
Systemic induction of traumatic resin ducts and resin flow in Austrian pine by wounding and...
10
ORIGINAL ARTICLE Nicola Luchi Rui Ma Paolo Capretti Pierluigi Bonello Systemic induction of traumatic resin ducts and resin flow in Austrian pine by wounding and inoculation with Sphaeropsis sapinea and Diplodia scrobiculata Received: 11 March 2004 / Accepted: 30 August 2004 / Published online: 10 November 2004 Ó Springer-Verlag 2004 Abstract The potential role of the resin system in the response of Austrian pine (Pinus nigra Arn.) seedlings to mechanical injury and fungal infection was studied in greenhouse experiments. Anatomical observations were performed on 2-year-old plants wounded at collar level and inoculated with Sphaeropsis sapinea (Fr.: Fr.) Dyko & Sutton in Sutton or Diplodia scrobiculata (J. de Wet, B. Slippers & M. J. Wingfield, sp. nov.; sensu de Wet et al. 2003), two fungal pathogens that cause shoot blight and canker on conifers, and that are characterized by different levels of aggressiveness. Histological exam- ination of host tissue taken from the stem at 0, 8, and 12 cm above the treatment site revealed significant treatment- and time-dependent effects on the course of locally and systemically induced traumatic resin duct (TRD) development. Occurrence of TRDs was observed after 4 days only in seedlings inoculated with D. scro- biculata. At 12 days, TRDs were present also in mock- inoculated controls. No TRDs appeared in seedlings inoculated with S. sapinea. However, S. sapinea caused loss of vacuolar phenolics, severe disruption of cambial tissue and invaded the host xylem quickly and appar- ently unimpeded, whereas D. scrobiculata was never detected in the host xylem. Five-year-old Austrian pines subjected to the same stem base treatments were used to determine the resin mass flowing from the stem 30 cm above the treatment sites. Wounding and/or inoculation induced a significant, 8.3-fold average increase in sys- temic resin flow over the untreated trees 3 weeks after basal treatment, suggesting that wounding is the sole prerequisite for systemic induction of resin flow. The results are discussed in the context of current disease resistance models. Keywords Canker Fungal pathogen Pinus Resin Systemic induced resistance Traumatic resin ducts Abbreviations: ANOVA: Analysis of variance LSD: Least significant difference PB: Polyphenolic body SIR: Systemic induced resistance TRD: Traumatic resin duct WA: Water agar Introduction Trees, like other plants, defend themselves against pathogens by deploying a combination of constitutive (pre-existing) and induced (post-infectional), structural and biochemical mechanisms. Constitutive structural defenses such as cuticle, periderm, and rhytidome act as physical barriers to microbial penetration. Constitutive biochemical defenses include toxic substances such as antimicrobial phenolics, terpenoids, and hydrolytic en- zymes (Blanchette and Biggs 1992). Induced structural defenses include necrophylactic periderms (Blanchette and Biggs 1992) and cell wall appositions (e.g. Bonello et al. 1991). In conifers, no de novo-inducible biochemical de- fenses, such as true phytoalexins, are known (Bonello and Blodgett 2003). However, the expression of many constitutive biochemical defense mechanisms is en- hanced following microbial challenge. This is true at both local and systemic levels, although understanding of systemic induced resistance (SIR) phenomena is ra- ther limited in conifers (Bonello and Blodgett 2003). One of the first lines of induced biochemical defense to fungal or insect invasion in conifers is the exudation of constitutive resin around the attack site (Berryman 1972). Resin is postulated to act primarily as a wound N. Luchi R. Ma P. Bonello (&) Department of Plant Pathology, The Ohio State University, 2021 Coffey Road, Columbus, OH 43210, USA E-mail: [email protected] Fax: +1-614-292-4455 N. Luchi P. Capretti Dipartimento di Biotecnologie Agrarie—Sez. Patologia Vegetale, Piazzale delle Cascine, 28-50144 Firenze, Italy Planta (2005) 221: 75–84 DOI 10.1007/s00425-004-1414-3
Transcript of Systemic induction of traumatic resin ducts and resin flow in Austrian pine by wounding and...

ORIGINAL ARTICLE
Nicola Luchi Æ Rui Ma Æ Paolo Capretti
Pierluigi Bonello
Systemic induction of traumatic resin ducts and resin flow in Austrianpine by wounding and inoculation with Sphaeropsis sapineaand Diplodia scrobiculata
Received: 11 March 2004 / Accepted: 30 August 2004 / Published online: 10 November 2004� Springer-Verlag 2004
Abstract The potential role of the resin system in theresponse of Austrian pine (Pinus nigra Arn.) seedlings tomechanical injury and fungal infection was studied ingreenhouse experiments. Anatomical observations wereperformed on 2-year-old plants wounded at collar leveland inoculated with Sphaeropsis sapinea (Fr.: Fr.) Dyko& Sutton in Sutton or Diplodia scrobiculata (J. de Wet,B. Slippers & M. J. Wingfield, sp. nov.; sensu de Wetet al. 2003), two fungal pathogens that cause shootblight and canker on conifers, and that are characterizedby different levels of aggressiveness. Histological exam-ination of host tissue taken from the stem at 0, 8, and12 cm above the treatment site revealed significanttreatment- and time-dependent effects on the course oflocally and systemically induced traumatic resin duct(TRD) development. Occurrence of TRDs was observedafter 4 days only in seedlings inoculated with D. scro-biculata. At 12 days, TRDs were present also in mock-inoculated controls. No TRDs appeared in seedlingsinoculated with S. sapinea. However, S. sapinea causedloss of vacuolar phenolics, severe disruption of cambialtissue and invaded the host xylem quickly and appar-ently unimpeded, whereas D. scrobiculata was neverdetected in the host xylem. Five-year-old Austrian pinessubjected to the same stem base treatments were used todetermine the resin mass flowing from the stem 30 cmabove the treatment sites. Wounding and/or inoculationinduced a significant, 8.3-fold average increase in sys-temic resin flow over the untreated trees 3 weeks afterbasal treatment, suggesting that wounding is the sole
prerequisite for systemic induction of resin flow. Theresults are discussed in the context of current diseaseresistance models.
Keywords Canker Æ Fungal pathogen Æ Pinus Æ ResinSystemic induced resistance Æ Traumatic resin ducts
Abbreviations: ANOVA: Analysis of variance Æ LSD:Least significant difference Æ PB: Polyphenolicbody Æ SIR: Systemic induced resistance Æ TRD:Traumatic resin duct Æ WA: Water agar
Introduction
Trees, like other plants, defend themselves againstpathogens by deploying a combination of constitutive(pre-existing) and induced (post-infectional), structuraland biochemical mechanisms. Constitutive structuraldefenses such as cuticle, periderm, and rhytidome act asphysical barriers to microbial penetration. Constitutivebiochemical defenses include toxic substances such asantimicrobial phenolics, terpenoids, and hydrolytic en-zymes (Blanchette and Biggs 1992). Induced structuraldefenses include necrophylactic periderms (Blanchetteand Biggs 1992) and cell wall appositions (e.g. Bonelloet al. 1991).
In conifers, no de novo-inducible biochemical de-fenses, such as true phytoalexins, are known (Bonelloand Blodgett 2003). However, the expression of manyconstitutive biochemical defense mechanisms is en-hanced following microbial challenge. This is true atboth local and systemic levels, although understandingof systemic induced resistance (SIR) phenomena is ra-ther limited in conifers (Bonello and Blodgett 2003).
