
Stable isotope analysis of well-preserved 120,000-year-old herbivore bone collagen from the Middle...
10
Stable isotope analysis of well-preserved 120,000-year-old herbivore bone collagen from the Middle Palaeolithic site of Neumark-Nord 2, Germany reveals niche separation between bovids and equids Kate Britton a, b, ⁎ , 1 , Sabine Gaudzinski-Windheuser c , Wil Roebroeks d , Lutz Kindler c , Michael P. Richards a, e a Department of Human Evolution, Max Planck Institute for Evolutionary Anthropology, Deutscher Platz 6, 04103 Leipzig, Germany b Department of Archaeology, University of Aberdeen, St. Mary's Building, Elphinstone Road, Aberdeen, AB24 3UF, UK c Römisch-Germanisches Zentralmuseum, Forschungsbereich Altsteinzeit Schloss Monrepos, 56567 Neuwied, Germany and Johannes Gutenberg–Universität Mainz, Institut für Vor- und Frühgeschichte, Germany d Faculty of Archaeology, Leiden University, P.O. Box 9515, 2300 RA Leiden, The Netherlands e Department of Anthropology, 6303 NW Marine Drive, Vancouver, BC, Canada V6T 1Z1 abstract article info Article history: Received 9 August 2011 Received in revised form 18 March 2012 Accepted 20 March 2012 Available online 29 March 2012 Keywords: Stable isotopes Herbivores Bone collagen Palaeoecology Diet Resource partitioning Herbivores from the Neumark-Nord 2 archaeological site, Germany, were analysed for bone collagen stable carbon (δ 13 C) and nitrogen (δ 15 N) isotope ratios in order to investigate feeding ecology at this early Last Interglacial (Eemian) shallow-lake site. Of 42 faunal samples selected, 23 yielded collagen, demonstrating re- markable preservation for material of this age. The results indicate clear inter-specific differences in δ 15 N and δ 13 C values, notably between equids (Equus) and bovids (Bos/Bison), with mean difference Δ 15 N of +2‰ measured in the bovids compared to the equids. The potential reasons for these differences are explored, in- cluding physiology, herbivore feeding ecology, biogeography and resource partitioning within the local envi- ronment. The data are compared to previously published archaeological data, and modern experimental and ecological data, suggesting that these inter-specific differences are not consistent and therefore unlikely to be solely the product of physiology or habitual forage preference. Data from this study are compared to the local vegetation (as reconstructed from pollen profiles), and it is suggested that these trends are likely the result of niche partitioning at the shallow lake site, reflecting the local diversity in vegetational zones. The evidence for resource partitioning amongst Pleistocene herbivore communities at Neumark-Nord 2 and elsewhere is dis- cussed. This study represents one of the largest data sets for collagen of this age, and the implications for our understanding of Late Pleistocene herbivore ecology, local herbivore community behaviour and hominin palaeodietary studies are explored. © 2012 Elsevier B.V. All rights reserved. 1. Introduction The landscapes of Mid- and Late Pleistocene Europe were domi- nated by large herbivores, including pachyderms (e.g. Mammuthus, Elephas), equids (Equus), cervids (Cervus), and bovids such as bison (Bison) and aurochs (Bos). The mechanisms that allow for the co- existence of different modern mid- and large-sized grazing herbivore guilds have been debated, but are largely thought to lie in their differ- ent digestive systems that permit them to adopt alternative foraging strategies where ranges overlap (Janis, 1976), and/or through the selective feeding on different plant communities or portions of plants within the same biome—known as resource partitioning (Krysl et al., 1984; Menard et al., 2002). These mechanisms lead to niche separa- tion, through the occupation of different sub-habitats by the selection of different plants within the same habitat, or different parts of the same plant; and therefore facilitating biodiversity in herbivore species (Janis, 1976). It has previously been suggested that the high diversity of herbivore taxa living in the steppic biomes of Pleistocene North Europe can be explained by such dietary specialisation (Guthrie, 1982), with a proposed high floral biodiversity allowing niche separation and the avoidance of direct inter-specific competition (Drucker et al., 2003). In this study, we investigate the feeding palaeoecology of equids (Equus) and bovids (Bos/Bison) through stable isotope data, and explore the possibility (and nature) of such niche separation in Late Pleistocene Europe. The stable isotope analysis of fossil animal remains is a power- ful tool for investigating ancient environments, ecosystems and feeding behaviours, and is a method that can be used to directly investigate dif- ferences in herbivore dietary specialisation. Stable isotope techniques Palaeogeography, Palaeoclimatology, Palaeoecology 333-334 (2012) 168–177 ⁎ Corresponding author at: Department of Archaeology, University of Aberdeen, St. Mary's Building, Elphinstone Road, Aberdeen, AB24 3UF, UK. Tel.: + 44 1224 273823. E-mail addresses: [email protected], [email protected] (K. Britton), [email protected] (S. Gaudzinski-Windheuser), [email protected] (W. Roebroeks), [email protected] (L. Kindler), [email protected], [email protected] (M.P. Richards). 1 Current address: Department of Archaeology, University of Aberdeen, St. Mary's Building, Elphinstone Road, Aberdeen, AB24 3UF, UK. 0031-0182/$ – see front matter © 2012 Elsevier B.V. All rights reserved. doi:10.1016/j.palaeo.2012.03.028 Contents lists available at SciVerse ScienceDirect Palaeogeography, Palaeoclimatology, Palaeoecology journal homepage: www.elsevier.com/locate/palaeo
Transcript of Stable isotope analysis of well-preserved 120,000-year-old herbivore bone collagen from the Middle...

