
The Dalbergioid Legumes (Fabaceae): Delimitation of a Pantropical Monophyletic Clade
31
503 American Journal of Botany 88(3): 503–533. 2001. THE DALBERGIOID LEGUMES (FABACEAE): DELIMITATION OF A PANTROPICAL MONOPHYLETIC CLADE 1 MATT LAVIN, 2,3 R. TOBY PENNINGTON, 4 BENTE B. KLITGAARD, 5 JANET I. SPRENT, 6 HAROLDO CAVALCANTE DE LIMA, 7 AND PETER E. GASSON 5 3 Department of Plant Sciences, Montana State University, Bozeman, Montana 59717 USA; 4 Tropical Biology Group, Royal Botanic Garden Edinburgh, 20a Inverleith Row, Edinburgh EH3 5LR, UK; 5 Royal Botanic Gardens, Kew, Richmond, Surrey TW9 3AB, UK; 6 Department of Biological Sciences, University of Dundee, Dundee DD1 4HN, UK; and 7 Jardim Bota ˆnico do Rio de Janeiro, Rua Pacheco Lea ˜o No. 915, Gavea 22.460 Rio de Janeiro—RJ, Brazil A monophyletic pantropical group of papilionoid legumes, here referred to as the ‘‘dalbergioid’’ legumes, is circumscribed to include all genera previously referred to the tribes Aeschynomeneae and Adesmieae, the subtribe Bryinae of the Desmodieae, and tribe Dalbergieae except Andira, Hymenolobium, Vatairea, and Vataireopsis. This previously undetected group was discovered with phy- logenetic analysis of DNA sequences from the chloroplast trnK (including matK) and trnL introns, and the nuclear ribosomal 5.8S and flanking internal transcribed spacers 1 and 2. All dalbergioids belong to one of three well-supported subclades, the Adesmia, Dalbergia, and Pterocarpus clades. The dalbergioid clade and its three main subclades are cryptic in the sense that they are genetically distinct but poorly, if at all, distinguished by nonmolecular data. Traditionally important taxonomic characters, such as arborescent habit, free stamens, and lomented pods, do not provide support for the major clades identified by the molecular analysis. Short shoots, glandular-based trichomes, bilabiate calyces, and aeschynomenoid root nodules, in contrast, are better indicators of relationship at this hierarchical level. The discovery of the dalbergioid clade prompted a re-analysis of root nodule structure and the subsequent finding that the aeschynomenoid root nodule is synapomorphic for the dalbergioids. Key words: aeschynomenoid nodule; dalbergioid legumes; Fabaceae; papilionoid legumes; root nodule. The ‘‘dalbergioid’’ legumes are a previously unrecognized monophyletic group of papilionoid legumes in spite of the ex- tensive taxonomic history of its four constituents: tribes Ades- mieae, Aeschynomeneae, Dalbergieae, and Desmodieae sub- tribe Bryinae. The formal recognition of this group represents a major rearrangement of papilionoid legumes. It combines elements conventionally considered disparate and character- ized as either ‘‘primitive’’ or having varying levels of ‘‘ad- vancement’’ (Fig. 1). The Dalbergieae originally included tropical trees with fused floral parts and indehiscent pods (Bentham, 1860). Three subtribes were recognized: Pterocarpeae with samaroid pods, Lonchocarpeae marked by pods having at most small marginal wings, and Geoffroyeae having drupaceous fruits. Polhill (1971, 1981d, 1994) revised this classification by combining morphological evidence with that of seed chemistry and wood 1 Manuscript received 11 January 2000; revision accepted 2 June 2000. The authors thank Angela Beyra-M., Alfonso Delgado, Colin Hughes, Jean- Noel Labat, Gwilym Lewis, Darien Prado, Mats Thulin, and Martin Wojcie- chowski for kindly providing seed or leaf material of many of the species analyzed during this study, Alfonso Delgado, Martin Wojciechowski, and an anonymous reviewer for providing comments that greatly improved the man- uscript, Mats Thulin for making available his observations on the nectary disk in Ormocarpum and close relatives, William Anderson for loaning copies of the figures taken from Flora Novo-Galiciana, Sergio Faria for providing un- published information on root nodule morphology, Karin Douthit, Shona McInroy, and Maureen Warwick for illustrating the figures, and Tom Turley for technical laboratory assistance. This study was supported by a grant from the United States National Science Foundation (DEB-9615203), the Lever- hulme Trust, and the Royal Botanic Garden Edinburgh Molecular Phyloge- netic project. 2 Author for reprint requests (e-mail: [email protected]). anatomy. This new Dalbergieae included 19 tropical woody genera mainly from Bentham’s Pterocarpeae and Geoffroyeae. Lonchocarpinae were relegated to a closer relationship with other legumes that accumulated nonprotein amino acids in seed (e.g., Evans, Fellows, and Bell, 1985). The revised Dal- bergieae were diagnosed by supposedly plesiomorphic flower morphologies (i.e., free keel petals, staminal filaments partly fused and without basal fenestrae), pods with specialized seed chambers, and seeds that accumulated alkaloids or other than nonprotein amino acids. Geesink (1981, 1984) accepted Pol- hill’s circumscription with slight modification, whereas Sousa and de Sousa (1981) proposed a classification similar to Ben- tham’s because Dalbergieae (sensu Polhill, 1981d) supposedly shared a determinate inflorescence with the Lonchocarpinae. The Aeschynomeneae (Rudd, 1981a) are one of five tribes traditionally characterized by lomented pods (Polhill, 1981a). Although some Aeschynomeneae lack such pods (e.g., Arach- is, Ormocarpopsis, Diphysa spp., Ormocarpum spp., Pictetia spp.), none of the members of this tribe have ever been con- fused or classified with the genera of Dalbergieae. Adesmieae (Polhill, 1981f) have a notable history independent of the other dalbergioid legumes. This is because this tribe combines a pre- sumed plesiomorphic trait, free staminal filaments, with a sup- posedly very derived one, lomented pods. This combination has suggested either a taxonomically isolated position or a relationship with other papilionoids also with free stamens (e.g., Burkart, 1952). Bryinae, with lomented pods, possess other traits confirming its placement in the tribe Desmodieae (e.g., explosive secondary pollen presentation; Ohashi, Polhill, and Schubert, 1981). However, Bryinae have seeds that do not accumulate nonprotein amino acids and lack a structural mu-
Transcript of The Dalbergioid Legumes (Fabaceae): Delimitation of a Pantropical Monophyletic Clade

503
American Journal of Botany 88(3): 503–533. 2001.
THE DALBERGIOID LEGUMES (FABACEAE):DELIMITATION OF A PANTROPICAL
MONOPHYLETIC CLADE1
MATT LAVIN,2,3 R. TOBY PENNINGTON,4 BENTE B. KLITGAARD,5
JANET I. SPRENT,6 HAROLDO CAVALCANTE DE LIMA,7 AND
PETER E. GASSON5
3Department of Plant Sciences, Montana State University, Bozeman, Montana 59717 USA;4Tropical Biology Group, Royal Botanic Garden Edinburgh, 20a Inverleith Row, Edinburgh EH3 5LR, UK;
5Royal Botanic Gardens, Kew, Richmond, Surrey TW9 3AB, UK;6Department of Biological Sciences, University of Dundee, Dundee DD1 4HN, UK; and
7Jardim Botanico do Rio de Janeiro, Rua Pacheco Leao No. 915, Gavea 22.460 Rio de Janeiro—RJ, Brazil
A monophyletic pantropical group of papilionoid legumes, here referred to as the ‘‘dalbergioid’’ legumes, is circumscribed to includeall genera previously referred to the tribes Aeschynomeneae and Adesmieae, the subtribe Bryinae of the Desmodieae, and tribeDalbergieae except Andira, Hymenolobium, Vatairea, and Vataireopsis. This previously undetected group was discovered with phy-logenetic analysis of DNA sequences from the chloroplast trnK (including matK) and trnL introns, and the nuclear ribosomal 5.8Sand flanking internal transcribed spacers 1 and 2. All dalbergioids belong to one of three well-supported subclades, the Adesmia,Dalbergia, and Pterocarpus clades. The dalbergioid clade and its three main subclades are cryptic in the sense that they are geneticallydistinct but poorly, if at all, distinguished by nonmolecular data. Traditionally important taxonomic characters, such as arborescenthabit, free stamens, and lomented pods, do not provide support for the major clades identified by the molecular analysis. Short shoots,glandular-based trichomes, bilabiate calyces, and aeschynomenoid root nodules, in contrast, are better indicators of relationship at thishierarchical level. The discovery of the dalbergioid clade prompted a re-analysis of root nodule structure and the subsequent findingthat the aeschynomenoid root nodule is synapomorphic for the dalbergioids.
Key words: aeschynomenoid nodule; dalbergioid legumes; Fabaceae; papilionoid legumes; root nodule.
The ‘‘dalbergioid’’ legumes are a previously unrecognizedmonophyletic group of papilionoid legumes in spite of the ex-tensive taxonomic history of its four constituents: tribes Ades-mieae, Aeschynomeneae, Dalbergieae, and Desmodieae sub-tribe Bryinae. The formal recognition of this group representsa major rearrangement of papilionoid legumes. It combineselements conventionally considered disparate and character-ized as either ‘‘primitive’’ or having varying levels of ‘‘ad-vancement’’ (Fig. 1).
The Dalbergieae originally included tropical trees withfused floral parts and indehiscent pods (Bentham, 1860). Threesubtribes were recognized: Pterocarpeae with samaroid pods,Lonchocarpeae marked by pods having at most small marginalwings, and Geoffroyeae having drupaceous fruits. Polhill(1971, 1981d, 1994) revised this classification by combiningmorphological evidence with that of seed chemistry and wood
1 Manuscript received 11 January 2000; revision accepted 2 June 2000.The authors thank Angela Beyra-M., Alfonso Delgado, Colin Hughes, Jean-
Noel Labat, Gwilym Lewis, Darien Prado, Mats Thulin, and Martin Wojcie-chowski for kindly providing seed or leaf material of many of the speciesanalyzed during this study, Alfonso Delgado, Martin Wojciechowski, and ananonymous reviewer for providing comments that greatly improved the man-uscript, Mats Thulin for making available his observations on the nectary diskin Ormocarpum and close relatives, William Anderson for loaning copies ofthe figures taken from Flora Novo-Galiciana, Sergio Faria for providing un-published information on root nodule morphology, Karin Douthit, ShonaMcInroy, and Maureen Warwick for illustrating the figures, and Tom Turleyfor technical laboratory assistance. This study was supported by a grant fromthe United States National Science Foundation (DEB-9615203), the Lever-hulme Trust, and the Royal Botanic Garden Edinburgh Molecular Phyloge-netic project.
2 Author for reprint requests (e-mail: [email protected]).
anatomy. This new Dalbergieae included 19 tropical woodygenera mainly from Bentham’s Pterocarpeae and Geoffroyeae.Lonchocarpinae were relegated to a closer relationship withother legumes that accumulated nonprotein amino acids inseed (e.g., Evans, Fellows, and Bell, 1985). The revised Dal-bergieae were diagnosed by supposedly plesiomorphic flowermorphologies (i.e., free keel petals, staminal filaments partlyfused and without basal fenestrae), pods with specialized seedchambers, and seeds that accumulated alkaloids or other thannonprotein amino acids. Geesink (1981, 1984) accepted Pol-hill’s circumscription with slight modification, whereas Sousaand de Sousa (1981) proposed a classification similar to Ben-tham’s because Dalbergieae (sensu Polhill, 1981d) supposedlyshared a determinate inflorescence with the Lonchocarpinae.
The Aeschynomeneae (Rudd, 1981a) are one of five tribestraditionally characterized by lomented pods (Polhill, 1981a).Although some Aeschynomeneae lack such pods (e.g., Arach-is, Ormocarpopsis, Diphysa spp., Ormocarpum spp., Pictetiaspp.), none of the members of this tribe have ever been con-fused or classified with the genera of Dalbergieae. Adesmieae(Polhill, 1981f) have a notable history independent of the otherdalbergioid legumes. This is because this tribe combines a pre-sumed plesiomorphic trait, free staminal filaments, with a sup-posedly very derived one, lomented pods. This combinationhas suggested either a taxonomically isolated position or arelationship with other papilionoids also with free stamens(e.g., Burkart, 1952). Bryinae, with lomented pods, possessother traits confirming its placement in the tribe Desmodieae(e.g., explosive secondary pollen presentation; Ohashi, Polhill,and Schubert, 1981). However, Bryinae have seeds that do notaccumulate nonprotein amino acids and lack a structural mu-

504 [Vol. 88AMERICAN JOURNAL OF BOTANY
Fig. 1. Putative relationships among tribes of the subfamily Papilionoideae according to Polhill (1981a). Tribes underscored include genera that are nowknown to be members of the dalbergioid clade (e.g., Desmodieae then included subtribe Bryinae, and Robinieae the genus Diphysa). Accumulation of nonproteinamino acids and fusion of floral parts occur frequently in Tephrosieae and all tribes positioned above it. The absence of such traits is traditionally viewed asprimitive and is most frequent in tribes positioned below Tephrosieae.
tation in the chloroplast rpl2 locus (Bailey et al., 1997). Bothare atypical of the rest of Desmodieae.
In spite of a taxonomic history of Dalbergieae that has beenseparate from those of Aeschynomeneae, Adesmieae, andBryinae, we present evidence that they collectively form amonophyletic group. The focus on these putatively disparatetaxa was motivated by the taxonomic distribution of the dis-tinctive aeschynomenoid root nodule (Corby, 1981; Faria etal., 1994) and four cladistic analyses: three involving non-molecular data (Lavin, 1987; Chappill, 1995; Beyra-M. andLavin, 1999), and one with rbcL sequence data (Doyle et al.,1997). We have expanded on these previous analyses by sam-pling exhaustively to reveal the exact constituents of the dal-bergioid clade and enumerate the nonmolecular characters thathave been used in the conventional tribal classification of theselegumes. As such, we demonstrate where molecular and non-molecular data are taxonomically concordant. We also showthat many traditionally important taxonomic characters in thisgroup are more homoplasious than previously considered. Be-cause taxon sampling has focused on just the putative mem-bers of the dalbergioid clade, a point to be briefly addressedhere but more thoroughly developed elsewhere is the higherlevel relationships of this newly recognized clade (Hu et al.,2000; Pennington et al., in press; M. Wojciechowski et al.,unpublished data).
MATERIALS AND METHODS
DNA sequence data—DNA isolations, polymerase chain reaction (PCR)amplifications, and template purifications were performed with Qiagen Kits(i.e., DNeasy Plant Mini Kit, Taq PCR Core Kit, QIAquick PCR PurificationKit; Qiagen, Santa Clarita, California, USA). DNA sequences analyzed werethe nuclear ribosomal 5.8S and flanking internal transcribed spacers (ITS1and ITS2), the chloroplast trnK intron, including matK, and the trnL intron.PCR and sequencing primers for ITS and 5.8S sequences are described in
Beyra-M. and Lavin (1999) and Delgado-Salinas et al. (1999). Primers formatK and flanking trnK intron sequences are described in Lavin et al. (2000).Primers for the trnL intron are described by Taberlet et al. (1991). DNAsequencing was performed on an automated sequencer at the Iowa State Uni-versity DNA Sequencing Facility (Ames, Iowa, USA) and Davis Sequencing(Davis, California, USA).
DNA sequences were aligned manually with Se-Al (Rambaut, 1996). Biasintroduced by the manual alignment was evaluated with a sensitivity analysis(cf. Whiting et al., 1997; Beyra-M. and Lavin, 1999; Delgado-Salinas et al.,1999). Alignment-variable regions were variably aligned or excluded, a stepmatrix (cf. Cunningham, 1997) was invoked or not, and gaps were treated asmissing data, a fifth state, or as separate characters. Each of the differentsensitivity analyses were subjected to the same heuristic search options. Miss-ing data included 12.9% of the matK/trnK data set, 5.4% of the trnL data set,1.5% of the ITS/5.8S data set, and 7.6% of the nonmolecular data set.
Maximum parsimony analyses were performed with PAUP* (Swofford,2000). Heuristic search options included 100 random-addition replicates, tree-bisection-reconnection branch swapping, and steepest descent. A maximumof 10 000 trees was allowed to accumulate, which is sufficient to capture alltopological variation (cf. Sanderson and Doyle, 1993). Clade stability testsinvolved bootstrap resampling (Felsenstein, 1985; Sanderson, 1995), whereeach of the 10 000 bootstrap replicates was subjected to heuristic search op-tions that included one random-addition sequence per replicate, swapping withtree-bisection-reconnection, and invoking neither steepest descent nor mul-pars.
Taxon sampling—Sampling of molecular and nonmolecular data was asexhaustive as possible at the generic level in order to determine membershipin the dalbergioid clade, as well as the principal phylogenetic structure withinthis clade. Molecular and nonmolecular data were obtained for at least onespecies from every genus ever placed in the Dalbergieae (Burkart, 1952; Pol-hill, 1981d), Aeschynomeneae (Rudd, 1981a), Adesmieae (Polhill, 1981f), orBryinae (Ohashi, Polhill, and Schubert, 1981). The only exception is the pre-sumably extinct genus Peltiera (Labat and Du Puy, 1997), where no success-ful PCR amplifications were obtained from the few available DNAs. In ad-dition to the advantages of being able to detail the taxonomic implications,

March 2001] 505LAVIN ET AL.—DALBERGIOID LEGUMES
exhaustive sampling for molecular data increases the probability of subdivid-ing long branches (e.g., Hillis, 1998).
Our original intent was to sample the same DNA accessions for each ofthe data sets. This proved impossible for DNA sequences because of incon-sistencies in DNA quality and quantity and PCR amplification. We conse-quently had to resort to multiple methods of sampling. The DNA sequencedata were sampled using the exemplar approach. Multiple species per terminaltaxon were sampled where possible (Appendix A). Because nonmolecular dataare generally open to visual inspection across all species of a particular ter-minal taxon, the ‘‘democratic’’ method of sampling (Bininda-Emonds, Bryant,and Russell, 1998) was used for nonmolecular data. In this approach, weincluded all possible character states represented by any one terminal, whichwas usually a traditionally recognized genus (i.e., multistate terminal taxawere coded). The reasoning is that in the evaluation of traditionally importanttaxonomic characters, the degree of polymorphisms within terminals shouldbe explicitly enumerated. For those few terminals in which species-level phy-logenetic analysis has been completed (e.g., Andira and Pictetia), we em-ployed the ancestral method of sampling nonmolecular data (Bininda-Emonds,Bryant, and Russell, 1998). The justification for ultimately combining datathat have been sampled differently is that a combined analysis should stillallow us to best estimate where the traditionally important taxonomic char-acters lie on the continuum from strongly phylogenetically constrained tomaximally homoplasious.
The genera Bergeronia, Dalbergiella, Lonchocarpus, and Muellera havebeen placed in the tribe Dalbergieae (e.g., Burkart, 1952; Geesink, 1981) andPongamiopsis has been synonymized with the genus Aeschynomene (Hutch-inson, 1964). However, they were not included in this analysis because otherphylogenetic analyses (Lavin et al., 1998; Hu et al., 2000) have shown thesegenera to be closely related to Millettia and relatives, all of which accumulatenonprotein amino acids in seed. Similarly, Poecilanthe and Cyclolobiumshould be allied with more basal Papilionoideae that accumulate alkaloids inseed (Greinwald et al., 1995; Lavin et al., 1998; Hu et al., 2000). This is thereason that Poecilanthe is retained as a designated outgroup.
Outgroups were sampled extensively as part of large-scale molecular phy-logenetic studies of the subfamily Papilionoideae (Hu et al., 2000; Penningtonet al., in press; M. Wojciechowski et al., unpublished data). Sampling out-groups was guided by phylogenetic studies involving nonmolecular data (e.g.,Chappill, 1995; Herendeen, 1995; Beyra-M. and Lavin, 1999). For example,all outgroups chosen have leaves with punctate glands, a trait common todalbergioids. In the end, the outgroups retained in this analysis included Acos-mium and Myrospermum (tribe Sophoreae; Polhill, 1981b), Dipteryx and Pter-odon (Dipterygeae; Polhill, 1981c), Poecilanthe (variously classified; see Lav-in and Sousa, 1995), and Apoplanesia, Amorpha, Eysenhardtia, and Marina(tribe Amorpheae; Barneby, 1977; Polhill, 1981e). This sampling was consid-ered sufficient to demonstrate membership in the dalbergioid clade. The find-ings reported here did not change with a more extensive sampling of out-groups.
Sampling for the molecular data was re-evaluated as aligned DNA sequenc-es accumulated. It became obvious that the matK/trnK sequences were by farthe most informative at higher taxonomic levels, as seen in increased reso-lution in the strict consensus and higher bootstrap values. The primary effortthen changed to sample as exhaustively as possible matK/trnK sequences and,secondarily, the ITS/5.8S and trnL intron sequences. Thus, the data analysisof this study centers on the matK/trnK data set. Sampling of ITS/5.8S se-quences was guided by species level analyses of certain dalbergioid genera(e.g., Beyra-M. and Lavin, 1999; Lavin et al., 2000). Sampling of the trnLintron data was guided by a phylogenetic analysis of putatively basal Papi-lionoideae (Pennington et al., in press). Unevenness in sampling was exac-erbated by inconsistencies in PCR amplifications (mentioned above). A com-bined molecular analysis was not attempted because unevenness in samplingwould result in a combined data set not exhaustively sampled at the genuslevel. Thus, consensus among the data sets was evaluated by congruence ofthe major clades resolved with high bootstrap values (cf. Huelsenbeck, Bull,and Cunningham, 1996).
Nonmolecular character analysis—A nonmolecular data set was devel-
oped from that in Beyra-M. and Lavin (1999) and is presented in AppendixB. Characters that have been considered traditionally important in the tax-onomy of Dalbergieae, Aeschynomeneae, Adesmieae, and Bryinae (e.g., Bur-kart, 1952; Ohashi, Polhill, and Schubert, 1981; Polhill, 1981d; Rudd, 1981a;Sousa and de Sousa, 1981) were targeted for analysis. As discussed above,multistate taxa were coded as polymorphic (cf. Weins, 1995; Weins and Ser-vedio, 1997), in spite of the recommendation of Nixon and Davis (1991).Although this can underestimate the degree of homoplasy (see individual char-acter discussions in Appendix B), splitting polymorphic terminals into two ormore monomorphic ones does not change our findings (e.g., as evaluated inthe fashion of a sensitivity analysis). This is because the focus is strictly atwide-scale relationships of groups of genera, and the potentially problematicpolymorphisms are at a different level, within genera. Polymorphisms arediscussed in the presentation of characters or ingroup terminal taxa (Appen-dices B and C). Inapplicable character states in certain terminals (e.g., leaftraits of Ramorinoa, a genus that doesn’t produce leaves) were variously treat-ed as a missing state, an uncertain state, or an extra state (as in a sensitivityanalysis). The nonmolecular data were gathered primarily from field obser-vations or herbarium specimens. Literature reports were usually verified byobservations of the plants.
RESULTS
Parsimony analysis of the 1266 informative sites from the95 taxa by 2966 sites matK/trnK data set produced 10 000 trees(the set maximum) each with a minimal length of 4352, aconsistency index of 0.570 and a retention index of 0.830. Themonophyly of the dalbergioid clade, including all genera ofAeschynomeneae, Adesmieae, Bryinae, and most Dalbergieae,was very well supported by bootstrap analysis (Fig. 2). Fourmembers of tribe Dalbergieae (Andira, Hymenolobium, Vatai-rea, and Vataireopsis) and two sampled genera of Dipterygeae(Dipteryx and Pterodon) were not included. Indeed, the sistergroup to the dalbergioid clade includes genera sampled fromthe tribe Amorpheae (Apoplanesia and Amorpha). Within thedalbergioid clade, there are three well-supported subcladesmarked as the Adesmia, Dalbergia, and Pterocarpus clades(Fig. 2). The earliest branching Adesmia clade includes thegenus Adesmia (sole member of the tribe Adesmieae) andmostly herbaceous to subshrubby genera of the tribe Aeschy-nomeneae (Poiretia, Amicia, Zornia, Chaetocalyx, and Nis-solia). The remaining two subclades each include members ofthe Aeschynomeneae and Dalbergieae. The Pterocarpus cladeadditionally includes two genera, Brya and Cranocarpus, ofDesmodieae (subtribe Bryinae).
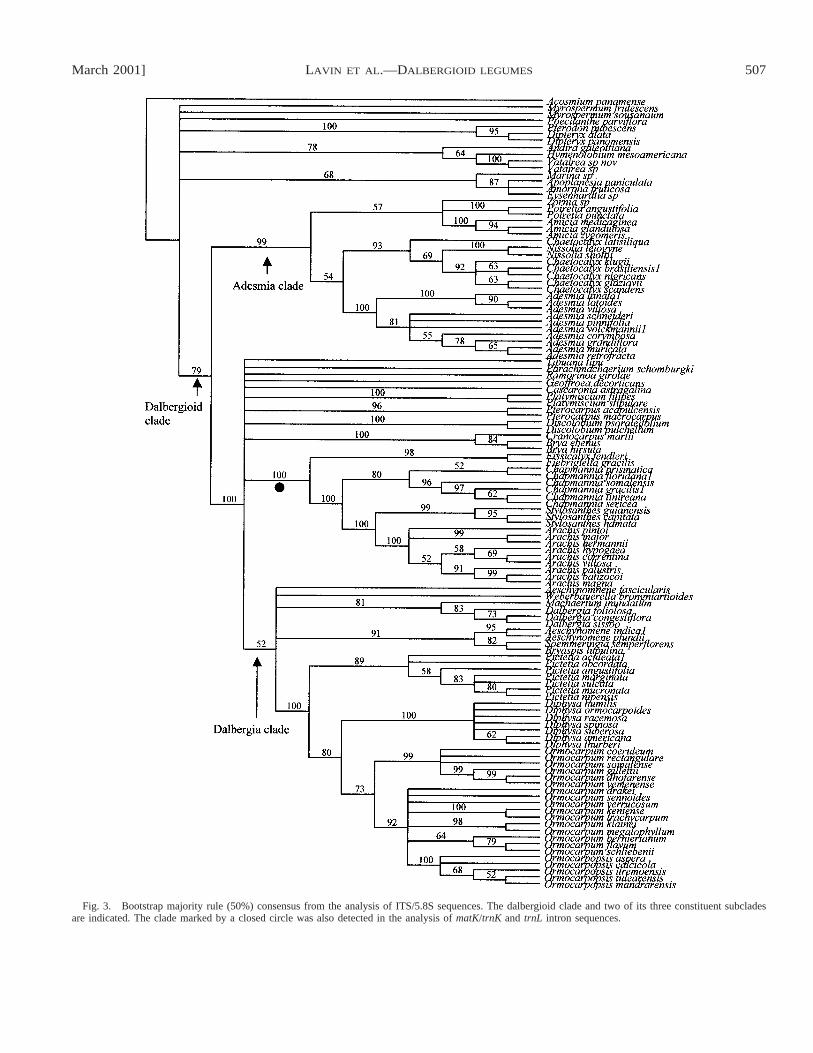
For the 481 informative sites from the 118 taxa by 719 sitesITS/5.8S data set, 120 trees were generated each with a min-imal length of 5009, a consistency index of 0.259, and a re-tention index of 0.714. The same higher level relationshipsdescribed for the matK/trnK analysis were resolved in thisanalysis, though with less bootstrap support (Fig. 3). Althoughthe Pterocarpus clade was resolved in the strict consensus ofthe parsimony analysis, it was resolved in less than 50% ofthe analyses of the bootstrap replicates. In no case (majority-rule bootstrap consensus or strict consensus of minimal lengthtrees) was the sister-group relationship of the Amorpheae sam-ples resolved.
Analysis of the 293 informative sites from the 93 taxa by737 sites trnL intron data set generated 10 000 trees each witha minimal length of 1102, a consistency index of 0.603, anda retention index of 0.804. Although the dalbergioid clade iswell resolved by bootstrap analysis, only the Adesmia cladeis further resolved (Fig. 4). Not in any case was the Dalbergiaor Pterocarpus clades resolved as monophyletic. Regardless,

506 [Vol. 88AMERICAN JOURNAL OF BOTANY
Fig. 2. Bootstrap majority rule (50%) consensus from the analysis of matK/trnK sequences. The dalbergioid clade and its three constituent subclades areindicated.
March 2001] 507LAVIN ET AL.—DALBERGIOID LEGUMES
Fig. 3. Bootstrap majority rule (50%) consensus from the analysis of ITS/5.8S sequences. The dalbergioid clade and two of its three constituent subcladesare indicated. The clade marked by a closed circle was also detected in the analysis of matK/trnK and trnL intron sequences.

