
The E3 ligase Topors induces the accumulation of polysumoylated forms of DNA topoisomerase I in...
7
The E3 ligase Topors induces the accumulation of polysumoylated forms of DNA topoisomerase I in vitro and in vivo Eva Hammer, Regine Heilbronn, Stefan Weger * Institute of Virology, Charite ´ Campus Benjamin Franklin, Free University of Berlin, Hindenburgdamm 27, 12203 Berlin, Germany Received 5 June 2007; revised 22 October 2007; accepted 22 October 2007 Available online 30 October 2007 Edited by Ivan Sadowski Abstract Human Topors has originally been identified as bind- ing partner of p53 and DNA topoisomerase I (TOP1). It can function as both an ubiquitin and SUMO-1 E3 ligase for p53. Here we demonstrate that Topors enhances the formation of high-molecular weight SUMO-1 conjugates of TOP1 in a recon- stituted in vitro system and also in human osteosarcoma cells, similar to treatment with CPT. In contrast to the situation ob- served with p53, overall sumoylation levels were rather unaf- fected. Experiments with TOP1 point mutants strongly suggest that the high-molecular weight conjugates represent SUMO-1 chains formed on a limited number of SUMO-1 acceptor sites. Ó 2007 Federation of European Biochemical Societies. Pub- lished by Elsevier B.V. All rights reserved. Keywords: Small ubiquitin-like modifier; Topors; DNA topoisomerase I; E3 ligase; CPT 1. Introduction Human SUMO-1 (the small ubiquitin-like modifier 1) is covalently attached as a post-translational modification to a variety of cellular target proteins. After activation by the hete- rodimeric E1 enzyme Aos-1/Uba2 and formation of a thioester with the E2 conjugating enzyme ubiquitin conjugating enzyme 9 (UBC9), SUMO-1 is attached to specific lysine residues with- in the target protein [1]. UBC9 physically interacts with many known SUMO-1 substrates and the E1 and E2 enzymes are suf- ficient for SUMO-1 modification of a variety of these substrates in reconstituted systems. However, several different types of SUMO-1 E3 ligases such as the protein inhibitors of activated STAT (PIAS) protein family, the nucleoporin RanBP2 or the Polycomb protein Pc2, which stimulate the SUMO-1 sumoyla- tion of specific substrates, have been identified [2]. Sumoylation has been implicated in various functions such as regulation of the stability, the intracellular localization and the transcrip- tional activity of the respective target protein [3]. Human DNA topoisomerase I (TOP1) is a major SUMO-1 target in cells treated with the TOP1-specific camptothecin (CPT) class of anticancer agents [4]. TOP1 catalyzes the relax- ation of torsional stress associated with DNA replication, recombination, transcription and chromatin condensation [5]. It transiently cleaves one strand of the double helix, creating a protein-linked nick, which allows rotation of the other strand, followed by religation of the phosphate backbone [5]. CPT and its derivates reversibly stabilize the covalent TOP1-DNA inter- mediate [6] accompanied by a rapid relocalization of TOP1 from the nucleolus to the nucleoplasm [7]. The hypersumoyla- tion of TOP1 observed after treatment with CPT and experi- ments with sumoylation-deficient TOP1 mutants suggested an involvement of TOP1 sumoylation in its partitioning between the nucleoli and the nucleoplasm [8,9]. However, this issue re- mains controversial in view of a more recent report concluding that sumoylation does not affect CPT-induced TOP1 relocaliza- tion [10]. TOP1 sumoylation sites have been mapped to lysine residues K103, K117 and K153 in the N-terminal part of the protein, with K117 as the major acceptor site [9]. The existence of polymeric SUMO-1 chains (polysumoylation) has long been debated. However, polysumoylation has been demonstrated in vitro for several substrates including a glutathione S-trans- ferase (GST) fusion protein harboring only the region sur- rounding the major TOP1 acceptor site at amino acid 117 [11]. Topors is a cellular RING finger protein, which binds to the N-terminus of DNA topoisomerase I [12] and also to p53 [13]. While normally located in nuclear speckles closely associated with the PML oncogenic domains (PODs) [14,15], it rapidly relocalizes to the nucleoplasm after CPT treatment [14]. It pos- sesses both RING finger dependent ubiquitin [16] and RING finger independent SUMO-1 E3 ligase activities for p53 [17]. The drosophila homologue dTopors interacts with and directs the nuclear organization of the gypsy chromatin insulator complex [18]. Whereas this process involves sumoylation of Mod(mdg4)2.2 and Cp190, dTopors does not seem to directly function as a SUMO-1 E3 ligase for these proteins [19]. In the present study we demonstrate a Topors induced accu- mulation of high-molecular weight SUMO-1 conjugates of TOP1 both in a reconstituted in vitro sumoylation system and in vivo. Point mutations of the TOP1 SUMO-1 acceptor sites show that these high-molecular weight forms are most likely due to polysumoylation at a very limited number of lysine residues. 2. Materials and methods 2.1. Plasmid construction Plasmid pBS-Top1 for in vitro transcription/translation of TOP1 was generated by subcloning of the corresponding BamHI/EcoRI frag- ment from plasmid pTI-2 [20] (kindly provided by William T. Beck) Abbreviations: SUMO, small ubiquitin-like modifier; UBC9, ubiquitin conjugating enzyme 9; GST, glutathione S-transferase; GFP, green fluorescent protein; YFP, yellow fluorescent protein; PIAS, protein inhibitors of activated STAT; HU, hydroxy-uracil * Corresponding author. Fax: +49 30 8445 4485. E-mail address: [email protected] (S. Weger). 0014-5793/$32.00 Ó 2007 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved. doi:10.1016/j.febslet.2007.10.040 FEBS Letters 581 (2007) 5418–5424
-
Upload
charite-de -
Category
Documents
-
view
2 -
download
0
Transcript of The E3 ligase Topors induces the accumulation of polysumoylated forms of DNA topoisomerase I in...

