christopher J. Lortie, David T. Ganey and Burt P. Kotler
Lortie, C. J., Ganey, D. T. and Kotler, B. P. 2000. The effects of gerbil foraging on the natural seedbank and consequences on the annual plant community. - Oikos 90: 399-407.
We explicitly tested the following predictions using Gerbillus allenbyi and G. pyra- midum foraging on artificial arrays of natural seedbank on a semi-stabilized sand dune: 1. that the gerbils will forage using a quitting harvest rate rule, 2. that larger seeds are preferred due to higher encounter rates, and 3. that there are community level consequences for the annual plants as a direct result of foraging by the desert rodents. Natural seedbank, separated into two size classes, was placed in seed trays in the field at four different densities (1/16 x, 1/4 x, 1 x, and 2 x normal). Follow- ing exposure to granivory, the remaining seeds were weighed and germinated to test for community level effects. Only the 1 x and 2 x normal density plots were heavily foraged, and at both seed sizes, which suggests that the gerbils employed a quitting harvest rate rule. In support of our second prediction, the two species of gerbils tended to consume more of the larger seeds, particularly at higher densities. The mean number and total number of plant species that germinated in plots exposed to granivores was not significantly different from unexposed samples. At the community level however, there was no net association of germinated plant species in the four treatment groups exposed to granivores, but a significant net positive association in the unexposed control seedbank. Gerbil foraging on annual plant seedbank may thus subtly change the entire community structure (from positive to neutral), although not necessarily shift the species distributions significantly. Our results corroborate other studies involving artificial food types such as millet and suggest that foraging decisions may affect the plant community.
C. J. Lortie, Dept of Botany, Univ. of British Columbia, Vancouver, BC, Canada V6T 1Z4 ([email protected]). - D. T. Ganey and B. P. Kotler, Ben-Gurion Univ. of the Negev, Blaustein Inst., for Desert Research, Mitrani Dept for Desert Ecology, Sede Boker Campus 84990, Israel.
Foraging theory and optimal patch use strategies are important avenues of research that significantly increase our understanding of competitive interactions and spe- cies coexistence (MacArthur and Pianka 1966, Tilman 1982, Kotler and Brown 1988, Werner 1992, Brown et al. 1994). Extensive work investigating desert rodent communities indicated that they are ideally suited to experiments that involve manipulating food resources and patch quality (Brown 1975, Rosenzweig 1977, Price and Brown 1983, Kotler and Brown 1988). An excellent example of this is provided by the desert rodents Gerbillus allenbyi (Allenby's gerbil) and G. pyramidum (greater
Egyptian sand gerbil) which have been shown to compete (Rosenzweig and Abramsky 1985, 1986, Abramsky and Pinshow 1989, Abramsky et al. 1990, 1991, Mitchell et al. 1990) and partition habitat use both temporally and spatially (Kotler et al. 1993a, Ziv et al. 1993, Brown et al. 1994). Nonetheless, these studies typically employed artificial food sources and often did not re-integrate foraging decisions back into the context of the actual plant community as a whole. Foraging may be particu- larly important to the community in arid systems since desert rodents have been shown to harvest up to 86.5% of total seed production (Chew and Chew 1970).
Accepted 15 February 2000 Copyright ? OIKOS 2000 ISSN 0030-1299 Printed in Ireland - all rights reserved
OIKOS 90:2 (2000) 399

A commonly used method to investigate mechanisms of species coexistence is the giving-up density (GUD). In essence, it is the resource density at which a forager leaves the foraging patch (Brown 1988). If the forager is able to assess patch quality, then the GUD should estimate foraging efficiency in terms of costs and benefits (Stephens and Krebs 1986, Brown 1988). GUDs have been successfully used to measure the foraging efficiency of a wide variety of organisms (i.e., Brown et al. 1992) and thereby infer patch use strate- gies (Valone and Brown 1989). The strategies proposed and tested to date have been 1. leaving a patch at a fixed quitting harvest rate (forager estimates patch quality), 2. after a fixed time, and 3. after a fixed amount of harvest (Brown and Mitchell 1989). Garb et al. (2000) showed that G. allenbyi and G. pyramidum used a quitting harvest rate rule when foraging in artificial patches containing millet. We feel that the next logical step is to test for patch use strategies in more complex resource patches. Incorporating elements such as different sized seeds and particularly use of the actual food types obtained from the natural seedbank in the field are highly suitable manipulations of patch complexity. Accordingly, we have focussed the experi- ment here on specifically increasing patch complexity to make the techniques more "realistic". In addition to the five mechanisms of coexistence proposed by Brown et al. (1994) (all habitat based), use of natural seedbank may reveal other mechanisms of coexistence involving diet selection, e.g. Kotler and Brown (1999).
