
Terrestrial adaptations in the postcranial skeletons of guenons
31
AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 93:341-371 (1994) Terrestrial Adaptations in the Postcranial Skeletons of Guenons DANIEL L. GEBO AND ERIC J. SARGIS Department of Anthropology, Northern Illinois University, DeKalb, Illinois 60115 KEY WORDS Limb morphology, Guenon evolution, Kibale Forest ABSTRACT Arboreal and semiterrestrial guenons show similar osteolog- ical features of the limbs across a wide range of species, environments, and geography, while the more terrestrially committed guenons exhibit greater morphological divergence. An ecomorphological comparison of two sympatric guenons living in Kibale Forest, Uganda, reveals an array of anatomical adaptations for terrestriality in the limbs of Cercopithecus lhoesti similar to those found in Erythrocebus patas. In contrast, Cercopithecus aethiops, al- though also frequent users of the terrestrial environment, generally exhibit fewer morphological adaptations characteristic of a terrestrial lifestyle. It appears that significant morphological modification for terrestriality has oc- curred twice within the diverse radiation of living guenons with C. aethiops perhaps representing a third group in the making. o 1994 Wiley-Liss, Inc. INTRODUCTION Although several investigations have fo- cused on gait analysis in guenons (e.g., Roll- inson and Martin, 1981; Hurov, 1985,1987; Vilensky et al., 1988, 1990; Vilensky and Gankiewicz, 1989, 1990a,b; Meldrum, 19911, field studies on positional behavior in this group of Old World monkeys have rarely been conducted. This lack of ecobe- havioral information has hindered work on form-function relations and has limited our understanding of the number of times a ter- restrial habitus has been adopted in this diverse radiation of Old World monkeys, and the variety of morphological adapta- tions to this habitus. Several previous studies on locomotor ad- aptation have succeeded in documenting os- teological differences in closely related spe- cies, and relating differences in skeletal morphology with particular movements and habitat preferences (Fleagle, 1976; Rodman, 1979; Fleagle and Meldrum, 1988; Mel- drum, 1989).This within-genus strategy has been adopted here since it offers an isolated comparative sample where environmental and phylogenetic differences can be mini- mized (particularly between sympatric spe- cies), or held essentially constant. In theory, this allows morphologists to better correlate differences in behavior with differences in morphology. This paper examines limb anatomy in arboreal, semiterrestrial, and terrestrial species of guenons. It highlights by comparison Cercopithecus mitis, an arbo- real guenon, with Cercopithecus aethiops, an open woodland terrestrial guenon, Cercop- ithecus lhoesti, a montane terrestrial gue- non, and Erythrocebus patas, a “savanna adapted” terrestrial guenon. All species of guenons utilize the arboreal environment, at least occasionally, to feed, rest or sleep. But guenons can be subdivided into three categories according to their use of this substrate. The bulk of the guenon species is highly arboreal with only occa- sional use of the forest floor. Guenons such as Cercopithecus diana, Cercopithecus mitis, and Cercopithecus ascanius are representa- tive of this diverse group. In the second cat- egory, a few guenon species like Cercopithe- cus neglectus, Cercopithecus hamlyni, and Allenopithecus nigrouiridis are more fre- Reeeived June 24,1992; accepted August 10,1993. 0 1994 WILEY-LISS, INC.
Transcript of Terrestrial adaptations in the postcranial skeletons of guenons

AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 93:341-371 (1994)
Terrestrial Adaptations in the Postcranial Skeletons of Guenons DANIEL L. GEBO AND ERIC J. SARGIS Department of Anthropology, Northern Illinois University, DeKalb, Illinois 60115
KEY WORDS Limb morphology, Guenon evolution, Kibale Forest
ABSTRACT Arboreal and semiterrestrial guenons show similar osteolog- ical features of the limbs across a wide range of species, environments, and geography, while the more terrestrially committed guenons exhibit greater morphological divergence. An ecomorphological comparison of two sympatric guenons living in Kibale Forest, Uganda, reveals an array of anatomical adaptations for terrestriality in the limbs of Cercopithecus lhoesti similar to those found in Erythrocebus patas. In contrast, Cercopithecus aethiops, al- though also frequent users of the terrestrial environment, generally exhibit fewer morphological adaptations characteristic of a terrestrial lifestyle. It appears that significant morphological modification for terrestriality has oc- curred twice within the diverse radiation of living guenons with C. aethiops perhaps representing a third group in the making. o 1994 Wiley-Liss, Inc.
INTRODUCTION
Although several investigations have fo- cused on gait analysis in guenons (e.g., Roll- inson and Martin, 1981; Hurov, 1985,1987; Vilensky et al., 1988, 1990; Vilensky and Gankiewicz, 1989, 1990a,b; Meldrum, 19911, field studies on positional behavior in this group of Old World monkeys have rarely been conducted. This lack of ecobe- havioral information has hindered work on form-function relations and has limited our understanding of the number of times a ter- restrial habitus has been adopted in this diverse radiation of Old World monkeys, and the variety of morphological adapta- tions to this habitus.
Several previous studies on locomotor ad- aptation have succeeded in documenting os- teological differences in closely related spe- cies, and relating differences in skeletal morphology with particular movements and habitat preferences (Fleagle, 1976; Rodman, 1979; Fleagle and Meldrum, 1988; Mel- drum, 1989). This within-genus strategy has been adopted here since it offers an isolated comparative sample where environmental and phylogenetic differences can be mini- mized (particularly between sympatric spe-
cies), or held essentially constant. In theory, this allows morphologists to better correlate differences in behavior with differences in morphology. This paper examines limb anatomy in arboreal, semiterrestrial, and terrestrial species of guenons. It highlights by comparison Cercopithecus mitis, an arbo- real guenon, with Cercopithecus aethiops, an open woodland terrestrial guenon, Cercop- ithecus lhoesti, a montane terrestrial gue- non, and Erythrocebus patas, a “savanna adapted” terrestrial guenon.
All species of guenons utilize the arboreal environment, at least occasionally, to feed, rest or sleep. But guenons can be subdivided into three categories according to their use of this substrate. The bulk of the guenon species is highly arboreal with only occa- sional use of the forest floor. Guenons such as Cercopithecus diana, Cercopithecus mitis, and Cercopithecus ascanius are representa- tive of this diverse group. In the second cat- egory, a few guenon species like Cercopithe- cus neglectus, Cercopithecus hamlyni, and Allenopithecus nigrouiridis are more fre-
Reeeived June 24,1992; accepted August 10,1993.
0 1994 WILEY-LISS, INC.

342 D.L. GEBO AND E.J. SARGIS
quent users of the terrestrial environment but are not habitually committed to life on the ground. Lastly, guenons such as the “sa- vanna-adapted Erythrocebus patas, the wide ranging Cercopithecus aethiops, and the “montane adapted” species of the Lhoesti group (Cercopithecus lhoesti, Cercopithecus preussi, and Cercopithecus solatus) are ha- bitual users of the terrestrial substrate.
The best study to date on guenon locomo- tion is by Rose (1979), who studied the posi- tional behavior of Cercopithecus aethiops in Kenya. He found that vervets locomote for 4.5% of their total daily time budget and only 20% of their locomotor time is spent on the ground. This implies that Cercopithecus aethiops, a relatively terrestrially oriented guenon compared to others, still moves most often in the arboreal environment. Cheney and Seyfarth (1990, p. 19), however, note that vervets “spend roughly equal amounts of time during daylight hours on the ground and in the trees.” Similarly, Tappen (1960, p. 96) noted that vervets “generally feed in trees, but may travel extensively in’ open country t o reach other woodlands.” Com- pared to patas monkeys, vervets specialize in exploiting more continuous rather than open woodlands such as along rivers (Chism and Rowell, 1988). Vervets, when moving, prefer to quadrupedally walWrun (53.9% of the time), climb (29.5%), and leap (9.6%), with 7% of time related to other locomotor activities (Rose, 1979). When in trees, vervets prefer to move on horizontal (56%) and sloping supports (41.6%) with vertical supports rarely being utilized (2.4%) (Rose, 1979). Leaps analyzed from takeoff show a similar pattern of support orientation use with medium-sized supports being pre- ferred. Vervets show a distinct bias for hori- zontally oriented supports (71%) while rest- ing (Rose, 1979). When moving on the ground, vervets spend 95% of their time walking or running quadrupedally (Rose, 1979).
Recently, Gebo and Chapman (1993a) have recorded quantitative observations on positional behavior on two arboreal guenons (Cercopithecus ascanius and Cercopithecus mitis) inhabiting Kibale Forest in Uganda. Cercopithecus ascanius’s overall locomotor profile shows that red-tailed monkeys prefer
to move most frequently by climbing (43%) and arboreal quadrupedalism (39%) (Table 1) and that they prefer to use the smallest diameter supports (<5 cm in circumfer- ence). Red-tailed monkeys utilize oblique and vertical supports more frequently and bound up vertical supports more often than do blue monkeys (Table 1). Although red- tailed monkeys generally prefer the middle canopy in Kibale Forest, they do frequent the lower canopy more often than do blue monkeys (Struhsaker, 1978; Thomas, 1991), although others have reported more fre- quent lower canopy use by blue monkeys (e.g., Schlichte, 1978). Both red-tailed and blue monkeys have very rarely been ob- served on the ground at Kibale Forest.
Cercopithecus mitis prefers to move using arboreal quadrupedalism (54%), and prefers larger and more horizontally oriented sup- ports relative to red-tailed monkeys (Table 1). Both red-tailed and blue monkeys leap with a similar style and across similar hori- zontal distances (see Gebo and Chapman, 1993a). When leaping, blue monkeys tend to fall shorter vertical distances downward than do red-tailed monkeys.
Semiterrestrial guenons utilize the ground more frequently than the more strictly arboreal guenons (e.g., Cercopithe- cus mitis and Cercopithecus ascanius) and include species such as Cercopithecus ne- glectus, Cercopithecus hamlyni, and Alleno- pithecus nigroviridis. Unfortunately, little is known concerning the positional behavior of these three semiterrestrial species.
Cercopithecus neglectus is known to fre- quently feed on the ground and to be a fast and an adept runner on the ground as well (Kingdon, 1974). Similarly, Gautier et al. (1980) record this species as semiterrestrial. Cercopithecus neglectus uses the lower for- est strata most often, 62% of the time, the middle strata 31%, and the high forest strata 7% (Gautier-Hion, 1988). De Brazza monkeys are noted to spend up to 70% of their time in forest levels under five meters and 20% of their time on the ground (Gau- tier-Hion, 1988). “When a predator is de- tected, monkeys flee silently onto the ground or alternatively freeze on a branch for up to five hours’’ (Gautier-Hion, 1988, p. 456). These monkeys tend to avoid leaping

343 LIMB MORPHOLOGY IN GUENONS
TABLE 1. Locomotor profile and habitat use for adult guenons at Kibale Forest, Uganda‘
Locomotor profile”
BW F/M Bouts Q (%I L (%I CL (%) Other (%)
Overall (Travel + Feeding combined) Cercopithecus ascanius 3.0/4.0 3,653 39 15 43 3 Cercopithecus mitis 3.516.0 3,413 54 11 35 <1 Cercopithecus lhoesti 3.0/6.0 + + + ~
Cercopithecus mitis 1,356 51 18 29 2
~
Travel Only Cercopithecus ascanius 1,121 42 25 27 6
Cercopithecus lhoesti - + - - -
Habitat Use” Support angle Support size - Canopy use
N UC(%) MC(%) LC(%) H(%) O(%) V(%) L(%) M(%) s(%) Cercopithecus ascanius
Q 1,421 24 64 12 64 36 0 12 64 24 L 548 20 70 11 46 52 2 9 63 28 CL 1,560 30 60 10 46 51 3 4 42 54
Q 1,833 37 57 6 80 21 0 15 64 21 Cercopithecus mitis
L 360 19 72 9 64 35 1 10 75 15 CL 1,178 26 65 9 28 69 3 11 57 31
’ Cercopithecus ascanius and Cercopithecus mitis are arboreal guenons and rarely come to the ground whereas Cercopithecus lhoesti is essentially a terrestrial guenon. In the Overall Profile, C. ascartius is a more frequent user of climhing (+8%) compared to C. mitis, which prefers quadrupedalism (+ 15%). Leaping and other differ by less than 5%. For other, all movements are rare except for vertical hounding in C. ascanius where i t represents almost 36 of the locomotor profile. During Travel, C. ascanius climbs about the same as, leaps more (+7%), and is a less frequent user of quadrupedalism (-9%) relative to C. mitis. Observations on Cercopithecus lhoesti are haphazard and nonquantifiahle. This species has been observed in the lower and middle canopy, on medium and small supports, and on oblique and horizontal branches. Cercopithecus lhoesti is usually observed on the ground either quadrupedally standing, walking, or running. Males will occasionally stand bipedally and peer about. ‘BW = body weights by females (F) and males (MI from Waser (1986); Bouts = locomotor bouts; Q = quadrupedalism; L = leaping; CL = climhing; Other = includes quadrupedal suspensory movements, bridging, himanual pull-ups, bipedalism, and vertical hounding. + = ob- served in Cercopithecus Ehoesti. ‘N = number of observations; UC = upper canopy; MC = middle canopy; LC = lower canopy; H = horizontal support; 0 = oblique support; V = vertical support; L = large diameter support; M = medium diameter support; S = small diameter support (see Gebo, 1992).
so that they may walk slowly and silently (Gautier-Hion, 1978). Gautier-Hion (1978) notes that the day range for De Brazza mon- keys is very small a t only 400-500 m.
Cercopithecus hamlyni is considered to be terrestrial by Cords (1987) and Thomas (19911, and Rahm (1970) also notes that it is often found on the ground. Little else has been reported.
Allenopithecus nigrouiridis is found in swamp forests (lower strata levels) and on the ground (Gautier-Hion, 1988). Gautier (1985) describes this species as semiterres- trial and occupying the lowest forest strata (<5 m). According to Gautier-Hion (1988), its antipredatory behavior for escape is by the ground.
The most terrestrially oriented guenons include vervets (as described above), patas monkeys, and the lhoesti monkey group, Cercopithecus lhoesti, Cercopithecus preussi, and Cercopithecus solatus.
Locomotor adaptations in patas monkeys have been viewed as specializations to a mainly terrestrial way of life (Hall, 1965). Both Struhsaker (1969, p. 82) and Chism and Rowell (1988) note that patas monkeys are “even more independent of trees than Cercopithecus aethiops.” However, Chism and Rowell (1988) have demonstrated that patas are not a species particularly adapted to open grassland conditions, but are more accurately viewed as primarily a woodland species that spends much of its time in or near trees (see also Hall, 1965; Hall and Gartlan, 1965; Rowell, 1966; Struhsaker, 1967). Part of Chism and Rowell’s evidence for this view is that patas monkeys strongly prefer open acacia woodland and only make brief forays into the grassland to feed (Chism and Rowell, 1988). Similarly, di- etary preferences show that patas rarely eat any part of grass plants and they seem to be specialists for patchy woodlands with adja-

