
Stochastic approach to determine spatial patterns of lizard community on a desert island
11
Original article Stochastic approach to determine spatial patterns of lizard community on a desert island Crystian Sadiel Venegas-Barrera 1 , Enrique Morales-Bojo ´rquez*, Gustavo Arnaud Centro de Investigaciones Biolo ´gicas del Noroeste (CIBNOR), Mar Bermejo 195, Col. Playa Palo de Santa Rita, La Paz, B.C.S. 23090, Mexico article info Article history: Received 23 May 2007 Accepted 30 November 2007 Published online 18 April 2008 Keywords: Spatial dependence Markov models Numerical dominance Species richness Spatial arrangement Nearby samples abstract One of the principal sources of error in identifying spatial arrangements is autocorrelation, since nearby points in space tend to have more similar values than would be expected by random change. When a Markovian approach is used, spatial arrangements can be mea- sured as a transition probability between occupied and empty spaces in samples that are spatially dependent. We applied a model that incorporates first-order Markov chains to an- alyse spatial arrangement of numerical dominance, richness, and abundance on a lizard community at different spatial and temporal scales. We hypothesized that if a spatial de- pendence on abundance and richness exists in a diurnal desert community, then the Mar- kov chains can predict the spatial arrangement. We found that each pair of values was dependent only on its immediate predecessor segment. In this sense, we found interge- neric differences at temporal and spatial scales of recurrence estimates. Also, in desert scrub, species show higher spatial aggregation and had lower species richness than at the island level; the inverse pattern occurred on rocky hillsides. At the species level, Uta stansburiana is the most abundant species in desert scrub, while Sauromalus slevini is the most abundant species on rocky hillsides. This report attempts to understand, using Mar- kovian spatial models, the effect of nearby samples on local abundance and richness on different scales and over several seasons. ª 2008 Elsevier Masson SAS. All rights reserved. 1. Introduction The spatial arrangement of several species in a community is not homogeneous because limiting factors that affect abun- dance and distribution change spatially and species tend to respond differentially to environmental heterogeneity. Gener- ally, species predominate in habitats where conditions are suitable, and are rare in unfavourable habitats (Kneitel and Chase, 2004). Therefore, spatial elements play a funda- mental role in most ecological processes, including spatial segregation, habitat selection, and territoriality (Legendre, 1993; Bevers and Flather, 1999). Patterns based on spatial ar- rangements are a first approximation for analysing the effect of biotic, that is, inter- or intraspecific interactions and abiotic environmental variations, such as, temperature, pH, topogra- phy, and soil on species. One of the principal sources of error in identifying spatial arrangements is autocorrelation, since nearby points tend to have more similar values than would be expected by random change (Lichstein et al., 2002). When a Markovian approach is used, spatial arrangements can be * Corresponding author. Tel.: þ52 612 123 8484x3351; fax: þ52 612 125 3625. E-mail address: [email protected] (E. Morales-Bojo ´ rquez). 1 Present address: Laboratorio de Ecologı´a Evolutiva, Centro de Investigaciones en Recursos Bio ´ ticos, Universidad Auto ´ noma del Estado de Me ´ xico, Instituto Literario 100, 50000, Toluca, Mexico. available at www.sciencedirect.com journal homepage: www.elsevier.com/locate/actoec 1146-609X/$ – see front matter ª 2008 Elsevier Masson SAS. All rights reserved. doi:10.1016/j.actao.2008.01.002 acta oecologica 33 (2008) 280–290
-
Upload
itvictoria -
Category
Documents
-
view
2 -
download
0
Transcript of Stochastic approach to determine spatial patterns of lizard community on a desert island

a c t a o e c o l o g i c a 3 3 ( 2 0 0 8 ) 2 8 0 – 2 9 0
ava i lab le a t www.sc iencedi rec t .com
journa l homepage : www.e lsev ie r . com/ loca te /ac toec
Original article
Stochastic approach to determine spatial patternsof lizard community on a desert island
Crystian Sadiel Venegas-Barrera1, Enrique Morales-Bojorquez*, Gustavo Arnaud
Centro de Investigaciones Biologicas del Noroeste (CIBNOR), Mar Bermejo 195, Col. Playa Palo de Santa Rita, La Paz, B.C.S. 23090, Mexico
a r t i c l e i n f o
Article history:
Received 23 May 2007
Accepted 30 November 2007
Published online 18 April 2008
Keywords:
Spatial dependence
Markov models
Numerical dominance
Species richness
Spatial arrangement
Nearby samples
* Corresponding author. Tel.: þ52 612 123 84E-mail address: [email protected] (E.
1 Present address: Laboratorio de Ecologıa Ede Mexico, Instituto Literario 100, 50000, Tol1146-609X/$ – see front matter ª 2008 Elsevidoi:10.1016/j.actao.2008.01.002
a b s t r a c t
One of the principal sources of error in identifying spatial arrangements is autocorrelation,
since nearby points in space tend to have more similar values than would be expected by
random change. When a Markovian approach is used, spatial arrangements can be mea-
sured as a transition probability between occupied and empty spaces in samples that are
spatially dependent. We applied a model that incorporates first-order Markov chains to an-
alyse spatial arrangement of numerical dominance, richness, and abundance on a lizard
community at different spatial and temporal scales. We hypothesized that if a spatial de-
pendence on abundance and richness exists in a diurnal desert community, then the Mar-
kov chains can predict the spatial arrangement. We found that each pair of values was
dependent only on its immediate predecessor segment. In this sense, we found interge-
neric differences at temporal and spatial scales of recurrence estimates. Also, in desert
scrub, species show higher spatial aggregation and had lower species richness than at
the island level; the inverse pattern occurred on rocky hillsides. At the species level, Uta
stansburiana is the most abundant species in desert scrub, while Sauromalus slevini is the
most abundant species on rocky hillsides. This report attempts to understand, using Mar-
kovian spatial models, the effect of nearby samples on local abundance and richness on
different scales and over several seasons.
ª 2008 Elsevier Masson SAS. All rights reserved.
1. Introduction segregation, habitat selection, and territoriality (Legendre,
The spatial arrangement of several species in a community is
not homogeneous because limiting factors that affect abun-
dance and distribution change spatially and species tend to
respond differentially to environmental heterogeneity. Gener-
ally, species predominate in habitats where conditions are
suitable, and are rare in unfavourable habitats (Kneitel
and Chase, 2004). Therefore, spatial elements play a funda-
mental role in most ecological processes, including spatial
84x3351; fax: þ52 612 125Morales-Bojorquez).volutiva, Centro de Invesuca, Mexico.er Masson SAS. All rights
1993; Bevers and Flather, 1999). Patterns based on spatial ar-
rangements are a first approximation for analysing the effect
of biotic, that is, inter- or intraspecific interactions and abiotic
environmental variations, such as, temperature, pH, topogra-
phy, and soil on species. One of the principal sources of error
in identifying spatial arrangements is autocorrelation, since
nearby points tend to have more similar values than would
be expected by random change (Lichstein et al., 2002). When
a Markovian approach is used, spatial arrangements can be
3625.
tigaciones en Recursos Bioticos, Universidad Autonoma del Estado
reserved.

a c t a o e c o l o g i c a 3 3 ( 2 0 0 8 ) 2 8 0 – 2 9 0 281
measured as a transition probability between occupied and
empty spaces in samples that are spatially dependent.