Palaeogeography, Palaeoclimatology, Palaeoecology 333-334 (2012) 168–177
Contents lists available at SciVerse ScienceDirect
Palaeogeography, Palaeoclimatology, Palaeoecology
j ourna l homepage: www.e lsev ie r .com/ locate /pa laeo
Stable isotope analysis of well-preserved 120,000-year-old herbivore bone collagenfrom the Middle Palaeolithic site of Neumark-Nord 2, Germany reveals nicheseparation between bovids and equids
Kate Britton a,b,⁎,1, Sabine Gaudzinski-Windheuser c, Wil Roebroeks d, Lutz Kindler c, Michael P. Richards a,e
a Department of Human Evolution, Max Planck Institute for Evolutionary Anthropology, Deutscher Platz 6, 04103 Leipzig, Germanyb Department of Archaeology, University of Aberdeen, St. Mary's Building, Elphinstone Road, Aberdeen, AB24 3UF, UKc Römisch-Germanisches Zentralmuseum, Forschungsbereich Altsteinzeit Schloss Monrepos, 56567 Neuwied, Germany and Johannes Gutenberg–Universität Mainz,Institut für Vor- und Frühgeschichte, Germanyd Faculty of Archaeology, Leiden University, P.O. Box 9515, 2300 RA Leiden, The Netherlandse Department of Anthropology, 6303 NW Marine Drive, Vancouver, BC, Canada V6T 1Z1
⁎ Corresponding author at: Department of ArchaeoloMary's Building, Elphinstone Road, Aberdeen, AB24 3UF
E-mail addresses: [email protected], [email protected] (S. Gaudzinski-Windheuser), w.ro(W. Roebroeks), [email protected] (L. Kindler), [email protected]@ubc.ca (M.P. Richards).
1 Current address: Department of Archaeology, UnivBuilding, Elphinstone Road, Aberdeen, AB24 3UF, UK.
0031-0182/$ – see front matter © 2012 Elsevier B.V. Alldoi:10.1016/j.palaeo.2012.03.028
a b s t r a c t
a r t i c l e i n f oArticle history:Received 9 August 2011Received in revised form 18 March 2012Accepted 20 March 2012Available online 29 March 2012
Keywords:Stable isotopesHerbivoresBone collagenPalaeoecologyDietResource partitioning
Herbivores from the Neumark-Nord 2 archaeological site, Germany, were analysed for bone collagen stablecarbon (δ13C) and nitrogen (δ15N) isotope ratios in order to investigate feeding ecology at this early LastInterglacial (Eemian) shallow-lake site. Of 42 faunal samples selected, 23 yielded collagen, demonstrating re-markable preservation for material of this age. The results indicate clear inter-specific differences in δ15N andδ13C values, notably between equids (Equus) and bovids (Bos/Bison), with mean difference Δ15N of +2‰measured in the bovids compared to the equids. The potential reasons for these differences are explored, in-cluding physiology, herbivore feeding ecology, biogeography and resource partitioning within the local envi-ronment. The data are compared to previously published archaeological data, and modern experimental andecological data, suggesting that these inter-specific differences are not consistent and therefore unlikely to besolely the product of physiology or habitual forage preference. Data from this study are compared to the localvegetation (as reconstructed from pollen profiles), and it is suggested that these trends are likely the result ofniche partitioning at the shallow lake site, reflecting the local diversity in vegetational zones. The evidence forresource partitioning amongst Pleistocene herbivore communities at Neumark-Nord 2 and elsewhere is dis-cussed. This study represents one of the largest data sets for collagen of this age, and the implications for ourunderstanding of Late Pleistocene herbivore ecology, local herbivore community behaviour and homininpalaeodietary studies are explored.
© 2012 Elsevier B.V. All rights reserved.
1. Introduction
The landscapes of Mid- and Late Pleistocene Europe were domi-nated by large herbivores, including pachyderms (e.g. Mammuthus,Elephas), equids (Equus), cervids (Cervus), and bovids such as bison(Bison) and aurochs (Bos). The mechanisms that allow for the co-existence of different modern mid- and large-sized grazing herbivoreguilds have been debated, but are largely thought to lie in their differ-ent digestive systems that permit them to adopt alternative foragingstrategies where ranges overlap (Janis, 1976), and/or through the
gy, University of Aberdeen, St., UK. Tel.: +44 1224 [email protected] (K. Britton),[email protected],
ersity of Aberdeen, St. Mary's
rights reserved.