FEBS Letters 581 (2007) 5418–5424
The E3 ligase Topors induces the accumulation of polysumoylatedforms of DNA topoisomerase I in vitro and in vivo
Eva Hammer, Regine Heilbronn, Stefan Weger*
Institute of Virology, Charite Campus Benjamin Franklin, Free University of Berlin, Hindenburgdamm 27, 12203 Berlin, Germany
Received 5 June 2007; revised 22 October 2007; accepted 22 October 2007
Available online 30 October 2007
Edited by Ivan Sadowski
Abstract Human Topors has originally been identified as bind-ing partner of p53 and DNA topoisomerase I (TOP1). It canfunction as both an ubiquitin and SUMO-1 E3 ligase for p53.Here we demonstrate that Topors enhances the formation ofhigh-molecular weight SUMO-1 conjugates of TOP1 in a recon-stituted in vitro system and also in human osteosarcoma cells,similar to treatment with CPT. In contrast to the situation ob-served with p53, overall sumoylation levels were rather unaf-fected. Experiments with TOP1 point mutants strongly suggestthat the high-molecular weight conjugates represent SUMO-1chains formed on a limited number of SUMO-1 acceptor sites.� 2007 Federation of European Biochemical Societies. Pub-lished by Elsevier B.V. All rights reserved.
Keywords: Small ubiquitin-like modifier; Topors;DNA topoisomerase I; E3 ligase; CPT
1. Introduction
Human SUMO-1 (the small ubiquitin-like modifier 1) is
covalently attached as a post-translational modification to a
variety of cellular target proteins. After activation by the hete-
rodimeric E1 enzyme Aos-1/Uba2 and formation of a thioester
with the E2 conjugating enzyme ubiquitin conjugating enzyme
9 (UBC9), SUMO-1 is attached to specific lysine residues with-
in the target protein [1]. UBC9 physically interacts with many
known SUMO-1 substrates and the E1 and E2 enzymes are suf-
ficient for SUMO-1 modification of a variety of these substrates
in reconstituted systems. However, several different types of
SUMO-1 E3 ligases such as the protein inhibitors of activated
STAT (PIAS) protein family, the nucleoporin RanBP2 or the
Polycomb protein Pc2, which stimulate the SUMO-1 sumoyla-
tion of specific substrates, have been identified [2]. Sumoylation
has been implicated in various functions such as regulation of
the stability, the intracellular localization and the transcrip-
tional activity of the respective target protein [3].
Human DNA topoisomerase I (TOP1) is a major SUMO-1
target in cells treated with the TOP1-specific camptothecin
(CPT) class of anticancer agents [4]. TOP1 catalyzes the relax-
Abbreviations: SUMO, small ubiquitin-like modifier; UBC9, ubiquitinconjugating enzyme 9; GST, glutathione S-transferase; GFP, greenfluorescent protein; YFP, yellow fluorescent protein; PIAS, proteininhibitors of activated STAT; HU, hydroxy-uracil
*Corresponding author. Fax: +49 30 8445 4485.E-mail address: [email protected] (S. Weger).
0014-5793/$32.00 � 2007 Federation of European Biochemical Societies. Pu
doi:10.1016/j.febslet.2007.10.040
ation of torsional stress associated with DNA replication,
recombination, transcription and chromatin condensation [5].
It transiently cleaves one strand of the double helix, creating
a protein-linked nick, which allows rotation of the other strand,
followed by religation of the phosphate backbone [5]. CPT and
its derivates reversibly stabilize the covalent TOP1-DNA inter-
mediate [6] accompanied by a rapid relocalization of TOP1
from the nucleolus to the nucleoplasm [7]. The hypersumoyla-
tion of TOP1 observed after treatment with CPT and experi-
ments with sumoylation-deficient TOP1 mutants suggested an
involvement of TOP1 sumoylation in its partitioning between
the nucleoli and the nucleoplasm [8,9]. However, this issue re-
mains controversial in view of a more recent report concluding
that sumoylation does not affect CPT-induced TOP1 relocaliza-
tion [10]. TOP1 sumoylation sites have been mapped to lysine
residues K103, K117 and K153 in the N-terminal part of the
protein, with K117 as the major acceptor site [9]. The existence
of polymeric SUMO-1 chains (polysumoylation) has long been
debated. However, polysumoylation has been demonstrated
in vitro for several substrates including a glutathione S-trans-
ferase (GST) fusion protein harboring only the region sur-
rounding the major TOP1 acceptor site at amino acid 117 [11].
Topors is a cellular RING finger protein, which binds to the
N-terminus of DNA topoisomerase I [12] and also to p53 [13].
While normally located in nuclear speckles closely associated
with the PML oncogenic domains (PODs) [14,15], it rapidly
relocalizes to the nucleoplasm after CPT treatment [14]. It pos-
sesses both RING finger dependent ubiquitin [16] and RING
finger independent SUMO-1 E3 ligase activities for p53 [17].
The drosophila homologue dTopors interacts with and directs
the nuclear organization of the gypsy chromatin insulator
complex [18]. Whereas this process involves sumoylation of
Mod(mdg4)2.2 and Cp190, dTopors does not seem to directly
function as a SUMO-1 E3 ligase for these proteins [19].
In the present study we demonstrate a Topors induced accu-
mulation of high-molecular weight SUMO-1 conjugates of
TOP1 both in a reconstituted in vitro sumoylation system
and in vivo. Point mutations of the TOP1 SUMO-1 acceptor
sites show that these high-molecular weight forms are most
likely due to polysumoylation at a very limited number of
lysine residues.