One of the first lines of induced biochemical defenseto fungal or insect invasion in conifers is the exudationof constitutive resin around the attack site (Berryman1972). Resin is postulated to act primarily as a wound
N. Luchi Æ R. Ma Æ P. Bonello (&)Department of Plant Pathology,The Ohio State University, 2021 Coffey Road, Columbus,OH 43210, USAE-mail: [email protected]: +1-614-292-4455
N. Luchi Æ P. CaprettiDipartimento di Biotecnologie Agrarie—Sez. PatologiaVegetale, Piazzale delle Cascine, 28-50144 Firenze, Italy
Planta (2005) 221: 75–84DOI 10.1007/s00425-004-1414-3

cleanser and sealant (Croteau and Johnson 1985; Phil-lips and Croteau 1999), but has been demonstrated toalso have antimicrobial activity (Shrimpton and Whit-ney 1968) and to possess direct toxic properties againstseveral insect guilds (e.g. Klepzig et al. 1995). Resin issynthesized in specialized secretory structures called‘resin cells’ and ‘resin ducts’. Resin cells are common infir (Abies spp.), cedar (Cedrus spp.) and hemlock (Tsugaspp.); resin ducts are characteristic of spruce (Piceaspp.), pine (Pinus spp.), larch (Larix spp.), and Douglas-fir (Pseudotsuga spp.; Hudgins et al. 2003). Resin ductsare further differentiated into two groups, ‘radial ducts’and ‘axial ducts’, depending on their orientation in theplant tissues. Radial ducts are generally more commonin the phloem, while axial ducts are usually scattered inthe xylem (Wu and Hu 1997; Hudgins et al. 2003).
For some conifer species, it is known that microbialinvasion, and wounding caused by insects or traumaticevents such as frost, as well as application of methyljasmonate (Bannan 1936; Franceschi et al. 2000, 2002;Nagy et al. 2000; Martin et al. 2002; Hudgins et al.2004), lead to local formation of ‘traumatic resin ducts’(TRDs), which produce secondary resin (Wong andBerryman 1977). TRDs are usually found in a single rowclose to the cambial zone (Bannan 1936; Franceschiet al. 2000; Nagy et al. 2000). In some cases, secondaryresin has been demonstrated to be more fungistatic thanconstitutive resin (Solheim 1991).
Studies carried out on Norway spruce (Picea abies L.)also showed that wounding and fungal infections induceTRD formation in tissues remotely located from theoriginal induction site (Franceschi et al. 2000; Nagyet al. 2000; Krekling et al. 2004). This demonstrates thatspruce possesses systemically inducible signaling path-ways that result in major anatomical reorganization.Whether systemic induction of TRDs or systemic effectsof fungal infection on resin flow occur in pine is notknown at present.
Recent studies have demonstrated that Sphaeropsissapinea (Fr.: Fr.) Dyko & Sutton in Sutton (ex Diplodiapinea) and Diplodia scrobiculata (J. de Wet, B. Slippers &M. J. Wingfield, sp. nov.; sensu de Wet et al. 2003) caninduce systemic resistance (SIR) to S. sapinea in Aus-trian pine (Pinus nigra Arn.; Blodgett et al. 2003a), andthat this phenomenon is associated with changes in se-lected pathways of phenolic metabolism (Bonello andBlodgett 2003). Both of these pathogens are mainly rainsplash-dispersed, but associations with putative insectvectors are also known (Feci et al. 2002). S. sapinea is aserious pathogen that causes significant damage onconifer species throughout the world by inciting shootblight, crown wilt, and canker diseases on plants rangingfrom seedling to mature tree size (Swart and Wingfield1991; Stanosz et al. 1996; Maresi et al. 2002). While S.sapinea is an aggressive shoot-blight pathogen, D. scro-biculata is non-aggressive on shoots of several conifers(Blodgett and Stanosz 1997, 1999), including Austrianpine. However, it can cause significant cankers on mainstems, although these cankers are always significantly
smaller than those caused by S. sapinea (Blodgett andBonello 2003). This aggressiveness differential betweenclosely related fungal pathogens provides a useful toolfor testing mechanistic hypotheses that attempt to ex-plain disease resistance in pine.
As an extension of earlier research, the study reportedhere tested the hypothesis that systemic induction ofTRDs and resin flow is positively correlated with theSIR phenotype. Induction of qualitative or quantitativechanges in terpenoid composition and resin biochemis-try were not the focus of this study, even though thesefactors might be important in conifer defense.
Materials and methods
Anatomy of resin system
Plant material In early spring 2003, 200 two-year-oldseedlings of Austrian pine (Pinus nigra Arn.) were pur-chased from the Ohio Department of Natural Re-sources’ Marietta Forest Nursery in Marietta, OH. Theseed originated from open-pollinated plants (mixed ge-netic background) and came from Turkey. The seedlingshad been stored at 4�C at the nursery and were kept atthat temperature until they were individually planted on20 March 2003 into 7.6-l plastic pots containing an or-ganic potting mix of pine bark and ComTil (Columbus,OH; municipal sewage sludge prepared by the aeratedstatic pile method; Finstein et al. 1983; 3:1, v:v). Seed-lings were maintained outdoors on drip irrigation untiluse. Two weeks before treatments were applied, 40seedlings were selected based on uniformity of size andhealth, and transferred to the greenhouse. The green-house temperature and relative humidity were measuredevery 5 min during the experiment and were26.4±0.03�C (error statistic here and thereafter isstandard error of the mean) and 63.9±0.17%, respec-tively. To avoid water stress effects the pots were wa-tered twice daily to field capacity (Blodgett and Bonello2003). The seedlings had a mean stem height of31.57±0.50 cm and a mean stem diameter of0.66±0.02 cm.
Inoculation and sampling procedures Twenty seedlingsand the following four treatments (five experimentalunits/treatment) were used in each of two experiments:inoculation with Sphaeropsis sapinea; inoculation withDiplodia scrobiculata; inoculation with 1.5% water agar(WA; mock inoculation); and unwounded controls. Thefirst experiment was started on 19 May 2003, the secondexperiment on 29 May 2003.
All seedlings, except those assigned to the un-wounded control treatment, were wounded on the basalportion of the stem (5 cm above soil level), down to thecambial area, by removing a 4-mm-diameter plug ofbark using a cork borer dipped in 95% ethanol. Three-millimeter-diameter mycelial plugs were taken from themargins of actively growing, 7-day-old WA cultures of
76

monoconidial isolates of S. sapinea and D. scrobiculata(Blodgett and Bonello 2003) using a flame-sterilized corkborer, and were inserted into the wounds of treesbelonging to the corresponding treatment groups. Un-colonized WA plugs were used in the mock inoculations.All treatment sites were sealed with Parafilm (AmericanNational Can Co., Chicago, IL, USA) to preventexcessive desiccation and limit entry by contaminants.