344 D.L. GEBO AND E.J. SARGIS
cent open areas (Chism and Rowell, 1988). Nakagawa (1989) noted that patas spend 59.6% of their time on the ground (versus 40.4% in trees) and that 90.5% of all feeding occurs on the ground. Hall (1965) similarly noted that less than 25% of feeding takes place in trees. Further, although fast for monkeys, patas could not outrun a cheetah and thus survival depends on vigilance, crypticity, and rapid flight to avoid preda- tors (Chism and Rowell, 1988). On the other hand, patas do possess very large home ranges which are similar in size to baboons and much larger than those of vervets (Chism and Rowell, 1988). Chism and Row- ell (1988) further believe that the cursorial adaptations in patas are linked to the long day ranges for efficient exploitation of the habitat rather than merely high speed es- cape from predators.
In terms of locomotion, patas will cross open grasslands and move from place to place on the ground most of the time (Chism and Rowell, 1988). On rare occasions patas will travel by leaping from tree to tree just as vervets do (Chism and Rowell, 1988). Pa- tas feed in tree crowns and will stand bipe- dally on the ground or cling to a tree trunk to gnaw on acacia gum (Chism and Rowell, 1988). Patas monkeys always sleep in scat- tered trees over a wide distance at night (Chism and Rowell, 1988; Nakagawa, 1989). Nakagawa (1989) notes that patas will climb tall trees and will spend 18-23% of the day moving. When moving on the ground, patas move quietly and vocalize infre- quently like De Brazza monkeys (Gautier- Hion and Gautier, 1978). Kingdon (1974) notes that patas monkeys have been clocked at 55 km/hour when running and that patas tend to flee from danger by scattering and running away on the ground rather than climbing trees.
Members of the lhoesti monkey group, Cercopithecus lhoesti, Cercopithecus preussi, and Cercopithecus solatus, also spend a great amount of time on the ground and are mainly observed within the lowest five meters of the forest (Gautier-Hion, 1988). Struhsaker (1969) has observed both Cerco- pithecus lhoesti and Cercopithecus preussi and noted their preference for terrestriality. Struhsaker (1969) stated that Cercopithecus
preussi had a distinct preference for the lower forest levels and was commonly seen on the ground in contrast to other guenons he studied in West Cameroon and East Af- rica. Cercopithecus preussi “invariably ran along the ground when fleeing from man and was often seen foraging on the ground. None of the other forest Cercopithecus fled on the ground.. . ” (Struhsaker, 1969, p. 100). Similarly, Gartlan and Struhsaker (1972) report Cercopithecus preussi almost always below 9 m in height and commonly on the ground. They note that Cercopithecus preussi “usually fled on the ground, whereas the other species typically fled in the trees” (Gartlan and Struhsaker, 1972, p. 258). Thus, antipredatory escape for Cercopithe- cus preussi is by the ground (Gautier-Hion, 1988). Cercopithecus preussi is reported to spend 35% of its time on the ground. (Gau- tier-Hion, 1988). In trees, Cercopithecus preussi spends 85% in the lower forest lev- els, 15% in the middle, and 5% in the high strata levels (Gautier-Hion, 1988). Cerco- pithecus preussi is strictly confined to the montane forests in Cameroon (Sanderson, 1940; Kmgdon, 1974).
Cercopithecus solatus was described in 1988 by Harrison where it is reported to be terrestrial by local hunters. Gautier-Hion (1988) reports this species to spend 40% of its time on the ground. When in the trees, 85% is spent in the lowest strata levels ( < l o m) with 20% in the middle levels (10-20 m) (Gautier-Hion, 1988). Antipredatory behav- ior escape is noted to be by the ground by Gautier-Hion (1988). Gautier et al. (1992) discuss the geographic distribution, habitat characteristics, and community structure for this rare and quiet semiterrestrial forest monkey.
Cercopithecus lhoesti is a fast moving ter- restrial guenon (Haddow, 1952; DeVore, 1963; Hill, 1966; Napier and Napier, 1967; Kingdon, 1974; Waser, 1980; Leland, 1987; Gautier-Hion, 1988; Fleagle, 1988; Thomas, 1991; DLG, personal observation) found in the montane forests of Uganda, Rwanda, Burundi, and eastern Zaire with its range extending into the lowland rain forests of Zaire (Haddow, 1952; Tappen, 1960; Hill, 1966; Kingdon, 1974; Lernould, 1988; Co- lyn, 1988). Cercopithecus lhoesti has been

345 LIMB MORPHOLOGY IN GUENONS
observed to commonly feed on the ground (Kingdon, 1974) but it does sleep in trees (Kingdon, 1974). Cercopithecus lhoesti “is very alert to danger and individuals con- stantly climb vantage points to peer around, this behavior may be a response to the greater risks involved in ground dwelling” (Kingdon, 1974, p. 232). Kingdon (1974, p. 231), crediting M~ller, further notes that Cercopithecus lhoesti “are more mobile and adventurous than the more strictly arboreal monkeys.” However, Cercopithecus lhoesti has been observed by Kingdon (1974) to use all levels of the canopy when feeding or rest- ing.
At Kibale, l’Hoest’s monkeys are occasion- ally observed in the forest but normally are very secretive and nonhabituated, and gen- erally flee from observers at a brisk pace and on the ground (DLG, personal observa- tion). As noted by Kingdon (1974), males are very wary and will stand bipedally and peer about being a sentry as the other monkeys run away on the ground. This contrasts with travel and fleeing responses in the other two guenons at Kibale (Cercopithecus ascanius and Cercopithecus mitis), which travel pre- dominately in trees and use trees to flee from observers. These guenons are rarely observed on the ground.
MATERIALS AND METHODS Skeletons of guenons (18 species, Table 2)
were examined at the Field Museum of Nat- ural History (Chicago, IL), the American Museum of Natural History (New York, Ny), the Museum of Comparative Zoology (Cambridge, MA), University of Wisconsin- Milwaukee (Milwaukee, WI), United States National Museum-Smithsonian (Washing- ton, DC), and at Northern Illinois Univer- sity (DeKalb, IL). One skeleton of Cerco- pithecus lhoesti, KF’P #13, an adult female, was borrowed from the skeletal collection housed at the Kibale Field Station with per- mission from the Makerere University Bio- logical Field Station (Drs. Andrew Johns and G.I. Basuta, codirectors). This female 1’Hoest’s monkey was collected by Drs. Rich- ard Wrangham and Liz Rogers in 1989 as a fresh carcass after it was killed by a crowned hawk-eagle.
TABLE 2. Number of guenon specimens examined
Species Males Females Total
Allenopithecus nigroviridis Cercopithecus aethiops Cercopithecus albogularis Cercopithecus ascanius Cercopithecus campbelli Cercopithecus cephus Cercopithecus diana Cercopithecus hanlyni Cercopithecus lhoesti Cercopithecus mitis Cercopithecus mona Cercopithecus nictitans Cercopithecus neglectus Cercopithecus petaurzsta Cercopithecus pogonias Cercopithecus preussi Cercopithecus talapoin Erythrocebus patas Total
1 6 1 7 1 3 3 1 0 7 1 1 3 1 0 1 1 4
1 2 5 11 1 2 3 10 1 2 1 4 3 6 0 1 1 1 7 14 0 1 0 1 3 6 0 1 1 1 1 2 4 5 2 6
76
All of the methods for the observations on guenon locomotion and habitat use (Table 1) are described in detail in Gebo and Chap- man (1993a). All variables such as locomotor categories, support sizes and orientations, and canopy height are defined in Gebo (1992).
Tables 3 and 4 list standard osteological measurements and indices for a number of guenon species. T-tests were performed on these indices to determine statistical signif- icance at the 0.05 level using Cercopithecus mitis as a standard of comparison, as well as testing the semiterrestrial and terrestrial guenons separately. Table 5 summarizes these statistical tests by index. Further, we add a cautionary note in that we did not, in our interpretation of these indices, compen- sate for the effects of body size (see Albrecht et al., 1993).
Due to inadequate samples for many of the taxa, detailed qualitative observations on a variety of osteological characters specif- ically relating to joint anatomy and joint an- gles were also evaluated across a broader range of guenon species (Table 6) . These os- teological characters were selected on the basis of what we perceived to be structures that might distinguish terrestrial and arbo- real taxa.
All of the CT scans were taken by Dr. Christopher Ruff (Johns Hopkins Univer- sity) and all calculations resulting from these images were performed in his lab and

GU
EN
ON
SP
EC
IES
(N)
I I
I I
I I
ME
AN
] S
D
MEA
N1
SD
M
EA
N/
SD
M
EA
N/
SD
M
EA
N]
SD
M
EAN
S
D
ME
AN
] S
D
I I
I I
I I
aeth
lops
(11)
_a
scan
lus (
10)
ceph
us (4
) dl
ana
(6)
ham
lyni
(1)
lho
estl
(1)
Ihoa
stU
pmus
si (3
) m
ltls
(14)
m
ona
(1)
negl
ectu
s (6
)
Tab
le 3

TAB
LE 3
. M
EA
NS
. I
WE
NO
N S
PEC
IES
(N)
lnrc
tftan
s (I)
nigm
vhid
is (2
) pa
tas (
6)
I M
EAN
S
D
MEA
N
Tabl
e 3.
(C
ontin
ued)
.
I I
pogo
nias
(I)
pr
euss
i (2)
ta
lapo
in (S
) M
EAN
1 SD
MEA
N1 S
D
SD
I I

348 D.L. GEBO AND E.J. SARGIS
Table 4
were kindly provided by him (see Ruff, 1989). All surface areas were derived from superior-inferior and anterior-posterior breadths and mediolateral depths using geo- metric formulae for partial spheres (Ruff, 1988). The cross-sectional slices begin dis- tally at 20% of bone length and proceed proximally up the shaft to 80% of bone length.
RESULTS Arboreal guenons
Cercopithecus ascanius and Cercopithecus mitis.
Red-tailed monkeys differ from blue mon- keys behaviorally in their overall higher climbing frequencies, higher leaping fre- quencies during travel, and more frequent use of vertical bounding (Table 1). The fre- quent use of oblique and vertical supports by red-tailed monkeys is in accord with their frequent climbing and bounding activities. Their differential use of the lower canopy and their smaller body size is related to their higher leaping frequency when traveling (see Fleagle and Mittermeier, 1980). Use of smaller diameter supports is also in accord with a smaller body size (Fleagle and Mitter- meier, 1980). Thus, from a morphological
point of view, Cercopithecus ascanius should differ from its closely related and sympatric relative, Cercopithecus mitis, in skeletal fea- tures which suggest increased climbing abil- ities and use of more angled supports, as well as features associated with increased leaping, especially during travel.
A morphological comparison of the limbs of red-tailed and blue monkeys, two arboreal guenons, uncovers very few anatomical fea- tures that distinguish them. Only five indi- ces distinguish Cercopithecus ascanius from Cercopithecus mitis in terms of statistical significance. These indices are a lower inter- membral index, a higher brachial index, a more proximally positioned tibia1 tubercle, a shorter navicular, and a longer third meta- tarsal for Cercopithecus ascanius (Table 5). Further, our qualitative assessments (Table 6) detect no differences in limb morphology between these two taxa. Thus, it appears that the limb morphology for these two arbo- real guenons is extremely similar.
Although the anatomical differences be- tween red-tailed and blue monkeys are few, some features could potentially be related to climbing and leaping, locomotor differences which distinguish Cercopithecus ascanius from Cercopithecus mitis (Table 1). For ex- ample, short naviculars are found in several

