
Sulcal Root” Generic Model: a Hypothesis to Overcome the Variability of the Human Cortex Folding...
17
1 Received June 16, 2004; Accepted July 12, 2004 R áegis and co-workers investigated the cerebral cortical folding pattern and proposed a generic sulcal root model based on MR imaging data of 20 normal subjects (4 females and 16 males). They tried to resolve the cortical folding pattern of the individuals using a computer three-dimensional visualization tool of the MR images. They tried to define `pli de passage' in French, which means a small gyri buried inside the deep main sulci. They hypothesized that the `pli de passage' subdivide in subunits (sulcal roots) large sulcus. Their observations are still under investigation, however, their approach using precise computer graphics is brand new. This method may be useful to understand brain function and to open new doors for brain mapping. Further improvement will be required not only for the resolution of the three dimensional MR imaging but also for the software of the image processing for the universal use of this technique. Tatsuya TANAKA, M.D. Department of Neurosurgery, Asahikawa Medical College, Asahikawa, Hokkaido The President of the 40th Annual Meeting of Japan Clinical Neurophysiology Society 1 Neurol Med Chir (Tokyo) 45, 1¿17, 2005 ``Sulcal Root'' Generic Model: a Hypothesis to Overcome the Variability of the Human Cortex Folding Patterns Jean R Á EGIS, Jean-François MANGIN*, Taku OCHIAI**, Vincent FROUIN*, Denis RIVI ÀERE*, Arnaud CACHIA*, Manabu TAMURA, and Yves SAMSON* Stereotactic and Functional Neurosurgery Department, Timone Hospital, A.P.M., Marseille, France & INSERM 99-26; *Service Hospitalier Fráedáeric Joliot, CEA, Orsay, France; ** Department of Neurosurgery, Neurological Institute, Tokyo Women's Medical University, Tokyo Abstract The great variability of cerebral cortical folding patterns raises major problems for the systematic study of functional-structural relationships. This paper describes a novel perspective for explaining this variability, a perspective that relies on gyri buried in the depth of the sulci. From this perspective we propose a generic model of folding, based on indivisible units, called sulcal roots, which correspond to the first folding locations during antenatal life. These units are organized according to a basic scheme allowing us to describe the cortical surface using a system of meridians and parallels. This scheme is thought to be stable across individuals at the fetal stage, and may be related to the protomap model. Variability at the adult stage is thought to result from the chaotic behavior of the folding process: inter-individual differences in cortical areas can lead to qualitatively different folding patterns. We have tested the capacity of this model to match actual cortical anatomy with a database of magnetic resonance images of 20 normal subjects, using new three-dimensional visualization tools giving access to shapes buried in the cortex. Key words: cerebral cortex, variability, brain mapping, sulcus, morphogenesis, tensegrity Introduction During recent decades, a large number of investiga- tional modalities yielding information on the func- tion of the brain have appeared. The usual strategy for interpreting such data consists of matching individual data with current knowledge of the normal functional organization of the brain. Using brain anatomy as a reference is the natural way of performing this matching. 3,7,18,22,23,38,39,43) While this strategy has been very successful for animal and human studies when in vitro microstructural data are available, dealing with in vivo human studies is much more problematic because magnetic resonance (MR) imaging provides only macroscopic anatomical information. With regard to the human cerebral cortex, the sulco-gyral patterns are the only available landmarks. Unfortunately, the spatial relationships between these landmarks and the underlying functional and architectonic maps are questionable. 53,60) Moreover, the large inter-subject variability of these cortical patterns raises major
Transcript of Sulcal Root” Generic Model: a Hypothesis to Overcome the Variability of the Human Cortex Folding...

1
Received June 16, 2004; Accepted July 12, 2004R áegis and co-workers investigated the cerebral cortical folding pattern and proposed a generic sulcal root model based onMR imaging data of 20 normal subjects (4 females and 16 males). They tried to resolve the cortical folding pattern of theindividuals using a computer three-dimensional visualization tool of the MR images. They tried to define `pli de passage'in French, which means a small gyri buried inside the deep main sulci. They hypothesized that the `pli de passage'subdivide in subunits (sulcal roots) large sulcus. Their observations are still under investigation, however, their approachusing precise computer graphics is brand new. This method may be useful to understand brain function and to open newdoors for brain mapping. Further improvement will be required not only for the resolution of the three dimensional MRimaging but also for the software of the image processing for the universal use of this technique.
Tatsuya TANAKA, M.D.Department of Neurosurgery, Asahikawa Medical College, Asahikawa, Hokkaido
The President of the 40th Annual Meeting of Japan Clinical Neurophysiology Society
1
Neurol Med Chir (Tokyo) 45, 1¿17, 2005
``Sulcal Root'' Generic Model: a Hypothesis to Overcomethe Variability of the Human Cortex Folding Patterns
Jean R ÁEGIS, Jean-François MANGIN*, Taku OCHIAI**, Vincent FROUIN*,Denis RIVI ÀERE*, Arnaud CACHIA*, Manabu TAMURA,
and Yves SAMSON*
Stereotactic and Functional Neurosurgery Department, Timone Hospital, A.P.M.,Marseille, France & INSERM 99-26; *Service Hospitalier Fr áed áeric Joliot, CEA, Orsay, France;
**Department of Neurosurgery, Neurological Institute, Tokyo Women's Medical University, Tokyo
Abstract
The great variability of cerebral cortical folding patterns raises major problems for the systematic studyof functional-structural relationships. This paper describes a novel perspective for explaining thisvariability, a perspective that relies on gyri buried in the depth of the sulci. From this perspective wepropose a generic model of folding, based on indivisible units, called sulcal roots, which correspond tothe first folding locations during antenatal life. These units are organized according to a basic schemeallowing us to describe the cortical surface using a system of meridians and parallels. This scheme isthought to be stable across individuals at the fetal stage, and may be related to the protomap model.Variability at the adult stage is thought to result from the chaotic behavior of the folding process:inter-individual differences in cortical areas can lead to qualitatively different folding patterns. Wehave tested the capacity of this model to match actual cortical anatomy with a database of magneticresonance images of 20 normal subjects, using new three-dimensional visualization tools giving accessto shapes buried in the cortex.
Key words: cerebral cortex, variability, brain mapping, sulcus, morphogenesis, tensegrity
Introduction
During recent decades, a large number of investiga-tional modalities yielding information on the func-tion of the brain have appeared. The usual strategyfor interpreting such data consists of matchingindividual data with current knowledge of thenormal functional organization of the brain. Usingbrain anatomy as a reference is the natural way ofperforming this matching.3,7,18,22,23,38,39,43) While thisstrategy has been very successful for animal and
human studies when in vitro microstructural dataare available, dealing with in vivo human studiesis much more problematic because magneticresonance (MR) imaging provides only macroscopicanatomical information. With regard to the humancerebral cortex, the sulco-gyral patterns are the onlyavailable landmarks. Unfortunately, the spatialrelationships between these landmarks and theunderlying functional and architectonic maps arequestionable.53,60) Moreover, the large inter-subjectvariability of these cortical patterns raises major
22
Neurol Med Chir (Tokyo) 45, January, 2005
J. R áegis et al.
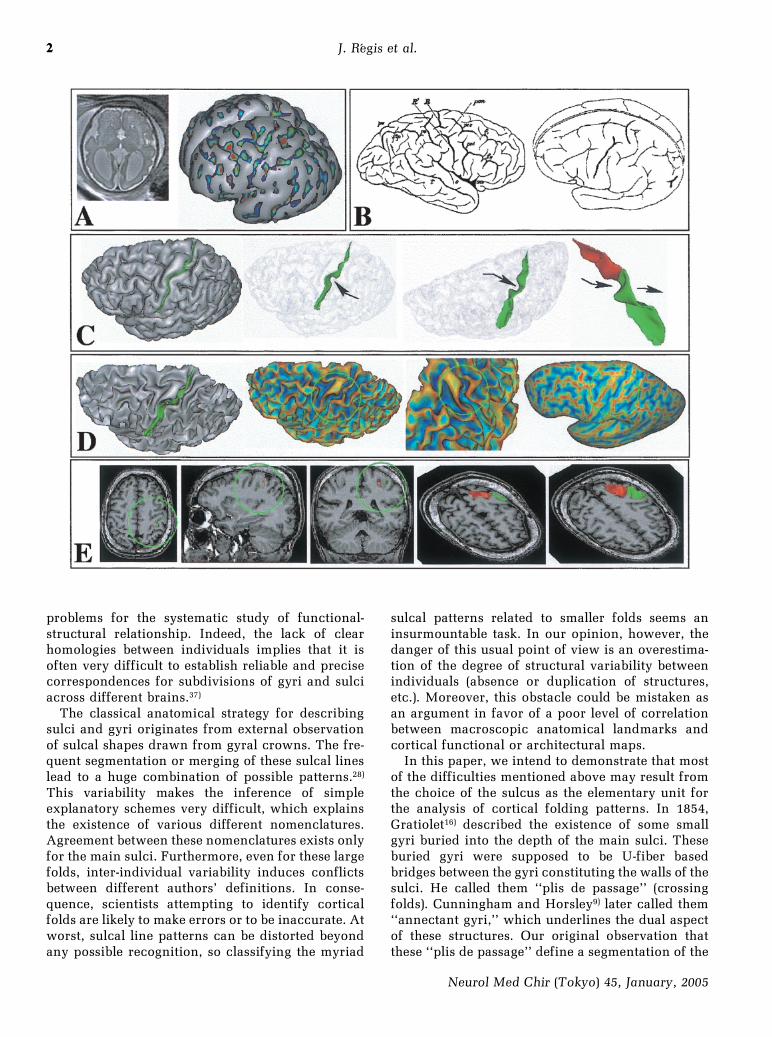
problems for the systematic study of functional-structural relationship. Indeed, the lack of clearhomologies between individuals implies that it isoften very difficult to establish reliable and precisecorrespondences for subdivisions of gyri and sulciacross different brains.37)
The classical anatomical strategy for describingsulci and gyri originates from external observationof sulcal shapes drawn from gyral crowns. The fre-quent segmentation or merging of these sulcal lineslead to a huge combination of possible patterns.28)
This variability makes the inference of simpleexplanatory schemes very difficult, which explainsthe existence of various different nomenclatures.Agreement between these nomenclatures exists onlyfor the main sulci. Furthermore, even for these largefolds, inter-individual variability induces conflictsbetween different authors' definitions. In conse-quence, scientists attempting to identify corticalfolds are likely to make errors or to be inaccurate. Atworst, sulcal line patterns can be distorted beyondany possible recognition, so classifying the myriad
sulcal patterns related to smaller folds seems aninsurmountable task. In our opinion, however, thedanger of this usual point of view is an overestima-tion of the degree of structural variability betweenindividuals (absence or duplication of structures,etc.). Moreover, this obstacle could be mistaken asan argument in favor of a poor level of correlationbetween macroscopic anatomical landmarks andcortical functional or architectural maps.
