
Subgenotype D5, BCP and MHR mutations in hepatic complications among hepatitis B virus infected...
9
(This is a sample cover image for this issue. The actual cover is not yet available at this time.) This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
-
Upload
utkaluniversityutkaluniversity -
Category
Documents
-
view
3 -
download
0
Transcript of Subgenotype D5, BCP and MHR mutations in hepatic complications among hepatitis B virus infected...

(This is a sample cover image for this issue. The actual cover is not yet available at this time.)
This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Subgenotype D5, BCP and MHR mutations in hepatic complicationsamong hepatitis B virus infected patients from Orissa, India
Rajesh Panigrahi a, Avik Biswas a, Arup Banerjee a, Shivaram Prasad Singh c, Manas K. Panigrahi c,Anne Marie Roque-Afonso e, Haribhakti Seba Das c, Pradip K. Mahapatra b, Sekhar Chakrabarti d,Runu Chakravarty a,⇑a ICMR Virus Unit, Kolkata, ID & BG Hospital Campus, Kolkata, Indiab Department of Chemistry, Jadavpur University, Kolkata, Indiac SCB Medical College, Dept. of Gastroenterology, Cuttack, Orissa, Indiad National Institute of Cholera and Enteric Diseases, Kolkata, Indiae INSERM U785, Univ Paris-Sud AP-HP, Hôpital Paul Brousse, Virologie Villejuif, 94804, France
a r t i c l e i n f o
Article history:Received 13 March 2012Received in revised form 30 June 2012Accepted 30 June 2012Available online 20 July 2012
Keywords:HBV subgenotype D5MHR mutationsBCP mutationsOrissa
a b s t r a c t
The study was undertaken to investigate the clinical implications of hepatitis B virus (HBV) genotypes,basal core promoter (BCP), precore (PC) and surface gene mutations in HBV infected patients from Orissa,southeastern India. HBV infections were identified by serology testing and HBV DNA amplification bypolymerase chain reaction among the 152 patients. After sequencing, surface gene mutation were studiedby sequence analysis as well as by using BLOSUM scores and BCP mutations were studied only bysequence analysis. A high proportion of HBV/D5 (66.0%) was found among the study samples having sig-nificant relation with liver cirrhosis (LC) and hepatocellular carcinoma (HCC) patients (p < 0.05). The BCPmutation, TA (81.4%) and C1753/TA (75.0%) was found in significant proportion (p < 0.05) among HCCcases and in fact a gradual increase in these mutations were noted between inactive carriers (IC) toHCC group and also showed higher viral load. An increasing trend of major hydrophilic region (MHR)mutations in S gene was also observed from IC (56.0%) to chronic liver disease (CLD) (60.4%) to LC(72.4%) to HCC (95.0%) patients. In conclusion, our study suggests that the predominant HBV subgenotypeHBV/D5 with high viral load and BCP mutations (double and triple) and high mutations in MHR regionwas significantly associated with advanced liver disease (LC and HCC) and might act as predictor ofsevere hepatic complications.
� 2012 Elsevier B.V. All rights reserved.
1. Introduction
Hepatitis B virus (HBV) affects 350–400 million people world-wide and accounts for one million deaths annually from cirrhosis,liver failure and hepatocellular carcinoma (HCC) (Lok and McMa-hon, 2001). HBV belongs to the family Hepadnaviridae and the sero-logical and genetic heterogeneity of HBV are well established.
Based on >8% divergence, the partially double-stranded HBVDNA approximately of 3.2 kb, with four partially overlapping openreading frames (ORF): surface (S), core (C), polymerase (P) and Xgenes, are divided into eight different genotypes, namely A–H(Kramvis et al., 2005) and several subgenotypes (Norder et al.,2004). HBV genotypes have a distinct geographical distribution.Genotypes B and C are mostly prevalent in Asia. Genotypes A and
D are commonly found in Europe, the Mediterranean region, theMiddle East and India (Norder et al., 2004). It was well documentedthat the clinical outcome of HBV differs in relation to varying geno-type distribution. In Asia HBV genotype C is associated with severeliver disease and a high risk of HCC (Ren et al., 2010).
HBV replicates using reverse transcriptase that lacks proof read-ing function and exhibits a mutation rate more than 10-fold higherthan that of other DNA viruses (Hunt et al., 2000). Among themutations identified in different ORFs, the most common naturallyoccurring mutations are the precore (PC) stop codon mutation atnucleotide (nt) 1896 resulting in A1896 and the basal core pro-moter (BCP) double mutation (T1762/A1764). Previous studieshave shown that the A1896 and T1762/A1764 mutations abolishor reduce the production of hepatitis B e antigen causing(HBeAg)-negative HBV infections (Lok et al., 1994). Studies alsoconfirmed the correlation of HBV genotypes with the developmentof PC and BCP mutations (Lindh et al., 1997; Lok et al., 1994). ThePC mutation is commonly found in genotypes B, D, and E andsometimes in genotype C but rarely in genotype A (Li et al.,
1567-1348/$ - see front matter � 2012 Elsevier B.V. All rights reserved.http://dx.doi.org/10.1016/j.meegid.2012.06.015
⇑ Corresponding author. Address: ICMR Virus Unit, Kolkata, ID & BG HospitalCampus, GB-4, 1st Floor (East Wing), Beliaghata, Kolkata 700 010, India. Tel.: +91 3323537425; fax: +91 33 23537424.
E-mail address: [email protected] (R. Chakravarty).
Infection, Genetics and Evolution 12 (2012) 1622–1629
Contents lists available at SciVerse ScienceDirect
Infection, Genetics and Evolution
journal homepage: www.elsevier .com/locate /meegid

Author's personal copy
1993). However, the BCP double mutation is commonly detected inHBV genotypes C and A but less frequently in genotypes B and D(Tanaka et al., 2004). In contrast, HBV/Ba had the highest frequencyof A1896 but the lowest of T1762/A1764 and HBV/C2 had interme-diate frequencies of these mutations (Wang et al., 2007). Thesemutations were frequently observed in chronically infected pa-tients with active liver disease, severe recurrent liver disease fol-lowing liver transplantation or HCC (Kidd-Ljunggren et al., 1997).Also HCC was found to be highly related with mutations in the coreregion (Huang and Lok, 2011). In addition, the mutations in X geneand BCP may increase the risk of HCC in patients with HBV geno-type C infections (Miyakawa and Mizokami, 2003).