A first-order Markov chain is a stochastic model in which
the future development of a system is dependent on the pres-
ent state of the system and is independent of the way in which
that state has developed (Formacion and Saila, 1994).
In ecology, Markov chains have been used principally in
temporal succession of ecological states (Tanner et al., 1996;
Wootton, 2001a; Hill et al., 2004), time of recovery and restora-
tion of forests (Orloci and Orloci, 1988; Hall et al., 1991; Tucker
and Anand, 2004), patterns of change in parental stock and re-
cruitment in fisheries (Roshchild and Mullen, 1985), estimates
of bird populations (Wileyto et al., 1994), and anthropogenic
impact on marine mammals (Lusseau, 2003). Markov chains
spatial models have been used to predict sequences of egg-
laying in butterflies (Root and Kareiva, 1984), ontogenic
change in habitat preference of cotton rats (Kincaid and
Cameron, 1985), movements of Canada geese (Hestbeck
et al., 1991), and spatial inhibition by allelopathy in plants
(Kenkel, 1993). These studies highlight the potential of Markov
chains in the study of population dynamics and importance of
proximity neighbourhoods in the state of system. However,
rarely has it been used to explore spatial changes in richness,
abundance, or dominance of species at a community scale,
despite the utility of that approach to analyse spatial depen-
dence between near samples and calculate the average dis-
tance between ecological states.
The advantages of Markov chains are that: (1) such models
are relatively easy to derive from continuous data; (2) the
model does not require deep insight into the mechanisms of
dynamic change; (3) the basic transition matrix summarizes
essential parameters of dynamic change in a systems in
a way that few others models achieve; and (4) a model has
much potential for identifying recent history in dynamic com-
munities and population dynamics (Formacion and Saila,
1994). These characteristics are ideal for calculating spatial ar-
rangement of species. Additionally, Markov chains can be
used to estimate the probability of any state of abundance or
richness occur in the space, assuming that it is dependent
on the preceding area. We assumed that: (1) contagious bio-
logical process that affect local abundance of individual spe-
cies are spatially dependent (Legendre, 1993), i.e. conditions
of growth, survival, and reproduction tend to be similar at
nearby sites, (2) well-selected habitats provide high fitness po-
tential (Railsback et al., 2003), (3) individuals in a population do
not show a random distribution, i.e. occurrence of an individ-
ual does not affect the presence of others, and (4) species with
similar adaptations will tend to occur together at the same
sites (Bell, 2001). In this study, we developed models that in-
corporate first-order Markov chains to analyse spatial changes
in states of numerical dominance (more abundant species by
unit area), richness (number of species by unit area), and
abundance of one lizard community on Isla Coronados in
the Gulf of California. We chose diurnal lizard species on
Isla Coronados because they are conspicuous, abundant, and
represent one of the four islands in Gulf of California with
high richness (ten species). We hypothesized that, if a pattern
of abundance and richness exist in a desert community, then
Markov chains can predict the spatial arrangement. If we
knew the sequences of presence-occurrence, richness, and
abundance of species, then we can estimate the average dis-
tance before we again observe the same species (recurrence)
and rate of change between species without the necessity of
evaluating the spatial structure of habitats. We predicted
high probabilities of replacement between species with simi-
lar habitat requirements as species with different require-
ments. Also, we can compare the numerical dominance and
richness at two spatial scales (island and landscape) to ana-
lyse the effect of scale on the estimation of spatial arrange-
ment. Finally, we calculated the spatial arrangement for
more abundant species on two landscapes.
2. Materials and methods
2.1. Study area
Isla Coronados is a volcanic, land bridge island (26�0801500 N,
111�160500 W) located w10 km northeast of Loreto, B.C.S.,
Mexico and w3 km from the closest shore of the Baja Califor-
nia Peninsula (Fig. 1). The climate is hot and arid in summer
(mean July temperature 33 �C) and warm in winter (mean Jan-
uary temperature 16 �C). Summer precipitation comes from
convectional storms on the Peninsula (average 190 mm/year)
(Grismer, 1994). Four types of habitats occur on Isla Coronado;
rocky hillsides, desert scrubland, a coastal zone, and a transi-
tion zone between rocky hillsides and desert scrub (Venegas,
2003). On rocky hillsides, approximately 45% of the surface is
covered with rocks (ranging in diameter from 20 to 100 cm),
15% is bare soil (includes fallen leaves, soil, and gravel
<2 cm), and 40% is covered with vegetation (Jatropha cuneata,
Euphorbia magdalenae, and Simmondsia chinensis). In desert
scrub, approximately 7% are rocks (<10 cm diameter), 38% is
bare soil, and 55% is covered with vegetation (principally Fou-
quieria diguetii, Gossypium harknessii, Hibiscus denudatus, and
Jatropha cuneata). The coastal zone has a ground surface that
is 5% rock (<10 cm diameter), 65% bare soil, and 30% covered
with vegetation (principally Salicornia virginica, Maytenus phyl-
lanthoides, and Atriplex barclayana). The transition habitat is
characterized by 30% rocks (5–50 cm diameter), 17% bare
soil, and 53% vegetation (Jatropha cuneata, Hibiscus denudatus,
Euphorbia magdalenae, and Lycium sp.).
2.2. Species
Isla Coronados is one of four islands in the Gulf of California
with a largest number of diurnal lizard species (Grismer,
2002). Of the ten species in the study area, four are ground-
dwelling species: desert iguana (Dipsosaurus dorsalis), zebra-
tailed lizard (Callisaurus draconoides), side-blotched lizard (Uta
stanburiana), and orange-throated whiptail (Aspidoscelis hyper-
ythra) and six are rock dwellers: slevin’s chuckwalla (Sauroma-
lus slevini), black-tailed brush lizard (Urosaurus nigricaudus),
central Baja California banded rock lizard (Petrosaurus repens),
granite spiny lizard (Sceloporus orcutti), western whiptail
(A. tigris), and Baja California spiny lizard (S. zosteromus)
(Grismer, 2002). Diurnal desert lizard species show different
sizes of home range, exhibit varying territoriality, and differ-
ent kinds of foraging behaviour (Pough et al., 2004). The foli-
vores, D. dorsalis and S. slevini, have a small home range,

Fig. 1 – Study area on Isla Coronado in the Gulf of California, off the east coast of the Baja California Peninsula. Transects and
principal habitats are shown.
a c t a o e c o l o g i c a 3 3 ( 2 0 0 8 ) 2 8 0 – 2 9 0282
where size tends to increase as the patchiness of food in-
creases, and defend polygynous territories (Kwiatkowski and
Sullivan, 2002). Phrynosomatidae species, genus Callisarus,
Uta, Urosaurus, Sceloporus, and Petrosaurus, commonly sit and
wait to prey on mobile prey and defend a small home range
(ambush, Pough et al., 2004). Finally, Teiidae species, Aspido-
scelis, are active predators that mostly feed on cryptic or
slow-moving prey, have larger home ranges with a high over-
lap with neighbours, but with very little overlap of their core
areas where they obtain most of their food (Eifler and Eifler,
1998). We assumed that changes in spatial distribution are
the result of availability of resources and reproductive events
that affect the size of the home range; therefore, the distance
between states of numerical dominance, richness, and abun-
dance depend on seasonality.