508 [Vol. 88AMERICAN JOURNAL OF BOTANY
Fig. 4. Bootstrap majority rule (50%) consensus from the analysis of trnL intron sequences. The dalbergioid clade and the Adesmia subclade are indicated.Clades marked by a closed circle were also detected in the analysis of matK/trnK and ITS/5.8S sequences.

March 2001] 509LAVIN ET AL.—DALBERGIOID LEGUMES
the relationships resolved by majority-rule bootstrap consensusdid not conflict with those similarly resolved in either thematK/trnK and ITS/5.8S analyses.
Analysis of the 55 nonmolecular characters (Appendix B)yielded poorly resolved and supported relationships, such thatthe majority-rule bootstrap consensus was largely unresolvedabove the genus level. Resolved intergeneric relationships in-clude a clade with Aeschynomene, Cyclocarpa, Bryaspis, Geis-saspis, Humularia, Kotschya, Smithia, and Soemeringia (60%bootstrap support), one with Chapmannia, Arachis, and Sty-losanthes (65%), Brya and Cranocarpus (67%), Chaetocalyxand Nissolia (100%), Amicia, Poiretia, and Zornia (67%), andOrmocarpopsis and Peltiera (93%). Because Peltiera is notrepresented by DNA sequence data, this nonmolecular dataprovide the only evidence for its relationships (the relation-ships of Peltiera are a focus of another study; M. Thulin andM. Lavin, unpublished data). The only well-supported cladethat was resolved during this analysis and that was not seenduring the previous molecular analyses was one with Etaballiaand Inocarpus (80%), apomorphically diagnosed as havingnearly regular flowers (characters 22–23 in Appendix B).
Because of the poorly resolved relationships obtained fromanalysis of the nonmolecular data set, it was combined withthe matK/trnK data set in order to explore the evolution of thetraditionally important taxonomic characters. Integration withjust the matK/trnK is justified by how well this data set canresolve relationships (discussed in MATERIALS ANDMETHODS) and because of noncompatibility of moleculardata sets with respect to sampling. Parsimony analysis of the1319 informative characters of the combined matK/trnK andnonmolecular data set (95 taxa by 3021 characters) produced2340 trees with a minimal length of 4664, each with a con-sistency index of 0.551 and a retention index of 0.821. Theresulting relationships are essentially those described previ-ously for the analysis of just the matK/trnK data set (Fig. 5).
Sensitivity analysis—Making different assumptions aboutthe molecular data sets, deleting characters with many missingentries (e.g., nonmolecular characters 50–54), splitting poly-morphic terminals into two or more monomorphic ones, orrecoding inapplicable nonmolecular characters to uncertainstates, missing data, or as an extra state, did not affect theresults described above (Figs. 2–5). The monophyly of thedalbergioid legumes was consistently resolved, as generallywas the monophyly of the three constituent subclades. Therewere no cases of clades with bootstrap values over 70% thatconflicted among the molecular data sets. Also, clades withhigh bootstrap values (i.e., .90%) in individual analyses ofthe matK/trnK, ITS/5.8S, trnL intron, or combined nonmolec-ular and matK/trnK data sets were consistently resolved re-gardless of the assumptions made about any one of the partic-ular data sets. This is exemplified by analysis of just the matKcoding region (i.e., excluding the flanking noncoding portionof the trnK intron), where some accessions in the data matrixwere missing either the 59 or 39 half of this locus (for a totalof 12.1% missing entries). The strict consensus of the parsi-mony analysis of the matK locus was essentially identical tothat of the analysis of the matK/trnK data set. Bootstrap anal-ysis resulted in values that were sometimes lower than in theanalysis of the entire matK/trnK data set: 80% for the Amor-pheae 1 dalbergioid clade, 100% for the dalbergioid clade,100% for the Adesmia clade, 94% for the Dalbergia clade, and71% for the Pterocarpus clade.
DISCUSSION
As now circumscribed, the dalbergioids comprise 44 genera(Appendix C) and ;1100 species of trees, shrubs, and peren-nial to annual herbs. Included are economically importanthardwoods (e.g., Dalbergia and Pterocarpus spp.), forage le-gumes (Stylosanthes spp.), and crops (e.g., Arachis spp.). Likemost pantropical legume taxa, the dalbergioids are concen-trated in the neotropics and subSaharan Africa. Although theposition of the dalbergioid clade within the Fabaceae is notfully developed here, its sister group is the tribe Amorpheae,which contains eight New World genera confined mostly towarm temperate and tropical North America. What is generallycertain of higher level relationships is that the dalbergioids aredistantly related to papilionoids that accumulate nonproteinamino acids in seed. This most notably includes Lonchocar-pus, Derris, Millettia, and Hologalegina (e.g., tribes Robi-nieae, Galegeae, etc.; Wojciechowski, Sanderson, and Hu,1999), which at times have been taxonomically confused withvarious elements now included in the dalbergioid clade.
Implications for traditional classifications—The classifi-cation of certain genera into tribes and subtribes of Papilion-oideae (e.g., Rudd, 1981a; Ohashi, Polhill, and Schubert,1981; Ohashi, 1999; Polhill, 1981a, d) needs to be greatlymodified in light of the evidence presented here. The generaBrya and Cranocarpus (subtribe Bryinae of tribe Desmodieae)share many unusual synapomorphies, such as periporate pollenand glochidiate trichomes, that have served to obscure higherlevel relationships. The explosive pollen presentation mecha-nism that Brya shares in common with Desmodieae is shownto have evolved independently. So have the lomented pods thatBrya and Cranocarpus share with Desmodieae.
Four of the five subtribes of Aeschynomeneae are eithermonotypic (e.g., Discolobiinae) or are polyphyletic. Aeschy-nomeneae subtribe Ormocarpinae includes three different el-ements: Diphysa, Ormocarpum, Ormocarpopsis (and Pelti-era), and Pictetia form one lineage in the Dalbergia clade,Fiebrigiella is in the Pterocarpus clade, and Chaetocalyx andNissolia are part of the Adesmia clade. The pod valves withdistinctive parallel venation that previously allied all of thesegenera now are considered to have evolved on three separateoccasions. Indeed, this derived pod trait is homologous amongFiebrigiella, Chapmannia, Arachis, and Stylosanthes.
Aeschynomeneae subtribe Poiretiinae includes two differentelements. Amicia, Poiretia, and Zornia form a monophyleticgroup within the Adesmia clade, and Weberbauerella is phy-logenetically isolated within the Dalbergia clade. The markedpustular glands of Weberbauerella are no longer consideredhomologous to those of Amicia, Poiretia, and Zornia. In therecent classification of Japanese legumes (Ohashi, 1999), Poir-etia and Zornia are classified as the sole members of the tribePoiretieae, a taxonomy that finds no support in this analysis.
Aeschynomeneae subtribe Aeschynomeninae includes eightgenera (Aeschynomene, Cyclocarpa, Soemmeringia, Kotschya,Smithia, Humularia, Bryaspis, and Geissaspis) that form avery well-supported monophyletic group. A nonmolecularcharacter supporting this relationship is the medifixed stipule,although it is not universal in this clade and has evolved in-dependently in Zornia. An extrapolation from our small sam-ple, however, suggests that species of Aeschynomene havingbasifixed stipules (e.g., A. fascicularis and A. purpusii) aremore closely related to Machaerium and Dalbergia than they

510 [Vol. 88AMERICAN JOURNAL OF BOTANY
Fig. 5. Bootstrap majority rule (50%) consensus from the analysis of combined nonmolecular and matK/trnK sequence data. The dalbergioid clade and itsthree constituent subclades are indicated.

March 2001] 511LAVIN ET AL.—DALBERGIOID LEGUMES
are to the species of Aeschynomene with medifixed stipules.Thus, the subtribe Aeschynomeninae includes two disparateelements.
Only Aeschynomeneae subtribe Stylosanthinae, with Arach-is, Stylosanthes, and Chapmannia (and the segregates Pache-coa and Arthrocarpum), has been long recognized as a distincttaxonomic group and is also revealed as monophyletic in thisanalysis. The well-known nonmolecular character supportingthe monophyly of this clade is a sessile papilionoid flower witha long hypanthium. However, these three genera are veryclosely related to Fiebrigiella and Fissicalyx and together allof these genera are set apart from other members of the Pter-ocarpus clade by large genetic distances. Notably, nonmolec-ular characters do not support most of the relationships in thisclade that are so well supported by independent moleculardata. For example, there are no known nonmolecular data thatsupport the monophyly of the genus Chapmannia (Thulin,2000) or the relationship of Fissicalyx and Fiebrigiella.
The tribe Dalbergieae also is not monophyletic. Excludedfrom the dalbergioid clade are Andira, with 30 species largelyconfined to the neotropics and with one species distributed inthe neotropics and tropical Africa (Lima, 1990; Pennington,1996; Pennington, Aymard, and Cuello, 1997), Hymenolobiumwith 10–15 species in tropical South America and one speciesin Central America (Polhill, 1981d; Lima, 1982a, 1990), Va-tairea with seven species from Mexico to Brazil (Lima, 1982b,1990), and Vataireopsis with three species in Brazil and theGuianas (Polhill, 1981d; Lima, 1990). The distinction of thesefour genera from others traditionally included in the tribe Dal-bergieae has been noted with wood anatomy (Baretta-Kuipers,1981) and estimates of overall similarity (Lima, 1990). Forexample, the wood of Andira, Hymenolobium, Vatairea, andVataireopsis lacks the storied structure and uniseriate rays thatare characteristic of dalbergioid wood and is generally of lesscommercial value.
The remaining genera of the tribe Dalbergieae belong toeither the Dalbergia or Pterocarpus clades. Only Dalbergia andMachaerium are part of the Dalbergia clade, where they aremost closely related to Aeschynomene species that have basi-fixed stipules. The rest of the genera previously classified inthe tribe Dalbergieae form the bulk of the Pterocarpus cladealong with some genera previously classified in the tribe Aes-chynomeneae (e.g., Fiebrigiella, Chapmannia, Arachis, Sty-losanthes, and Discolobium) and subtribe Bryinae of Desmo-dieae.
The genera of Dipterygeae (Taralea, Dipteryx, and Ptero-don; Polhill, 1981c) are not part of the dalbergioid clade. Bur-kart (1952) originally included Dipteryx (then Coumarouna)in the tribe Dalbergieae, and a phylogenetic analysis of non-molecular data by Beyra-M. and Lavin (1999) suggested Dip-terygeae was part of the dalbergioid clade. Even the combi-nation of paripinnate leaves bearing glandular punctae isknown only from Dipterygeae and the dalbergioid legumes.However, this analysis strongly suggests that the punctateglands are plesiomorphic because they are found in all generaincluded in this analysis. Paripinnate leaves evolved indepen-dently among Dipterygeae and various elements in the dal-bergioid clade.
Phylogenetic information among the various nonmolecularcharacters—While the matK/trnK phylogeny was not greatlyinfluenced by the addition of the 55 nonmolecular characters(compare Figs. 2 and 5), there is some phylogenetic infor-
mation in the nonmolecular characters, as evinced by high re-tention indices (Table 1). The consistency (CI) and retention(RI) indices for each of the 55 nonmolecular characters (Ap-pendix B) in the combined analysis were compared to thesame values obtained when each of the nonmolecular char-acters was mapped onto the matK/trnK phylogeny. In the com-bined analysis, the average CI and RI were 0.427 and 0.672,respectively. When mapped onto the matK/trnK trees, the av-erage CI and RI were 0.390 and 0.627, respectively. Regard-less of the small but significant differences (for RI, two-tailedt test, t 5 2.94, P 5 0.005, df 5 52), no character had ahigher consistency or retention index when mapped onto thematK/trnK phylogeny as when combined with the matK/trnKsequence data during parsimony analysis. This suggests thatmapping a few selected nonmolecular characters onto a mo-lecular phylogeny may involve a bias of excess levels of ho-moplasy.
Different classes of characters (e.g., vegetative, floral, andfruiting) were equally as prone to having homoplasy overes-timated when mapped onto a molecular phylogeny. These in-clude, for example, an asymmetric leaflet base (character 9 inAppendix B), persistent floral bracts (character 16), and a longpod stipe (character 33). The states of the leaflet base had anaverage retention index of 1.000 in the combined analysis and0.500 when mapped to the matK/trnK trees (Table 1). Thecorresponding values were 0.667 and 0.167 for the states ofthe floral bracts, and 0.647 and 0.559 for the pod stipe (Table1). Also, no particular class of characters (e.g., vegetative, flo-ral, and fruiting) was more informative than another. For veg-etative characters (1–13, 44–45, 50–55), the average retentionindex is 0.705. For floral characters (14–30, 46–49), it is0.604. For fruiting characters (31–43), the average retentionindex is 0.726. These differences are not significant (single-factor ANOVA, F 5 1.174, P 5 0.317, df 5 52). The lackof a difference in behavior among the various classes of char-acters, as also generally found by Bateman and Simpson(1998) for vascular plants, weakens the suggestion of Tuckerand Douglas (1994) that floral characters necessarily providethe best taxonomic information in Leguminosae. These find-ings also weaken the implication that pod morphology is proneto higher rates of convergent evolution than other types ofcharacters (e.g., Geesink, 1984; Hu et al., 2000).
Conventional taxonomic evidence—Some traditionally im-portant taxonomic characters are determined in this analysis tobe more homoplasious than previously considered. This is es-pecially true of the character states of growth habit, staminalfusion, and pod segmentation. Herbaceous and woody relativesgenerally are separated into different taxonomic groups whena temperate vs. tropical distinction correlates with habit (Judd,Sanders, and Donoghue, 1994). This is especially true of pap-ilionoid legumes where tribes have been categorized by habit(e.g., temperate herbaceous vs. tropical woody tribal divisionin Polhill, 1981a, 1994). An herbaceous habit (number 1 inAppendix B) has evolved at least three times in monomorphiccondition but more times than this in polymorphic condition(Table 1). The Adesmia clade contains mostly herbaceous spe-cies, although some species of Adesmia and Poiretia areshrubs. That an herbaceous growth form maps as the ancestralstate in the Adesmia clade stands in contrast to the conven-tional wisdom that woody taxa form basal clades in tropicalPapilionoideae (e.g., Polhill, 1981a; Tucker and Douglas,1994).

512 [Vol. 88AMERICAN JOURNAL OF BOTANY
TABLE 1. Average lengths (L) and consistency (CI) and retention (RI) indices for each of the 55 nonmolecular characters. These are comparedfor the combined analysis and when each of the 55 is mapped onto the matK/trnK phylogeny. An ‘‘5’’ indicates that the CI and RI of thecombined and mapped character are equal. A ‘‘.’’ signifies a higher CI and RI value for a character in the combined analysis compared towhen mapped. The reverse situation did not occur.
Number
Combined analysis
L CI RI
Mapped
L CI RI
12345
5.51.06.02.08.0
0.4171.0000.3330.5000.125
0.8501.0000.6670.8330.562
.5555
6.01.06.02.08.0
0.3331.0000.3330.5000.125
0.8001.0000.6670.8330.562
6789
10
5.04.08.01.02.0
0.2000.5000.1251.0000.500
0.7890.5000.7671.0000.833
555.5
5.04.08.02.52.0
0.2000.5000.1250.4170.500
0.7890.5000.7670.5000.833
1112131415
9.07.02.02.07.0
0.1110.1431.0000.5000.429
0.7240.6251.0000.6670.778
.555.
10.07.02.02.09.0
0.1000.1431.0000.5000.333
0.6900.6251.0000.6670.667
1617181920
2.010.010.06.04.0
0.5000.2000.2000.8330.500
0.6670.7780.7780.9550.833
.
.
.
.5
3.511.512.0
7.04.0
0.2920.1750.1670.7140.500
0.1670.7360.7220.9090.833
2122232425
4.05.02.08.05.0
0.2500.2000.5000.1250.200
0.8120.2000.0000.3640.429
55555
4.05.02.08.05.0
0.2500.2000.5000.1250.200
0.8120.2000.0000.3640.429
262728
15.02.0
10.0
0.2670.5000.200
0.6330.8570.800
555
15.02.0
10.0
0.2670.5000.200
0.6330.8570.800
2930313233
5.01.09.06.0
13.0
0.2001.0000.2500.1670.077
0.000
0.8250.5830.647
55...
5.01.0
10.07.0
16.0
0.2001.0000.2000.1430.062
0.000
0.8000.5000.559
3435363738
2.51.0
17.02.05.0
0.4171.0000.2350.5000.400
0.912
0.4090.8890.625
.5.55
3.01.0
17.52.05.0
0.3331.0000.2290.5000.400
0.882
0.3870.8890.625
3940414243
1.02.03.0
11.04.0
1.0001.0000.6670.1820.250
1.0001.0000.7500.5000.571
555.5
1.02.03.0
12.04.0
1.0001.0000.6670.1670.250
1.0001.0000.7500.4440.571
4445464748
5.07.01.0
14.53.0
0.2000.1431.0000.0690.333
0.6000.5001.0000.6250.333
.55.5
6.07.01.0
15.03.0
0.1670.1431.0000.0670.333
0.5000.5001.0000.6110.333
4950515253
6.02.04.07.03.0
0.5000.5000.2500.1430.333
0.5710.7500.4000.7390.000
.5555
8.02.04.07.03.0
0.3750.5000.2500.1430.333
0.2860.7500.4000.7390.000
5455MeanSD
1.07.05.43.9
1.0000.2860.4270.301
1.0000.6670.6720.254
.5
2.07.05.84.2
0.5000.2860.3900.279
0.6670.6670.6270.258
Genera containing both woody and herbaceous species alsooccur in the clade containing Aeschynomene sect. Aeschyno-mene, Kotschya, Humularia, and Geissaspis. The same is truefor the clade including Fiebrigiella, Chapmannia, Stylosan-
thes, and Arachis. Fissicalyx and some species of Chapmanniaare woody in a clade dominated by herbaceous to subshrubbyspecies. Representing yet two other clades, species of Ma-chaerium, Dalbergia, Brya, and Cranocarpus vary from trees

March 2001] 513LAVIN ET AL.—DALBERGIOID LEGUMES
Figs. 6–8. Selected nonmolecular characters (scale bar 5 1 cm for all figures). 6. Aeschynomenoid root nodule associated with lateral root (character number55, Appendix B). 7. Short shoots of Ormocarpum (character number 2). 8. Pseudopetiole of Arachis (character number 4).
or shrubs to weak subshrubs. Clearly, there is no evidencefrom this analysis that the ability to produce a strongly woodygrowth habit is a good indicator of relationship.
The staminal character number 26 (Appendix B) includesfive states that provide an average length of 15.0 to the mostparsimonious trees. The consistency index of 0.267 and theretention index of 0.633 demonstrate that this character ishomoplasious. Even state zero, free staminal filaments, addeda length of two because this state occurs ancestrally in someof the outgroup genera and represents a reversion in the genusAdesmia. That a legume group with free stamens can evolvethis condition secondarily from a fused condition (e.g., 9 1 1diadelphous) is not surprising. Four species of Pictetia havenearly free staminal filaments in a clade otherwise representedby species with fused filaments (Beyra-M. and Lavin, 1999).Also, Kass and Wink (1995, 1997) have implicitly shown inan unrelated papilionoid group that the evolution of staminalmorphology does not necessarily involve a unique transfor-mation from free filaments into the fused condition. Perhapsrelated to this issue, Klitgaard (1999a) showed that order ofinitiation and loss of stamens are more variable among thedalbergioids than previously appreciated. No doubt, the apriori view that free staminal filaments represent necessarily aplesiomorphic condition among papilionoid legumes will haveto be abandoned.
All papilionoid legumes with lomented pods were at onetime classified together, although more recently five tribes(Adesmieae, Aeschynomeneae, Coronilleae, Desmodieae, andHedysareae) were thought to have gained this pod type inde-pendently (Polhill, 1981a). We scored three states pertainingto articulation of pod segments (number 31 in Appendix B),which added an average length of 9.0 to the most parsimoni-ous trees. The consistency index of 0.250 and a retention indexof 0.825 suggest that, although homoplasious, this characterprovided phylogenetic resolution towards the tips of the tree.The Adesmia clade is uniform for lomented pods, but the Dal-bergia and Pterocarpus clades are variable, with a minimum
of three separate origins of this pod type in each of theseclades. What was thought to be two separate origins of lo-mented pods in Adesmieae and Aeschynomeneae is now con-sidered at least six origins combined with at least two rever-sals, and not counting polymorphisms.
New taxonomic evidence—In contrast to the above, a fewpreviously overlooked characters are shown by analysis ofcombined molecular and nonmolecular data to be taxonomi-cally informative. Short shoots (character 2 in Appendix B)evolved only once in the clade containing Pictetia, Ormocar-pum, and Ormocarpopsis (also Peltiera). However, the supportfor this clade is moderate (Fig. 5), both in this analysis, andin those of Beyra-M. and Lavin (1999) and Lavin et al. (2000).Bilabiate calyx lobes (state 2 of character 19 in Appendix B)mark the monophyly of the clade containing Aeschynomenesect. Aeschynomene, Smithia, Kotschya, Humularia, Cyclocar-pa, Soemmeringia, Bryaspis, and Geissaspis. In contrast toshort shoots, this calyx morphology marks a very well-sup-ported clade (Fig. 5). The other nonmolecular characters witha high retention index (Table 1), however, either mark smallclades (e.g., characters 13 and 46 and the clade with Brya andCranocarpus), or have homoplasy that was underestimated be-cause of scoring polymorphic taxa (e.g., see characters 39 and40 in Appendix B).
The aeschynomenoid root nodule (Fig. 6, character 55 inAppendix B) is the most notable nonmolecular character inthat it is inferred to be a synapomorphy for the dalbergioidclade. The idea that nodule morphology could be a usefulcharacter in legume taxonomy was pioneered by Corby(1981). He described a number of shapes, named according tothe genus from which he had most observations. The aeschy-nomenoid type has as its main feature a small oblate nodule(transverse diameter greater than axial) with determinategrowth. Corby noted that aeschynomenoid nodules are oftenassociated with fine rootlets, but his otherwise excellent draw-ings omitted these ‘‘for clarity.’’ Such nodules were found pri-