HBsAg is the first marker for the identification of HBV infection.Codons 100–169 is the major hydrophilic region (MHR) of HBsAg,contains the antigenic determinant region which has a conforma-tional structure stabilized by disulfide bridges located betweenconserved cysteine residues (Francois et al., 2001). Amino acid sub-stitutions within this region can lead to alterations in the bindingof HBsAg to anti-HBs that result in immune escape and failure ofimmune based detection (ELISA) of HBsAg.
India with estimated more than 40 million carriers is the secondlargest HBV carrier pool in the world. Genotype D is the predomi-nant HBV genotype in India, with varying prevalence of genotypesA and C in different parts of the country. Studies on disease pro-gression and their association with genotypes showed variable re-sults in different states of India (Datta et al., 2007; Chauhan et al.,2006; Gandhe et al., 2003a,b; Vivekanandan et al., 2004) but datafrom the state of Orissa with a population of was lacking. Further-more, none of the studies included HCC cases.
The present study tried to investigate the distribution of HBVgenotypes/subgenotypes among the different clinical group in Or-issa with special attention to their correlation with advance liverdisease and association with the variability of BCP, PC and MHR re-gion of surface gene and HBV genotype. Our study revealed pres-ence of a high proportion of HBV/D5 with BCP mutations amongthe study samples, having significant relation with liver cirrhosisand HCC. An increasing trend of S gene mutation was also observedfrom inactive carriers to chronic liver disease to liver cirrhosis toHCC patients.
2. Materials and methods
2.1. Study subjects
A total of 255 HBsAg positive patients from Orissa were in-cluded in the study, of which only 152 were HBV DNA positiveand are divided into two groups; Group I consists of fifty inactiveHBsAg carrier state (IC), who were mainly voluntary blood donorsincidentally detected as HBsAg positive. Group II has 102 symp-tomatic subjects who were referred from different clinics in Orissaincludes: thirteen acute hepatitis patients (AC), forty chronic liverdisease (CLD) cases 29 liver cirrhosis (LC) patients and 20 HCCpatients.
Inactive carriers (IC), were mainly asymptomatic carriers,majority being voluntary blood donors who were incidentally de-tected as HBsAg positive, with normal ALT (<40 IU/L; evaluated2–3 times at 3 months interval), Serum HBV DNA >2000 IU/mL(104 copies/mL) and normal liver ultrasonography for more than6 months.
Acute hepatitis was defined as having serum ALT 2.5 times theupper limit of normal as well as elevated serum bilirubin level anda positive IgM anti-HBc test.
Chronic liver disease was defined as symptomatic patients, withpersistently or intermittently elevated ALT (1.5 times upper limitof normal) for a period of more than 6 months, serum HBV DNA
>20,000 IU/mL (105 copies/mL), or >2000–20,000 IU/mL (104–105
copies/mL) for HBeAg negative cases and liver biopsy showing ne-cro-inflammatory changes and fibrosis without nodule formation,with or without ultrasonographic evidence of portal hypertension.
Liver cirrhosis was defined as patients, with clinical features ofliver cell failure; portal hypertension and liver biopsy showingfibrosis with nodule formation, with ultrasonographic evidenceof portal hypertension.
Hepatocellular carcinoma patients were with higher alpha feto-protein (AFP) above level >400 ng/ml and with ultrasound evi-dence of tumors lesions.
These patients were classified on the basis of total serum biliru-bin level, clinical examination, liver biochemistry, ultrasonogra-phy, and liver biopsy (Lok and McMahon, 2009; Kumar et al.,2008). Patients were admitted as and when necessary and wereevaluated in the respective Medical College, Orissa during thestudy. Samples coinfected with HIV or HCV were excluded fromstudy. Blood samples were collected, serum separated and storedin aliquots at �80 �C for further screening. Written informed con-sent was obtained from all patients or their legally responsibleguardians. The project was approved by the institutional ethicalcommittee.
2.2. Serological tests
Commercial ELISA kits were used for HBsAg (Biomerieux, Box-tel, The Netherlands), HBeAg/anti-HBe (Biomerieux, Boxtel, TheNetherlands and EQUIPAR, Soronno, Varese, Italy), anti-HCV(Ortho-Clinical Diagnostics, Raritan, NJ, USA) and anti- HIV (Bio-merieux) detection. ALT was measured by Ecoline ALAT kit (MerckSpecialties, Mumbai, India).
2.3. HBV DNA extraction, detection and quantification
HBV DNA was extracted from 200 ll serum using Qiagen DNAextraction Kit (Hilden, Germany). HBV DNA was detected by nestedPCR assay as previously described (Datta et al., 2006) and HBV-DNA was quantified by real-time detection PCR assay as previouslydescribed (Chandra et al., 2007). Guidelines for avoiding false posi-tive results were followed strictly (Kwok and Higuchi, 1991). Thenucleotide sequences used in this study have been submitted tothe DDBJ/EMBL/GenBank, Accession Numbers: JN164555–JN164648.
2.4. Amplification of the basal core promoter/precore gene (BCP-pC)regions
HBV-DNA amplification of BCP and PC regions were done as de-scribed earlier (Biswas et al., 2011), followed by direct sequencing.Variants in the BCP and PC region were identified by comparingwith HBV full genome sequences belonging to different genotypes.
2.5. Sequence analysis
Amplified products were directly sequenced in forward direc-tions for both the genome region using an ABI 3130xl sequencerand commercial kit (Applied Biosystems, Foster City, CA, USA) asdescribed (Datta et al., 2009).