In this paper, we intend to demonstrate that mostof the difficulties mentioned above may result fromthe choice of the sulcus as the elementary unit forthe analysis of cortical folding patterns. In 1854,Gratiolet16) described the existence of some smallgyri buried into the depth of the main sulci. Theseburied gyri were supposed to be U-fiber basedbridges between the gyri constituting the walls of thesulci. He called them ``plis de passage'' (crossingfolds). Cunningham and Horsley9) later called them``annectant gyri,'' which underlines the dual aspectof these structures. Our original observation thatthese ``plis de passage'' define a segmentation of the

3
"
Fig. 1 Tenets underlying the generic sulcal root model as illustrated by the example of the centralsulcus. The classical description reports that the majority of the subjects have one centralsulcus for each hemisphere (C) and that in 1% of the cases there are two central sulci in thesame hemisphere and very rarely in the two hemispheres. The figure shows such a casereported by Sernoff (B).9) This description introduces a structural variation of the centralsulcus model: one or two parts. In fact we know from ontology that in all the describedspecimens two primitive sulci (sulcal roots) appear at the age of 4 or 5 months, which can bevisualized in antenatal imaging from the curvature of the brain surface (A).5) The sulcal rootsare separated by a small transverse annectent gyrus. With the explosive development of thesurrounding gyri (pre- and postcentral gyri), these three primal elements are usually buriedinto the depth of the mature central sulcus. Rarely, however, the annectent gyrus is developedand superficial enough to be visible from outside, which leads to an interrupted centralsulcus aspect. In fact, a three-dimensional extraction and visualization of the central sulcuscomplex with Anatomist software demonstrates the systematic presence of a deep annectentgyrus splitting the mature central sulcus complex into two constant units (sulcal roots) (C, E).This buried gyrus can also be visualized from the curvature of the white matter surface (D).Usually, the inferior central sulcal root is separated from the sylvian fissure by a gyrus calledthe subcentral gyrus.28) In some cases, however, the inferior central sulcal root appears asconnected to this sylvian fissure (23%). In fact, the subcentral gyrus is still there but buried inthe depth of the folding. An identical comment can be made for the relation between thesuperior central sulcal root and the callosomarginal fissure. The sulcal root-based descrip-tion of the central sulcus, made up of two indivisible units, can account for the differentexternal aspects. Hence the same structural model is used to describe any pattern which leadsto straightforward structural homologies across subjects. Finally the difference between anannectent gyrus (as the intermediate parieto-frontal plis de passage) and a standard gyrus (asthe precentral gyrus) is only related to the fact that the former gyrus is usually buried. Precen-tral gyrus, however, may appear as an annectent gyrus when the superior precentral incisura,namely the crossing between superior precentral and frontal sulci, is superficially posteriorlyconnected to the central sulcus.
3
Neurol Med Chir (Tokyo) 45, January, 2005
Sulcal Root Generic Model
sulci into subunits led us to investigate the role ofthis phenomenon in the inter-individual variabilityof external sulcal patterns. We observed a clearcorrelation between sulci; interruptions describedin the literature and the locations of the plis depassage. We also observed a good correlationbetween the early fetal aspects of the sulci describedin the embryological literature and the interruptionsmentioned above. This set of observations led us tocall the sulcal subunits ``sulcal roots.''33)
We will clarify our approach using the simple caseof the central sulcus (CS) (see Fig. 1). First, a pli depassage called ``intermediate parieto-frontal'' hasbeen described in the middle of the CS.1,11,16,42,56,57)
Second, in 1% of brains, the CS is described as madeup of two distinct folds.12,15,34,51) Third, in the 5–6months old fetus,48) the CS appears as two different`primal sketches.'4) This consistent set of observa-tions leads to the definition of two sulcal roots thatwe call the inferior and superior central sulcal roots.The potential CS interruption is therefore interpret-ed as the nonoperculation (burying) of the pli depassage (annectent gyrus) which is visible during theearly stages of development. The degree of opercula-
tion depends on the expansion of the pli de passage'scortex relative to the expansion of the surroundingorthogonal gyri (pre- and postcentral gyri in thiscase). To conclude, depending on the depth of theusually buried pli de passage, the CS can appearpartially or completely interrupted, or continuous.
In order to further explore the potential of thesulcal root concept for overcoming the difficultiesinduced by variability of external observations ofcortical sulci, in 1992 we initiated a research pro-gram aimed at mapping the main sulcal roots andthe dual ``plis de passage'' of the human cortex.While the first sulcal root maps were mainlyinferred from observations in the literature,32) one ofour objectives was to rely as far as possible on MRimaging anatomical data. Therefore, since the usualsurface renderings of the cortex computed from MRimages suffer from the same weaknesses as externalobservations, we designed algorithmic methodsfor the analysis and manipulation of the buriedcortex.21) These new visualization tools have rev-ealed much information on sulcal shapes, whichhave allowed refinement of the sulcal root map.Difficulties for brain warping strategies induced by
44
Neurol Med Chir (Tokyo) 45, January, 2005
J. R áegis et al.
the variability of folding patterns have now raisedincreased interest in studies of the buried shapes ofcortical folds.10,19,20,35,40,49,58,59)
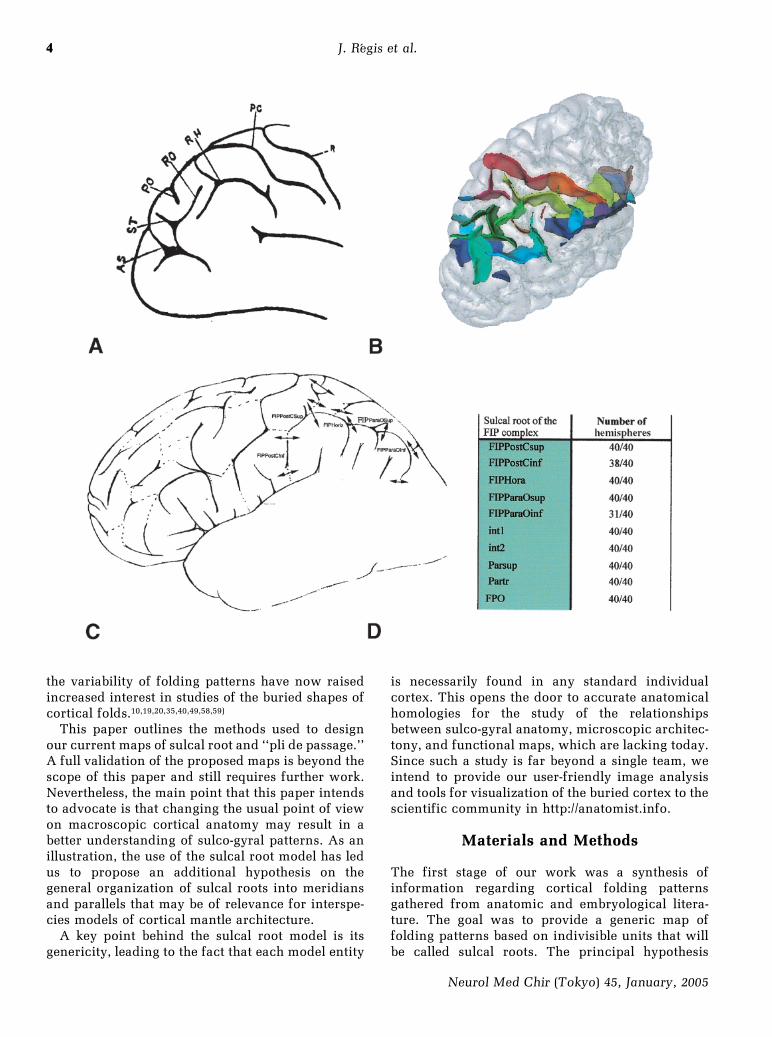
This paper outlines the methods used to designour current maps of sulcal root and ``pli de passage.''A full validation of the proposed maps is beyond thescope of this paper and still requires further work.Nevertheless, the main point that this paper intendsto advocate is that changing the usual point of viewon macroscopic cortical anatomy may result in abetter understanding of sulco-gyral patterns. As anillustration, the use of the sulcal root model has ledus to propose an additional hypothesis on thegeneral organization of sulcal roots into meridiansand parallels that may be of relevance for interspe-cies models of cortical mantle architecture.
A key point behind the sulcal root model is itsgenericity, leading to the fact that each model entity
is necessarily found in any standard individualcortex. This opens the door to accurate anatomicalhomologies for the study of the relationshipsbetween sulco-gyral anatomy, microscopic architec-tony, and functional maps, which are lacking today.Since such a study is far beyond a single team, weintend to provide our user-friendly image analysisand tools for visualization of the buried cortex to thescientific community in http://anatomist.info.
