The A17L Gene Product of Vaccinia Virus Is Exposed on the Surface of IMV
10
The A17L Gene Product of Vaccinia Virus Is Exposed on the Surface of IMV Kristina Wallengren,* Cristina Risco,† Jacomine Krijnse-Locker,‡ Mariano Esteban,† ,1 and Dolores Rodriguez† *Department of Biosciences at Novum, Karolinska Institute, S-141 57 Huddinge, Sweden; ‡Cell Biology Program, European Molecular Biology Laboratory, Meyerhofstrasse 1, 69118 Heidelberg, Germany; and †Centro Nacional de Biotecnologı ´a, CSIC, Campus Universidad Auto ´ noma, 28049 Madrid, Spain Received June 4, 2001; returned to author for revision July 12, 2001; accepted August 3, 2001 The p21 membrane protein of vaccinia virus (VV), encoded by the A17L gene, has been reported to localize on the inner of the two membranes of the intracellular mature virus (IMV). It has also been shown that p21 acts as a membrane anchor for the externally located fusion protein p14 (A27L gene). Since p14 is located on the surface of IMVs, it is hard to envision that p21 should be located only on the inner membrane. Our results from (i) immunoelectron microscopy, (ii) biotinylation, and (iii) protease treatment of purified IMVs showed that the N-terminus of p21 is exposed on the surface of virus particles, while the C-terminus is embedded in the membrane. Mono-specific antibodies to the N-terminus of p21 neutralize infection of VV while antibodies to the C-terminal domain do not. We suggest that p21 molecules are located both in the inner and in the outer membrane of IMV. © 2001 Academic Press Key Words: vaccinia virus; IMV membrane protein; neutralizing antibodies. INTRODUCTION Vaccinia virus (VV) is one of the largest and most complex animal viruses with its double-stranded DNA genome encoding about 200 genes. Approximately half of these are structural proteins, which are incorporated into the viral particles, while the rest are required in the complicated replication machinery applied by the virus or modulate, e.g., the immune response to the virus. The virus forms viral factories in the cytoplasm of the infected cell, where replication and assembly occur (reviewed in Moss, 1996). Two main infectious forms of virus are produced during the course of infection: the intracellular mature virus (IMV) and the extracellular enveloped virus (EEV) (Appleyard et al., 1971). The IMV consists of a nucleoprotein complex or core surrounded by an enve- lope. The IMV envelope is thought to derive from mem- branes of the intermediate compartment (ERGIC) be- tween the endoplasmic reticulum and the Golgi complex and thus consist of a double membrane (Sodeik et al., 1993). An alternative hypothesis for the IMV envelope is that it is a single membrane, which is synthesized de novo (Dales and Mosbach, 1968; Hollinshead et al., 1999). These membranes are modified by the insertion of viral proteins to give rise to the rigid, crescent-shaped membranes that assemble around the cytoplasmic viral factory (reviewed in Moss, 1996). During a process that is poorly understood, the membranes engulf electron- dense material from the factory to form spherical imma- ture virions (IVs) (Dales and Pogo, 1981; Dales and Simi- novitch, 1961; Ichihashi et al., 1971), which are enveloped by a double membrane (Risco et al., unpublished data). The spherical IVs then mature into brick-shaped IMVs, through a mostly unknown mechanism that involves pro- teolytic cleavage of VV proteins (Katz and Moss, 1970; Moss and Rosenblum, 1973; Vanslyke et al., 1993). The product of the A17L gene, p21, is essential for virus morphogenesis and constitutes one of the major components of the IMV membrane, together with p15 (A14L) and p8 (A13L) (Salmons et al., 1997). While p21 appears to be responsible for organizing the ERGIC- derived membranes into crescents, p15 seems to be responsible for the assembly of the crescent in close contact with the content of the factory (Rodrı ´guez et al., 1997, 1998; Traktman et al., 2000). p21 is synthesized in the endoplasmic reticulum as a 23-kDa precursor of 203 amino acids (aa). The precursor is proteolytically cleaved at aa 17 (Rodrı ´guez et al., 1993; Whitehead and Hruby, 1994) and has also been sug- gested to be processed in its C-terminus (Betakova et al., 1999b). The protein is phosphorylated (Betakova et al., 1999b; Derrien et al., 1999) and contains potential sites for myristylation. p21 has two long stretches of hydro- phobic aa residues, and it has been demonstrated that both ends of the protein are oriented toward the cyto- plasm (Krijnse-Locker et al., 1996; Betakova et al., 1999a). Two models for the topology of p21 have been sug- gested. One model predicts that p21 spans the mem- brane four times (Krijnse-Locker et al., 1996; Rodrı ´guez et al., 1993), whereas the other model suggests a two- transmembrane conformation (Betakova et al., 1999a). 1 To whom correspondence and reprint requests should be ad- dressed. Fax: 34 91 585 4506. E-mail: [email protected]. Virology 290, 143–152 (2001) doi:10.1006/viro.2001.1131, available online at http://www.idealibrary.com on 0042-6822/01 $35.00 Copyright © 2001 by Academic Press All rights of reproduction in any form reserved. 143
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of The A17L Gene Product of Vaccinia Virus Is Exposed on the Surface of IMV

nlbta1tn1vmfpd
d
Virology 290, 143–152 (2001)doi:10.1006/viro.2001.1131, available online at http://www.idealibrary.com on
The A17L Gene Product of Vaccinia Virus Is Exposed on the Surface of IMV
Kristina Wallengren,* Cristina Risco,† Jacomine Krijnse-Locker,‡ Mariano Esteban,†,1 and Dolores Rodriguez†
*Department of Biosciences at Novum, Karolinska Institute, S-141 57 Huddinge, Sweden; ‡Cell Biology Program, European Molecular BiologyLaboratory, Meyerhofstrasse 1, 69118 Heidelberg, Germany; and †Centro Nacional de Biotecnologıa,
CSIC, Campus Universidad Autonoma, 28049 Madrid, Spain
Received June 4, 2001; returned to author for revision July 12, 2001; accepted August 3, 2001
The p21 membrane protein of vaccinia virus (VV), encoded by the A17L gene, has been reported to localize on the innerof the two membranes of the intracellular mature virus (IMV). It has also been shown that p21 acts as a membrane anchorfor the externally located fusion protein p14 (A27L gene). Since p14 is located on the surface of IMVs, it is hard to envisionthat p21 should be located only on the inner membrane. Our results from (i) immunoelectron microscopy, (ii) biotinylation, and(iii) protease treatment of purified IMVs showed that the N-terminus of p21 is exposed on the surface of virus particles, whilethe C-terminus is embedded in the membrane. Mono-specific antibodies to the N-terminus of p21 neutralize infection of VVwhile antibodies to the C-terminal domain do not. We suggest that p21 molecules are located both in the inner and in the outer
membrane of IMV. © 2001 Academic PressKey Words: vaccinia virus; IMV membrane protein; neutralizing antibodies.