2.3. Data collection
Patterned annual activity of desert lizard species varies, which
affects the frequency of sightings. For example, Aspidoscelis
hyperythra and A. tigris are more abundant in warm months
(May–July), Dipsosaurus dorsalis and Sauromalus slevini are

a c t a o e c o l o g i c a 3 3 ( 2 0 0 8 ) 2 8 0 – 2 9 0 283
sighted more frequently after summer rainfall (September–
October), while Sceloporus spp. and Petrosaurus repens prefer
months with temperatures below 25 �C (November–Decem-
ber). We chose May, September, and November to evaluate
the frequency of sighting lizard species. We delimited homo-
geneous areas (landscape units) based on interpretations of
aerial photographs (scale 1:75,000) and prepared four perma-
nent transects (3 to 9.8 km long and 6 m wide) for censuses
of the four habitats in 2004 in four days in May, September,
and November (Fig 1). The 6-m width was close to the limit
of identifying species by sight. Transects were divided into
25-m segments because GPS resolution was 6–15 m with
MapInfo 5.0 software (MapInfo, New York). We analysed
data at two levels, island and landscape, for dominance, rich-
ness, and abundance of diurnal lizard species. At the island
level, 862 segments (21,745 m) were evaluated during each
sampling period, distributed among the four habitats. At the
landscape level, we evaluated only the two principal land-
scapes, rocky hillsides with 328 segments (8200 m) and desert
scrubland with 224 segments (5600 m). We sighted 446 lizards
in May, 691 in September, and 330 in November. Transects
were sampled from 08:00 to 14:00 h and again from 16:00 to
18:00 h because most of species have bimodal daily activity.
Observations were not taken at 14:00 to 16:00 h during the hot-
test time of day when most species reduce their activity
(Grismer, 2002). The species and the GPS-defined location of
every sighting were recorded.
2.4. Model
First-order Markovian models are based on probabilities of
transition from one state to another after a one-distance
step (segment), which depends only on the previous state
(Formacion and Saila, 1994; Wootton, 2001a; Hill et al., 2004).
We assumed that the population is closed during the month
of the survey, without loss or gain of individuals. Wootton
(2001b) found that the application of second-order Markov
chains showed similar results as first-order Markov chains;
therefore, we assumed that the present state of systems
only depend on the previous sample. We employed three
kinds of information: (1) numerical dominance, most abun-
dant species by segment and ecological states are the species
and empty segments; (2) richness or number of species by seg-
ment and states are the number of species and empty seg-
ments; and (3) abundance or number of lizards by species
and states are the number of lizards and empty segment.
For example, for numerical dominance, let the lizard’s com-
munity be sampled at regular intervals, and let there be a re-
cord of the state observed at each sampling.
We defined a state ‘i’ at segment ‘s’ of the lizard commu-
nity to mean that species ‘i’ is the dominate species present
in the lizard community at segment ‘f’, and let there be ‘m’
species that dominate the lizard community at one segment
or another on segments under study (Roshchild and Mullen,
1985). Let the one-step transition probability ‘piw’ be defined
as: Piw ¼ the probability that the dominant species in the lizard
communities is species w, given that the dominant species in
the lizard community during the immediately preceding seg-
ment was species i.
The matrix of one-step transition probabilities for all the m
states of the lizard community is given by:
P ¼
��������
Pii Piw Pim
Piw Pww Pwm
. . .Pim Pmw Pmm
��������
where Pii ¼ the dominant species in the lizard community
from one segment to the next remains Species i and Piw ¼ the
predominant species in the lizard community changes from
one segment dominated by Species i to Species w, where
isw. When the matrix is post-multiplied by vector w(f), repre-
senting the composition of states at segment s, according to
Equation 1,
Wðfþ 1Þ ¼ PwðfÞ; (1)
the resulting vector w(f þ 1) describes the composition of the
lizard’s community at segment s þ 1. The previous equation
can be used iteratively to simulate the composition of the
community over an area. The resulting vector is the steady
states probability ‘wi’, which means that the probability that
the lizard’s community will be in a particular state is indepen-
dent of its initial state (Hill et al., 2004). This is a relatively
simple model, in which probability distribution of the
model-states stabilizes after a given number of model steps
(Tucker and Anand, 2004). This property makes the model
ideal for presenting spatial pattern dominance.
We tested three Markovian assumptions of models. (1)
Transition probabilities are constant and depend only on pre-
vious states. We hypothesized that if each transition probabil-
ity was independent of the previous state, then we would
expect ‘n/y’, where ‘y’ is defined like a number of segments be-
tween a pair of states (cells), where ‘n’ is the total number of
segments, and ‘y’ is the total number of cells. The chi-square
criterion (P < 0.05) was used to test this hypothesis (Roshchild
and Mullen, 1985). (2) Irreducibility implies that every state is
possibly recurrent, that is, having occurred once, there is
a non-zero probability that it will occur again. (3) When ergo-
dicity, a recurrent state within a finite class of communicating
states, is not periodic, it means that a class of communicating
states need only contain at least one member i with piis0.
2.5. Parameters
Replacement Analysis (P) is the probability that a randomly-
selected point in a stationary lizard community is replaced
by a different species. This probability is the average of the
stationary distribution of the probability of replacement, de-
fined as:
P ¼ wið1� Pii � PeiÞ=wi; (2)
where wi is the stationary frequency of species i, and e is in an
empty state (Hill et al., 2004).
Expected turnover segments (E) measure the number of
segments in which a point changes state (Hill et al., 2004).
The turnover segments of species i is the distance that a point
changes to state j, defined as:
E ¼ di ¼ 1=ð1� PiiÞ: (3)
Recurrence segments (R) is a point in any state that will
eventually leave this state and then return to it after some
a c t a o e c o l o g i c a 3 3 ( 2 0 0 8 ) 2 8 0 – 2 9 0284
number of segments (Hill et al., 2004). A recurrence segment of
state i is the average distance elapsing between points leaving
state i and returning to it, defined as:
R ¼ qi ¼ ð1�wiÞ=wið1� PiiÞ: (4)
We analysed three spatial levels. (a) Island, which describes
spatial segregation between species and richness in all seg-
ments evaluated during the three months of sampling. We
expected high probabilities of replacement between species
with similar habitat requirements as species with different re-
quirements. We calculated the steady-state probabilities of
numerical dominance and richness for the three months,
since the species had different annual activity cycles (Grismer,
2002). We expected seasonal variations. To test this hypothe-
sis, we used the G test at P < 0.05 (Sokal and Rohlf, 1981). (b)
Landscape, which describes the properties of species with
similar habitat requirements. We analysed only the species
in the rocky hillside and flatland desert scrubland landscapes
in September because the largest number of lizards occur at
this time. We expected lower rates of replacement, turnover,
and distance of recurrence than at the island level. We used
the G test at P < 0.05. (c) Species, which describes the spatial
pattern of the most abundant species at the landscape level.
The estimates illustrate a potential use of Markov chains in
spatial arrangement studies because each species responds
differently to habitat and will show differences as distance be-
tween lizards.