selective feeding on different plant communities or portions of plantswithin the same biome—known as resource partitioning (Krysl et al.,1984; Menard et al., 2002). These mechanisms lead to niche separa-tion, through the occupation of different sub-habitats by the selectionof different plants within the same habitat, or different parts of thesame plant; and therefore facilitating biodiversity in herbivore species(Janis, 1976). It has previously been suggested that the high diversity ofherbivore taxa living in the steppic biomes of Pleistocene North Europecan be explained by such dietary specialisation (Guthrie, 1982), with aproposed high floral biodiversity allowing niche separation and theavoidance of direct inter-specific competition (Drucker et al., 2003). Inthis study, we investigate the feeding palaeoecology of equids (Equus)and bovids (Bos/Bison) through stable isotope data, and explore thepossibility (and nature) of such niche separation in Late PleistoceneEurope. The stable isotope analysis of fossil animal remains is a power-ful tool for investigating ancient environments, ecosystems and feedingbehaviours, and is a method that can be used to directly investigate dif-ferences in herbivore dietary specialisation. Stable isotope techniques

169K. Britton et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 333-334 (2012) 168–177
are based on the principle that an organism's tissues will reflect thechemical composition of food and water ingested during life (Kohn,1999). Isotopic methods have been used extensively in recent years toexplore the palaeobiology of different species, including predator–prey relationships (Richards et al., 2000; Bocherens, 2003); niche feed-ing behaviours/resource partitioning (Feranec, 2007; Feranec et al.,2010); and seasonal biogeography and range size (Hoppe et al., 1999;Feranec et al., 2007; Britton et al., 2011). Diachronic and spatial studiesallow the assessment of niche conservation and the stability of animalfeeding behaviours and biogeography through time (Feranec et al.,2007) and across different geographical regions (Szpak et al., 2010).
In modern case studies, numerous tissues can be used in stableisotope studies, including soft tissues sampled in vivo such as blood,hoof/nail, fur/wool/hair, horn and breath, as well as hard and soft tis-sues post-mortem such as bone, tooth and muscle. Except in rare casesof good preservation (such as permafrost environments), stable iso-tope investigations of archaeological and palaeontological materialsare normally restricted to skeletal materials, including the inorganicfraction of bone and tooth (so called ‘bioapatite’), and preserved pro-teins that may be extracted from these mineral matrices such as bonecollagen. Collagen is often the favoured analyte for stable isotopestudies on archaeological human and animal remains as modern ex-perimental data and published quality control criteria can be usedto evaluate extracted ‘collagen’ and assess its integrity prior to datainterpretation (Ambrose, 1990; van Klinken, 1999). Collagen mole-cules break-down through time in the burial environment, and theprocesses of diagenesis, hydrolysis and microbial attack can serve todegrade collagen, resulting in its loss or in the alteration of in vivo sta-ble isotope signatures (Ambrose, 1990; van Klinken, 1999; Collins etal., 2002; Nielsen-Marsh et al., 2007; Smith et al., 2007; Dobbersteinet al., 2009). Although the age of deposits can greatly influence thechances of collagen preservation, the rate of protein breakdownlargely depends on conditions within the local burial environment(Nielsen-Marsh et al., 2007; Smith et al., 2007) and, where conditionsare favourable, well-preserved collagen can be extracted and analysedfrom Late Pleistocene samples dating to older than 50,000 yr BP (Fizetet al., 1995; Bocherens et al., 1997, 1999, 2001; Jones et al., 2001).
Here, we report the results of stable carbon (δ13C) and nitrogen(δ15N) isotope analysis of herbivore bone collagen from the site ofNeumark-Nord 2, Germany (archaeological find layer NN2/2), withthe aim of investigating potential differences in herbivore feedingecology at this early Last Interglacial (Eemian) ‘water-hole’ site. Car-bon and nitrogen composition and mass ratios of ultra filtered bonecollagen show that the samples are mostly well preserved, indicatingextracted proteins are useful for reconstructing diet and feeding ecol-ogy. The data are explored for inter-specific differences and comparedto previously published bone collagen isotope data from a range ofother Late Pleistocene sites. Issues of herbivore digestive physiology,niche separation and resource partitioning amongst Late Pleistoceneequids and bovids are discussed, and the implications for homininpalaeodietary studies are addressed.
2. Reconstructing herbivore ecology using carbon and nitrogenisotope analysis
Carbon and nitrogen stable isotope techniques are based aroundthe principle that animal body tissues (e.g. bulk bone collagen) reflectthe isotopic composition of the food ingested throughout life. The rel-ative abundance of the stable isotopes of carbon, 13C and 12C (δ13C),varies characteristically between different biological communitiese.g. between plants of different photosynthetic pathway (Smith andEpstein, 1970; DeNiro and Epstein, 1978) or between terrestrial andmarine ecosystems (Schoeninger and DeNiro, 1984). There is a frac-tionation effect between plant δ13C and herbivore bone collagenδ13C values of approximately +5‰ (Bocherens and Drucker, 2003).Studies of resource partitioning and niche feeding ecology in modern
and ancient ecosystems have generally focused on environmentswhere both C3 and C4 plants can be found, such as sub-tropical Africa(Koch et al., 1995; Cerling et al., 1999) and North America (Gadbury etal., 2000; MacFadden, 2008). These plants have different photosyn-thetic pathways, selecting against 13C to different degrees, and there-fore display large differences in their δ13C. These variations are thenexpressed further up the food chain in herbivore tissues and, in turn,even in carnivores and humans, revealing resource partitioning inmixed C3/C4 environments. However, recent studies have also demon-strated the usefulness of carbon isotope analysis to reveal resourcepartitioning among modern ungulates in purely C3 environments(Feranec, 2007). Such variations stem from intra-ecosystem variationsin carbon and nitrogen isotope values of different plant communitiesand also reflect dietary choices of herbivores living within C3 environ-ments. For example, lichens exhibit higher δ13C values than other ter-restrial C3 plants (Park and Epstein, 1960; Teeri, 1981; Maguas andBrugnoli, 1996) and these less negative values can be exhibited inthe tissues of specialist lichen feeders such as modern barren groundcaribou (Ben-David et al., 2001; Drucker et al., 2010, 2001). Other car-bon isotope studies of modern ecosystems have demonstrated thatvegetation in densely forested areas has characteristically-low δ13Cvalues compared to those from more open areas (van der Merweand Medina, 1991; Heaton, 1999), leading to more negative δ13Cvalues in the tissues of leaf-foddered animals, due to the ‘canopy ef-fect’, also observed in archaeological case studies (Cerling et al.,2004; Noe-Nygaard et al., 2005). Conversely, woody plants are typical-ly enriched in 13C compared to herbaceous plants growing at the samesites, varying by 2‰ or more (Heaton, 1999). These modern casestudies demonstrate the potential for intra-ecosystem variations incarbon isotope values of different plant communities within purelyC3 environments. Where these are pronounced, differences in feedingbehaviours (resource partitioning) amongst different animals in thesame habitat can result in differences in the carbon isotope composi-tion of tooth enamel and bone collagen (Feranec, 2007).