A total of 76 sequences of the complete HBV genome were im-ported from GenBank and aligned with CLUSTAL X. A protein con-sensus sequence of the MHR, amino acids 101–163, was createdwith all the reference sequences by considering the most frequentamino acid at each of the position of the reference sequences. Alsoa consensus sequences was created with taking all the genotypes(including HBV/A-HBV/H). HBV genotypes were confirmed by phy-logenetic analysis using MEGA software Version 4.1.
R. Panigrahi et al. / Infection, Genetics and Evolution 12 (2012) 1622–1629 1623

Author's personal copy
BLOSUM (http://tandem.bu.edu/cgi-bin/mutationmaster/mut-mast.exe) scores for S gene analysis were used to define substitu-tions associated with altered antigenecity of HBsAg. The use ofBLOSUM score also helped us to identify the various substitutionsthat might be associated with diagnostic and/or vaccine and/orimmunoprophylaxis escape. Each amino acid position of clinicalsequences was compared to the consensus sequence with theMutation Master software. Our study includes previously reportedsubstitutions which are related with diagnostic or immune escapehaving BLOSUM scores6�3 (Roque-Afonso et al., 2007). This cutoffis selected because this score relates with G145R substitution, awell known immune escape substitution. This program allowsthe position and frequency of specific changes to be visualizedand enables variable and conserved positions to be identified start-ing from the multiple alignment data file of the 76 clinical se-quences. Mutation Master also allows the graphical presentationof BLOSUM scores (Blocks Matrix Substitution), reflecting changesin the chemical and structural properties of the amino acids(Henikoff and Henikoff, 1992). The BLOSUM score plot illustrateshow Mutation Master identifies each mutation and provides infor-mation about its conservative or nonconservative nature. The ma-jor substitutions at each position are scored according to theappropriate BLOSUM table. Positive numbers indicate a conserva-tive substitution, and negative numbers a nonconservativesubstitution.
2.6. Statistical analysis
Age, alanine aminotransferase (ALT) and HBV-DNA level wereexpressed as mean ± standard deviation. Student’s t-test was usedto compare continuous variables. The chi-squared test with Fishersexact test was used to compare categorical data. For all tests, P-va-lue <0.05 were considered significant.
3. Results
3.1. Demographic, biochemical and virological characteristics
The characteristic of 152 patients was explained in Table 1. Themean age is significantly higher in symptomatic patients than ininactive carrier’s cases (p < 0.05). Of them, 69 were found to beHBeAg positive and 83 were HBeAg negative. HBeAg positive pa-tients were older than HBeAg negative patients with no significantrelation (39.6 ± 15.6, vs. 35.6 ± 11.8, p = 0.14). The ALT level wassignificantly lower in inactive carriers group than chronic liver dis-ease and cirrhosis group (p < 0.05). Higher ALT was observedamong chronic liver disease patients in comparison with cirrhosispatients although statistically not significant (p = 0.64).
3.2. Predominance of HBV genotype D among the study samples
A total of 139 samples sequenced and analyzed for genotypingand mutational profiling in the study group. Of the total 139 sub-jects studied, 24 (17.2%), 4 (2.8%) and 109 (78.4%) were infectedwith HBV genotypes HBV/A, HBV/C and HBV/D, respectively (Ta-ble 1). Only one subgenotype was prevalent in HBV/A (A1), HBV/C (C1). Analysis of 109 HBV/D infected patients, showed four sub-genotypes where, HBV/D1 (4/109, 3.6%), HBV/D2 (11/109, 10.1%),HBV/D3 (22/109, 20.1%) and HBV/D5 (72/109, 66.0%) were identi-fied. Two samples were found to be C/D recombinant, most closelyrelated in terms of evolutionary distance to strains isolated fromchina (Accession Number AB675681 and AB675680). All HBV/D5samples were also confirmed by core gene analysis (data notshown). Fig. 1 showed a significant differential distribution of all
subgenotypes (A1, C1, D1, D2, D3, D5) with regard to the clinicalcourse of infection (p = 0.01).
Phylogenetic analysis showed different genotypes/subgenotypecirculating in the study population (Fig. 2). Notably, the presence ofHBV subgenotype D5 increased significantly with LC and HCCgroup compared with other groups (p < 0.05).
Considering the patients infected with HBV/D genotype a higherproportion of subgenotype HBV/D5 was found among the HCC (14/14; 100.0%) group, compared to LC (18/23; 78.2%), CLD (22/30;73.3%;) and IC (15/36; 41.6%) group. Among the acute patientsHBV/D5 and HBV/D3 were equally distributed (50.0%). The numberof CLD patients infected with HBV/D5 was significantly higher thanHBV/D3 (p < 0.05) and HBV/D1 (p < 0.05) infected ones.
3.3. Polymorphism at amino acid level in S region
Amino acid sequence of part of the S protein (amino acids 64–163) for various HBV isolates was compared. For comparison, allthe Indian HBV isolates sequenced, as well as isolates representingdifferent countries were used.
During S gene analysis by mutation master software positionswith BLOSUM scores 6�3 were highlighted in the Fig. 3; the upperpart of the figure represents the consensus amino acid sequence ofthe MHR and variation found in different genotypes (A–G). Thelower part represents the substitutions observed in the differentreference strains and clinical sequences from the three groups.Substitutions highlighted with red squares correspond to substitu-tions associated with diagnostic and/or vaccine and/or immuno-prophylaxis escape or to substitutions with BLOSUM scores 6�3,or to both. Numbers associated with amino acid symbols corre-spond to the number of occurrences of a given amino acid when>1. The most frequent substitutions among the studied sampleswere, L109P/Q, P111L, T116N (1.4%), Q101R, G112E/R, (2.2%) andF134N/S (23.6%), S/T114P/T (18.5%), S/T143L (27.4%).