We explicitly tested the following predictions using G. allenbyi and G. pyramidum foraging on artificial arrays of natural seedbank on a semi-stabilized sand dune in the Negev Desert, Israel: 1. that the gerbils will forage using a quitting harvest rate rule (similar to tests using millet, Brown et al. 1994, Garb et al. 2000), 2. that larger seeds are preferred due to higher encoun- ter rates (i.e., Garb et al. 2000), and 3. that there are community level consequences for the annual plants as a direct result of foraging by the desert rodents. Our primary objective was to assess whether optimal forag- ing decisions were made on seeds in their natural setting and to place this in the context of the emergent plant community. This is a considerable extension to the majority of studies in this body of literature as it tests whether seed tray techniques and GUDs on artifi- cial food and controlled conditions correspond to field data.
Methods
Description of study site The field component of this study was carried out at Bir Asluj (3100.2'N, 34?44.5E') in the Holot Mashabim Nature Reserve in the central Negev Desert. The site
for seed collection and implementation of feeding trials was a semi-stabilized sand dune dominated by annuals and sparsely distributed shrubs. The two main shrubs were Artemisia monosperma and Retama raetam (Abramsky et al. 1985). Typically, there were only 16 species of annual plants regularly represented at this particular site (Lortie and Turkington pers. comm.). Rainfall occurs mainly between December and April, with a long-term average of 110 mm per year. Mean daily low and high temperatures ranged from 9?C to 35?C. The following rodent species are commonly found in this community: Gerbillus allenbyi, G. pyra- midum, G. henleyi (pygmy gerbil), and Meriones sacra- menti (Buxton's jird). These rodents are all primarily granivorous and nocturnal (Bar et al. 1984).
Seed collection and seedbank properties The seed used in this experiment was almost exclusively from the winter annual plants. Seed was collected 17 May 1997, immediately after the last of the annual plants set seed and began primary seed dispersal. In desert annual grasslands, 99% of all seeds occur in the top 5 mm of sand (Venable 1989). The seedbank was collected using a sand collector, which is a metal wedge- shaped box open at the front with a sharpened edge at the bottom of the open face which effectively removes the top 1 cm of sand (thereby conservatively sampling the entire seedbank). The seed/sand mixture was stored in a fenced enclosure at the Mitrani Department of Ecology (25 km SE of the study site) for the duration of the summer. The seeds were thus subject to natural temperature and moisture fluctuations, but free from gerbil foraging.
In February 1998, before the beginning of the feeding trials, the sand was sieved with 2-mm and 0.5-mm mesh sieves to isolate the seeds. These two mesh sizes cap- tured virtually all the seeds (pers. obs. with dissecting scope), and sorted them into the following two cate- gories: large (> 2 mm), and small (> 0.5 mm but less than or equal to 2 mm) seeds. The collection surface area, total mass of sand, and total seedbank mass were recorded; the seed mass per m2 was calculated to esti- mate the gross normal seed density for each size class in the field. The normal seedbank density for 0.25-m2 experimental plots was 150.2 g of seed which was composed of 55.4 g of 2-mm seeds and 94.8 g of 0.5-mm seeds. These masses also include organic debris.
Seed consumption trials Four replicates of four densities of the natural seed- bank were exposed to granivores from 27 February to 1 March 1998 on the same sand dune at Bir Asluj where the seeds were collected. The experiment was conducted
OIKOS 90:2 (2000) 400

during the new moon cycle to maximize gerbil forag- ing (Kotler et al. 1993b) and timed to correspond with natural seedbank germination. Foraging immedi- ately before and during germination is likely the most important for the emergent plant community as it affects the composition of seeds at the surface that are in the process of germinating. Accordingly, only one set of replicates is possible that corresponds with both the new moon and germination which lasts ap- proximately three weeks (Lortie and Turkington pers. comm.). The densities were as follows: 1/16 x, 1/4 x, 1 x, and 2 x of normal density (as calculated for that sand dune - mass of seeds per m2). The alu- minum seed trays were 45 x 60 x 2.5 cm in size and were filled with 5 L of seedless sand (sieved to re- move seeds that may already be present) from the exact same location of the tray. The appropriate mass of seed for that density treatment was then added and thoroughly mixed throughout the tray. Transects were placed along the sand dune parallel to the crest, and the trays were all placed at least 1.5 m away from the nearest shrub. The remaining seeds were col- lected the following mornings at sunrise. Based on the activity pattern and spoor evident in the morning, the granviores were almost exclusively G. allenbyi and G. pyramidum.