Tissue processing for microscopy Four days (firstexperiment) and 12 days (second experiment) aftertreatments were applied, tissue samples were collected byremoving a 1.5-cm stem section from each of three sitesalong the stem: 0 cm (inoculation site), 8 and 12 cmabove the treatment site. Corresponding segments wereremoved from untreated controls. Each stem segmentwas cut longitudinally along two orthogonal, radialplanes into four equivalent pieces using a razor blade.One of the planes cut through the middle of the originalwound, so that the lesion produced by wounding and/orinoculations could be measured to the nearest millimeteron the exposed lesion profile. Lesion size was onlymeasured above the wound, starting at the margin of thewound. One of the two pieces containing the treatmentwound was randomly selected, together with one of thefour pieces taken at 8 and 12 cm. All selected stem pieceswere processed for fixation and resin embedding using amodification of the method of Nagy et al. (2000). Allsteps were carried out at room temperature unlessotherwise indicated. Briefly, samples were placed in 2-ml, screw-capped microfuge tubes containing a fixativesolution of 2% paraformaldehyde and 1.25% glutaral-dehyde in 50 mM PIPES buffer (pH 7.2) with10 drops l�1 of Tween 20 (Sigma, Aldrich). Tissues werefixed for 24 h, rinsed in buffer, and then dehydrated inan ethanol series (70–80–90–95–100%; each step 30 min,with the step at 100% repeated four times). Each samplewas then placed in a 1.5-ml microfuge tube and infil-trated with L.R. White acrylic resin (Ladd Research,Williston, VT, USA), with two resin changes (1 h each).The final infiltration was carried out for 12 h at roomtemperature and the samples were polymerized at 60�Cfor 20 h.
The embedded pine tissues were cut along a planeorthogonal to the stem axis by using a rotary microtome(AO Spencer n.820 Rotary Microtome). Randomlypicked, 10-lm-thick sections were stained with 0.05%toluidine blue O in sodium phosphate buffer (0.1 M;pH 6.5) on a glass slide placed on a hot plate at 60�C fora few minutes, rinsed, and mounted in deionized water(Bonello et al. 1991). Digital photographs of stainedsections were taken using a Nikon 990 Coolpix camerafitted to the eyepiece socket of a compound microscope.
Histological descriptions Microphotographs were usedfor histological descriptions and to estimate resin ductfrequencies in the different treatments. Frequencies wereestimated both in terms of cumulative number ofoccurrences (presence/absence) at each stem locationand in terms of number of resin ducts in reference to a
standard sample surface area. This standard samplesurface area was 25% of the stem cross-section and wasconsidered essentially constant across seedlings (approx.8.6 mm2), given the extremely small variance in stemdiameter. Thus, the number of resin ducts on each sec-tion was considered automatically normalized for sur-face area. A similar approach was used to estimate thedensity of cortical parenchyma cells containing phenolicbodies (see Results) in the same sections.
Resin-flow analysis
Plant material The small size of 2-year-old seedlingsprevented their use in resin-flow studies that involvemechanical wounding of the phloem. Thus, 5-year-oldAustrian pines were used. The plants (seed from open-pollinated families in Turkey) were obtained from RidgeManor Nursery (Madison, OH, USA) in early spring2003 and were grown in their original organic mix (5%hardwood bark, 60% pine bark and 35% peatmoss) in11-l plastic pots. At the time the first experiment wasconducted (see below) the pines had a mean height of108.33±1.92 cm and mean stem diameter of2.80±0.05 cm at 10 cm from the soil surface. They wereplaced in a greenhouse with a mean temperature of24±0.03�C and mean relative humidity of 76±0.15%(variables were monitored every 5 min during theexperiment). To exclude water stress as a factor, treeswere watered twice daily to field capacity (Blodgett andBonello 2003).
Inoculation and sampling procedures Plugs of S. sapineaand D. scrobiculata mycelium were cut from the marginsof actively growing colonies using a 12-mm-diametercork borer and were used as fungal inocula.
Twenty-four Austrian pines were assigned to each oftwo independent trials. One trial was initiated on 16 July2003, the other on 1 September 2003. The same fourtreatments used for the microscopy study were used ineach trial: inoculation with S. sapinea, inoculation withD. scrobiculata, inoculation with WA, and unwoundedcontrol. In the first three treatments, each tree waswounded at three equally spaced locations around thestem, 10 cm above the soil surface, by removing theouter bark and phloem using a 15-mm-diameter corkborer, and inoculated with plugs of either S. sapinea, D.scrobiculata, or WA. All trees, except in the unwoundedtreatment, were wrapped in Parafilm at the inoculationsites to reduce contamination and prevent excessivedrying.
To conform with previous studies on systemicinduction of secondary metabolism and associatedresistance (Blodgett et al. 2003a; Bonello and Blodgett2003), trees were incubated for 3 weeks, at which timethe outer bark and phloem were removed 30 cm abovethe induction treatments by using a 10-mm-diametercork borer. A V-shaped aluminum trough was fastenedjust below each punched hole using tacks. A pre-
77

weighed, 15-ml, conical plastic test tube was placedunderneath the aluminum trough to collect the resin.Tubes were collected and weighed 24 h after installa-tion. The resin was taken from the xylem–phloeminterface, because this is the area of main fungal col-onization. The lengths of the lesions above each inoc-ulation site were also measured (Blodgett and Bonello2003) and an average lesion length was calculated foreach tree.
Pathogen re-isolations We have isolated hundreds ofsamples of Austrian pine tissues inoculated with S.sapinea and D. scrobiculata in the greenhouse over thepast few years but have isolated the pathogens fromcontrol inoculations or untreated trees only rarely(Blodgett and Bonello 2003). Therefore, in this studypathogen re-isolation was attempted only at the end ofthe second resin-flow trial. Small pieces of tissue frominoculated areas, as well as corresponding areas oncontrol trees, were removed and processed using thetannic acid method as described by Blodgett et al.(2003b) to confirm or exclude the presence of thepathogen.
Statistical analysis
Data were analyzed using SPSS 11.5 for Windows. v2
tests were used to estimate the association betweentreatment and frequency of TRDs (presence/absence) ineach stem section.
Analysis of variance (ANOVA) of raw or squareroot-transformed data was used to test effects of treat-ments (fungal inoculation, mock inoculation, un-wounded control), date (4 and 12 days after treatment),and distance from treatment (0, 8, and 12 cm) on thenormalized density of TRDs in the xylem and phenolicbody-containing cells in cortical tissues. Post-ANOVAmean separation was conducted using the Least Signif-icant Difference (LSD) test.
Because the assumptions of normality and variancehomogeneity could not be satisfied by either the raw ortransformed data, differences in mean resin flow amongtreatments were tested using the non-parametric, uni-variate Kruskal–Wallis (KW) test.
When analyses showed no significant differences(P>0.05) between levels of a factor (e.g. different trials),data sets were combined.
Results
Pathogen re-isolation
The frequency of pathogen recovery from 18 sampledlocations in each treatment (three per tree) was 100% forS. sapinea (18/18) and 89% for D. scrobiculata (16/18).Neither pathogen was recovered from any mock inocu-lation or unwounded control treatment.