514 [Vol. 88AMERICAN JOURNAL OF BOTANY
marily in the tribes Adesmieae, and Aeschynomeneae, but alsoin some members of the Abreae, Dalbergieae, Phaseoleae, andRobinieae (Corby, 1988). On his retirement, Corby kindlygave the Sprent laboratory his collection of preserved nodules.These were used, together with new material, for more detailedstructural studies. As a result, the definition of an aeschyno-menoid nodule has been adapted to include additional features.In particular, this nodule is always associated with a lateral or(in the case of stem nodules) adventitious root. The centralinfected tissue contains few or no uninfected cells. Differen-tiated infection threads are not involved in the process of in-fection, which (where studied in detail) takes place at the lat-eral root junction (Sprent, Sutherland, and Faria, 1989). Allnodules of the tribe Aeschynomeneae that have been examinedconform to this description, together with ten genera of theDalbergieae: Centrolobium, Dalbergia, Etaballia, Geoffroea,Machaerium, Platymiscium, Platypodium, Riedeliella, Tipu-ana, and Pterocarpus (two Brazilian species, P. rohrii and P.santalinoides are not known to nodulate). The evidence forAdesmia, Brya, and Cranocarpus, although slightly less de-tailed, is entirely consistent with the revised description ofaeschynomenoid nodules.
Members of the Dalbergieae that have been omitted fromthe revised clade on morphological and molecular groundswould also be omitted on grounds of nodule structure (Andiraand Hymenolobium) or absence of nodules (Vatairea and Va-taireopsis; Sprent, Sutherland, and Faria, 1989). Two generaof the dalbergioid clade that do not nodulate are Chaetocalyxand Nissolia (Faria and Lima, 1998). Both of these are lianes.Notably, a group of species in Acacia with a semiscandenthabitat cannot nodulate (Harrier et al., 1997). These acaciashave retained some of the characters associated with nodula-tion, such as some of the nod genes, and the ability to stim-ulate rhizobial attachment to roots. It was thus suggested thatthey may have lost the ability to nodulate because, living onthe forest margins, they were not nitrogen limited (Harrier,1995). It would be interesting to carry out similar tests onChaetocalyx and Nissolia as one of their principal habitats isforest margins.
It is now generally agreed that nodulation in legumes mayhave evolved more than once (Sprent, 1994; Soltis et al.,1995). One of these nodulation events involved an infectionprocess through a wound, such as where a lateral or an ad-ventitious root emerges. Compared with the more familiar roothair infection pathway (see Sprent and Sprent, 1990 for de-tails), this pathway is simpler, involving less complex recog-nition systems. Apart from some species of the mimosoid ge-nus Neptunia (James et al., 1992), this wound infection path-way is associated with only aeschynomenoid nodules. In Nep-tunia, however, nodule processes subsequent to infectioninvolve production of infection threads and development of anindeterminate nodule.
Our phylogenetic results are in agreement with molecularand biochemical evidence that nodule structure and infectionsite are largely plant determined (e.g., Gualtieri and Bisseling,2000). Given a phylogenetic lineage, nodule morphology andinfection processes are generally the same regardless of whichspecies or genus of rhizobia is involved (six genera of bacterianodulating legumes are now recognized, and they are collec-tively known as rhizobia). Another general inference is derivedfrom the observation that all species of the genus Aeschyno-mene that have stem nodules are nodulated by photosyntheticrhizobia (Molouba et al., 1999). Given that the aeschynome-
noid root nodule has an unelaborated morphology and infec-tion mode, the ancestral rhizobial form could have been pho-tosynthetic. As legumes moved into drier areas, nodules de-veloped on roots and lost photosynthetic ability (Sprent, 1994).
A phylogenetic classification—The dalbergioid legumes aresimilar to a group of Papilionoideae that includes also Amor-pheae and Dipterygieae. They share a distinctive combinationof a base chromosome number of x 5 10 (Goldblatt, 1981),wood with uniseriate stored rays, vegetative growth with glan-dular punctae, flowers with fused keel petals or staminal fila-ments, and seeds that do not accumulate nonprotein aminoacids (derived from Beyra-M. and Lavin, 1999). The dalber-gioids differ and are apomorphically defined (sensu de Queirozand Gauthier, 1994) as having glandular-based trichomes onvegetative or floral organs, a well-developed abaxial calyxlobe, and the ‘‘aeschynomenoid’’ root nodule. All of thesetraits have been secondarily transformed in some constituentsof the dalbergioid clade (see characters 11, 19, and 55 in Ap-pendix B; also Table 1).
The dalbergioid clade is distinguished more by molecularthan nonmolecular data. It is another legume example of acryptic clade, like ‘‘Neo-Astragalus’’ (Wojciechowski et al.,1993) and the ‘‘temperate herbaceous clade’’ (Sanderson andWojciechowski, 1996). Regardless, it is informally recognizedhere as a distinctive taxonomic group. Furthermore, the threemajor constituent subclades are informally recognized and Ap-pendix C enumerates the 44 current dalbergioid genera ac-cordingly. The three subclades of dalbergioids are:
The Adesmia clade—This includes the genera Adesmia (oftribe Adesmieae; Polhill, 1981f) and Poiretia, Amicia, Zornia,Chaetocalyx, and Nissolia of the tribe Aeschynomeneae. Thisclade is apomorphically defined as having an herbaceousgrowth habit (modified in some descendants—character 1),leaves with few opposite leaflets (evolved in parallel in Ar-achis and close relatives—character 8), and pedicels confluentwith the calyx (modified only in a few species of Nissolia—character 17). A node-based definition (sensu de Queiroz andGauthier, 1994) includes all descendants from the commonancestor of Adesmia and Amicia.
The Dalbergia clade—This includes Dalbergia and Ma-chaerium (of tribe Dalbergieae; de Candolle, 1825; Polhill,1981d), and the following genera of Aeschynomeneae (sensuRudd, 1981a): Aeschynomene (all infrageneric taxa), Soem-meringia, Cyclocarpa, Kotschya, Smithia, Humularia, Bryas-pis, Geissaspis, Weberbauerella, Diphysa, Pictetia, Ormocar-pum, Ormocarpopsis, and Peltiera. This clade is apomorphi-cally defined as having diadelphous staminal filaments split-ting readily or tardily into two flanges, usually in a 5 1 5arrangement (polymorphic with a 9 1 1 diadelphous conditionin many species and occasionally monodelphous in Machaer-ium—character 26), and a persistent staminal flange that insome cases reflexes upward above the developing fruit (char-acter 28). A node-based definition includes all descendantsfrom the common ancestor of Dalbergia and Cyclocarpa.
The Pterocarpus clade—This includes Pterocarpus, Tipu-ana, Platypodium, Reideliella, Centrolobium, Grazieloden-dron, Paramachaerium, Ramorinoa, Inocarpus, Etaballia,Platymiscium, Cascaronia, Fissicalyx, Geoffroea from Dal-bergieae; Brya and Cranocarpus from Desmodieae; and Fie-

March 2001] 515LAVIN ET AL.—DALBERGIOID LEGUMES
brigiella, Chapmannia, Stylosanthes, Arachis, and Discolob-ium from Aeschynomeneae. This clade is apomorphically de-fined as having commonly caducous bracteoles (character 18)and seedlings producing a simplified eophyll (secondarilytransformed in Arachis and close relatives—character 45). Anode-based definition includes all descendants from the com-mon ancestor of Pterocarpus and Riedeliella.
Although data from matK/trnK, trnL, and ITS/5.8S were notcombined in a single analysis, results from individual analysesshowed significant consensus combined with no significantconflict. The combined matK/trnK and nonmolecular analysisyielded very robust results to support the conclusions outlinedabove. This study demonstrates that matK/trnK sequences pro-vide excellent resolution at the broadest phylogenetic levelsdealt with in this study. This same locus, along with ITS/5.8S,gives excellent resolution to within and among closely relatedgenera. In contrast, trnL provides the least resolution. Ulti-mately, this study provides a framework for future studies thatdeal taxonomically with individual dalbergioid genera. Thereis now sufficient data from which to guide the choice of po-tential sister groups or outgroups in such studies.
LITERATURE CITED
ANTONIO, R., AND M. SOUSA. 1991. Contribuciones al conocimiento delgenero Diphysa (Leguminosae) para la flora Mesoamericana. Anales delInstituto de Biologıa, Universidad Nacional Autonoma de Mexico, SerieBotanica 62: 115–120.
BAILEY, C. D., J. J. DOYLE, T. KAJITA, T. NEMOTO, AND H. OHASHI. 1997.The chloroplast rpl2 intron and ORF184 as phylogenetic markers in thelegume tribe Desmodieae. Systematic Botany 22: 133–138.
BARETTA-KUIPERS, T. 1981. Wood anatomy of Leguminosae: its relevanceto taxonomy. In R. M. Polhill and P. H. Raven [eds.], Advances in le-gume systematics, part 2, 677–705. Royal Botanic Gardens, Kew, Rich-mond, Surrey, UK.
BARNEBY, R. C. 1977. Daleae imagines. Memoirs of the New York BotanicalGarden 27: 1–891.
BATEMAN, R. M., AND N. J. SIMPSON. 1998. Comparing phylogenetic signalsfrom reproductive and vegetative organs. In S. J. Owens and P. J. Rudall[eds.], Reproductive biology in systematics, conservation and economicbotany, 231–253. Royal Botanic Gardens, Kew, Richmond, Surrey, UK.
BENTHAM, G. 1860. A synopsis of the Dalbergieae, a tribe of the Legumi-nosae. Journal of the Proceedings of The Linnean SocietyIV(Supplement): 1–134.
BEYRA-M., A., AND M. LAVIN. 1999. Monograph of Pictetia (Legumino-sae—Papilionoideae) and review of the Aeschynomeneae. SystematicBotany Monographs 56: 1–93.
BININDA-EMONDS, O. R. P., H. N. BRYANT, AND A. P. RUSSELL. 1998. Su-praspecific taxa as terminals in cladistic analysis: implicit assumptionsof monophyly and a comparison of methods. Biological Journal of theLinnean Society 64: 101–133.
BRIZICKY, G. K. 1960. A new species of Paramachaerium from Panama.Tropical Woods 112: 58–64.
BURKART, A. 1949. Contribucion al estudio del genero Adesmia (Legumi-nosae). Lilloa 15: 1–17.
———. 1952. Las Leguminosas Argentinas, 2nd ed. ACME Agency, Buen-uos Aires, Argentina.
———. 1954. Contribucion al estudio del genero Adesmia (Leguminosae),II. Darwiniana 10: 465–546.
———. 1957. El genero Pachecoa (Leguminosae). Darwiniana 11: 261–268.
———. 1960. Contribucion al estudio del genero Adesmia (Leguminosae),III. Darwiniana 12: 81–136.
———. 1962. Contribucion al estudio del genero Adesmia (Leguminosae),IV. Darwiniana 12: 309–364.
———. 1964. Contribucion al estudio del genero Adesmia (Leguminosae),V. Darwiniana 13: 9–66.
———. 1966. Contribucion al estudio del genero Adesmia (Leguminosae),VI. Darwiniana 14: 195–248.
———. 1967. Sinopsis del genero sudamericano de Leguminosas AdesmiaDC. Darwiniana 14: 463–568.
———, AND O. VILCHEZ. 1971. Valoracion e ilustracion del genero Fiebri-giella Harms (Leguminosae-Hedysareae). Darwiniana 16: 659–662.
CHAPPILL, J. 1995. Cladistic analysis of the Leguminosae. In M. D. Crispand J. J. Doyle [eds.], Advances in legume systematics, part 7, phylog-eny, 1–9. Royal Botanic Gardens, Kew, Richmond, Surrey, UK.
CORBY, H. D. L. 1981. The systematic value of leguminous root nodules. InR. M. Polhill and P. H. Raven [eds.], Advances in legume systematics,part 2, 657–669. Royal Botanic Gardens, Kew, Richmond, Surrey, UK.
———. 1988. Types of rhizobial nodule and their distribution among theLeguminosae. Kirkia 13: 53–123.
COZZO, D. 1949. Estudio anatomico sobre la posicion sistematica de algunosgeneros Argentinos de Leguminosas Papilionoideas. Lilloa 16: 97–124.
———. 1950. Anatomia del leno secundario de las Leguminosas Papilion-oideas Argentinas silvestres y cultivadas. Revista del Instituto Nacionalde Investigacion de las Ciencias Naturales ‘‘Bernadino Rivadavia’’. Cien-cias Botanicas I(7): 223–361.
CUMBIE, B. G. 1960. Anatomical studies in the Leguminosae. TropicalWoods 113: 1–47.
CUNNINGHAM, C. W. 1997. Is congruence between data partitions a reliablepredictor of phylogenetic accuracy? Empirically testing an interactiveprocedure for choosing among phylogenetic methods. Systematic Biology46: 464–478.
DE CANDOLLE, A. P. 1825. Memoires sur la famille de Legumineuses. Paris,France.
DE CARVALHO, A. M. 1997. A synopsis of the genus Dalbergia (Fabaceae:Dalbergieae) in Brazil. Brittonia 49: 87–109.
DE N. CARMO-BASTOS, M. 1987. Contribuicao ao estudo sistematico de al-gumas especies do genero Machaerium Persoon (Leguminosae—Papi-lionoideae) ocorrentes na Amazonia Brasileira. Boletim do Museu Par-aense Emılio Goeldi, Serie Botanica. 3: 183–279.
DELGADO-SALINAS, A., T. TURLEY, A. RICHMAN, AND M. LAVIN. 1999. Phy-logenetic analysis of the cultivated and wild species of Phaseolus (Fa-baceae). Systematic Botany 23: 438–460.
DE QUEIROZ, K., AND J. GAUTHIER. 1994. Toward a phylogenetic system ofbiological nomenclature. Trends in Ecology and Evolution 9: 27–31.
DETIENNE, P., AND P. JACQUET. 1983. Atlas d’identification des bois del’Amazonie et des regions voisines. Centre Technique Forestier Tropical,Nogent-sur-Marne, France.
DOYLE, J. J., J. L. DOYLE, J. A. BALLENGER, E. E. DICKSON, T. KAJITA, AND
H. OHASHI. 1997. A phylogeny of the chloroplast gene rbcL in theLeguminosae: Taxonomic correlations and insights into the evolution ofnodulation. American Journal of Botany 84: 541–554.
EVANS, S. V., L. E. FELLOWS, AND E. A. BELL. 1985. Distribution and sys-tematic significance of basic nonprotein amino acids and amines in theTephrosieae. Biochemical Systematics and Ecology 13: 271–302.
FARIA, S. M. DE, AND H. C. DE LIMA. 1998. Additional studies of the nod-ulation status of legume species in Brazil. Plant and Soil 200: 185–192.
———, H. C. DE LIMA, A. M. CARVALHO, V. F. CONCALVES, AND J. I.SPRENT. 1994. Occurrence of nodulation in legume species from Bahia,Minas Gerais, and Espirito Santo states of Brazil. In J. I. Sprent and D.McKey [eds.], Advances in legume systematics, part 5, the nitrogen fac-tor, 17–23. Royal Botanic Gardens, Kew, Richmond, Surrey, UK.
FELSENSTEIN, J. 1985. Confidence limits on phylogenies: an approach usingthe bootstrap. Evolution 38: 783–791.
FERGUSON, I. K., AND J. J. SKVARLA. 1981. The pollen morphology of thesubfamily Papilionoideae (Leguminosae). In R. M. Polhill and P. H. Ra-ven [eds.], Advances in legume systematics, part 2, 859–896. Royal Bo-tanic Gardens, Kew, Richmond, Surrey, UK.
FERNANDES, A. 1996. O Taxon Aeschynomene no Brasil. Fortaleza: EdicoesUFC.
FERREYRA, R. 1951. Una nueva Leguminosae del Peru. Publicaciones delMuseo de Historia Natural ‘‘Javier Prado,’’ Serie B, Botanica 3: 1–5.
GASSON, P. E. 1994. Wood anatomy of the tribe Sophoreae and related Cae-salpinioideae and Papilionoideae. In I. K. Ferguson and S. C. Tucker[eds.], Advances in legume systematics, part 6, 165–203. Royal BotanicGardens, Kew, Richmond, Surrey, UK.
———. 1999. Wood anatomy of the tribe Dipterygeae with comments onrelated Papilionoid and Caesalpinioid Leguminosae. IAWA Journal 20:361–375.
GEESINK, R. 1981. Tephrosieae. In R. M. Polhill and P. H. Raven [eds.],

516 [Vol. 88AMERICAN JOURNAL OF BOTANY
Advances in legume systematics, part 1, 245–260. Royal Botanic Gar-dens, Kew, Richmond, Surrey, UK.
———. 1984. Scala Millettiearum, a survey of the genera of the tribe Mil-lettieae (Legum.—Pap.) with methodological considerations. E. J. Brill/Leiden University Press, Leiden, The Netherlands.
GILLETT, J. B. 1966. The species of Ormocarpum Beauv. and ArthrocarpumBalf. f. (Leguminosae) in South-Western Asia and Africa (excludingMadagascar). Kew Bulletin 20: 323–355.
———, R. M. POLHILL, AND B. VERDCOURT. 1971. Leguminosae. In E.Milne-Redhead and R. M. Polhill [eds.], Flora of Tropical East Africa.Crown Agents, London, UK.
GLEDHILL, D. 1968. The Geissaspis, Bryaspis, Humularia complex. Boletimda Sociedade Broteriana, serie 2: 305–319.
GOLDBLATT, P. 1981. Cytology and the phylogeny of the Leguminosae. InR. M. Polhill and P. H. Raven [eds.], Advances in legumes systematics,part 2, 427–463. Royal Botanic Gardens, Kew, Richmond, Surrey, UK.
GREINWALD, R., P. BACHMANN, G. LEWIS, L. WITTE, AND F.-C. CZYGAN.1995. Alkaloids of the genus Poecilanthe (Leguminosae: Papiliono-ideae). Biochemical Systematics and Ecology 23: 547–553.
GUALTIERI, G., AND T. BISSELING. 2000. The evolution of nodulation. PlantMolecular Biology 42: 181–194.
GUINET, PH., AND I. K. FERGUSON. 1989. Structure, Evolution, and Biologyof Pollen in Leguminosae. In C. H. Stirton and J. L. Zarucchi [eds.],Advances in legume biology. Monographs in Systematic Botany from theMissouri Botanical Garden 29: 77–103.
GUNN, C. R., E. M. NORMAN, AND J. S. LASSETTER. 1980. Chapmanniafloridana Torrey & Gray (Fabaceae). Brittonia 32: 178–185.
HARLEY, R. M. 1978. A study of Cranocarpus in Brazil. Bradea, Bolitim doHerbarium Bradeanum 2: 265–272.
HARRIER, L. A. 1995. Non-nodulating African species of Acacia. Ph.D. dis-sertation, University of Dundee, Dundee, UK.
———, P. W. WHITTY, J. M. SUTHERLAND, AND J. I. SPRENT. 1997. Pheneticinvestigation of non-nodulating African species of Acacia (Leguminosae)using morphological and molecular markers. Plant Systematics and Evo-lution 205, 27–51.
HEPPER, F. N. 1958. Papilionaceae. In R. W. J. Keay [ed.], Flora of WestTropical Africa, 2nd ed. Crown Agents, London, UK.
HERENDEEN, P. S. 1995. Phylogenetic relationships of the tribe Swartzieae.In M. Crisp and J. J. Doyle [eds.], Advances in legume systematics, part7, phylogeny, 123–132. Royal Botanic Gardens, Kew, Richmond, Surrey,UK.
HILLIS, D. 1998. Taxonomic sampling, phylogenetic accuracy, and investi-gator bias. Systematic Biology 47: 3–8.
HU, J.-M., M. LAVIN, M. WOJCIECHOWSKI, AND M. J. SANDERSON. 2000.Phylogenetic systematics of the tribe Millettieae (Leguminosae) based onchloroplast trnK/matK sequences, and its implications for evolutionarypatterns in Papilionoideae. American Journal of Botany 87: 418–430.
HUELSENBECK, J. P., J. J. BULL, AND C. W. CUNNINGHAM. 1996. Combiningdata in phylogenetic analysis. Trends in Ecology and Evolution 11: 152–158.
HUTCHINSON, J. 1964. The genera of flowering plants, vol. 1. Oxford Uni-versity Press, Oxford, UK.
IRELAND, H., AND R. T. PENNINGTON. 1999. A revision of Geoffroea (Le-guminosae—Papilionoideae). Edinburgh Journal of Botany 56: 329–347.
JAMES, E. K., J. I. SPRENT, J. M. SUTHERLAND, S. G. MCINROY, AND F. R.MINCHIN. 1992. The structure of nitrogen fixing root nodules on theaquatic mimosoid legume Neptunia plena. Annals of Botany 69: 173–180.
JUDD, W. S., R. W. SANDERS, AND M. J. DONOGHUE. 1994. Angiospermfamily pairs: preliminary phylogenetic analyses. Harvard Papers in Bot-any 5: 1–51.
KASS, E., AND M. WINK. 1995. Molecular phylogeny of the Papilionoideae(Family Leguminosae): rbcL gene sequences versus chemical taxonomy.Botanica Acta 108: 149–162.
———, AND ———. 1997. Phylogenetic relationships in the Papilionoideae(Family Leguminosae) based on nucleotide sequences of cpDNA (rbcL)and ncDNA (ITS 1 and 2). Molecular Phylogenetics and Evolution 8:65–88.
KLITGAARD, B. B. 1999a. Floral ontogeny in tribe Dalbergieae (Legumino-sae: Papilionoideae): Dalbergia brasiliensis, Machaerium villosum senslat., Platymiscium floribundum, and Pterocarpus rotundifolius. Plant Sys-tematics and Evolution 219: 1–25.
———. 1999b. New species and nomenclatural changes in Neotropical Pla-
tymiscium (Leguminosae: Papilionoideae: Dalbergieae). Kew Bulletin 54:967–973.
KRAPOVICKAS, A., AND W. C. GREGORY. 1994. Taxonomıa del genero Ar-achis (Leguminosae). Bonplandia 8: 1–186.
LABAT J.-N., AND D. J. DU PUY. 1996. Two species of Ormocarpopsis R.Viguier and a new combination in Ormocarpum P. Beauvois (Legumi-nosae—Papilionoideae) from Madagascar. Novon 6: 54–58.
———, AND ———. 1997. A revision of Peltiera, a new, poorly knownand probably extinct genus of Leguminosae (Papilionoideae—Aeschy-nomeneae) from Madagascar. Adansonia, serie 3, 19: 85–91.
LAVIN, M. 1987. A cladistic analysis of the tribe Robinieae (Papilionoideae:Leguminosae). In C. H. Stirton [ed.], Advances in legumes systematics,part 3, 31–64. Royal Botanic Gardens, Kew, Richmond, Surrey, UK.
———. 1993. Biogeography and systematics of Poitea (Leguminosae). Sys-tematic Botany Monographs 37: 1–87.
———, E. ESHBAUGH, J.-M. HU, S. MATHEWS, AND R. A. SHARROCK. 1998.Monophyletic subgroups of the tribe Millettieae (Leguminosae) as re-vealed by phytochrome nucleotide sequence data. American Journal ofBotany 85: 412–433.
———, AND M. SOUSA. 1995. Phylogenetic systematics and biogeographyof the tribe Robinieae (Leguminosae). Systematic Botany Monographs45: 1–165.
———, M. THULIN, J.-N. LABAT, AND T. PENNINGTON. 2000. Africa, theodd man out: molecular biogeography of dalbergioid legumes suggestsotherwise. Systematic Botany 24: in press.
LEON, H., AND H. ALAIN. 1951. Leguminosas, Flore de Cuba, vol. 2, Con-tribuciones Ocasionales del Museo de la Historia Natural del Colegiode La Salle 10: 224–367.
LEONARD, J. 1954. Aeschynomene. Bulletin du Jardin Botanique de l’etatBruxelles 24: 63–84.
LEWIS, G. 1988. Four little-known species of Leguminosae from Cuba.Willdenowia 18: 223–229.
LIMA, H. C. DE. 1982a. Consideracoes taxonomicas sobre o genero Hymeno-lobium Bentham (Leguminosae—Faboideae). Acta Amazonica 12: 41–48.
———. 1982b. Revisao taxonomica do genero Vatairea Aublet (Legumi-nosae—Faboideae). Arquivos do Jardim Botanico do Rio de JaneiroXXVI: 173–214.
———. 1983. Novos taxa de Leguminosae Papilionoideae (tribo Dalber-gieae) do Brasil. Bradea, Bolitim do Herbarium Bradeanum 3: 399–405.
———. 1985. Centrolobium Martius ex Bentham (Leguminosae—Papilion-oideae) estudo taxonomico das especies Brasileiras extra-Amazonicas.Arquivos do Jardim Botanico do Rio de Janeiro XXVII: 177–191.
———. 1990. Tribo Dalbergieae (Leguminosae Papilionoideae)—morfologiado frutos, sementes e plantulas e sua aplicacao na sistematica. Arquivosdo Jardim Botanico do Rio de Janeiro 30: 1–42.
———, AND A. M. STUDART DA FANSECA VAZ. 1984. Revisao taxonomicado genero Riedeliella Harms (Leguminosae—Faboideae). Rodriguesia,Rio De Janeiro 36: 9–16.
MILES, A. 1978. Photomicrographs of world woods. Her Majesty’s StationeryOffice, London, UK.
MADDISON, W. P., AND D. R. MADDISON. 1999. MacClade: analysis of phy-logeny and character evolution. Version 3.08a. Sinauer, Sunderland, Mas-sachusetts, USA.
MOCTEZUMA, E., AND L. J. FELDMAN. 1998. Growth rates and auxin effectsin graviresponding gynophores of the peanut, Arachis hypogaea (Faba-ceae). American Journal of Botany 85: 1369–1376.
MOHLENBROCK, R. H. 1957. A revision of the genus Stylosanthes. Annalsof the Missouri Botanical Garden 44: 299–355.
———. 1960. Recent studies in the Leguminosae genus Stylosanthes. Rho-dora 62: 340–346.
———. 1961. A monograph of the Leguminous genus Zornia. Webbia 16:1–141.
———. 1962. Additional collections of the Leguminous genus Zornia. Web-bia 16: 649–655.
———. 1963. Further considerations in Stylosanthes (Leguminosae). Rho-dora 65: 245–258.
MOLOUBA, F., J. LORQUIN, A. WILLEMS, B. HOSTE, E. GIRAUD, B. DREYFUS,M. GILLIS, P. DE LAJUDIE, C. MASSON-BOIVIN. 1999. Photosyntheticbradyrhizobia from Aeschynomene spp. are specific to stem nodulatingspecies and form a separate 16S ribosomal DNA restriction fragmentlength polymorphism group. Applied and Environmental Microbiology65: 3084–3094.