Subsequent plant community The remaining seeds from each tray were sieved to the two size classes, weighed, recombined, and then planted along with control seedbank (unexposed to rodents) in a greenhouse at The Mitrani Department using a randomized block design. Every replicate of each seed density for each day along with controls was tested giving a total of 80 plots (5 treatment levels x 4 replicates x 4 days). The planting density for all treatment levels was 1/10th normal density in 0.5 x 0.5 m flats with clean sand from the same sand dune as substrate; the seeds were uniformly spread out within the trays and lightly covered. This planting density ensures that virtually all the seeds will germi- nate (Lortie and Turkington pers. comm.) and tests for differences in germination among seeds at con- stant densities. The trays were watered daily to satu- ration. After all the seeds had germinated, the seedlings were identified to species. Density for each species was recorded. Subsequent sieving verified that all seeds had indeed germinated.
Statistical analyses All statistics were performed with JMP Version 3.2.2 (SAS 1997). One-way ANOVAs were used to test for differences among the various densities in terms of
GUDs and proportion of seeds left by size following foraging. The ratios of the GUDs to each other were also tested with t-tests to determine if the initial pro- portional abundance of seeds was preserved even af- ter foraging, i.e., 2 x plots still had twice as many seeds compared to 1 x etc. This tests if the forager simply spent a fixed amount of time in each patch and reduced each by the same proportion (and hence the forager cannot adequately assess harvest rate; Iwasa et al. 1981) or whether it employed a quitting harvest rate rule and tended to equalize GUDs across different initial densities.
We used Manly's index for selectivities (1974), as modified for depletable resources (Chesson 1983), to determine if the gerbils preferred either of the two seed size classes we measured (> 2 mm; < 2 mm and > 0.5 mm) (Brown and Morgan 1995, Garb et al. 2000). The equation is:
S = ln(N1/R1)/(ln(N1/R,) + ln(N/2R2)),
where S is the selectivity for food type 1, N1 and N2 are the GUDs for food type 1 and 2, respectively, and RI and R2 are the initial abundances. The range of S is 0 to 1; S = indicates an absence of food type 1, S = 1 indicates an absence of food type 2, and S = 0.5 indicates no selectivity for either seed size in this case. Paired t-tests were used to test if the selec- tivity for small seeds was significantly different from 0.5 at each of the four densities we presented to ger- bils.
The density and species of the plants germinated from the remaining seeds were recorded. A one-way ANOVA was used to test for differences in total seedling density among the four densities and the control. Analysis of covariance was used to test for differences in mean frequency of species present and the total number of plant species present (with actual density of germinated seedlings as covariate). The to- tal number of germinated plants was also regressed against number of species. We used Schluter's (1984) variance ratio technique to assess if any competitive or facilitative effects likely existed among the seedlings. More specifically, we looked for net (+) or net (-) associations at the community level associ- ated with selective seed predation by rodents. The net interaction measurement simply uses a variance ratio (VR); a VR ratio of 1 is no net interaction, VR < 1 is net negative, and VR> 1 is net positive (for more details see Schluter 1984). If the gerbils foraged dif- ferently on different density patches or on different sized seeds, then the structure of the plant community may be altered as estimated by the net interactions. Since Sequential Bonferroni corrections (Rice 1989) for multiple comparisons did not change significance at the 0.05 level, unadjusted p-values are reported.