2. Materials and methods
2.1. Plasmid constructionPlasmid pBS-Top1 for in vitro transcription/translation of TOP1
was generated by subcloning of the corresponding BamHI/EcoRI frag-ment from plasmid pTI-2 [20] (kindly provided by William T. Beck)
blished by Elsevier B.V. All rights reserved.
Topors Protein
Assay-Mix
1 2 3 4 5 6 7
TOP1-n x SUMO-1
100
150
250
kDa
wildtype
8
ΔRING
TOP1
9
500
ng
UB
C9
Topors Protein
Assay-Mix
1 2 3 4 5 6
TOP1-n x SUMO-1
105
160
250
kDa
268/644
TOP1
500
ng
UB
C9
A
B
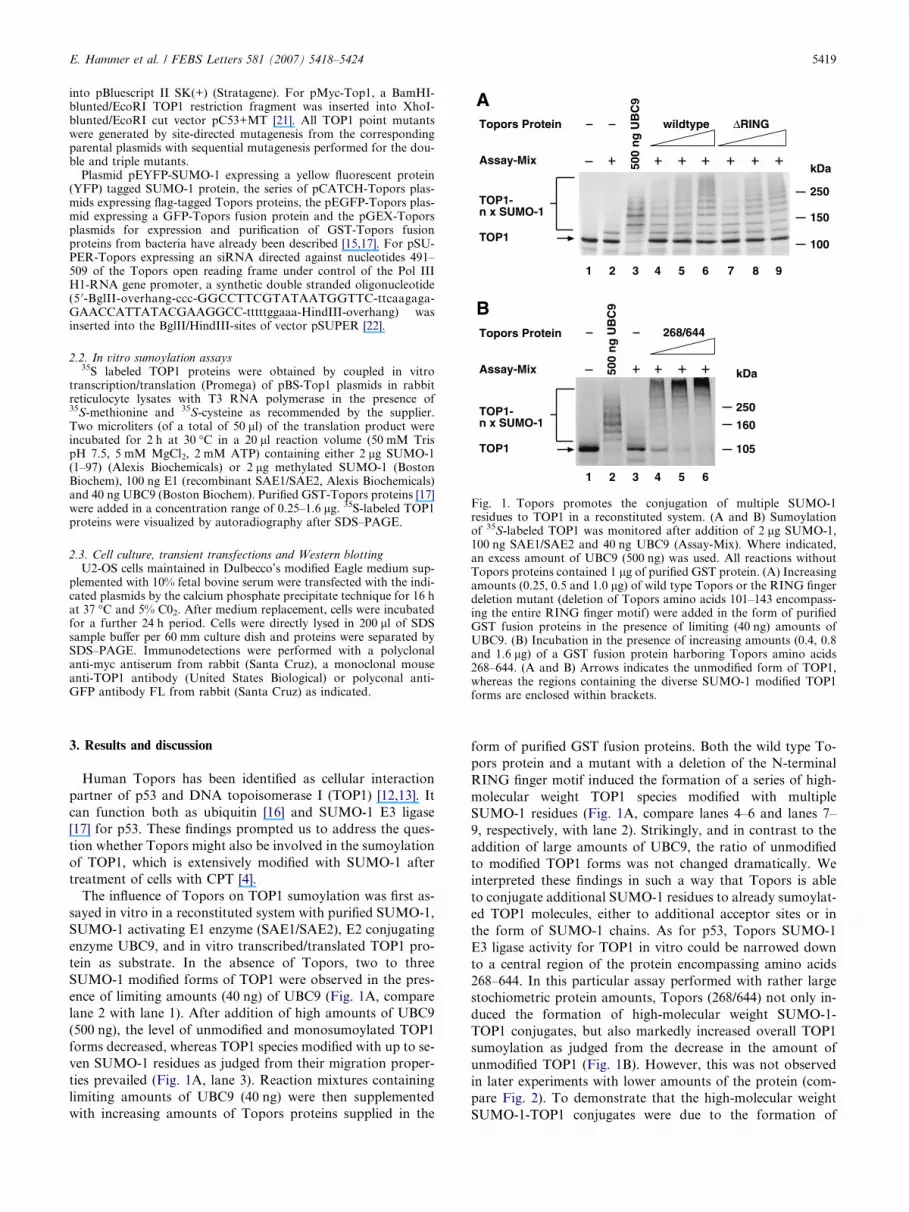
Fig. 1. Topors promotes the conjugation of multiple SUMO-1residues to TOP1 in a reconstituted system. (A and B) Sumoylationof 35S-labeled TOP1 was monitored after addition of 2 lg SUMO-1,100 ng SAE1/SAE2 and 40 ng UBC9 (Assay-Mix). Where indicated,an excess amount of UBC9 (500 ng) was used. All reactions withoutTopors proteins contained 1 lg of purified GST protein. (A) Increasingamounts (0.25, 0.5 and 1.0 lg) of wild type Topors or the RING fingerdeletion mutant (deletion of Topors amino acids 101–143 encompass-ing the entire RING finger motif) were added in the form of purifiedGST fusion proteins in the presence of limiting (40 ng) amounts ofUBC9. (B) Incubation in the presence of increasing amounts (0.4, 0.8and 1.6 lg) of a GST fusion protein harboring Topors amino acids268–644. (A and B) Arrows indicates the unmodified form of TOP1,whereas the regions containing the diverse SUMO-1 modified TOP1forms are enclosed within brackets.
E. Hammer et al. / FEBS Letters 581 (2007) 5418–5424 5419
into pBluescript II SK(+) (Stratagene). For pMyc-Top1, a BamHI-blunted/EcoRI TOP1 restriction fragment was inserted into XhoI-blunted/EcoRI cut vector pC53+MT [21]. All TOP1 point mutantswere generated by site-directed mutagenesis from the correspondingparental plasmids with sequential mutagenesis performed for the dou-ble and triple mutants.