tnbTttM
vc(adrc1
2iWg11fpbpTgb
INTRODUCTION
Vaccinia virus (VV) is one of the largest and mostcomplex animal viruses with its double-stranded DNAgenome encoding about 200 genes. Approximately halfof these are structural proteins, which are incorporatedinto the viral particles, while the rest are required in thecomplicated replication machinery applied by the virusor modulate, e.g., the immune response to the virus. Thevirus forms viral factories in the cytoplasm of the infectedcell, where replication and assembly occur (reviewed inMoss, 1996). Two main infectious forms of virus areproduced during the course of infection: the intracellularmature virus (IMV) and the extracellular enveloped virus(EEV) (Appleyard et al., 1971). The IMV consists of a
ucleoprotein complex or core surrounded by an enve-ope. The IMV envelope is thought to derive from mem-ranes of the intermediate compartment (ERGIC) be-
ween the endoplasmic reticulum and the Golgi complexnd thus consist of a double membrane (Sodeik et al.,993). An alternative hypothesis for the IMV envelope ishat it is a single membrane, which is synthesized deovo (Dales and Mosbach, 1968; Hollinshead et al.,999). These membranes are modified by the insertion ofiral proteins to give rise to the rigid, crescent-shapedembranes that assemble around the cytoplasmic viral
actory (reviewed in Moss, 1996). During a process that isoorly understood, the membranes engulf electron-ense material from the factory to form spherical imma-
at
1 To whom correspondence and reprint requests should be ad-ressed. Fax: 34 91 585 4506. E-mail: [email protected].
143
ure virions (IVs) (Dales and Pogo, 1981; Dales and Simi-ovitch, 1961; Ichihashi et al., 1971), which are envelopedy a double membrane (Risco et al., unpublished data).he spherical IVs then mature into brick-shaped IMVs,
hrough a mostly unknown mechanism that involves pro-eolytic cleavage of VV proteins (Katz and Moss, 1970;
oss and Rosenblum, 1973; Vanslyke et al., 1993).The product of the A17L gene, p21, is essential for
irus morphogenesis and constitutes one of the majoromponents of the IMV membrane, together with p15
A14L) and p8 (A13L) (Salmons et al., 1997). While p21ppears to be responsible for organizing the ERGIC-erived membranes into crescents, p15 seems to be
esponsible for the assembly of the crescent in closeontact with the content of the factory (Rodrıguez et al.,997, 1998; Traktman et al., 2000).
p21 is synthesized in the endoplasmic reticulum as a3-kDa precursor of 203 amino acids (aa). The precursor
s proteolytically cleaved at aa 17 (Rodrıguez et al., 1993;hitehead and Hruby, 1994) and has also been sug-
ested to be processed in its C-terminus (Betakova et al.,999b). The protein is phosphorylated (Betakova et al.,999b; Derrien et al., 1999) and contains potential sites
or myristylation. p21 has two long stretches of hydro-hobic aa residues, and it has been demonstrated thatoth ends of the protein are oriented toward the cyto-lasm (Krijnse-Locker et al., 1996; Betakova et al., 1999a).wo models for the topology of p21 have been sug-ested. One model predicts that p21 spans the mem-rane four times (Krijnse-Locker et al., 1996; Rodrıguez et
l., 1993), whereas the other model suggests a two-ransmembrane conformation (Betakova et al., 1999a).
0042-6822/01 $35.00Copyright © 2001 by Academic PressAll rights of reproduction in any form reserved.

aalmGSagaawps
omrb
ipa
144 WALLENGREN ET AL.
Recent results support the latter model (Betakova andMoss, 2000).
It has previously been shown that p21 forms a complexwith the 14-kDa protein (p14) encoded by the A27L gene(Rodrıguez et al., 1993) and with p15 (A14L) (Betakova et
l., 1999b; Rodrıguez et al., 1997). While p15 behaves asn integral membrane protein, p14 is a peripheral protein
ocated on the surface of IMV and has been postulated toediate fusion (Chung et al., 1998; Doms et al., 1990;ong et al., 1990; Lai et al., 1990; Rodrıguez et al., 1987;odeik et al., 1995; Vazquez and Esteban, 1999). p14 hasstructural organization much like the influenza hemag-
lutinin and gp41 of HIV, except that no transmembranenchor can be predicted from its sequence (Vazquez etl., 1998). Instead, it has a domain that binds to p21,hich thus is believed to serve as membrane anchor for14 (Rodrıguez et al., 1993). Through protein bindingtudies in vitro and in vivo of mutant forms of p14, the p21
binding domain has been predicted to involve a Leuzipper in the third a-helix in the C-terminus of the 14-kDaprotein (Vazquez et al., 1998).
The 14-kDa protein has been localized to the exteriorf IMV particles using immunoelectron microscopy (im-uno-EM) (Sodeik et al., 1995), while p21 has been
eported to localize to the innermost of the two mem-ranes of IV (Krijnse-Locker et al., 1996; Wolfe et al.,
1996). This conclusion is based on results from immu-nolabeled cryosections of infected cells. EM analysis ofIMV has not demonstrated any specific localization forp21 on the inner or the outer membrane (Betakova et al.,1999b). However, the fact that p21 interacts directly withthe externally located p14 suggests that p21 localizationshould be reexamined. Therefore we have studied thelocalization of p21 in IMV by different biochemical ap-proaches and by immunoelectron microscopy using an-tibodies that recognize different domains of the protein.
RESULTS
Characterization of a panel of anti-A17L antibodies
It has previously been shown that the A17L geneproduct is synthesized as a 23-kDa precursor that isproteolytically processed within cells at both the N-(Rodrıguez et al., 1993; Whitehead and Hruby, 1994) andthe C-termini (Betakova et al., 1999b). A set of polyclonalantibodies was generated to peptides corresponding todifferent regions of the A17L gene product. Both purifiedVV and lysate of VV-infected BSC-40 cells (collected at24 h postinfection (hpi)) were analyzed by sodium dode-cyl sulfate-polyacrylamide gel electrophoresis (SDS–PAGE) and Western blot to determine the specificity andpattern of recognition of these antibodies. As shown inFig. 1, all antibodies (aA17L N9, aA17L C9, and aA17L C)were able to identify the 23-kDa precursor in infected
cells (lanes 1, 3, and 5). In virus particles, however, the23-kDa precursor was not detected by any of the anti-bodies (lanes 2, 4, and 6). Antibodies aA17L N9 andaA17L C9 also recognized a protein of 21 kDa believed tobe the mature product of A17L (Betakova et al., 1999b;Rodrıguez et al., 1993; Whitehead and Hruby, 1994) bothn extracts of infected cells (lanes 1 and 3) and in virusarticles (lanes 2 and 4). However, the most C-terminalntibody, aA17L C, was unable to detect the 21-kDa
product, neither in cell lysates nor in virus particles(lanes 5 and 6).
To more thoroughly characterize the products recog-nized by the aA17L antibodies, a pulse–chase and im-munoprecipitation analysis was set up. VV-infected cellswere metabolically labeled with [35S]methionine for 5 minand then chased with an excess of cold methionine for15, 30, 60, and 120 min before collection in lysis buffer.Proteins were immunoprecipitated with aA17L N9, aA17LC9, or aA17L C and separated by SDS–PAGE. The pro-teins were transferred to a nitrocellulose filter, whichwas used for both autoradiography and visualization byECL after antibody recognition. All three antibodies de-tected the 23-kDa precursor, which could be visualizedas a doublet, and showed that this protein decreases inamount with time (Fig. 2). After immunoprecipitation withaA17L N9 and aA17L C9 antibodies, the 21-kDa product
FIG. 1. Processing of VV A17L gene product. Western blot analysis ofextracts from BSC-40 cells infected with VV (10 PFU/cell) for 24 h (C)and of purified virions (V). The proteins were separated by SDS–PAGE(12% polyacrylamide), transferred to nitrocellulose paper, and reactedwith different antibodies to p21: aA17L N9, aA17L C9, or aA17L C.Antibody reactivity was detected using ECL.
could be detected at the 30-min chase point and in-creases in amount with time. At 120 min postsynthesis,
145A17L GENE PRODUCT OF VACCINIA VIRUS
most 23-kDa precursor has been processed. Again,aA17L C is not able to immunoprecipitate p21. We alsoobserved protein bands, which migrated between theprecursor and the mature p21, that may represent cleav-age intermediates.