March 2001] 517LAVIN ET AL.—DALBERGIOID LEGUMES
NIXON, K. C., AND J. I DAVIS. 1991. Polymorphic taxa, missing values andcladistic analysis. Cladistics 7: 233–241.
OHASHI, H. 1999. The genera, tribes, and subfamilies of Japanese Legumi-nosae. The Science reports of the Tohoku University, 4th series Biology40: 187–268.
———, R. M. POLHILL, AND B. G. SCHUBERT. 1981. Desmodieae. In R. M.Polhill and P. H. Raven [eds.], Advances in legume systematics, part 1,292–300. Royal Botanic Gardens, Kew, Richmond, Surrey, UK.
PENNINGTON, R. T. 1996. Molecular and morphological data provide phy-logenetic resolution at different hierarchical levels in Andira. SystematicBiology 45: 496–515.
———, G. AYMARD, AND N. CUELLO. 1997. A new species of Andira (Le-guminosae, Papilionoideae) from the Venezuelan Guyana. Novon 7: 72–74.
———, M. LAVIN, H. IRELAND, B. KLITGAARD, AND J. PRESTON. In press.Phylogenetic relationships of primitive papilionoid legumes based uponsequences of the chloroplast intron trnL. Systematic Botany.
PITTIER, H. 1922. On the species of Dalbergia of Mexico and Central Amer-ica. J. Washington Academy of Science 12: 54–64.
POLHILL, R. M. 1971. Some observations on generic limits in Dalbergieae-Lonchocarpineae Benth. (Leguminosae). Kew Bulletin 25: 259–273.
———. 1981a. Papilionoideae. In R. M. Polhill and P. H. Raven [eds.],Advances in legume systematics, part 1, 191–208. Royal Botanic Gar-dens, Kew, Richmond, Surrey, UK.
———. 1981b. Sophoreae. In R. M. Polhill and P. H. Raven [eds.], Advancesin legume systematics, part 1, 213–230. Royal Botanic Gardens, Kew,Richmond, Surrey, UK.
———. 1981c. Dipterygeae. In R. M. Polhill and P. H. Raven [eds.], Ad-vances in legume systematics, part 1, 231–232. Royal Botanic Gardens,Kew, Richmond, Surrey, UK.
———. 1981d. Dalbergieae. In R. M. Polhill and P. H. Raven [eds.], Ad-vances in legume systematics, part 1, 233–242. Royal Botanic Gardens,Kew, Richmond, Surrey, UK.
———. 1981e. Amorpheae. In R. M. Polhill and P. H. Raven [eds.], Ad-vances in legume systematics, part 1, 244–246. Royal Botanic Gardens,Kew, Richmond, Surrey, UK.
———. 1981f. Adesmieae. In R. M. Polhill and P. H. Raven [eds.], Advancesin legume systematics, part 1, 355–356. Royal Botanic Gardens, Kew,Richmond, Surrey, UK.
———. 1994. Classification of the Leguminosae. In F. A. Bisby, J. Buck-ingham, and J. B. Harborne [eds.], Phytochemical dictionary of the Le-guminosae, vol. 1, XXXV–XLVIII. Chapman and Hall, London, UK.
PRAIN, D. 1904. The species of Dalbergia of South-eastern Asia. Annals ofthe Royal Botanical Gardens, Calcutta 10: 1–114, 91 pl.
RAMBAUT, A. 1996. Se-Al, sequence alignment editor version 1.0, alpha 1.University of Oxford, Oxford, UK.
RECORD, S. J. 1919. Storied or tier-like structure of certain dicot wood. Bul-letin of the Torrey Botanical Club 46: 253–273.
ROIG, F. A. 1986. The wood of Adesmia horrida and its modifications byclimatic conditions. IAWA Bulletin 7: 129–135.
ROJO, J. P. 1972. Pterocarpus (Leguminosae—Papilionaceae) revised for theworld. Verlag von J. Cramer, Lehre, Germany.
RUDD, V. E. 1954. Centrolobium (Leguminosae). Validation of a specificname and a brief review of the genus. Journal of the Washington Acad-emy of Science 44: 284–288.
———. 1955. The American species of Aeschynomene. Contributions of theUnited States National Herbarium 32: 1–172.
———. 1956. A revision of the genus Nissolia. Contributions of the UnitedStates National Herbarium 32: 173–206.
———. 1958. A revision of the genus Chaetocalyx. Contributions of theUnited States National Herbarium 32: 207–243.
———. 1959. The genus Aeschynomene in Malaysia (Leguminosae—Papi-lionatae). Reinwardtia 5: 23–36.
———. 1967. Supplementary studies in Aeschynomene, II: Series Pleuro-nerviae. Phytologia 15: 114–119.
———. 1970a. Revival of Nissolia microptera (Leguminosae). Phytologia20: 324.
———. 1970b. Etaballia dubia (Leguminosae), a new combination. Phyto-logia 20: 426–428.
———. 1972a. Reduction of Balisaea to Aeschynomene (Leguminosae).Phytologia 23: 321.
———. 1972b. Supplementary studies in Chaetocalyx I. (Leguminosae). In-cluding a new species from Brazil. Phytologia 24: 295–297.
———. 1972c. A new variety of Poiretia latifolia and a brief resume of thegenus Poiretia Vent. (Leguminosae). Phytologia 23: 141–148.
———. 1973. New taxa and combinations in Machaerium (Leguminosae).III. Phytologia 25: 398–403.
———. 1974. A resume of the genus Tipuana (Leguminosae). Phytologia28: 475–478.
———. 1975a. Supplementary studies in Aeschynomene III: Series Scopar-iae in Mexico and Central America. Phytologia 31: 431–434.
———. 1975b. Nissolia chiapensis, a new species of Leguminosae fromMexico. Phytologia 31: 427–430.
———. 1977. The genus Machaerium (Leguminosae) in Mexico. Boletın dela Sociedad Botanica de Mexico 37: 119–146.
———. 1981a. Aeschynomeneae. In R. M. Polhill and P. H. Raven [eds.],Advances in legume systematics, part 1, 347–354. Royal Botanic Gar-dens, Kew, Richmond, Surrey, UK.
———. 1981b. Two new species of Paramachaerium (Leguminosae) and abrief resume of the genus. Brittonia 33: 435–440.
———. 1986. A new species of Machaerium (Leguminosae) from Nicara-gua. Phytologia 60: 93–94.
———. 1987. Studies in Machaerium (Leguminosae) VII. Section II. Li-neata. Part I. Species with wingless fruit. Phytologia 64: 1–12.
———. 1996. Chaetocalyx longiloba (Fabaceae, Papilionoideae), a new spe-cies from Peru. Novon 6: 119.
SANDERSON, M. J. 1995. Objections to bootstrapping phylogenies: a critique.Systematic Biology 44: 299–320.
———, AND J. J. DOYLE. 1993. Phylogenetic relationships in North Amer-ican Astragalus (Fabaceae) based on chloroplast DNA restriction sitevariation. Systematic Botany 18: 395–408.
———, AND M. F. WOJCIECHOWSKI. 1996. Diversification rates in a tem-perate legume clade: are there ‘‘so many species of Astragalus (Faba-ceae)?’’ American Journal of Botany 83: 1488–1502.
SCHWEINGRUBER, F. H. 1990. Anatomy of European woods. Paul Haupt,Bern and Stuttgart Publishers, Stuttgard, Germany.
SOLTIS, D. E., P. S. SOLTIS, D. R. MORGAN, S. M. SWENSEN, B. C. MULLEN,J. M. DOWD, AND P. G. MARTIN. 1995. Chloroplast gene sequence datasuggest a single origin of the predisposition for symbiotic nitrogen fix-ation in angiosperms. Proceedings of the National Academy of Sciences,USA 92: 2647–2651.
SOUSA, M., AND M. P. DE SOUSA. 1981. New World Lonchocarpinae. In R.M. Polhill and P. H. Raven [eds.], Advances in legume systematics, part1, 261–281. Royal Botanic Gardens, Kew, Richmond, Surrey, UK.
SPRENT, J. I. 1994. Evolution and diversity in the legume-rhizobium sym-biosis: chaos theory? Plant and Soil 161: 1–10.
———, AND P. SPRENT. 1990. Nitrogen fixing organisms. Chapman andHall, London, UK.
———, J. M. SUTHERLAND, AND S. M. DE FARIA. 1989. Structure and func-tion of root nodules from woody legumes. In C. H. Stirton and J. L.Zarucchi [eds.], Advances in legume biology. Monographs in SystematicBotany from the Missouri Botanical Garden 29: 559–578.
SWOFFORD, D. L. 2000. PAUP*: phylogenetic analysis using parsimony(*and other methods), version 4.0b4a. Sinauer, Sunderland, Massachu-setts, USA.
TABERLET, P., L. GIELLY, G. PAUTOU, AND J. BOUVET. 1991. Universal prim-ers for amplification of three non-coding regions of chloroplast DNA.Plant Molecular Biology 17: 1105–1109.
THULIN, M. 1990. Two new species of Ormocarpum (Leguminosae) fromSomalia. Nordic Journal of Botany 10: 433–436.
———. 2000. Chapmannia (Leguminosae—Stylosanthinae) extended. Nor-dic Journal of Botany 19: 597–607.
TUCKER, S. C., AND A. W. DOUGLAS. 1994. Ontogenetic evidence and phy-logenetic relationships among basal taxa of legumes. In I. K. Fergusonand S. Tucker [eds.], Advances in legume systematics, part 6, structuralbotany, 11–32. Royal Botanic Gardens, Kew, Richmond, Surrey, UK.
ULIBARRI, E. A. 1978. Novedades en Adesmia (Leguminosae). Hickenia 1:121–124.
———. 1980. Notas sobre Adesmia I (Leguminosae—Papilionoideae). Dar-winiana 22: 493–498.
———. 1982a. Notas sobre Adesmia DC. II (Leguminosae—Papilionoideae).Darwiniana 24: 267–281.
———. 1982b. Nueva especie de Adesmia DC. (Leguminosae—Papiliono-ideae) de Chile. Hickenia 1: 277–290.
———. 1984. Notas sobre Adesmia DC. III (Leguminosae—Papilionoideae).Darwiniana 25: 355–360.

518 [Vol. 88AMERICAN JOURNAL OF BOTANY
———. 1987. Las especies de Adesmia de la serie Microphyllae (Legumi-nosae—Papilionoideae). Darwiniana 27: 315–388.
———. 1990. Notas sobre Adesmia IV. (Leguminosae—Papilionoideae).Darwiniana 30: 291–292.
VERDCOURT, B. 1971. Aeschynomene. In J. B. Gillett, R. M. Polhill, and B.Verdcourt [eds.], Flora of Tropical East Africa, Leguminosae, Papilion-oideae, 364–406. Royal Botanic Gardens, Kew, Richmond, Surrey, UK.
———. 1974. Summary of the Leguminosae-Papilionoideae-Hedysareae(sensu lato) of Flora Zambesiaca. Kirkia 9: 359–556.
WEINS, J. J. 1995. Polymorphic characters in phylogenetic systematics. Sys-tematic Biology 44: 482–500.
———, AND M. R. SERVEDIO. 1997. Accuracy of phylogenetic analysis in-cluding and excluding polymorphic characters. Systematic Biology 46:332–345.
WHEELER, E. A., P. BAAS, AND P. E. GASSON. 1989. IAWA list of micro-scopic features for hardwood identification. IAWA Journal 10: 219–332.
WHITING, M. F., J. C. CARPENTER, Q. D. WHEELER, AND W. C. WHEELER.1997. The strepsiptera problem: phylogeny of the holometabolous insectorders inferred from 18S and 28S ribosomal DNA sequences and mor-phology. Systematic Biology 46: 1–68.
WOJCIECHOWSKI, M. F., M. J. SANDERSON, B. G. BALDWIN, AND M. J. DON-OGHUE. 1993. Monophyly of anueploid Astragalus (Fabaceae): evidencefrom nuclear ribosomal DNA-ITS sequences. American Journal of Bot-any 80: 711–722.
———, ———, AND J.-M. HU. 1999. Evidence on the monophyly of As-tragalus (Fabaceae) and its major subgroups based on nuclear ribosomalDNA ITS and chloroplast DNA trnL intron data. Systematic Botany 24:409–437.
YAKOVLEV, G. P. 1972. A contribution to the system of the order FabalesNakai (Leguminales Jones). Botanical Journal 57: 585–594 (in Rus-sian—English translation at Royal Botanic Gardens, Kew, Richmond,Surrey, UK).

March 2001] 519LAVIN ET AL.—DALBERGIOID LEGUMESA
PP
EN
DIX
A.
Vou
cher
spec
imen
info
rmat
ion
for
the
mol
ecul
arda
ta.
Spe
cies
Loc
alit
yV
ouch
erb
Gen
Ban
kac
cess
ion
nos.
a
ITS
/5.8
Sm
atK
/trn
Ktr
nLin
tron
Aco
smiu
mpa
nam
ense
(Ben
th.)
Yak
ovle
vM
exic
o.O
axac
a.T
emas
cal
Hug
hes
1308
(FH
O)
GB
AN
-AF
1870
84G
BA
N-A
F14
2684
GB
AN
-AF
2088
91A
desm
iabo
rono
ides
Hoo
k.f.
Arg
enti
na.
Neu
quen
.L
asL
ajas
Lav
in82
52(M
ON
T)
GB
AN
-AF
2089
00A
desm
iaco
rym
bosa
Clo
sA
rgen
tina
.C
hubu
t.P
aso
deIn
dios
Lav
in82
83(M
ON
T)
GB
AN
-AF
1834
94A
desm
iagr
andi
flora
Gil
lies
exH
ook.
&A
rn.
Arg
enti
na.
Men
doza
.L
ujan
deC
uyo
Lav
in82
32b
(MO
NT
)G
BA
N-A
F18
3495
Ade
smia
lana
taH
ook.
f.A
rgen
tina
.N
euqu
en.
Las
Laj
asL
avin
8256
(MO
NT
)G
BA
N-A
F27
0863
GB
AN
-AF
2089
01A
desm
iala
nata
1A
rgen
tina
.C
hubu
t.T
ecka
Lav
in82
76(M
ON
T)
GB
AN
-AF
1870
97A
desm
ialo
toid
esH
ook.
f.A
rgen
tina
.C
hubu
t.P
aso
deIn
dios
Lav
in82
79(M
ON
T)
GB
AN
-AF
1870
98A
desm
iam
uric
ata
(Jac
q.)
DC
.A
rgen
tina
,M
endo
za,
Luj
ande
Cuy
oL
avin
8233
(MO
NT
)G
BA
N-A
F18
3493
Ade
smia
pinn
ifol
iaH
ook.
&A
rn.
Arg
enti
na.
Men
doza
.P
enit
ente
sL
avin
8235
(MO
NT
)G
BA
N-A
F18
3497
Ade
smia
retr
ofra
cta
Hoo
k.&
Arn
.A
rgen
tina
.N
euqu
en.
Cho
sM
alal
Lav
in82
49(M
ON
T)
GB
AN
-AF
1834
98A
desm
iasc
hnei
deri
Phi
l.A
rgen
tina
.C
hubu
t.P
aso
deIn
dios
Lav
in82
81(M
ON
T)
GB
AN
-AF
1834
96A
desm
iavi
llos
aH
ook.
f.A
rgen
tina
.C
hubu
t.T
ecka
Lav
in82
74(M
ON
T)
GB
AN
-AF
1834
99A
desm
iavo
lckm
anni
iP
hil.
Arg
enti
na.
Men
doza
.L
asL
enas
Lav
in82
45(M
ON
T)
GB
AN
-AF
1426
90A
desm
iavo
lckm
anni
i1
Arg
enti
na.
Neu
quen
.C
atan
Lil
Lav
in82
58(M
ON
T)
GB
AN
-U59
893
Aes
chyn
omen
eam
eric
ana
L.
Cub
a.C
amag
uey
Bey
ra-M
.55
4(M
ON
T)
GB
AN
-AF
2720
85G
BA
N-A
F27
2086
GB
AN
-AF
2720
87A
esch
ynom
ene
fasc
icul
aris
Cha
m.
&S
chle
ct.
Ven
ezue
la:
Mer
ida
Lav
in57
30(M
ON
T)
GB
AN
-AF
1890
25G
BA
N-A
F14
2695
GB
AN
-AF
2089
29
Aes
chyn
omen
ein
dica
L.
U.S
.A.
Lou
isia
na.
All
enT
hom
as&
All
enN
LU
3(L
SU
)G
BA
N-U
5989
2G
BA
N-A
F27
2083
GB
AN
-AF
2720
84G
BA
N-A
F20
8927
Aes
chyn
omen
ein
dica
1U
.S.A
.N
orth
Car
olin
a.H
yde
Co.
Car
ulli
58(L
SU
)G
BA
N-A
F06
8141
Aes
chyn
omen
epf
undi
iT
aub.
Zim
babw
e.N
atio
nal
Bot
anic
Gar
dens
Lav
ins.
n.(M
ON
T)
GB
AN
-AF
1890
26G
BA
N-A
F20
3587
GB
AN
-AF
2089
30A
esch
ynom
ene
purp
usii
Bra
ndeg
eeM
exic
o.O
axac
aL
avin
5833
(MO
NT
)G
BA
N-A
F27
0870
GB
AN
-AF
2089
28A
esch
ynom
ene
rudi
sB
enth
.U
.S.A
.F
lori
da.
Lak
eC
o.F
airb
roth
ers
etal
.82
(LS
U)
GB
AN
-AF
2708
69A
esch
ynom
ene
virg
inic
aB
ritt
on,
Ste
rn&
Pog
genb
.U
.S.A
.,L
ouis
iana
.B
eauf
ort
Car
ulli
42(L
SU
)G
BA
N-A
F20
3586
Am
icia
glan
dulo
saH
.B
.&
K.
Ecu
ador
.L
oja
R.
T.
Pen
ning
ton
654
(E)
GB
AN
-AF
1835
02G
BA
N-A
F20
3583
GB
AN
-AF
2089
02A
mic
iam
edic
agin
eaG
rise
b.A
rgen
tina
.T
ucum
an.
Am
aich
aL
avin
5773
(MO
NT
)G
BA
N-A
F18
3501
Am
icia
zygo
mer
isD
C.
Mex
ico.
Pue
bla.
Nau
zont
laD
elga
dos.
n.(M
EX
U)
GB
AN
-AF
2691
74A
mor
pha
frut
icos
aL
.U
.S.A
.W
yom
ing
Lav
in62
21(B
H)
GB
AN
-U59
890
GB
AN
-AF
2708
61G
BA
N-A
F20
8899
And
ira
gale
otti
ana
Sta
ndl.
Mex
ico.
Ver
acru
z.L
ago
Cat
emac
oL
avin
8214
(ME
XU
)G
BA
N-U
5988
9G
BA
N-A
F14
2681
GB
AN
-AF
2088
93A
popl
anes
iapa
nicu
lata
Pre
sl.
Gua
tem
ala.
El
Pro
gres
o.E
lR
auch
oH
ughe
s25
4(F
HO
)G
BA
N-A
F18
7093
GB
AN
-AF
2708
60G
BA
N-A
F20
8898
Ara
chis
bati
zoco
iK
rapo
v.&
W.
C.
Gre
g.B
oliv
ia.
San
taC
ruz.
Rio
Par
apet
ıC
IAT
2225
0(M
ON
T)
GB
AN
-AF
2035
53
Ara
chis
corr
enti
na(B
urka
rt)
Kra
pov.
Arg
enti
na.
Cor
rien
tes.
Utu
zain
goC
IAT
2224
9(M
ON
T)
GB
AN
-AF
2035
54A
rach
ishe
rman
nii
Kra
pov.
&W
.C
.G
reg
Bra
zil.
Mat
oG
ross
odo
Sul
.C
oxim
CIA
T22
227
(MO
NT
)G
BA
N-A
F20
3556
Ara
chis
hypo
gaea
L.
A.
Bha
gwat
,T.
G.
Kri
shna
,an
dR
.K
.M
itra
(unp
ubli
shed
)un
know
nG
BA
N-A
F15
6675
Ara
chis
mag
naK
rapo
v.,
W.
C.
Gre
g.&
Sim
pson
Bol
ivia
.S
anta
Cru
z.S
anIg
naci
oC
IAT
2224
8(M
ON
T)
GB
AN
-AF
2035
55G
BA
N-A
F20
8948
Ara
chis
maj
orK
rapo
v.&
W.
C.
Gre
g.B
razi
l.M
ato
Gro
sso
doS
ul.
Aqu
i-da
uana
CIA
T22
225
(MO
NT
)G
BA
N-A
F20
3552
GB
AN
-AF
2035
97G
BA
N-A
F20
8947
Ara
chis
palu
stri
sK
rapo
v.,
W.
C.
Gre
g.&
Val
lsB
razi
l.S
aoP
aulo
.M
irac
ema
CIA
T22
245
(MO
NT
)G
BA
N-A
F20
3557
Ara
chis
pint
oiK
rapo
v.&
W.
C.
Gre
g.B
razi
l.M
inas
Ger
ais.
Una
iC
IAT
2223
7(M
ON
T)
GB
AN
-AF
2035
51G
BA
N-A
F20
3596
GB
AN
-AF
2089
46A
rach
istr
isem
inat
aK
rapo
v.&
W.
C.
Gre
gB
razi
lC
IAT
2222
4(M
ON
T)
GB
AN
-AF
2042
33G
BA
N-A
F20
3599
GB
AN
-AF
2036
07A
rach
isvi
llos
aB
enth
.U
rugu
ayC
IAT
2225
4(M
ON
T)
GB
AN
-AF
2035
58B
rya
eben
us(L
.)D
C.
Cub
a.C
amag
uey.
Sab
anas
deC
rom
oB
eyra
-M.
523
(MO
NT
)G
BA
N-A
F06
8142
GB
AN
-AF
2708
76G
BA
N-A
F20
8950
Bry
ahi
rsut
aB
orhi
diC
uba.
Sie
rra
Nip
eL
avin
7110
(MO
NT
)G
BA
N-A
F20
3560