LIMB MORPHOLOGY IN GUENONS 349
Table 4 (continued)
primates that climb frequently (Gebo, 1988>, and the longer legs and metatarsals are fea- tures commonly found in leapers (Fleagle, 1976). The more proximal location of the tib- ial tubercle is also associated with frequent leaping or running animals (Howell, 1944; Maynard-Smith and Savage, 1956; Hilde- brand, 1974). However, these few and subtle anatomical differences are rather insub- stantial in our attempts to explain the noted behavioral differences between Cercopithe- cus mitis and Cercopithecus ascanius.
Other arboreal guenons
Comparing limb morphology among other arboreal guenons, where we lack quantita- tive observations on positional behavior, similarly shows few osteological differences across these taxa (Tables 5 and 6). Using Cercopithecus mitis as the standard of com- parison, Table 5 summarizes the statistical tests calculated on the indices reported in Table 4. The greatest number of statistically significant differences in these indices be- tween the arboreal Cercopithecus mitis and the other four arboreal guenons with ade- quate sample sizes is six out of the total twenty-six indices (Table 5). Six indice dif- ferences are noted between C. mitis and the talapoin monkey, five between C. mitis and
C. ascanius, four between C. mitis and C. cephus, and three between C. mitis and C. diana. Two indices, Index 4 (brachial index) and Index 12 (ulnar radial facet width) are shared by three of the four arboreal guenons and differ from Cercopithecus mitis. Both of these indices show higher values for the other arboreal guenons relative to Cerco- pithecus mitis.
Since we were unable to achieve sufficient sample sizes for all eighteen species of gue- nons listed in Table 2, we qualitatively eval- uated these species on a variety of osteologi- cal characters relating to joint anatomy, bony angles, and a few linear dimensions as listed in Table 6. These osteological charac- ters were selected on the basis of what we perceived to be structures that might distin- guish terrestrial and arboreal taxa. Out of the twenty-eight comparisons (Table 6), the arboreal guenons show a range of differ- ences from zero to six features, again using Cercopithecus mitis as the standard of com- parison. Thus, among arboreal guenons, morphological distinctions appear to be min- imal across these taxa.
Arboreal and semiterrestrial guenons Since both arboreal and semiterrestrial
guenons utilize the arboreal environment of-

350 D.L. GEBO AND E.J. SARGIS
TABLE 5. Summary of statistical significance of Table 4 Indices (t tests, significance at the 0.05 leuel)
Suecies comuarisons Total differences Specific indices2
mitislascanius lcephus ldiana ltalapoin lneglectus lnigrouiridis IZhoesti + preussi laethiops lpatas
lpatas lneglectus lnigrouiridis
aethiopsltalapoin lneglectus lnigrouiridis lpatas
pataslneglectus lni%rouiridis
lhoesti + preussilaethiops
3, 4, 17, 24, 26 4, 12, 14, 25 4, 12, 16 8, 11, 12, 16, 24, 25 12, 24, 26 11,26 4, 12, 17 1, 3, 5, 10, 11, 21, 24, 25, 26 1, 3, 4, 11, 12, 15, 16 1, 7, 9, 10, 11, 12, 16, 24, 26 1, 3, 4, 9, 15, 16, 17, 18 12, 20, 21, 24 12, 16, 19, 21, 22 5, 8, 10, 11, 12, 14, 25, 26 1, 5, 10, 11, 12, 14, 16, 19, 21, 22, 23, 24, 25, 26 1, 2, 4, 5, 8, 10, 11, 12, 14, 16, 19, 21, 22, 23, 24, 26 3, 4, 5, 7, 10, 11, 12, 14, 15, 16, 18, 22, 24, 26 3, 4, 12, 15, 24, 26 3, 12, 15, 23
'First, a comparison between Cercopithecw mitis, a n arboreal guenon, to other arboreal, semiterrestrial, and terrestrial guenons. Second, the combined lhoestilpreussi sample is compared against terrestrial and semiterrestrial guenons. Third, Cercopithecus aethiops, a terrestrial guenon, is compared to a close arboreal relative, Cercopithecus ialapoin, and then to other semiterrestrial and terrestrial guenons. Fourth, Erythrocebus $at,, a terrestrial guenon, is compared to the two semiterrestrial guenons.
Index 2 = supraspinatus fossa heighfflength. Index 3 = intennemhral index. Index 4 = brachial index. Index 5 = crural index. Index 6 = olecranon process lengthlulna length. Index 7 = ischium lengthiinnominate length. Index 8 = pubis lengthlinnominate length. Index 9 = ilium lengthhnnominate length. Index 10 = ischium lengthiilium length. Index 11 = ulnar humeral facet widthlsigmoid facet width. Index 12 = ulnar radial facet widthisigmoid facet width. Index 13 = greater trochanter heighvfemoral length. Index 14 = patellar facet widthiepicondylar width. Index 15 = femoral condylar heighvepicondylar width. Index 16 = medial condylar widtMateral femoral condylar width Index 17 = length to distal tibia1 cresutihial length. Index 18 = distal calcaneal lengthicalcaneal length. Index 19 = posterior calcaneal facet lengthlcalcaneal length. Index 20 = calcaneal heel widthicalcaneal length. Index 21 = talar head widthheight. Index 22 = talar trochlear widthltalar length. Index 23 = talar neck 1engtWtalar length. Index 24 = navicular lengthltalar trochlear width. Index 25 = cuboid lengthitalar trochlear width. Index 26 = third metatarsal lengthinavicular length
Index 1 = humeral head widthheight.
ten, any differences in limb morphology would most likely be the result of differing requirements of limb mechanics when on the ground. A comparison of the semiterres- trial guenons in Table 5 to Cercopithecus mi- tis, an arboreal guenon, shows only a few statistically significant indices. Cercopithe- cus nigroviridis differs in only two indices from Cercopithecus mitis while Cercopithe- cus neglectus differs by three (Table 5 ) . Both C. neglectus and C. nigroviridis share longer third metatarsals and a wider joint facet a t the elbow relative to Cercopithecus mitis.
Our qualitative assessments in Table 6 show four osteological differences between Cercopithecus mitis and Cercopithecus ne- glectus, five with Allenopithecus nigrouiri- dis, and seven with Cercopithecus hamlyni. The range of osteological differences be- tween the semiterrestrial guenons and C. mitis is very similar to the number of differ- ences noted between the arboreal guenons and Cercopithecus mitis (0-7 differences in Table 6). Thus, semiterrestrial guenons do not appear to be strikingly different in their overall limb morphology from that of the ar-

351 LIMB MORPHOLOGY IN GUENONS
boreal guenons. We assume that this assess- ment indicates that limb mechanics in both arboreal and semiterrestrial guenons must be generally similar.
Arboreal and terrestrial guenons
Cercopithecus mitis and Cercopithecus aethiops
Comparing Rose’s (1979) locomotor data on vervets with ours on blue monkeys re- veals no significant differences in frequen- cies for the different locomotor behaviors. Both species are quadrupedal 54% of the time; they climb about one-third of the time, and both leap about 10% of the time. The greatest difference in frequencies between the two studies is in climbing (29.5% vs. 35%). Both guenons also use arboreal sup- ports in the same way, showing a clear bias for horizontal and oblique supports. From these two quantitative studies of positional behavior, it is difficult to conclude that the osteological differences noted between these two species in Tables 5 and 6 are the result of behavioral differences in the arboreal mi- lieu. Rather, if these anatomical differences are related to locomotion and not other fac- tors like size (see Shea, 19921, they must be the result of differing mechanical demands in the terrestrial environment frequented more often by vervet monkeys.
Table 5 shows nine indices to be statisti- cally significant between c. aethiops and c. mitis. Relative to Cercopithecus mitis, Cer- copithecus aethiops possesses a wider hum- eral head (Index 1); a higher intermembral index (Index 3); a lower crural index (Index 5 ) ; a longer ischium (Index 10); a narrower ulnar facet for the humerus (Index 11); a wider talar head (Index 21); a longer navicu- lar and cuboid (Indices 24 and 25); and a shorter third metatarsal (Index 26). Five of these features can be associated with terres- triality while four cannot. A higher inter- membral index means more equal limb lengths and is associated with terrestrial primates as is reduced joint mobility a t the elbow (i.e., the narrow humeral facet on the ulna). Longer tarsals (navicular and cuboid) lengthen the leg for increased stride lengths, a characteristic frequently associated with cursors (Hildebrand, 1974). A short third
metatarsal may be associated with short- ened digitis as in terrestrial primates (Jolly, 1967). On the other hand, the wider hum- eral and talar heads, a shorter tibia, and a relatively longer ischium of Cercopithecus aethiops cannot be explained as adaptations for terrestriality since other terrestrial adapted mammals, including primates, tend to .display the opposite condition for these features (Maynard-Smith and Savage, 1956; Taylor, 1974,1976; Rodman, 1979).
When guenons are compared in Table 6, only five features of vervet limb anatomy appear to differ from that of blue monkeys, a relatively small number of differences com- pared to the other terrestrial guenons in Ta- ble 6. Yet, each different feature is shared with at least one of the other terrestrial gue- nons. The narrow patellar and ulnar facets and the narrow and tall talar head suggest reduced joint surfaces, a morphology associ- ated with terrestriality and more parasagit- tal limb movements. Although vervets are frequently terrestrial, vervet limb anatomy appears more intermediate with some fea- tures representing adaptations for terrestri- ality while others do not.
A case study
Cercopithecus mitis (3.5-6 kg) and Cerco- pithecus lhoesti (3.0-6 kg) (Waser, 1986) are two similar-sized congeneric guenon species which are sympatric at Kibale Forest, Uganda, and which differ in one important respect: Cercopithecus mitis is arboreal and Cercopithecus lhoesti is terrestrial. This comparison thus minimizes effects due to phylogenetic and habitat differences, and body size. Here KFP #13, an adult female of Cercopithecus lhoesti, will be compared ele- ment by element to a similar-sized adult fe- male of Cercopithecus mitis (FMNH 121598). All of the reported indices for Cer- copithecus mitis are from the sample listed in Table 2.
Limb proportions and other indices. Cercopithecus lhoesti possesses intermem- bra1 (801, brachial (971, and crural(96) indi- ces essentially similar to that of Cercopithe- cus mitis (80, 96, and 97, respectively; Table 4). A similar observation can be made for most of the other indices calculated in Table

Tab
le 6
.