3. Results
3.1. Island level
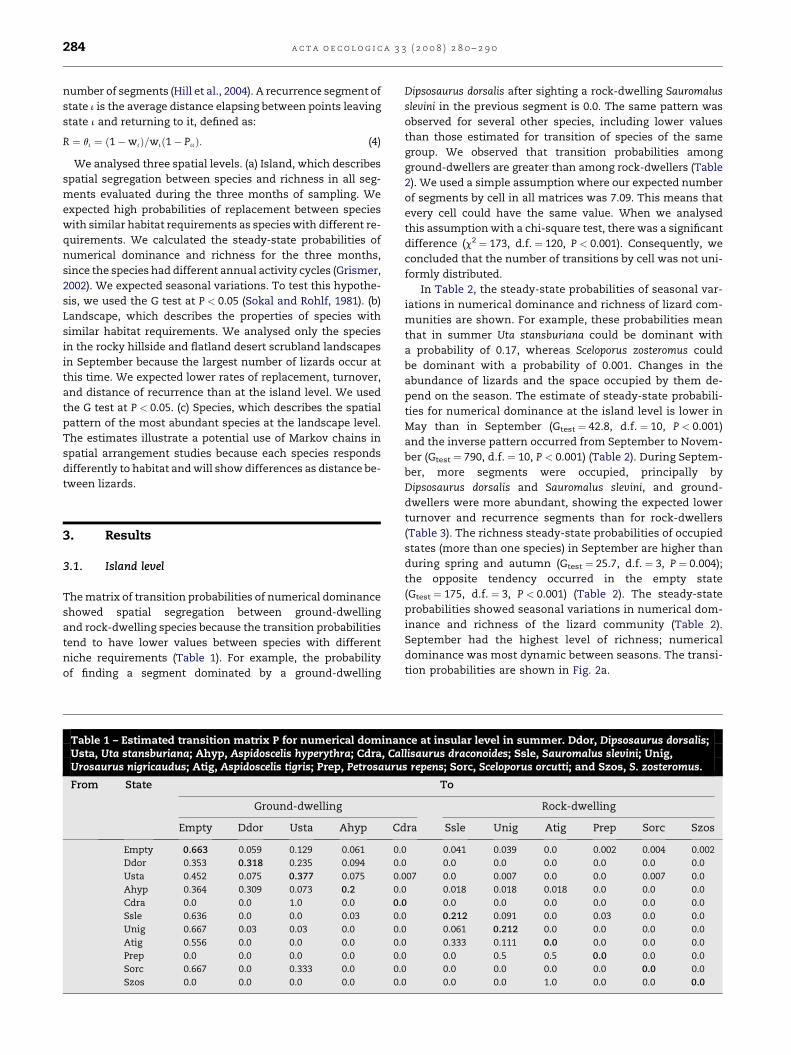
The matrix of transition probabilities of numerical dominance
showed spatial segregation between ground-dwelling
and rock-dwelling species because the transition probabilities
tend to have lower values between species with different
niche requirements (Table 1). For example, the probability
of finding a segment dominated by a ground-dwelling
Table 1 – Estimated transition matrix P for numerical dominanUsta, Uta stansburiana; Ahyp, Aspidoscelis hyperythra; Cdra, CaUrosaurus nigricaudus; Atig, Aspidoscelis tigris; Prep, Petrosauru
From State
Ground-dwelling
Empty Ddor Usta Ahyp Cd
Empty 0.663 0.059 0.129 0.061 0.
Ddor 0.353 0.318 0.235 0.094 0.
Usta 0.452 0.075 0.377 0.075 0.
Ahyp 0.364 0.309 0.073 0.2 0.
Cdra 0.0 0.0 1.0 0.0 0.
Ssle 0.636 0.0 0.0 0.03 0.
Unig 0.667 0.03 0.03 0.0 0.
Atig 0.556 0.0 0.0 0.0 0.
Prep 0.0 0.0 0.0 0.0 0.
Sorc 0.667 0.0 0.333 0.0 0.
Szos 0.0 0.0 0.0 0.0 0.
Dipsosaurus dorsalis after sighting a rock-dwelling Sauromalus
slevini in the previous segment is 0.0. The same pattern was
observed for several other species, including lower values
than those estimated for transition of species of the same
group. We observed that transition probabilities among
ground-dwellers are greater than among rock-dwellers (Table
2). We used a simple assumption where our expected number
of segments by cell in all matrices was 7.09. This means that
every cell could have the same value. When we analysed
this assumption with a chi-square test, there was a significant
difference (c2 ¼ 173, d.f. ¼ 120, P < 0.001). Consequently, we
concluded that the number of transitions by cell was not uni-
formly distributed.
In Table 2, the steady-state probabilities of seasonal var-
iations in numerical dominance and richness of lizard com-
munities are shown. For example, these probabilities mean
that in summer Uta stansburiana could be dominant with
a probability of 0.17, whereas Sceloporus zosteromus could
be dominant with a probability of 0.001. Changes in the
abundance of lizards and the space occupied by them de-
pend on the season. The estimate of steady-state probabili-
ties for numerical dominance at the island level is lower in
May than in September (Gtest ¼ 42.8, d.f. ¼ 10, P < 0.001)
and the inverse pattern occurred from September to Novem-
ber (Gtest ¼ 790, d.f. ¼ 10, P < 0.001) (Table 2). During Septem-
ber, more segments were occupied, principally by
Dipsosaurus dorsalis and Sauromalus slevini, and ground-
dwellers were more abundant, showing the expected lower
turnover and recurrence segments than for rock-dwellers
(Table 3). The richness steady-state probabilities of occupied
states (more than one species) in September are higher than
during spring and autumn (Gtest ¼ 25.7, d.f. ¼ 3, P ¼ 0.004);
the opposite tendency occurred in the empty state
(Gtest ¼ 175, d.f. ¼ 3, P < 0.001) (Table 2). The steady-state
probabilities showed seasonal variations in numerical dom-
inance and richness of the lizard community (Table 2).
September had the highest level of richness; numerical
dominance was most dynamic between seasons. The transi-
tion probabilities are shown in Fig. 2a.
ce at insular level in summer. Ddor, Dipsosaurus dorsalis;llisaurus draconoides; Ssle, Sauromalus slevini; Unig,s repens; Sorc, Sceloporus orcutti; and Szos, S. zosteromus.