520 [Vol. 88AMERICAN JOURNAL OF BOTANYA
PP
EN
DIX
A.
Con
tinu
ed.
Spe
cies
Loc
alit
yV
ouch
erb
Gen
Ban
kac
cess
ion
nos.
a
ITS
/5.8
Sm
atK
/trn
Ktr
nLin
tron
Bry
aspi
slu
puli
na(B
enth
.)D
uvig
n.S
ierr
aL
eone
.W
ater
loo
Daw
e42
4(K
)G
BA
N-A
F20
4234
GB
AN
-AF
2720
68G
BA
N-A
F20
8932
Cas
caro
nia
astr
agal
ina
Gri
seb.
Arg
enti
naK
litg
aard
100
(K)
GB
AN
-AF
2042
35G
BA
N-A
F27
2072
GB
AN
-AF
2089
58C
entr
olob
ium
sp.
Bra
zil.
Rio
deJa
neir
o.B
otan
icG
ar-
dens
H.
C.
Lim
as.
n.(E
)G
BA
N-A
F27
0883
GB
AN
-AF
2089
66
Cha
etoc
alyx
blan
chet
iana
(Ben
th.)
Rud
dB
razi
l.B
ahia
,R
ioL
ajen
doH
atsc
hbac
h56
922
(F)
GB
AN
-AF
2720
70G
BA
N-A
F20
8905
Cha
etoc
alyx
bras
ilie
nsis
(Vog
el)
Ben
th.
Mex
ico.
Ver
acru
zV
entu
ra14
977
(MO
)G
BA
N-A
F27
0866
Cha
etoc
alyx
bras
ilie
nsis
1A
rgen
tina
.M
isio
nes.
Row
ing
Clu
bP
rado
s.n.
(MO
NT
)G
BA
N-A
F18
3505
Cha
etoc
alyx
glaz
iovi
iT
aub.
Bra
zil.
Bah
iaC
orad
inet
al.
5741
(NY
)G
BA
N-A
F18
3507
Cha
etoc
alyx
klug
iiR
udd
Bra
zil.
Acr
eD
aly
6778
(NY
)G
BA
N-A
F18
3506
Cha
etoc
alyx
lati
sili
qua
(Poi
r.)B
enth
.ex
Hem
sl.
Cos
taR
ica.
Pun
tare
nas.
Cor
cova
doK
erna
n12
2(M
O)
GB
AN
-AF
2035
66
Cha
etoc
alyx
nigr
ican
sB
urka
rtA
rgen
tina
.M
isio
nes.
Igua
zuV
anni
2955
(F)
GB
AN
-AF
1835
08G
BA
N-A
F20
3585
Cha
etoc
alyx
scan
dens
(L.)
Urb
.B
razi
l.R
orai
ma.
Pac
arai
ma
CIA
T20
536
(MO
NT
)G
BA
N-A
F18
3509
GB
AN
-AF
2708
65C
hapm
anni
aflo
rida
naT
orr.
&G
ray
U.S
.A.
Flo
rida
.L
ake
Co.
Gun
ter
121
(FL
AS
)G
BA
N-A
F20
3543
Cha
pman
nia
flori
dana
1U
.S.A
.F
lori
da:
Hig
hlan
dsC
o.L
ake
Wal
esR
idge
Judd
3162
(FL
AS
)G
BA
N-A
F20
3562
GB
AN
-AF
2036
00G
BA
N-A
F20
3605
GB
AN
-AF
2089
42
Cha
pman
nia
grac
ilis
(Bal
f.f.
)T
huli
nY
emen
.S
ocot
raM
ille
r&
Ale
xand
er14
039
(E)
GB
AN
-AF
2035
45G
BA
N-A
F20
3592
GB
AN
-AF
2089
40C
hapm
anni
agr
acil
is1
Yem
en.
Soc
otra
Mil
ler
&A
lexa
nder
1401
6(E
)G
BA
N-A
F20
3546
GB
AN
-AF
2035
93G
BA
N-A
F20
8941
Cha
pman
nia
pris
mat
ica
(Ses
se&
Moc
i-no
)T
huli
nM
exic
o.M
icho
acan
.P
alos
Mar
ıas
Nun
ez11
153
(MO
)G
BA
N-A
F20
3542
GB
AN
-AF
2035
98G
BA
N-A
F20
3606
Cha
pman
nia
regh
iden
sis
Thu
lin
&M
c-K
ean
Yem
en.
Soc
otra
Thu
lin
&G
ifri
8829
(UP
S)
GB
AN
-AF
2042
32
Cha
pman
nia
regh
iden
sis
Yem
en.
Soc
otra
Thu
lin
&G
ifri
8807
(UP
S)
GB
AN
-AF
2606
44C
hapm
anni
ase
rice
aT
huli
n&
McK
ean
Yem
en.
Soc
otra
Mil
ler
&A
lexa
nder
1424
1(E
)G
BA
N-A
F20
3548
GB
AN
-AF
2035
91G
BA
N-A
F20
8943
Cha
pman
nia
som
alen
sis
(Hil
lcoa
t&
Gil
lett
)T
huli
nS
omal
iaK
ucha
r15
450
(UP
S)
GB
AN
-AF
2035
44
Cha
pman
nia
tini
rean
aT
huli
nY
emen
.S
ocot
raT
huli
n&
Gif
ri88
29(U
PS
)G
BA
N-A
F20
3547
Cra
noca
rpus
mar
tii
Ben
th.
Bra
zil.
Bah
ia.
Itac
are
Kal
lunk
iet
al.
540
(MO
)G
BA
N-A
F18
9060
GB
AN
-AF
2708
75G
BA
N-A
F20
8951
Cyc
loca
rpa
stel
lari
sB
aker
Tha
ilan
dM
axw
ell
89-1
487
(E)
GB
AN
-AF
2720
67D
albe
rgia
cong
esti
flora
Pit
tier
El
Sal
vado
r.S
anta
Ana
.M
etap
anH
ughe
s12
53(F
HO
)G
BA
N-A
F06
8140
GB
AN
-AF
1426
96G
BA
N-A
F20
8924
Dal
berg
iafo
liol
osa
Ben
th.
Bol
ivia
.Y
unga
sR
.T
.P
enni
ngto
n85
9(E
)G
BA
N-A
F18
9022
GB
AN
-AF
2035
81D
albe
rgia
mel
anox
ylon
Gui
llem
in&
Per
rott
etA
fric
aP
olhi
ll53
09(K
)G
BA
N-A
F20
8921
Dal
berg
iapa
chyc
arpa
(De
Wil
d.&
T.D
uran
d)D
eW
ild.
Afr
ica
Har
ris
1924
GB
AN
-AF
2089
19
Dal
berg
iasi
ssoo
Rox
b.In
dia.
Him
acha
lP
rade
shK
irkp
atri
ck17
59(E
)G
BA
N-A
F18
9023
GB
AN
-AF
2035
82D
albe
rgia
siss
oo1
Pak
ista
n.B
aluc
hist
anG
hafo
or&
Goo
dman
4435
(E)
GB
AN
-AF
1890
24D
albe
rgia
vacc
inif
olia
Vat
keA
fric
aP
olhi
ll53
08(K
)G
BA
N-A
F20
8920
Dal
berg
iasp
.E
lS
alva
dor.
Son
sona
teH
ughe
s12
37(F
HO
)G
BA
N-A
F20
3580
GB
AN
-AF
2035
80G
BA
N-A
F20
8923
Dal
berg
iasp
.1
Ecu
ador
.Z
amor
aR
.T
.P
enni
ngto
n66
8(E
)G
BA
N-A
F20
8922
Dip
hysa
amer
ican
a(M
ille
r)M
.S
ousa
Mex
ico.
Pue
bla.
San
tiag
oN
opal
aL
avin
5801
(MO
NT
)G
BA
N-A
F06
8160
GB
AN
-AF
2035
74D
iphy
saam
eric
ana
1M
exic
o.C
hiap
as.
San
Fer
nand
oM
acqu
een
309
(MO
NT
)G
BA
N-A
F06
8166
Dip
hysa
amer
ican
a2
Mex
ico.
Pue
bla.
San
tiag
oN
opal
aL
avin
5801
a(M
ON
T)
GB
AN
-U59
891
Dip
hysa
flori
bund
aB
enth
.&
Oer
st.
Mex
ico.
Oax
aca.
Put
laSo
usa
S.10
616
(MO
)G
BA
N-A
F06
8161
GB
AN
-AF
2035
75D
iphy
sahu
mil
isO
erst
.C
osta
Ric
a.P
unta
rena
s.S
anta
Ele
naH
aber
1322
(MO
)G
BA
N-A
F06
8162
Dip
hysa
mac
roph
ylla
Lun
dell
Mex
ico.
Oax
aca.
Sal
ina
Cru
zSo
usa
9115
(MO
)G
BA
N-A
F18
9029
Dip
hysa
orm
ocar
poid
es(R
udd)
M.
Sou
-sa
&R
.A
nton
ioM
exic
o.O
axac
a.S
anJo
seL
avin
5823
(ME
XU
)G
BA
N-A
F06
8167
Dip
hysa
orm
ocar
poid
es1
Mex
ico.
Oax
aca.
San
Ped
roT
otal
apan
Sayn
esV
.12
86(M
EX
U)
GB
AN
-AF
0681
68G
BA
N-A
F20
8912
Dip
hysa
orm
ocar
poid
es2
Mex
ico.
Oax
aca.
Teh
uant
epec
Tor
res
C.
997
(ME
XU
)G
BA
N-A
F06
8169
GB
AN
-AF
2036
01G
BA
N-A
F20
3608
GB
AN
-AF
2089
13
Dip
hysa
race
mos
aR
ose
Mex
ico.
Oax
aca.
El
Pue
nte
Sous
aS.
7070
(MO
)G
BA
N-A
F06
8163
Dip
hysa
race
mos
a1
Mex
ico.
Pue
bla.
El
Cor
oT
enor
io49
50(M
O)
GB
AN
-AF
1890
30

March 2001] 521LAVIN ET AL.—DALBERGIOID LEGUMESA
PP
EN
DIX
A.
Con
tinu
ed.
Spe
cies
Loc
alit
yV
ouch
erb
Gen
Ban
kac
cess
ion
nos.
a
ITS
/5.8
Sm
atK
/trn
Ktr
nLin
tron
Dip
hysa
race
mos
a2
Mex
ico.
Jali
sco.
La
Hue
rta
McV
augh
2304
3(M
O)
GB
AN
-AF
2089
11D
iphy
sase
nnoi
des
Ben
th.
Mex
ico.
Oax
aca.
Tep
osco
lula
Gar
cıa
M.
484
(MO
)G
BA
N-A
F06
8164
Dip
hysa
spin
osa
Ryd
berg
Hon
dura
s.F
ranc
isco
Mor
azan
.T
egu-
ciga
lpa
Nel
son
7754
(MO
)G
BA
N-A
F18
9031
Dip
hysa
spin
osa
1M
exic
o.C
hiap
as.
Am
aten
ango
deV
alle
Cab
rera
3024
(MO
)G
BA
N-A
F18
9032
Dip
hysa
sube
rosa
S.
Wat
son
Mex
ico.
Oax
aca.
San
taC
ruz
Mix
te-
pec
Sous
a62
64(M
O)
GB
AN
-AF
1890
33G
BA
N-A
F26
0645
Dip
hysa
sube
rosa
1M
exic
o.O
axac
a.Ju
chet
ango
Lav
in58
14(M
O)
GB
AN
-AF
1890
34D
iphy
sath
urbe
ri(A
.G
ray)
Ryd
berg
exS
tand
l.M
exic
o.S
onor
a.A
lam
osK
earn
set
al.
s.n.
(MO
NT
)G
BA
N-A
F06
8165
GB
AN
-AF
2035
76
Dip
tery
xal
ata
Vog
elB
razi
l.G
oias
R.
T.
Pen
ning
ton
494
(E)
GB
AN
-AF
1870
90G
BA
N-A
F27
2092
GB
AN
-AF
2088
96D
ipte
ryx
pana
men
sis
(Pit
tier
)R
ecor
d&
Mel
l.C
osta
Ric
a.H
ered
ia,
Sar
apiq
uiR
.T
.P
enni
ngto
n61
3(E
)G
BA
N-A
F18
3492
Dis
colo
bium
psor
alei
foli
umB
enth
.A
rgen
tina
.F
orm
osa.
For
mos
aC
rist
obal
&K
rapo
vick
as21
67(M
O)
GB
AN
-AF
1890
58G
BA
N-A
F27
0874
GB
AN
-AF
2089
64D
isco
lobi
umpu
lche
llum
Ben
th.
Bol
ivia
.S
anta
Cru
z.C
hiqu
itos
Fre
yet
al.
531
(MO
)G
BA
N-A
F18
9059
GB
AN
-AF
2708
73G
BA
N-A
F20
8963
Eta
ball
iagu
iane
nsis
Ben
th.
Guy
ana.
Kan
uku
Mts
.Ja
nson
-Jac
obs
107
(K)
GB
AN
-AF
2720
73G
BA
N-A
F27
2074
GB
AN
-AF
2089
60
Eys
enha
rdti
asp
.M
exic
o.Z
acat
ecas
.Ja
lapa
Lav
in50
52(M
ON
T)
GB
AN
-AF
1870
96F
iebr
igie
lla
grac
ilis
Har
ms
Bol
ivia
.T
arij
aE
hric
h65
(NY
)G
BA
N-A
F20
3541
Fie
brig
iell
agr
acil
is1
Ecu
ador
.L
oja
Lew
iset
al.
3823
(K)
GB
AN
-AF
2035
61G
BA
N-A
F20
3589
Fie
brig
iell
agr
acil
is2
Bol
ivia
.S
anta
Cru
z.V
illa
gran
deN
ee36
224a
(NY
)G
BA
N-A
F20
3590
GB
AN
-AF
2089
39F
issi
caly
xfe
ndle
riB
enth
.V
enez
uela
.B
oliv
arW
urda
ck34
436
(K)
GB
AN
-AF
1890
61G
BA
N-A
F27
2063
GB
AN
-AF
2089
38G
eiss
aspi
sde
scam
psii
De
Wil
d.&
T.D
uran
dM
alaw
iH
illi
ard
&B
urtt
4305
(E)
GB
AN
-AF
2720
64G
BA
N-A
F20
8931
Geo
ffro
eade
cort
ican
s(G
illi
esex
Hoo
k.&
Arn
.)B
urka
rtU
.S.A
.A
rizo
na(s
eed
sour
ce)
Lav
in75
0(M
ON
T)
GB
AN
-AF
1890
57G
BA
N-A
F27
0880
Geo
ffro
eade
cort
ican
s1
Chi
le.
Gar
dner
&K
nees
5823
(E)
GB
AN
-AF
2089
62G
eoff
roea
spin
osa
Jacq
.E
cuad
or.
Loj
aR
.T
.P
enni
ngto
n65
9(E
)G
BA
N-A
F27
0879
Gra
ziel
oden
dron
rio-
doce
nsis
H.
C.
Lim
aB
razi
l.R
iode
Jane
iro.
Bot
anic
Gar
-de
nL
ima
s.n.
(E)
GB
AN
-AF
2708
62G
BA
N-A
F20
8952
Hum
ular
iaco
rbis
ieri
(De
Wil
d.)
Du-
vign
.C
ongo
.K
atan
gaSy
moe
ns14
132
(K)
GB
AN
-AF
2720
69G
BA
N-A
F20
8936
Hym
enol
obiu
mm
esoa
mer
ican
aH
.C
.L
ima
Cos
taR
ica.
La
Sel
vaB
iolo
gica
lS
ta-
tion
R.
T.
Pen
ning
ton
614
(E)
GB
AN
-AF
1870
87G
BA
N-A
F27
2079
GB
AN
-AF
2720
80In
ocar
pus
fagi
fer
(Par
kins
on)
Fos
berg
U.K
.R
oyal
Bot
anic
aG
arde
ns,
Kew
R.
T.
Pen
ning
ton
s.n.
(E)
GB
AN
-AF
2708
78G
BA
N-A
F20
8965
Kot
schy
aae
schy
nom
enoi
des
(Bak
er)
J.D
ewit
&D
uvig
n.M
alaw
iSa
lube
ni30
60(E
)G
BA
N-A
F20
8934
Kot
schy
aoc
hrea
ta(T
aub.
)J.
Dew
it&
Duv
ign.
Fre
nch
Gui
nea
Arm
our
8400
(E)
GB
AN
-AF
2720
65G
BA
N-A
F20
8935
Mac
haer
ium
acut
ifol
ium
Vog
elB
razi
l.G
oias
R.
T.
Pen
ning
ton
487
(E)
GB
AN
-AF
2089
26M
acha
eriu
min
unda
tum
Duc
keB
razi
l.R
orai
ma
Lew
is15
98(E
)G
BA
N-A
F20
3564
Mac
haer
ium
sp.
Col
ombi
a.T
olim
a.Ib
ague
R.
T.
Pen
ning
ton
703
(E)
GB
AN
-AF
1426
92G
BA
N-A
F20
8925
Mar
ina
sp.
Mex
ico.
Oax
aca.
Cha
zum
baL
avin
5341
(MO
NT
)G
BA
N-A
F18
7095
Myr
ospe
rmum
frut
esce
nsJa
cq.
Nic
arag
ua.
Boa
co.
Ojo
deA
gua
Hug
hes
424
(FH
O)
GB
AN
-AF
1870
85G
BA
N-A
F14
2679
GB
AN
-AF
2088
92M
yros
perm
umso
usan
umA
.D
elga
do&
M.C
.Jo
hnst
.U
.S.A
.T
exas
.A
usti
n.cu
ltiv
ated
Tur
ner
s.n.
(MO
NT
)G
BA
N-A
F18
7086
Myr
ospe
rmum
sous
anum
1K
ewL
ivin
gC
olle
ctio
nA
cces
sion
1990
-901
novo
uche
rG
BA
N-A
F26
5559
Nis
soli
age
ntry
iR
udd
Mex
ico.
Son
ora.
Gua
yabo
Van
Dev
ende
r93
-189
(NY
)G
BA
N-A
F20
8906
Nis
soli
ahi
rsut
aD
C.
Mex
ico.
Mex
ico.
Ixta
pan
Roe
1904
(F)
GB
AN
-AF
2708
68G
BA
N-A
F20
8908
Nis
soli
ale
iogy
neS
andw
ith
Mex
ico.
Jali
sco.
Tap
alap
aM
agal
lane
s29
02(F
)G
BA
N-A
F20
3563
Nis
soli
ash
otti
iA
.G
ray
Mex
ico.
Son
ora
Joya
l20
94(N
Y)
GB
AN
-AF
1835
10G
BA
N-A
F27
0867
GB
AN
-AF
2089
07

522 [Vol. 88AMERICAN JOURNAL OF BOTANYA
PP
EN
DIX
A.
Con
tinu
ed.
Spe
cies
Loc
alit
yV
ouch
erb
Gen
Ban
kac
cess
ion
nos.
a
ITS
/5.8
Sm
atK
/trn
Ktr
nLin
tron
Orm
ocar
pops
isas
pera
R.
Vig
.M
adag
asca
r.M
ohob
oP
elti
er44
16(M
O)
GB
AN
-AF
0681
48O
rmoc
arpo
psis
calc
icol
aR
.V
ig.
Mad
agas
car.
Am
bong
o.A
ntsa
koam
a-ne
raC
apur
on24
240-
SF(K
)G
BA
N-A
F06
8145
GB
AN
-AF
2035
68
Orm
ocar
pops
isit
rem
oens
isD
uP
uy&
Lab
atM
adag
asca
r.F
iana
rant
soa;
Am
bato
fi-
nand
raha
naD
uPuy
2363
(K)
GB
AN
-AF
0681
49G
BA
N-A
F20
3567
GB
AN
-AF
2089
18
Orm
ocar
pops
ism
andr
aren
sis
Dum
az-l
e-G
rand
eM
adag
asca
r.T
olia
ra;
And
ohah
ela
Re-
serv
eP
hill
ipso
n29
24(K
)G
BA
N-A
F06
8147
Orm
ocar
pops
ispa
rvif
olia
Dum
az-l
e-G
rand
eM
adag
asca
r.T
olia
ra;
Tsi
ombe
Phi
llip
son
3508
(K)
GB
AN
-AF
0681
43
Orm
ocar
pops
ispa
rvif
olia
1M
adag
asca
r.T
olia
ra;
Bel
oha
Du
Puy
etal
.M
132
(K)
GB
AN
-AF
0681
44O
rmoc
arpo
psis
tule
aren
sis
Du
Puy
&L
abat
Mad
agas
car.
Tol
iara
Ker
audr
en13
69(K
)G
BA
N-A
F06
8146
Orm
ocar
pum
bern
ieri
anum
(Bai
ll.)
Du
Puy
&L
abat
Mad
agas
car.
Ant
sira
nana
Lab
atet
al.
2882
(P)
GB
AN
-AF
1890
35G
BA
N-A
F20
3570
Orm
ocar
pum
bern
ieri
anum
1M
adag
asca
r.A
ntsi
rana
naD
uP
uyet
al.
M71
6(P
)G
BA
N-A
F18
9036
Orm
ocar
pum
coer
uleu
mB
alf.
f.Y
emen
.S
ocot
raT
huli
n&
Gif
ri87
81(U
PS
)G
BA
N-A
F18
9037
GB
AN
-AF
2035
72G
BA
N-A
F20
8914
Orm
ocar
pum
dhof
aren
seH
illc
oat
&G
il-
lett
Yem
enT
huli
net
al.
9746
(UP
S)
GB
AN
-AF
1890
40G
BA
N-A
F26
0646
Orm
ocar
pum
drak
eiR
.V
ig.
Mad
agas
car.
Men
abe.
Ant
salo
vaC
apur
on24
625-
SF(P
)G
BA
N-A
F18
9038
Orm
ocar
pum
drak
ei1
Mad
agas
car.
Bek
opak
aR
akot
ozaf
y98
6(P
)G
BA
N-A
F18
9039
Orm
ocar
pum
flavu
mG
ille
ttT
anza
nia.
Irin
gaD
istr
.G
reen
way
1405
4(M
O)
GB
AN
-AF
1890
41O
rmoc
arpu
mgi
llet
tii
Thu
lin
Som
alia
Wie
land
4681
(UP
S)
GB
AN
-AF
1890
42G
BA
N-A
F26
0647
Orm
ocar
pum
gill
etti
i1
Som
alia
.H
obyo
Dis
t.W
iela
nd43
57(M
O)
GB
AN
-AF
1890
43O
rmoc
arpu
mke
nien
seG
ille
ttK
enya
.M
eru
Fad
en74
/958
(MO
)G
BA
N-A
F06
8155
GB
AN
-AF
2036
02G
BA
N-A
F20
8917
Orm
ocar
pum
kirk
iiS
.M
oore
Mal
awi.
Sou
ther
nR
egio
nC
hapm
an84
92(M
O)
GB
AN
-AF
0681
50G
BA
N-A
F20
3571
Orm
ocar
pum
kirk
ii1
Sou
thA
fric
a.K
loof
For
est
Bal
sinh
as28
42(M
O)
GB
AN
-AF
0681
51O
rmoc
arpu
mki
rkii
2M
ozam
biqu
e.M
omba
Tor
re94
58(M
O)
GB
AN
-AF
0681
52O
rmoc
arpu
mkl
aine
iT
isse
r.C
amer
oon.
Sou
thw
est.
Lak
eB
arom
biJ.
D.
Man
ning
747
(MO
)G
BA
N-A
F18
9044
GB
AN
-AF
2036
03O
rmoc
arpu
mm
egal
ophy
llum
Har
ms
Cam
eroo
n.F
ores
tP
rese
rve
Am
shof
f12
99(M
O)
GB
AN
-AF
0681
54O
rmoc
arpu
mm
uric
atum
Chi
ov.
Ken
ya.
Man
dera
Gil
bert
1309
(MO
)G
BA
N-A
F06
8156
Orm
ocar
pum
mur
icat
um1
Eth
iopi
a.H
arra
rD
eWil
de54
98(M
O)
GB
AN
-AF
0681
57G
BA
N-A
F20
3569
GB
AN
-AF
2089
16O
rmoc
arpu
mor
ient
ale
(Spr
eng.
)M
err.
Aus
tral
ia.
Que
ensl
and,
Nor
thK
en-
nedy
Roa
chs.
n.(Q
LD
)G
BA
N-A
F06
8159
Orm
ocar
pum
pube
scen
s(H
ochs
t.)
Cuf
odA
friq
ueoc
cide
ntal
eA
ubre
vill
es.
n.(P
)G
BA
N-A
F18
9045
Orm
ocar
pum
rect
angu
lare
Thu
lin
Som
alia
Thu
lin
etal
.68
91(U
PS
)G
BA
N-A
F18
9046
GB
AN
-AF
2606
48O
rmoc
arpu
msc
hlie
beni
iH
arm
sT
anza
nia.
Lin
diD
istr
.Sc
hlie
ben
5766
(P)
GB
AN
-AF
1890
47O
rmoc
arpu
mse
nnoi
des
(Wil
ld.)
DC
.T
anza
nia.
Zar
anin
jeF
ores
tC
FR
P10
43(M
O)
GB
AN
-AF
0681
53O
rmoc
arpu
mso
mal
ense
Gil
lett
Som
alia
Thu
lin
&W
arfa
5818
(UP
S)
GB
AN
-AF
1890
48G
BA
N-A
F26
0649
Orm
ocar
pum
trac
hyca
rpum
(Tau
b.)
Har
ms
Eth
iopi
a.H
arar
Am
shof
f98
87(M
O)
GB
AN
-AF
1890
49G
BA
N-A
F20
8915
Orm
ocar
pum
tric
hoca
rpum
(Tau
b.)
Bur
ttD
avy
Nat
al.
Gun
aman
ini
Pan
Step
hens
820
(MO
)G
BA
N-A
F06
8158
Orm
ocar
pum
verr
ucos
umP.
Bea
uv.
Cam
eroo
nJu
ngne
rs.
n.(U
PS
)G
BA
N-A
F18
9050
Orm
ocar
pum
yem
enen
seG
ille
ttY
emen
Thu
lin
etal
.92
67(U
PS
)G
BA
N-A
F18
9051
GB
AN
-AF
2035
73G
BA
N-A
F26
0650
Par
amac
haer
ium
scho
mbu
rgki
i(B
enth
.)D
ucke
Guy
ana.
Kan
uku
Mts
.Ja
nson
-Jac
obs
97(K
)G
BA
N-A
F20
4237
GB
AN
-AF
2720
62G
BA
N-A
F20
8959
Pic
teti
aac
ulea
ta(V
ahl)
Urb
.P
uert
oR
ico.
Cab
oR
ojo
Axe
lrod
4788
(NY
)G
BA
N-A
F06
8175
Pic
teti
aac
ulea
ta1
Pue
rto
Ric
o.G
uani
caA
xelr
od28
77(N
Y)
GB
AN
-AF
0681
74P
icte
tia
acul
eata
2S
t.Jo
hn.
Cor
alB
ayA
ceve
doet
al.
2046
(NY
)G
BA
N-A
F20
3577
Pic
teti
aac
ulea
ta3
Pue
rto
Ric
o.G
uani
caA
tha
&Z
anon
i73
0(N
Y)
GB
AN
-AF
2036
04G
BA
N-A
F26
0906
Pic
teti
aan
gust
ifol
iaG
rise
b.C
uba.
Cam
ague
y.L
oma
dela
Coc
aB
eyra
-M.
s.n.
(MO
NT
)G
BA
N-A
F06
8171
GB
AN
-AF
2035
79P
icte
tia
mar
gina
taS
auv.
Cub
a.H
oguı
n.S
ierr
aN
ipe
Lav
in71
08(M
ON
T)
GB
AN
-AF
0681
76G
BA
N-A
F20
3578
GB
AN
-AF
2089
10P
icte
tia
mar
gina
ta1
Cub
a.L
asV
illa
s-C
amag
uey
Bey
ra-M
.s.
n.(M
ON
T)
GB
AN
-AF
0681
77