Materials and Methods
The first stage of our work was a synthesis ofinformation regarding cortical folding patternsgathered from anatomic and embryological litera-ture. The goal was to provide a generic map offolding patterns based on indivisible units that willbe called sulcal roots. The principal hypothesis

5
"
Fig. 2 Construction of the model. Insight into the construction of the generic model, and example ofthe ``fissura intraparietalis (FIP).'' The FIP consists of five sulcal roots in our model (seeFig. 5). The main contributions of the anatomical literature on this area, which have beenused as the basic material for our model for this region, are reported here. Eberstaller12) hasgiven a detailed description of the deep gyri (pli de passage: PP) which bridge across the bot-tom of the FIP. The description has been confirmed by Cunningham and Horsley.9) Cunnin-gham also reported his observation of the development of the FIP from the analysis of 37 fetusbrains. The two ``postcentralis inferior'' precursors appear around 6.5 months (postovulato-ry), the two ``paraoccipitalis'' (superior and inferior) appear at 7 months, and the ``horizonta-lis'' appears at 7.5 months according to Cunningham and Horsley.9) According to theseauthors ``at first the ramus horizontalis and the ramus occipitalis stand well apart but as theyextend they gradually approach each other, and finally establish a superficial connection.''In 79 adult hemispheres, the same authors find that the four ``PPs'' separating these fivesulcal roots are superficial, namely give an interrupted pattern, in 50%, 30%, 37%, and 5% ofthe subjects (from rostral to caudal). These findings fit well with the observations of Ono etal.,28) who have described extensively the frequency of the interruption phenomenon accord-ing to the location. Based on these reports from the literature (A: Cunningham's drawing ofthe FIP9)) and in vivo study of the cortex of normal subjects (B: three-dimensional display ofthe FIP with Anatomist software), a generic description has been proposed (C: scheme of thesulcal roots and PP of the FIP for names legend see Fig. 5). The study of 40 ``testhemispheres'' (D) aiming at the identification of the FIP's sulcal roots and dual PPs led to aputative solution for all structures in all cases except for the inferior and superior paraoc-cipitalis roots, which were not clearly separated in nine of the 40 subjects. Interestingly theunderlying PP corresponds to the less frequent superficial interruption according to Cunnin-gham and Horsley.
5
Neurol Med Chir (Tokyo) 45, January, 2005
Sulcal Root Generic Model
underlying this work is the existence of theseindivisible units in any nonpathological maturecortex. Three main kinds of information were usedto infer this sulcal root map: 1) Interruptions ofthe main sulci described in the anatomical literatureduring observation of the outer aspect of thecortex.9,16,28) 2) Observations of some ``plis de pas-sage'' usually hidden in the depth of the main sulcimentioned by various authors.9,11,16,28) Our nomen-clature is based on Dejerine's.11) 3) The embryologi-cal literature has been browsed in order to poolinformation on date of onset of the development ofsulcal roots and annectent gyri, when available.6,13,48)
The main challenge during this work wasmatching the different elements found in the litera-ture. Although the current map required numerousiterations before achieving convergence, the logic ofour approach may be understood as follows:
1) Split each main sulcus into portions accordingto the different standard interruptions observed inthe literature. Use the relative shapes of the resultingportions and the relative locations of the neighbor-ing folds to match interruptions across authors. Themain source of information was the Ono and Kubikatlas, which provides a more recent and completework on the variability of the superficial aspect ofthe sulci in human cerebral cortex.28) They describea population of 25 postmortem brains, using a classi-
cal strategy for depicting individual patterns relyingon analogous references (Y shape, etc.) and structur-al variations (interruption, continuity, connection,pseudo-interruption, pseudo-continuity, supplemen-tary sulci, etc.). Their contribution has been used asa reference for the collection of these patterns andtheir frequencies.
2) Match the standard interruptions with the ``plisde passage'' using the relationship with surroundingcortical structures and the described frequencies ofsuperficiality the `nonburied situation.' Each inter-ruption is thought to be induced by a superficial ``plide passage.'' A new split of the sulcus is added foreach ``pli de passage'' without matched standardinterruption. These ``plis de passage'' are thought tobe buried most of the time.
3) Match the resulting units, namely the portionsof sulci separated by two splits, with the primal folds(the sulcal roots) described in embryological litera-ture. This was a more difficult task because someparts of the cortex have not been studied exten-sively. The chronology of the appearance of theroots, when available, is the main informationdriving the matching. Inferring the dynamic of thefolding process has led us to assume that the meandepth of the ``plis de passage'' in mature cortex iscorrelated with the date of apparition. Some newsubdivisions were added for sulci including fewer
6
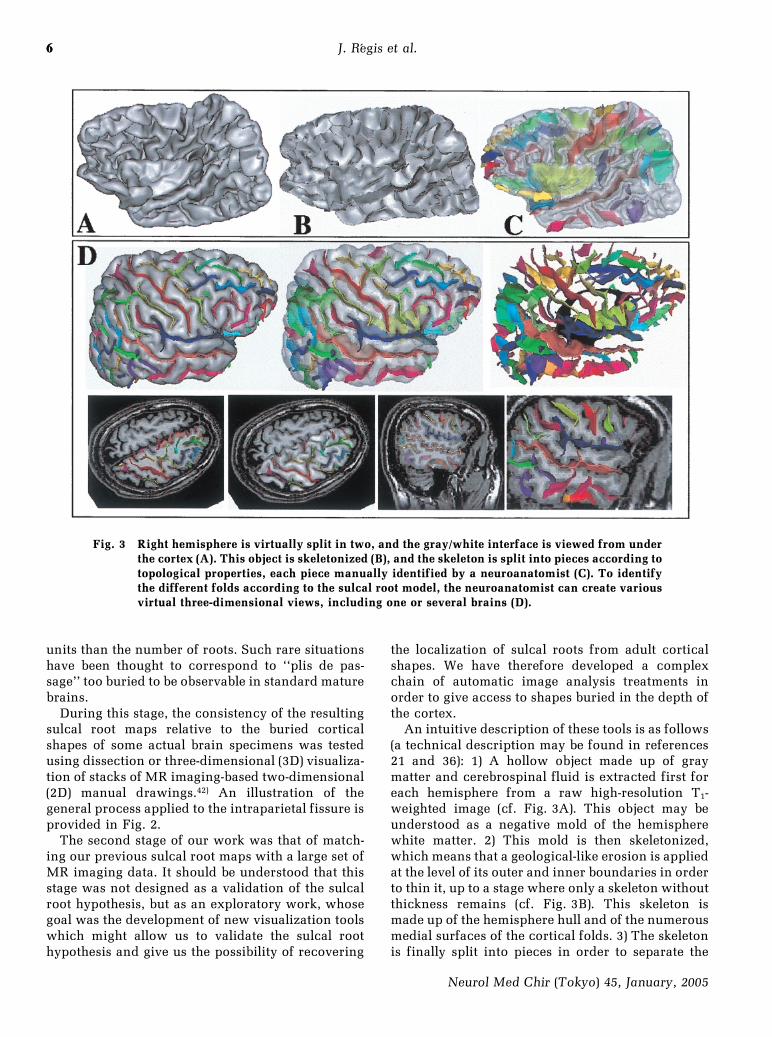
Fig. 3 Right hemisphere is virtually split in two, and the gray/white interface is viewed from underthe cortex (A). This object is skeletonized (B), and the skeleton is split into pieces according totopological properties, each piece manually identified by a neuroanatomist (C). To identifythe different folds according to the sulcal root model, the neuroanatomist can create variousvirtual three-dimensional views, including one or several brains (D).
6
Neurol Med Chir (Tokyo) 45, January, 2005
J. R áegis et al.
units than the number of roots. Such rare situationshave been thought to correspond to ``plis de pas-sage'' too buried to be observable in standard maturebrains.
During this stage, the consistency of the resultingsulcal root maps relative to the buried corticalshapes of some actual brain specimens was testedusing dissection or three-dimensional (3D) visualiza-tion of stacks of MR imaging-based two-dimensional(2D) manual drawings.42) An illustration of thegeneral process applied to the intraparietal fissure isprovided in Fig. 2.
The second stage of our work was that of match-ing our previous sulcal root maps with a large set ofMR imaging data. It should be understood that thisstage was not designed as a validation of the sulcalroot hypothesis, but as an exploratory work, whosegoal was the development of new visualization toolswhich might allow us to validate the sulcal roothypothesis and give us the possibility of recovering
the localization of sulcal roots from adult corticalshapes. We have therefore developed a complexchain of automatic image analysis treatments inorder to give access to shapes buried in the depth ofthe cortex.