To
Rock-dwelling
ra Ssle Unig Atig Prep Sorc Szos
0 0.041 0.039 0.0 0.002 0.004 0.002
0 0.0 0.0 0.0 0.0 0.0 0.0
007 0.0 0.007 0.0 0.0 0.007 0.0
0 0.018 0.018 0.018 0.0 0.0 0.0
0 0.0 0.0 0.0 0.0 0.0 0.0
0 0.212 0.091 0.0 0.03 0.0 0.0
0 0.061 0.212 0.0 0.0 0.0 0.0
0 0.333 0.111 0.0 0.0 0.0 0.0
0 0.0 0.5 0.5 0.0 0.0 0.0
0 0.0 0.0 0.0 0.0 0.0 0.0
0 0.0 0.0 1.0 0.0 0.0 0.0
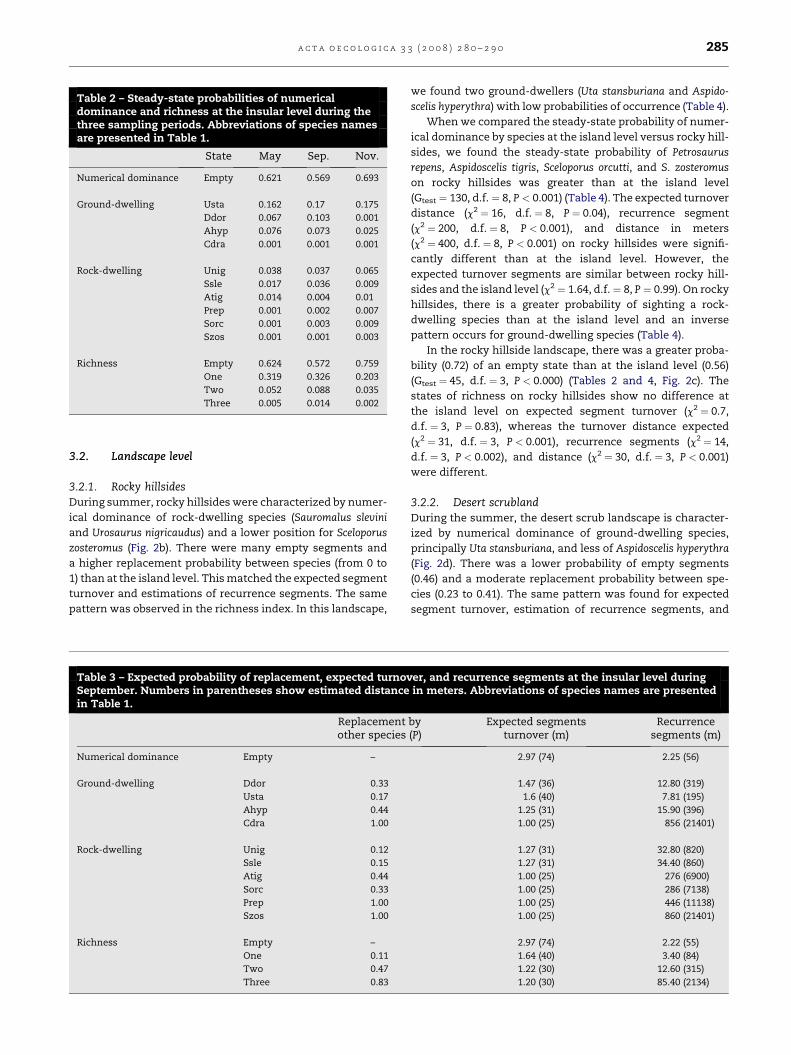
Table 2 – Steady-state probabilities of numericaldominance and richness at the insular level during thethree sampling periods. Abbreviations of species namesare presented in Table 1.
State May Sep. Nov.
Numerical dominance Empty 0.621 0.569 0.693
Ground-dwelling Usta 0.162 0.17 0.175
Ddor 0.067 0.103 0.001
Ahyp 0.076 0.073 0.025
Cdra 0.001 0.001 0.001
Rock-dwelling Unig 0.038 0.037 0.065
Ssle 0.017 0.036 0.009
Atig 0.014 0.004 0.01
Prep 0.001 0.002 0.007
Sorc 0.001 0.003 0.009
Szos 0.001 0.001 0.003
Richness Empty 0.624 0.572 0.759
One 0.319 0.326 0.203
Two 0.052 0.088 0.035
Three 0.005 0.014 0.002
a c t a o e c o l o g i c a 3 3 ( 2 0 0 8 ) 2 8 0 – 2 9 0 285
3.2. Landscape level
3.2.1. Rocky hillsidesDuring summer, rocky hillsides were characterized by numer-
ical dominance of rock-dwelling species (Sauromalus slevini
and Urosaurus nigricaudus) and a lower position for Sceloporus
zosteromus (Fig. 2b). There were many empty segments and
a higher replacement probability between species (from 0 to
1) than at the island level. This matched the expected segment
turnover and estimations of recurrence segments. The same
pattern was observed in the richness index. In this landscape,
Table 3 – Expected probability of replacement, expected turnovSeptember. Numbers in parentheses show estimated distancein Table 1.
Replacementother species
Numerical dominance Empty –
Ground-dwelling Ddor 0.33
Usta 0.17
Ahyp 0.44
Cdra 1.00
Rock-dwelling Unig 0.12
Ssle 0.15
Atig 0.44
Sorc 0.33
Prep 1.00
Szos 1.00
Richness Empty –
One 0.11
Two 0.47
Three 0.83
we found two ground-dwellers (Uta stansburiana and Aspido-
scelis hyperythra) with low probabilities of occurrence (Table 4).
When we compared the steady-state probability of numer-
ical dominance by species at the island level versus rocky hill-
sides, we found the steady-state probability of Petrosaurus
repens, Aspidoscelis tigris, Sceloporus orcutti, and S. zosteromus
on rocky hillsides was greater than at the island level
(Gtest ¼ 130, d.f. ¼ 8, P < 0.001) (Table 4). The expected turnover
distance (c2 ¼ 16, d.f. ¼ 8, P ¼ 0.04), recurrence segment
(c2 ¼ 200, d.f. ¼ 8, P < 0.001), and distance in meters
(c2 ¼ 400, d.f. ¼ 8, P < 0.001) on rocky hillsides were signifi-
cantly different than at the island level. However, the
expected turnover segments are similar between rocky hill-
sides and the island level (c2 ¼ 1.64, d.f. ¼ 8, P ¼ 0.99). On rocky
hillsides, there is a greater probability of sighting a rock-
dwelling species than at the island level and an inverse
pattern occurs for ground-dwelling species (Table 4).
In the rocky hillside landscape, there was a greater proba-
bility (0.72) of an empty state than at the island level (0.56)
(Gtest ¼ 45, d.f. ¼ 3, P < 0.000) (Tables 2 and 4, Fig. 2c). The
states of richness on rocky hillsides show no difference at
the island level on expected segment turnover (c2 ¼ 0.7,
d.f. ¼ 3, P ¼ 0.83), whereas the turnover distance expected
(c2 ¼ 31, d.f. ¼ 3, P < 0.001), recurrence segments (c2 ¼ 14,
d.f. ¼ 3, P < 0.002), and distance (c2 ¼ 30, d.f. ¼ 3, P < 0.001)
were different.
3.2.2. Desert scrublandDuring the summer, the desert scrub landscape is character-
ized by numerical dominance of ground-dwelling species,
principally Uta stansburiana, and less of Aspidoscelis hyperythra
(Fig. 2d). There was a lower probability of empty segments
(0.46) and a moderate replacement probability between spe-
cies (0.23 to 0.41). The same pattern was found for expected
segment turnover, estimation of recurrence segments, and
er, and recurrence segments at the insular level duringin meters. Abbreviations of species names are presented
by(P)
Expected segmentsturnover (m)
Recurrencesegments (m)
2.97 (74) 2.25 (56)
1.47 (36) 12.80 (319)
1.6 (40) 7.81 (195)
1.25 (31) 15.90 (396)
1.00 (25) 856 (21401)
1.27 (31) 32.80 (820)
1.27 (31) 34.40 (860)
1.00 (25) 276 (6900)
1.00 (25) 286 (7138)
1.00 (25) 446 (11138)
1.00 (25) 860 (21401)
2.97 (74) 2.22 (55)
1.64 (40) 3.40 (84)
1.22 (30) 12.60 (315)
1.20 (30) 85.40 (2134)

Fig. 2 – Diagram of transition probabilities of: (a) richness at the insular level, (b) numerical dominance; and (c) richness in
rocky hillsides, (d) numerical dominance, and (e) richness in desert scrub.
a c t a o e c o l o g i c a 3 3 ( 2 0 0 8 ) 2 8 0 – 2 9 0286
richness index. In this landscape, we did not find rock-
dwelling species (Table 4).
Steady-state probabilities for numerical dominance in des-
ert scrub differ significantly from the island level (Gtest ¼ 15,
d.f. ¼ 3, P ¼ 0.001). In desert scrubland, Uta stansburiana had
a steady-state probability of 0.23 as a dominant species and
Aspidoscelis hyperytrhus has a steady-state probability of 0.11.
