
Terrestrial adaptations in the hands of Equatorius africanus revisited
10
Terrestrial adaptations in the hands of Equatorius africanus revisited Biren A. Patel a, * , Randall L. Susman a , James B. Rossie b , Andrew Hill c a Department of Anatomical Sciences, Stony Brook University, Stony Brook, NY 11794, USA b Department of Anthropology, Stony Brook University, Stony Brook, NY 11794, USA c Department of Anthropology, Yale University, New Haven CT 06520, USA article info Article history: Received 8 January 2009 Accepted 22 July 2009 Available online xxx Keywords: Metacarpal Phalanx Knuckle-walk Digitigrade Hominoid Miocene Palmigrade abstract Interpretations of the postcranial anatomy of East African early and middle Miocene large-bodied hominoids (e.g., Proconsul, Afropithecus, Turkanapithecus, Nacholapithecus) have suggested that these diverse primates utilized positional behaviors dominated by arboreal quadrupedalism. Preliminary descriptions of the Equatorius africanus partial skeleton (KNM-TH 28860) and other forelimb specimens, however, have argued that this animal relied more on terrestrial locomotion compared to its contem- poraries, possibly similar to extant large papionin monkeys. In this paper, we reevaluate this interpre- tation by examining intrinsic hand proportions based on the lengths of the third proximal phalanx and fifth metacarpal in Equatorius in reference to a large sample of extant catarrhine primate taxa. We focused on the lengths of these hand bones because the ratio between phalanx and metacarpal lengths has been previously documented to discriminate terrestrial from arboreal mammalian taxa, including primates. The Equatorius hand displays semi-terrestrial hand proportions with a relatively shorter proximal phalanx compared to most arboreal monkeys. Its proximal phalanx, however, is relatively longer than those of habitually terrestrial monkeys (e.g., Theropithecus, Papio). Accordingly, although Equatorius retains some arboreal quadrupedal characteristics, these results corroborate the previous inference that it engaged in more terrestrial locomotion than earlier Miocene apes such as Proconsul. We suggest that the postcranial skeleton of Equatorius evinces the earliest signs of semi-terrestriality in the hominoid fossil record. It is likely that the terrestrial specialization utilized by living hominoids, e.g., knuckle-walking, evolved separately. Ó 2009 Elsevier Ltd. All rights reserved. Introduction Functional interpretations of the postcranial anatomy of early Miocene hominoids from East Africa (Proconsul, Afropithecus, Tur- kanapithecus) have suggested that these diverse large-bodied primates utilized positional behaviors dominated by arboreal quadrupedal locomotion (Napier and Davis, 1959; Rose, 1983,1993,1994,1997; Leakey et al., 1988a,b; Ward et al., 1993; Leakey and Walker, 1997; Richmond et al., 1998; Ward, 1998; MacLatchy et al., 2000; MacLatchy, 2004). It appears that they did not engage in the suspensory behaviors characteristic of all extant hominoids and did not utilize knuckle-walking hand postures like those adopted by extant African apes during terrestrial quadru- pedal locomotion. Accordingly, these hominoids engaged in posi- tional behaviors more similar to their Oligocene ancestors, such as Aegyptopithecus (Fleagle and Simons, 1982; Fleagle, 1983; Ankel- Simons et al., 1998). In the past decade, significant postcranial remains from two middle Miocene hominoid taxa have been described from deposits in East Africa: Nacholapithecus kerioi and Equatorius africanus (Rose et al., 1996; Ishida et al., 1999; Ward et al., 1999). Both Nacholapi- thecus and Equatorius come from deposits dated to w15 Ma (Ishida et al., 1999; Ward et al., 1999). Like other Miocene hominoids from East Africa, Nacholapithecus was reconstructed as an arboreal quadruped similar to Proconsul and Afropithecus but may have also relied on more orthograde positional behaviors such as vertical climbing (Rose et al., 1996; Nakatsukasa et al., 2003,2007; Ishida et al., 2004; Senut et al., 2004; Nakatsukasa and Kunimatsu, 2009). In contrast, the expanded sample of Equatorius africanus (formerly Kenyapithecus africanus) has revealed some indications of semi- terrestrial quadrupedal behavior, like that of Chlorocebus and some species of Macaca (McCrossin, 1994a,b; Benefit and McCrossin, 1995; Ward et al., 1996; McCrossin et al., 1998; Sherwood et al., 2002; Allen, 2008). Equatorius may therefore provide the earliest evidence of the shift away from the ancestral arboreal adaptations * Corresponding author. E-mail address: [email protected] (B.A. Patel). Contents lists available at ScienceDirect Journal of Human Evolution journal homepage: www.elsevier.com/locate/jhevol 0047-2484/$ – see front matter Ó 2009 Elsevier Ltd. All rights reserved. doi:10.1016/j.jhevol.2009.08.005 Journal of Human Evolution 57 (2009) 763–772
Transcript of Terrestrial adaptations in the hands of Equatorius africanus revisited

lable at ScienceDirect
Journal of Human Evolution 57 (2009) 763–772
Contents lists avai
Journal of Human Evolution
journal homepage: www.elsevier .com/locate/ jhevol
Terrestrial adaptations in the hands of Equatorius africanus revisited
Biren A. Patel a,*, Randall L. Susman a, James B. Rossie b, Andrew Hill c
a Department of Anatomical Sciences, Stony Brook University, Stony Brook, NY 11794, USAb Department of Anthropology, Stony Brook University, Stony Brook, NY 11794, USAc Department of Anthropology, Yale University, New Haven CT 06520, USA
a r t i c l e i n f o
Article history:Received 8 January 2009Accepted 22 July 2009Available online xxx
Keywords:MetacarpalPhalanxKnuckle-walkDigitigradeHominoidMiocenePalmigrade
* Corresponding author.E-mail address: [email protected] (B.A.
0047-2484/$ – see front matter � 2009 Elsevier Ltd.doi:10.1016/j.jhevol.2009.08.005
a b s t r a c t
Interpretations of the postcranial anatomy of East African early and middle Miocene large-bodiedhominoids (e.g., Proconsul, Afropithecus, Turkanapithecus, Nacholapithecus) have suggested that thesediverse primates utilized positional behaviors dominated by arboreal quadrupedalism. Preliminarydescriptions of the Equatorius africanus partial skeleton (KNM-TH 28860) and other forelimb specimens,however, have argued that this animal relied more on terrestrial locomotion compared to its contem-poraries, possibly similar to extant large papionin monkeys. In this paper, we reevaluate this interpre-tation by examining intrinsic hand proportions based on the lengths of the third proximal phalanx andfifth metacarpal in Equatorius in reference to a large sample of extant catarrhine primate taxa. Wefocused on the lengths of these hand bones because the ratio between phalanx and metacarpal lengthshas been previously documented to discriminate terrestrial from arboreal mammalian taxa, includingprimates. The Equatorius hand displays semi-terrestrial hand proportions with a relatively shorterproximal phalanx compared to most arboreal monkeys. Its proximal phalanx, however, is relativelylonger than those of habitually terrestrial monkeys (e.g., Theropithecus, Papio). Accordingly, althoughEquatorius retains some arboreal quadrupedal characteristics, these results corroborate the previousinference that it engaged in more terrestrial locomotion than earlier Miocene apes such as Proconsul. Wesuggest that the postcranial skeleton of Equatorius evinces the earliest signs of semi-terrestriality in thehominoid fossil record. It is likely that the terrestrial specialization utilized by living hominoids, e.g.,knuckle-walking, evolved separately.
� 2009 Elsevier Ltd. All rights reserved.
Introduction
Functional interpretations of the postcranial anatomy of earlyMiocene hominoids from East Africa (Proconsul, Afropithecus, Tur-kanapithecus) have suggested that these diverse large-bodiedprimates utilized positional behaviors dominated by arborealquadrupedal locomotion (Napier and Davis, 1959; Rose,1983,1993,1994,1997; Leakey et al., 1988a,b; Ward et al., 1993;Leakey and Walker, 1997; Richmond et al., 1998; Ward, 1998;MacLatchy et al., 2000; MacLatchy, 2004). It appears that they didnot engage in the suspensory behaviors characteristic of all extanthominoids and did not utilize knuckle-walking hand postures likethose adopted by extant African apes during terrestrial quadru-pedal locomotion. Accordingly, these hominoids engaged in posi-tional behaviors more similar to their Oligocene ancestors, such as
Patel).
All rights reserved.
Aegyptopithecus (Fleagle and Simons, 1982; Fleagle, 1983; Ankel-Simons et al., 1998).
In the past decade, significant postcranial remains from twomiddle Miocene hominoid taxa have been described from depositsin East Africa: Nacholapithecus kerioi and Equatorius africanus (Roseet al., 1996; Ishida et al., 1999; Ward et al., 1999). Both Nacholapi-thecus and Equatorius come from deposits dated to w15 Ma (Ishidaet al., 1999; Ward et al., 1999). Like other Miocene hominoids fromEast Africa, Nacholapithecus was reconstructed as an arborealquadruped similar to Proconsul and Afropithecus but may have alsorelied on more orthograde positional behaviors such as verticalclimbing (Rose et al., 1996; Nakatsukasa et al., 2003,2007; Ishidaet al., 2004; Senut et al., 2004; Nakatsukasa and Kunimatsu, 2009).In contrast, the expanded sample of Equatorius africanus (formerlyKenyapithecus africanus) has revealed some indications of semi-terrestrial quadrupedal behavior, like that of Chlorocebus and somespecies of Macaca (McCrossin, 1994a,b; Benefit and McCrossin,1995; Ward et al., 1996; McCrossin et al., 1998; Sherwood et al.,2002; Allen, 2008). Equatorius may therefore provide the earliestevidence of the shift away from the ancestral arboreal adaptations

B.A. Patel et al. / Journal of Human Evolution 57 (2009) 763–772764
of early hominoids. Because both taxa are temporally andgeographically similar, they would represent an increase inecological and locomotor diversity that took place among homi-noids in the middle Miocene of East Africa (Ishida et al., 2004).