An intuitive description of these tools is as follows(a technical description may be found in references21 and 36): 1) A hollow object made up of graymatter and cerebrospinal fluid is extracted first foreach hemisphere from a raw high-resolution T1-weighted image (cf. Fig. 3A). This object may beunderstood as a negative mold of the hemispherewhite matter. 2) This mold is then skeletonized,which means that a geological-like erosion is appliedat the level of its outer and inner boundaries in orderto thin it, up to a stage where only a skeleton withoutthickness remains (cf. Fig. 3B). This skeleton ismade up of the hemisphere hull and of the numerousmedial surfaces of the cortical folds. 3) The skeletonis finally split into pieces in order to separate the
7
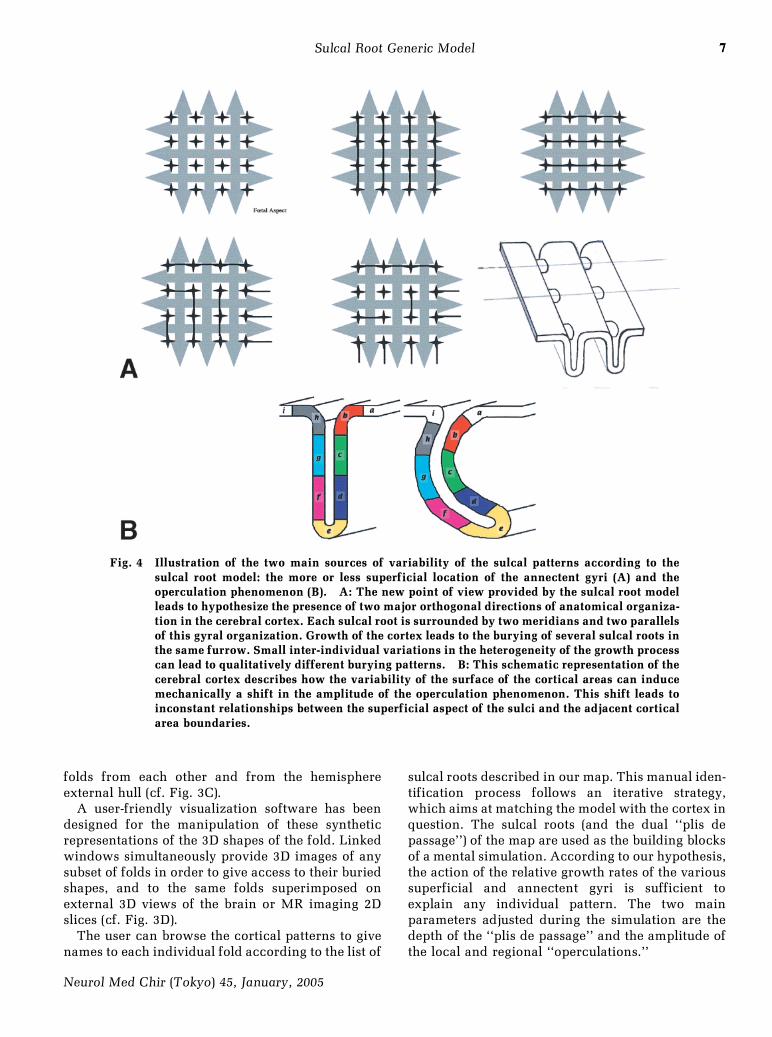
Fig. 4 Illustration of the two main sources of variability of the sulcal patterns according to thesulcal root model: the more or less superficial location of the annectent gyri (A) and theoperculation phenomenon (B). A: The new point of view provided by the sulcal root modelleads to hypothesize the presence of two major orthogonal directions of anatomical organiza-tion in the cerebral cortex. Each sulcal root is surrounded by two meridians and two parallelsof this gyral organization. Growth of the cortex leads to the burying of several sulcal roots inthe same furrow. Small inter-individual variations in the heterogeneity of the growth processcan lead to qualitatively different burying patterns. B: This schematic representation of thecerebral cortex describes how the variability of the surface of the cortical areas can inducemechanically a shift in the amplitude of the operculation phenomenon. This shift leads toinconstant relationships between the superficial aspect of the sulci and the adjacent corticalarea boundaries.
7
Neurol Med Chir (Tokyo) 45, January, 2005
Sulcal Root Generic Model
folds from each other and from the hemisphereexternal hull (cf. Fig. 3C).
A user-friendly visualization software has beendesigned for the manipulation of these syntheticrepresentations of the 3D shapes of the fold. Linkedwindows simultaneously provide 3D images of anysubset of folds in order to give access to their buriedshapes, and to the same folds superimposed onexternal 3D views of the brain or MR imaging 2Dslices (cf. Fig. 3D).
The user can browse the cortical patterns to givenames to each individual fold according to the list of
sulcal roots described in our map. This manual iden-tification process follows an iterative strategy,which aims at matching the model with the cortex inquestion. The sulcal roots (and the dual ``plis depassage'') of the map are used as the building blocksof a mental simulation. According to our hypothesis,the action of the relative growth rates of the varioussuperficial and annectent gyri is sufficient toexplain any individual pattern. The two mainparameters adjusted during the simulation are thedepth of the ``plis de passage'' and the amplitude ofthe local and regional ``operculations.''

88
Neurol Med Chir (Tokyo) 45, January, 2005
J. R áegis et al.
Operculation is a growth phenomenon that isclosely related to the ``pli de passage'' notion.Operculation occurs when actively developingcortical regions grow outward faster than doadjacent zones, which lag behind in the depth (cf.Fig. 4). The most striking example of operculation isthe formation of the sylvian fissure induced by themore rapid growth of the temporal and frontal lobes.Operculation is also implied in the creation of someof the sulci, but is far from being the only processbehind gyration.50) The variability of the numeroussulcal branches and incisurae, nevertheless, may berelated to variability of local operculation ampli-tude.55) Some branches, for instance, may becomeburied into sulci or may not.
The matching process relies particularly on ournew visualization tool. Browsing virtual images ofthe deepest part of the sulci gives access to manyclues to the organization of sulcal roots and ``plis depassages.'' For each fold portion, topographicalrelationship to other folds, morphology, dimensionsof the segments, incisura, and annectent gyri werecarefully checked and compared to the rest of thebrain database in order to obtain consistent labeling.In fact, the software allows the user to simultaneous-ly browse any number of hemispheres.
MR imaging was performed in a group of 20normal subjects. These subjects had no previousneurological history. Clinical neurological examina-tion was normal in all. The Edinburgh test26) wasperformed to determine the laterality quotient (LQ).There were four females, who were right handed(LQ= 100), and 16 males, of whom four were lefthanded (mean LQ= 87.5) and 12 right handed(mean LQ= 93.3). The mean age was 30.4 years.Our sulcal root model was matched with the individ-ual cortices following the manual labeling strategymentioned above. Our attention was focused on thefollowing main sulcal complexes: the central sulcalcomplex, the frontal superior sulcal complex, theprecentral sulcal complex, the frontal inferior sulcalcomplex, the callosomarginal sulcal complex, theinterparietal sulcal complex, the temporal superiorsulcal complex, the parieto-occipital sulcal complex,and the calcarine sulcal complex. For each of thesesulcal complexes, we identified the sulcal rootsubunits and the ``plis de passage.''
Results
The topography of the main sulcal roots of ourgeneric model are displayed in the Talairach gridsystem in Fig. 5. The 3D visualization of the sulciwith Anatomist software has dramatically improvedour capacity to quickly and simply recognize these
subunits (sulcal roots). This improvement is mainlydue to the availability of the abundant visual infor-mation on the 3D shape of the sulci, the enhance-ment of our capability to locate the plis de passageeven when deeply buried, direct visualization of thepoints of depth variation and the points of change inthe orientation of sulcal planes, and the impressionof annectent gyri on the inner aspect of the sulcalwalls.
We will detail our results on the parietal cortex. InFig. 2 we can see that all subunits are not identifiedin 100% of the brains. The inferior root of thepostcentral segment of the interparietal fissurecould be distinguished in 38 of 40 hemispheres. Theinferior root of the paraoccipital segment of theinterparietal fissure could be distinguished in 31 of40 hemispheres. In two hemispheres for the formerand nine for the latter, the identification of the plisde passage was not clear enough to locate the limitbetween the two putative sulcal roots.
For the great majority of each sulcal complexstudied, we have been able to find a possible labelingof every putative sulcal root of the generic model.When the Anatomist software had extracted severalsubunits of the sulcal complex (sulcal roots) as onesingle object, by reason of some weaknesses of thecurvature-based image analysis process, the putativeannectent gyrus was identified either as a point ofreduced depth, or as a discontinuity in the orienta-tion of the medial surface of the sulcus. The labelingof the external part of 20 right frontal hemispheres isproposed in Fig. 6.
The main result of analyzing the 3D morphologyof the cerebral cortex according to this strategy wasto allow the description of two main directions oforganization of gyri. The identification of annectentgyri allowed us to follow the hidden continuity ofsuperficial gyri. There were preferential trends oforganization according to two axes: rostrocaudaland dorsoventral. The two extremities or poles ofthe dorsoventral ``meridians'' are the cingulategyrus on the inner surface and the insular corelaterally. For example there is a clear continuitybetween the precentral insular gyrus, the precentralgyrus, the gyrus of the paracentral lobulus, and theparacentral annectent gyrus.