Plasmid pEYFP-SUMO-1 expressing a yellow fluorescent protein(YFP) tagged SUMO-1 protein, the series of pCATCH-Topors plas-mids expressing flag-tagged Topors proteins, the pEGFP-Topors plas-mid expressing a GFP-Topors fusion protein and the pGEX-Toporsplasmids for expression and purification of GST-Topors fusionproteins from bacteria have already been described [15,17]. For pSU-PER-Topors expressing an siRNA directed against nucleotides 491–509 of the Topors open reading frame under control of the Pol IIIH1-RNA gene promoter, a synthetic double stranded oligonucleotide(5 0-BglII-overhang-ccc-GGCCTTCGTATAATGGTTC-ttcaagaga-GAACCATTATACGAAGGCC-tttttggaaa-HindIII-overhang) wasinserted into the BglII/HindIII-sites of vector pSUPER [22].
2.2. In vitro sumoylation assays35S labeled TOP1 proteins were obtained by coupled in vitro
transcription/translation (Promega) of pBS-Top1 plasmids in rabbitreticulocyte lysates with T3 RNA polymerase in the presence of35S-methionine and 35S-cysteine as recommended by the supplier.Two microliters (of a total of 50 ll) of the translation product wereincubated for 2 h at 30 �C in a 20 ll reaction volume (50 mM TrispH 7.5, 5 mM MgCl2, 2 mM ATP) containing either 2 lg SUMO-1(1–97) (Alexis Biochemicals) or 2 lg methylated SUMO-1 (BostonBiochem), 100 ng E1 (recombinant SAE1/SAE2, Alexis Biochemicals)and 40 ng UBC9 (Boston Biochem). Purified GST-Topors proteins [17]were added in a concentration range of 0.25–1.6 lg. 35S-labeled TOP1proteins were visualized by autoradiography after SDS–PAGE.
2.3. Cell culture, transient transfections and Western blottingU2-OS cells maintained in Dulbecco’s modified Eagle medium sup-
plemented with 10% fetal bovine serum were transfected with the indi-cated plasmids by the calcium phosphate precipitate technique for 16 hat 37 �C and 5% C02. After medium replacement, cells were incubatedfor a further 24 h period. Cells were directly lysed in 200 ll of SDSsample buffer per 60 mm culture dish and proteins were separated bySDS–PAGE. Immunodetections were performed with a polyclonalanti-myc antiserum from rabbit (Santa Cruz), a monoclonal mouseanti-TOP1 antibody (United States Biological) or polyconal anti-GFP antibody FL from rabbit (Santa Cruz) as indicated.
3. Results and discussion
Human Topors has been identified as cellular interaction
partner of p53 and DNA topoisomerase I (TOP1) [12,13]. It
can function both as ubiquitin [16] and SUMO-1 E3 ligase
[17] for p53. These findings prompted us to address the ques-
tion whether Topors might also be involved in the sumoylation
of TOP1, which is extensively modified with SUMO-1 after
treatment of cells with CPT [4].
The influence of Topors on TOP1 sumoylation was first as-
sayed in vitro in a reconstituted system with purified SUMO-1,
SUMO-1 activating E1 enzyme (SAE1/SAE2), E2 conjugating
enzyme UBC9, and in vitro transcribed/translated TOP1 pro-
tein as substrate. In the absence of Topors, two to three
SUMO-1 modified forms of TOP1 were observed in the pres-
ence of limiting amounts (40 ng) of UBC9 (Fig. 1A, compare
lane 2 with lane 1). After addition of high amounts of UBC9
(500 ng), the level of unmodified and monosumoylated TOP1
forms decreased, whereas TOP1 species modified with up to se-
ven SUMO-1 residues as judged from their migration proper-
ties prevailed (Fig. 1A, lane 3). Reaction mixtures containing
limiting amounts of UBC9 (40 ng) were then supplemented
with increasing amounts of Topors proteins supplied in the
form of purified GST fusion proteins. Both the wild type To-
pors protein and a mutant with a deletion of the N-terminal
RING finger motif induced the formation of a series of high-
molecular weight TOP1 species modified with multiple
SUMO-1 residues (Fig. 1A, compare lanes 4–6 and lanes 7–
9, respectively, with lane 2). Strikingly, and in contrast to the
addition of large amounts of UBC9, the ratio of unmodified
to modified TOP1 forms was not changed dramatically. We
interpreted these findings in such a way that Topors is able
to conjugate additional SUMO-1 residues to already sumoylat-
ed TOP1 molecules, either to additional acceptor sites or in
the form of SUMO-1 chains. As for p53, Topors SUMO-1
E3 ligase activity for TOP1 in vitro could be narrowed down
to a central region of the protein encompassing amino acids
268–644. In this particular assay performed with rather large
stochiometric protein amounts, Topors (268/644) not only in-
duced the formation of high-molecular weight SUMO-1-
TOP1 conjugates, but also markedly increased overall TOP1
sumoylation as judged from the decrease in the amount of
unmodified TOP1 (Fig. 1B). However, this was not observed
in later experiments with lower amounts of the protein (com-
pare Fig. 2). To demonstrate that the high-molecular weight
SUMO-1-TOP1 conjugates were due to the formation of

B
C D
Topors proteinAssay-Mix
1 2 3 4 5 6 7
TOP1-n x SUMO-1
105
160
250
kDa
wildty
pe
8
TOP1
TOP1-wt TOP1-K117R
268/
644
wildty
pe
TOP1 protein
268/
644
**
Assay-Mix
1 2 3 4 5 6
TOP1-n x SUMO-1
105
160
250
kDa
7
TOP1
SUMO-1 Me-SUMO-1
Topors
268
/644
SUMO protein
500
ng UBC9
500
ng UBC9
Topors
268
/644
Assay-Mix
1 2 3 4 5 6
TOP1-n x SUMO-1
105
160
250
kDa
7
TOP1
8
wildtype triple mutant
Topors
268
/644
TOP1 protein
500
ng UBC9
500
ng UBC9
Topors
268
/644
ATopors proteinAssay-Mix
1 2 3 4 5 6 7
TOP1-n x SUMO-1
105
160
250
kDa
wildty
pe
8
ΔRING
TOP1
9 10
TOP1-wt TOP1-K103R/K153R
268/
644
wildty
pe
ΔRING
268/
644
TOP1 protein
Fig. 2. Topors induced polysumoylation of TOP1 point mutants in vitro. (A–D) 35S-labeled wild type or mutant TOP1 proteins were assayed forsumoylation in the presence of either 2 lg SUMO-1 (A–C) or 2 lg methylated SUMO-1 (right panel in part D), 100 ng SAE1/SAE2 and 40 ng UBC9(Assay-Mix). Purified GST fusion proteins of wild type Topors (1.0 lg), the RING finger deletion mutant (1.0 lg) or the Topors 268/644 deletionmutant (0.5 lg) were added to the reaction mixture as indicated. The control reactions contained 1.0 lg of purified GST protein. Arrows indicate theunmodified forms of TOP1, whereas the regions containing the SUMO-1 modified TOP1 forms are enclosed within brackets. (C and D) Whereindicated, reactions were performed in the presence of an excess amount of UBC9 (500 ng) or GST-Topors (268/644) (1.6 lg).