TAB
LE 6
. O
STE
OLO
GtC
AL
FE
AN
RE
S A
CR
OSS
GU
EN
ON
SP
EC
IES
33
d
__
__
R
AD
IUS
ID
. di
rt81 f
acet
s 11
. bra
chia
l ind
ex
nkuh
rns
pets
urltd
a nla
r ts
lapo
in
mon
a
SCA
PUIA
I ,
WN
ed
W
NE
d C
UN
Ed
lCUrVed
CU
Ne
d
CU
NE
d
GU
N&
C
UN
ed
W
Ned
m
oder
ate
lmod
erat
e m
oder
ate
lmod
erat
e m
oder
ate
mod
erat
e m
cder
ate
mod
erat
e m
oder
ate
I I
__
__
__
_ _
__
FE
MU
R
14. a
ngle
of f
emor
al h
ead
16. a
rtic
uhr r
ulf.
cefe
mor
al
head
17.
p&et
tarf
acat
wid
th
__
_
19. m
edla
l con
dyle
Wid
th
-
16. s
upra
pate
llar w
sam
oid
depr
essi
on
18. k
no
heQ
M
I
I .-
----A
-
obliq
ue
ob
liq
r-o
bli
qu
e
perp
endi
cula
r' ob
hsue
/o
bliq
ue
obliq
ue
perp
endi
cula
r' ob
lique
m
oder
ate
mE
m
oder
ate
_mod
erat
e m
oder
ate
mod
eral
e m
oder
ate
mod
erat
e m
oder
ate
shal
low
Sh
allo
w
shal
low
S
hallo
w
Snal
lGW
Sh
allG
W
shal
low
sh
allo
w
shal
low
d
de
w
ide
wid
e w
ide _
__
wid
e na
rrow
' w
ide
wid
e w
ide
mod
erat
e m
oder
ate
mod
erat
e m
oder
ate
mod
erat
e ta
ll *
mod
erat
e m
oder
ate
mod
erat
e m
oder
ate
mod
erat
e m
oder
ate
wid
e.
mod
erat
e na
rrow
* m
oder
ate
mod
erat
e w
ide
* _
__
_~
--
TIB
IA
20.
crur
al in
dex
~_
__
_
TALU
S
21.ta
lnrh
rnd
shap
e 22
. tro
chka
r wid
th
23.
tala
r nec
k le
ngth
I
mod
erat
e - m
oder
ate
mod
erat
e m
oder
ate
mod
erat
e m
oder
ate
mod
erat
e m
oder
ate
mod
erat
e
oval
ov
al
oval
ov
al
oval
ov
a1
oval
ov
al
oval
m
oder
ate
mod
erat
e m
oder
ate
mod
erat
e m
oder
ate
mod
erat
e m
oder
ate
mod
erat
e m
oder
ate
mod
erat
e m
oder
ale
mod
erat
e lo
ng *
mod
erat
e lo
ng *
mod
erat
e m
oder
ate
mod
erat
e
CA
LCA
NE
US
24
. ant
erio
r cal
cane
al fa
cet s
ize
21. p
oste
rior c
alca
neal
face
t len
gth
26.
pe- 27
. cal
cana
ocub
oid
pivo
t dep
th
28.
heel
wid
th
I I
I 1
I I
1
t t
l2
lZ
l
I TO
TAL-
DlfE
RE
NC
ES
6
31
71
31
3
3
Tab
le 6
. (C
ontin
ued)
.
1 I
mod
erat
e m
od
er
m
oder
ate
mod
erat
e m
oder
aie
~ m
oder
ate
mod
erat
e m
oder
ate
mod
erat
e m
oder
ate
mod
erat
e m
oder
ate
mod
erat
e m
od
d m
oder
ate
mod
erat
e m
oder
ate
mod
erat
e la
me
* m
oder
ate
mod
erat
e m
oder
ate
larg
e'
mod
erat
e p--
smal
l' m
oder
ate
mod
erat
e m
oder
ate
mod
erat
e m
oder
ate
mod
eret
e m
oder
ate
mod
erat
e m
oder
ate
mod
erat
e m
oder
ate
mod
erat
e m
oder
ate
mod
erat
e m
oder
ate
mod
erat
e m
oder
ate
mod
erat
e m
oder
ate
mod
erat
e I
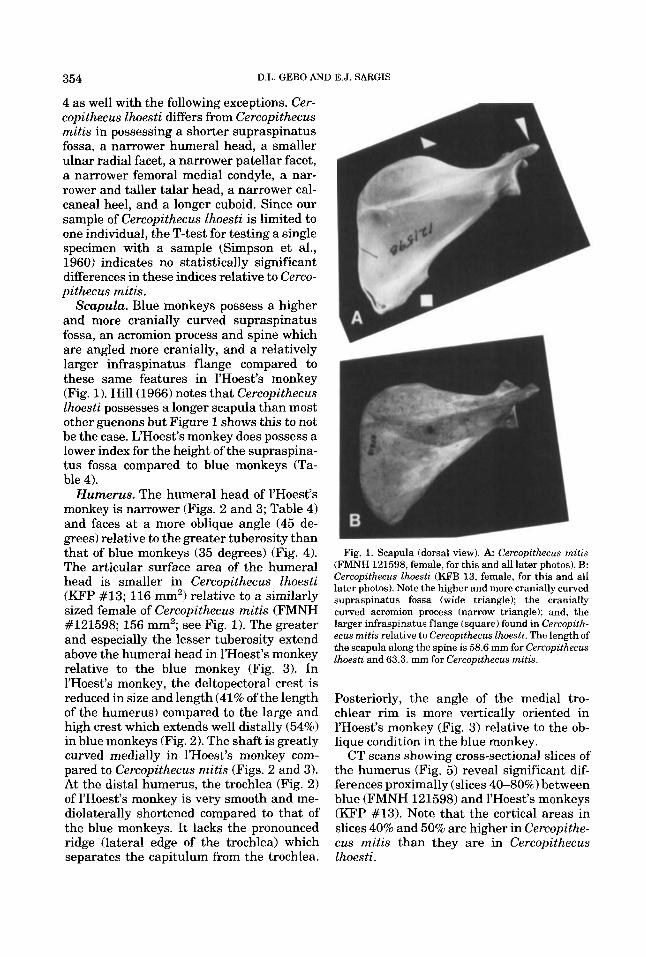
354 D.L. GEBO AND E.J. SARGIS
4 as well with the following exceptions. Cer- copithecus lhoesti differs from Cercopithecus mitis in possessing a shorter supraspinatus fossa, a narrower humeral head, a smaller ulnar radial facet, a narrower patellar facet, a narrower femoral medial condyle, a nar- rower and taller talar head, a narrower cal- caneal heel, and a longer cuboid. Since our sample of Cercopithecus lhoesti is limited to one individual, the T-test for testing a single specimen with a sample (Simpson et al., 1960) indicates no statistically significant differences in these indices relative to Cerco- pithecus mitis.
Scapula. Blue monkeys possess a higher and more cranially curved supraspinatus fossa, an acromion process and spine which are angled more cranially, and a relatively larger infraspinatus flange compared to these same features in l’Hoest’s monkey (Fig. 1). Hill (1966) notes that Cercopithecus lhoesti possesses a longer scapula than most other guenons but Figure 1 shows this to not be the case. L’Hoest’s monkey does possess a lower index for the height of the supraspina- tus fossa compared to blue monkeys (Ta- ble 4).
Humerus. The humeral head of l’Hoest’s monkey is narrower (Figs. 2 and 3; Table 4) and faces at a more oblique angle (45 de- grees) relative to the greater tuberosity than that of blue monkeys (35 degrees) (Fig. 4). The articular surface area of the humeral head is smaller in Cercopithecus lhoesti (KFP #13; 116 mm2) relative to a similarly sized female of Cercopithecus mitis (FMNH #121598; 156 mm2; see Fig. 1). The greater and especially the lesser tuberosity extend above the humeral head in I’Hoest’s monkey relative to the blue monkey (Fig. 3). In l’Hoest’s monkey, the deltopectoral crest is reduced in size and length (41% of the length of the humerus) compared to the large and high crest which extends well distally (54%) in blue monkeys (Fig. 2). The shaft is greatly curved medially in 1’Hoest’s monkey com- pared to Cercopithecus mitis (Figs. 2 and 3). At the distal humerus, the trochlea (Fig. 2) of l’Hoest’s monkey is very smooth and me- diolaterally shortened compared to that of the blue monkeys. It lacks the pronounced ridge (lateral edge of the trochlea) which separates the capitulum from the trochlea.
Fig. 1. Scapula (dorsal view). A Cercopithecus mitis (FMNH 121598, female, for this and all later photos). B: Cercopithecus lhoesti (KFB 13, female, for this and all later photos). Note the higher and more cranially curved supraspinatus fossa (wide triangle); the cranially curved acromion process (narrow triangle); and, the larger infraspinatus flange (square) found in Cercopith- ecus mitis relative to Cercopithecus Zhoesti. The length of the scapula along the spine is 58.6 mm for Cercopzthecus lhoestz and 63.3. mm for Cercopithecus rnitis.
Posteriorly, the angle of the medial tro- chlear rim is more vertically oriented in l’Hoest’s monkey (Fig. 3) relative to the ob- lique condition in the blue monkey.
CT scans showing cross-sectional slices of the humerus (Fig. 5) reveal significant dif- ferences proximally (slices 40-80%) between blue (FMNH 121598) and 1’Hoest’s monkeys (KFP #13). Note that the cortical areas in slices 40% and 50% are higher in Cercopithe- cus mitis than they are in Cercopithecus lhoesti .

LIMB MORPHOLOGY IN GUENONS 355
Fig. 2. Humerus. Anterior view of a right humerus (A), a superior view of the left humeral head (B), and an anterior view of a left distal humerus (C,D). Cercopithe- cus nitis is on the left for all views. Humeral length is 120.5 mm for Cercopithecus mi& and 124.5 mm for Cer- copithecus lhoesti. Note the greater medial curvature of
the humeral shaft (arrow, panel A) and the narrower humeral head (arrow, panel B) in Cercopithecus lhoesti. The distal humeri show the smooth trochlea of Cerco- pithecus lhoesti (D) relative to the pronounced ridge (ar- row, panel C) which separates the trochlea from the zona conoidea in Cercopithecus mitis (C).
Ulna and radius. The proximal ulna shows a reduced facet for the radial head (Fig. 6; Table 3) and the ulnar shaft is twisted distally in 1’Hoest’s monkeys rela- tive to that of blue monkeys (Fig. 6). In 1’Hoest’s monkeys, the head of the radius is smaller compared to this region in Cerco- pithecus mitis (Fig. 6) . The distal facets on the radius are very flattened in Cercopithe- cus lhoesti rather than concave (dished- shaped) as in blue monkeys (Fig. 6).
Femur and tibia. In Cercopithecus lhoe- sti, the femoral head and neck are oriented at a right angle relative to the shaft com- pared to the more obliquely oriented femoral head and neck of blue monkeys (a difference of 30 degrees; Fig. 7). The femoral head articular surface area is smaller in size in Cercopithecus lhoesti (KFP #13; 389 mm2)
relative to a similarly sized female of Cerco- pithecus mitis (FMNH 121598; 449 mm2> and the articular surface of the femoral head is reduced, especially posteriorly. The greater trochanter is situated above the femoral head in Cercopithecus lhoesti (Fig. 7 ) but its height relative to the length of the femur (Table 4) does not differentiate be- tween 1’Hoest’s and blue monkeys. The lesser trochanter is located more posteriorly in Cercopithecus lhoesti relative to Cercop- ithecus mitis. Distally, the patellar facet is very narrow (Table 4) compared to that of blue monkeys and the medial rim is much sharper rather than rounded (Fig. 7). CT scans taken on the right femur (Fig. 5) fail to show any significant differences in cortical areas between the two specimens. On the tibia, the distal tibia1 facet is anteroposteri-

356 D.L. GEBO AND E.J. SARGIS
Fig. 3. Humerus. Posterior views of a right humerus (panel A), a left proximal humerus (panel B), and a left distal humerus (panels C,D). In all views, Cercopithecus mitis is on the left and Cercopithecus lhoesti is on the right. Again, note the greater medial curvature of the humeral shaft in Cercopithecus lhoesti (arrow, panel A).
Note the smaller humeral head and the more elevated position of the greater tuberosity (arrow, panel B), as well as the vertical alignment of the medial trochlear rim (arrow, panel D) for Cercopithecus lhoesti relative to Cercopithecus mitis.
mitis I’hoesti
Fig. 4. Angle of the humeral head. The humeral head is twisted in Cercopithecus lhoesti relative to Cercopithe- cus mitis and therefore differs in its orientation to the scapula.
orly deeper and the medial part of this facet is small in 1’Hoest’s monkey compared to that in blue monkeys. The tibial tubercle is
more proximally located in 1’Hoest’s mon- keys than it is in blue monkeys (Table 4).
Tarsals. The talar trochlea is very square in appearance and the talar head is narrow and tall in Cercopithecus lhoesti (Fig. 8 ) com- pared to these same features in Cercopithe- cus mitis. The tibial cup (medial talar facet for the tibia) is very deep and pronounced compared to the blue monkey (Fig. 8). Blue monkeys possess much larger anterior cal- caneal facets (distal part), larger peroneal tubercles, and deeper calcaneocuboid pivots than do YHoest’s monkeys (Fig. 9). The heel (calcaneal tuber) is less angled (medially) in 1’Hoest’s monkey than it is in blue monkeys. There is a very deep groove distolaterally on the calcaneus of blue monkeys (attachment for ligamentum cervicis) compared to Cerco- pithecus lhoesti and the interosseous groove

LIMB MORPHOLOGY IN GUENONS 35 7
20 35 40 50 65 80
20 35 50 65 80
FEMUR
Fig. 5. CT scans. The numbers at the top of the cross-sectional slices through the humerus and femur represent percentages of bone length beginning at the distal end. Thus, the section at 35 represents a section which is taken at 35% of the length of the bone starting a t the distal end. The numbers within the cross-sectional slices represent cortical areas (mrn'). The black bar = 5 mm.
(attachment area for the anterior and poste- rior interosseous ligaments) is very tiny in Cercopithecus lhoesti.
Limb function in Cercopithecus lhoesti. In the shoulder, the shorter supraspinatus fossa height of Cercopithecus lhoesti is indic- ative of more terrestrial species which infre- quently raise their arms above the head (Roberts, 1974; Larson and Stern, 1989). The lack of curvature and lack of bending of the acromion process and spine toward the cranium also reflects limited leverage for the deltoid during arm-raising activities in Cercopithecus lhoesti. The infraspinatus
flange along the inferior angle of the scapula provides origin for the teres major muscle, a major retractor of the humerus. Its smaller size in Cercopithecus lhoesti implies less fre- quent climbing activities in this guenon (see Fleagle and Meldrum, 1988).
All cercopithecines possess proximally flattened and narrow humeral heads com- pared to New World monkeys (Fleagle and Simons, 1982; Gebo et al., 1988; Rose, 1989). The even narrower humeral head of Cerco- pithecus lhoesti suggests an even more re- stricted path of shoulder mobility in favor of fore and aft movements within the parasag-