Perpendicular to the above, the rostrocaudalgradient is organized in ``parallel'' following theembryologic curvature of the telencephalon. Forexample, the gyri of F2 follow this organizationin parallel with the intermediate parieto-frontalannectent gyrus, separating the two sulcal roots ofthe CS hidden in the depth of the CS, and with theintermediate parieto-postcentral annectent gyrus,separating the superior and the inferior postcentral
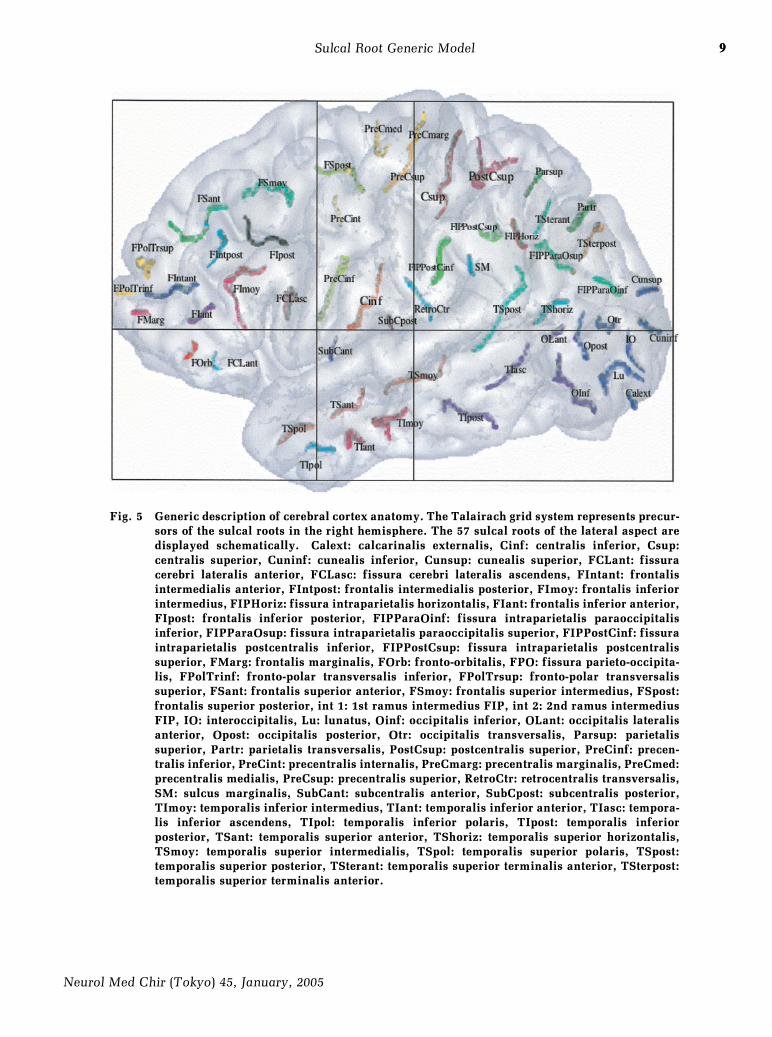
9
Fig. 5 Generic description of cerebral cortex anatomy. The Talairach grid system represents precur-sors of the sulcal roots in the right hemisphere. The 57 sulcal roots of the lateral aspect aredisplayed schematically. Calext: calcarinalis externalis, Cinf: centralis inferior, Csup:centralis superior, Cuninf: cunealis inferior, Cunsup: cunealis superior, FCLant: fissuracerebri lateralis anterior, FCLasc: fissura cerebri lateralis ascendens, FIntant: frontalisintermedialis anterior, FIntpost: frontalis intermedialis posterior, FImoy: frontalis inferiorintermedius, FIPHoriz: fissura intraparietalis horizontalis, FIant: frontalis inferior anterior,FIpost: frontalis inferior posterior, FIPParaOinf: fissura intraparietalis paraoccipitalisinferior, FIPParaOsup: fissura intraparietalis paraoccipitalis superior, FIPPostCinf: fissuraintraparietalis postcentralis inferior, FIPPostCsup: fissura intraparietalis postcentralissuperior, FMarg: frontalis marginalis, FOrb: fronto-orbitalis, FPO: fissura parieto-occipita-lis, FPolTrinf: fronto-polar transversalis inferior, FPolTrsup: fronto-polar transversalissuperior, FSant: frontalis superior anterior, FSmoy: frontalis superior intermedius, FSpost:frontalis superior posterior, int 1: 1st ramus intermedius FIP, int 2: 2nd ramus intermediusFIP, IO: interoccipitalis, Lu: lunatus, Oinf: occipitalis inferior, OLant: occipitalis lateralisanterior, Opost: occipitalis posterior, Otr: occipitalis transversalis, Parsup: parietalissuperior, Partr: parietalis transversalis, PostCsup: postcentralis superior, PreCinf: precen-tralis inferior, PreCint: precentralis internalis, PreCmarg: precentralis marginalis, PreCmed:precentralis medialis, PreCsup: precentralis superior, RetroCtr: retrocentralis transversalis,SM: sulcus marginalis, SubCant: subcentralis anterior, SubCpost: subcentralis posterior,TImoy: temporalis inferior intermedius, TIant: temporalis inferior anterior, TIasc: tempora-lis inferior ascendens, TIpol: temporalis inferior polaris, TIpost: temporalis inferiorposterior, TSant: temporalis superior anterior, TShoriz: temporalis superior horizontalis,TSmoy: temporalis superior intermedialis, TSpol: temporalis superior polaris, TSpost:temporalis superior posterior, TSterant: temporalis superior terminalis anterior, TSterpost:temporalis superior terminalis anterior.
9
Neurol Med Chir (Tokyo) 45, January, 2005
Sulcal Root Generic Model
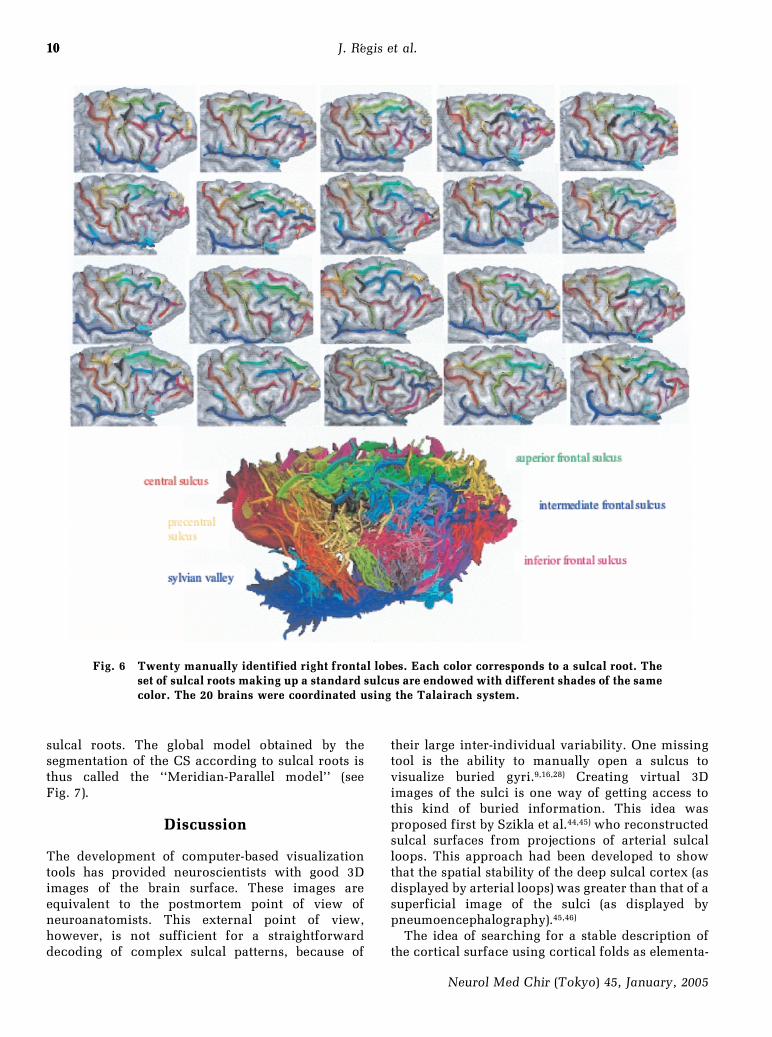
10
Fig. 6 Twenty manually identified right frontal lobes. Each color corresponds to a sulcal root. Theset of sulcal roots making up a standard sulcus are endowed with different shades of the samecolor. The 20 brains were coordinated using the Talairach system.
10
Neurol Med Chir (Tokyo) 45, January, 2005
J. R áegis et al.
sulcal roots. The global model obtained by thesegmentation of the CS according to sulcal roots isthus called the ``Meridian-Parallel model'' (seeFig. 7).
Discussion
The development of computer-based visualizationtools has provided neuroscientists with good 3Dimages of the brain surface. These images areequivalent to the postmortem point of view ofneuroanatomists. This external point of view,however, is not sufficient for a straightforwarddecoding of complex sulcal patterns, because of
their large inter-individual variability. One missingtool is the ability to manually open a sulcus tovisualize buried gyri.9,16,28) Creating virtual 3Dimages of the sulci is one way of getting access tothis kind of buried information. This idea wasproposed first by Szikla et al.44,45) who reconstructedsulcal surfaces from projections of arterial sulcalloops. This approach had been developed to showthat the spatial stability of the deep sulcal cortex (asdisplayed by arterial loops) was greater than that of asuperficial image of the sulci (as displayed bypneumoencephalography).45,46)
The idea of searching for a stable description ofthe cortical surface using cortical folds as elementa-
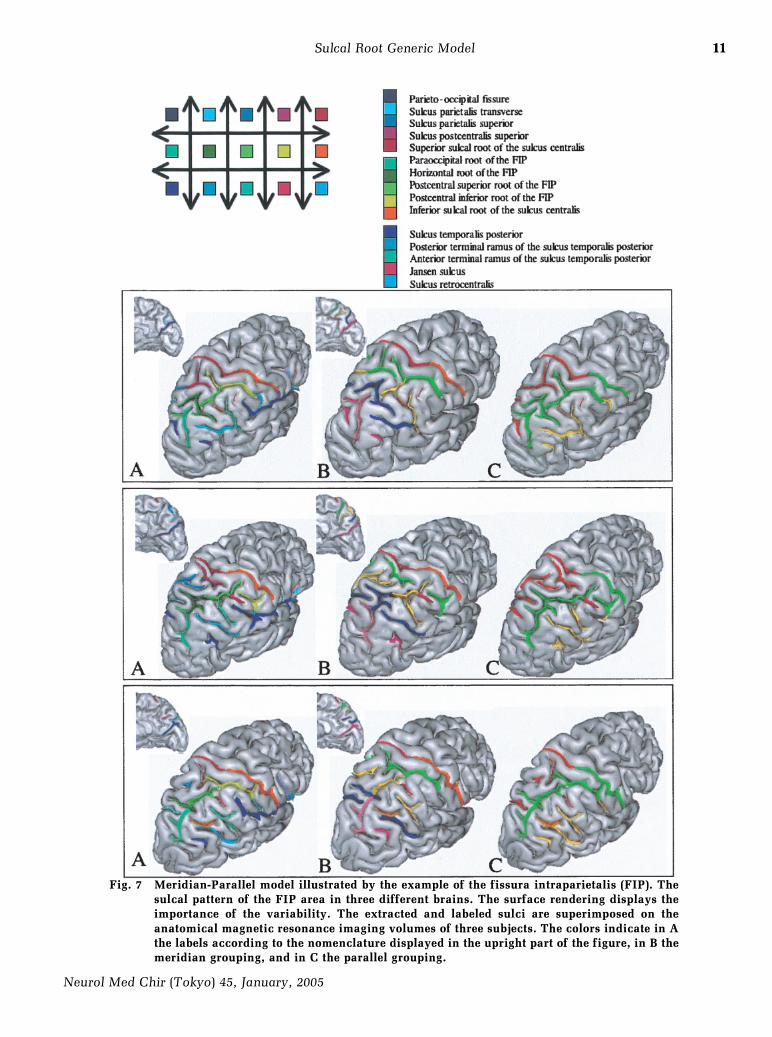
11
Fig. 7 Meridian-Parallel model illustrated by the example of the fissura intraparietalis (FIP). Thesulcal pattern of the FIP area in three different brains. The surface rendering displays theimportance of the variability. The extracted and labeled sulci are superimposed on theanatomical magnetic resonance imaging volumes of three subjects. The colors indicate in Athe labels according to the nomenclature displayed in the upright part of the figure, in B themeridian grouping, and in C the parallel grouping.