5420 E. Hammer et al. / FEBS Letters 581 (2007) 5418–5424
SUMO-1 chains, as has recently been suggested by Yang et al.
[11], we point-mutated the three described SUMO-1 acceptor
sites of TOP1 located in the N-terminal part of the protein
at amino acid positions 103, 117 and 153, respectively. Inacti-
vation of the major SUMO-1 acceptor site at lysine residue 117
(Fig. 2A, K117R) resulted in reduced basal sumoylation with
two putative monosumoylated TOP1 forms migrating with
slightly different mobilities (marked by asterisks in Fig. 2A),
probably due to modification at the two different remaining
acceptor sites. Addition of Topors proteins again induced
the accumulation of high-molecular weight conjugates,
although with lower intensity as compared to the wild type
protein (Fig. 2B), corresponding to the lower basal level of
sumoylation. With a TOP1 mutant retaining only the major
sumoylation site (TOP1-K103R/K153R), Topors induced the
accumulation of TOP1 species modified with multiple
SUMO-1 residues to a similar extent as with the wild type
TOP1 protein (Fig. 2B). Thus these conjugates most likely rep-
resent SUMO-1 chains linked to lysine residue 117. An alterna-
tive explanation, the conjugation of single SUMO-1 moieties
to additional, yet unidentified, acceptor sites in TOP1, seems
very unlikely in view of the results obtained after additional
point mutation of lysine residue 117. With this triple mutant,
only a weak residual sumoylation, detectable exclusively in
the presence of high amounts (500 ng) of UBC or an excess
amount (1.6 lg) of GST-Topors (268/644) (Fig. 2C, lanes 7
and 8), was observed. To further manifest the Topors-medi-
ated formation of SUMO-1 chains on TOP1, sumoylation as-
says with the TOP1 K103/153 mutant were performed in the
presence of methylated SUMO-1 (Fig. 2D). Reductive methyl-
ation of the lysine residues renders SUMO-1 unable to form
chains via lysine linkage with other SUMO-1 molecules.
Whereas enhancement of TOP1 sumoylation by an excess
amount of UBC9 was rather unaffected by SUMO-1 methyla-
tion (Fig. 2D, compare lanes 6 and 3), the induction of high-
molecular weight TOP1-SUMO-1 conjugates was abolished
with methylated SUMO-1 (compare lanes 7 and 4). In sum-
mary, the results of the in vitro experiments provided firm evi-
dence that Topors can induce the formation of SUMO-1
chains on all three TOP1-SUMO-1 acceptor sites.
The influence of Topors on TOP1 sumoylation in mamma-
lian cells was first monitored by cotransfection of the human
osteosarcoma cell line U2-OS with myc-tagged TOP protein,
YFP-tagged SUMO-1 and increasing amounts of Topors
expression constructs. Whereas sumoylation of myc-tagged
TOP1 could not unequivocally be detected in the absence of

E. Hammer et al. / FEBS Letters 581 (2007) 5418–5424 5421
SUMO-1 overexpression, YFP or green fluorescent protein
(GFP) tagged SUMO-1 proteins have already been used in a
variety of previous studies examining sumolation of cellular
proteins [17,23] and GFP-SUMO can functionally replace
endogenous SUMO (pmt3) in fission yeast [24]. In the absence
of ectopic Topors expression, one main additional band corre-
sponding in size to TOP1 modified with a single YFP-SUMO-
1 entity was observed after YFP-SUMO-1 cotransfection
(Fig. 3A, compare lanes 2 and 3). Several weak high-molecular
weight myc-reactive bands corresponding to TOP1 species
modified with several YFP-SUMO-1 molecules were also visi-
ble. After cotransfection of increasing amounts of the full-
length wild type Topors protein the intensity of these polysu-
moylated TOP1 bands increased, whereas the amount of the
monosumoylated TOP1 form rather decreased (Fig. 3A, lanes
A
B
no
ne
160
250
105
kDa
wildtyp
1 2 3 4 5
Toporsprotein n
on
e
YFP-SUMO-1
Myc-Top1
no
ne
160
250
105
kDa
1 2 3 4 5
YFP-SUMO-1
Myc-TOP1
2510 1
5 μμM CPTtime (min)
10tim
Fig. 3. A central domain of the Topors protein promotes the conjugation of mand B) U2-OS cells were cotransfected with 2 lg of plasmid pMyc-TOP1 expexpression constructs for either the YFP protein or an YFP-SUMO-1 fusionamounts of pCATCH-Topors constructs (0.25, 0.5 and 1.0 lg, respectively) enadded. Total DNA content was adjusted to 4 lg with pCATCH vector DNA.extracts from equal number of cells were analyzed by immunoblotting wiunmodified and mono-YFP-SUMO-1 modified forms of TOP1, while hiExpression of Topors proteins was controlled by anti-Flag immunoblotting (nTOP1 and pEYFP-SUMO-1 were treated with the indicated concentrationsDMSO/PBS for 25 min for the controls.