The nitrogen isotope composition of animal collagen largely reflectsthe animal's position within food webs, with the ratio of 15N and 14N(δ15N) increasing by 3–5‰ with each trophic step (Bocherens andDrucker, 2003). However, although occupying the same trophic level,absolute δ15N values of herbivore tissues depend on a number of fac-tors, most notably on the varying 15N levels of the plants they consume(and ultimately the soils) at the base of the food chain. Nitrogenmetab-olism in certain animal species may also influence the δ15N of their tis-sues, although this has been poorly characterized to date. The majorityof terrestrial plants obtain their nitrogen from soil ammonium (NH4
+)or nitrate (NO3
−), and therefore the abundance of 15N in their vegetationreflects the 15N abundance of nitrogen sources in the soils (Shearer andKohl, 1986). A range of variables can lead to depletion or enrichment insoil and therefore plant 15N,which, in turn affects the tissue δ15N valuesof animal feeders—even herbivores at the same trophic level. Onemajorinfluence is that of local temperatures and water availability, which isinversely correlated with plant δ15N values in modern studies (Heatonet al., 1986; Schwarcz et al., 1999). The relationship between long-term climatic changes and δ15N of archaeological herbivore collagenhas been established in archaeological studies (van Klinken et al.,2000; Richards and Hedges, 2003; Hedges et al., 2004; Stevens andHedges, 2004; Stevens et al., 2008), likely due to soil water availabilitybeing reduced during cooler, drier climatic phases. Salinity has alsobeen demonstrated to positively correlate with δ15N values in studieson modern plants (Heaton, 1987; van Groenigen and van Kessel,2002), possibly inducing a form of ‘pseudo-aridity’. Variation in plantδ15N values in modern studies has also been demonstrated to varywith soil type (Schwertl et al., 2005) and proximity to coast (Virginiaand Delwiche, 1982; Heaton, 1987; Hicks et al., 2005); fire regime(Grogan et al., 2000); nitrogen availability (Hobbie et al., 2000);andthe addition of modern and ancient fertilizers organic and inorganic(Choi et al., 2003; Bogaard et al., 2007), amongst other factors including

2 An additional 19 samples from Horizons A, B and C (Bos/Bison, n=14; Equus sp.,n=3; Elephas antiquus, n=2) were also processed but did not yield sufficient materialfor analysis in either the >30 kDa or >10 kDa fraction, or (in a small number of cases)had immeasurable or extremely low C% (b5%), and/or N% (b2%). None of these samplesare included in the following discussion.
170 K. Britton et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 333-334 (2012) 168–177
plant physiology (Evans, 2001). These different conditions can meanthat there can be large variations in plant δ15N (>10‰) over relativelysmall areas (Sponheimer et al., 2003: 81). Data from studies onmodernmaterials and ecosystems therefore demonstrate the potential for localvariation in soil and plant δ15N and δ13C isotope composition to bereflected in local herbivores, and that the stable isotope analysis of ani-mal bone collagen can be used to investigate resource partitioning andunderlying isotopic variation in ancient ecosystems.