11
Neurol Med Chir (Tokyo) 45, January, 2005
Sulcal Root Generic Model

1212
Neurol Med Chir (Tokyo) 45, January, 2005
J. R áegis et al.
ry units may be surprising at first glance, becausethese objects are not pieces of cortex but ``empty''space separating two gyri. The elementary function-al modules embedded into the cortical surface,indeed, are usually supposed to be related to gyri.No simple morphological definition, however, canallow a segmentation of the cortical surface intogyral units, whereas cortical folds can be associatedwith simple topological objects. Here, cortical foldsare considered as indicators of the meeting linebetween the cortex of the two neighboring gyriburied in the fold's depth rather than the line corre-sponding with the superficial drawing of the foldson the external brain surface (cf. Fig. 5). Conse-quently, various algorithmic approaches can bedevised to convert a sulcal-based representation intoa gyral-based division.5) Finally, one key idea under-lying our work is the possible limiting value of mostof the cortical folds relative to a functional level thatis yet unknown. This point will be developed furtherbelow.
The putative sulcal root map proposed in thispaper is the synthesis of 10 years of work on variousdata from the literature and our own experiments.Since such data are more abundant for some regionsthan for some others, the quality of this map may notbe uniform. Hence, our current sulcal root model islikely to be further refined during the coming years.The manual matching performed during ourexperiments between some individual brains andour putative sulcal root map involves a measure ofsubjectivity. Therefore, the results cannot beconsidered as a definitive validation of our initialhypothesis on the existence of a stable map ofindivisible cortical folds. The validation of this idearequires a long-term research program consisting ofcorrelating manual identifications of the putativesulcal roots with individual activation, cytoarchitec-tony, or connectivity-based cortical maps. Thus, thework presented in this paper aims mainly to showthat the new tools dedicated to the manipulation ofsulcal patterns are now sufficiently mature tosupport the testing of anatomical models, such asthe sulcal root model. A typical underlying neu-roscience question would be that of the possible linkbetween the fronto-parietal `pli de passage' splittingthe CS and the hand-related functional areas.1,42,57)
Another direction of research that may providesome validation of the sulcal root idea relies inmathematical analysis of the shape of the corticalsurface. For instance, recent results have shown thata curvature study might be able to automaticallyhighlight the fronto-parietal pli de passage splittingthe CS.5) Parceling of the cortical surface usingsulcal depth has recently led another team to
propose a map of ``sulcal basins,'' namely areas ofmaximal depth. This shares striking similarities withour sulcal root map.20) Further efforts should consistof retrograde simulation of the cortical surfacefolding process in order to recover the chronology ofthe merging of sulcal roots. The data of choice forreaching a better understanding of this process, ofcourse, would be longitudinal acquisitions of braingrowth using antenatal MR or ultrasound images.Such studies, however, may be ethically questiona-ble.
One key point to be discussed is whether a sulcalroot type model is stable across individuals. Study ofcerebral cortical folding patterns has often beenconsidered as uninteresting insofar as the foldingprocess only resulted from the need to fit a growingsmooth organ (entero äƒde process) into the skull.Hence, the sulcal patterns were thought to embedlittle information regarding the underlying organiza-tion of the cortex. Another point of view, however,which is advocated in this paper, stems from thepossible link between cortical organization and thevarious growth and mechanical processes resultingin folding.
The cerebral cortex is divided into anatomicallyand functionally distinct areas, forming a species-specific area map across the cortical sheet. For thegrowth processes to be able to induce folding atspecific locations relative to these cortex areas, thearea map has to exist before the folding begins. Thetraditional ``tabula rasa'' model presumes that theembryonic telencephalon first forms an equipoten-tial sheet of cells that then becomes specified byinput from subcortical centers,27) which raises thequestion of the relative timing between folding andthe development of connectivity. The alternative``protomap'' model, however, for which cells of theembryonic vesicle themselves carry intrinsic pro-grams for species-specific cortical regionalization,has been recently bolstered by many new data.14,25,31)
Therefore, it seems reasonable to hypothesize thatthe first stage of the folding process might be deeplyinfluenced by such a protomap. Assuming thestability of the protomap across individuals, becauseof its species-specificity, this should imply stabilityof the beginning of the folding process duringgestational, something which has effectively beenobserved.48) Hence, our quest for a sulcal root typemodel is highly motivated by the hypothesis that thefolding process gives clues to the protomap bounda-ries.
Striking histological differences have been ob-served between the fundi, walls, and gyral crowns ofsulci.53) Assuming that the cortex buried in thebottom of the folds at the adult stage corresponds to

1313
Neurol Med Chir (Tokyo) 45, January, 2005
Sulcal Root Generic Model
the places where the folding process begins at thefetal stage leads us to hypothesize the existence ofprosulcal and progyral maps embedded in theprotomap. This hypothesis does not imply that thewhole prosulcal map has to turn into a fold, but thatfolds are bound to occur somewhere into this maprelated to ``boundaries'' between cortical areas. Ofcourse, the notion of boundaries is not straightfor-ward and may embed for instance multiple scales ofdescription. In any event, while it is far from asystematic rule, some boundaries between architec-tonic maps do clearly correspond to some folds.60)
Patterns of connectivity are also related to corticalfolding. For instance, some secondary folds fre-quently split a gyrus into areas with distinct connec-tivity.54) It is finally interesting to note that primaryareas which are stable relative to cytoarchitectonyusually correspond to the bottom of main sulci. Incontrast, it is obvious that the majority of themultimodal associative areas, which develop later inontogenesis and phylogenesis, are located in thecortex of gyral crowns and that of second levelsulcal walls. More precisely, when one considers thearchitectonic borders inside a region or a gyrus thehierarchy of complexity of the areas or the function-al hierarchy of these map generally follows an in-outgradient (with external areas more integrative).These areas, whose link with architectony is farfrom clear, are then surrounded by very variablesulcal patterns.
Welker and Seidenstein,55) describing a segmenta-tion of the sensorimotor areas of the raccoon basedon morphological analysis of the sulco-gyralvariability, have demonstrated a good correspon-dence between these anatomical subunits andsomatotopic functional maps as defined fromevoked potentials. In spite of large differences insize and shape, the cortical folds always lay betweengyri that contained peripherally distinct representa-tion. The conclusion of the authors was that inter-animal variations in gyral-sulcal arrangement doreflect individual differences in underlying connec-tivity and functional organization. Following themodel introduced previously, our own interpreta-tion of these variable folding patterns is related tothe variable development of each somestheticrepresentation. So, on an individual basis, the partof the prosulcal map turning into folds variesbecause of different pressures for folding.
Extrapolation from raccoons to humans isdifficult, because the human cortex is made up ofseveral levels of folding. In our opinion, the stabilityof the first folding level corresponding to the puta-tive sulcal root map is hidden behind the variabilityof the second folding level. Fortunately, information
buried in sulcal depths reveals this first folding level,which is thought to represent the gross organizationof the protomap. Following the raccoon results,55)
the second level of folding is also thought to befunctionally significant as a subset of the boundariesof the full cortical area map. Thus, we firmly believethat there is a functional meaning of the convolu-tions in the sulci and gyri. This point of view isrelated to the principle of tensegrity, which can bedefined as the way a biological system reaches itsfinal shape via stabilization of the sum of tensionsand compressions stemming from its differentparts.17) This principle is supposed to be highlyinfluential during morphogenesis. Such ideas havebeen developed recently by Van Essen50) for thebrain through the hypothesis that fiber-relatedtension may be the main explanation of the pattern-ing of cortical folds.
Accepting the hypothesis that the main architec-tural constraints are related to white matter tracts(connection patterns of axons and dendrites) leadsrapidly to the conclusion that the local gyrogenesismust be dependent on underlying functionalorganization. Considering the complexity of corticalconnectivity, however, one can guess that severalqualitatively different folding patterns might resultfrom the stabilization process-driving morphogene-sis. We can also hypothesize that slight differencesin the sizes of cortical areas might lead to radicallydifferent folding solutions during morphogenesis,because of chaotic behavior in the system,47) whichwould explain the variability of adult corticalfolding patterns. Hence, simulating the process ofcortical morphogenesis from the tensegrity point ofview seems a promising research avenue, whichmight improve our understanding of corticalorganization. Recent progress in diffusion-weightedMR imaging provides information on fiber bundletrajectories30) which may open the door to retro-grade simulation of the morphogenesis stabilizationprocess.