A protein of approximately 15 kDa was found to co-precipitate with p21, using all of the above antibodies. Itappears in samples from the 30-min chase point andonward. The protein was identified as p15 (A14L geneproduct) by incubating the filter with the correspondingantibody (not shown). This is in agreement with previousresults showing that this interaction occurs even beforeC-terminal truncation of the A17L precursor (Betakova etal., 1999b).
Localization of p21 on the surface of IMVs byimmunoelectron microscopy
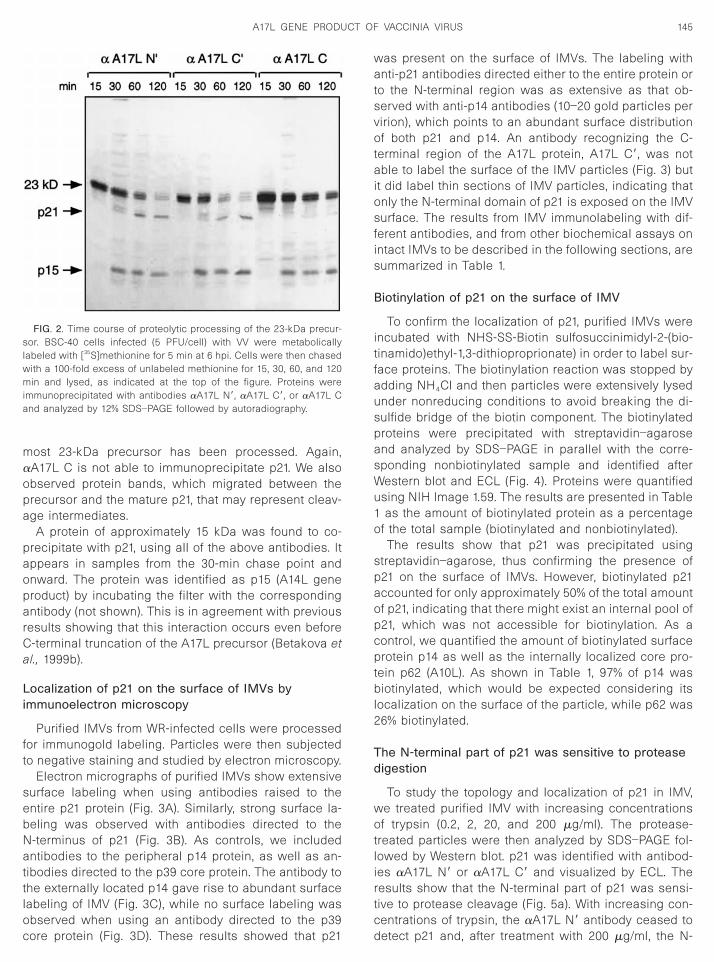
Purified IMVs from WR-infected cells were processedfor immunogold labeling. Particles were then subjectedto negative staining and studied by electron microscopy.
Electron micrographs of purified IMVs show extensivesurface labeling when using antibodies raised to theentire p21 protein (Fig. 3A). Similarly, strong surface la-beling was observed with antibodies directed to theN-terminus of p21 (Fig. 3B). As controls, we includedantibodies to the peripheral p14 protein, as well as an-tibodies directed to the p39 core protein. The antibody tothe externally located p14 gave rise to abundant surfacelabeling of IMV (Fig. 3C), while no surface labeling was
FIG. 2. Time course of proteolytic processing of the 23-kDa precur-sor. BSC-40 cells infected (5 PFU/cell) with VV were metabolicallylabeled with [35S]methionine for 5 min at 6 hpi. Cells were then chasedwith a 100-fold excess of unlabeled methionine for 15, 30, 60, and 120min and lysed, as indicated at the top of the figure. Proteins wereimmunoprecipitated with antibodies aA17L N9, aA17L C9, or aA17L Cand analyzed by 12% SDS–PAGE followed by autoradiography.
observed when using an antibody directed to the p39core protein (Fig. 3D). These results showed that p21
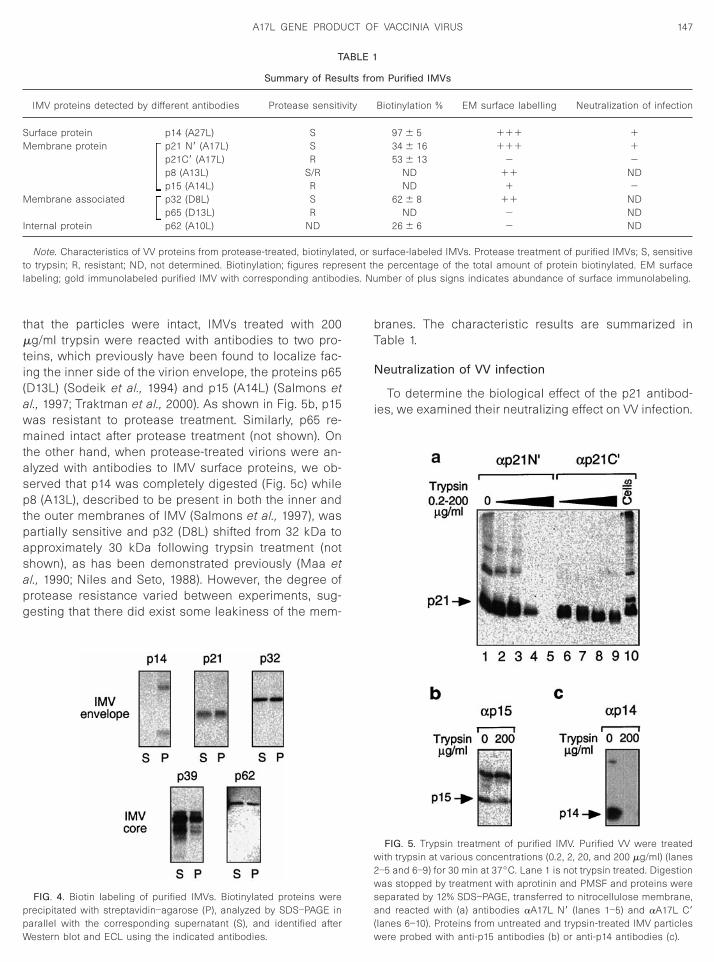
was present on the surface of IMVs. The labeling withanti-p21 antibodies directed either to the entire protein orto the N-terminal region was as extensive as that ob-served with anti-p14 antibodies (10–20 gold particles pervirion), which points to an abundant surface distributionof both p21 and p14. An antibody recognizing the C-terminal region of the A17L protein, A17L C9, was notable to label the surface of the IMV particles (Fig. 3) butit did label thin sections of IMV particles, indicating thatonly the N-terminal domain of p21 is exposed on the IMVsurface. The results from IMV immunolabeling with dif-ferent antibodies, and from other biochemical assays onintact IMVs to be described in the following sections, aresummarized in Table 1.