3. The site of Neumark-Nord 2
The site of Neumark-Nord 2 is located approximately 35 km westof Leipzig in Saxony-Anhalt, Germany (51°19′28″N, 11°53′56″E).First discovered in the 1980s, this area of the Geisel Valley contains aseries of shallow Middle and Late Pleistocene basins, rich with faunalremains and Palaeolithic artefacts (Strahl et al., 2010; Sier et al.,2011). The basins, formed by isostatic movements induced by lignitediapirism (Eissman, 2002; Mania and Mania, 2008), include the prox-imal sites of Neumark-Nord 1 (Mania et al., 1990; Mania and Mania,2008; Strahl et al., 2010) and Neumark-Nord 3 (Laurat and Brühl,2006). Extensive excavations were carried out at Neumark-Nord 2 be-tween 2004 and 2008, yielding c.20,000 Middle Palaeolithic flint arte-facts and c.120,000 faunal remains, dominated by warm-temperatespecies (Sier et al., 2011). A recent multi-disciplinary study ofNeumark-Nord 2 basin infill has produced detailed climatic and chro-nological proxy records, which have shown that the archaeologicalfind level NN2/2 preserved in the basin dates to the early part of an in-terglacial. The age of the interglacial succession is constrained by theunderlying diamicton, a till of Late Saalian/Drenthe age (Eissman,2002) and by the overlying Weichselian deposits. Amino acid racemi-sation analysis of Bithynia tentaculata opercula (Penkman in Sier et al.,2011: Supplementary Material [SM]) suggest that the interglacialdeposits are contemporaneous with those at the Amersfoort andAmsterdam basin in the Netherlands, the Eemian type localities area(e.g. Zagwijn, 1961; Cleveringa et al., 2000; Van Leeuwen et al.,2000). Thermoluminescene (TL) dating of five heated flint artefactsyielded a weighted mean age of 126±6 ka (Richter in Sier et al.,2011: SM). Study of the basin infill also yielded a high-resolution re-cord of the palaeomagnetic Blake-event (Sier et al., 2011).
Sedimentological and soil micromorphological studies indicate arapid infilling of the basin with fine-grained calcareous silts, a nearcontinuous process with very little evidence of soil formation duringperiods of non-deposition (Sier et al., 2011). Pollen are well preservedand data demonstrate an interglacial succession that is typical for theEemian interglacial in northern Europe, the succession starting withPollen Zone I and ending with Zones VI/VII, (sensu Menke and Tynni,1984) at the top of the sections. This further constrains the durationof deposition represented in these deposits (Sier et al., 2011), withthe small basin most likely reflecting strictly local vegetation. Thishigh sedimentation rate, and subsequent lack of soil formation andbioturbation, and these local favourable local taphonomic conditionshave resulted in a well-stratified, well-preserved assemblage of most-ly anthropogenically-modified faunal remains (Sier et al., 2011).
The zooarchaeological record at Neumark-Nord 2 is dominated bylarge sized mammals; bovids including aurochs (Bos primigenius) andbison (Bison priscus), and horse (Equus sp.), and medium sized cervidssuch as red deer (Cervus elaphus) and fallow deer (Dama dama). Theassemblage also includes other warm-temperate species such asstraight-tusked elephant (Elephas antiquus), bear (Ursus sp.), smallcarnivores and pond tortoise (Emys orbicularis). Bones show abundanttraces of butchery and marrow extraction, indications that they arethe product of hominin (Neanderthal) activity. The numerous flint ar-tefacts found are the by-product of various simple ways of producingcores and flakes, with simple scrapers, notches and denticulates form-ing major tool categories. The faunal samples selected for this studyoriginated fromHorizon A to C (predominantly B) from archaeological
find level Level 2/2. This sequence broadly corresponds to the Corylus-phase (IVa sensu Menke and Tynni, 1984) and is calculated to repre-sent less than 2000 yr of sediment accumulation (Sier et al., 2011). Itshould be noted that the faunal remains were disarticulated, brokenup and probably not in situ, although there is very little evidence ofpost-deposition disturbance in the profile (Sier et al., 2011). The con-temporary pollen record at the site includes many plant species in-cluding aquatic and riparian species, and grasses from open-land andwoodland species, with evidence for the presence of much troddenareas. Steppe indicators are present too, whereas the presence ofIlex (holly) and Hedera (Ivy) indicate that the interglacial was of an“oceanic” character. Pollen also suggests the presence of forestedareas, further away from the small basin (Bakels in Sier et al., 2011:SM).
4. Methods
In this study, collagen was successfully extracted and analysedfrom 232 samples (Bos/Bison, n=15; Equus sp., n=7; cervid, n=1)fromearly Eemian deposits at Neumark-Nord 2 (Level NN2/2). Speciesdeterminationswere conducted by one of the authors (LK). The horsesat Neumark-Nord 2 are all caballoid, with the larger, less gracile mo-lars indicative of the Caballine clade. Bovids included Bos primigenius,as well as a number of samples of Bos/Bison in which genus or speciescould not be determined. The single cervid sample could not be iden-tified to genus level (Cervus orDama) but is likely to be Cervus elaphus.
Samples (~1 g) were removed from bones with a circular saw andexternal surfaces were removed using air abrasion. All bone sampleswere prepared following the collagen extraction method of Longin(1971), with modifications based on the recommendations of Collinsand Galley (1998), with the addition of an ultrafiltration step (Brownet al., 1988). Samples were demineralized in 0.5 M hydrochloric acidat 6–8 °C for 3–10 days with acid changed at regular intervals. Theywere then rinsed to neutrality with de-ionized water and gelatinizedin a weak acidic (pH 3) HCl solution at 70 °C for 48 h. The liquid fractioncontaining the gelatinized protein was isolated through filtration using5–8 μm Ezee® mesh filters (Elkay Laboratory Products), and then puri-fied using a series of ultrafiltration steps to isolate both the >30 kDaand 10–30 kDa fraction for each sample where possible, the >30 kDafraction being preferred due to the presence of larger (and thereforepresumably more intact) protein molecules. The remaining solutions(i.e. one or two filtrates per sample) were then frozen and lyophilized.