Anatomy of control seedlings
Transverse sections from three locations on the stems ofunwounded (control) seedlings were used as reference(normal stem anatomy), with samples taken at both timepoints (4 and 12 days after the start of the experiment,respectively). At 4 days, the cortex consisted of irregularlyshaped parenchymatous cells. These cells often had whatappeared to be yellow–green–brown vacuolar bodies intheir cytoplasm (Fig. 1a). These vacuoles likely containedsoluble or insoluble phenolic or polyphenolic substances(Franceschi et al. 2000; Nagy et al. 2000; Krekling et al.2004), and therefore we refer to them as ‘Phenolic Bodies’(PBs), and the cells containing them as ‘PB cells’. Con-stitutive resin ducts in the cortex had swollen epithelialcells surrounded by small parenchymatous cells contain-ing PBs (Fig. 1b). Constitutive resin ducts were neverobserved in the secondary phloem. In the secondaryphloem we observed normal phloem elements and radialparenchyma ray cells with large nuclei. The cambial zonehad regular rows of small rectangular cells (Fig. 1c). Thexylem showed regular rows of tracheids, parenchyma raysand a scattered distribution of constitutive axial resinducts with swollen epithelial cells (Fig. 1d). The anatomyof control seedlings at 12 days from inoculation was en-tirely consistent with that observed after 4 days.
Anatomy after treatment
Seedlings inoculated with D. scrobiculata
Four days post inoculation, all pine seedlings had anecrotic band (3±0.1 mm) around the wound,accompanied by enhanced resinosis. Cortex, cambium,and secondary phloem from all three sites had similaranatomy to that described for seedlings inoculated
Fig. 1a–l Constitutive and traumatic resin ducts in control andinoculated Austrian pine (Pinus nigra) seedlings. a Cortical cells incontrol seedlings showing accumulation of phenolic bodies (arrow).b Constitutive resin duct (rdl) in the cortex of a control seedlingshowing PBs in the surrounding cells. c Appearance of the cambialarea in control seedlings. d Constitutive axial resin duct in thesecondary xylem of a control seedling. Note the non-lignifiednature of the resin duct cells (magenta color indicates acidicpolysaccharides) and the lignified walls of the tracheids (blue–greencoloration). e,f Early stage formation of TRD cell clusters (stage 1,arrows) in the mother xylem area of a seedling inoculated withDiplodia scrobiculata (J. de Wet, B. Slippers & M. J. Wingfield, sp.nov.; sensu de Wet et al. 2003). The blue cytoplasm coloration ofthe cluster cells is due to the presence of large nuclei. g,h Stage-2TRDs (arrows) in seedlings inoculated with D. scrobiculata. i,jStage-3 TRDs in seedlings inoculated with D. scrobiculata.k Disruption of host tissue in the cambial area of a seedlinginoculated with Sphaeropsis sapinea (Fr.: Fr.) Dyko & Sutton inSutton. Fungal hyphae are visible in the disrupted area. l Fungalhyphae arranged around the lumen periphery of a constitutive axialresin duct in the xylem of a seedling inoculated with S. sapinea. cCambial area, cx cortex, h hypha, n host cell nucleus, rdlconstitutive resin duct lumen, sp secondary phloem, sx secondaryxylem, trdl traumatic resin duct lumen. Bars = 25 lm (a,d,f–h,j–l),100 lm (b,c,e,i)
c
78

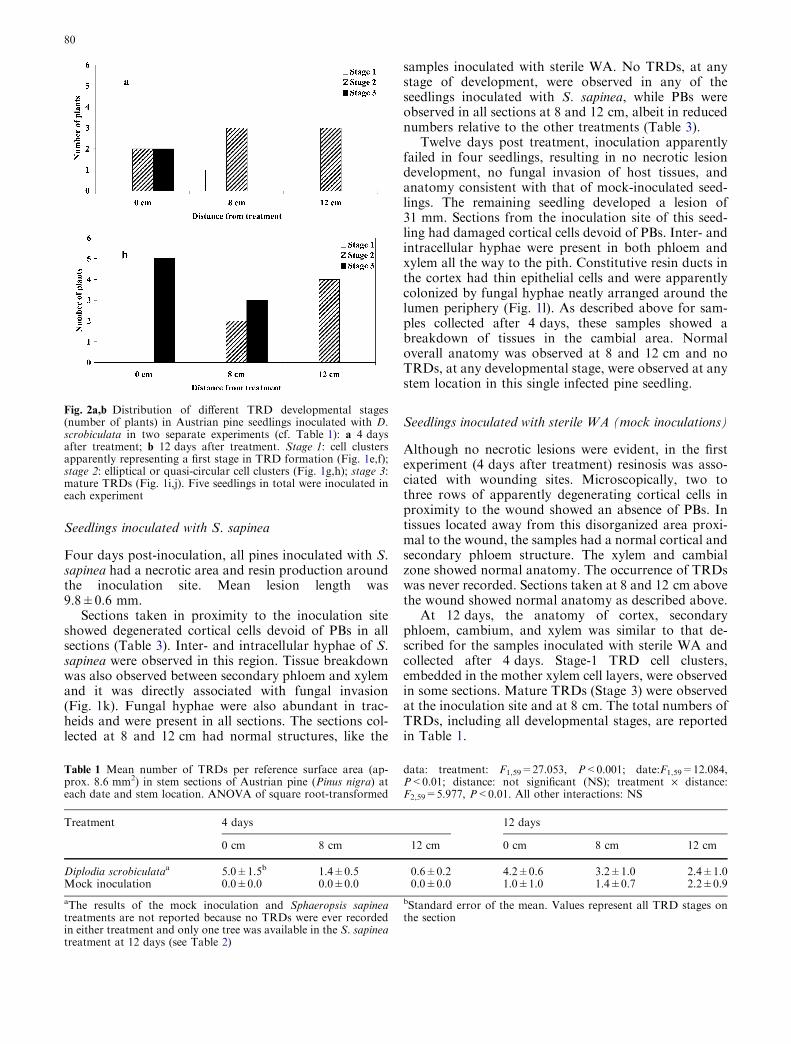
with sterile WA. However, the mother xylem con-tained irregularly shaped cell aggregations that wereabsent from mock-inoculated trees and unwoundedcontrols. These cell aggregations were found at allthree stem sites. Such cells had large nuclei and wereseparated along irregular planes. These cell clustersapparently represent a first stage in TRD formation(stage 1; Fig. 1e,f). At apparently later stages, thesestructures developed into elliptical or quasi-circularcell clusters (stage 2; Fig. 1g,h). Finally, these cellclusters developed into what appeared to be matureTRDs (stage 3), with swollen epithelial cells containingswollen nuclei, surrounding small resin duct lumina(Fig. 1i,j). These mature TRDs were smaller than theconstitutive axial resin ducts and were embedded inthe mother xylem. Stages 1 and 2 were also observedin sections taken at 8 and 12 cm from the inoculation
site, but stage 3 was only present at the inoculationsite (Fig. 2a). No fungal hyphae were ever observedgrowing inter- or intracellularly in any of the sectionsfrom any of the three stem locations.
Twelve days post inoculation, all pines had a ne-crotic area (mean lesion length 13±0.7 mm) and resinproduction around the inoculation site. The generalanatomy of cortex, secondary phloem, cambium, andxylem was similar to that described for the samplesinoculated with D. scrobiculata collected after 4 days.However, all five samples from the inoculation site hadonly mature TRDs (Fig. 2b). Furthermore, the numberof TRDs at various stages was higher across all threesampling sites (Table 1) than in samples inoculatedwith sterile WA, as well as samples inoculated with D.scrobiculata and collected after 4 days (first experi-ment).
79
Seedlings inoculated with S. sapinea
Four days post-inoculation, all pines inoculated with S.sapinea had a necrotic area and resin production aroundthe inoculation site. Mean lesion length was9.8±0.6 mm.