4–6). The in vitro requirement of a central Topors domain for
its SUMO-1 E3 ligase activity towards TOP1 could be con-
firmed in U2-OS cells with a Topors polypeptide harboring
amino acids 437–574, which induced a comparable polysumoy-
lation of TOP1 as the wild type Topors protein (Fig. 3A, lanes
7–9). With the myc-tagged TOP1 protein, the forced expres-
sion of Topors had a very similar effect as treating the cells
with CPT, which also mainly increased the abundance of the
high-order YFP-SUMO-1 conjugates of TOP1 (Fig. 3B).
In analogy to the in vitro assays, the transfection experiments
were repeated with TOP1 lysine point mutants to show that the
high-molecular weight YFP-SUMO-1 modified forms of TOP1
consist of SUMO-1 chains. Both with the TOP1 mutant retaining
only the major sumoylation site at lysine residue 117 (Fig. 4A,
TOP1-K103R/K153R) and the mutant harboring the minor sites
e
TOP1
TOP1-SUMO-1
6
TOP1-n x SUMO-1
7 8 9
437/574
TOP1
TOP1-SUMO-1
6
TOP1-n x SUMO-1
7
250
μμM CPTe (min)
ultiple YFP-SUMO-1 residues to exogenous TOP1 in U2-OS cells. (Aressing a full-length N-terminal myc-tagged TOP1 protein and 1 lg ofprotein. (A) To study the effect of forced Topors expression, increasingcoding either full-length Topors or Topors amino acids 437 to 574 wereUnmodified and YFP-SUMO-1 modified forms of TOP1 in whole cell
th a polyclonal anti-myc antiserum. Arrows indicate the position ofgher-order YFP-SUMO-1 conjugates are enclosed within brackets.ot shown). (B) Directly before cell lysis, cells cotransfected with pMyc-
of CPT (in 1% DMSO/PBS) for 10 or 25 min, respectively, or with 1%

A
B
no
ne
160
250
105
kDa
437/574
1 2 3 4
Toporsprotein
TOP1-(K103R-K153R)
TOP1-SUMO-1
YFP-SUMO-1
5
no
ne
TOP1-n x SUMO-1
C
no
ne
160
250
105
kDa
1 2 3 4
TOP1protein
TOP1-
TOP1-SUMO-1
YFP-SUMO-1
5
TOP1-n x SUMO-1
6 7 8 9
wildtype K117R K103R/K153R
triplemutant
no
ne
160
250
kDa
437/574Toporsprotein
TOP1-(K117R)
TOP1-SUMO-1
YFP-SUMO-1
no
ne
TOP1-n x SUMO-1
1051 2 3 4 5
Fig. 4. Topors induced formation of high-order SUMO-1 conjugates on TOP1 mutants in vivo. (A–C) U2-OS cells were cotransfected with 2 lg ofexpression constructs for the indicated myc-tagged TOP1 point mutants and 1 lg of expression constructs for either the YFP protein or an YFP-SUMO-1 fusion protein. In (A and B), increasing amounts of pCATCH-Topors (437–574) plasmid (0.25, 0.5 and 1.0 lg, respectively) encoding thecentral Topors domain were cotransfected. Total DNA content was adjusted to 4 lg with pCATCH vector DNA in all transfections. Unmodified andYFP-SUMO-1 modified forms of TOP1 in whole cell extracts from equal number of cells were analyzed by immunoblotting with a polyclonal anti-myc antiserum. Arrows indicate the position of unmodified and mono-YFP-SUMO-1 modified forms of TOP1, while higher-order YFP-SUMO-1conjugates are enclosed within brackets.
5422 E. Hammer et al. / FEBS Letters 581 (2007) 5418–5424
at lysine residues 103 and 153 (Fig. 4B, TOP1-K117R), the cen-
tral Topors domain induced the accumulation of high-order
YFP-SUMO-1 conjugates. YFP-SUMO-1 modification in cells
closely mimicked the in vitro experiments also with regard to
the pattern of monosumoylated TOP1 species, with the K117R
mutant showing two bands with slightly different mobility’s. In
line with published results [9], the triple mutant K103R/
K117R/K153R was sumoylation-deficient (Fig. 4C).