A terrestrial locomotor component in Equatorius has beeninferred principally from postcranial specimens recovered fromMaboko Island, Kenya (specimens formerly attributed to Kenyapi-thecus africanus [McCrossin, 1994a,b,1997; Benefit and McCrossin,1995; McCrossin and Benefit, 1997; McCrossin et al., 1998; Allenand McCrossin, 2007; Allen, 2008]). While preliminary study ofa partial forelimb skeleton (KNM-TH 28860) from Kipsaramon(Tugen Hills), Kenya largely concurred (Ward et al., 1996,1999;Sherwood et al., 2002), more detailed analysis of this new materialis needed to further refine our understanding of Equatorius loco-motor behavior. Some of the terrestrial features of the Equatoriusforelimb include: a greater tubercle extending above the humeralhead; distal humerus with a retroflexed medial epicondyle;a proximal ulna with a dorsally extending olecranon process, deepolecranon fossa, and well-developed coronoid process; metacarpalheads with well-developed transverse dorsal ridges and palmarlybroad articular surface; and phalanges that are short, robust, andstraight (McCrossin and Benefit, 1997; McCrossin et al., 1998;Sherwood et al., 2002; Allen and McCrossin, 2007; Allen, 2008).Most of these features are found in the material at Maboko Island(McCrossin et al., 1998) but some are also apparent in the Kipsar-amon specimens (Sherwood et al., 2002).
Although many of these characters can successfully discriminatebetween extant arboreal and terrestrial primate taxa, many of thesecharacters in Equatorius have not yet been quantitatively comparedto habitually arboreal and terrestrial quadrupedal primates(McCrossin et al., 1998; Allen, 2008). Furthermore, some character-istics of the Equatorius forelimb appear primitive (i.e., Proconsul-like), which is suggestive of some degree of arboreal quadrupedalismfor this fossil hominoid (or may not be functionally informative;Rose, 1983,1993,1994,1997; Sherwood et al., 2002; Allen, 2008).Some of these characters include: a humeral shaft that is retroflexedand has a distinct lateral supracondylar ridge; a straight and gracileradius; circular radial head; metacarpals that are short, witha dorsally wasted proximal articular surface (Ward et al., 1999;Sherwood et al., 2002; Allen, 2008).
Therefore, it is unclear whether Equatorius was as terrestrial asextant large-bodied cercopithecine monkeys (Papio, Mandrillus,Theropithecus, and Erythrocebus) and African apes (Gorilla and Pan),or if it was more semi-terrestrial like extant small- and medium-bodied cercopithecine monkeys (Chlorocebus, Cercocebus, andMacaca). Body size estimates from dental and postcranial elementsfor Equatorius are approximately 27 kg for the Kipsaramon material(Ward et al., 1999) and range between 17.3–36.3 kg for the MabokoIsland material (McCrossin et al., 1998). Therefore, these body sizeestimates place Equatorius at the higher range of extant Old Worldmonkeys (e.g., Papio, Mandrillus) and at the lower range of extantAfrican apes (i.e., Pan paniscus). Accordingly, this relatively largesize for a catarrhine primate in the middle Miocene could havebeen one plausible cause for invading a terrestrial niche. Whileprevious characterizations of positional behavior for Equatoriushave favored a macaque-like mode of semi-terrestrial locomotion(McCrossin 1994b; McCrossin and Benefit, 1997; Sherwood et al.,2002), many individual traits of the postcranium resemble condi-tions found in more committed terrestrialists, such as Papio, Ther-opithecus, and Mandrillus (McCrossin, 1994a; McCrossin et al., 1998)or Pan (McCrossin et al., 1998; Allen and McCrossin, 2007, Allen,2008). Accordingly, the degree and manner of terrestriality inEquatorius africanus remains unclear.
The fauna from Kipsaramon and Maboko are similar and suggestthe presence of more wooded than forest environments (Van
Couvering and Van Couvering, 1976; McCrossin et al., 1998; Beh-rensmeyer et al., 2002), a result that is also supported by evidencefor C4 vegetation (Kingston et al., 1994). Such a change from themore forested early Miocene East African sites provides a fittingcontext for a shift to semi-terrestrial behavior in the residentcatarrhines. Of course, the reconstructed environment does not, initself, constitute evidence for the locomotor adaptations of theprimates (McCrossin et al., 1998). Moreover, the non-primate fossilsat Kipsaramon derive from a stratigraphic layer (mostly BPRP #89)that is separated by around 200,000 years from the Equatoriuslocality (BPRP #122), making it difficult to argue that they samplethe same environment.
This paper compares Equatorius africanus hand proportions toa large sample of terrestrial and arboreal catarrhine primates. Wefocused on relative finger length (i.e., intrinsic hand proportion)because it has been shown to discriminate terrestrial from arborealmammalian taxa, including primates (Midlo, 1934; Gabis, 1960;Napier, 1967; Napier and Napier, 1967; Etter, 1973; Susman, 1979;Lemelin, 1999; Jungers et al., 2005; Lemelin and Schmitt, 2007; Kirket al., 2008; Meachen-Samuels and Van Valkenburgh, 2009).Terrestrial primates tend to have shorter phalanges relative toeither metacarpal length or total hand length than do their morearboreal counterparts (both quadrupedal and suspensory; e.g.,Midlo, 1934; Etter, 1973; Susman, 1979). Because ground reactionforces tend to be higher during terrestrial compared to arboreallocomotion (Schmitt, 1999; Schmitt and Hanna, 2004; Franz et al.,2005; Young, 2009), shorter phalanges may attenuate bendingmoments by effectively shortening the load arm for the higherground reaction force vector to act on (Nieschalk and Demes, 1993).In contrast, relative elongation of phalanges serves as an arborealadaptation because longer fingers facilitate grasping of branchesduring arboreal quadrupedalism and/or suspension (Washburn,1951; Rose, 1988; Napier, 1993; Lemelin, 1999). Accordingly, ifEquatorius also preferred terrestrial substrates, we predict that ittoo should have phalanges that are relatively short.
Materials and methods
Hand bones from a total of 315 adult individuals were measuredin a sample of extant catarrhine primates; at least two individualsof each taxon were sampled (Table 1). Adults are defined as indi-viduals with complete epiphyseal fusion of all forelimb long bonesincluding the metacarpal heads. The comparative sample includestaxa that can be grouped as: 1) AQC: arboreal quadrupedal cerco-pithecoids (n¼ 108), 2) SQC: semi-terrestrial quadrupedal cerco-pithecoids (n¼ 41), 3) TQC: terrestrial quadrupedal cercopithecoids(n¼ 41), 4) STQAH: suspensory/terrestrial quadrupedal Africanhominoids (n¼ 71), or 5) SAH: suspensory Asian hominoids(n¼ 54; Fleagle, 1999). These groups provide an appropriatecomparative set since details of the positional behaviors for eachindividual species sampled have not yet been thoroughly described(cf. Kirk et al., 2008). We acknowledge the fact that while all cer-copithecoid primates utilize arboreal substrates, some species aresubstantially more terrestrial than others, and that both arborealityand terrestriality are gradients of a range of substrate types that canbe utilized by living primates (Fleagle, 1999). Therefore we groupedspecies into broader categories, even though such categories canobscure potential continuity between behaviors (e.g., Hunt et al.,1996). The comparative sample was obtained from the AmericanMuseum of Natural History (New York, NY), the Department ofAnatomical Sciences, Stony Brook University (Stony Brook, NY), theCleveland Museum of Natural History (Cleveland, OH), the TervurenMuseum (ex-Musee Royale de l’Afrique Centrale,Tervuren,Belgium), the Museum of Comparative Zoology (Cambridge, MA),and the National Museum of Natural History (Washington, DC).