March 2001] 523LAVIN ET AL.—DALBERGIOID LEGUMESA
PP
EN
DIX
A.
Con
tinu
ed.
Spe
cies
Loc
alit
yV
ouch
erb
Gen
Ban
kac
cess
ion
nos.
a
ITS
/5.8
Sm
atK
/trn
Ktr
nLin
tron
Pic
teti
am
ucro
nata
(Gri
seb.
)B
eyra
-M.
&L
avin
Cub
a.C
amag
uey.
La
Min
aB
eyra
-M.
s.n.
(MO
NT
)G
BA
N-A
F06
8172
Pic
teti
am
ucro
nata
1C
uba.
Cam
ague
y.T
agar
roB
eyra
-M.
s.n.
(MO
NT
)G
BA
N-A
F06
8173
Pic
teti
ani
pens
is(U
rb.)
Bey
ra-M
.&
Lav
inC
uba.
Sie
rra
Nip
eE
kman
1000
1(N
Y)
GB
AN
-AF
1890
52
Pic
teti
aob
cord
ata
DC
.D
omin
ican
Rep
ubli
c.C
ambi
taG
ara-
bita
Zan
oni
4048
8(N
Y)
GB
AN
-AF
0681
70
Pic
teti
asp
inos
a(A
.R
ich.
)B
eyra
-M.
&L
avin
Cub
a.S
anti
ago
deC
uba
Ekm
an84
03(N
Y)
GB
AN
-AF
2035
65
Pic
teti
asu
lcat
a(P
.B
eauv
.)B
eyra
-M.
&L
avin
Dom
inic
anR
epub
lic.
Bao
ruco
Gar
cıa
etal
.62
3(N
Y)
GB
AN
-AF
0681
78
Pla
tym
isci
umfil
ipes
Ben
th.
Bra
zil
Kli
tgaa
rd35
(K)
GB
AN
-AF
1890
53P
laty
mis
cium
stip
ular
eB
enth
.E
cuad
or.
Nap
o.Ja
tun
Sac
haR
.T
.P
enni
ngto
n64
9(E
)G
BA
N-A
F18
9054
GB
AN
-AF
2708
72P
laty
mis
cium
sp.
Col
ombi
a.A
ntio
quia
.R
ioC
laro
R.
T.
Pen
ning
ton
692
(E)
GB
AN
-AF
2708
71G
BA
N-A
F20
8955
Pla
typo
dium
eleg
ans
Vog
elC
olom
bia.
Ant
ioqu
ia.
Rio
Cla
roR
.T
.P
enni
ngto
n68
8(E
)G
BA
N-A
F27
0877
Pla
typo
dium
eleg
ans
1B
razi
l.G
oias
.C
hapa
dado
sV
eade
iros
R.
T.
Pen
ning
ton
488
(E)
GB
AN
-AF
2089
61P
oeci
lant
hepa
rvifl
ora
Ben
th.
Bra
zil.
Rio
deJa
neir
o.B
otan
icG
ar-
den
Lim
as.
n.(R
B)
GB
AN
-AF
1870
89G
BA
N-A
F14
2687
GB
AN
-AF
2088
97
Poi
reti
aan
gust
ifol
iaV
ogel
Bra
zil.
Goi
as.
Niq
uela
ndia
Fon
seca
etal
.14
19(M
O)
GB
AN
-AF
1835
03G
BA
N-A
F27
0864
GB
AN
-AF
2089
04P
oire
tia
punc
tata
(Wil
ld.)
Des
v.E
cuad
or.
Gua
yas
Mad
sen
6349
1(M
O)
GB
AN
-AF
1835
04G
BA
N-A
F27
2081
GB
AN
-AF
2720
82P
tero
carp
usac
apul
cens
isR
ose
Mex
ico.
Oax
aca.
San
tiag
oA
stat
aL
avin
5325
(MO
NT
)G
BA
N-A
F26
9175
Pte
roca
rpus
indi
cus
Wil
ld.
Phi
lipp
ines
.L
uzon
.M
t.Iz
arog
Pen
ning
ton
718
(E)
GB
AN
-AF
2089
53P
tero
carp
usin
dicu
s1
unkn
own
loca
lity
Hen
ders
ons.
n.(N
Y)
GB
AN
-AF
2691
77G
BA
N-A
F14
2691
Pte
roca
rpus
mac
roca
rpus
Kur
zP
uert
oR
ico.
Faj
ardo
.cu
ltiv
ated
Lav
in72
1(M
ON
T)
GB
AN
-AF
2691
76G
BA
N-A
F20
3588
Pte
roca
rpus
sp.
Col
ombi
a.A
ntio
quia
.R
ioC
laro
R.
T.
Pen
ning
ton
690
(E)
GB
AN
-AF
2089
54P
tero
don
pube
scen
sB
enth
.B
razi
l.G
oias
R.
T.
Pen
ning
ton
476
(E)
GB
AN
-AF
1870
91G
BA
N-A
F27
2094
GB
AN
-AF
2720
95G
BA
N-A
F20
8895
Ram
orin
oagi
rola
eS
peg.
U.K
.R
oyal
Bot
anic
Gar
dens
,K
ew.
cult
ivat
edno
vouc
her
GB
AN
-AF
2042
36G
BA
N-A
F27
0881
GB
AN
-AF
2089
57
Rie
deli
ella
grac
ilifl
ora
Har
ms
Bra
zil.
Mat
oG
ross
odo
Sul
Rat
ter
etal
.74
94(E
)G
BA
N-A
F27
2090
GB
AN
-AF
2720
91G
BA
N-A
F20
8949
Smit
hia
cili
ata
Roy
leN
epal
Stai
nton
etal
.40
48(E
)G
BA
N-A
F27
2066
GB
AN
-AF
2089
33So
emm
erin
gia
sem
perfl
oren
sM
art.
Bra
zil.
Ror
aim
a,Il
hada
Mar
aca
Lew
is16
00(E
)G
BA
N-A
F18
9027
GB
AN
-AF
2720
88G
BA
N-A
F27
2089
GB
AN
-AF
2089
37
Styl
osan
thes
angu
stif
olia
Vog
el(V
ande
rS
tapp
enet
al.,
1998
)G
BA
N-A
J230
726
Styl
osan
thes
calc
icol
aS
mal
l(V
ande
rS
tapp
enet
al.,
1998
)G
BA
N-A
J230
728
Styl
osan
thes
capi
tata
Vog
elB
razi
l.M
ato
Gro
sso.
Ron
dono
poli
sC
IAT
1693
(MO
NT
)G
BA
N-A
F20
3549
GB
AN
-AF
2035
95G
BA
N-A
F20
8945
Styl
osan
thes
ham
ata
(L.)
Tau
b.C
uba.
San
tiag
ode
Cub
aB
eyra
-M.
595
(MO
NT
)G
BA
N-A
F20
3550
GB
AN
-AF
2035
94G
BA
N-A
F20
8944
Styl
osan
thes
hum
ilis
Kun
th(V
ande
rS
tapp
enet
al.,
1998
)G
BA
N-A
J230
738
Styl
osan
thes
frut
icos
a(R
etz.
)A
lsto
n(V
ande
rS
tapp
enet
al.,
1998
)G
BA
N-A
J230
731
Styl
osan
thes
grac
ilis
H.
B.
&K
.1
(Van
der
Sta
ppen
etal
.,19
98)
GB
AN
-Y13
483
Styl
osan
thes
grac
ilis
2(V
ande
rS
tapp
enet
al.,
1998
)G
BA
N-Y
1348
8St
ylos
anth
esgr
andi
flora
M.
B.
Fer
reir
a&
S.
Cos
ta1
(Van
der
Sta
ppen
etal
.,19
98)
GB
AN
-Y13
486
Styl
osan
thes
gran
diflo
ra2
(Van
der
Sta
ppen
etal
.,19
98)
GB
AN
-Y13
487
Styl
osan
thes
guia
nens
is(A
ubl.
)S
war
tz1
(Van
der
Sta
ppen
etal
.,19
98)
CIA
T13
6G
BA
N-Y
1348
0St
ylos
anth
esgu
iane
nsis
2(V
ande
rS
tapp
enet
al.,
1998
)C
IAT
1283
GB
AN
-Y13
481
Styl
osan
thes
guia
nens
is3
(Van
der
Sta
ppen
etal
.,19
98)
Sch
ofiel
dD
NA
GB
AN
-Y13
482
Styl
osan
thes
guia
nens
is4
(Van
der
Sta
ppen
etal
.,19
98)
CP
I34
906
GB
AN
-Y13
485
Styl
osan
thes
guia
nens
is5
(Van
der
Sta
ppen
etal
.,19
98)
CP
I18
750
GB
AN
-Y13
489
Styl
osan
thes
guia
nens
is6
(Van
der
Sta
ppen
etal
.,19
98)
CIA
T12
83G
BA
N-Y
1349
0St
ylos
anth
esgu
iane
nsis
7(V
ande
rS
tapp
enet
al.,
1998
)C
IAT
184
GB
AN
-Y13
491

524 [Vol. 88AMERICAN JOURNAL OF BOTANY
AP
PE
ND
IXA
.C
onti
nued
.
Spe
cies
Loc
alit
yV
ouch
erb
Gen
Ban
kac
cess
ion
nos.
a
ITS
/5.8
Sm
atK
/trn
Ktr
nLin
tron
Styl
osan
thes
guia
nens
is8
(Van
der
Sta
ppen
etal
.,19
98)
cult
ivar
SG
220
GB
AN
-AJ2
3073
3St
ylos
anth
esha
mat
a(L
.)T
aub.
Cub
a.S
anti
ago
deC
uba
Bey
raM
.59
5(M
ON
T)
GB
AN
-AF
2035
50G
BA
N-A
F20
3594
GB
AN
-AF
2089
44St
ylos
anth
eshi
ppoc
ampo
ides
Moh
lenb
r.(V
ande
rS
tapp
enet
al.,
1998
)G
BA
N-Y
1348
4St
ylos
anth
eshu
mil
isH
.B
.&
K.
(Van
der
Sta
ppen
etal
.,19
98)
GB
AN
-AJ2
3073
8St
ylos
anth
eshu
mil
is1
(Van
der
Sta
ppen
etal
.,un
publ
ishe
dda
ta)
GB
AN
-AJ1
0132
5
Styl
osan
thes
hum
ilis
2(V
ande
rS
tapp
enet
al.,
unpu
blis
hed
data
)G
BA
N-A
J101
326
Styl
osan
thes
hum
ilis
3(V
ande
rS
tapp
enet
al.,
unpu
blis
hed
data
)G
BA
N-A
J101
327
Styl
osan
thes
hum
ilis
4(V
ande
rS
tapp
enet
al.,
unpu
blis
hed
data
)G
BA
N-A
J101
328
Styl
osan
thes
hum
ilis
5(V
ande
rS
tapp
enet
al.,
unpu
blis
hed
data
)G
BA
N-A
J101
329
Styl
osan
thes
hum
ilis
6(V
ande
rS
tapp
enet
al.,
unpu
blis
hed
data
)G
BA
N-A
J010
330
Tip
uana
tipu
(Ben
th.)
Kun
tze
Arg
enti
na.
Sal
ta.
La
Cru
zL
avin
5796
(MO
NT
)G
BA
N-A
F18
9056
GB
AN
-AF
2708
82T
ipua
nati
pu1
Spa
in.
Bar
celo
na.
Cul
tiva
ted
R.
T.
Pen
ning
ton
s.n.
(E)
GB
AN
-AF
2089
56V
atai
rea
mac
roca
rpa
(Ben
th.)
Duc
keB
razi
l.G
oias
.C
hapa
dado
sV
eade
iros
R.
T.
Pen
ning
ton
495
(E)
GB
AN
-AF
2655
60V
atai
rea
sp.
Col
ombi
a.A
ntio
quia
.R
ioC
laro
R.
T.
Pen
ning
ton
689
(E)
GB
AN
-AF
1834
91G
BA
N-A
F27
0859
Vat
aire
asp
.no
v.C
osta
Ric
a.P
unta
rena
s,P
enın
sula
deO
saR
.T
.P
enni
ngto
n58
7(E
)G
BA
N-A
F18
7088
Vat
aire
opsi
ssu
rina
men
sis
H.
C.
Lim
aG
uyan
a.Iw
okra
ma
Res
erve
Are
aR
.T
.P
enni
ngto
n38
5(E
)G
BA
N-A
F14
2680
GB
AN
-AF
2088
94W
eber
baue
rell
abr
ongn
iart
ioid
esU
lbr.
Per
u.A
requ
ipa.
Lom
asde
Mol
lend
oD
illo
n39
09(F
)G
BA
N-A
F18
9028
GB
AN
-AF
2720
75G
BA
N-A
F27
2076
GB
AN
-AF
2089
09
Web
erba
uere
lla
bron
gnia
rtio
ides
1P
eru.
Are
quip
a.L
omas
deM
ejıa
Dil
lon
3742
(F)
GB
AN
-AF
2720
71W
eber
baue
rell
abr
ongn
iart
ioid
es2
Per
u.A
requ
ipa.
Lom
asde
Mej
ıaD
illo
n48
18(F
)G
BA
N-A
F27
2077
GB
AN
-AF
2720
78Z
orni
asp
.M
exic
o.Z
acat
ecas
.F
resn
illo
Lav
in50
39(M
ON
T)
GB
AN
-AF
1835
00G
BA
N-A
F20
3584
GB
AN
-AF
2089
03Z
orni
asp
.1
(Van
der
Sta
ppen
etal
.,un
publ
ishe
dda
ta)
GB
AN
-AJ2
3074
8
Not
e:N
exus
file
sof
nonm
olec
ular
and
mol
ecul
arda
taar
elo
cate
dat
http
://g
emin
i.os
cs.
mon
tana
.edu
/;m
lavi
n/da
ta/d
albd
at.h
tm.
aT
hepr
efix
GB
AN
-ha
sbe
enad
ded
toal
lG
enB
ank
acce
ssio
nnu
mbe
rsto
link
the
onli
neve
rsio
nof
Am
eric
anJo
urna
lof
Bot
any
toG
enB
ank
but
isno
tpa
rtof
the
actu
alac
cess
ion
num
ber.
bC
IAT
acce
ssio
nsw
ere
grow
nfr
omse
ed,
and
ahe
rbar
ium
spec
imen
was
prep
ared
from
the
gree
nhou
sepl
ant
and
depo
site
dat
MO
NT.

March 2001] 525LAVIN ET AL.—DALBERGIOID LEGUMES
APPENDIX B
Nonmolecular characters and character states. All references to clades arethose derived from the combined matK/trnK and nonmolecular analysis (Fig.5). References to ancestral states were inferred with the reconstruct tree optionin PAUP (Swofford, 2000) and the trace option in MacClade (Maddison andMaddison, 1999).
Vegetative characters
1. Habit: 0) woody (trees to shrubs), 1) herbaceous (subshrubs to herbs),2) twining and herbaceous, 3) twining and woody. Predominantly herbaceousgenera sometimes include subshrubby species, whereas woody genera usuallydo not, thus explaining the coding for state number 1. A herbaceous habitarose independently in the following clades: one represented by Fiebrigiellaand Arachis, another by Chaetocalyx and Poiretia, and one by Weberbauer-ella and Kotschya. The twining herbaceous habit is restricted to the Adesmiaclade where it is known from some species of Poiretia (Rudd, 1972c) and allChaetocalyx (Rudd, 1958) and Nissolia (Rudd, 1956). A twining woody habitoccurs in polymorphic condition in the clade with Dalbergia and Machaer-ium.
2. Short shoots: 0) absent or not regularly present and then not covered bypersistent stipules, 1) regularly present and covered by distichously arrangedpersistent stipules from the axils of which are born the inflorescence (Fig. 7).The short shoot condition is restricted to the clade including all descendantsof the most recent common ancestor of Pictetia and Ormocarpopsis. Verysimilar short shoots were described for Poitea (tribe Robinieae; Lavin, 1993),which is also from the Greater Antilles.
3. Stipule modifications: 0) attached to stem at base (basifixed) and folia-ceous, 1) attached to stem in the middle and foliaceous (peltate or medifixed),2) basifixed and lignescent. Medifixed stipules are referred to as appendiculate(e.g., Rudd, 1981a) and are evolved independently in a clade including Aes-chynomene sect. Aeschynomene, Cyclocarpa, Humularia, Geissaspis, Smithia,and another including just Zornia. Lignescent stipules evolved independentlyin polymorphic condition in the liana-forming species of Machaerium, in mostspecies of Brya, and in all species of Pictetia. In Brya, the leaves of the longshoot are entirely transformed into a single spine.
4. Pseudopetiole: 0) absent, 1) present (Fig. 8). A pseudopetiole is tradi-tionally defined as a petiole with stipules attached. It is here described as apulvinus (leaf base) that is projected away from the main axis of the stem.The stipules are attached to this projected portion of the stem, and they su-perficially appear as if they are adnate to the petiole. The pseudopetioleevolved independently in a clade including just Adesmia, and another includ-ing Arachis and Stylosanthes.
5. Leaf rachis in cross section: 0) terete, 1) with a single continuous groove(canaliculate). A terete leaf rachis is recorded from Discolobium, Dalbergia,Machaerium, and Ormocarpopsis, Peltiera, Platymiscium, Centrolobium,Grazielodendron, Etaballia, Fissicalyx, Peltiera, and Pterocarpus, and inpolymorphic condition from Ormocarpum, Aeschynomene (all subgroups) andclosely related genera (Cyclocarpa, etc.). Grooved leaf rachises occur in therest of the genera, except where the leaves are uniformly sessile, as in Bryaand Inocarpus, and this trait is then scored as inapplicable. Otherwise, leafrachises vary continuously between narrowly grooved and distinctly canalic-ulate. The motivation for using this trait is that terete leaf rachises are shownto be derived (but in polymorphic condition) in two clades: that including alldescendents but Pictetia of the most recent common ancestor of Dalbergiaand Ormocarpopsis, and that including most descendants of the recent com-mon ancestor of Platymiscium and Pterocarpus.
6. Distal end of leaf rachis: 0) terminated by a leaflet, 1) not terminated bya leaflet (a mucro is often present). A leaf rachis not terminated by a leafletis found in the large clade including Aeschynomene sect. Aeschynomene, Cy-clocarpa, Humularia, Soemmeringia, Kotschya, Smithia, Geissaspis, andBryaspis. This type of leaf also has evolved independently in the outgroupsamples of Dipterygeae (Dipteryx and Pterodon), the clade including Amicia,Zornia, Adesmia, Arachis, and Poiretia, the clade including just Aeschyno-mene sect. Ochopodium, and the clade including Stylosanthes and Arachis.
7. Number of leaflets per leaf: 0) leaves unifoliolate/simple, 1) leaves tri-to 20-foliolate, 2) leaves more than 20-foliolate. State zero occurs uniformlyin Etaballia, Inocarpus, and Brya, and in polymorphic condition in Crano-carpus. State two is restricted to just the Dalbergia clade where it occursuniformly in Weberbauerella, and predominantly so (i.e., polymorphic) inMachaerium, Dalbergia, and all the sections and series of Aeschynomene(Aeschynomene, Viscidulae, Pleuronerviae, and Scopariae). This state is cap-turing ‘‘fern-like’’ leaves where the leaflets abut laterally, are narrowly ellip-
tic, and have parallel lateral margins. Simple leaves are scattered throughoutbut with most occurrences (usually in polymorphic condition) in the Ptero-carpus clade (Discolobium, Etaballia, Inocarpus, Platypodium, Byra, andCranocarpus).
8. Leaflet arrangement: 0) alternate, 1) opposite. Two large clades haveevolved opposite leaflets independently. One includes Adesmia, Chaetocalyx,Nissolia, Poiretia, Amicia, Zornia, and the other includes Fissicalyx, Fiebri-giella, Chapmannia, Stylosanthes, and Arachis. Opposite leaflets have evolvedsporadically mostly within the Pterocarpus clade (Grazielodendron, Riede-liella, Cranocarpus, Paramachaerium), and rarely in the Dalbergia clade(Smithia). The genera with uniformly simple or unifoliolate leaves (e.g., Eta-ballia, Inocarpus, Brya, and Ramorinoa) were marked inapplicable. The spe-cies of Cranocarpus with imparipinnate leaves have opposite leaflets, and thiscondition is used to represent the genus. A terminal taxon is scored for op-posite leaflets if all constituent species predominate with this condition. Aterminal taxon is scored for a polymorphic condition only if some constituentspecies have uniformly opposite leaflets and others have uniformly alternateleaflets.
9. Leaflet base: 0) symmetric, 1) asymmetric. The asymmetric state is re-stricted to the Dalbergia clade, where it has evolved independently and poly-morphically in Pictetia and Aeschynomene (all subgroups except Scoparia)and uniformly in Humularia, Bryaspis, Geissaspis, Kotschya, and Smithia. Anasymmetric base of the leaflet is correlated with an eccentric midrib and prob-ably related to a nyctinastic leaflet movement that involves a forward twistingand folding of each leaflet. This ‘‘forward-folding’’ type is very similar tothe leaflet movements in legume subfamilies Mimosoideae and Caesalpinioi-deae, as well as the papilionoid genus Sesbania, and it has been observed inspecies of Aeschynomene, Arachis, Diphysa, Dalbergia, and Machaerium.
10. Tannin deposits on the abaxial surface of dried leaflets: 0) absent, 1)present. Tanniniferous patches on dried leaflets have evolved independentlyin the clade including Arachis, Stylosanthes (polymorphic in these first two),and Chapmannia (this is the subtribe Stylosanthinae of Rudd, 1981a) and intwo genera endemic to Madagascar, Ormocarpopsis and Peltiera (Labat andDu Puy, 1996, 1997). Reddish tannin deposits usually occur in reticulate pat-terns demarcating individual epidermal cells. In Ormocarpopsis and Peltiera,they can be concentrated along the leaflet midrib.
11. Glandular-based trichomes: 0) absent, 1) present (Figs. 9, 10). This typeof trichome is a synapomorphy for the dalbergioid group, where it is foundon the stems, leaves, inflorescence, or ovary. Although synapomorphic, theglandular-based trichomes have been secondarily lost several times in each ofthe Adesmia, Dalbergia, and Pterocarpus clades. In addition to most generaof the formally recognized tribe Aeschynomeneae, glandular-based trichomesare found in Centrolobium, Grazielodendron, Ramorinoa, Etaballia, Riede-liella, Fissicalyx, Paramachaerium, Peltiera, and polymorphic in Brya, Cran-ocarpus, Dalbergia, and Machaerium.
12. Pustular glands: 0) absent, 1) present (Fig. 11). The latter condition isthought to be a derivation of the general dalbergioid trait of punctate glands(all members of the ingroup and outgroup possess punctate glands on theleaflets). There has been further development in the size and color of thecommon punctate gland such that they protrude outward from the plane ofthe leaflet, calyx, or ovary and are brownish red to blackish in color. Pustularglands are known from genera outside the dalbergioid clade (Acosmium, My-rospermum, Amorpha, Apoplanesia, Dipteryx, and Pterodon), and haveevolved in four separate instances within the dalbergioid clade (Geoffroea andCascaronia; Poiretia, Amicia, and Zornia; Weberbauerella; and Centrolob-ium).
13. Stipitate glands: 0) absent, 1) present from non-glochidiate trichomes,2) present from microscopically glochidiate trichomes (Fig. 12). Such glan-dular trichomes are usually present on stems or leaves, but can also occur onovaries and pods. Stipitate glands have evolved in the clade including Brya,Cranocarpus, and Grazielodendron (uniquely from glochidiate trichomes inthe first two genera), and in polymorphic condition in the clade includingAdesmia, Chapmannia, and Stylosanthes. In Brya and Cranocarpus, stipitateglands are found, in addition to the foliage, on the ovary where they persistwith the mature fruit. The high tree scores for this character (Table 1) do notaccount for the optimizations of polymorphic codings where Chapmannia,Stylosanthes, and Adesmia were assigned state zero during parsimony analy-sis.
Inflorescence characters
14. Inflorescence position: 0) axillary, 1) terminal. The first state corre-sponds to leafy flowering branches that are indeterminate with vegetativegrowth from the apical meristem. The second refers to leafy flowering branch-
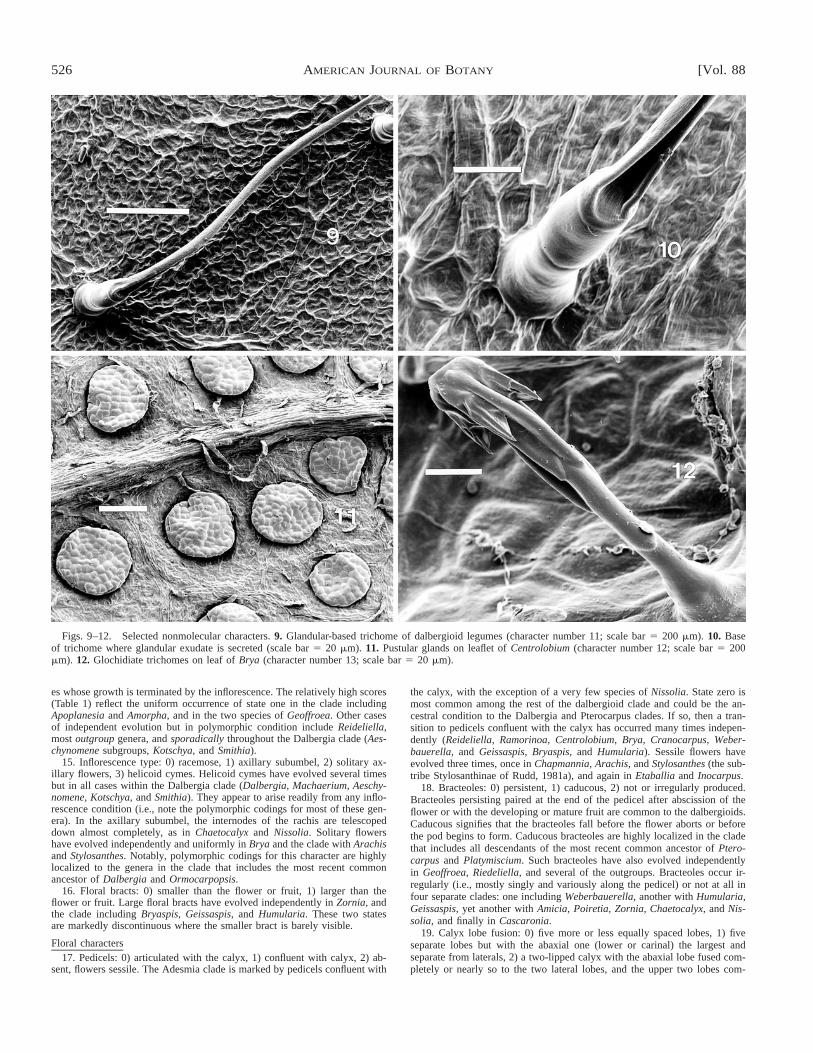
526 [Vol. 88AMERICAN JOURNAL OF BOTANY
Figs. 9–12. Selected nonmolecular characters. 9. Glandular-based trichome of dalbergioid legumes (character number 11; scale bar 5 200 mm). 10. Baseof trichome where glandular exudate is secreted (scale bar 5 20 mm). 11. Pustular glands on leaflet of Centrolobium (character number 12; scale bar 5 200mm). 12. Glochidiate trichomes on leaf of Brya (character number 13; scale bar 5 20 mm).
es whose growth is terminated by the inflorescence. The relatively high scores(Table 1) reflect the uniform occurrence of state one in the clade includingApoplanesia and Amorpha, and in the two species of Geoffroea. Other casesof independent evolution but in polymorphic condition include Reideliella,most outgroup genera, and sporadically throughout the Dalbergia clade (Aes-chynomene subgroups, Kotschya, and Smithia).
15. Inflorescence type: 0) racemose, 1) axillary subumbel, 2) solitary ax-illary flowers, 3) helicoid cymes. Helicoid cymes have evolved several timesbut in all cases within the Dalbergia clade (Dalbergia, Machaerium, Aeschy-nomene, Kotschya, and Smithia). They appear to arise readily from any inflo-rescence condition (i.e., note the polymorphic codings for most of these gen-era). In the axillary subumbel, the internodes of the rachis are telescopeddown almost completely, as in Chaetocalyx and Nissolia. Solitary flowershave evolved independently and uniformly in Brya and the clade with Arachisand Stylosanthes. Notably, polymorphic codings for this character are highlylocalized to the genera in the clade that includes the most recent commonancestor of Dalbergia and Ormocarpopsis.
16. Floral bracts: 0) smaller than the flower or fruit, 1) larger than theflower or fruit. Large floral bracts have evolved independently in Zornia, andthe clade including Bryaspis, Geissaspis, and Humularia. These two statesare markedly discontinuous where the smaller bract is barely visible.
Floral characters17. Pedicels: 0) articulated with the calyx, 1) confluent with calyx, 2) ab-
sent, flowers sessile. The Adesmia clade is marked by pedicels confluent with
the calyx, with the exception of a very few species of Nissolia. State zero ismost common among the rest of the dalbergioid clade and could be the an-cestral condition to the Dalbergia and Pterocarpus clades. If so, then a tran-sition to pedicels confluent with the calyx has occurred many times indepen-dently (Reideliella, Ramorinoa, Centrolobium, Brya, Cranocarpus, Weber-bauerella, and Geissaspis, Bryaspis, and Humularia). Sessile flowers haveevolved three times, once in Chapmannia, Arachis, and Stylosanthes (the sub-tribe Stylosanthinae of Rudd, 1981a), and again in Etaballia and Inocarpus.
18. Bracteoles: 0) persistent, 1) caducous, 2) not or irregularly produced.Bracteoles persisting paired at the end of the pedicel after abscission of theflower or with the developing or mature fruit are common to the dalbergioids.Caducous signifies that the bracteoles fall before the flower aborts or beforethe pod begins to form. Caducous bracteoles are highly localized in the cladethat includes all descendants of the most recent common ancestor of Ptero-carpus and Platymiscium. Such bracteoles have also evolved independentlyin Geoffroea, Riedeliella, and several of the outgroups. Bracteoles occur ir-regularly (i.e., mostly singly and variously along the pedicel) or not at all infour separate clades: one including Weberbauerella, another with Humularia,Geissaspis, yet another with Amicia, Poiretia, Zornia, Chaetocalyx, and Nis-solia, and finally in Cascaronia.
19. Calyx lobe fusion: 0) five more or less equally spaced lobes, 1) fiveseparate lobes but with the abaxial one (lower or carinal) the largest andseparate from laterals, 2) a two-lipped calyx with the abaxial lobe fused com-pletely or nearly so to the two lateral lobes, and the upper two lobes com-