We then examined the influence of Topors on sumoylation
of endogenous TOP1 in HeLa cells by cotransfection of To-
pors (437/574) with YFP-SUMO-1, followed by immunoblot
analysis with a TOP1 antibody. Several putative YFP-SUMO

E. Hammer et al. / FEBS Letters 581 (2007) 5418–5424 5423
modified forms of endogenous TOP1 were identified (Fig. 5A,
indicated by arrows). Some bands with a higher mobility than
the non-modified TOP1 (marked by asterisks in Fig. 5) were
present in all transfection and some faint signals in the range
of the putative YFP-SUMO-1 modified forms of TOP1 were
also present in the control without YFP-SUMO-1. However,
the mobilities of the later differed slightly from the YFP-
no
ne
160
250
105
kDa
43
1 2 3
Toporsprotein
YFP-SUMO-1
no
ne
160
250
105
kDa
1 2 3 4 5
ToporssiRNA
TOP1
TOP1-SUMO-1
YFP-SUMO-1
TOP1-n x SUMO-1
TOP1-2 x SUMO-1
no
ne
+ 10 μμM CPTB
A
**
Fig. 5. Topors induces formation of high-order SUMO-1 conjugates on endYFP-SUMO-1 (or YFP only for the control) and increasing amounts of erespectively) or (B) Topors-si-RNA expression construct (1.0 and 2.0 lg, repCATCH vector DNA or (B) pSUPER vector DNA in all transfections. Unwhole cell extracts from equal number of cells were analyzed by immunobposition of unmodified, mono- and di-YFP-SUMO-1 modified forms of TObrackets. Cellular protein cross-reacting with the antibody are indicated by asincreasing amounts of Topors-si-RNA expression construct (1.0 and 2.0 lg, rcell extracts from equal number of cells was analyzed by immunoblotting w
SUMO-1 specific bands. Thus these bands most probably pres-
ent cellular protein cross-reacting with the antibody. After
cotransfection of increasing amounts of Topos (437/574), the
intensities of the high-molecular weight forms of YFP-
SUMO-1 modified TOP1 were enhanced and additional spe-
cies with an increased size could be detected (Fig. 5A, lanes
3–5). In a complementary approach, endogenous Topors
7/574
4 5
TOP1
TOP1-SUMO-1
TOP1-n x SUMO-1
TOP1-2 x SUMO-1
no
ne
160
250
105
kDa
1 2 3 4
ToporssiRNA
GFP-Topors
GFP-Topors
75
no
ne
C
**
ogenous TOP1. (A and B) HeLa cells were cotransfected with 2 lg ofither (A) pCATCH-Topors (437–574) plasmid (0.25, 0.5 and 1.0 lg,spectively). Total DNA content was adjusted to 4 lg with either (A)modified and YFP-SUMO-1 modified forms of endogenous TOP1 inlotting with a monoclonal anti-TOP1 antibody. Arrows indicate theP1, while higher-order YFP-SUMO-1 conjugates are enclosed within
terisks. (C) HeLa cells were cotransfected with 2 lg of GFP-Topors andespectively). GFP-Topors expression (indicated by an arrow) in whole
ith a polyclonal anti-GFP antibody.

5424 E. Hammer et al. / FEBS Letters 581 (2007) 5418–5424
expression was knocked out in HeLa cells transfected with
YFP-SUMO-1 and treated with CPT by means of a cotrans-
fected Topors-siRNA expression construct. The selected To-
pors-siRNA efficiently downregulated Topors expression as
monitored in parallel by cotransfection with a GFP-tagged To-
pors construct (Fig. 5C). Whereas the high-order YFP-
SUMO-1 modified forms of endogenous TOP1 were enhanced
after treatment with CPT (Fig. 5B, compare lanes 2 and 3), this
effect was efficiently repressed by delivery of the Topors-siR-
NA. In contrast, knockdown of Topors seems to have only lit-
tle effect on mono- and di-sumoylation of Top1. Therefore it
seems that Topors is not involved in the initial sumoylation
of Top1 after CPT treatment, which probably involves a
SUMO ligase other than Topors, but that the role of Topors
is to facilitate chain formation on already sumoylated lysines.
In conclusion, Topors induced accumulation of high-order
SUMO-1 conjugates of TOP1 did not depend on the presence
of multiple lysine acceptor within the TOP1 sequence, neither
in the reconstituted nor in the cellular system, strongly arguing
for the formation of SUMO-1 chains. The apparently larger
number of SUMO-1 molecules conjugated to individual
TOP1 molecules in the reconstituted in vitro system as com-
pared to the cellular system may be due to the presence of
SUMO-1 specific isopeptidases in the later system. For the
yeast Smt3/SUMO isopeptidase Ulp2p, a function in the edit-
ing of the SUMO chains has been demonstrated and accumu-
lation of SUMO chains in Ulp2p negative strains induces cell
death and correlates with increased sensitivity to hydroxy-ura-
cil (HU) [25]. These effects are enhanced in the presence of
TOP1 [26], implying TOP1 as a major target for SUMO chain
formation in yeast. TOP1 sumoylation enhances CPT induced
apoptosis in human cells [27] and Topors has also been impli-
cated in cell cycle arrest and induction of apoptosis in mamma-
lian cells [28,29]. Of note, Topors is both induced [28] and
relocated from PML nuclear bodies to the nucleoplasm [14]
after CPT treatment. Further studies will help to clarify the
role of Topors induced formation of SUMO-1 chains on
TOP1 in CPT induced apoptosis.
References
[1] Melchior, F. (2000) SUMO–nonclassical ubiquitin. Ann. Rev.Cell Dev. Biol. 16, 591–626.
[2] Melchior, F., Schergaut, M. and Pichler, A. (2003) SUMO:ligases, isopeptidases and nuclear pores. Trends Biochem. Sci. 28,612–618.
[3] Johnson, E.S. (2004) Protein modification by sumo. Ann. Rev.Biochem. 73, 355–382.
[4] Mao, Y., Sun, M., Desai, S.D. and Liu, L.F. (2000) SUMO-1conjugation to topoisomerase I: a possible repair response totopoisomerase-mediated DNA damage. Proc. Natl. Acad. Sci.USA 97, 4046–4051.
[5] Champoux, J.J. (2001) DNA topoisomerases: structure, function,and mechanism. Ann. Rev. Biochem. 70, 369–413.
[6] Hertzberg, R.P., Caranfa, M.J. and Hecht, S.M. (1989) On themechanism of topoisomerase I inhibition by camptothecin:evidence for binding to an enzyme-DNA complex. Biochemistry28, 4629–4638.