The resulting extracted fractions were analysed for stable carbon(δ13C) and nitrogen (δ15N) isotope ratios at the Department ofHuman Evolution, Max Planck Institute for Evolutionary Anthropology(Leipzig, Germany). Samples were measured in duplicate (aside froms-eva-14786 and 14789, due to lack of material) on a Delta XP massspectrometer coupled to a Flash EA 2112 elemental analyzer. The δ13Cvalues and δ15N values are reported relative to the V-PDB standardand AIR standards respectively. Analytical error for the δ13C and δ15Nmeasurements was calculated from repeat measurements of internaland international standards and was determined to be ±0.1‰ (1σ) orbetter. Mean difference between isotope measurements gained fromduplicate analyses of samples was ±0.1‰ (1σ) or better for both δ13Cand δ15N values.
5. Results
Isotopic results from the animal bone collagen extracted and ana-lysed from Neumark-Nord 2 are given in Table 1 (n=23). The C:N

Table 1Results of whole bone collagen stable carbon and nitrogen isotope analysis from herbi-vores from Neumark-Nord 2 (* = >30 KDa fraction, others were all the b30 to>10 KDa; § denotes samples containing b26.2% C and b11% N). The stratigraphic se-quence (archaeological find level Level 2/2; Horizon A to C) represents b2000 yr ofsediment accumulation (Sier et al., 2011).
Samplenumber
Species Level Yield(%)
δ13C(‰)
δ15N(‰)
%C %N C:N
14828 Bos or Bison A b0.1 −20.5 2.0 32.5 12.0 3.214795 Bos or Bison B2 0.6 −20.7 6.2 31.4 11.8 3.114826 Bos or Bison B2 0.1 −21.1 6.7 29.9 10.3 3.414789 Bos or Bison B2 0.5 −20.8 6.3 31.1 10.9 3.314799 Equus B2a/B3oT 0.2 −22.1 2.6 15.4 5.2 3.4 §14803 Equus B3oT 0.8 −20.5 4.6 25.9 9.5 3.2 §14820 Equus B3oT b0.1 −21.7 3.9 27.0 9.3 3.4 §14794 Bos sp. B3oT 0.5 −20.4 5.4 32.0 11.6 3.214830 Bos or Bison B3oT 1.0 −20.9 6.7 35.9 13.1 3.214814 Equus B3mT 1.6 −21.3 3.6 35.2 12.5 3.3 *14807 Equus B3mT b0.1 −21.7 3.1 23.0 7.9 3.4 §14793 Equus B3mT 0.2 −21.9 4.0 14.3 5.2 3.2 §14786 Bos or Bison B3mT b0.1 −21.0 5.0 16.7 6.2 3.114824 Bos or Bison B3mT 0.4 −20.7 6.5 33.7 11.9 3.3 §14815 Bos or Bison B3mT b0.1 −20.8 5.9 41.1 15.1 3.214805 Cervid B3mT 0.1 −22.1 3.9 12.6 4.2 3.5 §14809 Bos or Bison B3uT 0.3 −20.9 5.6 20.7 7.6 3.2 §14811 Bos or Bison B3uT 0.4 −21.0 5.4 33.0 12.1 3.214821 Equus C1 0.4 −21.6 4.3 33.0 11.8 3.314796 Bos or Bison C 0.9 −22.3 6.4 40.4 14.2 3.3 *14810 Bos or Bison C 2.4 −21.6 6.0 41.6 14.7 3.3 *14816 Bos or Bison C 0.5 −21.4 5.6 41.0 14.1 3.4 *14813 Bos primigenius C 1.2 −20.9 5.5 33.8 12.0 3.3 *
Fig. 1. A) Atomic C:N ratios plotted against sample δ15N values, and B) Atomic C:Nratios plotted against sample δ13C values for herbivore bone collagen from Neumark-Nord 2.
171K. Britton et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 333-334 (2012) 168–177
ratio, %C, %N and collagen yield for all samples are also presented.In each instance, it is indicated whether data was obtained fromthe smaller (10–30 kDa) or larger (>30 kDa) ultrafilter fraction. Theatomic C:N ratios of the samples analysed ranges between the rangeof 3.1 and 3.5, and are therefore within the expected range for well-preserved bone (DeNiro, 1985; Ambrose, 1990; van Klinken, 1999),confirmed by recent experimental data (Dobberstein et al., 2009).The collagen yields are relatively low compared to modern and morerecent archaeological bone (Ambrose, 1990; van Klinken, 1999), al-though comparable to previously published data for samples of a sim-ilar age (e.g. Bocherens et al., 1999). Here the filtration process, whichisolates only the largest preserved molecules and excludes smallermolecules, has helped in the retrieval of intact collagen (as evidencedby the C:N ratios of the samples). It is also likely that ultrafiltration hasserved to further reduce overall yield. Themajority of samples have %Cand %N resembling that of modern collagen, C=~35±8.8 wt.%,N=11–16 wt.% (van Klinken, 1999). δ15N and δ13C values are not sig-nificantly correlatedwith C:N ratios (see Fig. 1A and B), and although aminority of samples have b26.2% C and/or b11%N (see Table 1), theirC:N ratios all lie within the acceptable range. Furthermore, the δ-valuesof these samples all fall within themain group of the samples, suggestingit is likely that original isotope values have been preserved. When thesamples with low C and N % are excluded from the data set (e.g. sampless-eva-14786, 14793, 14799, 14803, 14805, 14807, 14809 and14820), themean δ13C and δ15N for each species remain the same or very similar.3
The bovids (aurochs/bison; n=15) have δ13C values ranging from−22.3‰ to−20.4‰, with a mean of−21.0‰±0.5‰ (1σ). Associatedδ15N values range from2.0‰ to 6.7‰with amean value of 5.7‰±1.1‰(1σ). Horses from the site (n=7)have δ13C values ranging from−22.1‰to −20.5‰ with a mean of −21.5‰±0.5‰ (1σ). Corresponding δ15Nvalues ranged from 2.6‰ to 4.6‰ with a mean of 3.7‰±0.7‰ (1σ).