Biotinylation of p21 on the surface of IMV
To confirm the localization of p21, purified IMVs wereincubated with NHS-SS-Biotin sulfosuccinimidyl-2-(bio-tinamido)ethyl-1,3-dithioproprionate) in order to label sur-face proteins. The biotinylation reaction was stopped byadding NH4Cl and then particles were extensively lysedunder nonreducing conditions to avoid breaking the di-sulfide bridge of the biotin component. The biotinylatedproteins were precipitated with streptavidin–agaroseand analyzed by SDS–PAGE in parallel with the corre-sponding nonbiotinylated sample and identified afterWestern blot and ECL (Fig. 4). Proteins were quantifiedusing NIH Image 1.59. The results are presented in Table1 as the amount of biotinylated protein as a percentageof the total sample (biotinylated and nonbiotinylated).
The results show that p21 was precipitated usingstreptavidin–agarose, thus confirming the presence ofp21 on the surface of IMVs. However, biotinylated p21accounted for only approximately 50% of the total amountof p21, indicating that there might exist an internal pool ofp21, which was not accessible for biotinylation. As acontrol, we quantified the amount of biotinylated surfaceprotein p14 as well as the internally localized core pro-tein p62 (A10L). As shown in Table 1, 97% of p14 wasbiotinylated, which would be expected considering itslocalization on the surface of the particle, while p62 was26% biotinylated.
The N-terminal part of p21 was sensitive to proteasedigestion
To study the topology and localization of p21 in IMV,we treated purified IMV with increasing concentrationsof trypsin (0.2, 2, 20, and 200 mg/ml). The protease-treated particles were then analyzed by SDS–PAGE fol-lowed by Western blot. p21 was identified with antibod-ies aA17L N9 or aA17L C9 and visualized by ECL. Theresults show that the N-terminal part of p21 was sensi-tive to protease cleavage (Fig. 5a). With increasing con-
centrations of trypsin, the aA17L N9 antibody ceased todetect p21 and, after treatment with 200 mg/ml, the N-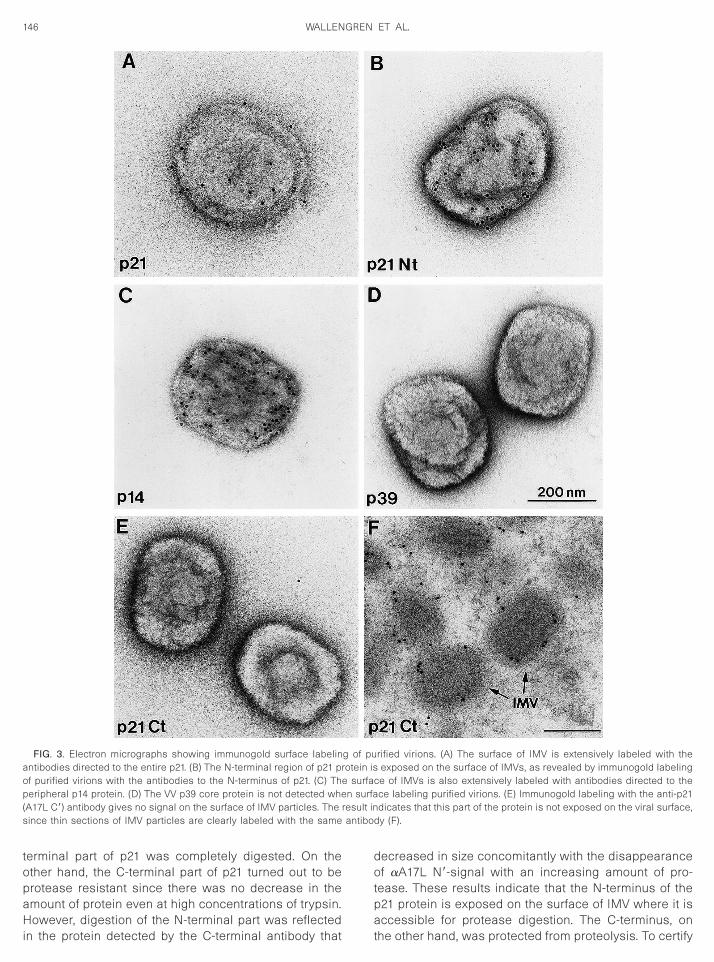
aop( result in
antibo
146 WALLENGREN ET AL.
terminal part of p21 was completely digested. On theother hand, the C-terminal part of p21 turned out to beprotease resistant since there was no decrease in theamount of protein even at high concentrations of trypsin.
FIG. 3. Electron micrographs showing immunogold surface labelinntibodies directed to the entire p21. (B) The N-terminal region of p21 prf purified virions with the antibodies to the N-terminus of p21. (C) Theripheral p14 protein. (D) The VV p39 core protein is not detected whe
A17L C9) antibody gives no signal on the surface of IMV particles. Thesince thin sections of IMV particles are clearly labeled with the same
However, digestion of the N-terminal part was reflectedin the protein detected by the C-terminal antibody that
decreased in size concomitantly with the disappearanceof aA17L N9-signal with an increasing amount of pro-tease. These results indicate that the N-terminus of thep21 protein is exposed on the surface of IMV where it is
rified virions. (A) The surface of IMV is extensively labeled with theexposed on the surface of IMVs, as revealed by immunogold labeling
ce of IMVs is also extensively labeled with antibodies directed to thece labeling purified virions. (E) Immunogold labeling with the anti-p21dicates that this part of the protein is not exposed on the viral surface,
dy (F).
g of puotein is
e surfan surfa
accessible for protease digestion. The C-terminus, onthe other hand, was protected from proteolysis. To certify
SM
sent thies. Nu
ppW
2wsa
147A17L GENE PRODUCT OF VACCINIA VIRUS
that the particles were intact, IMVs treated with 200mg/ml trypsin were reacted with antibodies to two pro-teins, which previously have been found to localize fac-ing the inner side of the virion envelope, the proteins p65(D13L) (Sodeik et al., 1994) and p15 (A14L) (Salmons etal., 1997; Traktman et al., 2000). As shown in Fig. 5b, p15was resistant to protease treatment. Similarly, p65 re-mained intact after protease treatment (not shown). Onthe other hand, when protease-treated virions were an-alyzed with antibodies to IMV surface proteins, we ob-served that p14 was completely digested (Fig. 5c) whilep8 (A13L), described to be present in both the inner andthe outer membranes of IMV (Salmons et al., 1997), waspartially sensitive and p32 (D8L) shifted from 32 kDa toapproximately 30 kDa following trypsin treatment (notshown), as has been demonstrated previously (Maa etal., 1990; Niles and Seto, 1988). However, the degree ofprotease resistance varied between experiments, sug-gesting that there did exist some leakiness of the mem-
T
Summary of Res
IMV proteins detected by different antibodies Protease sensiti
urface protein p14 (A27L) Sembrane protein p21 N9 (A17L) S
p21C9 (A17L) Rp8 (A13L) S/Rp15 (A14L) R
Membrane associated p32 (D8L) Sp65 (D13L) R
Internal protein p62 (A10L) ND
Note. Characteristics of VV proteins from protease-treated, biotinylatto trypsin; R, resistant; ND, not determined. Biotinylation; figures reprelabeling; gold immunolabeled purified IMV with corresponding antibod
FIG. 4. Biotin labeling of purified IMVs. Biotinylated proteins wererecipitated with streptavidin–agarose (P), analyzed by SDS–PAGE in
arallel with the corresponding supernatant (S), and identified afterestern blot and ECL using the indicated antibodies.branes. The characteristic results are summarized inTable 1.