OIKOS 90:2 (2000) 401
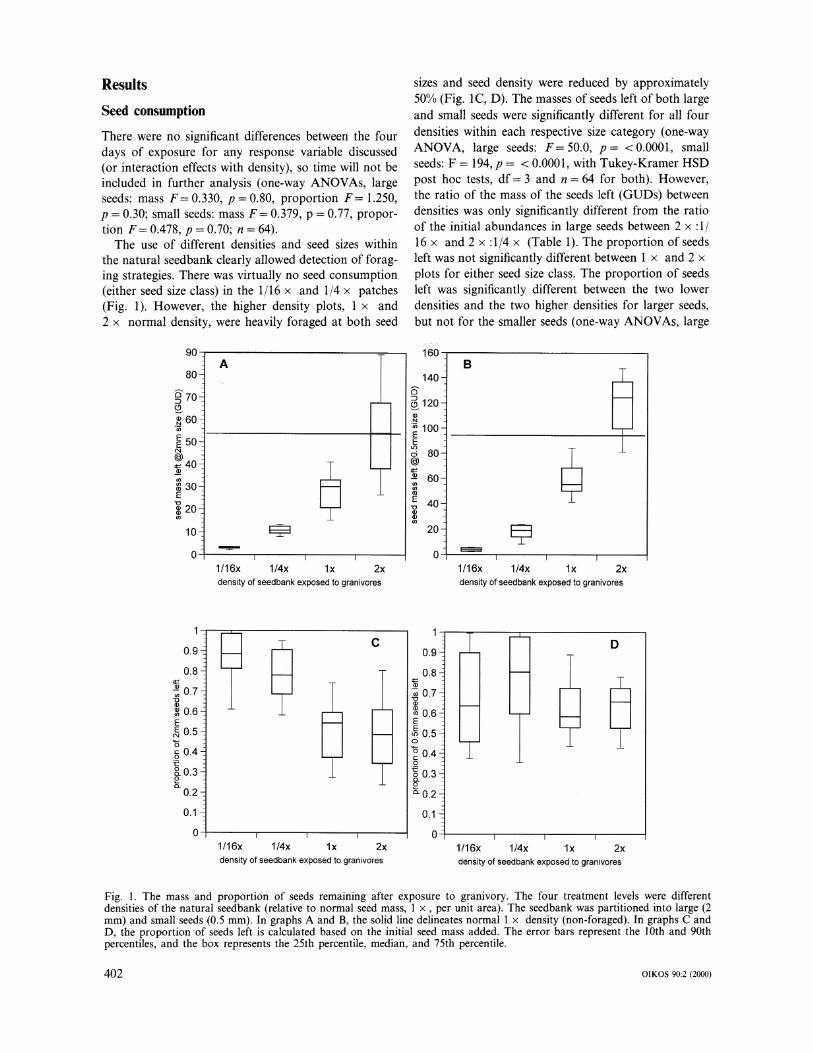
Results Seed consumption There were no significant differences between the four days of exposure for any response variable discussed (or interaction effects with density), so time will not be included in further analysis (one-way ANOVAs, large seeds: mass F= 0.330, p = 0.80, proportion F= 1.250, p = 0.30; small seeds: mass F= 0.379, p = 0.77, propor- tion F= 0.478, p = 0.70; n = 64).
The use of different densities and seed sizes within the natural seedbank clearly allowed detection of forag- ing strategies. There was virtually no seed consumption (either seed size class) in the 1/16 x and 1/4 x patches (Fig. 1). However, the higher density plots, 1 x and 2 x normal density, were heavily foraged at both seed
density of seedbank exposed to granivores
1/16x 1/4x 1x 2x density of seedbank exposed to granivores
sizes and seed density were reduced by approximately 50% (Fig. 1C, D). The masses of seeds left of both large and small seeds were significantly different for all four densities within each respective size category (one-way ANOVA, large seeds: F=50.0, p= <0.0001, small seeds: F = 194, p = < 0.0001, with Tukey-Kramer HSD post hoc tests, df= 3 and n = 64 for both). However, the ratio of the mass of the seeds left (GUDs) between densities was only significantly different from the ratio of the initial abundances in large seeds between 2 x :1/ 16 x and 2 x :1/4 x (Table 1). The proportion of seeds left was not significantly different between 1 x and 2 x plots for either seed size class. The proportion of seeds left was significantly different between the two lower densities and the two higher densities for larger seeds, but not for the smaller seeds (one-way ANOVAs, large
density of seedbank exposed to granivores
1/16x 1/4x 1x 2x density of seedbank exposed to granivores
Fig. 1. The mass and proportion of seeds remaining after exposure to granivory. The four treatment levels were different densities of the natural seedbank (relative to normal seed mass, 1 x, per unit area). The seedbank was partitioned into large (2 mm) and small seeds (0.5 mm). In graphs A and B, the solid line delineates normal 1 x density (non-foraged). In graphs C and D, the proportion of seeds left is calculated based on the initial seed mass added. The error bars represent the 10th and 90th percentiles, and the box represents the 25th percentile, median, and 75th percentile.
402 OIKOS 90:2 (2000)

Table 1. A representative sample of the ratio of GUDs as compared to the initial abundances of seed masses for the four different densities.