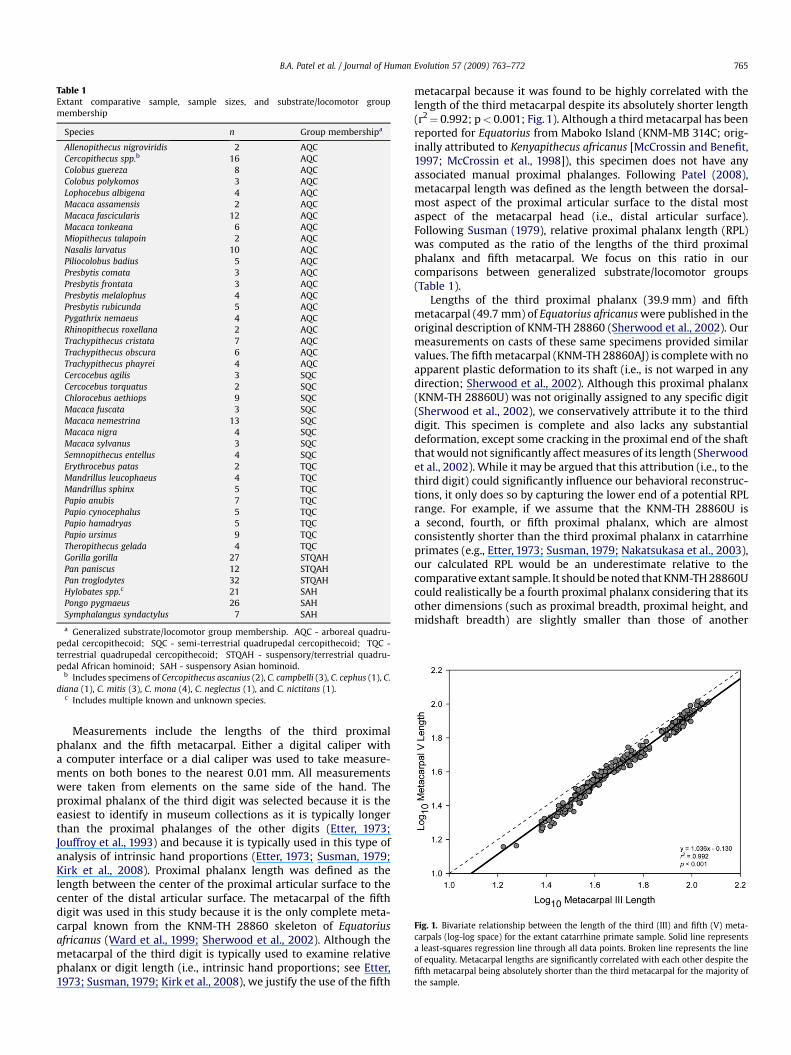
Table 1Extant comparative sample, sample sizes, and substrate/locomotor groupmembership
Species n Group membershipa
Allenopithecus nigroviridis 2 AQCCercopithecus spp.b 16 AQCColobus guereza 8 AQCColobus polykomos 3 AQCLophocebus albigena 4 AQCMacaca assamensis 2 AQCMacaca fascicularis 12 AQCMacaca tonkeana 6 AQCMiopithecus talapoin 2 AQCNasalis larvatus 10 AQCPiliocolobus badius 5 AQCPresbytis comata 3 AQCPresbytis frontata 3 AQCPresbytis melalophus 4 AQCPresbytis rubicunda 5 AQCPygathrix nemaeus 4 AQCRhinopithecus roxellana 2 AQCTrachypithecus cristata 7 AQCTrachypithecus obscura 6 AQCTrachypithecus phayrei 4 AQCCercocebus agilis 3 SQCCercocebus torquatus 2 SQCChlorocebus aethiops 9 SQCMacaca fuscata 3 SQCMacaca nemestrina 13 SQCMacaca nigra 4 SQCMacaca sylvanus 3 SQCSemnopithecus entellus 4 SQCErythrocebus patas 2 TQCMandrillus leucophaeus 4 TQCMandrillus sphinx 5 TQCPapio anubis 7 TQCPapio cynocephalus 5 TQCPapio hamadryas 5 TQCPapio ursinus 9 TQCTheropithecus gelada 4 TQCGorilla gorilla 27 STQAHPan paniscus 12 STQAHPan troglodytes 32 STQAHHylobates spp.c 21 SAHPongo pygmaeus 26 SAHSymphalangus syndactylus 7 SAH
a Generalized substrate/locomotor group membership. AQC - arboreal quadru-pedal cercopithecoid; SQC - semi-terrestrial quadrupedal cercopithecoid; TQC -terrestrial quadrupedal cercopithecoid; STQAH - suspensory/terrestrial quadru-pedal African hominoid; SAH - suspensory Asian hominoid.
b Includes specimens of Cercopithecus ascanius (2), C. campbelli (3), C. cephus (1), C.diana (1), C. mitis (3), C. mona (4), C. neglectus (1), and C. nictitans (1).
c Includes multiple known and unknown species.
Fig. 1. Bivariate relationship between the length of the third (III) and fifth (V) meta-carpals (log-log space) for the extant catarrhine primate sample. Solid line representsa least-squares regression line through all data points. Broken line represents the lineof equality. Metacarpal lengths are significantly correlated with each other despite thefifth metacarpal being absolutely shorter than the third metacarpal for the majority ofthe sample.
B.A. Patel et al. / Journal of Human Evolution 57 (2009) 763–772 765
Measurements include the lengths of the third proximalphalanx and the fifth metacarpal. Either a digital caliper witha computer interface or a dial caliper was used to take measure-ments on both bones to the nearest 0.01 mm. All measurementswere taken from elements on the same side of the hand. Theproximal phalanx of the third digit was selected because it is theeasiest to identify in museum collections as it is typically longerthan the proximal phalanges of the other digits (Etter, 1973;Jouffroy et al., 1993) and because it is typically used in this type ofanalysis of intrinsic hand proportions (Etter, 1973; Susman, 1979;Kirk et al., 2008). Proximal phalanx length was defined as thelength between the center of the proximal articular surface to thecenter of the distal articular surface. The metacarpal of the fifthdigit was used in this study because it is the only complete meta-carpal known from the KNM-TH 28860 skeleton of Equatoriusafricanus (Ward et al., 1999; Sherwood et al., 2002). Although themetacarpal of the third digit is typically used to examine relativephalanx or digit length (i.e., intrinsic hand proportions; see Etter,1973; Susman, 1979; Kirk et al., 2008), we justify the use of the fifth
metacarpal because it was found to be highly correlated with thelength of the third metacarpal despite its absolutely shorter length(r2¼ 0.992; p< 0.001; Fig. 1). Although a third metacarpal has beenreported for Equatorius from Maboko Island (KNM-MB 314C; orig-inally attributed to Kenyapithecus africanus [McCrossin and Benefit,1997; McCrossin et al., 1998]), this specimen does not have anyassociated manual proximal phalanges. Following Patel (2008),metacarpal length was defined as the length between the dorsal-most aspect of the proximal articular surface to the distal mostaspect of the metacarpal head (i.e., distal articular surface).Following Susman (1979), relative proximal phalanx length (RPL)was computed as the ratio of the lengths of the third proximalphalanx and fifth metacarpal. We focus on this ratio in ourcomparisons between generalized substrate/locomotor groups(Table 1).
Lengths of the third proximal phalanx (39.9 mm) and fifthmetacarpal (49.7 mm) of Equatorius africanus were published in theoriginal description of KNM-TH 28860 (Sherwood et al., 2002). Ourmeasurements on casts of these same specimens provided similarvalues. The fifth metacarpal (KNM-TH 28860AJ) is complete with noapparent plastic deformation to its shaft (i.e., is not warped in anydirection; Sherwood et al., 2002). Although this proximal phalanx(KNM-TH 28860U) was not originally assigned to any specific digit(Sherwood et al., 2002), we conservatively attribute it to the thirddigit. This specimen is complete and also lacks any substantialdeformation, except some cracking in the proximal end of the shaftthat would not significantly affect measures of its length (Sherwoodet al., 2002). While it may be argued that this attribution (i.e., to thethird digit) could significantly influence our behavioral reconstruc-tions, it only does so by capturing the lower end of a potential RPLrange. For example, if we assume that the KNM-TH 28860U isa second, fourth, or fifth proximal phalanx, which are almostconsistently shorter than the third proximal phalanx in catarrhineprimates (e.g., Etter, 1973; Susman, 1979; Nakatsukasa et al., 2003),our calculated RPL would be an underestimate relative to thecomparative extant sample. It should be noted that KNM-TH 28860Ucould realistically be a fourth proximal phalanx considering that itsother dimensions (such as proximal breadth, proximal height, andmidshaft breadth) are slightly smaller than those of another

Table 2Descriptive statistics for relative proximal phalanx length (RPL) for each substrate/locomotor group and fossil specimensa
Substrate/Locomotor Group n Mean SD Min Max
AQC 108 92.47 6.00 78.17 112.29SQC 41 82.00 6.46 65.28 100.15TQC 41 61.15 6.03 47.91 76.91STQAH 71 76.37 4.91 65.34 88.36SAH 54 90.09 6.59 75.41 106.48Equatorius africanus 1 80.28Theropithecus brumpti 1 59.02Archaeolemur edwardsi 1 55.56
a AQC - arboreal quadrupedal cercopithecoid; SQC - semi-terrestrial quadrupedalcercopithecoid; TQC - terrestrial quadrupedal cercopithecoid; STQAH - suspensory/terrestrial quadrupedal African hominoid; SAH - suspensory Asian hominoid.
B.A. Patel et al. / Journal of Human Evolution 57 (2009) 763–772766
associated proximal phalanx specimen, KNM-TH 28860V (see Table3 in Sherwood et al., 2002). But, if KNM-TH 28869U does in factbelong to the fourth digit, then actual RPL would also be longer thanreported here.
In addition to the extant comparative sample and the Equatoriusafricanus specimens, we included in our analysis associated handbones belonging to two additional fossil primates inferred to beadapted for terrestrial locomotion: Theropithecus brumpti andArchaeolemur edwardsi (Jablonski et al., 2002; Jungers et al., 2005).For Theropithecus brumpti, lengths of the third proximal phalanx(27.8 mm) and fifth metacarpal (47.1 mm) were taken from pub-lished values of the near complete male skeleton KNM-WT 39368(Jablonski et al., 2002). For Archaeolemur edwardsi, lengths of thethird proximal phalanx (19.0 mm) and fifth metacarpal (34.2 mm)were obtained from either published values (i.e., mean of twometacarpals) or estimated (using ImageJ software) from publishedfigures (i.e., proximal phalanx) of a near complete hand USNM447012 (Jungers et al., 2005). Although Archaeolemur is nota catarrhine primate, it is often regarded as the most monkey-likesubfossil lemur (Walker, 1974; Jungers et al., 2005) and thus makesfor an appropriate fossil to include in this comparative study offunctional morphology.
While it would be ideal to also compare intrinsic hand propor-tions in Equatorius with that of other Miocene hominoids, includingthose from the early Miocene of Africa (e.g., Proconsul, Afropithecus),the later Miocene of Europe (e.g., Pierolapithecus, Dryopithecus/Hispanopithecus), and even contemporary taxa in the middleMiocene of Africa and Europe (e.g., Nacholapithecus, Griphopithe-cus), this is not currently possible because a fifth metacarpalassociated with any proximal phalanges has yet to be documentedfor any of these taxa. We do, however, report estimates of intrinsichand proportions for some of these taxa based on other handelements, but these data are not directly comparable to the Equa-torius material or the extant comparative sample.
For all analyses, we used data for each individual specimenrather than species means in order to maximize the variation thatcould be found for each substrate/locomotor group. For the extantsample only, differences in RPL group means were examined withanalysis of variance (ANOVA) followed by Hochberg’s GT2 post-hoctests (Sokal and Rohlf, 1995). Differences in RPL among the extantgroups and the fossil sample were also evaluated graphically withbox-and-whiskers plots.