The amino acids representing the MHR region were assessed forthe comparison of the nature of ancestral state to their derivedstate. When all the residues of the corresponding clinical groupswere analyzed it was observed that maximum of the amino acidsubstitutions show a change from hydrophobic to hydrophilicgroup (S114P, Y134H, L104S, L109Q, T126I, S143L, W156R,A159E). Amino acids substitutions from uncharged group tocharged group were also found (Q129R, Q101R/H, G112E, G112R,and G130R).
A total of 92 amino acid residues were found to have differentsubstitutions in different clinical categories. Compared to IC groupthe prevalence of amino acid substitutions showing change fromhydrophobic to hydrophilic and uncharged to charged group(L104S, T118V, F134N, W161R, Q101R, G130R, N146H), prevalenceof foresaid change were higher among CLD patients (Q101H,S114P, F134S, F134N, A159E, G159A, F161Y), LC and HCC patients(L101Q, C101R, G112R, Q129R, S143M, T126I, P127T, C149Y,C107R, W156R, F161Y).
Analysis of partial S gene of HBV revealed that about 30.9% ofthe study samples showed variation in the MHR region. It was ob-served from this study that the MHR variants analysed were foundto be in an increasing order from IC (28/50; 56.0%) to CLD (24/40;60.4%) to LC (21/29; 72.4%) to HCC (19/20; 95.0%). T125M was themost frequent in both inactive carriers and symptomatic patients.In our study group S143L/M was most prevalent.
Analyzing the sequence similarity identity matrix of surfacegene for HBV/D5 compared with reference sequences from Japan(AB033558), Paharia (GQ205379–85) and Eastern India(DQ315779–80) was analyzed using BioEdit v 7.0.9. The similaritypercentage observed with all the reference sequences were 98.7%with Japan, 98.3% with Eastern India and 98.1% with Paharia group.
1624 R. Panigrahi et al. / Infection, Genetics and Evolution 12 (2012) 1622–1629
Author's personal copy
3.4. Mutations in the TA rich BCP and PC region
Only 87 samples could be sequenced and analyzed for BCP re-gion of which 60/87; 68.9% belongs to HBV/D genotype. Eight sam-ples were not analyzed as for poor sequencing but used in BCPcalculation.
Among the HBV/D5 infected patients prevalence of T1762/A1764 (22/27; 81.4%) and C1753/T1762/A1764 (12/16, 75.0%)was found to be significantly higher in our study than other subge-notypes of HBV/D (p < 0.05). Also, T1762/A1764/C1753 mutationwas significantly higher in LC and HCC group compared to CLDgroup (p < 0.05). A higher viral load was found among the patientswith triple mutation than the patients with wild type (Table 1).
T1862 was found among 24 (27.5%) of 87 samples analyzed forBCP/PC region with a majority of HBV/A genotype. Our study sam-ples presented a specific pattern of tetra mutations T1809, T1812,and A1888 along with T1862 in 22 (91.6%) of 24 patients. The PCstop codon mutation, A1896 (24/87; 27.5%), was seen to be associ-ated with HBV/D5 (15/24; 62.5%).
3.5. HBV viral load
HBV-DNA viral load was quantified in 94 samples (Table 1).With a statistically insignificant relation the average HBV-DNAwas found to be higher in chronic liver disease (p = 0.3) and cirrho-sis patients (p = 0.6) in comparison with inactive carriers carriercases. Difference in viral load was considerably higher amongHBeAg positive group, but the difference was statistically insignif-icant (p = 0.2) compared to HBeAg negative group. A higher viralload was found among patients infected with HBV/D5 of LC groupcompared to others.
4. Discussion