Some have argued that what are called axial sulci,namely large folds located by definition in themiddle of homogeneous cytoarchitectonic areas, aregood examples of sulci without function- limitingvalue. In fact, in the majority of such cases, someother types of functional aspect can be found tosegregate the two areas of identical cytoarchitec-tony. For instance, the split induced by the calcarinesulcus in area 17 (V1 in the visual system) distin-guishes upper and lower hemifields, which havevery different roles for the visual system.8) Cytoar-chitectony is only one of the features that can beused to distinguish cortical areas. Connectivitypattern, for instance, is another key feature,43) which

1414
Neurol Med Chir (Tokyo) 45, January, 2005
J. R áegis et al.
often leads to gyral subdivisions in accordance withthe second level of folding.53)
Another misleading argument against the func-tional limiting value of cortical folds results from thepoor correlation between sulcal lines drawn on thebrain external surface and architectonic subdivi-sions (Fig. 4B). Thus, architectonic borders must becompared with the cortical surface in full 3Drepresentations of the cortex including the corticalsurface bordered in the sulci.60) The tensegrity pointof view, however, does not give any meaning to thisline. Morphogenesis, indeed, is only thought to keepcortical areas inside a gyrus, in order to favor shortaxonal connections. The lines of the bottoms of thesulci are the only landmarks thought to give cluesabout functional organization. Another similarargument consists of claiming that most of thearchitectonic transitions are not marked by a fold.Such situations, in fact, are predicted by the ten-segrity point of view, which links folding to a localpressure to increase cortical surface. Hence, foldinghas no reason to occur along any boundary.
A last argument that may be raised against anyrelevant meaning of the folding patterns is the lownumber of experimental results linking functionalactivation with specific folds outside primary areas.It should be noted, however, that the currentstrategy of the brain mapping community, whichconsists of averaging brains without perfect defor-mation of the folds, prevents such discoveries.Furthermore, our limited capacity to identifyreliably generic sulco-gyral patterns across individ-uals is a huge handicap for such studies. In thispaper, we intend to provide a new framework fordealing with sulco-gyral patterns, that may allowscientists to analyze functional data, as in the studyof Watson et al.52) A positron emission tomographystudy of the V5 area showed large inter-individualvariability in the location of this visual area inTalairach coordinates. A careful study of corticalfolding patterns, however, revealed a stable localiza-tion of V5 at the crossing point between theposterior root of the sulcus temporalis inferior andthe anterior root of the sulcus occipitalis lateralis.52)
The sulcal root concept proposed in this paperand the related notion of ``pli de passage'' lead us toreconsider the organization of the cortical surface.The attempt to elaborate general principles thatmight underlie the sulcal root map led us to proposethe Meridian-Parallel model, named by analogy withearth's latitude and longitude system (Fig. 4). Thismodel considers the cortical surface as a network oforthogonal gyri, which leads to a new definition ofsulcal roots as the deep sulcal cortex surrounded bytwo orthogonal pairs of gyri. One pair follows the
meridian direction, the first gyrus being locatedrostrally to the sulcal root and the second onecaudally. The other pair follows the paralleldirection, the first gyrus being located dorsally tothe sulcal root and the second one ventrally.
These two orthogonal axes of gyral organizationare especially visible at the fetal stage (Fig. 1). At theadult stage, however, one axis is usually dominantand the other axis buried (annectent gyri), because itseems to be a good folding solution to stabilize thevarious forces underlying the growing process. Forinstance, the ``meridian'' dorsoventral axis is usuallydominant in the central region, while the ``parallel''rostrocaudal axis is usually dominant in the frontalregion. According to the predominance of one gyralaxis, several sulcal roots corresponding to a meridi-an or a parallel direction can be buried in the samelong furrow. The Meridian-Parallel model interpretsunusual folding patterns as a flip of dominant gyraldirection compared to the majority of individuals(Fig. 7). Opening the sulci, however, should alwaysreveal the dual orthogonal gyri. Hence, in suchflipped cases, the sulcal roots buried in the depth ofthe furrows would be the reliable landmarksproviding the link with standard folding patterns.
Sanides41) and Pandya and Yeterian29) havedescribed in the monkey preferential trends ofcytoarchitectonic organization according to tworostrocaudal and dorsoventral axes. The dorsoven-tral axis of the stream of differentiation fits very wellwith this orientation of our meridians going fromthe limbic archeocortical cingulate core to the lim-bic paleocortical insular core.29) These phylogeneticand cytoarchitectonic observations fit well with ourstructural analysis of the organization of the foldingof the human cerebral cortex. The region of theinsula corresponds to the paleocortical pole mergingwith neocortex through cytoarchitectonic waves.24)
These waves are perpendicular to the dorsoventralaxis but the sulci of the ``insular lobe'' diverge like afan and indicate the radial direction of the dorsoven-tral axis. As far as functional systems are concerned,the Meridian-Parallel model also seems pertinent.Thus, in the central region the dorsoventral axiscorresponds to the somatotopic level of organiza-tion. The rostrocaudal axis is orientated orthogonal-ly, following the curvature of the telencephalon.This axis in the central region corresponds to agradient of modality.
Some of the Meridian-Parallel model gyral linesdo not stem from straightforward architecturalsubdivisions. Architectony, however, may stillembed much hidden information. In the frontal lobe,for instance, the parallel direction is usually theprominent one (F1, F2, F3) but in fact some other

1515
Neurol Med Chir (Tokyo) 45, January, 2005
Sulcal Root Generic Model
information such as the maps defined by Braak,2)
relying on the pigmentoarchitectonical method,correlates well with the existence of a dual meridiandirection in these areas. In any event, the Meridian-Parallel model is only a general principle of organi-zation, which cannot provide an explanation foradditional local gyral subdivision, which may cor-respond to further segregation of the cortical sur-face. This principle, however, helps for instance todevelop interspecies comparisons. For instance, thefrontal lobe of macaques can be split into precentral,retroarcuate, and prearcuate meridian sectors anddorsomesial, dorsolateral, ventromesial, and ven-trolateral parallel sectors. If we regard the inter-mediate frontal sulcus as the equivalent of the sulcusprincipalis, we can propose it as the boundarybetween the dorsal and the ventral part of themeridian sectors. Much work remains to be done tovalidate or negate the pertinence of this kind ofcomparison.
Conclusions
This paper suggests the reappraisal of a classicalanatomical phenomenon well described during theXIXth century and practically forgotten during theXXth century: that of the plis de passage, namely thegyri buried in the depth of the sulci. We have shownthat this phenomenon may be of great importancefor the understanding (decryption) of variability incerebral cortical gyration. Taking into account theseburied gyri has led us to propose a generic model offolding patterns based on a constant number ofsulcal roots. These sulcal roots are separated by gyriorientated according to two orthogonal directions(rostrocaudal and dorsoventral). Usually, onedirection of gyri is buried in the depth of the furrowsconstituted by the gyri of the orthogonal prominentdirection. This burying process is thought to resultfrom a trade-off between the various folding pres-sures occurring during brain growth. Thus, thevariability of the folding patterns is thought to stemfrom differential expression of cortical areas (interms of surface) from one individual to another,leading to variably buried gyri and sometimes to aflip of the prominent gyral direction. According tothis hypothesis, the huge variability of the superfi-cial appearance of the cortex only results from thechaotic behavior of the folding process, and does notimply a large variability of cortical organization.Our point of view, indeed, presupposes a constantprotomap with constant topography of cortical areasin each individual. This new approach enables us touse the same nomenclature for the description ofany nonpathological individual, and to endow the
cortical surface with a 2D reference system that mayopen new doors to brain mapping projects.
Acknowledgments
We thank Dr. Roderick Duncan (Lead Clinician,West of Scotland Regional Epilepsy Service,Southern General Hospital, Glasgow, Scotland,U.K.) for his help.
This work was supported by the French Ministryof Health.
References
1) Boling W, Olivier A, Bittar RG, Reutens D: Localiza-tion of hand motor activation in Broca's pli depassage moyen. J Neurosurg 91: 903–910, 1999
2) Braak H: The pigment architecture of the humanfrontal lobe precentral, subcentral and frontal region.Anat Embryol (Berl) 154: 35–68, 1979
3) Brett M, Johnsrude IS, Owen AM: The problem offunctional localization in the human brain. Nat RevNeurosci 3: 243–249, 2002
4) Cachia A, Mangin J, Rivi àere D, Kherif F, Boddaert N,Andrade A, Papadopoulos-Orfanos D, Poline JB,Bloch I, Zilbovicius M, Sonigo P, Brunelle F, R áegis J:A primal sketch of the cortex mean curvature: amorphogenesis based approach to study the variabil-ity of the folding patterns. IEEE Trans Med Imaging22: 754–765, 2003
5) Cachia A, Mangin JF, Rivi àere D, Kherif F,Papadopoulos-Orfanos D, Bloch I, R áegis J: A Genericframework for parcellation of the cortical surfaceinto gyri using geodesic Vorono äƒ diagrams. MedicalImage Analysis 7: 403–416, 2003
6) Chi JG, Dooling EC, Gilles FH: Gyral development ofthe human brain. Ann Neurol 1: 86–93, 1977
7) Chicurel M: Databasing the brain. Nature 406:822–825, 2000
8) Crawford MLJ: Stimulus-specific columns in monkeyvisual cortex as revealed by the (14C)-2-deoxyglucosemethod, in Peters A, Jones DW (eds): Cerebral Cortex.New York, Plenum, 1985, pp 331–349
9) Cunningham DJ, Horsley V: Contribution to thesurface anatomy of the cerebral hemispheres. Dublin,Royal Irish Academy, 1892
10) Davatzikos C, Bryan RN: Morphometric analysis ofcortical sulci using parametric ribbons: a study of thecentral sulcus. J Comput Assist Tomogr 26: 298–307,2002
11) Dejerine J: [Anatomy of Central Nervous System].Paris, Masson, 1895 (Fre)
12) Eberstaller O: [The Frontal Lobe. A Contribution forthe Anatomy of the Surface of the Rough Brain].Vienna, Urban & Schwarzenberg, 1890 (Ger)
13) Feess-Higgins A, Larroche J-C: Development of theHuman Foetal Brain. An Anatomical Atlas. Paris,Masson, 1987