Seed size and ratio Expected ratio Mean ratio t-test statistic p-value
Large seeds 2 x :1 x 2 3.33 -0.992 0.83 2 x :/4 x 8 5.37 4.955 <0.0001 2x:1/16x 32 18.46 6.786 <0.0001 Small seeds 2x:1 x 2 2.04 -0.378 0.64 2 x:1/4 x 8 7.56 0.454 0.33 2x:1/16 x 32 34.02 -0.594 0.72
seeds: F= 17.3, p= <0.0001, small seeds: F= 1.51, p = 0.22, df = 3, n = 64, with Tukey-Kramer post hoc tests). The selectivity for small seeds was not signifi- cantly different from a null of 0.5 (paired t-test, t= - 0.127, p = 0.45, n = 4).
Subsequent annual plant community
Although all treatment levels were planted in the green- house at the same low density to promote maximum germination (1/10th normal seed mass for 0.5 x 0.5-m plot), the seeds taken from the plots that were exposed to granivores versus those from unexposed seedbank had significantly less germinated plants (Fig. 2, one-way ANOVA, F= 4.46, p = 0.0027, df= 4, n = 80). Interest- ingly, the samples exposed to granivory did not differ in numbers of germinated seeds regardless of initial den- sity. This is strong evidence that granivores equalized GUDs. The mean number of plant species that germi- nated in plots exposed to granivores was not signifi- cantly different from unexposed samples (Fig. 3, ANCOVA, F= 0.556, p = 0.70), and the total number of species present was also not significantly different between the different densities (Fig. 4, ANCOVA, F= 0.556, p = 0.70). However, in general when all germina- tion data were pooled, density does act as a significant predictor of number of species (positive relationship, Simple linear regression, R2 = 0.641, p< 0.0001, n= 80).
At the community level, there was no net association of germinated plant species in the four treatment groups exposed to granivores, but a significant net positive association in the unexposed control seedbank (Table 2, variance ratio technique, significance at p = 0.05 level). Granivory therefore significantly changed the community structure of the plants from net positive (control seedbank) to no net association between plant species (all four exposed densities) thereby removing consistent positive associations of species within the community. Inspection of correlation matrices also showed that no particular set of positive associations of species were removed but that there was a general reduction in the frequency of positive correlations par- ticularly between large and small seeded plants.
Discussion The first objective of this study was to assess if the findings involving foraging decisions of gerbils with millet as a food item (e.g., Brown et al. 1994, Garb et al. 2000) corresponds to that of the actual food type, the natural seedbank. Possible potential foraging strate- gies available to gerbils include leaving a patch at a fixed quitting harvest rate, at a fixed time, or at a fixed amount of harvest (Brown and Mitchell 1989). In gen- eral, the two gerbils G. allenbyi and G. pyramidum have been found to use quitting harvest rate rules (Brown et al. 1994, Garb et al. 2000) and can therefore assess patch quality. We also found that gerbils foraged differ- ently on different density patches and likely employed quitting harvest rate rules in that relatively higher den- sity patches (high, 2 x, and normal, 1 x ) were heavily consumed while lower density patches were not (Fig. 1). This indicates that the gerbils tended to equalize the GUDs.
250
200
., 150 cn
-o -a 100
1/16x 1/4x 1x 2x control treatment levels exposed to granivores
Fig. 2. Plant density of different treatment levels exposed to granivores. The four treatment levels were different densities of the seedbank, and the control was not exposed to grani- vores. Note that all treatment levels were planted at 1/10th normal density.
OIKOS 90:2 (2000) 403

._0.4 __ 10. - ' 0.3- c 0 C
te 0.2-
E I 0.1 -
0-I I I I 1/16x 1/4x 1x 2x control
treatment levels exposed to granivores
Fig. 3. The mean frequency of plant species germinated after exposure to granivores. The four treatment levels were differ- ent densities of the seedbank, and the control was unexposed. All treatment levels were planted at the same density, 1/10th of normal.