Accumulating evidences suggest the association of HBV geno-types and subgenotypes with disease progression (Sumi et al.,2003; Wang et al., 2007). India is a vast country with a history ofpopulation migration from different countries resulting in variableethnicity and different patterns of chronic diseases in differentparts of the country (Sen et al., 2002). HBV genotype distributionand disease association has been reported to vary in different partsof India (Datta et al., 2008). In the present study HBV/D is the majorgenotype found among the HBV infected patients of Orissa, a statein southeastern India. Reports from northern India reveal thatHBV/A is associated with more severe liver disease than HBV/D(Kumar et al., 2005), contradicted by another study (Chattopadhy-ay et al., 2006). On the other hand, reports from western andSouthern India showed that HBV/D did not influence the clinicaloutcome of the HBV infected patients (Gandhe et al., 2003a,b;Vivekanandan et al., 2004). Previous reports from Kolkata, EasternIndia revealed that genotype HBV/A and HBV/C were associatedwith chronic liver disease, while genotype D was common amongasymptomatic carriers (Datta et al., 2008; Biswas et al., 2011). Fur-thermore, studies among the subgenotypes prevalent among thepredominant genotype D in eastern India suggested that there isdifferential association of disease prevalence among the subgeno-types of HBV/D. Subgenotype D5 was recently identified fromKolkata, Eastern India with 20% prevalence among HBV/D infectedpatients (Chandra et al., 2009). Later, a study among the tribal’s of aneighboring state of Jharkhand, showed HBV/D5 to be prevalentamong the Paharia tribal’s and reported HBV/D5 to be the most an-cient of the HBV/D subgenotypes. The present study, conductedfrom Orissa (Fig. 4), a state that borders both of these eastern In-dian states showed that HBV/D5 with a prevalence of about 66%
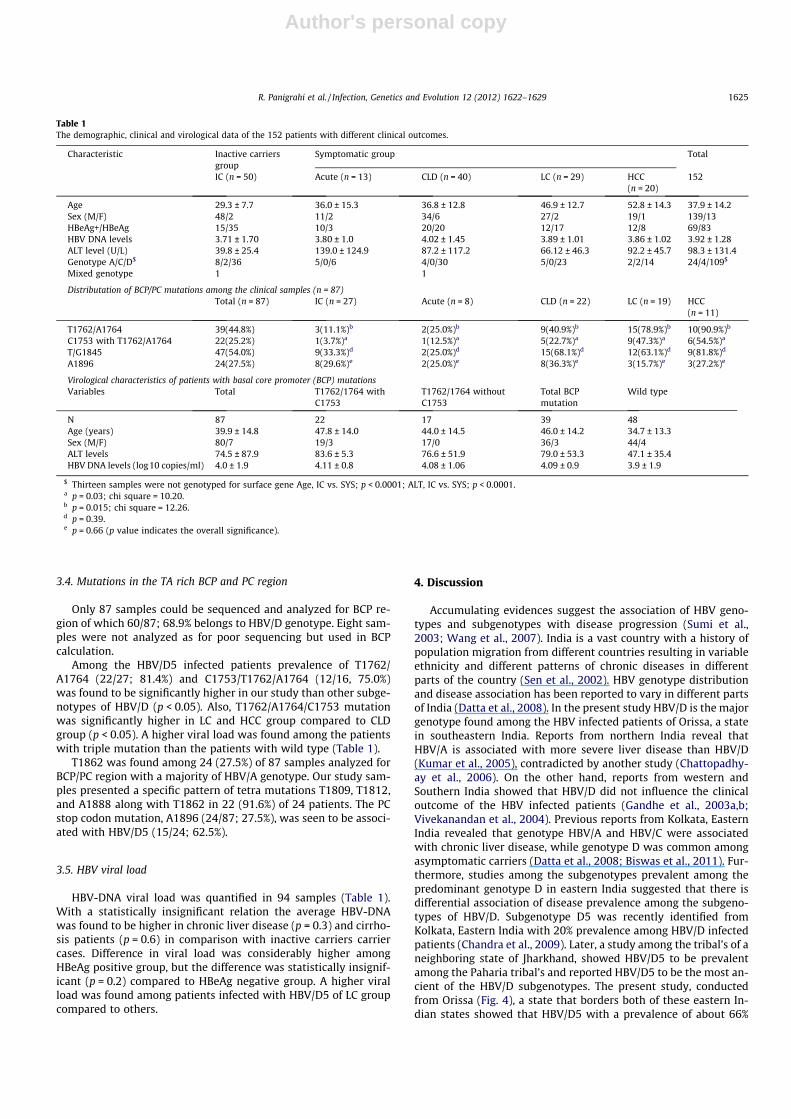
Table 1The demographic, clinical and virological data of the 152 patients with different clinical outcomes.
Characteristic Inactive carriersgroup
Symptomatic group Total
IC (n = 50) Acute (n = 13) CLD (n = 40) LC (n = 29) HCC(n = 20)
152
Age 29.3 ± 7.7 36.0 ± 15.3 36.8 ± 12.8 46.9 ± 12.7 52.8 ± 14.3 37.9 ± 14.2Sex (M/F) 48/2 11/2 34/6 27/2 19/1 139/13HBeAg+/HBeAg� 15/35 10/3 20/20 12/17 12/8 69/83HBV DNA levels 3.71 ± 1.70 3.80 ± 1.0 4.02 ± 1.45 3.89 ± 1.01 3.86 ± 1.02 3.92 ± 1.28ALT level (U/L) 39.8 ± 25.4 139.0 ± 124.9 87.2 ± 117.2 66.12 ± 46.3 92.2 ± 45.7 98.3 ± 131.4Genotype A/C/D$ 8/2/36 5/0/6 4/0/30 5/0/23 2/2/14 24/4/109$
Mixed genotype 1 1
Distributation of BCP/PC mutations among the clinical samples (n = 87)Total (n = 87) IC (n = 27) Acute (n = 8) CLD (n = 22) LC (n = 19) HCC
(n = 11)
T1762/A1764 39(44.8%) 3(11.1%)b 2(25.0%)b 9(40.9%)b 15(78.9%)b 10(90.9%)b
C1753 with T1762/A1764 22(25.2%) 1(3.7%)a 1(12.5%)a 5(22.7%)a 9(47.3%)a 6(54.5%)a
T/G1845 47(54.0%) 9(33.3%)d 2(25.0%)d 15(68.1%)d 12(63.1%)d 9(81.8%)d
A1896 24(27.5%) 8(29.6%)e 2(25.0%)e 8(36.3%)e 3(15.7%)e 3(27.2%)e
Virological characteristics of patients with basal core promoter (BCP) mutationsVariables Total T1762/1764 with
C1753T1762/1764 withoutC1753
Total BCPmutation
Wild type
N 87 22 17 39 48Age (years) 39.9 ± 14.8 47.8 ± 14.0 44.0 ± 14.5 46.0 ± 14.2 34.7 ± 13.3Sex (M/F) 80/7 19/3 17/0 36/3 44/4ALT levels 74.5 ± 87.9 83.6 ± 5.3 76.6 ± 51.9 79.0 ± 53.3 47.1 ± 35.4HBV DNA levels (log10 copies/ml) 4.0 ± 1.9 4.11 ± 0.8 4.08 ± 1.06 4.09 ± 0.9 3.9 ± 1.9
$ Thirteen samples were not genotyped for surface gene Age, IC vs. SYS; p < 0.0001; ALT, IC vs. SYS; p < 0.0001.a p = 0.03; chi square = 10.20.b p = 0.015; chi square = 12.26.d p = 0.39.e p = 0.66 (p value indicates the overall significance).
R. Panigrahi et al. / Infection, Genetics and Evolution 12 (2012) 1622–1629 1625

Author's personal copy
among HBV infected patients, was found to be associated with ad-vanced liver disease, particularly with HCC.
A recent study from Kolkata, Eastern India described significantassociation of HBV/D1 with chronic liver disease, while HBV/D5was associated with higher T1762/A1764 mutation and higher vir-al load, but no significant clinical correlation was found (Chandraet al., 2007). Interestingly, in Orissa, frequency of HBV/D1 is lowand HBV/D5 was associated with severity of liver disease, elevatedALT and higher viral load. On the other hand among the inactivecarriers of Paharia tribes of Jharkhand, HBV/D5 was associatedwith wild type BCP and low ALT level.