Neutralization of VV infection
To determine the biological effect of the p21 antibod-ies, we examined their neutralizing effect on VV infection.
m Purified IMVs
Biotinylation % EM surface labelling Neutralization of infection
97 6 5 111 134 6 16 111 153 6 13 2 2
ND 11 NDND 1 2
62 6 8 11 NDND 2 ND
26 6 6 2 ND
urface-labeled IMVs. Protease treatment of purified IMVs; S, sensitivee percentage of the total amount of protein biotinylated. EM surfacember of plus signs indicates abundance of surface immunolabeling.
FIG. 5. Trypsin treatment of purified IMV. Purified VV were treatedwith trypsin at various concentrations (0.2, 2, 20, and 200 mg/ml) (lanes
–5 and 6–9) for 30 min at 37°C. Lane 1 is not trypsin treated. Digestionas stopped by treatment with aprotinin and PMSF and proteins were
eparated by 12% SDS–PAGE, transferred to nitrocellulose membrane,nd reacted with (a) antibodies aA17L N9 (lanes 1–5) and aA17L C9
ABLE 1
ults fro
vity
ed, or s
(lanes 6–10). Proteins from untreated and trypsin-treated IMV particleswere probed with anti-p15 antibodies (b) or anti-p14 antibodies (c).
npct(Nrtn
148 WALLENGREN ET AL.
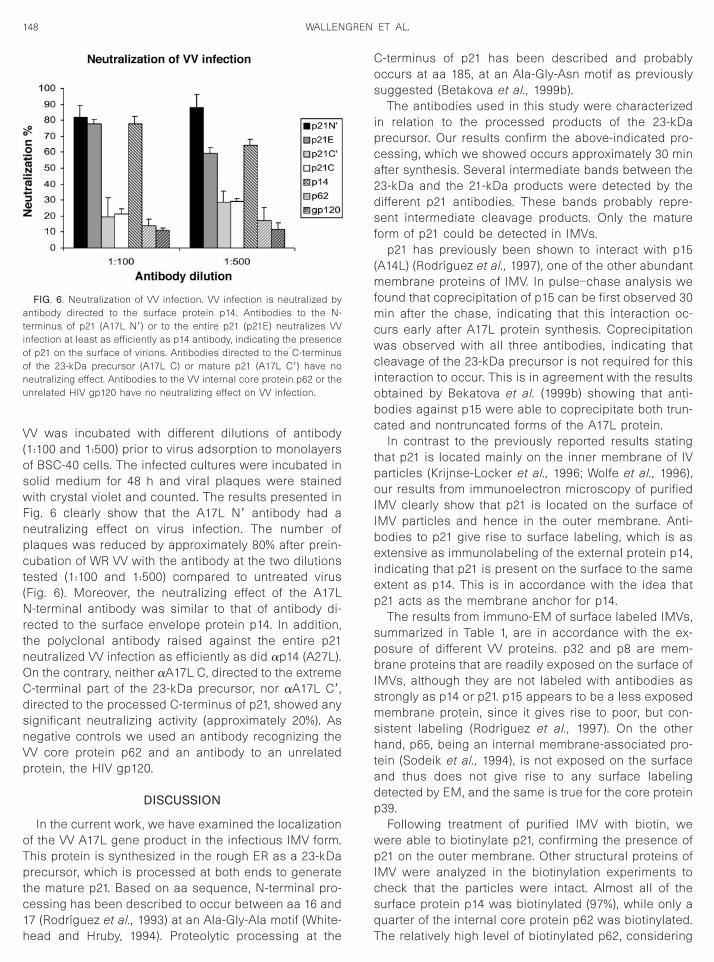
VV was incubated with different dilutions of antibody(1:100 and 1:500) prior to virus adsorption to monolayersof BSC-40 cells. The infected cultures were incubated insolid medium for 48 h and viral plaques were stainedwith crystal violet and counted. The results presented inFig. 6 clearly show that the A17L N9 antibody had a
eutralizing effect on virus infection. The number oflaques was reduced by approximately 80% after prein-ubation of WR VV with the antibody at the two dilutions
ested (1:100 and 1:500) compared to untreated virusFig. 6). Moreover, the neutralizing effect of the A17L
-terminal antibody was similar to that of antibody di-ected to the surface envelope protein p14. In addition,he polyclonal antibody raised against the entire p21eutralized VV infection as efficiently as did ap14 (A27L).
On the contrary, neither aA17L C, directed to the extremeC-terminal part of the 23-kDa precursor, nor aA17L C9,directed to the processed C-terminus of p21, showed anysignificant neutralizing activity (approximately 20%). Asnegative controls we used an antibody recognizing theVV core protein p62 and an antibody to an unrelatedprotein, the HIV gp120.
DISCUSSION
In the current work, we have examined the localizationof the VV A17L gene product in the infectious IMV form.This protein is synthesized in the rough ER as a 23-kDaprecursor, which is processed at both ends to generatethe mature p21. Based on aa sequence, N-terminal pro-cessing has been described to occur between aa 16 and
FIG. 6. Neutralization of VV infection. VV infection is neutralized byantibody directed to the surface protein p14. Antibodies to the N-terminus of p21 (A17L N9) or to the entire p21 (p21E) neutralizes VVinfection at least as efficiently as p14 antibody, indicating the presenceof p21 on the surface of virions. Antibodies directed to the C-terminusof the 23-kDa precursor (A17L C) or mature p21 (A17L C9) have noneutralizing effect. Antibodies to the VV internal core protein p62 or theunrelated HIV gp120 have no neutralizing effect on VV infection.
17 (Rodrıguez et al., 1993) at an Ala-Gly-Ala motif (White-head and Hruby, 1994). Proteolytic processing at the
C-terminus of p21 has been described and probablyoccurs at aa 185, at an Ala-Gly-Asn motif as previouslysuggested (Betakova et al., 1999b).
The antibodies used in this study were characterizedin relation to the processed products of the 23-kDaprecursor. Our results confirm the above-indicated pro-cessing, which we showed occurs approximately 30 minafter synthesis. Several intermediate bands between the23-kDa and the 21-kDa products were detected by thedifferent p21 antibodies. These bands probably repre-sent intermediate cleavage products. Only the matureform of p21 could be detected in IMVs.
p21 has previously been shown to interact with p15(A14L) (Rodrıguez et al., 1997), one of the other abundantmembrane proteins of IMV. In pulse–chase analysis wefound that coprecipitation of p15 can be first observed 30min after the chase, indicating that this interaction oc-curs early after A17L protein synthesis. Coprecipitationwas observed with all three antibodies, indicating thatcleavage of the 23-kDa precursor is not required for thisinteraction to occur. This is in agreement with the resultsobtained by Bekatova et al. (1999b) showing that anti-bodies against p15 were able to coprecipitate both trun-cated and nontruncated forms of the A17L protein.
In contrast to the previously reported results statingthat p21 is located mainly on the inner membrane of IVparticles (Krijnse-Locker et al., 1996; Wolfe et al., 1996),our results from immunoelectron microscopy of purifiedIMV clearly show that p21 is located on the surface ofIMV particles and hence in the outer membrane. Anti-bodies to p21 give rise to surface labeling, which is asextensive as immunolabeling of the external protein p14,indicating that p21 is present on the surface to the sameextent as p14. This is in accordance with the idea thatp21 acts as the membrane anchor for p14.