Interestingly however, there may be different strate- gies with respect to large and small seeds. The ratio of the GUDs compared to the initial abundance of seeds for each density was significantly reduced for larger seeds (i.e., the ratio of 2 x :1/16 x was reduced to 18.5 versus the initial proportionate difference of 32) while smaller seeds did not significantly differ from the initial relative ratios (Table 1). Similarly, the proportion of seeds left was significantly different between the high and normal densities and the two lower densities for larger seeds, but not for smaller seeds (Fig. 1C, D). Not surprisingly, there was no significant difference between the 1 x and 2 x plots for large seeds (or small seeds) as the magnitude of difference of mass is only two; whereas all other comparisons made here are at least a factor of eight (Table 1). There are at least three plausible explanations. 1. The gerbils do not discrimi- nate to this level (2 x differences) of seed mass. Valone and Brown (1989) showed that several granivores em- ployed a quitting harvest rate strategy when foraging on patches with high variance, but a fixed time strategy while foraging on patches that had low initial variance. This suggests that 2 x vs 1/4 x is sufficient variation, but 2 x vs 1 x may not be sufficient to detect a quitting harvest rate. 2. The gerbils cannot discriminate between these two densities of seed mass; or 3. the differences in seed mass between 1 x and 2 x are not actually two-fold. The measured seed masses also in- cluded some debris which may decrease the ability to compare masses this closely. Furthermore, equal num- bers of seeds were germinated from the various densi-
ties exposed to granivores (Fig. 2). This also strongly suggests that the GUDs between the two higher densi- ties (1 x and 2 x ) are not actually different but for the organic matter and that gerbils may even be equalizing GUDs across lower densities (1/4 x and 1/16 x ). What is most important here is that the gerbils clearly as- sessed patch quality. It is therefore reasonable to sug- gest that a quitting harvest rate rule is used for larger seeds, but not for smaller seeds. A fixed amount of search time may be used for small seeds at all patch qualities, or more likely, small seeds are simply eaten incidentally since encounter probabilities are much higher for larger seeds. Using cracked wheat seeds on the same species of gerbils, Garb et al. (2000) did find that encounter probabilities were almost 20 times higher on large versus medium seeds and 2-5 times higher on medium seeds relative to small seeds.
The apparent lack of significant differences in selec- tivity for smaller seeds with density also suggests that smaller seeds are consumed for a fixed amount of time. Hence, there is no selectivity against smaller seeds per se. Consumption of the natural seedbank by gerbils may generate a (-, 0) interaction between the two seed sizes tested here. Brown and Mitchell (1989) and Garb et al. (2000) however found that subsequent community structure of two predator species may be strongly influ- enced by short-term apparent competition (-, -) for patchy shared resources whereby foraging by a shared predator (granivore) for one food type depresses the abundance of all food types. This suggests that different species of large seeded plants may have a (-, -) interaction. Nonetheless, the preference here for larger seeds (based on the proportion of seeds remaining, see Fig. 1) by the gerbils supports a similar trend across a broad range of animals (sparrows: Pulliam 1975, squir- rels: Thorson 1995, gerbils: Garb et al. 2000, and granivorous birds: Howe and Brown 1999).
Experiments involving tests of foraging strategies such as those proposed by Brown and Mitchell (1989) and tests of mechanisms of coexistence as presented by Brown et al. (1994) have all relied on the use of GUDs on artificial food sources (such as millet) as the key tool to measure foraging behavior (i.e., Mitchell et al. 1990, Kotler et al. 1993a, Ziv et al. 1993, Brown et al. 1994, Garb et al. 2000). To the best of our knowledge, this study is the first to test if the same foraging strategies apply to the granivore in the field with the natural food type. Our results using natural seed support the general trend in the literature in that these two gerbil species employed an optimal foraging strategy and exhibited a seed size preference that corresponds to predictions based on energetics and encounter rates (Abramsky and Pinshow 1989, Mitchell et al. 1990, Abramsky et al. 1991, Kotler et al. 1993a, Garb et al. 2000). More specifically, the findings here correspond with the study by Garb et al. (2000) which used millet seeds and showed that the gerbils harvested a higher proportion
OIKOS 90:2 (2000) 404

Fig. 4. The total number of plant species germinated after seeds were exposed to granivores (except control). The number of germinated plants is at 1/10th normal seed density and germinated in 0.5 x 0.5-m plots. The four different density curves represent the different seed densities that were exposed to granivores in the field.
0 50 100 150 200 250 300 number of germinated plants
c- U, o 0
a)
0 0 E 2~ t- 0
0 50 100 150 200 250 300 number of germinated plants
of larger seeds than smaller seeds when the two seed sizes co-occurred within the same patch. Hence, the use of GUDs also appears to be an appropriate method for foraging on natural seed since differences were detectable even with plant material present in the sample and under field conditions.