To assess whether hand bones attributed to each fossil reveala preference for a specific substrate and locomotor behavior basedon its relative proximal phalanx length, we regressed, using ordinaryleast-squares (OLS) methods (see Smith, 2009), the log (base 10) ofproximal phalanx length against the log (base 10) of fifth metacarpallength. Regression analyses were undertaken for each substrate/locomotor group: arboreal quadrupedal cercopithecoid, semi-terrestrial quadrupedal cercopithecoid, terrestrial quadrupedalcercopithecoid, suspensory/terrestrial quadrupedal African homi-noid, and suspensory Asian hominoid. Using the derived regressionequations, we computed 95% prediction intervals (PI; see Equation17.29 in Zar, 2009) for the fossil sample in reference to eachsubstrate/locomotor group and determined if the actual value foreach fossil was within the calculated 95% PIs.
Results and discussion
Table 2 provides descriptive statistics of relative proximalphalanx length (RPL) for each extant substrate/locomotor groupand the fossil specimens. Box-and-whiskers plots including eachextant group’s median value, interquartile range (50% of values),total range excluding outliers, and outliers are presented inFigure 2. ANOVA reveals significant differences in RPL among the
extant groups (df¼ 4, 310; F¼ 248.344; p< 0.001), and Hochberg’sGT2 post-hoc tests show that all groups are significantly differentfrom each other (p< 0.001) except for the arboreal quadrupedalcercopithecoids and suspensory Asian hominoids (p¼ 0.159). Thedifferences between extant groups are further highlighted in thebivariate plots between proximal phalanx length and metacarpallength (Fig. 3). As documented by Etter (1973) using the thirdmetacarpal, and here using the fifth metacarpal, terrestrialquadrupedal cercopithecoids (Theropithecus, Papio, Mandrillus,Erythrocebus) have the shortest proximal phalanges relative tometacarpal length (Figs. 2–4). In contrast, arboreal quadrupedalcercopithecoids (most colobines, some macaques and mangabeys,and most guenons) and habitually suspensory Asian hominoids(Hylobates, Symphalangus, and Pongo) have the longest proximalphalanges relative to metacarpal length. As would be predicted,semi-terrestrial quadrupedal cercopithecoids (some species ofMacaca and Cercocebus, Chlorocebus, and Semnopithecus) haveproximal phalanges that are intermediate in length between thoseof the more terrestrial and the more arboreal groups (Figs. 2–4).African hominoids (Pan and Gorilla) that engage in both suspensorybehaviors and terrestrial quadrupedal locomotion have proximalphalanges that are relatively longer than terrestrial quadrupedalcercopithecoids and relatively shorter than their more suspensoryAsian cousins (Figs. 2–4).
The Theropithecus brumpti and Archaeolemur edwardsi speci-mens studied here have relatively short proximal phalangessimilar to extant terrestrial quadrupedal cercopithecoid monkeys(Figs. 2–4). In fact, Archaeolemur appears to have an extremely shortproximal phalanx for its metacarpal length. These observations arefurther supported by the regression results and calculations of 95%PI (Table 3). Proximal phalanx lengths for both Theropithecusbrumpti and Archaeolemur edwardsi fall within the 95% PI for onlyterrestrial quadrupedal cercopithecoids and are significantlyoutside the 95% PI of all other extant groups (Table 3). Accordingly,our behavioral reconstructions presented here for both Ther-opithecus brumpti and Archaeolemur edwardsi based on handproportions support previous interpretations that both fossil taxaare adapted for terrestrial locomotion (Walker, 1974; Jablonskiet al., 2002; Jungers et al., 2005).
We find that the proximal phalanx of Equatorius africanus isrelatively longer than that of terrestrial quadrupedal cercopithe-coids (Figs. 2, 4, and 5), with its relative length falling significantlyoutside the 95% PI for that group (Table 3; Fig. 3). Rather, relativeproximal phalanx length of Equatorius cannot be distinguishedfrom semi-terrestrial quadrupedal cercopithecoid monkeys givenits metacarpal length (Table 3; Fig. 3). In fact, it also falls within the95% PI for arboreal quadrupedal cercopithecoids but at theextremes of its lower bound (Table 3). This finding agrees withprevious reconstructions of Equatorius as being capable of

Fig. 2. Box-and-whiskers plots of relative proximal phalanx length (RPL) for each substrate/locomotor group. Higher values of RPL indicated relatively longer proximal phalangesrelative to the fifth metacarpal. Horizontal lines within each box illustrate the median of the distribution. Boxes envelope the interquartile range (50% of values) of the sampledistribution, and whiskers encompass the range excluding outliers. Solid circles beyond whiskers indicate outliers. Solid black squares represent RPL values for the fossils. Groupcompositions for the extant sample are provided in Table 1.
B.A. Patel et al. / Journal of Human Evolution 57 (2009) 763–772 767
macaque-like or vervet-like semi-terrestrial quadrupedalism(McCrossin, 1994a,b; McCrossin and Benefit, 1997; McCrossin et al.,1998; Sherwood et al., 2002).
If Equatorius africanus was habitually terrestrial and used digi-tigrade hand postures like large-bodied cercopithecine monkeys(e.g., Papio, Theropithecus, Erythrocebus), its long fingers would have
Fig. 3. Bivariate relationships between fifth metacarpal length and proximal phalanx lengtrepresent least-squares regression lines through each extant group. Equatorius africanus issample regression line. Theropithecus brumpti and Archaeolemur edwardsi are within the 95statistics are presented in Table 3. Color version of this figure is available online.
likely experienced relatively large and costly bending moments(Patel, 2008). It is more likely that Equatorius would have adopteda palmigrade hand posture as originally suggested by McCrossin(1994a) in order to lower pressures on the fingers as it walked onterrestrial substrates (cf. Patel and Wunderlich, 2008,in review).Since habitually terrestrial extant cercopithecoid monkeys have
h (log-log space) for each extant substrate/locomotor group and the fossils. Solid linesoutside the 95% prediction interval (PI) of the terrestrial quadrupedal cercopithecoid% PI of the terrestrial quadrupedal cercopithecoid sample regression line. Regression
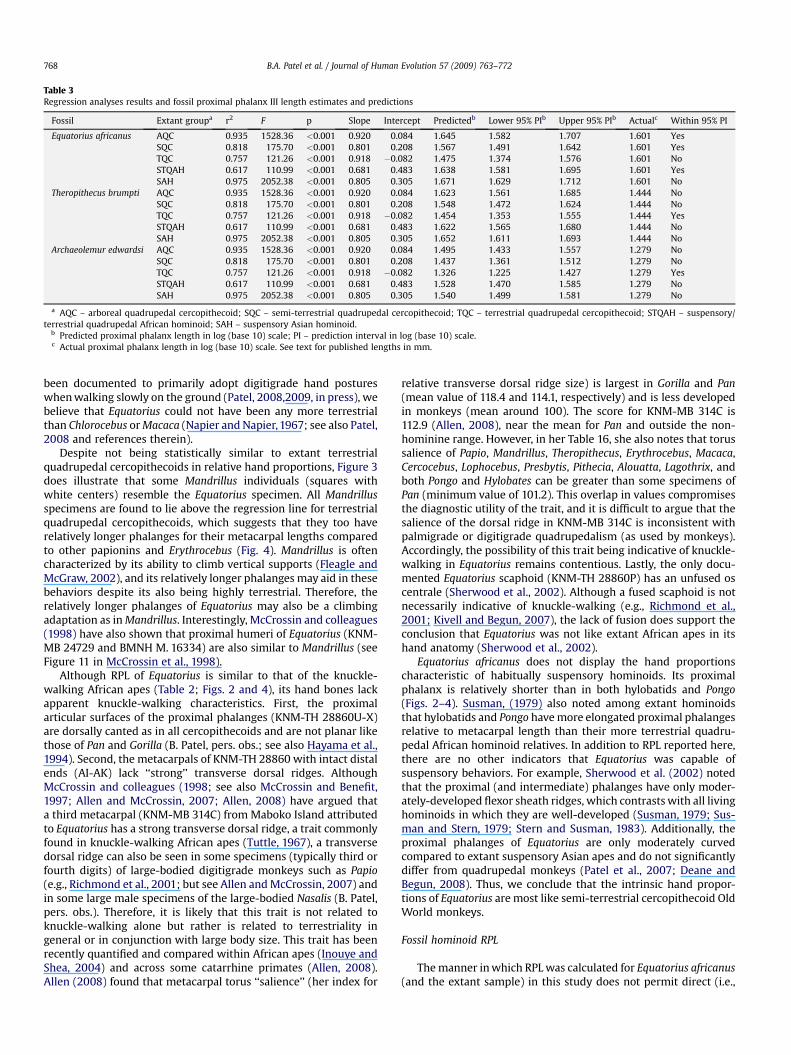
Table 3Regression analyses results and fossil proximal phalanx III length estimates and predictions
Fossil Extant groupa r2 F p Slope Intercept Predictedb Lower 95% PIb Upper 95% PIb Actualc Within 95% PI
Equatorius africanus AQC 0.935 1528.36 <0.001 0.920 0.084 1.645 1.582 1.707 1.601 YesSQC 0.818 175.70 <0.001 0.801 0.208 1.567 1.491 1.642 1.601 YesTQC 0.757 121.26 <0.001 0.918 �0.082 1.475 1.374 1.576 1.601 NoSTQAH 0.617 110.99 <0.001 0.681 0.483 1.638 1.581 1.695 1.601 YesSAH 0.975 2052.38 <0.001 0.805 0.305 1.671 1.629 1.712 1.601 No
Theropithecus brumpti AQC 0.935 1528.36 <0.001 0.920 0.084 1.623 1.561 1.685 1.444 NoSQC 0.818 175.70 <0.001 0.801 0.208 1.548 1.472 1.624 1.444 NoTQC 0.757 121.26 <0.001 0.918 �0.082 1.454 1.353 1.555 1.444 YesSTQAH 0.617 110.99 <0.001 0.681 0.483 1.622 1.565 1.680 1.444 NoSAH 0.975 2052.38 <0.001 0.805 0.305 1.652 1.611 1.693 1.444 No
Archaeolemur edwardsi AQC 0.935 1528.36 <0.001 0.920 0.084 1.495 1.433 1.557 1.279 NoSQC 0.818 175.70 <0.001 0.801 0.208 1.437 1.361 1.512 1.279 NoTQC 0.757 121.26 <0.001 0.918 �0.082 1.326 1.225 1.427 1.279 YesSTQAH 0.617 110.99 <0.001 0.681 0.483 1.528 1.470 1.585 1.279 NoSAH 0.975 2052.38 <0.001 0.805 0.305 1.540 1.499 1.581 1.279 No
a AQC – arboreal quadrupedal cercopithecoid; SQC – semi-terrestrial quadrupedal cercopithecoid; TQC – terrestrial quadrupedal cercopithecoid; STQAH – suspensory/terrestrial quadrupedal African hominoid; SAH – suspensory Asian hominoid.
b Predicted proximal phalanx length in log (base 10) scale; PI – prediction interval in log (base 10) scale.c Actual proximal phalanx length in log (base 10) scale. See text for published lengths in mm.