Previous studies have shown that T1762/A1764 double muta-tions (Orito et al., 2001) are associated with progressive liver dis-ease and may even predict the development of HCC (Huang andLok, 2011; Yang et al., 2008). Biswas et al. from eastern Indiashowed association of triple mutation (C1753/T1762/A1764) in
Fig. 1. Distributation of overall subgenotypes of HBV among the clinical groups.
Fig. 2. The phylogenetic tree of HBV based on the partial S gene sequences of 139 isolates, from southeastern India (marked with solid shapes). A phylogenetic treeconstructed using NJ method, based on partial S gene sequences retrieved from the DDBJ/EMBL/GenBank. The number represents the percentages of bootstrap replicates (of1000 totals) for the node. The capital letters A to H designate HBV genotypes and the subgenotypes for each genotype were also included during the construction of the tree.Reference sequences are denoted by their accession number.
1626 R. Panigrahi et al. / Infection, Genetics and Evolution 12 (2012) 1622–1629

Author's personal copy
BCP region with advance liver disease irrespective of HBV geno-types or subgenotypes. These mutations alone and in combinationmay be predictive for hepatocarcinogenesis (Liu et al., 2009). Takentogether the difference in disease profile among the Paharia andthe patients from Orissa may be related to the prevalence of wildtype and mutant BCP. In the present study, the T1762/A1764 muta-tion occurred frequently in patients with CLD and cirrhosis than ICcases (Table 1). Moreover, the prevalence of triple mutation(C1753/T1762/A1764) was significantly higher among the LC andHCC group than the CLD group.
It is also clear from Table 1 that the triple mutation results sig-nificantly higher ALT and HBV-DNA load. Hence considering theassociation of BCP region mutations with advanced liver diseaseson one hand and with genotypes HBV/D5 on the other, the presentresults clearly support that genotype HBV/D with HBV/D5 subge-notype are associated more frequently with progressive liver dis-ease in southeastern Indian population. Interestingly, studiesfrom western India showed that HBV/D with C1753 mutationwas rare (Gandhe et al., 2003a).
In this study, 91.6% of the patients with T1862 mutation was of-ten associated with other mutations, that is, T1809, T1812, and
A1888 and was detected predominantly in HBeAg �ve patients.A study from South Africa has reported the presence of this muta-tion in 17% HBeAg-positive and 36% HBeAg-negative HBV/A pa-tients (Kramvis et al., 1998). These mutations were well studiedfrom north India also (Chauhan et al., 2006). In our study the pres-ence of this mutation was seen among 63.6% of HBeAg negativeisolates, however, maximum of them are in IC group. The low prev-alence of precore A1896 mutation among the southeastern Indianpatients is similar to the reports from eastern India (Banerjee et al.,2005), with no apparent clinical correlation.
Previous studies have shown that patients with advancing ageand end-stage liver disease exhibited higher S gene variability thanpatients with acute self-limited hepatitis (Song et al., 2005). Ourstudy finding shows an increasing trend in the variability of MHRregion and ‘‘a’’ determinant region also with increasing diseaseseverity. The presence of G145R was found only in one sample,suggesting that this well known mutation is rare among HBV in-fected patients in this study population. The BLOSUM scoreshelped to identify most commonly occurring mutations at each po-sition (Walewski et al., 2002). Interestingly, the present study re-vealed the presence of several substitutions with BLOSUM score
Fig. 3. Representing BLOSUM scores of sequence variation in major hydrophilic region (MHR) region. The upper part of the figure represents the consensus amino acidsequence of the major hydrophilic region (MHR) and variation found in different genotypes (A–G). The lower part represents the substitutions observed in the differentreference strains and clinical sequences from the three groups. Substitutions highlighted with red squares correspond to substitutions associated with diagnostic and/orvaccine and/or immunoprophylaxis escape or to substitutions with BLOSUM scores 6�3, or to both. Numbers associated with amino acid symbols correspond to the numberof occurrences of a given amino acid when >1. The frequency of variation is given as a percentage. The positions with highest degrees of polymorphism were 118, 121, 125and 127 for the 76 reference strains and 110, 114, 116, 125, 127 and 159 for the 152 clinical sequences. (For interpretation of the references to color in this figure legend, thereader is referred to the web version of this article.)
R. Panigrahi et al. / Infection, Genetics and Evolution 12 (2012) 1622–1629 1627

Author's personal copy
6�3. It distinguishes positions with many different amino acidsfrom those with a high frequency of one or two substitutions. Thisis very useful because a position with only one or two naturallyoccurring substitutions with high frequency may be may be sub-ject to more constraints than positions with a higher variabilityAlthough the association of S gene variability with disease severitycan only be partially explained with the help of BLOSUM score plotbut it was possible to highlight that the frequency of mutationswas related with advanced liver disease. The substitution of aminoacids in the MHR region might alter the 3D structure of the anti-genic determinant region and thus might play a role in immuneescape.
In conclusion, we established that the predominant HBV subge-notype HBV/D5 with high viral load and BCP mutations was signif-icantly associated with advanced liver disease (LC and HCC) andmight act as a predictor of severe hepatic complications. Howeverthese results should be confirmed by further prospective studies.Comparison of HBV/D5 from different geographical region suggeststhat the ultimate clinical manifestation associated with a particu-lar HBV genotype even at subgenotype level may differ geograph-ically due to the pattern of mutations. Therefore, prior toapplication of HBV genetic information in disease prognosis andmanagement, studies among local population are necessary. Inaddition, the study underscores the increasing trend of S genemutation and their association with severity of liver disease, whichrequires a further analysis in larger scale.
Acknowledgments
The authors thank Tapan Chakraborty, for their excellent tech-nical assistance as well as the doctors and technicians of M.K.C.Gmedical college and SCB medical college for their help in blood col-lection. This research was supported by Grants from the IndianCouncil of Medical Research (ICMR), New Delhi, India, from whichRajesh Panigrahi got senior research fellowship.