Sections taken in proximity to the inoculation siteshowed degenerated cortical cells devoid of PBs in allsections (Table 3). Inter- and intracellular hyphae of S.sapinea were observed in this region. Tissue breakdownwas also observed between secondary phloem and xylemand it was directly associated with fungal invasion(Fig. 1k). Fungal hyphae were also abundant in trac-heids and were present in all sections. The sections col-lected at 8 and 12 cm had normal structures, like the
samples inoculated with sterile WA. No TRDs, at anystage of development, were observed in any of theseedlings inoculated with S. sapinea, while PBs wereobserved in all sections at 8 and 12 cm, albeit in reducednumbers relative to the other treatments (Table 3).
Twelve days post treatment, inoculation apparentlyfailed in four seedlings, resulting in no necrotic lesiondevelopment, no fungal invasion of host tissues, andanatomy consistent with that of mock-inoculated seed-lings. The remaining seedling developed a lesion of31 mm. Sections from the inoculation site of this seed-ling had damaged cortical cells devoid of PBs. Inter- andintracellular hyphae were present in both phloem andxylem all the way to the pith. Constitutive resin ducts inthe cortex had thin epithelial cells and were apparentlycolonized by fungal hyphae neatly arranged around thelumen periphery (Fig. 1l). As described above for sam-ples collected after 4 days, these samples showed abreakdown of tissues in the cambial area. Normaloverall anatomy was observed at 8 and 12 cm and noTRDs, at any developmental stage, were observed at anystem location in this single infected pine seedling.
Seedlings inoculated with sterile WA (mock inoculations)
Although no necrotic lesions were evident, in the firstexperiment (4 days after treatment) resinosis was asso-ciated with wounding sites. Microscopically, two tothree rows of apparently degenerating cortical cells inproximity to the wound showed an absence of PBs. Intissues located away from this disorganized area proxi-mal to the wound, the samples had a normal cortical andsecondary phloem structure. The xylem and cambialzone showed normal anatomy. The occurrence of TRDswas never recorded. Sections taken at 8 and 12 cm abovethe wound showed normal anatomy as described above.
At 12 days, the anatomy of cortex, secondaryphloem, cambium, and xylem was similar to that de-scribed for the samples inoculated with sterile WA andcollected after 4 days. Stage-1 TRD cell clusters,embedded in the mother xylem cell layers, were observedin some sections. Mature TRDs (Stage 3) were observedat the inoculation site and at 8 cm. The total numbers ofTRDs, including all developmental stages, are reportedin Table 1.
Treatment 4 days 12 days
0 cm 8 cm 12 cm 0 cm 8 cm 12 cm
Diplodia scrobiculataa 5.0±1.5b 1.4±0.5 0.6±0.2 4.2±0.6 3.2±1.0 2.4±1.0Mock inoculation 0.0±0.0 0.0±0.0 0.0±0.0 1.0±1.0 1.4±0.7 2.2±0.9
Fig. 2a,b Distribution of different TRD developmental stages(number of plants) in Austrian pine seedlings inoculated with D.scrobiculata in two separate experiments (cf. Table 1): a 4 daysafter treatment; b 12 days after treatment. Stage 1: cell clustersapparently representing a first stage in TRD formation (Fig. 1e,f);stage 2: elliptical or quasi-circular cell clusters (Fig. 1g,h); stage 3:mature TRDs (Fig. 1i,j). Five seedlings in total were inoculated ineach experiment
aThe results of the mock inoculation and Sphaeropsis sapineatreatments are not reported because no TRDs were ever recordedin either treatment and only one tree was available in the S. sapineatreatment at 12 days (see Table 2)
bStandard error of the mean. Values represent all TRD stages onthe section
Table 1 Mean number of TRDs per reference surface area (ap-prox. 8.6 mm2) in stem sections of Austrian pine (Pinus nigra) ateach date and stem location. ANOVA of square root-transformed
data: treatment: F1,59=27.053, P<0.001; date:F1,59=12.084,P<0.01; distance: not significant (NS); treatment · distance:F2,59=5.977, P<0.01. All other interactions: NS
80
Cumulative TRD frequencies
The morphological analyses described above showedthat no TRDs were ever recorded in unwounded controland S. sapinea-inoculated seedlings. Cumulative fre-quencies and statistical analysis of TRD occurrences interms of presence/absence at each stem location are re-ported in Table 2. The analysis of contingency tablesdemonstrates that treatment was a significant factor inthe appearance of TRDs in two of four treatments,whether data were grouped by distance from treatmentsite or date.
Density of PB cells
Four days after treatment, all sections from the inocu-lation site (0 cm) of seedlings treated with S. sapineashowed a complete absence of PBs; all other sectionscontained PB cells in various densities (Table 3).ANOVA revealed significant effects of treatment but notdistance from treatment site, and a significant treatment· distance interaction (Table 3). Post-hoc mean separa-tion by LSD analysis showed that the S. sapinea treat-
ment contained fewer PB cells than any other treatment,whereas the other treatments did not significantly differfrom each other (Table 3). No significant differences indensity of PB-containing cortical cells were observedamong the D. scrobiculata, mock-inoculation, and un-wounded treatments at 12 days at any of the threelocations (data not shown).
Resin flow
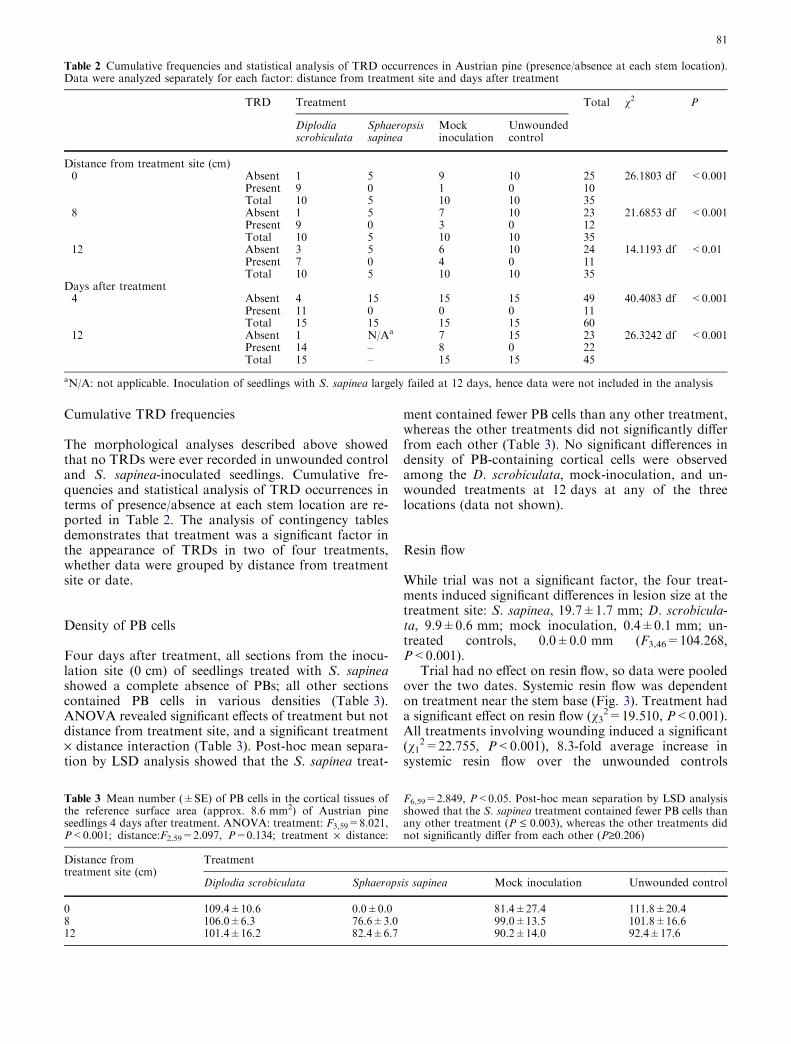
While trial was not a significant factor, the four treat-ments induced significant differences in lesion size at thetreatment site: S. sapinea, 19.7±1.7 mm; D. scrobicula-ta, 9.9±0.6 mm; mock inoculation, 0.4±0.1 mm; un-treated controls, 0.0±0.0 mm (F3,46=104.268,P<0.001).