358 D.L. GEBO AND E.J. SARGIS
Fig. 6. Ulna and radius. Panels A and B are anterior views of the ulna while the panels C and D show a superior view of the proximal radius (C) and an inferior view of the distal radius (D). Lengths for the ulna and radius for Cercopithecus lhoesti are 135 mm and 120.1 mm, respectively, and 131.6 mm and 117.5 mm for Cer- copithecus mitis. Panel A shows a closeup view of the proximal joint surface ofthe ulna for Cercopithecus lhoe- sti (right) and Cercopithecus mitis (left). Note the tiny radial facet (arrow) for Cercopithecus lhoesti relative to
ittal plane. The greater and lesser tuberosi- ties extend further above the humeral head in l'Hoest's monkey, further limiting arm ro- tation and abduction (Fleagle and Simons, 1982; Gebo et al., 1988; Rose, 1989) com- pared to blue monkeys. Larson and Stern (1989,1992) note that for terrestrial species, a higher greater tuberosity provides stabil- ity at the glenohumeral joint for a limited muscle mass (supraspinatus) and acts to limit mobility. The reduction and more prox- imal limit of the deltopectoral crest implies a proximal insertion for the deltoid and a more speed-oriented versus power function in protraction of the arm (Howell, 1944;
Cercopithecus mitis. Panel B shows the entire ulna (an- terior view) with Cercopithecus lhoesti (below) and Cer- copithecus mitis (above). Note the twist of the distal ulnar facet (arrow) in Cercopithecus lhoesti relative to Cercopithecus mitis. Panel C (superior view of the radial head), note the smaller radial head (arrow) for Cerco- pithecus lhoesti (left) relative to Cercopithecus mitis (right). In panel D (inferior view of the distal radius), note the flat radial facets (arrow) for Cercopithecus lhoe- sti (left) relative to Cercopithecus mitis (right).
Maynard-Smith and Savage, 1956; Hilde- brand, 1974; but see Stern, 1974). The angle of the humeral head, the curvature of the humeral shaft, and the twisting of the ulnar shaft suggest that the arm and forearm are more aligned in the parasagittal plane and brought more directly beneath the body than is characteristic of other arboreal gue- nons.
At the elbow, the flattened capitulum and the shortened and smooth humeral trochlea with a shallow zona and conoidea in Cerco- pithecus lhoesti, as well as the greatly re- duced ulnar facets for the radius and hu- merus and the small radial head, all

LIMB MORPHOLOGY IN GUENONS 359
Fig. 7. Femur. The Panels A, B, and C are anterior and inferior views of the proximal and distal femur, respectively. Cercopithecus lhoesti is on the right and Cercopithecus mitis is on the left in the panels. Femoral length is 156.6 mm for Cercopithecus lhoesti and 151.5 mm for Cercopithecus mitis. Note the perpendicular an- gle of the femoral head (narrow triangle, panel A), the more elevated greater trochanter relative to the femoral head (wide triangle, panel A), and the narrower patellar facet (triangle, panel C) in Cercopithecus lhoesti relative to Cercopithecus mitis.
function to reduce rotational or side-to-side movements a t the elbow in favor of the fore- and-aft direction of limb flexion and exten- sion (Rose, 1988; Harrison, 1989).
Likewise, at the wrist, the very flattened versus dish-shaped distal radial facets in 1’Hoest’s monkeys reduce side-to-side move- ments and function to resist compressive forces from terrestrial quadrupedalism.
Comparative information from the CT scans shows ratios for humeral and femoral head surface areas relative to cortical areas (50% slice) to be smaller in our single speci- men of a female I’Hoest’s monkey (KFP #13; 3.8 and 9.7, respectively), compared to a similar-sized female blue monkey (FMNH 121598; 4.1 and 11.3). Lower ratios are in-
Fig. 8. Talus. Dorsal (Panel A) and distal (Panel B) views of tali of Cercopithecus lhoesti (left) and Cerco- pithecus mitis (right). Talar length is 18.9 mm for Cerco- pithecus lhoesti and 19.0 mm for Cercopithecus rnitis. Note the pronounced tibia1 cup (triangle, panel A) and the narrow and tall talar head (triangle, panel B) in Cercopithecus lhoesti relative to Cercopithecus mitis.
dicative of terrestrial species (see Ruff, 1988). Likewise, comparing cortical areas of the humerus to the femur shows our female Cercopithecus lhoesti to possess a smaller value (0.75) relative to our female blue mon- key (0.94), indicating that forelimb loading is enhanced in the arboreal guenon. Calcu- lating A-P to M-L bending rigidity (IxAy) as well as bending and torsional rigidities (J) for the humerus relative to the femur at midlength shows our female Cercopithecus lhoesti to possess lower values (0.57 and 0.53, respectively) relative to these same values for a similarly sized female blue mon- key (1.08 and 0.61). Again, the J values are indicative of higher forelimb loading in the arboreal guenon (Schaffler et al., 1985; Ruff, 1987). IdIy values show a proximal to mid- shaft decline in the arboreal species, Cerco- pithecus mitis, which could reflect more fre- quent leaping (Ruff, 1987).

360 D.L. GEBO AND E.J. SARGIS
The perpendicular angle of the femoral head and neck to the shaft, as well as the greater height of the greater trochanter, the more posterior location of the lesser tro- chanter, and the small size and restricted joint surface of the femoral head, all imply an increased emphasis on fore and aft move- ments rather than enhanced rotational movements at the hip (Grand and Lorenz, 1968; Walker, 1974; Jenkins and Camazine, 1977; Fleagle, 1976). The very narrow patel- lar facet at the distal femur and the sharp medial rim imply fewer twisting movements at the knee than is likely the case for blue monkeys. The more proximal tibial tubercle position also implies a more speed-oriented lower limb (Howell, 1944; Hildebrand, 1974).
At the ankle, the deep distal tibial facet and the small medial facet in 1’Hoest’s mon-
Fig. 9. Calcaneus. Dorsal (Panel A), medial (Panel B) and plantar (Panel C) views of cal- canei of Cercopithecus Zhoesti (left in A and C; above in B) and Cercopithecus mitis (right in A and C; below in B). Calcaneal length is 28.6 mm for Cercopithecus Zhoesti and 28.4 mm for Cercopithecus mitis. Note the small peroneal tubecle (arrow, panel A) for Cercopithecus lhoesti, the large anterior calcaneal facet (ar- row, panel B) for Cercopithecus mitis, and the deeper calcaneocuboid pivot (arrow, panel C) for Cercopithecus mitis.
key function to restrict movements to the parasagittal direction compared to that in blue monkeys. Likewise, the shallow calca- neocuboid pivot (Kingdon, 19881, the narrow but taller talar head, and the small distal calcaneal facet suggest reduced helical and rotational movements at the subtalar and transverse tarsal joints in 1’Hoest’s monkeys (see Szalay, 1975; Lewis, 1980a,b; Langdon, 1984; Rose, 1986; Gebo and Simons, 1987; Strasser, 1988; Gebo, 1989, for references to other cercopithecoids). The small areas for ligament attachments imply a reduced need for talocrural stability corresponding with a reduced emphasis on the complex foot move- ments utilized in arboreality (Langdon, 1984). The smaller peroneal tubercle on the calcaneus implies a smaller tendon for the peroneus longus muscle, an important mus- cle in flexion and grasping of the big toe. The

LIMB MORPHOLOGY IN GUENONS 361
less angled (medially curved) calcaneal heel suggests a reduction in size and in the role of the intrinsic foot abductors in YHoest’s mon- key. All of these foot features suggest fur- ther limb modifications for flexion and ex- tension of the foot and a reduction in complex foot movements like inversion, an important foot movement during arboreal locomotion.
In sum, most of the observed morphologi- cal differences between these two guenons, the arboreal Cercopithecus mitis and the ter- restrial Cercopithecus lhoesti, are the result of smaller joint surfaces in the terrestrial guenon and thus a reduction of mobility a t the major joints of the limbs. This type of skeletal design is exactly what is to be ex- pected in a terrestrial primate emphasizing a more “cursorial” lifestyle. Thus, skeletal design in two closely related and sympatric species of Kibale Forest guenous corre- sponds with the behavioral and ecological distinctions, arboreality versus terrestrial- ity, observed in these two species.
Cercopithecus mitis and Cercopithecus Ihoestifpreussi
By combining the small samples of Cerco- pithecus lhoesti and Cercopithecus preussi, two closely related and terrestrial guenons, we are able to provisionally test the indices calculated in Table 4 for these two terres- trial guenons relative to the arboreal Cerco- pithecus mitis. The combined sample of Cer- copithecus lhoesti and Cercopithecus preussi shows three significant indices relative to Cercopithecus mitis including a lower bra- chial index (Index 4), a narrower ulnar facet for the radius (Index 12), and a shorter or more proximally located tibial tubercle (In- dex 17; Table 5). Indices 12 and 17 are indic- ative of cursors (Howell, 1944; Hildebrand, 1974). The reduced size of the ulnar facet restricts mobility at the elbow and is charac- teristically observed in more parasagittally oriented limb movements. Similarly, the more proximal position of the tibial tubercle reflects a more proximal muscle insertion and thus a greater distance of movement at the knee (Howell, 1944; Maynard-Smith and Savage, 1956; Hildebrand, 1974). Thus, al- though the samples are small and the over-
all differences from Cercopithecus mitis are few, these structural differences do suggest adaptations for terrestriality.
Cercopithecus mitis and Erythrocebus patas
The terrestrial guenon, Erythrocebus pa- tas, differs from the arboreal Cercopithecus mitis in a variety of aspects in Tables 5 and 6 which confirm an association between limb anatomy and terrestriality in patas mon- keys. Table 5 shows patas monkeys to differ from blue monkeys in seven indices. The higher intermembral (Index 3) and brachial indices (Index 4) for patas monkeys support their lengthened limbs, especially the distal elements as is true for terrestrial cursors (Howell, 1944; Hildebrand, 1974). Similarly, a high femoral condylar height (Index 15) is associated with limb mechanics for running and leaping animals (Hildebrand, 1974). The narrower ulnar facet for the radius (In- dex 12) suggests a reduced joint surface, a feature associated with terrestriality. The wider, humeral head (Index I), ulnar facet for the humerus (Index 111, and medial condyle of the femur (Index 161, relative to Cercopithecus mitis, are unusual features for a terrestrial primate to possess. Surpris- ingly, the total number of significantly dif- ferent indices for patas monkeys, relative to Cercopithecus mitis, is only slightly greater than for the arboreal talapoin and red-tailed monkeys (Table 5). However, Table 5 does show that both patas and vervet monkeys exhibit the greatest number of statistical differences compared to the other guenons.
Table 6 shows that Erythrocebus patas is very different from Cercopithecus mitis with 14 different qualitative features noted. Many of these features are shared with Cercopithecus lhoesti and Cercopithecus preussi, two terrestrial guenons. All of the features shared in common among C. lhoe- sti, C. preussi, and E . patas, compared to the arboreal Cercopithecus mitis, are related to reductions in joint sizes (features 7, 10, 15, 24, and 27 in Table 6) or limb mechanics for running (e.g., a narrower scapula and the angle of curvature for the humeral shaft). Thus, the limb anatomy of Erythrocebus pa-