1616
Neurol Med Chir (Tokyo) 45, January, 2005
J. R áegis et al.
14) Fukuchi-Shimogori T, Grove E: Neocortex patterningby the secreted signaling molecule FGF8. Science294: 1071–1074, 2001
15) Giacomini C: [Pattern of Gyration in Human CerebralCortex]. Torino, Arch Ital Biol, 1882 (Ita)
16) Gratiolet LP: [On the Folding of Cortical Folding of theHuman and Primates Brain]. Paris, Bertrand, 1854(Fre)
17) Ingber DE: Cellular tensegrity: defining new rules ofbiological design that govern the cytoskeleton. J CellSci 104: 613–627, 1993
18) Kotter R: Neuroscience databases: tools for exploringbrain structure-function relationships. Philos Trans RSoc Lond B Biol Sci 356: 1111–1120, 2001
19) Le Goualher G, Procyk E, Collins DL, Venugopal R,Barillot C, Evans AC: Automated extraction andvariability analysis of sulcal neuroanatomy. IEEETrans Med Imaging 18: 206–217, 1999
20) Lohmann G, von Cramon DY: Automatic labelling ofthe human cortical surface using sulcal basins. MedImage Anal 4: 179–188, 2000
21) Mangin JF, Frouin V, Bloch I, R áegis J, Lopez-Krahe J:From 3D magnetic resonance images to structuralrepresentations of the cortex topography usingtopology preserving deformations. Journal of Mathe-matical Imaging and Vision 5: 297–318, 1995
22) Mazziotta J, Toga A, Evans A, Fox P, Lancaster J,Zilles K, Woods R, Paus T, Simpson G, Pike B,Holmes C, Collins L, Thompson P, MacDonald D,Iacoboni M, Schormann T, Amunts K, Palomero-Gallagher N, Geyer S, Parsons L, Narr K, Kabani N,Le Goualher G, Boomsma D, Cannon T, KawashimaR, Mazoyer B: A probabilistic atlas and referencesystem for the human brain: International Consorti-um for Brain Mapping (ICBM). Philos Trans R SocLond B Biol Sci 356: 1293–1322, 2001
23) Mazziotta JC, Toga AW, Evans A, Fox P, Lancaster J:A probabilistic atlas of the human brain: theory andrationale for its development. The InternationalConsortium for Brain Mapping (ICBM). Neuroimage2: 89–101, 1995
24) Mesulam MM, Mufson EJ: Insula of the old worldmonkey. I. Architectonics in the insulo-orbito-temporal component of the paralimbic brain. J CompNeurol 212: 1–22, 1982
25) Miyashita-Lin EM, Hevner R, Wassarman KM,Martinez S, Rubenstein JR: Early neocorticalregionalization in the absence of thalamic innerva-tion. Science 285: 906–909, 1999
26) Oldfield RC: The assessment and analysis of handed-ness: the Edinburgh inventory. Neuropsychologia 9:97–113, 1971
27) O'Leary DD: Do cortical areas emerge from aprotocortex? Trends Neurosci 12: 400–406, 1989
28) Ono M, Kubik S, Abernathey CD: Atlas of theCerebral Sulci. Stuttgart, Georg Thieme Verlag, 1990
29) Pandya DN, Yeterian EH: Architecture and connec-tions of cortical association areas, in Peters A, JonesEG (eds): Cerebral Cortex. New York, Plenum Press,1985, pp 3–61
30) Poupon C, Clark CA, Frouin V, R áegis J, Bloch I, LeBD, Mangin J: Regularization of diffusion-baseddirection maps for the tracking of brain white matterfascicles. Neuroimage 12: 184–195, 2000
31) Rakic P: Neurobiology. Neurocreationism — makingnew cortical maps. Science 294: 1011–1012, 2001
32) R áegis J: [Sulcal roots anatomy and functionalmapping of the cerebral cortex]. Marseille, Univer-sit áe d'Aix Marseille II, 1994, p 272 (thesis) (Fre)
33) R áegis J, Mangin JF, Frouin V, Sastre F, Peragut JC,Samson Y: Generic model for the localization of thecerebral cortex and preoperative multimodalintegration in epilepsy surgery. Stereotact FunctNeurosurg 65: 72–80, 1995
34) Reitzus: [The Human Brain. Studies in the Macroscop-ic Morphology]. Stockholm, Norstedt & S äorner, 1896(Ger)
35) Rettmann ME, Han X, Xu C, Prince JL: Automatedsulcal segmentation using watersheds on the corticalsurface. Neuroimage 15: 329–344, 2002
36) Rivi àere D, Mangin JF, Papadopoulos-Orfanos D,Martinez JM, Frouin V, R áegis J: Automatic recogni-tion of cortical sulci of the human brain using acongregation of neural networks. Med Image Anal 6:77–92, 2002
37) Roland PE, Geyer S, Amunts K, Schormann T,Schleider A, Malikovic A, Zilles K: Cytoarchitecturalmaps of the human brain in standard anatomicalspace. Hum Brain Mapp 5: 222–227, 1997
38) Roland PE, Svensson G, Lindeberg T, Risch T,Baumann P, Dehmel A, Frederiksson J, HalldorsonH, Forsberg L, Young J, Zilles K: A database genera-tor for human brain imaging. Trends Neurosci 24:562–564, 2001
39) Roland PE, Zilles K: Brain atlases — a new researchtool. Trends Neurosci 17: 458–467, 1994
40) Royackkers N, Desvignes M, Fawal H, Revenu M:Detection and statistical analysis of human corticalsulci. Neuroimage 10: 625–641, 1999
41) Sanides F: Comparative architectonics of the neocor-tex of mammals and their evolutionary interpreta-tion. Ann N Y Acad Sci 167: 404–423, 1969
42) Sastre-Janer FA, R áegis J, Belin P, Mangin JF,Dormont D, Masure MC, Remy P, Frouin V, SamsonY: Three-dimensional reconstruction of the humancentral sulcus reveals a morphological correlate ofthe hand area. Cereb Cortex 8: 641–647, 1998
43) Stephan KE, Kamper L, Bozkurt A, Burns GA, YoungMP, Kotter R: Advanced database methodology forthe Collation of Connectivity data on the Macaquebrain (CoCoMac). Philos Trans R Soc Lond B Biol Sci356: 1159–1186, 2001
44) Szikla G, Bouvier G, Hori T: In vivo localization ofbrain sulci by arteriography: a stereotacticanatomoradiological study. Brain Res 95: 497–502,1975
45) Szikla G, Bouvier G, Hori T, Petrov V: Angiography ofthe Human Brain Cortex. Berlin, Springer-Verlag,1977
46) Talairach J, Szikla G, Tournoux P, Prosalentis A,

1717
Neurol Med Chir (Tokyo) 45, January, 2005
Sulcal Root Generic Model
Bordas-Ferrier M: [Stereotactic Atlas of the Telen-cephalon]. Paris, Masson, 1967 (Fre)
47) Thom R: Structural Stability and Morphogenesis: anOutline of a General Theory of Models. New York,Benjamin Addison Wesley, 1975
48) Turner OA: Growth and development of the cerebralpattern in man. Arch Neurol Psychiatry 59: 1–12,1948
49) Vaillant M, Davatzikos C: Finding parametricrepresentations of the cortical sulci using an activecontour model. Med Image Anal 1: 295–315, 1997
50) Van Essen DC: A tension-based theory of morpho-genesis and compact wiring in the central nervoussystem. Nature 385: 313–318, 1997
51) Waterston D: Complete bilateral interruption of thefissure of Rolando. J Anat XLI: 143–146, 1907
52) Watson JD, Myers R, Frackowiak RS, Hajnal JV,Woods RP, Mazziotta JC, Shipp S, Zeki S: Area V5 ofthe human brain: evidence from a combined studyusing positron emission tomography and magneticresonance imaging. Cereb Cortex 3: 79–94, 1993
53) Welker W: Why does cerebral cortex fissure and fold?A review of determinants of gyri and sulci, in JonesEG, Peters A (eds): Comparative Structure andEvolution of Cerebral Cortex. New York, PlenumPress, 1990, pp 3–136
54) Welker WI: Correlation between nuclear morphologyand somatotopic organisation in ventro-basal com-plex of the raccoon's thalamus. J Anat 99: 761–790,1965
55) Welker WI, Seidenstein S: Somatic sensory represen-tation in the cerebral cortex of the raccoon. J CompNeurol 111: 469–501, 1959
56) White LE, Andrews TJ, Hulette C, Richards A,Groelle M, Paydarfar J, Purves D: Structure of thehuman sensorimotor system. I: Morphology andcytoarchitecture of the central sulcus. Cereb Cortex 7:18–30, 1997
57) Yousry TA, Schmid UD, Alkadhi H, Schmidt D,Peraud A, Buettner A, Winkler P: Localization of themotor hand area to a knob on the precentral gyrus. Anew landmark. Brain 120: 141–157, 1997
58) Zeng X, Staib LH, Schultz RT, Duncan JS: Segmenta-tion and measurement of the cortex from 3-D MRimages using coupled-surfaces propagation. IEEETrans Med Imaging 18: 927–937, 1999
59) Zhou Y, Thompson PM, Toga AW: Extracting andrepresenting the cortical sulci. IEEE Comput GraphAppl 19: 49–55, 1999
60) Zilles K, Schleicher A, Langemann C, Amunts K,Morosan P, Palomero-Gallagher N, Schormann T,Mohlberg H, B äurgel U, Steinmetz H, Schlaug G,Roland PE: Quantitative analysis of sulci in humancerebral cortex: Development, regional heterogenei-ty, gender difference, asymmetry, intersubjectvariability and cortical architecture. Hum BrainMapp 5: 218–221, 1997
Address reprint requests to: J. R áegis, M.D., Service deNeurochirurgie Fonctionnelle et St áer áeotaxique,C.H.U. La Timone, 264 rue Saint Pierre, 13385Marseille CEDEX 05, France.e-mail: jregis@ap-hm.fr




