Trial had no effect on resin flow, so data were pooledover the two dates. Systemic resin flow was dependenton treatment near the stem base (Fig. 3). Treatment hada significant effect on resin flow (v3
2=19.510, P<0.001).All treatments involving wounding induced a significant(v1
2=22.755, P<0.001), 8.3-fold average increase insystemic resin flow over the unwounded controls
Table 2 Cumulative frequencies and statistical analysis of TRD occurrences in Austrian pine (presence/absence at each stem location).Data were analyzed separately for each factor: distance from treatment site and days after treatment
TRD Treatment Total v2 P
Diplodiascrobiculata
Sphaeropsissapinea
Mockinoculation
Unwoundedcontrol
Distance from treatment site (cm)0 Absent 1 5 9 10 25 26.1803 df <0.001
Present 9 0 1 0 10Total 10 5 10 10 35
8 Absent 1 5 7 10 23 21.6853 df <0.001Present 9 0 3 0 12Total 10 5 10 10 35
12 Absent 3 5 6 10 24 14.1193 df <0.01Present 7 0 4 0 11Total 10 5 10 10 35
Days after treatment4 Absent 4 15 15 15 49 40.4083 df <0.001
Present 11 0 0 0 11Total 15 15 15 15 60
12 Absent 1 N/Aa 7 15 23 26.3242 df <0.001Present 14 – 8 0 22Total 15 – 15 15 45
aN/A: not applicable. Inoculation of seedlings with S. sapinea largely failed at 12 days, hence data were not included in the analysis
Table 3 Mean number (±SE) of PB cells in the cortical tissues ofthe reference surface area (approx. 8.6 mm2) of Austrian pineseedlings 4 days after treatment. ANOVA: treatment: F3,59=8.021,P<0.001; distance:F2,59=2.097, P=0.134; treatment · distance:
F6,59=2.849, P<0.05. Post-hoc mean separation by LSD analysisshowed that the S. sapinea treatment contained fewer PB cells thanany other treatment (P £ 0.003), whereas the other treatments didnot significantly differ from each other (P‡0.206)
Distance fromtreatment site (cm)
Treatment
Diplodia scrobiculata Sphaeropsis sapinea Mock inoculation Unwounded control
0 109.4±10.6 0.0±0.0 81.4±27.4 111.8±20.48 106.0±6.3 76.6±3.0 99.0±13.5 101.8±16.612 101.4±16.2 82.4±6.7 90.2±14.0 92.4±17.6
81

(Fig. 3), with no significant differences among the mock-inoculated, D. scrobiculata, and S. sapinea treatments.
Discussion
Our study describes a novel phenomenon in which theconstitutive resin system of pines is augmented system-ically by abiotic and biotic factors. Greatly enhanced (8-fold) systemic resin flows of comparable magnitudeswere achieved by wounding, alone and in combinationwith infection by fungal pathogens of different aggres-siveness. Thus, it appears that wounding is sufficient forthis enhanced resin flow. However, the histological datasuggest that wounding alone and wounding plus infec-tion result in different morphogenetic trajectories for thetraumatic resin system, and these differ according to theaggressiveness of the pathogen. TRDs were visible just4 days after infection with D. scrobiculata at all threelocations along the stem, up to 12 cm away from theinfection court. In contrast, TRDs appeared only12 days after mock inoculation and the available seed-lings inoculated with S. sapinea showed no TRD for-mation. This lack of TRD formation was correlated withextensive tissue invasion and disruption by the pathogenin S. sapinea-inoculated seedlings, whereas no fungalhyphae were observed in tissues of D. scrobiculata-inoculated seedlings. The timing of TRD induction byD. scrobiculata observed in our study is comparable tothe timing observed in Norway spruce. Nagy et al.(2000) observed formation of early-stage TRDs 9 dayspost-inoculation with Ceratocystis polonica (Siem.) C.Moreau (a bark beetle-associated wilt pathogen ofNorway spruce) and their development into matureTRDs after 18 days.
Our observations on the presence/absence of PBs isconsistent with a recent study involving infection of
Norway spruce with the root and butt rot pathogenHeterobasidion annosum (Fr. Bref.), in which fungalinoculation induced cellular disorganization in thephloem resulting in discharge of the vacuolar content ofpolyphenolic parenchyma cells onto the cell walls(Krekling et al. 2004). Although not in the same detail,we have also documented localized disappearance ofPBs from disorganized cells when the host is infectedwith S. sapinea, as well as a systemic reduction of thenumber of PB cells in the same seedlings, compared withthe other treatments (Table 3). In addition to this evi-dence for mobilization of phenolics within the host cells,previous studies have also shown that infection ofAustrian pine with S. sapinea elevates the concentrationsof soluble phenolics in the phloem, both locally andsystemically (Bonello and Blodgett 2003). The signifi-cance of the PB response to S. sapinea in Austrian pine[functionally a hypersensitive response (HR)—Ham-merschmidt and Nicholson 1999], is unclear at present,as S. sapinea is a necrotrophic pathogen unlikely to besignificantly affected by HR, and its growth rate doesnot seem to be negatively affected by elevated concen-trations of soluble phenolics in infected seedlings (Bo-nello and Blodgett 2003).
In order to understand a plant’s defense responses toa pathogen, for example to identify resistance genes, it isnecessary to select for host lines that are either resistantor susceptible. Such lines are generally not available forconifers, with the notable exception of some pine speciesthat possess major resistance genes to rust diseases (e.g.Kinloch et al. 1999). In the absence of such host popu-lations, as with Austrian pine, the use of pathogenpopulations differing in aggressiveness may be a usefulsubstitute to identify host defense responses that may becritical in disease resistance. Hence, our decision tocompare host responses to S. sapinea and D. scrobicu-lata, two closely related pathogens that were consideredthe same species until recently (de Wet et al. 2003), butare characterized by different levels of aggressiveness.Our results suggest the traumatic resin system is animportant component in pine resistance to S. sapineaand D. scrobiculata, because we found a negative cor-relation between formation of TRDs and actual hosttissue invasion by the fungi. However, our study alsosuggests that resin flow may not be a good predictor ofhost resistance against these canker pathogens.Wounding alone and wounding plus fungal infectionwere equally effective in the induction of systemicallyenhanced resin flow, but wounding alone does not in-duce SIR in Austrian pine (Blodgett et al. 2003a) orMonterey pine (Bonello et al. 2001). If resin flow playeda central role in resistance, SIR would be equally ex-pressed in all treatments.