362 D.L. GEBO AND E.J. SARGIS
tas supports a substantial anatomical in- vestment for terrestriality.
Semiterrestrial and terrestrial guenons When we compare the semiterrestrial and
terrestrial guenons to each other, we find that the combined sample of Cercopithecus lhoestilpreussi is more similar to the semi- terrestrial guenons (C. neglectus and A. ni- groviridis) with four and five significant dif- ferences, respectively, relative to the other terrestrial guenons (Table 5). Likewise, a to- tal of four and six indices separate E. patas from the two semiterrestrial guenons (Table 5). This range of differences is again similar to our observations for the arboreal-arboreal and arboreal-semiterrestrial guenons. Two indices (Index 12 and Index 21) are shared in common by C. neglectus and A. nigroviri- dis but differ from Cercopithecus lhoestil preussi while Indices 3, 12, and 15 are shared by both semiterrestrial guenons but differ from E. patas (Table 5). C. neglectus and A. nigroviridis possess wider ulnar fac- ets (Index 12) and talar heads (Index 12) compared to C. lhoestilpreussi, the opposite condition of more terrestrially committed primates. Likewise, the lower intermembral (Index 3) and femoral condylar height (In- dex 15) indices represent the opposite condi- tion from patas monkeys. C. neglectus and A. nigroviridis differ more markedly from C. aethiops with 14 and 16 significant differ- ences reported in Table 5. Both C. neglectus and A. nigroviridis, share thirteen statisti- cally different indices relative to c. aethiops (Indices 1,5,10-12,14,16,19,21-24 and 26 in Table 5). Thus, the features which separate the terrestrial guenons from the semiterres- trial guenons serve only to highlight the dis- tinctive features of the terrestrial guenons. In fact, no single index is held in common by all of the semiterrestrial and terrestrial gue- nons relative to the arboreal Cercopithecus mitis (Table 5).
Table 6 shows the semiterrestrial gue- nons (C. hamlyni, C. neglectus and A. nigro- viridis) to differ greatly from the terrestrial guenons like Erythrocebus patas and Cerco- pithecus lhoesti. Comparing patas monkeys to C. hamlyni shows 16 differences, 14 for C. neglectus, and 12 for A. nigroviridis (Table
6). The number of limb differences which appear to separate Cercopithecus lhoesti and Cercopithecus preussi from the semiterres- trial guenons is much greater in Table 6 compared to Table 5. Of the terrestrial gue- nons, only vervets appear to be fairly similar to the semiterrestrial guenons (Table 6). However, C. hamlyni does share a few fea- tures with C. lhoesti (i.e., a narrower scap- ula, a short supraspinatus height, a narrow and tall talar head; Table 6) and all these features are interpreted to be related to ter- restriality. Likewise, C. neglectus and A. ni- groviridis share both a narrow medial femo- ral condyle and a short supraspinatus fossa, respectively, with C. lhoesti. Thus, Table 6 does show that semiterrestrial guenons can and do share a few features with other more terrestrially committed guenons but that overall, semiterrestrial guenons are ana- tomically most similar to arboreal guenons, with perhaps the exception of vervets. No osteological features listed in Table 6 appear to be held in common by all of the semiter- restrial or terrestrial guenons.
Terrestrial guenons
Cercopithecus aethiops
Cercopithecus aethiops shares with at least one other terrestrial guenon the follow- ing four features: a narrow humeral facet on the ulna, a narrow patellar facet, a short tibia, and a narrow and tall talar head (Ta- ble 5). The first two features reflect reduced joint size and limit mobility, and are fea- tures associated with terrestriality. A short tibia runs counter to the trend in other cur- sorially adapted animals which tend to lengthen the distal limb elements (Howell, 1944; Hildebrand, 1974). The functional sig- nificance of a tall versus round talar head is likely related to lessened mediolateral rota- tions at the transverse tarsal joint and the greater forces being applied in a more an- teroposterior direction. This interpretation is consistent for terrestrial primates which utilize parasagittal limb movements.
Of all the guenons in Table 5, Cercopithe- cus aethiops is certainly the most different in these postcranial indices. When Cerco- pithecus aethiops is compared to E. patas,

LIMB MORPHOLOGY IN GUENONS 363
TABLE 7. Terrestrial cursorial features associated with Erythrocebus patas
Limb Indices Higher intermembral index' Higher brachial index'
Scapula Long and narrower scapula' Squarer glenoid fossa' Shorter vertebral border' Short supraspinatus fossa (moderate-sized in C. preussi) Short infraspinatus fossa'
Humerus Strongly retroflexed humerus' Strong medial curvature of the humeral shaft Oblique angle of the humeral head Flattened anterior trochlea Narrower posterior trochlea with a high medial edge' Deep radial and ulnar fossas
Ulna Separate and small radial facets Straight ulna'
Radius Straight radius'
Femur Strongly retroflexed femur' Higher lateral patellar rim' Higher knee'
Calcaneus
Digitigrade Feet'
' Different from Cercopithecus lhoesti and Cercopithecus preussi.
Shallow calcaneocuboid pivot
fourteen statistically significant indices are uncovered (Table 5). Similarly, C. aethiops differs from the C. lhoestilpreussi sample by nine indices (Table 5). Table 6 also suggests that C. aethwps is markedly different in its limb morphology from that of E. patas, C. lhoesti, and C. preussi.
Erythrocebus patas
Limb features in Erythrocebuspatas, rela- tive to other guenons, show patas monkeys to differ morphologically most from both ar- boreal and semiterrestrial guenons and vervets, but patas monkeys do share several anatomical features with Cercopithecus lhoesti and Cercopithecus preussi (see Table 7). Both C. Ihoesti, C. preussi, and E. patas show joints (i.e., at the shoulder, elbow, hip, knee, and foot) that have been reduced in size and restrict mobility in a more or less parasagittal plane relative to other guenons (Table 7). Further Hurov (1987) has noted
longer limbs for increased stride length in patas monkeys while Kingdon (19881, Mel- drum (19891, and Strasser (1992) have docu- mented several features in the feet of patas monkeys associated with terrestriality. Also, patas monkeys display hindfoot digiti- grady, which increases stride length, in con- trast to the semiplantigrade foot of other guenons (Meldrum, 1991).
On the other hand, patas monkeys differ anatomically in a variety of ways from the lhoesti monkey group in possessing: a nar- rower scapula, a wider humeral head, a prominent lateral trochlear ridge on the dis- tal humerus, a wider ulnar facet for both the humerus and radius, a longer radius, an ob- lique femoral head, a wider patellar facet, a taller knee, a wider medial condyle, a longer talar neck, an oval talar head, a moderate- sized anterior calcaneal facet, a moderate- sized peroneal tubercle, and patas monkeys lack a distal twist to the ulna (Tables 6 and 7). Three of these features, the narrower scapula, the longer radius, and the taller knee, relate to limb mechanics for terres- trial cursors (Howell, 1944; Roberts, 1974; Hildebrand, 1974) and are more pronounced in patas monkeys than in Cercopithecus Zhoesti. However, the overall large number of anatomical differences between patas monkeys and the lhoesti monkey group, given their evolutionary distance (Fig. lo), are best explained by independent adapta- tional events for terrestriality.
DISCUSSION Locomotor behavior and habitat use
Comparing postcranial anatomy with po- sitional behavior in Cercopithecus lhoesti and C. mitis, C. aethiops and C. mitis, and C. ascanius and C. mitis, shows that the best morphobehavioral associations exist be- tween C. lhoesti and mitis. Unfortunately, Cercopithecus lhoesti is behaviorally one of the least known of all of the guenons com- pared in this paper. Both the C. aethwps-C. mitis, (terrestrial-arboreal) comparison and the C. ascanius-C. mitis (arboreal-arboreal) comparison demonstrate few substantial anatomical distinctions reflecting the be-

364 D.L. GEBO AND E.J. SARGIS
A/T Allenopithecus
T Erythrocebus
aethiops
A/T harnlyni
Fig. 10. Guenon cladogram. T = terrestrial habitus; A = arboreal habitus; and APT = semiterrestrial habitus.
havioral differences noted in the quantita- tive behavioral studies. Why might this be SO?
First, the behavioral distinctions between the two arboreal guenons, Cercopithecus as- canius and Cercopithecus mitis, are not as great as in the Presbytis and pitheciine stud- ies by Fleagle (1978) and Fleagle and Mel- drum (1988). For example, Presbytis melalo- phus leaps 68% during travel compared to
40% for Presbytis obscura, a difference of 28% compared to a 7% difference noted to occur between red-tailed and blue monkeys (Table 1). Even the more terrestrially ori- ented vervets show an extremely similar lo- comotor profile to the arboreal blue mon- keys. Therefore, one possible explanation for the lack of anatomical congruence with loco- motor behavior is that the behavioral differ- ences noted in guenons are generally far less

LIMB MORPHOLOGY IN GUENONS 365
functionally disparate relative to those ana- lyzed in other within-genera morphological studies. Thus, limb morphology should be expected to be generally more similar in overall structure.
Second, the accuracy of the locomotor fre- quencies, which is the basis of these morpho- logical comparisons, can be questioned. In- creased sampling for larger sample sizes, at different seasons, in different types of for- ests, and by different observers shows con- siderable variation in the reported frequen- cies for positional behavior (See Gebo and Chapman, 1993b, for red colobus). Some types of locomotor categories can vary by 10% during travel and up to 25% during feeding. Thus, given the small frequency dif- ferences noted to occur in the two arboreal guenons at Kibale Forest, a less than spe- cies-wide level of anatomical distinctiveness may be implied.
Third, the same morphological features can fulfill the mechanical demands of more than one biological role (Bock and von Wah- lert, 1965; Ruff and Runestad, 1992). For example, the perpendicular femoral head and neck angle in 1’Hoest’s and diana mon- keys may be due to cursoriality in the former and leaping in the latter, since both behav- iors require rapid movements of the femur in a fore-and-aft direction.
Fourth, although many guenon species live in a wide variety of habitats, few show any significant morphological divergence. For example, vervets utilize a very wide habitat range (savanna woodland, forests, semideserts) and yet no vervet group shows any significant morphological adaptations which can be specifically linked to the use of these very different habitats. Thus, one might postulate that the mechanical de- mands on vervet limbs in these different habitats are likely to be essentially similar.
Lastly, the influence of phylogeny or past adaptational events may confound mor- phobehavioral analysis. Although the radia- tion of guenon species is large, time is short (Pliocene; Leakey, 19881, and therefore one would expect morphological differences to be minimal. This appears to be the case in gue- non evolution since even semiterrestrial species like Cercopithecus hamlyni and Cer- copithecus neglectus look morphologically
very similar to the more arboreal guenons, especially in joint anatomy. In fact, all Old World monkeys share a very uniform post- cranial morphology (see for example, Schultz, 1970; Cortright, 1983; Strasser and Delson, 1987; Harrison, 1989; Meldrum, 1989). Thus, at least for guenon evolution and probably for other primate lineages as well, the influence of a historical past may well hinder our “current use” mechanical- functional associations in our attempts to explain ecobehavioral distinctions in living species of primates.
Guenon evolution Guenon systematics is still complex and
unsettled. The most recent systematic inter- pretations of this group are in the 1988 volume by Gautier-Hion et al. (See Colyn, Dutrillaux et al., Lernould, Martin and MacLarnon, and Ruvolo in that volume). One way to deal with the complex nature of guenon systematics is to simply combine the work of Ruvolo (1988), a molecular approach with a consensus cladogram, with Lernould (1988), a general review of guenon classifi- cation (Fig. 10). This allows groups of spe- cies to be compared by outgroups and thus test whether any morphological differences in guenons can best be explained as associa- tions with habitat use and locomotor abili- ties, particularly terrestriality.
Starting in the middle of the cladogram with the lhoesti monkey group, Figure 10 shows that the closest phyletic comparisons for Cercopithecus lhoesti are with Cerco- pithecus preussi and the newly described Cercopithecus solatus (Harrison, 1988). The next outgroup is Cercopithecus hamlyni fol- lowed by the cephus and mitis groups as well as Cercopithecus neglectus, Cercopithecus diana, and the mona group. Farther out- group comparisons include Cercopithecus aethiops and Cercopithecus talapoin, Eryth- rocebus patas and finally the most primitive guenon, Alleltopithecus nigroviridis.
Comparing the postcranial elements of Cercopithecus lhoesti to Cercopithecus preussi shows few morphological differences (Table 6 ) even though the two species are completely separated and isolated from each other in Western and Eastern Africa. Both species live in montane forests, are terres-

366 D.L. GEBO AND E.J. SARGIS
trial, and frequent the lower canopy most often when in trees (Lernould, 1988; Gau- tier-Hion, 1988). Cercopithecus preussi dif- fers in limb morphology from Cercopithecus lhoesti in possessing a lower brachial index, a wider humeral head, wider radial and hu- meral facets (ulna), a longer olecranon pro- cess, a wider patellar facet, a lower knee height, a shorter cuboid, and a longer third metatarsal (Table 4). Almost all of the inno- vative features for terrestriality noted to oc- cur in Cercopithecus lhoesti relative to blue monkeys are also present in Cercopithecus preussi (Table 6). This raises the question whether the morphological features which Cercopithecus preussi and Cercopithecus Zhoesti share are inherited from a common terrestrial ancestor, or have these innova- tive features associated with terrestriality evolved independently in two closely related species inhabiting similar habitats? Further outgroup comparisons suggest inheritance (see also Dutrillaux et al., 1988) and later geographical isolation from each other since few other guenons have adopted such a ter- restrial lifestyle. If correct, then the node which unites Cercopithecus lhoesti, Cerco- pithecus preussi, and Cercopithecus solatus in Figure 10 represents an ancestral terres- trial guenon.
Expanding our outgroup analysis out- ward shows the limb anatomy of the semi- terrestrial Cercopithecus hamlyni t o be more similar to that of blue monkeys and other arboreal guenons (Table 6) rather than being similar to the lhoesti monkey group. Likewise, the cephus group, the mitis group, the mona group, and Cercopithecus diana (all arboreal guenons) and Cercupithe- cus neglectus (a semiterrestial guenon), all show limb and joint anatomies more similar to those of arboreal guenons than to the lhoesti monkey group (Table 6). The sim- plest interpretation of these ancestral nodes linking all of these taxa within the clado- gram in Figure 10 is that these nodes, in- cluding the node for Cercopithecus hamlyni, represent arboreal ancestors and that the terrestrial features found to occur in the lhoesti group are unique to this group alone.
Comparing the limb anatomy of the ter- restrial Cercopithecus aethiops with its close arboreal relative Cercopithecus talapozn,
shows both species to retain joint features which are most similar to those of the arbo- real guenons (i.e., like blue monkeys, Figs. 1-9). Yet, metrical comparisons for Cerco- pithecus aethiops reveal several distinctive limb features relative to other arboreal gue- nons and to the talapoin monkey (Table 5; see also Manaster, 1979; Hurov, 1987; Mel- drum, 1989; Anapol and Bischoff, 1992). Therefore, since both vervets and the tal- apoin monkey do not share these distinctive features in common, and yet their overall joint anatomy is similar to arboreal gue- nons, it is simpler to designate their com- mon ancestral node to be that of an arboreal ancestor (Fig. 10).
Terrestrial features in the limbs of Eryth- rocebus patus are highly developed relative to other guenons and to the closely related Allenopithecus. Since patas monkeys differ anatomically from Allenopithecus and other arboreal and semiterrestrial guenons, the line which unites Erythrocebus to the other guenons is best viewed as a terrestrial lin- eage.
This leaves the most primitive guenon, Al- lenopithecus, a semiterrestrial guenon, and the ancestral condition for all guenons to be considered. Allenopithecus compares best anatomically with all of the other arboreal and semiterrestrial guenons. Allenopithe- cus, although semiterrestrial, lacks signifi- cant joint modifications in its limbs for the terrestrial environment as observed in pa- tas monkeys and the lhoesti monkey group (Table 6). The limb anatomy of Allenopithe- cus and the commonality of arboreality in guenon species throughout the cladogram strongly suggests that the ancestral guenon was likely arboreal rather than semiterres- trial (Fig. 10). It certainly was not habitu- ally terrestrial. Whether this ancestral node for guenons is viewed as being a semiterres- trial ancestor similar to Allenopithecus, or more like other arboreal guenons, matters very little from a morphological perspective since both arboreal and semiterrestrial gue- nons exhibit a similar limb structure (Ta- bles 5 and 6). Since the majority of living guenon species are in fact arboreal, it seems best to emphasize the arboreal nature of this ancestor rather than promote its semiter- restrial status. Only with a better fossil