Comparing expected segments and distance turnover, we
did not find significant statistical differences between desert
scrubland and the island level (c2 ¼ 0.54, d.f. ¼ 3, P ¼ 0.96;
c2 ¼ 7.9, d.f. ¼ 3, P ¼ 0.09). We found lower expected recur-
rence segments and distance for richness states within desert
scrubland than at the island level (c2 ¼ 7.9, d.f. ¼ 3, P ¼ 0.04;
c2 ¼ 80, d.f. ¼ 8, P ¼ 0.001). In this landscape, there were less
empty segments than at the island level (Gtest ¼ 8.6, d.f. ¼ 3,
P ¼ 0.035, see Tables 2 and 4, Fig. 2e). Number of species by
segment in desert scrubland were no different than at the is-
land level, as in the case for expected segment turnover and
distance (c2 ¼ 0.27, d.f. ¼ 3, P ¼ 0.96; c2 ¼ 5.16, d.f. ¼ 3,
P ¼ 0.16). However, recurrence segments and distance were
different (c2 ¼ 8, d.f. ¼ 3, P ¼ 0.04; c2 ¼ 30, d.f. ¼ 3, P ¼ 0.001).
3.3. Species level
This analysis was made for the most abundant species in both
landscapes. They were defined according to estimates of
steady-state probabilities that are >0.095; the value among
species ranged from 0.003 to 0.235 (Table 4); therefore, we
selected five species (Table 5). Table 5 shows that recurrence
for empty segments is, on average, between 1.2 to 1.8 seg-
ments (from 30 m to 45 m) for all species. When a species is
sighted in a segment, we expect to sight it again within 4.7
to 27.7 segments (117 to 692 m), depending on the species.
The state with four lizards per segment varied from 111 to
115 (2787 to 2890 m). Some species were absent from this
state, with different combinations of this description applying
to states, as shown in Table 5. According to these results, Uta
stansburiana had the most individuals by segment and the
shortest recurrence segment in the desert scrublands. On
rocky hillsides, Sauromalus slevini had more members by seg-
ment and Urosaurus nigricaudus had the shortest recurrence
segments and distances.
4. Discussion
Markov chains predict the most probable state of a system and
it depends only on the previous state of system before an
elapsed interval of time or space (Formacion and Saila,
1994). In temporal models, Markov chains can predict behav-
iour, dominance, or ecological succession, while spatial
models have been used to explain movements of animals. In
this sense, Hestbeck et al., 1991 noted that Markov chains
are not strictly appropriate for projected equilibrium regional
abundance of Canada geese; however, Markov chains are use-
ful for the study of population dynamics. Root and Kareiva

Table 4 – Expected probability during September of steady states, replacement, expected turnover, and recurrence segmentsfor numerical dominance and richness for numerical dominance and richness at the landscape level. Numbers inparentheses indicate estimated distance in meters. Abbreviations of species names are presented in Table 1.
Landscape Model State Steadystates
Replacement byother species (P)
Expected segmentsturnover (m)
Recurrencesegments (m)
Rocky hillside Numerical dominance Empty 0.720 – 4.18 (104) 1.62 (40)
Ssle 0.097 0.63 1.28 (32) 11.90 (297)
Unig 0.095 0.68 1.29 (32) 12.30 (307)
Ahyp 0.024 0.75 1.00 (25) 40.30 (1007)
Atig 0.024 0.63 1.00 (25) 40.30 (1007)
Usta 0.024 0.33 2.00 (50) 80.30 (2007)
Prep 0.006 0.00 1.00 (25) 164 (4100)
Sorc 0.006 1.00 1.00 (25) 164 (4100)
Szos 0.003 0.00 1.00 (25) 330 (8250)
Richness Empty 0.720 – 4.18 (104) 1.63 (40)
One 0.240 0.04 1.44 (36) 4.53 (113)
Two 0.034 0.50 1.11 (27) 31.50 (787)
Three 0.006 1.00 1.00 (25) 154 (3850)
Desert scrub Numerical dominance Empty 0.460 – 2.20 (55) 2.58 (64)
Usta 0.235 0.23 1.410 (35) 4.57 (114)
Ddor 0.190 0.34 1.66 (41) 6.99 (174)
Ahyp 0.115 0.41 1.23 (30) 9.71 (242)
Richness Empty 0.570 – 2.10 (74) 2.07 (51)
One 0.330 0.11 1.60 (40) 2.58 (64)
Two 0.090 0.48 1.20 (30) 11.50 (287)
Three 0.010 1.00 1.00 (25) 233 (5833)
a c t a o e c o l o g i c a 3 3 ( 2 0 0 8 ) 2 8 0 – 2 9 0 287
(1984) suggested that a flight sequence of cabbage butterflies
was Markovian. Kenkel (1993) demonstrated that individual
plant performance is determined in part by proximity of
neighbourhood, i.e. spatially dependent. Finally, Kincaid and
Cameron (1985) found that movements of cotton rats Sismodus
hispidus between habitats follow a Markovian pattern and
were dependent on age. Results in this study are similar to re-
sults reported in seasonal modelsdpreceding samples can
predict the state of the system (Wootton, 2001a,b; Hill et al.,
2004).
Transition probabilities of numerical dominance and rich-
ness at the island, landscape, and species levels can be used to
compress data into a few predictive quantities. Transition
Table 5 – Recurrence segments for the most abundant species inNumbers in parentheses indicate estimated distance in meters
State
Desert scru
Usta Ddor
Number of lizards/segment
0 1.6 (40) 1.7 (45)
1 4.7 (117) 8.5 (212)
2 7.5 (187) 18.4 (460)
3 58.7 (1467) 43.2 (1080)
4 115.6 (2890) –
Number expected for cell 9.5 14.6
d.f. 24 15
c2 81 75
P 0.00001 0.00001
probabilities, steady-state probabilities, replacement analysis,
expected turnover, and recurrence show different useful
information of the characteristics of spatial arrangement of
the lizard community. These include: (1) a view of the possible
interactions between species and their environment without
deeper evaluation (in the form of transition probabilities),
the interactions occurring in areas where growth, survival,
and reproduction are suitable for maintaining population dy-
namics in the absence of immigration (Pulliam, 2000)dalso,
dominance and spatial recurrence change, depending on avail-
ability of resources and habitat use where some species tend to
be dominant in certain areas and rare in other; (2) an index
of dominance and richness (steady-states probabilities);
desert scrub and rocky hillside landscapes. –, not recorded.. Abbreviations of species names are presented in Table 1.
Recurrence segments (m)
b Rocky hillside
Ahyp Unig Ssle
1.6 (40) 1.2 (30) 1.8 (45)
9.7 (242) 10.5 (262) 27.7 (692)
36.5 (912) 41.7 (1043) 116 (2900)
74.0 (1850) – 176 (4400)
111.5 (2787) – –
9 19 11.1
24 8 15
81 60 73
0.00001 0.00001 0.00001

a c t a o e c o l o g i c a 3 3 ( 2 0 0 8 ) 2 8 0 – 2 9 0288
(3) importance of seasonal activity and scale in the spatial dispo-
sition of species; (4) an estimate of spatial change in dominance
and richness (replacement and turnover segments); and (5)
observation of abundance of individual species (recurrence
segments). Also, since we include four different kinds of habi-
tats, we can analyse effect of transition probabilities between
pairs of species among diverse environmental conditions.