References
Banerjee, A., Banerjee, S., Chowdhury, A., Santra, A., Chowdhury, S., Roychowdhury,S., Panda, C.K., Bhattacharya, S.K., Chakravarty, R., 2005. Nucleic acid sequenceanalysis of BCP/precore/core region of hepatitis B virus isolated from chroniccarriers of the virus from Kolkata, Eastern India: low frequency of mutation inthe precore region. Intervirology 48, 389–399.
Biswas, A., Banerjee, A., Chandra, P.K., Datta, S., Panigrahi, R., Dutta, D., De, B.K., Pal,M., Guha, S.K., Chakrabarti, S., Chakravarty, R., 2011. Variations in the functionaldomain of basal core promoter of hepatitis B virus among Eastern Indianpatients with prevalence of genotypes A, C, and D among the same ethnicpopulation. J. Med. Virol. 83 (2), 253–260.
Chauhan, R., Kazim, S.N., Bhattacharjee, J., Sakhuja, P., Sarin, S.K., 2006. Basal corepromoter, precore region mutations of HBV and their association with eantigen, genotype, and severity of liver disease in patients with chronichepatitis B in India. J. Med. Virol. 78, 1047–1054.
Chandra, P.K., Banerjee, A., Datta, S., Chakravarty, R., 2007. G1862T mutation amongHepatitis B virus infected individuals: association with viral genotypes anddisease outcome in Kolkata, Eastern India. Intervirology 50, 173–180.
Chandra, P.K., Biswas, A., Datta, S., Banerjee, A., Panigrahi, R., Chakrabarti, S., De, B.K.,Chakravarty, R., 2009. Subgenotypes of hepatitis B virus genotype D (D1, D2, D3
Fig. 4. Map of India showing the neighboring states with which Orissa shares the borders. The region of Paharia population and Delhi was also shown in the map.
1628 R. Panigrahi et al. / Infection, Genetics and Evolution 12 (2012) 1622–1629

Author's personal copy
and D5) in India: differential pattern of mutations, liver injury and occult HBVinfection. J. Viral Hepat. 16 (10), 749–756.
Chattopadhyay, S., Das, B.C., Kar, P., 2006. Hepatitis B virus genotypes in chronicliver disease patients from New Delhi, India. World J. Gastroenterol. 12 (41),6702–6706.
Datta, S., Banerjee, A., Chandra, P.K., Chakravarty, R., 2007. Pin1-HBx interaction: astep towards understanding the significance of HBV genotypes inhepatocarcinogenesis. Gastroenterology 133, 727–728.
Datta, S., Banerjee, A., Chandra, P.K., Chowdhury, A., Chakravarty, R., 2006.Genotype, phylogenetic analysis, and transmission pattern of occult hepatitisB virus (HBV) infection in families of asymptomatic HBsAg carriers. J. Med.Virol. 78, 53–59.
Datta, S., Panigrahi, R., Biswas, A., Chandra, P.K., Banerjee, A., Mahapatra, P.K., Panda,C.K., Chakrabarti, S., Bhattacharya, S.K., Biswas, K., Chakravarty, R., 2009. Geneticcharacterization of hepatitis B virus in peripheral blood leukocytes: evidence forselection and compartmentalization of viral variants with the immune escapeG145R mutation. J. Virol. 83 (19), 9983–9992.
Datta, S., Biswas, A., Chandra, P.K., Banerjee, A., Panigrahi, R., Mahapatra, P.K.,Chakrabarti, S., Panda, C.K., Chakravarty, R., 2008. Molecular epidemiology andclinical significance of hepatitis B virus genotypes, core promoter and precoremutation in eastern India. Intervirology 51, 275–284.
Francois, G., Kew, M., Van Damme, P., Mphahlele, M.J., Meheus, A., 2001. Mutanthepatitis B viruses: a matter of academic interest only or a problem with far-reaching implications? Vaccine 19, 3799–3815.
Gandhe, S.S., Chadha, M.S., Arankalle, V.A., 2003a. Hepatitis B virus genotypes andserotypes in western India: lack of clinical significance. J. Med. Virol. 69, 324–330.
Gandhe, S.S., Chadha, M.S., Walimbe, A.M., Arankalle, V.A., 2003b. Hepatitis B virus:prevalence of precore/core promoter mutations in different clinical categoriesin Indian patients. J. Viral Hepat. 10, 367–382.
Henikoff, S., Henikoff, J.G., 1992. Amino acid substitution matrices from proteinblocks. Proc. Natl. Acad. Sci. USA 89, 10915–10919.
Huang, Y., Lok, A.S., 2011. Viral factors and outcomes of chronic HBV infection. Am.J. Gastroenterol. 106 (1), 93–95.
Hunt, C.M., McGill, J.M., Allen, M.I., Condreay, L.D., 2000. Clinical relevance ofhepatitis B viral mutations. Hepatology 31 (5), 1037–1044.
Kidd-Ljunggren, K., Oberg, M., Kidd, A.H., 1997. Hepatitis B virus X gene 1751 to1764 mutations: implications for HBeAg status and disease. J. Gen. Virol. 78,1469–1478.
Kramvis, A., Kew, M., Francois, G., 2005. Hepatitis B virus genotypes. Vaccine 23,2409–2423.
Kramvis, A., Kew, M.C., Bukofzer, S., 1998. Hepatitis B virus precore mutants inserum and liver of Southern African Blacks with hepatocellular carcinoma. J.Hepatol. 28, 132–141.
Kumar, R., Saraswat, M.K., Sharma, B.C., Sakhuja, P., Sarin, S.K., 2008. Characteristicsof hepatocellular carcinoma in India: a retrospective analysis of 191 cases. QJM101 (6), 479–485.
Kumar, A., Kumar, S.I., Pandey, R., Naik, S., Aggarwal, R., 2005. Hepatitis B virusgenotype A is more often associated with severe liver disease in northern Indiathan is genotype D. Ind. J. Gasroenterol. 24, 19–22.
Kwok, S., Higuchi, R., 1991. Avoiding false positives with PCR. Nature 8 (339),237.