LIMB MORPHOLOGY IN GUENONS 367
record and further anatomical work will we be better able to clarify the ancestral state of guenon locomotor evolution.
CONCLUSION This paper examines limb anatomy in ar-
boreal, semiterrestrial, and terrestrially adapted species of guenons in the within- genus morphological tradition. Few morpho- logical features could be identified to ex- plain the small differences in locomotor and habitat use observed between sympatric red-tailed and blue monkeys living at Kibale Forest as well as between other arboreal guenons. Likewise, very few anatomical fea- tures could be found to distinguish semiter- restrial from arboreal guenon positional be- havior. Further, the more terrestrially committed vervet monkey possesses an in- termediate morphology between that of ar- boreal and terrestrial guenons, reflecting its mixed substrate usage. Although vervet limb structure generally resembles that of arboreal guenons, vervets do possess some features associated with terrestriality and their limb indices differ dramatically from other guenons. Thus, our success in explain- ing vervet anatomy relative to vervet posi- tional behavior is somewhat better than the arboreal and semiterrestrial guenon com- parisons.
On the other hand, a host of morphological features could readily be identified in the terrestrial guenons Cercopithecus lhoesti, Cercopithecus preussi, and Erythrocebus pa- tas relative to arboreal guenons, and these features support habitual use of the terres- trial habitat. Many of these terrestrial fea- tures can be broken into three categories: linear lengths of bones, bone angles or orien- tation to joints, and reduction of joint sur- faces. Thus, limb elements are modified by terrestrial primates by increasing or de- creasing lengths or by changing the angles of bones and joints for more efficient limb mechanics (i.e., to increase stride length and muscle function). Further, joint surfaces are reduced in terrestrial primates to limit mo- bility to the parasagittal plane. These types of structural modifications in guenon limbs are to be expected in terrestrial mammals (see e.g., Howell, 1944; Maynard-Smith and Savage, 1956; Jolly, 1967; Hildebrand, 1974;
Taylor, 1974, 1976; Jenkins and Camazine, 1977; Rodman, 1979).
Although several species of guenons are reported as semiterrestrial, only vervets, patas monkeys, and the lhoesti monkeys ap- pear to share morphological modifications in their limb anatomy for this habitat rela- tive to other guenons. Since the lhoesti mon- key group is not closely related to patas monkeys, nor do they occupy a similar environment, it appears that terrestrially adapted guenons with substantial morpho- logical changes have evolved twice in gue- non evolution, with vervets representing a third group in the making.
The significance of terrestriality in Old World monkey evolution is not to be under- rated. The consensus of the literature ap- pears to be that the ancestral cercopithecoid was a terrestrially adapted form (Delson, 1975; Delson and Andrews, 1975; Strasser and Delson, 1987; Andrews and Aiello, 1984; Leakey, 1988; Pickford and Senut, 1988; Strasser, 1988; Harrison, 1989; Gebo, 1989; McCrossin and Benefit, 1992) and that this explains the drastic skeletal modifications for reduced mobility in the joints observed in all Old World monkeys, cercopitbecines and colobines alike. Yet, even with the peculiar limb specializations associated with a cerco- pithecoid terrestrial past, all species of Old World monkeys, including the large terres- trial baboons, are still quite capable climb- ers and arborealists t o a degree. The ances- tral guenon appears to have committed itself to an arboreal existence. Within the guenon lineage, a t least two different groups, the lhoesti group and patas mon- keys, have readopted a more terrestrial life- style. Thus, unlike other primates, Old World monkeys represent an evolutionary lineage in which transitions between the ar- boreal and terrestrial habitus are quite com- mon.
ACKNOWLEDGMENTS We thank all of the personnel at the fol-
lowing institutions for their help and kind- ness: Drs. G. Musser (American Museum of Natural History, New York), B. Patterson (Field Museum of Natural History, Chi- cago), R. Thorington (US National Museum, Washington, D.C.), and N. Tappen (Univer-

368 D.L. GEBO AND E.J. SmGIS
sity of Wisconsin-Milwaukee, Milwaukee, WI), and the staff at the Museum of Compar- ative Zoology at Harvard University (Cam- bridge, MA). We especially thank Dr. Chris Ruff (Johns Hopkins University) for his help and effort in taking the CT scans and for his calculations. We appreciate the help of Drs. Andrew Johns and G. Isabirye-Basuta in ob- taining permission to borrow and work on the skeleton of Cercopzthecus lhoesti from the collection housed at the Makerere Uni- versity Biological Field Station (Kibale For- est, Uganda). We also acknowledge the help and support from the Office of the President, Uganda, and the National Research Coun- cil. Finally, we also thank the three review- ers, Dr. Marian Dagosto, and Dr. Matt Cart- mill for their many helpful suggestions.
LITERATURE CITED Albrecht GH, Gelvin BR, and Hartman SE (1993) Ratios
as a size adjustment in morphometrics. Am. J. Phys. Anthrop. 9It441468.
Anapol F, and Bischoff LK (1992) Comparative skeletal adaptations of Cercopithecus ascanius and Cerco- pithecus aethiops to locomotor behavior. Am. J . Phys. Anthrop. Supplement 14t43 (abstract).
Andrews P, and Aiello L (1984) An evolutionary model for feeding and positional behavior. In DJ Chivers, BA Wood, and A Bilsborough (eds.): Food Acquisition and Processing in Primates. New York Plenum Press, pp. 429-466.
Bock WJ, and von Wahlert G (1965) Adaptation and the form-function complex. Evolution 19t269-299.
Cheney DL, and Seyfarth RM (1990) How Monkeys See the World. Chicago: University of Chicago Press.
Chism J , and Roweel TE (1988) The natural history of patas monkeys. In A Gautier-Hion, F Bourliere, J-P Gautier, and J Kindon (eds.): A Primate Radiation- Evolutionary Biology of The African Guenons. Cam- bridge: Cambridge Press, pp. 412438.
Colyn MM (1988) Distribution of guenons in the Zaire- Lualaba-Lomani river system. In A Gautier-Hion, F Bourliere, J-P Gautier, and J Kingdon (eds.): A Pri- mate Radiation-EvolutionaIy Biology of the Africian Guenons. Cambridge: Cambridge Press, pp. 104-124.
Cords M (1987) Forest guenons and patas monkeys. In BB Smuts, DL Cheney, RM Seyfarth, RW Wrangham, and TT Struhsaker (eds.): Primate Societies. Chicago: University of Chicago Press, pp. 98-1 11.
Cortright GW (1983) The relative mass of hindlimb muscles in anthropoid primates: Functional and evo- lutionary implications. Ph.D Dissertation, University of Chicago.
Delson E (1975) Evolutionary history of the Cercopithe- cidae. In FS Szalay (ed.): Approaches to Primate Pale- ontology. Basel: Karger, pp. 167-217.
Delson E, and Andrews P (1975) Evolution and interre- lationships of the catarrhine primates. In WP Luckett and FS Szalay (eds. ): Phylogeny of the Primates: An
Interdisciplinary Approach, New York Plenum Press, pp. 405-466.
DeVore I (1963) A comparison of the ecology and behav- ior of monkeys and apes. In SL Washburn (ed.): Clas- sification and Human Evolution. Chicago: Aldine Press, pp. 301-319.
Dutrillaux B, Muleris M, and Couturier J (1988) Chro- mosomal evolution of Cercopithecinae. In A Gautier- Hion, F Bourliere, J-P Gautier, and J Kindon (eds.): A Primate Radiation-Evolutionary Biology of the Afri- can Guenons. Cambridge: Cambridge Press, pp. 150- 159.
Fleagle JG (1976) Locomotor behavior and skeletal anatomy of sympatric Malaysian leak-monkeys (Pres- bytis obscura and Presbytis melalophos). Yrbk. Phys. Anthrop. 20:440-453.
Fleagle JG (1978) Locomotion, posture, and habitat uti- lization in two sympatric, Malaysian leaf-monkeys (Presbytis obscura and Presbytis melalophos). In GG Montgomery (ed.): The Ecology of Arboreal Folivores. Washington, D.C.: Smithsonian Press, pp. 243-251.
Fleagle J G (1988) Primate Adaptation and Evolution. New York: Academic Press.
Fleagle JG, and Simons EL (1982) The humerus of Ae- gyptopithecus zeuxis: A primitive anthropoid. Am. J . Phys. Anthrop. 59t175-193.
Fleagle JG, and Mittermeier RA (1980) Locomotor be- havior, body size, and comparative ecology of severn Surinam monkeys. Am. J . Phys. Anthrop. 52t301- 314.
Fleagle JG, and Meldrum DJ (1988) Locomotor behavior and skeletal morphology of two sympatric pitheciine monkeys, Pithecia pithecia and Chiropotes satanas. Am. J . Primatol. 16t227-249.
Gartland JS, and Struhsaker TT (1972) Polyspecific as- sociations and niche separation of rain-forest anthro- poids in Cameroon, West Africa. J. 2001. Lond. 168: 221-266.
Gautier J P (1985) Quelques caracteristiques ecologiques du singe des Marais: Allenopithecus ni- grouirzdis Lang 1923. Rev. Ecol. (Terre Vie) 40:l-11.
Gautier JP, Moysan F, Feistner ATC, Loireau JN, and Cooper RW (1992) The distribution of Cercopithecus fIhoestil solatus, an endemic guenon of Gabon. Rev. Ecol. (Terre Vie) 47:367-381.
Gautier-Hion A (1978) Food niches and coexistence in sympatric primates in Gabon. In DJ Chivers and J Hebert (eds.): Recent Advances in Primatology, Vol- ume 2. New York: Academic Press, pp. 270-286.
Gautier-Hion A (1988) Polyspecific associations among forest guenons: Ecological, behavioral and evolution- ary aspects. In A Gautier-Hion, F Bourliere, J-P Gau- tier, and J Kingdon (eds.): A Primate Radiation-Evo- lutionary Biology of the African Guenons. Cambridge: Cambridge Press, pp. 452-476.
Gautier-Hion A, and Gautier J P (1978) Le singe de Brazza: Une strategie originale. Z. Tierpsychol. 46: 84-104.
Gautier-Hion A, Emmons LH, and Dubost G (1980) A comparison of the diets of three major groups of pri- mary consumers of Gabon (primates, squirrels and ruminants), Oecologia (Berl.) 45t182-189.
Gebo DL (1988) Foot morphology and locomotor adapta- tion in Eocene primates. Folia Primatol. 50t3-41.