[7] Danks, M.K., Garrett, K.E., Marion, R.C. and Whipple, D.O.(1996) Subcellular redistribution of DNA topoisomerase I inanaplastic astrocytoma cells treated with topotecan. Cancer Res.56, 1664–1673.
[8] Mo, Y.Y., Yu, Y., Shen, Z. and Beck, W.T. (2002) Nucleolardelocalization of human topoisomerase I in response to topotecancorrelates with sumoylation of the protein. J. Biol. Chem. 277,2958–2964.
[9] Rallabhandi, P., Hashimoto, K., Mo, Y.Y., Beck, W.T., Moitra,P.K. and D’Arpa, P. (2002) Sumoylation of topoisomerase I isinvolved in its partitioning between nucleoli and nucleoplasm andits clearing from nucleoli in response to camptothecin. J. Biol.Chem. 277, 40020–40026.
[10] Christensen, M.O., Krokowski, R.M., Barthelmes, H.U., Hock,R., Boege, F. and Mielke, C. (2004) Distinct effects of topoiso-merase I and RNA polymerase I inhibitors suggest a dualmechanism of nucleolar/nucleoplasmic partitioning of topoiso-merase I. J. Biol. Chem. 279, 21873–21882.
[11] Yang, M., Hsu, C.T., Ting, C.Y., Liu, L.F. and Hwang, J. (2006)Assembly of a polymeric chain of SUMO1 on human topoiso-merase I in vitro. J. Biol. Chem. 281, 8264–8274.
[12] Haluska Jr., P., Saleem, A., Rasheed, Z., Ahmed, F., Su, E.W.,Liu, L.F. and Rubin, E.H. (1999) Interaction between humantopoisomerase I and a novel RING finger/arginine–serine protein.Nucl. Acids Res. 27, 2538–2544.
[13] Zhou, R., Wen, H. and Ao, S.Z. (1999) Identification of a novelgene encoding a p53-associated protein. Gene 235, 93–101.
[14] Rasheed, Z.A., Saleem, A., Ravee, Y., Pandolfi, P.P. and Rubin,E.H. (2002) The topoisomerase I-binding RING protein, topors,is associated with promyelocytic leukemia nuclear bodies. Exp.Cell Res. 277, 152–160.
[15] Weger, S., Hammer, E. and Engstler, M. (2003) The DNAtopoisomerase I binding protein topors as a novel cellular targetfor SUMO-1 modification: characterization of domains necessaryfor subcellular localization and sumoylation. Exp. Cell Res. 290,13–27.
[16] Rajendra, R. et al. (2004) Topors functions as an E3 ubiquitinligase with specific E2 enzymes and ubiquitinates p53. J. Biol.Chem. 279, 36440–36444.
[17] Weger, S., Hammer, E. and Heilbronn, R. (2005) Topors acts as aSUMO-1 E3 ligase for p53 in vitro and in vivo. FEBS Lett. 579,5007–5012.
[18] Capelson, M. and Corces, V.G. (2005) The ubiquitin ligasedTopors directs the nuclear organization of a chromatin insula-tor. Mol. Cell 20, 105–116.
[19] Capelson, M. and Corces, V.G. (2006) SUMO conjugationattenuates the activity of the gypsy chromatin insulator. EmboJ. 25, 1906–1914.
[20] Mo, Y.Y., Wang, P. and Beck, W.T. (2000) Functional expressionof human DNA topoisomerase I and its subcellular localization inHeLa cells. Exp. Cell Res. 256, 480–490.
[21] Turner, D.L. and Weintraub, H. (1994) Expression of achaete-scute homolog 3 in Xenopus embryos converts ectodermal cells toa neural fate. Genes Dev. 8, 1434–1447.
[22] Brummelkamp, T.R., Bernards, R. and Agami, R. (2002) Asystem for stable expression of short interfering RNAs inmammalian cells. Science 296, 550–553.
[23] Pichler, A., Gast, A., Seeler, J.S., Dejean, A. and Melchior, F.(2002) The nucleoporin RanBP2 has SUMO1 E3 ligase activity.Cell 108, 109–120.
[24] Tanaka, K. et al. (1999) Characterization of a fission yeastSUMO-1 homologue, pmt3p, required for multiple nuclearevents, including the control of telomere length and chromosomesegregation. Mol. Cell Biol. 19, 8660–8672.
[25] Bylebyl, G.R., Belichenko, I. and Johnson, E.S. (2003) TheSUMO isopeptidase Ulp2 prevents accumulation of SUMOchains in yeast. J. Biol. Chem. 278, 44113–44120.
[26] Jacquiau, H.R., van Waardenburg, R.C., Reid, R.J., Woo, M.H.,Guo, H., Johnson, E.S. and Bjornsti, M.A. (2005) Defects inSUMO (small ubiquitin-related modifier) conjugation and decon-jugation alter cell sensitivity to DNA topoisomerase I-inducedDNA damage. J. Biol. Chem. 280, 23566–23575.
[27] Horie, K., Tomida, A., Sugimoto, Y., Yasugi, T., Yoshikawa, H.,Taketani, Y. and Tsuruo, T. (2002) SUMO-1 conjugation tointact DNA topoisomerase I amplifies cleavable complex forma-tion induced by camptothecin. Oncogene 21, 7913–7922.
[28] Lin, L. et al. (2005) Topors, a p53 and topoisomerase I-bindingRING finger protein, is a coactivator of p53 in growth suppres-sion induced by DNA damage. Oncogene 24, 3385–3396.
[29] Saleem, A. et al. (2004) The topoisomerase I- and p53-bindingprotein topors is differentially expressed in normal and malignanthuman tissues and may function as a tumor suppressor. Oncogene23, 5293–5300.




