The results of the island and landscape models provide
some interesting insights into the importance of seasonal
change in the spatial arrangement of lizard species. Four
main results were found. (1) In contrast to sedentary species
(Hill et al., 2004), season influenced the estimates of parame-
ters of numerical dominance and richness that can be
explained by interspecific differences in periods of activity,
thermal preferences, or foraging behaviour. For example, Dip-
sosaurus dorsalis is active in warmer months and reduces its
activity at temperatures below 25 �C (Muth, 1980) as do Petro-
saurus repens and Sceloporus orcutti (Grismer, 1994). (2) There
were higher transition probabilities within a habitat group
(ground-dwelling or rock-dwelling) than between species.
This spatial segregation could reflect choice of different mi-
croclimates, reflecting physiological tolerance or choice of dif-
ferent vegetation (Pianka, 1982; Vitt, 1991). (3) Since steady-
state probabilities, expected turnover, and recurrence seg-
ments were lower at the island than the landscape level, it is
necessary to select the appropriate scale for evaluation in sur-
vey methods to obtain a more realistic estimate of abundance
and richness of species (Collins and Glenn, 1995; Bradbury
et al., 2001). In this case, the island level was used to contrast
species with different niche requirements and the landscape
level to analyse species with similar niche requirement. (4)
We found lower probabilities of steady states for rock-dwell-
ing species at the island level and rocky hillsides than species
in the desert scrubland.
We found that Uta stansburiana will probability become the
dominant species at the island level and in the desert scrub-
land and Urosaurus nigricaudus will continue to be the domi-
nant species on rocky hillsides. In contrast, Sceloporus
zosteromus and Petrosaurus repens have the lowest probabilities
to become dominant species at the island level and in the
rocky hillside landscape. Our results seem to be consistent
with observations about generalist species tending to achieve
dominance (Formacion and Saila, 1994; Brown et al., 1995).
According to Hanski and Gyllenberg, 1997, generalist species,
such as U. stansburiana, or species using broad resources are
common and widely distributed; whereas specialist species,
such as P. repens, are narrowly distributed.
We found that three species co-exist in high abundance
and low recurrence in segments of the desert scrubland, while
eight species are present in low abundance and high recur-
rence in segments of rocky hillside. This result is consistent
with the models of He and Legendre (2002) and Kneitel and
Chase (2004) that higher species richness in a sampling area
occurs if the species are spatially more regularly distributed
in the community, while high spatial aggregation of individ-
uals of a species result in lower species richness in a study
area. When environmental factors or resources are spatially
variable, different species find suitable microhabitats in dif-
ferent localities and co-exist regionally (Morris, 1990; He and
Legendre, 2002).
At the species level in desert scrubland, Aspidoscelis hyper-
ythus had the highest number of recurrence segments for their
states of abundance, possibly because this species actively
forages (Pianka, 1982) and defends a mobile territory from
other conspecifics (Eifler and Eifler, 1998). Uta stansburiana is
a passive, general predator, using broad spatial resources by
sitting and waiting (Grismer, 2002), which could explain its
high abundance and low recurrence in this landscape. On
the rocky hillside, Sauromalus slevini is the rock-dwelling spe-
cies with more lizards per segment, possibly because the
males protect their polygynous territories (Kwiatkowski and
Sullivan, 2002). In general, inter- and intraspecific differences
in abundance depend on spatially-fixed environmental varia-
tions and seasonal environmental variations (Ives and
Klopfer, 1997; Pulliam, 2000). In lizards, abundance autocorre-
lation can result from social organization (Glinsky and Krekor-
ian, 1985; Eifler and Eifler, 1998), foraging mode (Pianka, 1982),
mating systems (Sinervo et al., 2001; Kwiatkowski and Sulli-
van, 2002), or food resources (Vitt, 1991).
Our estimates of first-order Markovian chains are domi-
nated by empty segments and these probably play a funda-
mental role in inter- and intraspecific interactions. Gilpin
and Hanski, 1991 proposed that unoccupied, but habitable,
areas may also be important in the dynamics of the spatial
structure of populations. In this sense, populations may be-
come locally extinct, even in perfectly suitable habitats, and
the delay between extinction and re-colonization should leave
some fraction of suitable habitats unoccupied at any given
time (Thomas and Kunin, 1999). Patches and travelling waves
can be produced when populations are influenced by densities
of neighbouring populations (Sinervo et al., 2001), where spa-
tial-temporal patterns occur because local dispersal of indi-
viduals link the dynamics of adjacent areas (Morris, 1990;
Johnson, 2000).
Legendre (1993) explained that spatial structuring is an im-
portant component of ecosystems, where heterogeneity is
functional, and not the result of some random, noise-generat-
ing process. Because the spatial pattern in the desert lizard
community is variable, with spatial patterns that change sea-
sonally (Vitt, 1991; Case, 2002; Grismer, 2002), we quantified
the probability that a random sighting of a lizard at an insular
and landscape level in different periods of year. To predict
spatial behaviour in biological communities, it was necessary
to quantify the complete distribution of probabilities that
describe movements and sightings along all possible land-
scapes and seasons (Bradbury et al., 2001). If the dynamics of
movement are even moderately complex, it is not possible
to describe complete sight probability of movement in conve-
nient, closed forms, such as those found by other researchers
(Yoccoz et al., 2001; MacKenzie et al., 2003). We need to know
not only what physical boundaries delineate lizard species,
but also to show how rapidly the components of a species
mix change within those boundaries.
In conclusion, we could establish a model where each pair
of values was dependent only upon its immediate predecessor
segment. According to Liebhold and Gurevitch, 2002, spatial
dependence of ecological data has typically been a problem
that can reduce a field observer’s capacity to understand the
spatial pattern of the population or community. This effort
illustrates how first-order Markovian spatial chains may be

a c t a o e c o l o g i c a 3 3 ( 2 0 0 8 ) 2 8 0 – 2 9 0 289
used to examine spatial dependence of abundance and rich-
ness at different scales and seasons. However, understanding
how neighbouring elements affect each other or how they af-
fect a process is quite different from classical ecological
concerns for the structure and function of discrete communi-
ties, populations, or ecosystems (Pickett and Cadenasso, 1995).
Acknowledgements
We thank Abelino Cota, Franco Cota, Israel Guerrero, and
Armando Tejas for assistance with fieldwork. This research
was supported by grants from Consejo Nacional de la Ciencia
y Tecnologıa (CONACYT) and Fondo Mexicano para la Conser-
vacion de la Naturaleza (FMCN). Thanks are extended to one
anonymous reviewer who made useful comments and sug-
gestions to improve the manuscript.
r e f e r e n c e s
Bell, G., 2001. Neutral macroecology. Science 293, 2413–2418.Bevers, M., Flather, C.H., 1999. The distribution and abundance of
population limited at multiple spatial scales. J. Anim. Ecol. 68,976–987.
Bradbury, R.B., Payne, R.J.H., Wilson, J.D., Krebs, J.B., 2001.Predicting population responses to resource management.Trends Ecol. Evol. 16, 440–445.
Brown, J., Mehlman, D., Stevens, G.C., 1995. Spatial variation inabundance. Ecology 76, 2028–2043.
Case, T.J., 2002. Ecology of reptiles. In: Case, T.J., Cody, M.C. (Eds.),New Island Biogeography in the Sea of Cortez. OxfordUniversity Press, New York, pp. 221–270.
Collins, S., Glenn, S., 1995. Effects of organismal and distancescaling on analysis of species distribution and abundance.Ecol. Appl. 7, 543–551.