March 2001] 527LAVIN ET AL.—DALBERGIOID LEGUMES
Figs. 13–16. Petal characters (scale bar 5 5 mm for all figures). Figs. 13–15. Petals differentiated into blade and claw in Geoffroea (character number 23).13. Standard. 14. Wing. 15. Keel. 16. Petals not differentiated into a blade and claw in Inocarpus.
pletely fused, 3) Dipteryx type, 4) Fissicalyx type, 5) Inocarpus type. Char-acter state one is synapomorphic for the dalbergioid clade, and it is mostdistinctive developmentally with the abaxial sepal initiating with the largersize and faster growth rate relative to the other sepals (Klitgaard, 1999a). Themost notable derivation from this condition within the dalbergioids is statetwo, which occurs in the clade with Aeschynomene sect. Aeschynomene, Cy-clocarpa, Soemmeringia, Kotschya, Smithia, Geissaspis, Bryaspis, and Hu-mularia (Aeschynomeneae subtribe Aeschynomeninae of Rudd, 1981a). TheDipteryx type occurs in Dipteryx, Pterodon, and Amicia zygomeris, where theupper two calyx lobes are greatly enlarged, contrasting with the diminutivelower three lobes. The Fissicalyx type evolved only in Fissicalyx, where allthe calyx lobes occur as an upper lip (spathaceous). The Inocarpus type hasthree lips, the lower formed by the carinal lobe, and two lateral lips formedby one lateral and one vexillar lobe. Also, the Socotran species of Chapman-nia have yet another type where the upper lip of a bilabiate calyx comprisesthe two upper and two lateral calyx lobes, and the lower lip comprises justthe carinal lobe. Chapmannia, however, was scored for state one because itrepresents the ancestral state for the genus (see the Chapmannia phylogenyin Lavin et al., 2000). Similarly, the ancestral state for Pictetia is state one(Beyra-M. and Lavin, 1999).
20. Hypanthium: 0) not well developed, petals and stamens arising at thebase of the ovary, 1) short-tubular, petals and stamens arising from a rimpositioned above the ovary base but not above the ovary itself, 2) long-tubular,where petals and stamens arise from a rim located above the ovary. Thecalyces of Acosmium, Apoplanesia, Amorpha, and Etaballia have a poorlydeveloped hypanthium (the last genus represents the only reversion to a lossof the hypanthium among dalbergioid legumes). Among the dalbergioids, stateone predominates and is ancestral. State two is confined to the clade includingChapmannia, Arachis, and Stylosanthes.
21. Petal coloration: 0) predominantly whitish to reddish or violet, 1) pre-dominantly yellow. The large majority of dalbergioid legumes have yellowpetals, and this is inferred to be ancestral. Notable exceptions include theclade with Dalbergia and Machaerium (polymorphic), as well as Grazielo-dendron, Paramachaerium, Ormocarpum (polymorphic), and Adesmia (thespecies with solitary axillary flowers) where whitish to violet petals are com-mon.
22. Corolla symmetry: 0) bilateral (papilionoid), 1) nearly radial. State zeropredominates among dalbergioids and indeed most papilionoids. Notably, anearly radially symmetric flower has evolved independently four times: onceeach in Inocarpus, Etaballia, Reideliella, and the clade with the samples ofAmorpheae (Apoplanesia and Amorpha). Nearly radially symmetric flowershave also evolved independently in the four species of Pictetia where statezero is considered ancestral (Beyra-M. and Lavin, 1999).
23. Petal morphology: 0) petals abruptly differentiated into a blade andclaw (Figs. 13–15), 1) petals ligulate, the claw and blade not distinguishable(Fig. 16). State one evolved separately in Etaballia and Inocarpus. This char-acter is not dependent on character number 22 because Amorpheae, Reide-liella, and four species of Pictetia have a nearly radial flower symmetry withpetals differentiated into a blade and claw.
24. Keel petals: 0) free, 1) connate, at least along the carinal margin if notto near the tip. Free keel petals in papilionoid legumes have been the hallmarkof the tribes Swartzieae and Sophoreae. Acosmium and Myrospermum, tra-ditionally placed in the tribe Sophoreae, have free keel petals. Among thedalbergioid legumes, fused keel petals represent the ancestral condition thathas reverted back to the free condition only in Etaballia, Geoffroea, Riede-liella, Platypodium, and Tipuana (all confined to the Pterocarpus clade).
25. Wing petals: 0) smooth, 1) crimped. Crimped wing petals are distinctlymuch broader than the adjacent keel petals. The evolution of such wing petalshas occurred in the clade including Paramachaerium, Pterocarpus, Ramori-noa, Paramachaerium, Tipuana, and Platypodium, and separately in that in-cluding Geoffroea.
Androecial characters
26. Staminal filaments: 0) all free from the base, 1) diadelphous 9 1 1, 2)open monodelphous, 3) closed monodelphous, 4) diadelphous [5 1 5, 5 1 41 1, or 4 1 1 1 4 1 1] with at least two phalanges of fused filaments. Statetwo is the inferred ancestral condition of the dalbergioid legumes. State fouris synapomorphic for the Dalbergia clade (though species of Ormocarpum,Pictetia, Diphysa, Machaerium, and Dalbergia are polymorphic). State four,however, has evolved independently in Platypodium and Discolobium (bothof the Pterocarpus clade). The free stamens of Adesmia are inferred to rep-resent a reversion from an open monodelphous condition. State one is uncom-mon among dalbergioids and is monomorphic only among some members thePterocarpus clade (Grazielodendron, Geoffroea, and Ramorinoa). The di-adelphous 9 1 1 condition is associated with weakly developed basal fenes-trae (Klitgaard, 1999a).
27. Anther size and attachment: 0) monomorphic, basi- to dorsi-fixed, 1)dimorphic, the smaller anthers usually dorsifixed, the larger basifixed. Char-acter state one evolved independently in the clade with Amicia (polymorphic),Poiretia, and Zornia, and again in that with Arachis and Stylosanthes. Di-morphic anthers also have other sporadic occurrences, such as in Aeschyno-mene genistioides (Aeschynomene sect. Ochopodium; Rudd, 1967, 1972a), themonotypic Aeschynomene subgen. Bakerophyton (Verdcourt, 1971), and in aMesoamerican clade of Platymiscium species (B. Klitgaard, unpublished data).
28. Staminal flange and filaments post-anthesis: 0) readily caducous, not

528 [Vol. 88AMERICAN JOURNAL OF BOTANY
persisting with the maturing fruit, 1) persistent on the abaxial side of fruit, 2)persistent on adaxial side of fruit. This trait is only partially conditional uponcharacter number 27 (see Beyra-M. and Lavin, 1999). Persistent stamens arediagnostic of the Dalbergia clade where the predominant condition is stateone. State two occurs in a clade with Aeschynomene sect. Aeschynomene,Kotschya, etc. Persistent stamens evolved separately in Brya, Cascaronia, andseveral outgroup genera.
Gynoecial characters
29. Locule: 0) encompassing nearly the entire length of the ovary, 1) con-fined to the basal end of the ovary. The locule is situated just above the stipein state one, and a large portion of the distal end is solid. All five occurrencesof this state are inferred to be cases of independent evolution (Vatairea, Va-taireopsis, Tipuana, Centrolobium, and Paramachaerium).
30. Nectary disk: 0) absent; 1) present. A nectary disk surrounding the baseof the ovary is known from Paramachaerium (Rudd, 1981a), most species ofOrmocarpum (M. Thulin and M. Lavin, unpublished data), and occasionallyin Machaerium (Klitgaard, 1999a). The retention index is undefined in thischaracter (Table 1) because Machaerium and Ormocarpum were assigned anancestral state of zero.
Fruit and seed characters
31. Pod valves: 0) loments present during early stages of fruit development,1) loments present by late stages, 2) valves continuous. Articulations forminglate during pod development occur in Ormocarpum, Pictetia, Diphysa, Chae-tocalyx, and Nissolia. Some species in the first of these three genera forminarticulate pods. Only in the Adesmia clade is the lomented condition uni-formly present. Both of the Dalbergia and Pterocarpus clades combine generawith articulate and inarticulate pods.
32. Pod margins: 0) straight, with no marginal constrictions between seeds1) constricted between seeds. State one has evolved numerous times indepen-dently in Discolobium, Fiebrigiella, Brya and Cranocarpus (polymorphic),Amicia, and Giessaspis, Bryaspis, Humularia, Kotschya, and Smithia. Aes-chynomene, Ormocarpum, and Diphysa are distinctively polymorphic for thischaracter.
33. Stipe of mature pod: 0) absent to less than half the length of the calyxtube, 1) surpassing the length of the calyx tube. State one has evolved mostuniformly in the clade with Dalbergia, Machaerium, Diphysa, Ormocarpum,Ormocarpopsis, Peltiera, and Pictetia. Among dalbergioids, state zero pre-dominates only in the Adesmia clade. Otherwise, both states have been gainedand lost on many separate occasions, particularly in the Pterocarpus clade.
34. Nervation of the mature pod valve in the region of the seed chamber:0) primarily reticulate, 1) primarily longitudinally parallel. This trait hasevolved once in the clade with Arachis, Stylosanthes, Chapmannia, and Fie-brigiella, (not Fissicalyx, however), and again in the clade with Chaetocalyxand Nissolia. Some species of Diphysa, Ormocarpum, and Pictetia have podswith strong longitudinal nerves, but these genera were optimized during anal-ysis as having state zero.
35. Replum: 0) placental margin disarticulating with the pod valves orarticles, 1) the valves or articles disarticulating separately from the persistentplacental margin. The last state is gained independently in Adesmia sect. Mur-icatae, Cyclocarpa, and in species of Aeschynomene sect. Aeschynomene (e.g.,A. villosa). This character had an undefined retention index (Table 1) becauseAdesmia and Aeschynomene were optimized for state zero.
36. Development of pod wings: 0) not winged, 1) wing from expansion ofthe ovary wall, 2) wing from expansion of the ovary sutures, 3) wing fromattenuation of the distal end of the ovary (i.e., the style), 4) winged fromattenuation of the proximal end of the ovary (i.e., the stipe). This characteris derived from Lima’s (1990) developmental work on samaroid fruits of tribeDalbergieae. He distinguished wings whose area of origin was the ovary walls(state one); wings with an origin of the ovary sutures (state two); and wingswith an origin from the distal end of the ovary (state three). Lima consideredVatairea and Vataireopsis to have a separate state, with wings derived fromthe solid distal portion of the ovary. This reflects the different morphology ofthe gynoecium in these two genera (see state one of character 29). We con-sider that the origin of the wing in these taxa is merely from the distal endof the ovary, and thus they are coded with state three. A further modificationis that we consider the basal wing of Platypodium to be derived from anexpansion of the stipe (state four). Developmental anatomical work couldconfirm these distinctions. State one has evolved independently in both theDalbergia (Dalbergia and Weberbauerella) and Pterocarpus clades (Platym-iscium, Cranocarpus, Grazielodendron, Ramorinoa, Pterocarpus, Fissicalyx,
and Riedeliella). State three has evolved in every occurrence separately (Va-tairea, Vataireopsis, Nissolia, Machaerium (polymorphic), Tipuana, Centro-lobium, and Paramachaerium).
37. Inner epidermis and endocarp: 0) lignescent, 1) spongy and adheringto the mature seeds. State one evolved in the clade with Chaetocalyx andNissolia, and again in that with Pictetia, and perhaps again in Peltiera (thesister genus of Ormocarpopsis—see Labat and Du Puy, 1997).
38. Mesocarp: 0) lignescent, 1) spongy, 2) fleshy. State one arose indepen-dently and uniformly in Pictetia and Fiebrigiella, and sporadically in Or-mocarpum, Chapmannia, Nissolia, and Chaetocalyx. State two occurs in thefleshy vertebrate dispersed fruits of Dipteryx, Andira and Geoffroea, all in-stances of independent evolution. Polymorphic terminals were optimized forstate zero, effectively underestimating the actual levels of homoplasy.
39. Exocarp: 0) adnate to the mesocarp, 1) loosely attached to mesocarp,2) separate from the mesocarp by the formation of a distinct air chamber.This last trait occurs in many species of Diphysa and a few species of Nissolia(e.g., N. leiogyne). State one is characteristic of Ormocarpopsis. The high treescores for this character (Table 1) resulted from state zero being assigned tothe polymorphic terminals Diphysa and Nissolia during parsimony analysis.
40. Pod coiling: 0) coiled in a forward directed manner, 1) not coiled, 2)coiled in a laterally directed manner. The forward coil of the pod is confinedto Discolobium, whereas the lateral coil has evolved in Cyclocarpa and var-ious species of Aeschynomene sect. Aeschynomene and Ormocarpum. Al-though the lateral coil is restricted to members of the Dalbergia clade, stateone was assigned to the polymorphic terminals Aeschynomene and Ormocar-pum during parsimony analysis, resulting in high tree scores (Table 1).
41. Pod valve ornamentation: 0) not present, 1) multiseriate trichomes, 2)crests and bumps. Multiseriate trichomes persisting on the mature pod valvehave evolved in many separate occasions throughout the dalbergioid legumes,as in Adesmia (polymorphic), Ormocarpum (polymorphic), Brya, and Centro-lobium (where they become spinose). Pod valves with crests or bumps haveevolved once in the clade with Poiretia, Amicia, and Zornia, and again inpolymorphic condition among various species of Aeschynomene sect. Aeschy-nomene. The polymorphic terminals Adesmia, Ormocarpum, and Aeschyno-mene were assigned state zero during parsimony analysis, thus resulting inrelatively high tree scores (Table 1).
42. Seed shape: 0) lenticular to spherical with a centrally placed hilum, 1)reniform with a central recessed hilum, 2) longitudinally elongate with thehilum placed toward the end toward the style. The last condition is charac-teristic of most dalbergioids and indeed ancestral to that clade. However, statetwo has evolved independently in Dipterygeae (Dipteryx and Pterodon), Hy-menolobium, Vatairea (polymorphic), and Vataireopsis. Among dalbergioids,Aeschynomene, Kotschya, Smithia, and Platymiscium have reniform seeds(three separate origins), and Ormocarpopsis and Peltiera have spherical seeds.
43. Orientation of the seed in the fruit: 0) longitudinal, 1) oblique to trans-verse. Oblique to transverse orientation of seeds is confined to a subcladeincluding Platymiscium, Centrolobium, Paramachaerium, Pterocarpus, Ra-morinoa, and Tipuana (Lima, 1990). Such seeds have also evolved indepen-dently in Hymenolobium (polymorphic).
Seedling characters
44. Position of the eophylls: 0) alternate, 1) opposite. Among dalbergioids,opposite eophylls are confined to the Pterocarpus clade where they are knownfrom Platymiscium, Grazielodendron, Ramorinoa, Centrolobium, and Geof-froea.
45. Number of leaflets in the first eophyll: 0) one, 1) more than one. Dal-bergioids commonly have eophylls that are not strongly differentiated fromthe adult leaves (i.e., multifoliolate). The Pterocarpus clade is exceptional inhaving all known instances where the eophylls are unifoliolate. Data for char-acters 44 and 45 for Ramorinoa came from Burkart (1952, p. 238).
Pollen characters
46. Aperture type: 0) tricolporate, 1) periporate. Tricolporate apertures arethe general and most common type in legumes. Among the dalbergioids, per-iporate pollen is known from only Brya and Cranocarpus.
47. Pollen pore: 0) without an operculum, 1) with an operculum. An oper-culum is a distinctly delimited ectexinous structure that covers the ectoaper-ture, which in the case of all dalbergioids means the colpus. State one hasbeen gained independently many times throughout all three principal cladesof the dalbergioid legumes.
48. Colpi (the polar region): 0) colpi short, not anastomosing at the polesof the pollen grain, the polar region entire, 1) colpi longer, the ends of colpi

March 2001] 529LAVIN ET AL.—DALBERGIOID LEGUMES
anastomosing, forming syncolpi. State one evolved independently and uni-formly in Humularia and Vataireopsis. This state is polymorphic for Aeschy-nomene, Kotschya, and Smithia. Thus, state one is confined to the Dalbergiaclade among the dalbergioid legumes.
49. Wall stratification (Guinet and Ferguson, 1989): 0) well-developed end-exine and footlayer, 1) thickening of the endexine at least at the aperturescombined with a reduction in the foot layer, 2) reduction of the endexinecombined with a thickening of the foot layer, 3) reduction of the endexineand foot layer combined with an elongation of the columellae. State one ismost common among the dalbergioids and is inferred to be ancestral in thisclade. State two evolved independently in Adesmia and the outgroup Myros-permum. State three evolved once in the clade with Geissaspis and Bryaspis.
Wood characters
50. Ray size: 0) three or more cells wide, and taller than 20 cells high, 1)1–2 cells wide and less than 15–20 cells high. Many of the dalbergioid generahave narrow short rays (state one). Good examples include Platymiscium,Fissicalyx, and the nondalbergioid Dipteryx. Outgroup genera have storiedrays that are larger than this. Wood characters were scored from examinationof slides in the collections at the Jodrell Laboratory, Kew, and at the ForestProducts Laboratory, Madison, Wisconsin, USA, and by reference to descrip-tions and photographs in Baretta-Kuipers (1981), Gasson (1994, 1999), Miles(1978), and Detienne and Jacquet (1983). Wheeler, Bass, and Gasson (1989)provide thorough definitions for all of the wood characters used in this anal-ysis. Details on the wood anatomy of Chaetocalyx, Nissolia, Poiretia, Amicia,Zornia, Chapmannia, Arachis, Stylosanthes, Soemmeringia, Smithia and Geis-saspis come from Cumbie (1960). Unfortunately, the information is presentedin such a way that only a few character states can be coded. Amorpha fruti-cosa has been illustrated and described by Schweingruber (1990), Adesmiahorrida by Roig (1986), Discolobium by Cozzo (1949, 1950), and Parama-chaerium by Brizicky (1960). No information on the wood anatomy is avail-able for Riedeliella, Cranocarpus, Fiebrigiella, Cyclocarpa, Kotschya, Bryas-pis, Humularia, Weberbauerella, Ormocarpum, Ormocarpopsis, and Peltiera.
51. Ray arrangement: 0) not storied, 1) storied. In legumes, storied rays,axial parenchyma, and adjacent vessels are common and can be observed intangential longitudinal section. Although considered a very useful anatomicalcharacter, both diagnostically and cladistically, there are many legume generawith storied rays that are irregular, or obvious in short rays and less so intaller rays which may be axially fused. Storied rays are particularly stronglydeveloped in Dipteryx, Pterocarpus, Platymiscium, Grazielodendron, Etabal-lia, Inocarpus, Dalbergia, Machaerium, and Aeschynomene, all of which haveshort rays. The taxa with larger rays often do not exhibit such regular storiedarrangement, and axial fusion is often the cause, as in species of Acosmium.
52. Composition of cells in rays: 0) homocellular, 1) heterocellular. Thisfeature is observed in radial longitudinal section. Homocellular rays are com-posed entirely of procumbent ray cells. Heterocellular rays in legumes arecomposed mainly of procumbent cells, but there are also some square orupright cells, usually in a row or rows at the ray margins (i.e., at the top orbottom of a ray). These two character states are not mutually exclusive. Ju-venile wood often tends to be more heterocellular than mature wood, and itis not always apparent where exactly a wood sample was taken from if thepith in the stem is not included.
53. Crystals in ray cells: 0) absent, 1) present in some ray cells. Prismaticcrystals of calcium oxalate are found in many, if not most legumes. They arealmost ubiquitous in chambered axial parenchyma strands, but in a few generacan also be found in ray cells. The main difficulty with this character is thatif the crystals are rare they can be overlooked. They are searched for in radiallongitudinal section, because they are even more difficult to find in tangentiallongitudinal section.
54. Axial parenchyma: 0) not abundantly aliform and confluent, 1) abun-dantly aliform and confluent. Axial parenchyma patterns in legumes are verydifficult to code. All the legumes in this study have predominantly paratra-cheal parenchyma, with the addition of some apotracheal diffuse parenchymain particularly Dalbergia and Platymiscium. This ranges continuously fromscanty paratracheal, vasicentric, aliform, to confluent. In the opinion of oneof us (Gasson), these all constitute one character. They could each be codedas character states, but virtually all wood samples in the legumes exhibit morethan one condition. Unilaterally paratracheal parenchyma is found in some ofthe taxa, and probably forms part of this continuum. Banded parenchyma,which could be treated separately, may be an extreme form of confluent pa-renchyma, particularly if the bands are several cells wide. Some taxa in thestudy group do have narrow bands, but they are not distinguished here. Thechoice of the two character states above serves to separate four of the genera
very well, but does not distinguish all the other complicated variations on theparatracheal theme exhibited by the taxa coded as zero. Aeschynomene is verydifferent, in that it has such abundant parenchyma, that the fibers exhibit awinged-aliform appearance.
Nitrogen fixation character
55. Root nodule: 0) none produced, 1) produced as a non-aeschynomenoidnodule, 2) produced as an aeschynomenoid nodule (Fig. 6; Corby, 1981; seediscussion). State two is synapomorphic for the dalbergioid clade, althoughsome genera in this clade are known not to produce nodules at all (i.e., Chae-tocalyx and Nissolia), as is the case for some nondalbergioids (e.g., Myros-permum, Dipteryx, Pterodon, Vatairea, and Vataireopsis). Some outgroupsproduce nodules of a type other than aeschynomenoid (e.g., Acosmium, Poe-cilanthe, Andira, Hymenolobium, and Amorpha). Although lost within thedalbergioid clade (Table 1), the aeschynomenoid root nodule is not encoun-tered elsewhere in the legume family. The stem (but not root) nodules ofSesbania rostrata (tribe Robinieae) are superficially similar, but they differfrom the aeschynomenoid type in having an apical meristem, albeit ephemeral(J. Sprent, unpublished data). Three dalbergioid genera, Cyclocarpum, Geis-saspis, and Paramachaerium, are known to produce nodules, but the exacttype is unknown. These genera were variously coded as having missing dataor state one. Such alternative coding did not affect how these genera wererelated with respect to the three major subclades of the dalbergioid clade.Future studies incorporating nodule morphology in a phylogenetic analysiswill do well to recognize specific morphologies independent of nodule cate-gories. Specifically, these would include characters such as the apical meri-stem (absent vs. present), infection site (associated with emergent rootlet ornot), infection threads (absent vs. present), and central tissue (uniformly in-fected vs. uninfected). The aeschynomenoid type is defined as having the firststate of each of these four characters. Regardless, this coding strategy wouldnot change our findings because the dalbergioids would be nearly uniform inoccurrence for the states of these four characters.
APPENDIX C
Enumeration of the constituent genera of the dalbergioid clade. The em-phasis in the discussion of each of the dalbergioid genera is on the diagnostictraits that are presumably autapomorphic.
The Adesmia clade
Adesmia DC. is diagnosed by stipules attached a pseudopetiole. Althoughalso found in Stylosanthes and Arachis, the projected portions of the nodesof these two genera are nearly as long as the petiole. In Adesmia, the nodalprojections extend to much less than half the length of the petiole. In addition,Adesmia uniquely combines free staminal filaments and lomented pods (Pol-hill, 1981f). Adesmia comprises about 230 species centered in Chile and Ar-gentina (Burkart, 1949, 1954, 1960, 1962, 1964, 1966, 1967; Ulibarri, 1978,1980, 1982a, b, 1984, 1987, 1990). The genus contains two distinct mono-phyletic subgroups, according to ITS/5.8S sequence analysis (Fig. 3). One ismarked by inflorescences of usually solitary axillary flowers with pedicelsconfluent with the calyx, stipules (or at least scars) that are connate aroundthe stem, and pods that lack glandular-based trichomes, multiseriate trichomes,or the raised pericarp reticulations (e.g., Adesmia lanata and A. villosa). Thesecond clade is characterized by inflorescences of terminal racemes, subum-bels, or panicles, flowers articulated with the pedicel, stipules (or scars) thatare not connate around the stem, and pod loments that commonly bear sometype of ornamentation, for example, large glandular-based trichomes, longmultiseriate plumose trichomes, or very prominent reticulate venation (e.g.,Adesmia muricata and A. volckmannii). Phylogenetic analysis of ITS/5.8Ssequence data strongly supports the monophyly of Adesmia, as does matK/trnK.
Chaetocalyx DC. (Figs. 17–23) is paraphyletic with respect to Nissolia, anissue that is the focus of another study (M. Lavin and D. Prado, unpublisheddata). It possesses no autapomorphic traits and is characterized like Nissolia(with twining herbaceous stems and ebracteolate flowers) but lacking the ster-ile (usually samaroid) terminal loment of the mature pod. The glandular-basedtrichomes on the calyx of most species of Chaetocalyx are not diagnostic andthe species of Chaetocalyx form a rather homogeneous assemblage. The sup-posedly obvious division between species with laterally flattened or wingedfruits vs. those with terete fruits (as coded in Beyra-M. and Lavin, 1999) isnot resolved with 5.8S/ITS sequence analysis. Chaetocalyx includes ;13 neo-tropical species centered in dry forests of South America (Rudd, 1958, 1972b,1996).