Lindh, M., Andersson, A.S., Gusdal, A., 1997. Genotypes, nt 1858 variants, andgeographic origin of hepatitis B virus – large-scale analysis using a newgenotyping method. J. Infect. Dis. 175, 1285–1293.
Li, J.S., Tong, S.P., Wen, Y.M., Vitvitski, L., Zhang, Q., Trepo, C., 1993. Hepatitis B virusgenotype A rarely circulates as an HBe-minus mutant: possible contribution of asingle nucleotide in the precore region. J. Virol. 67, 5402–5410.
Liu, S., Zhang, H., Gu, C., Yin, J., He, Y., Xie, J., Cao, G., 2009. Associations betweenhepatitis B virus mutations and the risk of hepatocellular carcinoma: a meta-analysis. J. Natl. Cancer Inst. 101 (15), 1066–1082.
Lok, A.S., McMahon, B.J., 2001. Practice Guidelines Committee, AmericanAssociation for the Study of Liver Diseases. Chronic hepatitis B. Hepatology34, 1225–1241.
Lok, A.S., Akarca, U., Greene, S., 1994. Mutations in the pre-core region of hepatitis Bvirus serve to enhance the stability of the secondary structure of the pre-genome encapsidation signal. Proc. Natl. Acad. Sci. USA 91, 4077–4081.
Lok, A.S., McMahon, B.J., 2009. Chronic hepatitis B: update. Hepatology 50 (3), 661–662.
Miyakawa, Y., Mizokami, M., 2003. Classifying hepatitis B virus genotypes.Intervirology 46, 329–338.
Norder, H., Couroucé, A.M., Coursaget, P., Echevarria, J.M., Lee, S.D., Mushahwar, I.K.,Robertson, B.H., Locarnini, S., Magnius, L.O., 2004. Genetic diversity of hepatitisB virus strains derived worldwide: genotypes, subgenotypes, and HBsAgsubtypes. Intervirology 47 (6), 289–309.
Orito, E., Ichida, T., Sakugawa, H., Sata, M., Horiike, N., Hino, K., Okita, K., Okanoue, T.,Iino, S., Tanaka, E., Suzuki, K., Watanabe, H., Hige, S., Mizokami, M., 2001.Geographical distribution of hepatitis B virus (HBV) genotype in patients withchronic HBV infection in Japan. Hepatology 34, 590–594.
Ren, X., Xu, Z., Liu, Y., Li, X., Bai, S., Ding, N., Zhong, Y., Wang, L., Mao, P., Zoulim, F.,Xu, D., 2010. Hepatitis B virus genotype and basal core promoter/precoremutations are associated with hepatitis B-related acute-on-chronic liver failurewithout pre-existing liver cirrhosis. J. Viral Hepat. 17 (12), 887–895.
Roque-Afonso, A.M., Férey, M.P., Ly, T.D., Graube, A., Costa-Faria, L., Samuel, D.,Dussaix, E., 2007. Viral and clinical factors associated with surface gene variantsamong hepatitis B virus carriers. Antiviral Ther. 12 (8), 1255–1263.
Sen, U., Sankaranarayanan, R., Mandal, S., Ramanakumar, A.V., Parkin, D.M., Siddiqi,M., 2002. Cancer patterns in eastern India: the first report of the Kolkata Cancerregistry. Int. J. Cancer 100, 86–91.
Song, B.C., Kim, S.H., Kim, H., Ying, Y.H., Kim, H.J., Kim, Y.J., Yoon, J.H., Lee, H.S., Cha,C.Y., Kook, Y.H., Kim, B.J., 2005. Prevalence of naturally occurring surfaceantigen variants of hepatitis B virus in Korean patients infected chronically. J.Med. Virol. 76, 194–202.
Sumi, H., Yokosuka, O., Seki, N., Arai, M., Imazeki, F., Kurihara, T., Kanda, T., Fukai, K.,Kato, M., Saisho, H., 2003. Influence of hepatitis B virus genotypes on theprogression of chronic type b liver disease. Hepatology 37, 19–26.
Tanaka, Y., Hasegawa, I., Kato, T., Orito, E., Hirashima, N., Acharya, S.K., Gish, R.G.,Kramvis, A., Kew, M.C., Yoshihara, N., Shrestha, S.M., Khan, M., Miyakawa, Y.,Mizokami, M., 2004. A case–control study for differences among hepatitis Bvirus infections of genotypes A (subtypes Aa and Ae) and D. Hepatology 40,747–755.
Vivekanandan, P., Abraham, P., Sridharan, G., Chandy, G., Daniel, D., Raghuraman, S.,Daniel, H.D., Subramaniam, T., 2004. Distribution of hepatitis B virus genotypesin blood donors and chronically infected patients in a tertiary care hospital insouthern India. Clin. Infect. Dis. 38, e81–e86.
Wang, Z., Tanaka, Y., Huang, Y., Kurbanov, F., Chen, J., Zeng, G., Zhou, B., Mizokami,M., Hou, J., 2007. Clinical and virological characteristics of hepatitis B virussubgenotypes Ba, C1 and C2 in China. J. Clin. Microbiol. 45, 1491–1496.
Walewski, J.L., Gutierrez, J.A., Branch-Elliman, W., Stump, D.D., Keller, T.R.,Rodriguez, A., Benson, G., Branch, A.D., 2002. Mutation master: profiles ofsubstitutions in hepatitis C virus RNA of the core, alternate reading frame, andNs2 coding regions. RNA 8, 557–571.
Yang, H.I., Yeh, S.H., Chen, P.J., Iloeje, U.H., Jen, C.L., Su, J., Wang, L.Y., Lu, S.N., You,S.L., Chen, D.S., Liaw, Y.F., Chen, C.J.REVEAL-HBV Study Group, 2008.Associations between hepatitis B virus genotype and mutants and the risk ofhepatocellular carcinoma. J. Natl. Cancer Inst. 100, 1134–1143.
R. Panigrahi et al. / Infection, Genetics and Evolution 12 (2012) 1622–1629 1629




