3 δ13C=−21.0‰, δ15N=5.7‰ for Bos/Bison, which is the same as the means includ-ing low % samples; δ13C=−21.5‰, δ15N=4.0‰ for the Equus samples which is only0.3‰ higher in δ15N compared to the unadjusted mean value and equal to the meanδ13C value.
The single cervid sample that yielded collagen had a δ13C value of−22.1‰ and a δ15N value of 3.9‰.
Themean herbivore values (±1σ) are shown in Fig. 2. Two-sampleKolmogorov–Smirnov (K–S) tests indicate that the bovid and equidisotope data sets are drawn from the same distribution for both δ13C(Maximal Difference=0.58095; p=0.04733), and δ15N (MaximalDifference=0.85714; p=0.00058). One-way ANOVA suggests differ-ences between the horse and bovid data sets for both δ13C (F(1,20)=5.723; p=0.027) and δ15N (F(1,20)=17.520; pb0.001) values. There-fore, pairwise comparisons between the horse and bison data (bothfor carbon and nitrogen) reveal differences significant at the 0.05level (pb0.05). Post-hoc analysis by Tukey–Kramer test, which is con-servative and robust against the effects of unbalanced group size (seeKeppel and Wickens, 2004), also reveals significant differences be-tween horse and bison δ13C (p=0.0267) and δ15N (p=0.00055) ra-tios. The results of a non-parametric test (Mann–Whitney U test) alsosuggest significant differences between horse and bison δ15N ratios(p=0.00134) and, again to a lesser extent, δ13C ratios (p=0.02639)The cervid is very similar to the horses, with lower carbon and nitrogenvalues compared to the bovids, although given that it is only a singlesample statistical analysis was not undertaken.
6. Discussion
6.1. Quality and preservation of organic material
The C:N ratios, %C and %N data of the majority of the samples indi-cate good collagen preservation. Thus, we can consider the samples tocarry reliable isotopic signatures and therefore these can be used toelucidate palaeoenvironmental, palaeoecological and palaeodietaryinformation. Given the age of these samples, this preservation is re-markable. Such preservation is considered rare, and there are veryfew other published examples of isotope studies on collagen of
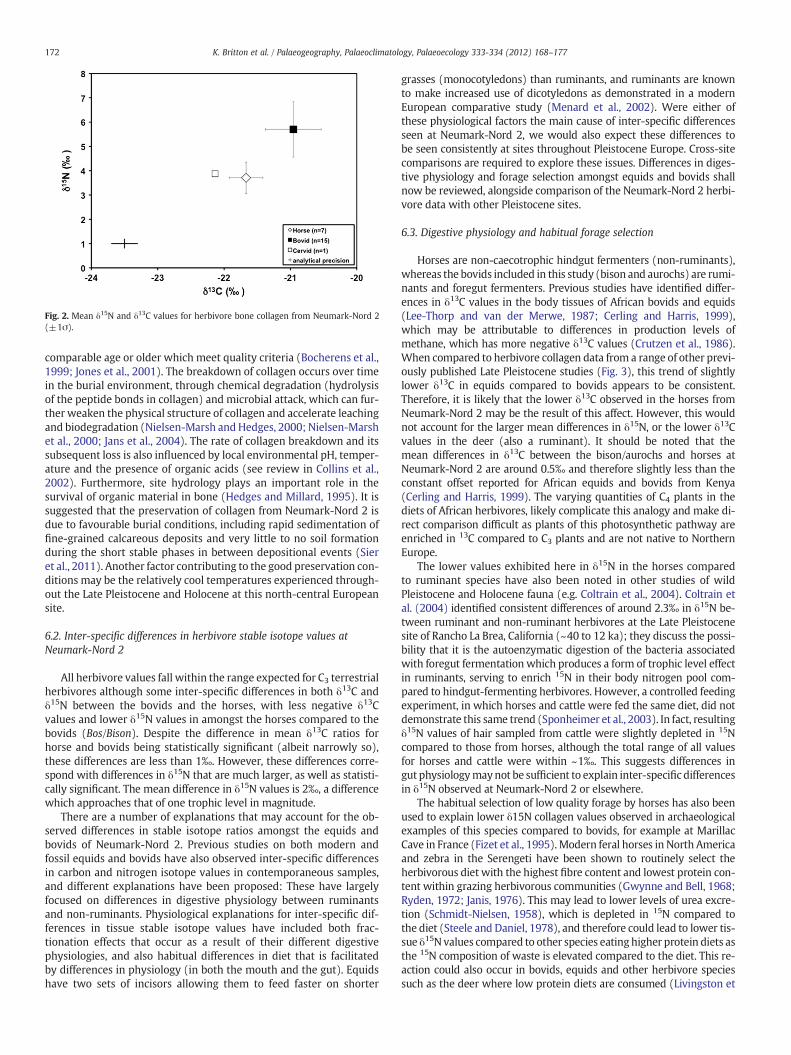
Fig. 2. Mean δ15N and δ13C values for herbivore bone collagen from Neumark-Nord 2(±1σ).