If resin plays a role in Austrian pine defense againstcanker pathogens, as suggested by the negative correla-tion between host tissue invasion and production ofTRDs, timing of induction of the resin system, ratherthan resin flow, may be crucial in the host resistanceresponse. Timing, rather than the magnitude of the
Fig. 3 Systemic resin flow from 5-year-old Austrian pine saplingswounded and treated near the stem base with WA (Mockinoculated), one of two fungal pathogens, or left untreated(Unwounded). The chart shows data combined over the two trials.Treatments involving wounding did not differ from each other, butcollectively induced an 8.3-fold average increase in resin flow overthe untreated controls. Different letters above bars indicatesignificantly different systemic resin flows (P<0.001). Error barsare standard errors of the means
82

response, is often the critical factor in host–pathogeninteractions (Walton 1997). Thus, early production ofTRDs against D. scrobiculata may explain the host’shigher resistance against this pathogen, particularly iftraumatic resin is more antimicrobial than constitutiveresin (Solheim 1991; Blodgett and Stanosz 1997), ahypothesis we will test in future investigations.
Due to limitations of the experimental system usedhere, it is presently impossible to state with confidencewhether systemic enhancement of resin flow is the resultof systemic TRD induction or augmentation of resinflow from the constitutive resin system. However, in-duced enzyme activities and transcript accumulation forgenes involved in terpenoid biosynthesis in Norwayspruce and Sitka spruce were closely associated withtraumatic resinosis and with induced increase of resinformation (Martin et al. 2002, 2003; Faldt et al. 2003;McKay et al. 2003). Induced enzyme and gene activitiescould also be a mechanism for induced resin flow in theAustrian pine system described here.
While no TRDs were observed in the S. sapinea ormock-inoculated treated seedlings at 4 days, TRDs wereobserved systemically in mock-inoculated seedlings at12 days (Tables 1, 2), and no conclusive statements canbe made for S. sapinea-treated seedlings at 12 days. Thissuggests that all treatments may be able to elicit TRDformation, but that D. scrobiculata elicited the mostrapid host response, followed by mock inoculation andperhaps S. sapinea. Because resin flow was measured inlarger plants 3 weeks after treatment, it is possible thatdelayed TRD formation in the mock inoculation treat-ment, and perhaps in the S. sapinea treatment, resultedin equalization of resin flows observed in that timeframe. This suggests that increased flow is due to sec-ondary resin produced in TRDs; however, a partial roleby activated constitutive cortical, radial, and xylemducts cannot be excluded at this point.
The anatomical observations on D. scrobiculata-in-fected seedlings indicate a time-dependent inductionprocess in TRD formation, in which a signal originatingat the infection site propagates up the stem and inducesdifferentiation of mother xylem cells into TRDs(Fig. 2a,b). This finding is consistent with recent evi-dence for a similar phenomenon in Norway spruce(Krekling et al. 2004), in which a putative signal triggersthe propagation of a developmental wave in the axialdirection at about 2.5 cm per day. This evidence, cou-pled with our data on systemically reduced PB cells in S.sapinea-infected seedlings (Table 3) and work showingsystemic induction of phenolic metabolism (Bonello andBlodgett 2003) and associated expression of SIR(Blodgett et al. 2003a), suggests that Austrian pine’sresponses to canker pathogens are mediated by endog-enous signals, resembling processes known to occur inherbaceous plants. While strong evidence exists for aputative signaling role of salicylic and jasmonic acids,and other molecules, in the SIR of herbaceous hosts(Metraux 2001), the signaling mechanisms in conifersare unknown at present. However, evidence shows that
both salicylic and jasmonic acids can be inducers ofdefense responses when applied exogenously in highconcentrations to conifer tissues (e.g. Davis et al. 2002;Hudgins et al. 2004).
Whether or not resin plays a role in host defenseagainst canker pathogens, systemically enhanced resinflows induced by wounding and fungal infection mayhave important consequences in the host’s response toinsect attack, particularly against bark- and xylem-inhabiting insects such as bark beetles and wood borers.Indeed, ‘‘pitching out’’ of bark beetles is considered oneof the fundamental mechanisms by which bark beetleattacks often fail in healthy trees (e.g. Nebeker et al.1993). Our study suggests that under circumstances inwhich the host is not stressed abiotically (e.g. bydrought), pathogen infection, e.g. by root pathogens,may increase resin flow systemically and consequentlyenhance host resistance to these insect guilds, at leasttemporarily. Only once the host’s physiology is impairedby pathogenic invasion to the point that the resin re-sponse is no longer expressed at levels antagonistic to theinsects will the insect attacks be successful. While themechanisms involved are different, similar outcomes ofcross-induction of systemic resistance to insects bypathogens are well known in many herbaceous plantsystems (Felton et al. 1989; Blua and Perring 1992; Stoutand Duffey 1996; Inbar et al. 1998), but we know of onlyone study with coniferous trees to date (McNee et al.2003). Further investigations in this area will contributesignificantly to our understanding of the effects of sys-temic cross-effects between pathogens and herbivores onoverall conifer host fitness and ecosystem sustainability.
Acknowledgements The authors thank M. Bellizzi for technicalassistance, and John Freudenstein (Ohio State University) andTom Gordon (University of California, Davis) for pre-submissionreviews. Salaries and research support were provided by state andfederal funds appropriated to the Ohio Agricultural Research andDevelopment Center and The Ohio State University.