B.A. Patel et al. / Journal of Human Evolution 57 (2009) 763–772768
been documented to primarily adopt digitigrade hand postureswhen walking slowly on the ground (Patel, 2008,2009, in press), webelieve that Equatorius could not have been any more terrestrialthan Chlorocebus or Macaca (Napier and Napier, 1967; see also Patel,2008 and references therein).
Despite not being statistically similar to extant terrestrialquadrupedal cercopithecoids in relative hand proportions, Figure 3does illustrate that some Mandrillus individuals (squares withwhite centers) resemble the Equatorius specimen. All Mandrillusspecimens are found to lie above the regression line for terrestrialquadrupedal cercopithecoids, which suggests that they too haverelatively longer phalanges for their metacarpal lengths comparedto other papionins and Erythrocebus (Fig. 4). Mandrillus is oftencharacterized by its ability to climb vertical supports (Fleagle andMcGraw, 2002), and its relatively longer phalanges may aid in thesebehaviors despite its also being highly terrestrial. Therefore, therelatively longer phalanges of Equatorius may also be a climbingadaptation as in Mandrillus. Interestingly, McCrossin and colleagues(1998) have also shown that proximal humeri of Equatorius (KNM-MB 24729 and BMNH M. 16334) are also similar to Mandrillus (seeFigure 11 in McCrossin et al., 1998).
Although RPL of Equatorius is similar to that of the knuckle-walking African apes (Table 2; Figs. 2 and 4), its hand bones lackapparent knuckle-walking characteristics. First, the proximalarticular surfaces of the proximal phalanges (KNM-TH 28860U-X)are dorsally canted as in all cercopithecoids and are not planar likethose of Pan and Gorilla (B. Patel, pers. obs.; see also Hayama et al.,1994). Second, the metacarpals of KNM-TH 28860 with intact distalends (AI-AK) lack ‘‘strong’’ transverse dorsal ridges. AlthoughMcCrossin and colleagues (1998; see also McCrossin and Benefit,1997; Allen and McCrossin, 2007; Allen, 2008) have argued thata third metacarpal (KNM-MB 314C) from Maboko Island attributedto Equatorius has a strong transverse dorsal ridge, a trait commonlyfound in knuckle-walking African apes (Tuttle, 1967), a transversedorsal ridge can also be seen in some specimens (typically third orfourth digits) of large-bodied digitigrade monkeys such as Papio(e.g., Richmond et al., 2001; but see Allen and McCrossin, 2007) andin some large male specimens of the large-bodied Nasalis (B. Patel,pers. obs.). Therefore, it is likely that this trait is not related toknuckle-walking alone but rather is related to terrestriality ingeneral or in conjunction with large body size. This trait has beenrecently quantified and compared within African apes (Inouye andShea, 2004) and across some catarrhine primates (Allen, 2008).Allen (2008) found that metacarpal torus ‘‘salience’’ (her index for
relative transverse dorsal ridge size) is largest in Gorilla and Pan(mean value of 118.4 and 114.1, respectively) and is less developedin monkeys (mean around 100). The score for KNM-MB 314C is112.9 (Allen, 2008), near the mean for Pan and outside the non-hominine range. However, in her Table 16, she also notes that torussalience of Papio, Mandrillus, Theropithecus, Erythrocebus, Macaca,Cercocebus, Lophocebus, Presbytis, Pithecia, Alouatta, Lagothrix, andboth Pongo and Hylobates can be greater than some specimens ofPan (minimum value of 101.2). This overlap in values compromisesthe diagnostic utility of the trait, and it is difficult to argue that thesalience of the dorsal ridge in KNM-MB 314C is inconsistent withpalmigrade or digitigrade quadrupedalism (as used by monkeys).Accordingly, the possibility of this trait being indicative of knuckle-walking in Equatorius remains contentious. Lastly, the only docu-mented Equatorius scaphoid (KNM-TH 28860P) has an unfused oscentrale (Sherwood et al., 2002). Although a fused scaphoid is notnecessarily indicative of knuckle-walking (e.g., Richmond et al.,2001; Kivell and Begun, 2007), the lack of fusion does support theconclusion that Equatorius was not like extant African apes in itshand anatomy (Sherwood et al., 2002).
Equatorius africanus does not display the hand proportionscharacteristic of habitually suspensory hominoids. Its proximalphalanx is relatively shorter than in both hylobatids and Pongo(Figs. 2–4). Susman, (1979) also noted among extant hominoidsthat hylobatids and Pongo have more elongated proximal phalangesrelative to metacarpal length than their more terrestrial quadru-pedal African hominoid relatives. In addition to RPL reported here,there are no other indicators that Equatorius was capable ofsuspensory behaviors. For example, Sherwood et al. (2002) notedthat the proximal (and intermediate) phalanges have only moder-ately-developed flexor sheath ridges, which contrasts with all livinghominoids in which they are well-developed (Susman, 1979; Sus-man and Stern, 1979; Stern and Susman, 1983). Additionally, theproximal phalanges of Equatorius are only moderately curvedcompared to extant suspensory Asian apes and do not significantlydiffer from quadrupedal monkeys (Patel et al., 2007; Deane andBegun, 2008). Thus, we conclude that the intrinsic hand propor-tions of Equatorius are most like semi-terrestrial cercopithecoid OldWorld monkeys.
Fossil hominoid RPL
The manner in which RPL was calculated for Equatorius africanus(and the extant sample) in this study does not permit direct (i.e.,

Fig. 4. Box-and-whiskers plots of relative proximal phalanx length (RPL) for each taxonomic group in Table 1 displayed in descending order of mean values. Higher values of RPLindicated relatively longer proximal phalanges relative to the fifth metacarpal. Horizontal lines within each box illustrate the median of the distribution. Boxes envelope theinterquartile range (50% of values) of the sample distribution, and whiskers encompass the range excluding outliers. Open circles beyond whiskers indicate outliers. RPL forEquatorius africanus lies comfortably among the semi-terrestrial quadrupedal cercopithecoids. Color version of this figure is available online and color codes follow Fig. 3.
Fig. 5. Dorsal view photographs of the right fifth metacarpal and third proximal phalanx in select extant catarrhine species and Equatorius africanus. Each species is scaled to thesame metacarpal length. Note that Equatorius’ proximal phalanx is relatively longer than Papio and is more similar to Chlorocebus and Nasalis. Scale bar¼ 1 cm.
B.A. Patel et al. / Journal of Human Evolution 57 (2009) 763–772 769

Table 4Relative proximal phalanx length in Miocene fossil hominoids and Theropithecus brumptia.
Taxon ID # Age (Ma) RPL (%) PP# PPL (mm) MCIIIL (mm) MCVL (mm) Reference
Proconsul heseloni KNM-RU 2036 17–18.5 93.43 IV 30.00 38.00 32.11 Napier and Davis (1959)Equatorius africanus KNM-TH 28860 15 80.28 III 39.90 49.70 Sherwood et al. (2002)Dryopithecus (Hispanopithecus) laietanus IPS 18800 9.5 99.70 III 62.70 72.70b 62.89 Almecija et al. (2007)Theropithecus brumpti KNM-WT 39368 3.3 59.02 III 27.80 47.10 Jablonski et al. (2002)Theropithecus brumpti KNM-WT 39368 3.3 57.75 IV 27.20 47.10 Jablonski et al. (2002)
a RPL: relative proximal phalanx length (PPL/MCVL� 100); PP#: digit from which RPL was calculated; PPL: proximal phalanx length; MCIIIL: metacarpal III length; MCVL:metacarpal V length. For Proconsul and Dryopithecus, MCVL estimated from MCIIIL and regression equation in Figure 1.
b Based on an estimated length reported in Almecija et al. (2007).