530 [Vol. 88AMERICAN JOURNAL OF BOTANY
Figs. 17–28. Representative species of the Adesmia clade (scale bar 5 1 cm for all figures). Figs. 17–23. Chaetocalyx brasiliensis. 17. Habit. 18. Calyx.19. Gynoecium. 20. Androecium. 21. Keel petal. 22. Wing petal. 23. Standard. Figs. 24–27. Nissolia wislizenii. 24. Habit. 25. Cauline leaf. 26. Flower. 27.Fruits. 28. Nissolia microptera, leafy stem with fruits. Reproduced from Volume 5 of Flora Novo-Galiciana by Rogers McVaugh.
Nissolia Jacq. is derived from within Chaetocalyx (Figs. 2, 3, and 5) andcharacterized by the autapomorphy of pods with a sterile (usually samaroid)terminal loment (Figs. 24–28). This genus contains ;13 species centered intropical dry forests of Mexico and Central America (Rudd, 1956, 1970a,1975b). Chaetocalyx and Nissolia lack floral bracteoles, otherwise occurringamong dalbergioids in Poiretia, Amicia, Zornia, Cyclocarpa, Humularia,Geissaspis, and Bryaspis.
Amicia Kunth occurs in Mexico, Ecuador, Peru, Bolivia, and Argentina(Rudd, 1981a). This genus is closely related to Poiretia and Zornia. All threehave legumes with crests or bristles on each pod article and leaves that areusually paripinnate (a few species of Poiretia have imparipinnate leaves).Amicia differs from Poiretia and Zornia in having blunt keel petals, a staminalsheath that is split open above, and anthers that are mostly uniform. A recentattempt to segregate Poiretia and Zornia from Amicia (Ohashi, 1999) is notsupported by this analysis.
Poiretia Vent. is confined to the Neotropics but with most species fromBrazil to northern Argentina (Rudd, 1972c). The genus is similar to Zornia,but differs in its usually twining habit and racemose inflorescences with small,single flower bracts at each node.
Zornia J. F. Gmel. occurs in southeastern United States, the Neotropics witha center of diversity in Brazil, and throughout sub-Saharan Africa (Mohlen-brock, 1961, 1962). It is marked by medifixed stipules (independently evolvedin Aeschynomene sect. Aeschynomene and relatives), leaves with digitatelyarranged few leaflets, and sessile flowers in axils of large paired bracts.
The Pterocarpus clade
Discolobium Benth. is readily diagnosed by its pod that coils in a forwarddirection with each of three turns compressed together into a single disc. Only
the middle loment is fertile. Its 4 1 1 1 4 1 1 diadelphous staminal columnis not unique and is found sporadically among the dalbergioids. Discolobiumcomprises eight species distributed from northern Argentina to adjacent Braziland Paraguay (Rudd, 1981a).
Riedeliella Harms comprises three species endemic to southeastern Braziland Paraguay (Lima and Studart da Fanseca Vaz, 1984). Like Inocarpus,Etaballia, and some species of Pictetia, the flowers of Riedeliella are nearlyradially symmetric. Lima and Studart da Fanseca Vaz (1984) propose the closerelationship of Etaballia and Riedeliella in the tribe Acosmieae (Yakovlev,1972), a group also with essentially radially symmetric flowers, although withfree staminal filaments. Riedeliella differs from Etaballia and Inocarpus inhaving paripinnate leaves and a long exerted style, and in this analysis it issuggested to be not most closely related to Etaballia, but rather to Discolob-ium.
Brya P. Br. is recorded to have explosive pollen release (Leon and Alain,1951, p. 315, fig. 131), a form of pollen presentation that is unique amongdalbergioid legumes. Also, Brya is characterized by its leaves from the longshoots being transformed into spines. Brya is sister to Cranocarpus, as evi-denced by the shared occurrence of leaves, stems, inflorescences, and podsbearing capitate glandular trichomes that are microscopically glochidiate, andby periporate pollen (Ferguson and Skvarla, 1981). Brya includes four speciesendemic to the Greater Antilles (Ohashi, Polhill, and Schubert, 1981; Lewis,1988).
Cranocarpus Bentham comprises three species endemic to Brazil (Harley,1978; Ohashi, Polhill, and Schubert, 1981). In all respects Cranocarpus islike Brya but the leaves from the long shoots are not transformed into spines.The yellow petals, base chromosome number of x 5 10, storied wood struc-ture (Record, 1919), and simple axillary racemes or solitary flowers of Brya

March 2001] 531LAVIN ET AL.—DALBERGIOID LEGUMES
Figs. 29–47. Representative species of the Pterocarpus clade (scale bar 5 1 cm for all figures). Figs. 29–36. Platymiscium trifoliolatum. 29. Floweringbranch. 30. Branch of fruiting inflorescences with wall of one fruit cut away to show seed. 31. Calyx. 32. Androecium. 33–34. Wing petals. 35. Keel petals.36. Standard. Figs. 37–47. Pterocarpus orbiculatus. 37. Detached leaf. 38. Inflorescence. 39. Mature fruits. 40. Immature fruits. 41. Calyx. 42. Androecium.43. Gynoecium. 44. Keel petals. 45–46. Wing petals. 47. Standard. Reproduced from Volume 5 of Flora Novo-Galiciana by Rogers McVaugh.
and Cranocarpus are traits strongly suggestive of a relationship with the dal-bergioid legumes.
Platymiscium Vogel (Figs. 29–36) comprises 18 neotropical species cen-tered in Mexico and northeastern Brazil. The genus is unique in having op-posite leaves with interpetiolar stipules (Lima, 1990; Klitgaard, 1999a, b).
Centrolobium Mart. ex Benth. comprises six tropical species from Panamato Colombia, Ecuador, Venezuela, Brazil, and Bolivia (Rudd, 1954; Lima,1985). The genus is well marked by its orange peltate glands covering theleaves and inflorescences, and winged pods in which the seed-bearing portionis covered with spines.
Grazielodendron Lima is a monotypic genus endemic to Brazil (Lima,1983, 1990). The laterally compressed pod of Grazielodendron is distin-guished by having an additional wing-like extension of the dorsal margin. Thewinged dorsal margin is markedly delineated from the main winged body ofthe pod.
Pterocarpus Jacq. (Figs. 37–47) comprises 20 species distributed pantrop-ically (Rojo, 1972). Of the genera with wide, crimped wing petals (i.e., Par-amachaerium, Geoffroea, Pterocarpus, Ramorinoa, Paramachaerium, Tipu-ana, and Platypodium), Pterocarpus is diagnosed by its pod that is wingedfrom an attenuation of the pod body all around the seed chamber (Polhill,1981d; Lima, 1990). The pods are variable in this genus with some wingedand bristly (e.g., P. angolensis), others winged and not bristly (e.g., P. indi-cus), and rarely not winged and not bristly (i.e., P. amazonum).
Tipuana (Benth.) Benth. is a monotypic genus of subtropical forests inBolivia and northwestern Argentina (Rudd, 1974). Of the genera with wide,crimped wing petals (see description of Pterocarpus), Tipuana is diagnosedby its pod that is winged from the style, the seed chambers being proximalto the wing (Lima, 1990; Polhill, 1981d).
Platypodium Vogel includes one or two species in Panama, Guatemala,Venezuela, Colombia, Bolivia, Brazil, and Paraguay. Of the genera with wide,
crimped wing petals (see description of Pterocarpus), Platypodium is diag-nosed by its pod that is winged from the stipe, the seed chambers being distalto the wing (Polhill, 1981d; Lima, 1990).
Paramachaerium Ducke includes five species from Panama, Guyana, Peru,and Brazil (Rudd, 1981a, b; Lima, 1990). Of the dalbergioid tree genera withlaterally broadened and crimped wing petals, Paramachaerium has reddish toviolet petals rather than the typical yellow pigment. This genus is unusual inits nectariferous disk surrounding the base of the ovary, a trait independentlyevolved in certain species of Machaerium and Ormocarpum.
Ramorinoa Speg. is a monotypic genus from west-central Argentina. Thegenus is very well marked by its leafless pungent branches (Burkart, 1952;Polhill, 1981d; Lima, 1990). As remarked by Burkart (1952), the genus is sohighly modified vegetatively that morphology provides few clues to its closestrelationships.
Inocarpus J. R. & G. Forster is very distinctive in having all five ligulatepetals fused at base, and with the ten stamens fused by their filaments to thecorolla tube (similar to some genera of Amorpheae). Inocarpus comprises oneto three species and is geographically distinctive in being restricted to Ma-laysia and adjacent Pacific islands (Polhill, 1981d). The only other dalbergioidgenus restricted to Asia is Geissaspis.
Etaballia Benth. is very similar to Inocarpus, except that its leaves areunifoliolate rather than simple, and the staminal filaments are monodelphouswith no split along the adaxial side. Etaballia is monotypic and unlike Ino-carpus is neotropical, occurring in Guyana, Venezuela, and Brazil (Rudd,1970b).
Geoffroea Jacq. comprises two species from Colombia and Venezuela southto Chubut, Argentina, and also on the Galapagos Islands possibly due tocultivation (Ireland and Pennington, 1999). Of the genera with wide, crimpedwing petals (see description of Pterocarpus), Geoffroea is diagnosed by itssessile ovary that develops into a fleshy drupe (Polhill, 1981d; Lima, 1990).

532 [Vol. 88AMERICAN JOURNAL OF BOTANY
Figs. 48–69. Representative species of the Dalbergia clade (scale bar 5 1 cm for all figures except where noted). Figs. 48–58. Dalbergia congestiflora. 48.Leafy branch. 49. Flowering branch. 50. Fruits. 51. Seed. 52. Androecium. 53. Anther (scale bar 5 1 mm). 54. Calyx. 55. Keel petals. 56–57. Wing petals. 58.Standard. Figs. 59–69. Machaerium kegelii. 59. Flowering branch. 60–61. Nodes with stipular spines. 62. Androecium. 63. Gynoecium. 64. Calyx. 65. Keelpetals. 66. Wing petal. 67. Standard. 68. Flower. 69. Fruit. Reproduced from Volume 5 of Flora Novo-Galiciana by Rogers McVaugh.
Cascaronia Griseb. is not readily diagnosed, but the combination of itsleaves and pods with dark pustular glands, pods with strong longitudinalnerves, arborescent habit, inflorescences of axillary racemes, and small yellowpetals is unique. Cascaronia is a monotypic genus from northern Argentinaand adjacent Paraguay and Bolivia (Polhill, 1981d).
Fissicalyx Bentham is a monotypic genus from Venezuela and Guyana, andmarked by its spathaceous calyx (all five lobes are on the adaxial lip), porateanthers, and pods with a fusiform seed chamber bearing a closely veinedmembranous wing on both margins (Polhill, 1981d; Lima, 1990). There is nomorphological evidence to suggest that this genus is closely related to Fie-brigiella, as revealed by DNA sequence analysis.
Fiebrigiella Harms is a monotypic genus from Bolivia and Ecuador (Bur-kart and Vilchez, 1971). The pods of Fiebrigiella have prominent continuousparallel venation on the lateral walls, once suggesting an affinity to Chaeto-calyx and Nissolia (Rudd, 1981a), but now considered homologous to suchpods of the genera Chapmannia, Stylosanthes, and Arachis.
Chapmannia Torr. & Gray is recently expanded from monotypic (Gunn,Norman, and Lassetter, 1980) to include seven species of seasonally dry veg-etation, two New World (Florida and Mesoamerica), and five Old World (So-malia and the Yemeni island Socotra; Thulin, 2000). Arthrocarpum Balf. f.(Gillett, 1966) and Pachecoa Standl. & Steyerm. (Burkart, 1957) are synon-ymized. The genus is diagnosed by its dried (herbarium preserved) leafletswith uniformly reddish reticulate tannin deposits on the abaxial surface. Chap-mannia is sister to Arachis, and Stylosanthes; the species of these two lattergenera do not consistently show the reddish tannin reticulations. Chapmannia,Arachis, and Stylosanthes form a monophyletic group marked in part by theirsessile flowers with long hypanthia. Within this group, Chapmannia maintains
the plesiomorphic spicate inflorescence, whereas Arachis and Stylosantheshave inflorescences of solitary axillary flowers.
Stylosanthes Swartz and Arachis share the synapomorphy of stipules unitedto nodal projections, which in turn are superficially continuous with the petiole(a trait known also from Adesmia). Stylosanthes is distinguished from Arachisby having lomented, nongeocarpic pods, which are presumably plesiomorphic,as well as ovaries that are uniformly covered by uniseriate trichomes, anautapomorphy. The ;25 species of Stylosanthes are distributed in warm tem-perate to tropical regions of the world, but with a center of diversity in theneotropics (Mohlenbrock, 1957, 1960, 1963; Rudd, 1981a).
Arachis L. is distinguished from Stylosanthes by its flowers with a verylong and narrow hypanthium, a gynophore (Moctezuma and Feldman, 1998)that renders the pods geocarpic, nonlomented glabrous pods, and mostly fourleaflets per leaf. The 69 species of Arachis originate in South America froma region including Brazil south to northern Argentina (Krapovickas and Greg-ory, 1994).
The Dalbergia clade
Dalbergia L. f. (Figs. 48–58) is diagnosed by small ovate to obovate an-thers with short transverse slits at dehiscence. The genus includes over 100species distributed pantropically, but with centers of diversity in Amazoniaand Indo-Asia (Prain, 1904; Pittier, 1922; Polhill, 1981d; Lima, 1990; de Car-valho, 1997).
Machaerium Pers. (Figs. 59–69) includes ;120 neotropical species, al-though M. lunatum (L. f.) Ducke also occurs in western Africa. Machaeriumis related to Dalbergia (Polhill, 1981d; Doyle et al., 1997) and Aeschynomene

March 2001] 533LAVIN ET AL.—DALBERGIOID LEGUMES
sect. Ochopodium, as evinced in part by inflorescences of helicoid cymes (butpolymorphic in all three taxa). Machaerium differs in its spinescent recurvedstipules (on at least the climbing species) and pods that are usually distallywinged, or at least have the seed chamber toward the base (Rudd, 1973, 1977,1986, 1987; Polhill, 1981d; de N. Carmo-Bastos, 1987; Lima, 1990).
Aeschynomene L. sect. Ochopodium. Aeschynomene sensu lato includesspecies that do not fit the diagnosis of the other dalbergioid genera. It is forthis reason that the genus is treated with two terminal taxa, sects. Ochopodium(with basifixed stipules) and Aeschynomene (with medifixed stipules). SectionOchopodium, according to DNA sequence analysis, is more closely related toMachaerium than to sect. Aeschynomene (e.g., section Ochopodium is rep-resented by Aeschynomene purpusii and A. fasicularis in Fig. 5). Regardless,it is not certain if either of these two sections are monophyletic, a topic thatwill have to be taken up elsewhere given their large taxonomic size. As such,sect. Ochopodium includes ;101 species distributed pantropically (Rudd,1955, 1967, 1975a).
Aeschynomene sect. Aeschynomene is diagnosed by medifixed stipules,which are also characteristic of the closely related Smithia and Geissaspis.Thus, this taxon (represented by Aeschynomene americana, A. indica, A. pfun-dii, A. rudis, and A. virginica in Figs. 2 and 5), potentially lacking any obviousmorphological apomorphy, could be paraphyletic with respect to at least someof the genera listed immediately below. It is beyond the scope of this analysisto address this potential problem. As such, sect. Aeschynomene comprises;50 species with a pantropical distribution (Leonard, 1954; Rudd, 1955,1959, 1972a; Verdcourt, 1971; Fernandes, 1996).
Soemmeringia Mart. is characterized by a scarious standard petal that per-sists with the mature pod, which is independently evolved in some species ofOrmocarpum. Soemmeringia is a monotypic, neotropical genus from Brazil,Bolivia, and Venezuela (Rudd, 1981a). Soemmeringia, along with Cyclocarpa,Kotschya, Smithia, Geissaspis, Bryaspis, and Humularia (below), are allclosely related to sect. Aeschynomene because of their paripinnate leaves,usually alternate leaflets, and bilabiate calyces (Rudd, 1981a).
Cyclocarpa Afz. ex Bak. is diagnosed by pods that have one lateral spiral,the pod articles of which disarticulate from a persistent placental margin orreplum. This monotypic genus is locally common across tropical Africa, andin southeast Asia (Laos and Borneo) and northern Australia (Hepper, 1958).
Kotschya Endl. and Smithia (below) are characterized by an inflorescenceof a dense strobilate helicoid cyme, a pod enclosed by the calyx and in whichthe articles are folded against each other. Kotschya differs in having alternateleaflets that each bear 2–7 basal nerves, as well as basifixed stipules. Kotschyacomprises 31 species restricted to tropical Africa and Madagascar (Gillett,Polhill, and Verdcourt, 1971; Verdcourt, 1974; Rudd, 1981a).
Smithia Ait. differs from Kotschya by its medifixed stipules and oppositeleaflets each bearing one main nerve. Smithia comprises ;30 species mainlyin Asia and Madagascar (Gillett, Polhill, and Verdcourt, 1971; Verdcourt,1974; Rudd, 1981a).
Geissaspis Wight & Arn. together with Bryaspis and Humularia (below)are characterized by large inflorescence bracts that completely envelop thesubtending flower and fruit (independently evolved in Zornia). Geissaspis andBryaspis differ by ebracteolate flowers, and Geissaspis differs from Bryaspisby its medifixed stipules. Geissaspis comprises three species confined to trop-ical and subtropical central and southeast Asia, but not crossing Wallace’s line(Gledhill, 1968; Rudd, 1981a)
Bryaspis Duvign. includes two species from tropical west Africa (Gledhill,1968; Hepper, 1958; Gillett, Polhill, and Verdcourt, 1971; Rudd, 1981a). Un-like Geissaspis, the inflorescence bracts of Bryaspis are markedly imbricate.
Humularia Duvign. differs from Geissaspis and Bryaspis by emarginateinflorescence bracts and panduriform standard petals. Humularia comprises;40 species confined to central Africa (Gledhill, 1968; Gillett, Polhill, andVerdcourt, 1971; Verdcourt, 1974; Rudd, 1981a).
Weberbauerella Ulbrich is diagnosed by the combination of its herbaceoushabit, pustular glands densely covering the stems, leaves, and inflorescences(including petals), and leaves with well over 40 leaflets. Similar pustularglands on the petals are known from Poiretia, but this genus is marked byleaves with four leaflets, and a sometimes climbing habit. Weberbauerellacontains two species confined to sand in southern coastal Peru (Ferreyra,1951; Rudd, 1981a).
Pictetia DC. is characterized by spiny stipules, short shoots bearing disti-chously arranged stipules (shared with Ormocarpum, Ormocarpopsis, andPeltiera), coriaceous leaflets that in all but two species have spinescent mu-cros, and pods with two-ribbed placental margins. Pictetia includes eight spe-cies confined to Cuba, Hispaniola, Puerto Rico, and the Virgin Islands ex-cluding St. Croix (Beyra-M. and Lavin, 1999).
Diphysa Jacq. has been characterized by its mature pods that have an exo-carp distinctly inflated and separated from the mesocarp. However, Diphysaormocarpoides and D. spinosa have laterally flattened lomented pods verysimilar to species of Ormocarpum and Pictetia (Antonio and Sousa, 1991).The monophyly of this genus is strongly supported, however, by phylogeneticanalysis of molecular data (see also Beyra-M. and Lavin, 1999; Lavin et al.,2000). The genus includes about ten species centered in Mexico and CentralAmerica (M. Lavin, unpublished data).
Ormocarpum P. Beauv. is diagnosed by most species forming a cylindricalnectary disk surrounding the base of the ovary (M. Thulin and M. Lavin,unpublished data). This trait otherwise is known in a few species of Ma-chaerium and Paramachaerium. This genus of ;20 species is primarily Af-rican. Three species occur on the southern Arabian Peninsula in Yemen (in-cluding Socotra) and Oman, and one to two species occur in tropical Asiaand Australia (Gillett, 1966; Rudd, 1981a; Thulin, 1990). According to ITS/5.8S sequence data (see also Lavin et al., 2000), Ormocarpum comprises twolineages (one with and one without the intrastaminal disk) that are collectivelyparaphyletic with respect to Ormocarpopsis (and Peltiera). This issue is beingaddressed in a separate study (M. Thulin and M. Lavin, unpublished data).
Ormocarpopsis R. Viguier has short shoots with persistent distichously ar-ranged stipules shared with Ormocarpum, Peltiera, and Pictetia. In this con-text, its non-lomented pod with a smooth exocarp (no evidence of prominentparallel nervation on the pod valves) and tannin patches on the abaxial surfaceof dried leaflets are diagnostic. Ormocarpopsis comprises six species endemicto Madagascar (Labat and Du Puy, 1996).
Peltiera Labat & Du Puy includes two endemic Madagascan species thatare sister to Ormocarpopsis (Labat and Du Puy, 1997). These two generashare a distinctive tannin patterning on the abaxial surface of herbarium-driedleaflets where tannin deposits are concentrated along the midrib. Like Or-mocarpopsis, the flowers of Peltiera lack a nectary disk (M. Thulin and M.Lavin, unpublished data), and the pods, though lomented and with all but oneloment aborting, contain spherical seeds. The pod valves in the seed-bearingarticle are dehiscent. Unfortunately, both species of Peltiera are probablyextinct due to the clearing of forests from which they were known.




