The second objective of this study was to assess if foraging decisions made by the gerbils could affect the subsequent emergent plant community. The seeds that were exposed to gerbils had significantly fewer individual plants germinate (Fig. 2). This is likely due to a reduction in the amount of actual seed present relative to the control. Simply put, the gerbils consumed the seeds and left behind the other organic matter such as floral parts, flower heads, and seed cases or pods. Therefore, the control seeds we planted actually had more seeds per gram. The mean frequency of the average number of species present or total number of species present in
foraged plots was not significantly different from control seedbank (Figs 3, 4). Nonetheless, there was weak evidence for a general reduction in both species estimates in the treatment plots.
Table 2. The variance ratio (VR) technique (Schluter 1984) to estimate net interactions between germinated plants exposed to gerbils at four different densities and with a control that was not subject to granivory. A VR ratio of 1 is no net interaction, VR < 1 is net negative, and VR > 1 is net positive. Significance is at the 0.5 level.
Initial density Calculated variance Test statistic ratio
2x 0.7621 12.19 1 x 0.6436 10.30 1/4 x 0.6625 10.60 1/16 x 0.6008 9.61 control 1.2666 20.56*
*p<0.05.
OIKOS 90:2 (2000)
c.
0
0. C,
(I) 0 0 .0 E c
405

The change from net positive to no significant associ- ation between plants when exposed to gerbils (Table 2) suggests that either a general reduction in seed density per patch or the selective foraging for larger seeds by gerbils may significantly restructure the annual plant community. For instance, a net positive association suggests that the plants may either be selecting similar micro-sites (commonly occur together due to abiotic factors) or that certain key species within the commu- nity may be facilitating or interacting positively with other species to increase germination. The preferential removal of larger seeds from the seedbank by gerbils shifted the positive interaction between large and small seeds from facilitating increased establishment to com- petition between more similar sized smaller individuals. Depending on gerbil movements, foraging intensity, potential dispersal, and movement of seeds via wind and sand movement, foraging may shift the balance point within the annual community from facilitation to competition.
In summary, these two species of gerbils do employ harvest rate rules in foraging on patches of annual seeds. A seed size preference exists in favour of larger seeds which may subtly influence the balance within the annual plant community. Although more time consum- ing and perhaps more difficult, further work that in- volves the use of natural diet items over artificial food types is suggested as preferences and foraging decisions likely affect the community at other scales. This experi- ment did however serve as a "check" on artificial food types and showed that there is a strong correspondence with natural seed in the field. The interaction between the preference for different seed sizes and the annual plants is particularly interesting, and experiments that explicitly test for these interactions will broaden the scope and impact of optimal foraging theory.
Acknowledgements - This work was supported by the United States-Israel Bi-national Science Foundation (grant 97-0427 to BPK), and CJL was funded by the Blaustein Center for Scientific Cooperation. This is publication no. 288 of the Mitrani Dept of Desert Ecology.
References Abramsky, Z. and Pinshow, B. 1989. Changes in foraging
effort in two gerbil species with habitat type and intra- and interspecific activity. - Oikos 56: 43-53.
Abramsky, Z., Brand, B. and Rosenzweig, M. L. 1985. Geo- graphical ecology of gerbilline rodents in the sand dune habitats of Israel. - J. Biogeogr. 12: 363-372.
Abramsky, Z., Rosenzweig, M. L., Pinshow, B. et al. 1990. Habitat selection: an experimental field test with two gerbil species. - Ecology 71: 2358-2369.
Abramsky, Z., Rosenzweig, M. L. and Pinshow, B. 1991. The shape of a gerbil isocline measured using principles of optimal selection. - Ecology 72: 329-340.
Bar, Y., Abramsky, Z. and Gutterman, Y. 1984. Diet of gerbilline rodents of the Israeli desert. - J. Arid Environ. 7: 371-376.
Brown, J. H. 1975. Geographical ecology of desert rodents. - In: Cody, M. L. and Diamonds, J. M. (eds), Ecology and evolution of communities. Harvard Univ. Press, pp. 315- 341.
Brown, J. S. 1988. Patch use as an indicator of habitat preference, predation risk, and competition. - Behav. Ecol. Sociobiol. 22: 37-47.
Brown, J. S. and Mitchell, W. A. 1989. Diet selection on depletable resources. - Oikos 54: 33-43.
Brown, J. S. and Morgan, R. A. 1995. Effects of foraging behavior and spatial scale on diet selectivities: a test with fox squirrels. - Oikos 74: 122-136.
Brown, J. S., Morgan, R. A. and Dow, B. D. 1992. Patch use under predation risk II: a test with fox squirrels, Sciurus niger. - Ann. Zool. Fenn. 29: 311-318.