Eifler, D.A., Eifler, M.A., 1998. Foraging behavior and spacingpatterns of the lizard Cnemidophorus uniparens. J. Herpetol. 1,24–33.
Formacion, S.P., Saila, S.B., 1994. Markov chain properties totemporal dominance changes in a Philippine pelagic fishery.Fish. Res. 19, 241–256.
Gilpin, M.E., Hanski, I., 1991. Metapopulation Dynamics: Empiricaland Theoretical Investigations. Academic Press, London.
Glinsky, T.H., Krekorian, C., 1985. Individual recognition in free-living adult male desert iguanas, Dipsosaurus dorsalis.J. Herpetol. 19, 541–554.
Grismer, L.L., 1994. The origin and evolution of the peninsularherpetofauna of Baja California, Mexico. Herpet. Nat. Hist. 1,51–106.
Grismer, L.L., 2002. Amphibians and Reptiles of Baja California, ItsPacific Islands, and the Islands in the Sea of Cortes. Universityof California Press, Berkeley.
Hanski, I., Gyllenberg, M., 1997. Uniting two general patterns inthe distribution of species. Science 275, 397–400.
Hall, F.G., Botkin, D.B., Strebel, D.E., Woods, K.D., Goetz, S.J., 1991.Large-scale patterns of forest succession as determined byremote sensing. Ecology 72, 628–640.
He, F., Legendre, P., 2002. Species diversity pattern derived fromspecies-area models. Ecology 83, 1185–1198.
Hestbeck, J.B., Nichols, J.D., Malecki, R.A., 1991. Estimates ofmovement and sites fidelity using mark-resight data ofwintering Canada Geese. Ecology 72, 523–533.
Hill, M.F., Witman, J.D., Caswell, H., 2004. Markov chain analysisof succession in rocky subtidal community. Am. Nat. 164,E46–E61.
Ives, A.R., Klopfer, E.D., 1997. Spatial variation in abundance createdby stochastic temporal variation. Ecology 78, 1907–1913.
Johnson, M.P., 2000. Scale of density dependence as an alternativeto local dispersal in spatial ecology. J. Anim. Ecol. 69, 536–540.
Kenkel, N.C., 1993. Modeling Markovian dependence inpopulations of Aralia nudicaulis. Ecology 74, 1700–1708.
Kincaid, M.B., Cameron, G.N., 1985. Interactions of cotton ratswith a patchy environment: dietary responses and habitatselection. Ecology 66, 1769–1783.
Kneitel, J.M., Chase, J.M., 2004. Trade-offs in community ecology:linking spatial scales and species coexistence. Ecol. Lett. 7,69–80.
Kwiatkowski, A.A., Sullivan, B.K., 2002. Mating system structureand population density in a polygynous lizards, Sauromalusobesus (¼ater). Inter. Soc. Behav. Ecol. 2, 201–208.
Legendre, P., 1993. Spatial autocorrelation: trouble or newparadigm. Ecology 73, 1659–1673.
Lichstein, J.M., Simons, T.R., Shriner, S.A., Franzreb, K.E., 2002.Spatial autocorrelation and autoregressive models in ecology.Ecol. Monogr. 72, 445–463.
Liebhold, A.M., Gurevitch, J., 2002. Integrating the statisticalanalysis of spatial data in ecology. Ecography 25, 553–557.
Lusseau, D., 2003. Effects of tour boats on the behaviour ofbottlenose dolphins: using Markov chain to modelanthropogenic impacts. Conserv. Ecol. 17, 1785–1793.
MacKenzie, D.I., Nichols, J.D., Hines, J.E., Knutson, M.G.,Franklin, A.A., 2003. Estimating site occupancy, colonization,and local extinction when a species in detected imperfectly.Ecology 84, 2200–2207.
Morris, D.W., 1990. Temporal variation, habitat selection andcommunity structure. Oikos 59, 303–312.
Muth, A., 1980. Physiological ecology of desert iguana (Dipsosurusdorsalis) eggs: temperature and water relations. Ecology 61,1335–1343.
Orloci, L., Orloci, M., 1988. On recovery, Markov chains, andcanonical analysis. Ecology 69, 1260–1265.
Pianka, E.R., 1982. Niche relations of desert lizards. In: Cody, M.,Diamond, J. (Eds.), Ecology and Evolution of Communities.Harvard University Press, Cambridge, MA, pp. 292–314.
Pickett, S.T.A., Cadenasso, M.L., 1995. Landscape ecology: spatialheterogeneity in ecological systems. Science 269, 331–334.
Pough, F., Andrews, R., Cadle, J., Crump, M., Savitsky, A., Wells, K.,2004. Herpetology. Pearson Prentice Hall, New Jersey.
Pulliam, H.R., 2000. On the relationship between niche anddistribution. Ecol. Lett. 3, 349–361.
Railsback, S.F., Stauffer, H.B., Harvey, B.C., 2003. What can habitatpreference models tell us? test using a virtual troutpopulation. Ecol. Appl. 13, 1580–1594.
Roshchild, R.S., Mullen, A.J., 1985. The information content ofstock-and-recruitment data and its non-parametricclassification. J. Cons. Int. Explor. Mer. 42, 116–124.
Root, R.B., Kareiva, P.M., 1984. The search resources by cabbagebutterflies (Pieris rapae) ecological consequences andadaptative significance of Markovian movements in a patchyenvironment. Ecology 65, 147–168.
Sokal, R.R., Rohlf, F.J.R., 1981. Biometry, second ed. Freeman, SanFrancisco, CA.
Sinervo, B., Svensson, E., Comendant, T., 2001. Condition genotype-by-environment interaction, and correlational selection inlizards life-history morphs. Evolution 10, 2053–2069.
Tanner, J.E., Hughes, T.P., Connell, J.H., 1996. The role of history incommunity dynamics: a modelling approach. Ecology 77,108–117.
Thomas, C.D., Kunin, W.E., 1999. The spatial structure ofpopulations. J. Anim. Ecol. 68, 647–657.

a c t a o e c o l o g i c a 3 3 ( 2 0 0 8 ) 2 8 0 – 2 9 0290
Tucker, B., Anand, C.M., 2004. The applications of Markov modelsin recovery and restoration. Int. J. Ecol. Environ. Sci. 30,131–140.
Venegas, B.C.S., 2003. Abundancia, distribucion y nicho de laslagartijas diurnas de Isla Coronados, Baja California Sur,Mexico. Master’s Thesis. Centro de Investigaciones Biologicasdel Noroeste, La Paz, B.C.S., Mexico.
Vitt, L.J., 1991. Desert reptile communities. In: Polis, G.A. (Ed.), TheEcology of Desert Communities. University of Arizona Press,Tucson, pp. 249–277.
Wileyto, E.P., Ewens, W.J., Mullen, M.A., 1994. Markov-recapturepopulation estimates: a tool for improving interpretations oftrapping experiments. Ecology 75, 1109–1117.
Wootton, J.T., 2001a. Prediction in complex communities: analysisof empirically derived Markov models. Ecology 82, 580–598.
Wootton, J.T., 2001b. Causes of species diversity differences:a comparative analysis of Markov models. Ecol. Lett. 4, 46–56.
Yoccoz, N.G., Nichols, J.D., Boulinier, T., 2001. Monitoring ofbiological diversity in space and time. Trends Ecol. Evol. 16,446–453.




