B.A. Patel et al. / Journal of Human Evolution 57 (2009) 763–772770
statistical) comparisons with other Miocene hominoids becausethey are not represented by fifth metacarpals with associatedproximal phalanges. However, based on an examination of differenthand bones not used in the statistical analysis presented above (i.e.,metacarpals and proximal phalanges of other digits; Table 4), it ispossible to speculate whether the inferred semi-terrestrial-likehand proportions found in Equatorius is a derived condition relativeto earlier Miocene hominoid taxa reconstructed as strict arborealquadrupeds (e.g., Proconsul) and whether it differs from laterMiocene hominoid taxa that have been inferred to utilize suspen-sory behaviors (e.g., Dryopithecus). The older hominoid Proconsulheseloni from the early Miocene of Kenya (Napier and Davis, 1959)has relatively longer proximal phalanges than Equatorius: 93.43 forProconsul; 80.28 for Equatorius. If the generalized arboreal quad-rupedalism of Proconsul is primitive (e.g., Napier and Davis, 1959;Rose, 1983, 1993, 1994; Begun et al., 1994), then the shortenedproximal phalanx in Equatorius is a derived condition indicatinga shift toward increased terrestriality (McCrossin et al., 1998;Sherwood et al., 2002; Allen, 2008; present study). The youngerhominoid Dryopithecus (Hispanopithecus) laietanus from the lateMiocene of Spain (Moya-Sola and Kohler, 1999; Almecija et al.,2007, 2009) also has an RPL (99.70) substantially longer thanEquatorius and slightly longer than Proconsul. These results accordwith the pattern seen in our extant sample in which arborealquadrupeds overlap broadly with suspensory apes (Tables 2 and 4).The RPL values reported here, therefore, support previous behav-ioral reconstructions of Dryopithecus (e.g., Moya-Sola and Kohler,1999; Moya-Sola et al., 2004; Almecija et al., 2007, 2009). All of thefossil hominoids evaluated here are found to have relatively longerproximal phalanges than Theropithecus brumpti (Table 4), clearlyindicating that none were habitually terrestrial like large-bodieddigitigrade cercopitheoid monkeys.
Unfortunately, no metacarpals have been described for anyother Miocene hominoid taxa that are from a similar time period,a similar geographic location, or are closely related to Equatorius(i.e., Nacholapithecus [Nakatsukasa et al., 2003] and Griphopithecus[Ersoy et al., 2008]). Thus, a full assessment of the phylogentic andfunctional utility of RPL in fossil Miocene hominoids must awaitnew fossil material. Despite this, the clear discrimination betweenthe extant sample presented in this study provides substantialevidence that RPL is a measure with a robust functional signal.Furthermore, the clear separation between all three fossil apes andthe highly terrestrial fossil cercopithecoid Theropithecus brumpti,using a variety of different hand bone combinations to calculate RPL(Table 4), suggests that the method may not be highly sensitive toa particular phalanx/metacarpal combination.
Evolutionary implications
The results of this study speak to several contentious issuesplaguing our understanding of hominoid evolution. Is thereevidence for terrestriality in the Miocene hominoid fossil record?Were the African apes (in a cladistic sense) the first to begin
exploiting such a niche? And, is Equatorius one of them? When thehominoid fossil record was sparser, it was intelligently argued thatthe unique knuckle-walking adaptations of Pan and Gorilla repre-sented a solution to the problem of acquiring terrestrial quadru-pedal locomotion without conceding the long and curvedphalanges and metacarpals that aided in arboreal (particularlysuspensory) locomotion (Tuttle, 1967,1969). Implicit in this scenariois an arboreal and suspensory/brachiating ancestry for the homi-noid, or at least hominine, clade (e.g., Keith, 1923; Schultz,1930,1936; Harrison, 1982; Gebo, 1996; see Richmond et al., 2001for a more detailed review). Thus, the terrestrial component ofAfrican ape locomotion, as well as its requisite cheiridial adapta-tions, was seen as unique in hominoid evolution. The growingsample of Miocene ape postcranial fossils (e.g., Pilbeam et al., 1990;McCrossin, 1994a; McCrossin et al., 1998; Sherwood et al., 2002) hasunfortunately complicated this elegant hypothesis.
While parallel acquisition of suspensory postcranialmorphology in the three principal hominoid lineages from a moregeneralized arboreal quadrupedal condition has been advocated inthe past (e.g., Le Gros Clark and Leakey, 1951; Simons and Pilbeam,1965; reviewed in Larson, 1998), many current researchers arguethat consideration of extant ape anatomy favors the homology ofsuch traits (e.g., Harrison, 1982; Benefit and McCrossin, 1995; Gebo,1996; Pilbeam, 1996; MacLatchy, 2004; Begun, 2007). Comparativestudy of the extant hominoid postcranium within a cladisticframework results in only one parsimonious hypothesis: thatextant apes share a common ancestor from which they inheritedhomologous traits related to some mixture of climbing andbrachiation (Begun, 2007). The fossil record has largely failed to tellthis story (Benefit and McCrossin, 1995; Pilbeam, 1996; Larson,1998). Taxa such as Sivapithecus, Kenyapithecus, Nacholapithecus,and Equatorius are unmistakably hominoid in their craniodentalmorphology (Andrews, 1992). Yet none exhibit the full suite ofsuspensory features predicted to characterize fossil crown homi-noids (e.g., Pilbeam et al., 1990; McCrossin, 1994a; Nakatsukasaet al., 1998; Larson, 1998; Ward et al., 1999; Sherwood et al., 2002;MacLatchy, 2004; Begun, 2007). In particular, they lack modifica-tions of the shoulder that permit increased circumduction, which iscritical for ape-like brachiation (Gebo, 1996, MacLatchy, 2004). Ourbest-known middle Miocene apes were quadrupeds (Rose,1983,1993,1994,1997), and the present study adds to growingevidence that some of them were semi-terrestrial (McCrossin,1994b; Gebo, 1996; McCrossin et al., 1998; Sherwood, et al., 2002;Patel et al., 2007; Allen, 2008).
If Equatorius (or Kenyapithecus) were semi-terrestrial, thenwere they also members of a semi-terrestrial African ape clade?Theoretical, as well as empirical, evidence argues against such aninference. Although the transverse dorsal ridge present on onemetacarpal specimen of Equatorius raised the question of knuckle-walking in this taxon (McCrossin et al., 1998; Allen, 2008), thepreponderance of evidence indicates a palmigrade form oflocomotion (McCrossin and Benefit, 1997; McCrossin et al., 1998;Patel et al., 2007; present study). But could Equatorius represent

B.A. Patel et al. / Journal of Human Evolution 57 (2009) 763–772 771
an early stage in the shift toward terrestriality in the African apeclade? Such a hypothesis, were it to find future support, wouldhave to offer a new explanation for why knuckle-walking arose. Itis difficult to imagine why knuckle-walking would evolve from anancestor capable of palmigrade terrestrial locomotion (see Tuttle,1969). Without any evidence for knuckle-walking, we need onlyto entertain this hypothesis if Equatorius exhibits traits thatdemonstrate a phyletic relationship with Pan and Gorilla, and nosuch features have been found (Ward and Pilbeam, 1983;Andrews, 1992; Ward et al., 1999; Begun, 2007; Nakatsukasa andKunimatsu, 2009). Moreover, as reported here, Equatoriushad likely already undergone the digital shortening that obviatesthe need for orangutan-like fist walking or African ape-likeknuckle-walking.
It appears, then, that some middle Miocene apes (probably stemgreat apes), who were not members of the African ape clade, andwho had arboreal quadrupedal ancestors like Proconsul, began toexploit semi-terrestrial niches. Furthermore, it appears thatterrestrial adaptations among most Miocene hominoids areconvergent (Begun, 2007). The bearing of this early semi-terrestrialphase on the homology of suspensory adaptations in crownhominoids remains unresolved. Our inability to find brachiatingmiddle Miocene apes may indicate that they did not exist, or it maymean that our meager sample of functionally informative post-cranial fossils has missed them (Pilbeam and Young, 2004).Regardless, the preceding discussion suggests that the adaptivecontext of the knuckle-walking adaptations of African apes isreason enough to cling to the supposition of homology in hominoidsuspensory traits (Begun, 2007). If African apes did not descendfrom an arboreal brachiating great ape common ancestor, then whywould knuckle-walking have been favored?
Summary
This paper presents the first functional and comparative analysisof the Equatorius africanus postcranial material from Kipsaramon(KNM-TH 28860) since its original preliminary description (Wardet al., 1999; Sherwood et al., 2002). We conclude that Equatoriuswas likely capable of terrestrial quadrupedal locomotion, aspreviously suggested by analyses of its shoulder and elbow joints. Italso clearly retained many fundamentally primitive (i.e., arborealquadrupedal) traits in its forelimb, including the hand (e.g., gracileradius, gracile metacarpals, and minimal phalangeal curvature; seeSherwood et al., 2002; Deane and Begun, 2008). Therefore, it ismost likely that Equatorius was semi-terrestrial, and among extantcatarrhines, most like macaques, mangabeys, and vervet monkeys.These semi-terrestrial adaptations are distinct from themorphology associated with extant knuckle-walkers. The foregoingdiscussion further confirms the primitive nature of the earlyhominoid postcranial skeleton and implies that exploitation ofterrestrial substrates likely occurred in more than one clade duringthe course of ape evolution. Whether suspensory apes were derivedfrom these semi-terrestrial apes or their wholly arboreal counter-parts remains to be seen.
Acknowledgements
We thank the curators at the following museums and theircurators for access to comparative material: American Museum ofNatural History, Cleveland Museum of Natural History, TervurenMuseum, Museum of Comparative Zoology, and National Museumof Natural History. Ari Grossman, Andrew Lee, and Anne Suprovided valuable feedback during the preparation of this manu-script. We thank Susan Anton, an Associate Editor, and our
anonymous reviewers for providing constructive comments andsuggestions. Thanks to Luci Betti-Nash for assembling Figure 5. Thisstudy was funded in part by the National Science Foundation (BCS0524988 awarded to BAP), Stony Brook University IDPAS, and OhioUniversity College of Osteopathic Medicine RSAC.
References
Allen, K.L., 2008. The hand of Kenyapithecus africanus from Maboko Island, Kenya.M.A. Thesis, New Mexico State University.
Allen, K.L., McCrossin, M.L., 2007. Functional morphology of the Kenyapithecus handfrom Maboko Island (Kenya). Am. J. Phys. Anthropol. 44 (Suppl.), 62.
Almecija, S., Alba, D.M., Moya-Sola, S., Kohler, M., 2007. Orang-like adaptations inthe fossil hominoid Hispanopithecus laietanus: first steps towards great apesuspensory behaviours. Proc. R. Soc. B 274, 2375–2384.