References
Bannan MW (1936) Vertical resin ducts in the secondary wood ofthe Abietineae. New Phytol 35:11–46
Berryman AA (1972) Resistance in conifers to invasion by barkbeetle–fungus associations. Bioscience 22:598–602
Blanchette RA, Biggs AR (1992) Defense mechanisms of woodyplants against fungi. Springer, Berlin Heidelberg New York,pp 458
Blodgett JT, Bonello P (2003) The aggressiveness of Sphaeropsissapinea on Austrian pine varies with isolate group and site ofinfection. For Pathol 33:15–19
Blodgett JT, Stanosz GR (1997) Sphaeropsis sapinea morphotypesdiffer in aggressiveness, but both infect nonwounded red or jackpines. Plant Dis 81:143–147
Blodgett JT, Stanosz GR (1999) Differences in aggressiveness ofSphaeropsis sapinea RAPD marker group isolates on severalconifers. Plant Dis 83:853–856
Blodgett JT, Bellizzi M, Bonello P (2003a) The induction of systemicresistance and systemic susceptibility in Sphaeropsis sapineainoculated Pinus nigra. In: Stone J, Maffei H (eds) 50th WesternInt Forest Disease Working Conference, USDA Forest Service,Powell River, British Columbia, Canada, pp 46–47
83

Blodgett JT, Bonello P, Stanosz GR (2003b) An effective mediumfor isolating Sphaeropsis sapinea from asymptomatic pines. ForPathol 33:395–404
Blua MJ, Perring TM (1992) Effects of zucchini yellow mosaic-virus on colonization and feeding-behavior of Aphis gossypii(Homoptera, Aphididae) alatae. Environ Entomol 21:578–585
Bonello P, Blodgett JT (2003) Pinus nigra–Sphaeropsis sapinea asa model pathosystem to investigate local and systemic effectsof fungal infection of pines. Physiol Mol Plant Pathol63:249–261
Bonello P, Pearce RB, Watt F, Grime GW (1991) An inducedpapilla response in primary roots of Scots pine challenged invitro with Cylindrocarpon destructans. Physiol Mol Plant Pathol39:213–228
Bonello P, Gordon TR, Storer AJ (2001) Systemic induced resis-tance in Monterey pine. For Pathol 31:99–106
Croteau RB, Johnson MA (1985) Biosynthesis of terpenoidwood extractives. In: Higuchi T (ed) Biosynthesis and bio-degradation of wood components. Academic Press, NewYork, pp 379–439
Davis JM, Wu H, Cooke JEK, Reed JM, Luce KS, Michler CH(2002) Pathogen challenge, salicylic acid, and jasmonic acidregulate expression of chitinase gene homologs in pine. MolPlant Microbe Interact 15:380–387
de Wet J, Burgess T, Slippers B, Preisig O, Wingfield BD, WingfieldMJ (2003) Multiple gene genealogies and microsatellite markersreflect relationships between morphotypes of Sphaeropsis sapi-nea and distinguish a new species of Diplodia. Mycol Res107:557–566
Faldt J, Martin D, Miller B, Rawat S, Bohlmann J (2003) Trau-matic resin defense in Norway spruce (Picea abies): methyljasmonate-induced terpene synthase gene expression, andcDNA cloning and functional characterization of (+)-3-carenesynthase. Plant Mol Biol 51:119–133
Feci E, Battisti A, Capretti P, Tegli S (2002) An association be-tween the fungus Sphaeropsis sapinea and the cone bug Gas-trodes grossipes in cones of Pinus nigra in Italy. For Pathol32:241–247
Felton GW, Broadway RM, Duffey SS (1989) Inactivation ofprotease inhibitor activity by plant-derived quinones—compli-cations for host-plant resistance against Noctuid herbivores.J Ins Phys 35:981–990
Finstein MS, Miller FC, Strom PF, Macgregor ST, Psarianos KM(1983) Composting ecosystem management for waste treat-ment. Bio Technol 1:347–353
Franceschi VR, Krokene P, Krekling T, Christiansen E (2000)Phloem parenchyma cells are involved in local and distant de-fense responses to fungal inoculation or bark-beetle attack inNorway spruce (Pinaceae). Am J Bot 87:314–326
Franceschi VR, Krekling T, Christiansen E (2002) Application ofmethyl jasmonate on Picea abies (Pinaceae) stems induces de-fense-related responses in phloem and xylem. Am J Bot 89:578–586
Hammerschmidt R, Nicholson RL (1999) A survey of plant defenseresponses to pathogens. In: Agrawal AA, Tuzun S, Bent E (eds)Induced plant defenses against pathogens and herbivores. APSPress, St. Paul, MN, pp 55–71
Hudgins JW, Christiansen E, Franceschi VR (2003) Methyl jasm-onate induces changes mimicking anatomical defenses in di-verse members of the Pinaceae. Tree Physiol 23:361–371
Hudgins JW, Christiansen E, Franceschi VR (2004) Induction ofanatomically based defense responses in stems of diverse coni-fers by methyl jasmonate: a phylogenetic perspective. TreePhysiol 24:251–264
Inbar M, Doostdar H, Sonoda RM, Leibee GL, Mayer RT (1998)Elicitors of plant defensive systems reduce insect densities anddisease incidence. J Chem Ecol 24:135–149
Kinloch BB, Sniezko RA, Barnes GD, Greathouse TE (1999) Amajor gene for resistance to white pine blister rust in westernwhite pine from the Western Cascade Range. Phytopathology89:861–867
Klepzig KD, Kruger EL, Smalley EB, Raffa KF (1995) Effects ofbiotic and abiotic stress on induced accumulation of terpenesand phenolics in red pines inoculated with bark beetle-vectoredfungus. J Chem Ecol 21:601–626
Krekling T, Franceschi VR, Krokene P, Solheim H (2004) Differ-ential anatomical response of Norway spruce stem tissues tosterile and fungus infected inoculations. Trees Struct Func18:1–9
Maresi G, Ambrosi P, Battisti A, Capretti P, Danti R, Feci E,Minerbi S, Tegli S (2002) Pine dieback by Sphaeropsis sapinea inNorthern and Central Italy. In: Shoot and foliage diseases.Forest research institute research papers. Proceedings of theIUFRO Working Party 7.02.02. Hyytiala, Finland, pp 60–67
Martin D, Tholl D, Gershenzon J, Bohlmann J (2002) Methyljasmonate induces traumatic resin ducts, terpenoid resin bio-synthesis, and terpenoid accumulation in developing xylem ofNorway spruce stems. Plant Physiol 129:1003–1018
Martin DM, Gershenzon J, Bohlmann J (2003) Induction of vol-atile terpene biosynthesis and diurnal emission by methyljasmonate in foliage of Norway spruce. Plant Physiol 132:1586–1599
McKay SAB, Hunter WL, Godard KA, Wang SX, Martin DM,Bohlmann J, Plant AL (2003) Insect attack and wounding in-duce traumatic resin duct development and gene expression of(�)-pinene synthase in Sitka spruce. Plant Physiol 133:368–378
McNee WR, Bonello P, Wood DL, Storer AJ, Gordon TR (2003)Feeding response of Ips paraconfusus to phloem and phloemmetabolites of Heterobasidion annosum-inoculated ponderosapine, Pinus ponderosa. J Chem Ecol 29:1183–1202
Metraux JP (2001) Systemic acquired resistance and salicylic acid:current state of knowledge. Eur J Plant Pathol 107:13–18
Nagy NE, Franceschi VR, Solheim H, Krekling T, Christiansen E(2000) Wound-induced traumatic resin duct development instem of Norway spruce (Pinaceae): anatomy and cytochemicaltraits. Am J Bot 87:302–313
Nebeker TE, Hodges JD, Blanche CA (1993) Host response to barkbeetle and pathogen colonization. In: Schowalter TD, GM Filip(eds) Beetle-pathogen interactions in conifer forests. AcademicPress, London, pp 157–173
Phillips MA, Croteau RB (1999) Resin-based defenses in conifers.Trends Plant Sci 4:184–190
Shrimpton DM, Whitney HS (1968) Inhibition of growth of bluestain fungi by wood extractive. Can J Bot 46:757–761
Solheim H (1991) Oxygen deficiency and spruce resin inhibition ofgrowth of fungi associated with Ips typographus. Mycol Res95:1387–1392
Stanosz GR, Smith DR, Guntmiller MA (1996) Characterizationof Sphaeropsis sapinea from the west central United States bymeans of random amplified polymorphic DNA marker analy-sis. Plant Dis 80:1175–1178
Stout MJ, SS Duffey (1996) Characterization of induced resistancein tomato plants. Entomol Exp Appl 79:273–283
Swart WJ, Wingfield MJ (1991) Biology and control of Sphaeropsissapinea on Pinus species in South Africa. Plant Dis 75:761–766
Walton JD (1997) Biochemical plant pathology. In: Dey PM,Harborne JB (eds) Plant biochemistry. Academic Press, SanDiego, pp 487–502
Wong BL, Berryman AA (1977) Host resistance to the fir engraverbeetle. 3. Lesion development and containment of infection byresistant Abies grandis inoculated with Trichosporium symbiot-icum. Can J Bot 55:2358–2365
Wu H, Hu Z (1977) Comparative anatomy of resin ducts of thePinaceae. Trees 11:135–143
84




