The results from immuno-EM of surface labeled IMVs,summarized in Table 1, are in accordance with the ex-posure of different VV proteins. p32 and p8 are mem-brane proteins that are readily exposed on the surface ofIMVs, although they are not labeled with antibodies asstrongly as p14 or p21. p15 appears to be a less exposedmembrane protein, since it gives rise to poor, but con-sistent labeling (Rodrıguez et al., 1997). On the otherhand, p65, being an internal membrane-associated pro-tein (Sodeik et al., 1994), is not exposed on the surfaceand thus does not give rise to any surface labelingdetected by EM, and the same is true for the core proteinp39.
Following treatment of purified IMV with biotin, wewere able to biotinylate p21, confirming the presence ofp21 on the outer membrane. Other structural proteins ofIMV were analyzed in the biotinylation experiments tocheck that the particles were intact. Almost all of thesurface protein p14 was biotinylated (97%), while only a
quarter of the internal core protein p62 was biotinylated.The relatively high level of biotinylated p62, considering
149A17L GENE PRODUCT OF VACCINIA VIRUS
that it is a core protein and was not expected to belabeled, suggests that the particles, to some degree,were disrupted. The results from the biotinylation studiesare still indicative of the position of proteins in the par-ticles, since there is a clear gradient of degree of bio-tinylation corresponding to the different localizations ofthe proteins in the particles.
Our results showed that only approximately half of thep21 was biotinylated, which suggests the presence oftwo equivalent pools of p21. Another explanation couldbe that p21 was incompletely biotinylated due to pooraccessibility, indicating that p21 might very well exist in asingle pool on the surface of IMV. However, since previ-ous results have determined the localization of p21 onthe inner membrane of IV (Krijnse-Locker et al., 1996), ourresults should be interpreted as p21 being more or lessevenly distributed on both the inner and the outer mem-branes of IMV. After treatment of purified IMV with tryp-sin, we found that while the C-terminal part of the maturep21 was protease resistant, the N-terminal part wasdigested, which suggests that this domain is exposed onthe surface of IMV. Purified virions used in the experi-ments were shown to be intact by determining proteasesensitivity of other VV proteins. The surface protein p14(A27L) was completely digested, while the more internalproteins p15 (A14L) and p65 (D13L) were protease resis-tant. P8 (A13L) was sensitive, but it was not alwaysdigested, indicating the presence of a bipolar fraction. P8is a membrane protein believed to localize to both theinner and the outer membranes of IMV (Salmons et al.,1997), which is supported by our results.
p21 is thought to span the membrane four times,adopting a topology in which it exposes both termini onthe cytosolic side of the membrane (Krijnse-Locker et al.,1996). In accordance with this model, the N-terminus isexposed on the surface of IMV and hence is accessiblefor protease digestion. Protection of the C-terminus ofp21 from protease digestion could be explained if thisdomain was embedded in the membrane or in closecontact with it or simply does not protrude on the surfaceof the particle. Another possibility could be that a pro-tease-resistant protein masks it. This masking could notoccur through the interaction with p14, since p14 wascompletely digested in the protease assay. Our resultsdemonstrating that the N-terminal end of p21 was com-pletely digested when increased amounts of trypsinwere used indicate that the protease would gain accessto the interior of the particle at high concentrations,which was also demonstrated by the difficulty in achiev-ing consistent results for internal proteins, since theywere more or less protease resistant in different exper-iments. Since we did not find a protease-resistant sub-population of the N-terminus of p21, our results indicatethat p21 exposes its N-terminus in both the outer and the
inner membranes. Thus, p21 would adopt the same, butinverse, topology in both membranes. According to thefour-transmembrane topology, the two transmembranelinking sequences (aa 78–81 and aa 139–141) would beprotected in between the tightly apposed membranes,corresponding to the lumen of ERGIC. The region (aa99–117) separating the second and the third hydrophobicstretches would thus form a linking loop located on thecytosolic side. This region possesses an arginine (aa108), which is susceptible to cleavage by trypsin, butnevertheless it was not affected by protease digestion inour experiments. In the two-transmembrane model ofp21, this arginine-containing region would be protectedfrom protease digestion in between the two membranes.Thus, our results support the latter model (Betakova andMoss, 2000).
All of the above results taken together strongly sug-gest that p21 is located on the outer membrane of IMV,as well. Further support was provided by results ob-tained with antibodies directed to the N-terminus of p21as well as antibodies directed to the entire p21, whichwere capable of neutralizing VV infection. The neutraliz-ing activity of these p21 antibodies was as strong as thatof the antibody directed to p14. Antibodies to the C-terminal part of p21 had no effect on virus infection.
Previous studies of p21 localization were done whenthe proteolytic cleavage of the protein was not wellcharacterized. VV-infected cells were studied in immu-noelectron microscopy using antibodies that were di-rected to both the N- and the C-terminal regions of the23-kDa precursor, which we now know are not present inthe processed protein of the IMV. In these experiments,preferentially the inner, concave membrane of assem-bling viral crescents and IV were immunolabeled, whichled to the conclusion that p21 was located mainly on theinner membrane (Krijnse-Locker et al., 1996; Wolffe et al.,1996). These results could indicate that proteolytic pro-cessing of the 23-kDa precursor is initiated on the exte-rior of IV. This would be in agreement with the resultshowing that processing was not completed until after120 min postsynthesis.
p21 is essential for membrane morphogenesis and inits absence the membranes derived from ERGIC do notform the rigid crescents that line up around the viralfactories (Rodrıguez et al., 1995; Wolfe et al., 1996). Thisindicates that p21 is involved in shaping the membranesas well as being responsible for anchoring p14 to theIMV surface. The possibility also exists that interaction ofp21, localized in the external IMV membrane, with itscounterpart, or the A14L protein, located in the innermembrane, may contribute to bringing these adjacentmembranes together.
In conclusion, the A17L gene product is synthesizedas a 23-kDa precursor, which is processed in both the N-and the C-termini to produce the final 21-kDa form, whichis present in IMV particles. Using protease sensitivity,
surface labeling assays, and neutralization of virus infec-tion, we have shown that p21 is located on the surface of
i
B
Bt
150 WALLENGREN ET AL.
IMV exposing its N-terminus on the exterior of the mem-brane. Hence, our results and previous results takentogether suggest that p21 exists in two pools and is thuslocated on both the outer and the inner membranes ofIMV.
MATERIALS AND METHODS
Cells and virus
African green monkey kidney cells (BSC-40) weregrown in Dulbecco’s modified Eagle’s medium (DMEM)supplemented with 10% newborn calf serum (NCS). TheVV WR strain was propagated and titrated in BSC-40cells as previously described (Joklik, 1962). IMV particleswere purified by banding on a sucrose gradient (Joklik,1962).
Antisera
Several antibodies directed against different regionsof the A17L gene product were raised in rabbits byimmunization with synthetic peptides. The following poly-clonal antibodies were used: aA17L N9, which recog-nizes aa 17–37; aA17L C9 recognizes aa 165–185; aA17LC recognizes aa 193–203 (Rodrıguez et al., 1996); andap21E is raised to the entire p21 (referred to as anti-21-kDa polyclonal antibody in Vazquez et al. (1998)). Theregions of the protein recognized by the different anti-bodies are represented in Fig. 1A. Other antibodies to VVproteins were ap14 (A27L), ap39 (A5L), ap62 (A10L), ap8(A13L), ap15 (A14L), ap65 (D13L), and ap32 (D8L). Anunrelated antibody directed to HIV agp120 was used asa control in the neutralization experiments.