Brown, J. S., Kotler, B. P. and Mitchell, W. A. 1994. Foraging theory, patch use, and the structure of a Negev desert granivore community. - Ecology 75: 2286-2300.
Chesson, J. 1983. The estimation and analysis of preference and its relation to foraging models. - Ecology 64: 1297- 1304.
Chew, R. M. and Chew, A. 1970. Energy relationships of the mammals of a desert shrub (Larrea tridentata) community. - Ecol. Monogr. 40: 1-21.
Garb, J., Kotler, B. P. and Brown, J. S. 2000. Foraging and community consequences of seed size for coexisting Negev Desert granviores. - Oikos 88: 291-300.
Howe, H. F. and Brown, J. S. 1999. Effects of birds and rodents on synthetic tallgrass communities. - Ecology 80: 1776-1781.
Iwasa, Y. M., Higashi, M. and Yamamura, N. 1981. Prey distribution as a factor determining the choice of optimal foraging strategy. - Am. Nat. 117: 710-723.
Kotler, B. P. and Brown, J. S. 1988. Environmental hetero- geneity and the coexistence of desert rodents. - Annu. Rev. Ecol. Syst. 19: 281-307.
Kotler, B. P. and Brown, J. S. 1999. Mechanisms of coexis- tence of optimal foragers as determinants of the local abundance and distribution of desert granivores. - J. Mammal. 80: 361-374.
Kotler, B. P., Brown, J. S. and Subach, A. 1993a. Mechanisms of species coexistence of optimal foragers: temporal parti- tioning by two species of sand dune gerbils. - Oikos 67: 548-556.
Kotler, B. P., Brown, J. S. and Mitchell, W. A. 1993b. Environmental factors affecting patch use in gerbilline rodents. - J. Mammal. 74: 614-620.
MacArthur, R. and Pianka, E. 1966. On optimal use of a patchy environment. - Am. Nat. 100: 603-609.
Manly, B. F. 1974. A model for certain types of selection experiments. - Biometrics 30: 281-294.
Mitchell, W. A., Abramsky, Z., Kotler, B. P. et al. 1990. The effect of competition on foraging activity in desert rodents: theory and experiments. - Ecology 71: 844-854.
Price, M. V. and Brown, J. H. 1983. Patterns of morphology and resource use in North American desert rodent commu- nities. - Great Basin Nat. Mem. 7: 117-134.
Pulliam, H. R. 1975. Coexistence of sparrow: a test of commu- nity theory. - Science 189: 474-476.
Rice, W. R. 1989. Analysing tables of statistical tests. - Evolution 43: 223-225.
Rosenzweig, M. L. 1977. Coexistence and diversity in het- eromyid rodents. - In: Stonehouse, B. and Perrins, C. (eds), Evolutionary ecology. Macmillan, pp. 84-99.
Rosenzweig, M. L. and Abramsky, Z. 1985. Detecting density dependent habitat selection. - Am. Nat. 126: 405-417.
Rosenzweig, M. L. and Abramsky, Z. 1986. Centrifugal com- munity structure. - Oikos 46: 339-348.
SAS 1997. Jmp Version 3.2.2. - SAS Inst., Inc., Cary, NC. Schluter, D. 1984. A variance test for detecting species associ-
ations, with some example applications. - Ecology 65: 998-1005.
OIKOS 90:2 (2000) 406

Stephens, D. W. and Krebs, J. R. 1986. Foraging theory. - Princeton Univ. Press.
Thorson, J. M. 1995. Fear and food: a squirrel's perception of predatory risk and diet selection. - MSc thesis, Univ. of Illinois at Chicago.
Tilman, D. 1982. Resource competition and community struc- ture. - Princeton Univ. Press.
Valone, T. J. and Brown, J. S. 1989. Measuring patch assess- ment abilities of desert granivores. - Ecology 70: 1800- 1810.
Venable, D. L. 1989. Modelling the evolutionary ecology of seedbanks. - In: Leck, M. A., Parker, V. T. and Simpson, R. L. (eds), Ecology of soil seedbanks. Academic Press, pp. 67-90.
Werner, E. E. 1992. Individual behavior and higher-order species interactions. - Am. Nat. 140: S5-S32.
Ziv, Y., Abramsky, Z., Kotler, B. P. and Subach, A. 1993. Interference competition and temporal and habitat partitioning in two gerbil species. - Oikos 66: 237- 246.
OIKOS 90:2 (2000) 407
Copyright © 2022 FDOKUMEN




