Almecija, S., Alba, D.M., Moya-Sola, S., 2009. Pierolapithecus and the functionalmorphology of Miocene ape hand phalanges: paleobiological and evolutionaryimplications. J. Hum. Evol. 57, 284–297.
Andrews, P.J., 1992. Evolution and environment in Miocene hominoids. Nature 360,641–646.
Ankel-Simons, F., Fleagle, J.G., Chatrath, P.S., 1998. Femoral anatomy of Aegyptopi-thecus zeuxis, an early Oligocene anthropoid. Am. J. Phys. Anthropol. 106, 413–424.
Begun, D.R., 2007. How to identify (as opposed to define) a homoplasy: examplesfrom fossil and living great apes. J. Hum. Evol. 52, 559–572.
Begun, D.R., Teaford, M.F., Walker, A., 1994. Comparative and functional anatomy ofProconsul phalanges from the Kaswanga Primate Site, Rusinga Island, Kenya.J. Hum. Evol. 26, 89–165.
Behrensmeyer, A.K., Deino, A.L., Hill, A., Kingston, J., Saunders, J.J., 2002. Geologyand geochronology of the middle Miocene Kipsaramon site complex, MuruyurBeds, Tugen Hills, Kenya. J. Hum. Evol. 42, 11–38.
Benefit, B.R., McCrossin, M.L., 1995. Miocene hominoids and hominid origins. A. Rev.Anthropol. 24, 237–256.
Deane, A.S., Begun, D.R., 2008. Broken fingers: retesting locomotor hypotheses forfossil hominoids using fragmentary proximal phalanges and high-resolutionpolynomial curve fitting (HR-PCF). J. Hum. Evol. 55, 691–701.
Ersoy, A., Kelley, J., Andrews, P., Alpagut, B., 2008. Hominoid phalanges from themiddle Miocene site of Pasalar, Turkey. J. Hum. Evol. 54, 518–529.
Etter, H.F., 1973. Terrestrial adaptations in the hands of cercopithecinae. Folia Pri-matol. 20, 331–350.
Fleagle, J.G., 1983. Locomotor adaptations of Oligocene and Miocene hominoidsand their phyletic implications. In: Ciochon, R.L., Corruccini, R.S. (Eds.), NewInterpretations of Ape and Human Ancestry. Plenum Press, New York, pp.301–324.
Fleagle, J.G., 1999. Primate Adaptation and Evolution, second ed.. Academic Press,San Diego.
Fleagle, J.G., McGraw, W.S., 2002. Skeletal and dental morphology of Africanpapionins: unmasking a cryptic clade. J. Hum. Evol. 42, 267–292.
Fleagle, J.G., Simons, E.L., 1982. The humerus of Aegyptopithecus zeuxis, a primitiveanthropoid. Am. J. Phys. Anthropol. 59, 175–194.
Franz, T.M., Demes, B., Carlson, K.J., 2005. Gait mechanics of lemurid primates onterrestrial and arboreal substrates. J. Hum. Evol. 48, 199–217.
Gabis, R.V., 1960. Les os des members des singes cynomorphs. Mammalia 24,577–607.
Gebo, D.L., 1996. Climbing, brachiation, and terrestrial quardupedalism: historicalprecursors of hominid bipedalism. Am. J. Phys. Anthropol. 101, 55–92.
Harrison, T., 1982. Small-bodied apes from the Miocene of East Africa. Ph.D.Dissertation, University College London.
Hayama, S., Chatani, K., Nakatsukasa, M., 1994. The digitigrade hand and terrestrialadaptation in Japanese macaques. Anthropol. Sci. 102, 115–125.
Hunt, K.D., Cant, J.G.H., Gebo, D.L., Rose, M.D., Walker, S.E., Youlatos, D., 1996.Standardized descriptions of primate locomotor and postural modes. Primates37, 363–387.
Inouye, S.E., Shea, B.T., 2004. The implications of variation in knuckle-walkingfeatures for models of African hominoid locomotor evolution. J. Anthropol. Sci.82, 67–88.
Ishida, H., Kunimatsu, Y., Takano, T., Nakano, Y., Nakatsukasa, M., 2004. Nacholapi-thecus skeleton from the Middle Miocene of Kenya. J. Hum. Evol. 46, 67–101.
Ishida, H., Kunimatsu, Y., Nakatsukasa, M., Nakano, Y., 1999. New hominoid genusfrom the middle Miocene of Nachola, Kenya. Anthropol. Sci. 107, 189–191.
Jablonski, N., Leakey, M., Kiarie, C., Anton, M., 2002. A new skeleton of Theropithecusbrumpti (Primates: Cercopithecidae) from Lomekwi, West Turkana, Kenya.J. Hum. Evol. 43, 887–923.
Jouffroy, F.K., Godinot, M., Nakano, Y., 1993. Biometrical characteristics of primatehands. In: Preuschoft, H., Chivers, D.J. (Eds.), Hands of Primates. Springer-Verlag,Wien, pp. 133–173.
Jungers, W.L., Lemelin, P., Godfrey, L.R., Wunderlich, R.E., Burney, D.A., Simons, E.L.,Chatrath, P.S., James, H.F., Randria, G.F.N., 2005. The hands and feet of Archae-olemur: metrical affinities and their functional significance. J. Hum. Evol. 49, 36–55.
Keith, A., 1923. Man’s posture: its evolution and disorders. Br. Med. J. 1, 451–454.Kingston, J.D., Marino, B.D., Hill, A., 1994. Isotopic evidence for Neogene hominid
paleoenvironments in the Kenya Rift Valley. Science 264, 955–959.

B.A. Patel et al. / Journal of Human Evolution 57 (2009) 763–772772
Kirk, E.C., Lemelin, P., Hamrick, M.W., Boyer, D.M., Bloch, J.I., 2008. Intrinsic handproportions of euarchontans and other mammals: implications for the loco-motor behavior of plesiadapiforms. J. Hum. Evol. 55, 278–299.
Kivell, T.L., Begun, D.R., 2007. Frequency and timing of scaphoid-centrale fusion inhominoids. J. Hum. Evol. 52, 321–340.
Larson, S.G., 1998. Parallel evolution in the hominoid trunk and forelimb. Evol.Anthropol. 6, 87–99.
Le Gros Clark, W., Leakey, L.S.B., 1951. The Miocene Hominoidea of East Africa: FossilMammals of Africa. British Museum (Natural History), London, pp. 1–117.
Leakey, M., Walker, A., 1997. Afropithecus: function and phylogeny. In: Begun, D.R.,Ward, C.V., Rose, M.D. (Eds.), Function, Phylogeny, and Fossils. Plenum Press,New York, pp. 225–240.
Leakey, R.E.F., Leakey, M.G., Walker, A.C., 1988a. Morphology of Afropithecus turka-nensis from Kenya. Am. J. Phys. Anthropol. 76, 289–307.
Leakey, R.E.F., Leakey, M.G., Walker, A.C., 1988b. Morphology of Turkanapithecuskalakolensis from Kenya. Am. J. Phys. Anthropol. 76, 277–288.
Lemelin, P., 1999. Morphological correlates of substrate use in didelphid marsupials:implications for primate origins. J. Zool. Lond. 247, 165–175.
Lemelin, P., Schmitt, D., 2007. Origins of grasping and locomotor adaptations inprimates: comparative and experimental approaches using an opossum model.In: Ravosa, M.J., Dagosto, M. (Eds.), Primate Origins: Adaptations and Evolution.Springer, New York, pp. 329–380.
MacLatchy, L., 2004. The oldest ape. Evol. Anthropol. 13, 90–103.MacLatchy, L., Gebo, D., Kityo, R., Pilbeam, D., 2000. Postcranial functional
morphology of Morotopithecus bishopi, with implications for the evolution ofmodern ape locomotion. J. Hum. Evol. 39, 159–183.
McCrossin, M.L., 1994a The Phylogenetic Relationships, Adaptations, and Ecology ofKenyapithecus. Ph.D. Dissertation, University of California, Berkeley.
McCrossin, M.L., 1994b. Semi-terrestrial adaptations of Kenyapithecus. Am. J. Phys.Anthropol. Suppl. 18, 142–143.
McCrossin, M.L., 1997. New postcranial remains of Kenyapithecus and their impli-cations for understanding the origins of hominoid terrestriality. Am. J. Phys.Anthropol. Suppl. 24, 164.
McCrossin, M.L., Benefit, B.R., 1997. On the relationships and adaptations of Kenyapi-thecus, a large-bodied hominoid from the Middle Miocene of eastern Africa. In:Begun, D.R., Ward, C.V., Rose, M.D. (Eds.), Function, Phylogeny, and Fossils: MioceneHominoid Evolution and Adaptations. Plenum Press, New York, pp. 241–267.
McCrossin, M.L., Benefit, B.R., Gitau, S.N., Palmer, A.K., Blue, K.T., 1998. Fossilevidence for the origins of terrestriality among Old World higher primates. In:Strasser, E., Fleagle, J., Rosenberger, A., McHenry, H. (Eds.), Primate Locomotion:Recent Advances. Plenum Press, New York, pp. 353–396.
Meachen-Samuels, J., Van Valkenburgh, B., 2009. Forelimb indicators of prey-sizepreference in the Felidae. J. Morph. 270, 729–744.
Midlo, C., 1934. Form of the hand and foot in primates. Am. J. Phys. Anthropol. 19,337–389.
Moya-Sola, S., Kohler, M., 1999. A Dryopithecus skeleton and the origins of great-apelocomotion. Nature 379, 156–159.
Moya-Sola, S., Kohler, M., Alba, D.M., Casanovas-Vilar, I., Galindo, J., 2004. Pier-olapithecus catalaunicus, a new Middle Miocene great ape from Spain. Science306, 1339–1344.