Pulse–chase analysis and immunoprecipitation
BSC-40 cells were infected with VV at 5 PFU per cell.At 5.5 hpi cells were washed and incubated in methi-onine-free DMEM for 30 min. Cells were then pulse-labeled for 5 min with 100 mCi/ml [35S]methionine(Amersham) in the same medium and chased with a100-fold excess of unlabeled methionine for the indi-cated times. Cells were lysed in lysis buffer A (1% Non-idet-P40 (NP-40), 80 mM NaCl, 20 mM Tris–HCl (pH 8.0),20 mM EDTA) and nuclei were removed by 5 min ofcentrifugation at 13,000 rpm in an eppendorf centrifuge.Proteins were immunoprecipitated from the lysate for 1 hat room temperature (RT) or overnight at 4°C, usingindicated antibodies and 50 ml of 50% protein A–Sepha-rose beads (Pharmacia) in 10 mM Tris–HCl (pH 7.5)containing 1% nonfat dry milk. The immunoprecipitateswere washed three times with lysis buffer A and threetimes with PBS and resuspended in 23 sample buffer(1.25 M Tris–HCl, pH 6.8, 4% SDS, 0.25% bromophenolblue, and 10% 2-mercaptoethanol). Samples were boiled
and proteins were resolved by SDS–PAGE and visualizedafter fluorography.Electron microscopy
Purified IMV particles from the WR strain of VV wereadsorbed to the surface of EM grids and subjected toimmunogold labeling. The IMV particles were incubatedwith different antibodies against the A17L gene productand against other VV envelope and core proteins andsubsequently reacted with secondary antibodies conju-gated with 10-nm colloidal gold particles (Risco et al.,1996; Rodrıguez et al., 1997). Negative staining with 2%uranyl acetate was performed as previously described(Rodrıguez et al., 1996). Immunogold labeling on thinsections of VV-infected Hela cells was performed asdescribed (Rodriguez et al., 1997). Samples were studiedn a Jeol 1200EX II electron microscope.
iotinylation of purified virions
Purified virions were treated with 1 mg/ml NHS-SS-iotin (Pierce) for 30 min on ice. The biotinylation reac-
ion was stopped by adding NH4Cl to a concentration of50 mM. Virus particles were subjected to centrifugationfor 30 min at 13,000 rpm in an eppendorf centrifuge at4°C and washed with 50 mM NH4Cl. After a secondcentrifugation they were washed with 1 mM NH4Cl andresubjected to centrifugation. The resulting pellet wasresuspended in lysis buffer B (1% NP-40, 150 mM NaCl,50 mM Tris–HCl (pH 7.6), 2 mM EDTA), incubated for 30min at 37°C, and sonicated twice for 10 s. The lysate wassubjected to centrifugation for 5 min at 13,000 rpm in aneppendorf centrifuge and the resulting pellet was resus-pended in lysis buffer and treated as above. The twosupernatants were pooled and incubated with 50%streptavidin–agarose (Sigma) for 1 h at RT or overnight at4°C. The sample was subjected to centrifugation for 1min at 13,000 rpm in an eppendorf centrifuge. The su-pernatant was recovered and mixed (vol:vol) with 23sample buffer to analyze the material that had not boundto the streptavidin. The streptavidin–agarose pellet con-taining the bound proteins was washed once with buffer1 (50 mM Tris–HCl (pH 8.0), 100 mM NaCl, 1% TX-100,0.1% SDS, 0.2% bovine serum albumin (BSA), 0.2 mMphenylmethylsulfonyl fluoride (PMSF)), three times withbuffer 2 (200 mM Tris–HCl (pH 8.0), 150 mM NaCl, 0.5%TX-100, 0.1% SDS, 0.2% BSA, 0.2 mM PMSF), three timeswith buffer 3 (50 mM Tris–HCl (pH 8.0), 1 M NaCl, 1%TX-100, 0.1% SDS, 0.2% BSA, 0.2 mM PMSF), and twicewith buffer 4 (10 mM Tris–HCl (pH 8.0)) by subjecting thesample to centrifugation for 1 min at 13,000 rpm in aneppendorf centrifuge between each wash. Sample bufferwas added to the streptavidin–agarose and boiled for 5min. Samples were analyzed by SDS–PAGE and Westernblot.
Protease digestion of purified virions
Purified virions were treated with trypsin at variousconcentrations (0.2, 2, 20, and 200 mg/ml) for 30 min at

R
R
R
S
151A17L GENE PRODUCT OF VACCINIA VIRUS
37°C. The trypsin reaction was stopped by adding apro-tinin and PMSF to a concentration of 4 and 100 mg/ml,respectively. Then 23 sample buffer was added andsamples were boiled immediately for 5 min. Sampleswere analyzed by 12% SDS–PAGE and transferred tonitrocellulose membranes. The membranes were incu-bated with specific antibodies and immunoreactivebands were detected by chemiluminescence using ECL(Amersham Pharmacia Biotech).
Neutralization of VV infection
Aliquots of VV containing 100 PFU were incubated withdifferent dilutions (1:100, 1:500) of indicated antibodies inDMEM for 30 min at 37°C before being added to conflu-ent monolayers of BSC-40 cells. After 1 h of virus adsorp-tion, the inoculum was removed and cells were overlaidwith 0.9% agarose in DMEM supplemented with 2% NCS.At 48 hpi, the solid medium was removed and the cellswere stained with 1% crystal violet in 2% methanol.
ACKNOWLEDGMENTS
We thank Victoria Jimenez and Ana Garzon for expert technicalassistance. This investigation was supported by grants from the Co-munidad de Madrid 08.2/0042/2000 to D. Rodrıguez and C. Risco andfrom the European Union (CT98-0225) to M. Esteban.
REFERENCES
Appleyard, G., Hapel, A. J., and Boulter, E. A. (1971). An antigenicdifference between intracellular and extracellular rabbitpox virus.J. Gen. Virol. 13(1), 9–17.
Betakova, T., and Moss, B. (2000). Disulphide bonds and membranetopology of the vaccinia virus A17L envelope protein. J. Virol. 74(5),2438–2442.
Betakova, T., Wolffe, E., and Moss, B. (1999a). Membrane topology ofthe vaccinia virus A17L envelope protein. Virology 261, 347–356.
Betakova, T., Wolffe, E. J., and Moss, B. (1999b). Regulation of vacciniavirus morphogenesis: Phosphorylation of the A14L and A17L mem-brane proteins and C-terminal truncation of the A17L protein aredependent on the F10L kinase. J. Virol. 73, 3534–3543.
Chung, C. S., Hsiao, J. C., Chang, Y. S., and Chang, W. (1998). A27Lprotein mediates vaccinia virus interaction with cell surface heparansulfate. J. Virol. 72(2), 1577–1585.
Dales, S., and Mosbach, E. H. (1968). Vaccinia as a model for mem-brane biogenesis. Virology 35, 564–583.
Dales, S., and Pogo, B. G. T. (1981). Biology of poxviruses. In “VirologyMonographs,” Vol. 18, pp. 54–64. Springer-Verlag, Vienna/New York.
Dales, S., and Siminovitch, L. (1961). The development of vaccinia virusin Earle’s L strain cells as examined by electron microscopy. J. Bio-phys. Biochem. Cytol. 10, 475–503.