LIMB MORPHOLOGY IN GUENONS 369
Gebo DL (1989) Locomotor and phylogenetic consider- ations in anthropoid evolution. J. Hum. Evol. 18:201- 233.
Gebo DL (1992) Locomotor and postural behavior in AZ- ouattapalliata and Cebus capucinus. Am. J. Primatol.
Gebo DL, Beard KC, Teaford MF, Walker A, Larson SG, Jungers WL and Fleagle JG (1988) A hominoid proxi- mal humerus from the Early Miocene of Rusinga Is- land, Kenya. J. Hum. Evol. 17:393401.
Gebo DL, and Simons EL (1987) Morphology and loco- motor adaptations of the foot in early Oligocene an- thropoids. Am. J . Phys. Anthrop. 74:83-101.
Gebo DL, and Chapman CA (1993a) Positional behavior and habitat use in five sympatric Old World monkeys. Am. J. Phys. Anthrop. (in press)
Gebo DL and Chapman CA (1993b) Seasonal, annual, and forest effects on positional behavior in red colobus monkeys. Am. J. Phys. Anthrop. (in press)
Grand TI, and Lorenz R (1968) Functional analysis of the hip joint in Tarsius bancanus (Horsfield, 1821) and Tarsius syrichta (Linnaeus, 1758). Folia Prima-
Haddow AJ (1952) Field and laboratory studies on an African monkey, Cercopithecus ascanius schmidti Matschie. Proc. Zool. SOC. Lond. 122:297-394.
Hall KRL (1965) Behaviour and ecoogy of the wild patas monkey, Erythrocebus patus, in Uganda. J. 2001. Lond. 148: 15-87.
Hall KRL, and Gartlan JS (1965) Ecology and behavior of the vervet monkey, Cercopithecus aethiops, Loli Island, Lake Victoria. Proc. Zool. Soc. Lond. 145.37-56.
Harrison MJS (1988) A new species of guenon (genus Cercopithecus) from Gabon. J. 2001. Lond. 215.561- 575.
Harrison T (1989) New postcranial remains of Victoria- pithecus from the middle Miocene of Kenya. J. Hum.
Hildebrand M (1974) Analysis of Vertebrate Structure. New York: John Wiley and Sons.
Hill WCO (1966) Primates-Comparative Anatomy and Taxonomy, Vol VI, Catarrhini, Cercopithecoidea, Cercopithecinae. Edinburgh: Edinburgh University Press.
Howell AE3 (1944) Speed in Animals, their Specialization for Running and Leaping. Chicago: University of Chi- cago Press.
Hurov JR (1985) Monkey locomotion during gait transi- tions: How do interlimb time intervals, step se- quences, and kinematics change? Am. J. Phys. An- throp. 66.417427.
Hurov JR (1987) Terrestrial locomotion and back anat- omy in vervets (Cercopithecus aethiops) and patas monkeys (Erythrocebus patus). Am. J. Phys. Anthrop.
Jenkins FA, and Camazine SM (1977) Hip structure and locomotion in ambulatory and cursorial carnivores. J. Zool. Lond. 181:351-370.
Jolly CJ (1967) The evolution of the baboons. In H Vopt- borg (ed.): The Baboon in Medical Research, Vol. 11. Austin: University of Texas, pp. 23-50.
Kingdon J (1974) East African Mammals, Volume 1. Chicago: University of Chicago Press.
Kingdon J (1988) Comparative morphology of hands and
26:277-290.
tol. 161-181.
Evo~. 18:3-54.
13~297-311.
feet in the genus Cercopithecus. In A Gautier-Hion, F Bourliere, J-P Gautirr, and J Kindon (eds.): A Primate Radiation-Evolutionary Biology of the African Gue- nons. Cambridge: Cambridge Press, pp. 184-193.
Langdon JH (1984) The Miocene horninoid foot. Ph.D. Dissertation. Yale University.
Larson SG, and Stern J T (1989) Role of supraspinatus in the quadrupedal locomotion of vervets (Cercopithecus aethiopds k Implications for interpretation of humeral morphology. Am. J. Phys. Anthrop. 79:369-377.
Larson SG, and Stern JT (1992) Further evidence for the role of supraspinatus in quadrupedal monkeys. Am. J. Phys. Anthrop. 87:359-363.
Leakey M (1988) Fossil evidence for the evolution of the guenons. In A Gautier-Hion, F Bourliere, J-P Gautier, and J Kindon (eds.): A Primate Radiation-Evolution- ary Biology of The African Guenons. Cambridge: Cambridge Press, pp. 7-12.
Leland L (1987) Monkey Business. Animal Kingdom. MayIJune, pp. 24-37.
Lernould J-M (1988) Classification and geographical distribution of guenons: A review. In A Gautier-Hion, F Bourliere, J-P Gautier, and J Kindon (eds.): A Pri- mate Radiation-Evolutionary Biology of the African Guenons. Cambridge: Cambridge Press, pp. 54-78.
Lewis OJ (1980a) The joints of the evolving foot. Part I. The ankle joint. J. Anat. 130:527-543.
Lewis OJ (1980b) The joints of the evolving foot. Part 11. The intrinsic joints. J Anat. 131:275-298.
Manaster BJ (1979) Locomotor adaptations within the Cercopithecus genus: Multivariate approach. Am. J. Phys. Anthrop. 50:169-182.
Martin RD, and MacLarnon AM (1988) Quantitative comparisons of the skull and teeth in guenons. In A Gautier-Hion, F Bourliere, J-P Gautier, and J Kindon (eds.): A Primate Radiation-Evolutionary Biology of the African Guenons. Cambridge: Cambridge Press, pp. 160-183.
Maynard-Smith J , and Savage RJG (1956) Some locomo- tor adaptation in mammals. J. Linn. Soc. Lond. Zool. 42t603422.
McCrossin ML, and Benefit BR (1992) Comparative as- sessment of the ischial morphology of Vzctorzapithecus macinnesi. Am. J. Phys. Anthrop. 87:277-290.
Meldrum DJ (1989) Terrestrial adaptations in the feet of African cercopithecines. Ph.D. Dissertation, State University of New York at Stony Brook.
Meldrum DJ (1991) Kinematics of the cercopithecine foot on arboreal and terrestrial substrates with impli- cations for the interpretation of hominid terrestrial adaptations. Am. J. Phys. Anthrop. 84t273-289.
Nakagawa N (1989) Activity budget and diet of patas monkeys in Kala Maloue National Park, Cameroon: A preliminary report. Primates 30r27-34.
Napier JR, and Napier PH (1967) A Handbook of Living Primates. New York: Academic Press.
Pickford M, and Senut B (1988) In A Gautier-Hion, F Bourliere, J-P Gautier, and J Kindon (eds.): A Primate Radiation-Evolutionary Biology of the African Gue- nons. Cambridge: Cambridge Press, pp. 3553.
Rahm UH (1970) Ecology, zoogeography, and systemat- ics of some African forest monkeys. In J Napier (ed.): Old World Monkeys. New York Academic Press, pp. 591-626.

370 D.L. GEBO AND E.J. SAFlGIS
Roberts D (1974) Structure and function of the primate scapula. In F Jenkins (ed.): Primate Locomotion. New York: Academic Press, pp. 171-200.
Rodman PS (1979) Skeletal differentiation of Macaca fascicularis and Macaca nemestrina in relation to ar- boreal and terrestrial quadrupedalism. Am. J . Phys. Anthrop. 51:51-62.
Rollinson J and Martin RD (1981) Comparative aspects of primate locomotion with special reference to arbo- real cercopithecines. Symp. Zool. Soc. Land. 48,377- 427.
Rose MD (1979) Positional behavior . natural popula- tions: Some quantitative results t,f a field study of Colobus guereza and Cercopithecus aethiops. In ME Morbeck, H Preuschoft, and N Gomberg (eds.): Envi- ronment, Behavior, and Morphology, New York Gustav Fischer, pp. 7S94.
Rose MD (1986) Further hominoid postcranial speci- mens from the late Miocene Nagri Formation of Paki- stan. J . Hum. Evol. 15:333-348.
Rose MD (1988) Another look at the anthropoid elbow. J. Hum. Evol. I7:193-224.
Rose MD (1989) New postcranial specimens of catar- rhines from the Middle Miocene Chinji Formation, Pakistan: Descriptions and a discussion of proximal humeral functional morphology in anthropoids. J . Hum. Evol. 18:131-162.
Rowel1 TE (1966) Forest-living baboons in Uganda. J. Zool. Lond. 1493344-364.
Ruff C (1987) Structural allometry of the femur and tibia in Hominoidea and Macaca. Folia Primatol. 48: 9-49.
Ruff C (1988) Hindlimb articular surface allometry in Hominoidea andMacma, with comparisons to diaphy- seal scaling. J . Hum. Evol. 17:687-714.
Ruff C (1989) New approaches to structural evolution of limb bones in primates. Folia Primatol. 53:142-159.
Ruff C, and Runestad JA (1992) Primate limb bone structural adaptations. Ann. Rev. Anthropol. 21 :433- 459.
Ruvolo M (1988) Genetic evolution in the African gue- nons. In A Gautier-Hion, F Bourliere, J-P Gautier, and J Kindon (eds.): A Primate Radiation-Evolution- ary Biology of the African Guenons. Cambridge: Cam- bridge Press, pp. 127-139.
Sanderson IT (1940) The mammals of the North Cameroons forest area. Trans. Zool. SOC. Lond. 24: 623-725.
Schaffler MB, Burr DB, Jungers WL, and Ruff CB (1985) Structural and mechanical indicators of limb specialization in primates. Folia Primatol45:61-75.
Schlichte H-J (1978) The ecology of two groups of blue monkeys, Cercopithecus rnitis stuhlmanni, in an iso- lated habitat of poor vegetation. In GG Montgomery (ed.): Ecology of Arboreal Folivores. Washington, D.C.: Smithsonian Institution Press, pp. 505-517.
Schultz AH (1970) The comparative uniformity of the Cercopithecoidea. In JR Napier and PH Napier (eds.): Old World Monkeys: Evolution, Systematics and Be- havior. New York: Academic Press, pp. 39-51.
Shea BT (1992) Ontogenetic scaling of skeletal propor-
tions in the talapoin monkey. J. Hum. Evol. 23:283- 307.
Simpson GG, Roe A, and Lewontin RC (1960) Quantita- tive Zoology. New York: Harcourt Brace and World, Inc.
Stern JT (1974) Computer modelling of gross muscle dynamics. J. Biomech. 7t411-428.
Strasser E (1988) Pedal evidence for the origin and di- versification of cercopithecoid clades. J. Hum. Evol.
Strasser E (1992) Hindlimb proportions, allometry, and biomechanics in Old World monkeys (Primates, Cer- copithecidade). Am. J. Phys. Anthrop. 87:187-213.
Strasser E and Delson E (1987) Cladistic analysis of cercopithecoid relationships. J . Hum. Evol. 16:81- 100.
Struhsaker TT (1967) Social structure among vervet monkeys (Cercopithecus aethiops). Behaviour 29183- 121.
Struhsaker TT (1969) Correlates of ecology and social organization among African cercopithecines. Folia and Primatol. 11:8@118.
Struhsaker TT (1978) Food habits of five monkey spe- cies in the Kibale Forest, Uganda. In D Chivers and J Herbert (eds.): Recent Advances in Primatology. New York Academic Press, pp. 226-248.
Szalay FS (1975) Haplorhine phylogeny and the status of the Anthropoidea. In R Tuttle (ed.): Functional Morphology and Evolution. The Hague: Mouton, pp. 3-22.
Tappen NC (1960) Problems of distribution and adapta- tion of the African monkeys. Curr. Anthrop. 1:91-120.
Taylor ME (1974) The functional anatomy of the fore- limb of some African Viverridae (Carnivora). J.
Taylor ME (1976) The functional anatomy of the hind- limb of some African Viverridae (Carnivora). J . Morph. 148:227-254.
Thomas SC (1991) Population densities and patterns of habitat use among anthropoid primates of the Ituri Forest, Zaire. Biotropica 23:6%83.
Vilensky JA, and Gankiewicz E (1989) Early develop- ment of locomotor behavior in vervet monkeys. Am. J. Primatol. 17:ll-25.
Vilensky JA, and Gankiewicz E (1990a) Effects of growth and speed on hindlimb joint angular displace- ment patterns in vervet monkeys. fCercopithecus aethiops). Am. J . Phys. Anthrop. 81:441449.
Vilensky JA, and Gankiewicz E (1990b) Effects of speed on forelimb joint angular displacement patterns in vervet monkeys (Cercopithecus uethiops). Am. J. Phys. Anthrop. 83:203-210.
Vilensky JA, Gandiewicz E, and Townsend DW (1988) Effects of size on vervet (Cercopithecus aethiops) gait parameters: A cross-sectional approach. Am. J. Phys. Anthrop. 76:463480.
Vilensky JA, Moore-Kuhns M, and Moore AM (1990) Angular displacement patterns of leading and trailing limb joints during galloping in monkeys. Am. J . Pri- matol. 22:227-239.
Walker AC (1974) Locomotor adaptations in past and
17:225-246.
Movh. 143:307-336.

LIMB MORPHOLOGY IN GUENONS 371
Waser PM (1986) Interactions among primate species. In BB Smuts, DL Cheney, RM Seyfarth, RW Wrang- ham, and TT Struhsaker (eds.): Primate Societies. Chicago: Chicago University Press, pp. 210-226.
present prosimian primates. In FA Jenkins (ed.): Pri- mate Locomotion. New York: Academic Press, pp. 349-382.
Waser PM (1980) Polyspecific associations of Cercocebus albigena: Geographic variation and ecological corre- lates. Folia Primatol. 33~57-76.




