Nakatsukasa, M., Kunimatsu, Y., 2009. Nacholapithecs and its importance forunderstanding hominoid evolution. Evol. Anthropol. 18, 103–119.
Nakatsukasa, M., Kunimatsu, Y., Nakano, Y., Ishida, H., 2007. Vertebral morphologyof Nacholapithecus kerioi based on KNM-BG 35250. J. Hum. Evol. 52, 347–369.
Nakatsukasa, M., Kunimatsu, Y., Nakano, Y., Takano, T., Ishida, H., 2003. Comparativeand functional anatomy of phalanges in Nacholapithecus kerioi, a MiddleMiocene hominoid from northern Kenya. Primates 44, 371–412.
Nakatsukasa, M., Yamanaka, A., Kunimatsu, Y., Shimizu, D., Ishida, H., 1998. A newlydiscovered Kenyapithecus skeleton and its implications for the evolution ofpositional behavior in Miocene East African hominoids. J. Hum. Evol. 34, 657–664.
Napier, J.R., 1967. Evolutionary aspects of primate locomotion. Am. J. Phys.Anthropol. 27, 333–342.
Napier, J.R., 1993. Hands (revised edition). Princeton University Press, Princeton.Napier, J.R., Davis, P.R., 1959. The fore-limb skeleton and associated remains of
Proconsul africanus. Br. Mus. Nat. His. Fossil Mam. Afr. 16, 1–69.Napier, J.R., Napier, P.H.,1967. A Handbook of Living Primates. Academic Press, New York.Nieschalk, U., Demes, B., 1993. Biomechanical determinants of reduction of the
second ray in Lorisinae. In: Preuschoft, H., Chivers, D.J. (Eds.), Hands of Primates.Springer-Verlag, Wien, pp. 225–234.
Patel, B.A., 2008. Functional morphology and biomechanics of digitigrade handpostures in cercopithecoid primates. Ph.D. Dissertation, Stony Brook University.
Patel, B.A., 2009. Not so fast: speed effects on forelimb kinematics in cercopithecinemonkeys and implications for digitigrades postures in primates. Am. J. Phys.Anthropol. 140, 92–112.
Patel, B.A., in press. The interplay between speed, kinetics and hand postures duringprimate terrestrial locomotion. Am. J. Phys. Anthropol. doi:10.1002/ajpa.21138.
Patel, B.A., Rossie, J.B., Biechele, M., Hill, A., 2007. Functional morphology of theEquatorius africanus hand. J. Vert. Paleo. Suppl. 27, 128A.
Patel, B.A., Wunderlich, R.E., 2008. Speed effects on palmar pressure in digitigradebaboons (Papio anubis). Am. J. Phys. Anthropol. Suppl. 46, 168.
Patel, B.A., Wunderlich, R.E., Dynamic pressure patterns in the hands of olivebaboons (Papio anubis) during terrestrial locomotion: implications for cerco-pithecoid primate hand morphology. Anat Rec. (in review).
Pilbeam, D.R., 1996. Genetic and morphological records of the Hominoidea andhominid origins: a synthesis. Mol. Phyl. Evol. 5, 155–168.
Pilbeam, D.R., Rose, M.D., Barry, J.C., Shah, S.M.I., 1990. New Sivapithecus humeri fromPakistan and the relationship of Sivapithecus and Pongo. Nature 348, 237–239.
Pilbeam, D.R., Young, N.M., 2004. Hominoid evolution: synthesizing disparate data.C.R. Palevol. 3, 305–321.
Richmond, B.G., Begun, D.R., Strait, D.S., 2001. Origin of human bipedalism: theknuckle-walking hypothesis revisited. Yrbk. Phys. Anthropol. 44, 70–105.
Richmond, B.G., Fleagle, J.G., Kappelman, J., Swisher III, C.C., 1998. First hominoidfrom the Miocene of Ethiopia and the evolution of the catarhine elbow. Am. J.Phys. Anthropol. 105, 257–277.
Rose, M.D., 1983. Miocene hominoid postcranial morphology: monkey-like, ape-like, neither, or both? In: Ciochon, R.L., Corruccini, R.S. (Eds.), New Interpreta-tions of Ape and Human Ancestry. Plenum Press, New York, pp. 405–417.
Rose, M.D., 1988. Functional anatomy of the cheiridia. In: Schwartz, J.H. (Ed.), Orang-utan Biology. Oxford University Press, New York, pp. 299–310.
Rose, M.D., 1993. Locomotor anatomy of Miocene hominoids. In: Gebo, D.L. (Ed.),Postcranial Adaptations in Nonhuman Primates. University of Illinois Press,DeKalb, pp. 252–272.
Rose, M.D., 1994. Quadrupedalism in some Miocene catarrhines. J. Hum. Evol. 26,387–411.
Rose, M.D., 1997. Functional and phylogenetic features of the forelimb in miocenehominoids. In: Begun, D.R., Ward, C.V., Rose, M.D. (Eds.), Function, Phylogenyand Fossils: Miocene Hominoid Evolution and Adaptations. Plenum, New York,pp. 79–100.
Rose, M.D., Nakano, Y., Ishida, H., 1996. Kenyapithecus postcranial specimens fromNachola, Kenya. Afr. Stud. Monographs Suppl. 24, 3–56.
Schmitt, D., 1999. Compliant walking in primates. J. Zool. 248, 149–160.Schmitt, D., Hanna, J.B., 2004. Substrate alters forelimb to hind limb peak force
ratios in primates. J. Hum. Evol. 46, 239–254.Schultz, A.H., 1930. The skeleton of the trunk and limbs of higher primates. Hum.
Biol. 2, 303–438.Schultz, A.H., 1936. Characters common to higher primates and characters specific
to man. Q. Rev. Biol. 11, 259–283.Senut, B., Nakatsukasa, M., Kunimatsu, Y., Nakano, Y., Takano, T., Tsujikawa, H.,
Shimizu, D., Kagaya, M., Ishida, H., 2004. Preliminary analysis of Nacholapithecusscapula and clavicle from Nachola, Kenya. Primates 45, 97–104.
Sherwood, R.J., Ward, S., Hill, A., Duren, D.L., Brown, B., Downs, W., 2002. Prelimi-nary description of the Equatorius africanus partial skeleton (KNM-TH 28860)from Kipsaramon, Tugen Hills, Baringo District, Kenya. J. Hum. Evol. 42, 63–73.
Simons, E.L., Pilbeam, D.R., 1965. Preliminary revision of the Dryopithecinae (Pon-gidae, Anthropoidea). Folia. Primatol. 3, 81–152.
Smith, R.J., 2009. Use and misuse of the reduced major axis for line-fitting. Am. J.Phys. Anthropol. 140, 476–486.
Sokal, R.R., Rohlf, F.J., 1995. Biometry. W.H. Freeman and Co., New York.Stern Jr., J.T., Susman, R.L., 1983. The locomotor anatomy of Australopithecus afar-
ensis. Am. J. Phys. Anthropol. 60, 279–317.Susman, R.L., 1979. Comparative and functional morphology of hominoid fingers.
Am. J. Phys. Anthropol. 50, 215–235.Susman, R.L., Stern Jr., J.T., 1979. Telemetered electromyography of flexor digitorum
profundus and flexor digitorum superficialis in Pan troglodytes and implicationsfor interpretation of the O.H. 7 hand. Am. J. Phys. Anthropol. 50, 565–574.
Tuttle, R.H., 1967. Knuckle-walking and the evolution of hominoid hands. Am. J.Phys. Anthropol. 26, 171–206.
Tuttle, R.H., 1969. Knuckle-walking and the problem of human origins. Science 166,953–961.
Van Couvering, J.A.H., Van Couvering, J.A., 1976. Early Miocene mammal fossils fromEast Africa: aspects of geology, faunistics, and paleoecology. In: Isaac, G.L.,McCown, E.R. (Eds.), Human Origins: Louis Leakey and the East AfricanEvidence. W.A. Benjamin, Menlo Park, CA, pp. 155–207.
Walker, A., 1974. Locomotor adaptations in past and present prosimians. In:Jenkins, F.A. (Ed.), Primate Locomotion. Academic Press, New York, pp. 349–381.
Ward, C.V., 1998. Afropithecus, Proconsul, and the Primitive Hominoid skeleton. In:Strasser, E., Fleagle, J., Rosenberger, A., McHenry, H. (Eds.), Primate Locomotion:Recent Advances. Plenum Press, New York, pp. 337–352.
Ward, C.V., Walker, A.C., Teaford, M.F., Odhiambo, I., 1993. Partial skeleton ofProconsul nyanzae from Mfangano Island, Kenya. Am. J. Phys. Anthropol. 90,77–111.
Ward, S.C., Pilbeam, D.R., 1983. Maxillofacial morphology of Miocene hominoidsfrom Africa and Indo-Pakistan. In: Ciochon, R.S., Corruccini, R.L. (Eds.), NewInterpretations of Ape and Human Ancestry. Plenum Press, New York, pp.211–238.
Ward, S.C., Brown, B., Hill, A., 1996. Forelimb of Kenyapithecus africanus from theTugen Hills, Baringo District, Kenya. Am. J. Phys. Anthropol. Suppl. 22, 240.
Ward, S.C., Brown, B., Hill, A., Kelley, J., Downs, W., 1999. Equatorius: a new homi-noid genus from the Middle Miocene of Kenya. Science 285, 1382–1386.
Washburn, S.L., 1951. The analysis of primate evolution with particular reference tothe origin of man, Cold Spring Harbor. Symp. Quant. Biol. 15, 67–77.
Young, J.W., 2009. Substrate determines asymmetrical gait dynamics in marmosets(Callithrix jacchus) and squirrel monkeys (Saimiri boliviensis). Am. J. Phys.Anthropol. 138, 403–420.
Zar, J.H., 2009. Biostatistical Analysis, fifth ed. Prentice Hall, Upper Saddle River.




