Derrien, M., Punjabi, A., Khanna, M., Grubisha, O., and Traktman, P.(1999). Tyrosine phosphorylation of A17 during vaccinia virus infec-tion: Involvement of the H1 phosphatase and the F10 kinase. J. Virol.73(9), 7287–7296.
Doms, R. W., Blumenthal, R., and Moss, B. (1990). Fusion of intra- andextracellular forms of vaccinia virus with the cell membrane. J. Virol.64(10), 4884–4892.
Gong, S. C., Lai, C. F., and Esteban, M. (1990). Vaccinia virus inducescell fusion at acid pH and this activity is mediated by the N-terminus
of the 14-kDa virus envelope protein. Virology 178, 81–91.Hollinshead, M., Vanderplasschen, A., Smith, G. L., and Vaux, D. J.S
(1999). Vaccinia virus intracellular mature virions contain only onelipid membrane. J. Virol. 73(2), 1503–1617.
Ichihashi, Y., Matsumoto, S., and Dales, S. (1971). Biogenesis of poxvi-ruses: Role of A-type inclusions and host cell membranes in virusdissemination. Virology 46, 507–532.
Joklik, W. K. (1962). The preparation and characteristics of highly puri-fied radioactively labelled poxvirus. Biochim. Biophys. Acta 61, 290–301.
Katz, E., and Moss, B. (1970). Formation of a vaccinia virus structuralpolypeptide from a higher molecular weight precursor: Inhibition byrifampicin. Proc. Natl. Acad. Sci. USA 66(3), 677–684.
Krijnse-Locker, J., Schleich, S., Rodrıguez, D., Goud, B., Snijder, E. J., andGriffiths, G. (1996). The role of a 21-kDa viral membrane protein in theassembly of vaccinia virus from the intermediate compartment.J. Biol. Chem. 271(25), 14950–14958.
Lai, C. F., Gong, S. C., and Esteban, M. (1990). Structural and functionalproperties of the 14-kDa envelope protein of vaccinia virus synthe-sized in Escherichia coli. J. Biol. Chem. 265(36), 22174–22180.
Maa, J. S., Rodrıguez, J. F., and Esteban, M. (1990). Structural andfunctional characterization of a cell surface binding protein of vac-cinia virus. J. Biol. Chem. 265(3), 1569–1577.
Moss, B. (1996). Poxviridae: The viruses and their replication. In “FieldsVirology” (B. N. Fields, D. M. Knipe, P. M. Howley, et al., Eds.), 3rd ed.,pp. 2637–2671. Raven Press, Philadelphia.
Moss, B., and Rosenblum, E. N. (1973). Protein cleavage and poxvirusmorphogenesis: Tryptic peptide analysis of core precursors accu-mulated by blocking assembly with rifampicin. J. Mol. Biol. 81, 267–269.
Niles, E. G., and Seto, J. (1988). Vaccinia virus gene D8 encodes a viriontransmembrane protein. J. Virol. 62(10), 3772–3778.
Risco, C., Anton, I. M., Enjuanes, L., and Carrascosa, J. L. (1996). Thetransmissible gastroenteritis coronavirus contains a spherical coreshell consisting of M and N proteins. J. Virol. 70, 4773–4777.
Rodrıguez, D., Esteban, M., and Rodrıguez, J. R. (1995). Vaccinia virusA17L gene product is essential for an early step in virion morpho-genesis. J. Virol. 69, 4640–4648.
odrıguez, D., Risco, C., Rodrıguez, J. R., Carrascosa, J. L., and Esteban,M. (1996). Inducible expression of the vaccinia virus A17L geneprovides a synchronized system to monitor sorting of viral proteinsduring morphogenesis. J. Virol. 70(11), 7641–7653.
odrıguez, D., Rodrıguez, J., and Esteban, M. (1993). The vaccinia virus14-kilodalton fusion protein forms a stable complex with the pro-cessed protein encoded by the vaccinia virus A17L gene. J. Virol.67(6), 3435–3440.
odrıguez, J. F., Paez, E., and Esteban, M. (1987). A 14,000-M r envelopeprotein of vaccinia virus is involved in cell fusion and forms co-valently linked trimers. J. Virol. 61(2), 395–404.
Rodrıguez, J. R., Risco, C., Carrascosa, J. L., Esteban, M., and Rodrıguez,D. (1997). Characterization of early stages in vaccinia virus mem-brane biogenesis: Implications of the 21-kilodalton protein and anewly identified 15-kilodalton envelope protein. J. Virol. 71, 1821–1833.
Rodrıguez, J. R., Risco, C., Carrascosa, J. L., Esteban, M., and Rodrıguez,D. (1998). Vaccinia virus 15-kilodalton (A14L) protein is essential forassembly and attachment of viral crescents to virosomes. J. Virol.72(2), 1287–1296.
Salmons, T., Kuhn, A., Wylie, F., Schleich, S., Rodrıguez, J. R., Rodrıguez,D., Esteban, M., Griffiths, G., and Krijnse-Locker, J. (1997). Vacciniavirus membrane proteins p8 and p16 are cotranslationally insertedinto the rough endoplasmic reticulum and retained in the intermedi-ate compartment. J. Virol. 71, 7404–7420.
odeik, B., Cudmore, S., Ericsson, M., Esteban, M., Niles, E. G., andGriffiths, G. (1995). Assembly of vaccinia virus: Incorporation of p14and p32 into the membrane of the intracellular mature virus. J. Virol.69(6), 3560–3574.
odeik, B., Doms, R. W., Ericsson, M., Hiller, G., Machamer, C. E., van’tHof, W., van Meer, G., Moss, B., and Griffiths, G. (1993). Assembly of

W
W
152 WALLENGREN ET AL.
vaccinia virus: Role of the intermediate compartment between theendoplasmic reticulum and the Golgi stacks. J. Cell Biol. 121, 521–541.
Sodeik, B., Griffiths, G., Ericsson, M., Moss, B., and Doms, R. W. (1994).Assembly of vaccinia virus: Effects of rifampin on the intracellulardistribution of viral protein p65. J. Virol. 68(2), 1103–1114.
Traktman, P., Liu, K., DeMasi, J., Rollins, R., Jesty, S., and Unger, B.(2000). Elucidating the essential role of the A14 phosphoprotein invaccinia virus morphogenesis: Construction and characterization ofa tetracycline-inducible recombinant. J. Virol. 74(8), 3682–3695.
Vanslyke, J. K., Lee, P., Wilson, E. M., and Hruby, D. E. (1993). Isolation
and analysis of vaccinia virus previrions. Virus Genes 7(4), 311–324.Vazquez, M.-I., and Esteban, M. (1999). Identification of functional do-
mains in the 14-kilodalton envelope protein (A27L) of vaccinia virus.J. Virol. 73(11), 9098–9109.
Vazquez, M.-I., Rivas, G., Cregut, D., Serrano, L., and Esteban, M. (1998).The vaccinia virus 14-kilodalton (A27L) fusion protein forms a triplecoiled-coil structure and interacts with the 21-k (A17L) virus mem-brane protein through a C-terminal a-helix. J. Virol. 72, 10126–10137.hitehead, S. S., and Hruby, D. E. (1994). Differential utilization of aconserved motif for the proteolytic maturation of vaccinia virus pro-teins. Virology 200, 154–161.
olffe, E. J., Moore, D. M., Peters, P. J., and Moss, B. (1996). Vacciniavirus A17L open reading frame encodes an essential component of
nascent viral membranes that is required to initiate morphogenesis.J. Virol. 70(5), 2797–2808.