172 K. Britton et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 333-334 (2012) 168–177
comparable age or older which meet quality criteria (Bocherens et al.,1999; Jones et al., 2001). The breakdown of collagen occurs over timein the burial environment, through chemical degradation (hydrolysisof the peptide bonds in collagen) and microbial attack, which can fur-ther weaken the physical structure of collagen and accelerate leachingand biodegradation (Nielsen-Marsh and Hedges, 2000; Nielsen-Marshet al., 2000; Jans et al., 2004). The rate of collagen breakdown and itssubsequent loss is also influenced by local environmental pH, temper-ature and the presence of organic acids (see review in Collins et al.,2002). Furthermore, site hydrology plays an important role in thesurvival of organic material in bone (Hedges and Millard, 1995). It issuggested that the preservation of collagen from Neumark-Nord 2 isdue to favourable burial conditions, including rapid sedimentation offine-grained calcareous deposits and very little to no soil formationduring the short stable phases in between depositional events (Sieret al., 2011). Another factor contributing to the good preservation con-ditions may be the relatively cool temperatures experienced through-out the Late Pleistocene and Holocene at this north-central Europeansite.
6.2. Inter-specific differences in herbivore stable isotope values atNeumark-Nord 2
All herbivore values fall within the range expected for C3 terrestrialherbivores although some inter-specific differences in both δ13C andδ15N between the bovids and the horses, with less negative δ13Cvalues and lower δ15N values in amongst the horses compared to thebovids (Bos/Bison). Despite the difference in mean δ13C ratios forhorse and bovids being statistically significant (albeit narrowly so),these differences are less than 1‰. However, these differences corre-spond with differences in δ15N that are much larger, as well as statisti-cally significant. The mean difference in δ15N values is 2‰, a differencewhich approaches that of one trophic level in magnitude.
There are a number of explanations that may account for the ob-served differences in stable isotope ratios amongst the equids andbovids of Neumark-Nord 2. Previous studies on both modern andfossil equids and bovids have also observed inter-specific differencesin carbon and nitrogen isotope values in contemporaneous samples,and different explanations have been proposed: These have largelyfocused on differences in digestive physiology between ruminantsand non-ruminants. Physiological explanations for inter-specific dif-ferences in tissue stable isotope values have included both frac-tionation effects that occur as a result of their different digestivephysiologies, and also habitual differences in diet that is facilitatedby differences in physiology (in both the mouth and the gut). Equidshave two sets of incisors allowing them to feed faster on shorter
grasses (monocotyledons) than ruminants, and ruminants are knownto make increased use of dicotyledons as demonstrated in a modernEuropean comparative study (Menard et al., 2002). Were either ofthese physiological factors the main cause of inter-specific differencesseen at Neumark-Nord 2, we would also expect these differences tobe seen consistently at sites throughout Pleistocene Europe. Cross-sitecomparisons are required to explore these issues. Differences in diges-tive physiology and forage selection amongst equids and bovids shallnow be reviewed, alongside comparison of the Neumark-Nord 2 herbi-vore data with other Pleistocene sites.
6.3. Digestive physiology and habitual forage selection
Horses are non-caecotrophic hindgut fermenters (non-ruminants),whereas the bovids included in this study (bison and aurochs) are rumi-nants and foregut fermenters. Previous studies have identified differ-ences in δ13C values in the body tissues of African bovids and equids(Lee-Thorp and van der Merwe, 1987; Cerling and Harris, 1999),which may be attributable to differences in production levels ofmethane, which has more negative δ13C values (Crutzen et al., 1986).When compared to herbivore collagen data from a range of other previ-ously published Late Pleistocene studies (Fig. 3), this trend of slightlylower δ13C in equids compared to bovids appears to be consistent.Therefore, it is likely that the lower δ13C observed in the horses fromNeumark-Nord 2 may be the result of this affect. However, this wouldnot account for the larger mean differences in δ15N, or the lower δ13Cvalues in the deer (also a ruminant). It should be noted that themean differences in δ13C between the bison/aurochs and horses atNeumark-Nord 2 are around 0.5‰ and therefore slightly less than theconstant offset reported for African equids and bovids from Kenya(Cerling and Harris, 1999). The varying quantities of C4 plants in thediets of African herbivores, likely complicate this analogy and make di-rect comparison difficult as plants of this photosynthetic pathway areenriched in 13C compared to C3 plants and are not native to NorthernEurope.
The lower values exhibited here in δ15N in the horses comparedto ruminant species have also been noted in other studies of wildPleistocene and Holocene fauna (e.g. Coltrain et al., 2004). Coltrain etal. (2004) identified consistent differences of around 2.3‰ in δ15N be-tween ruminant and non-ruminant herbivores at the Late Pleistocenesite of Rancho La Brea, California (~40 to 12 ka); they discuss the possi-bility that it is the autoenzymatic digestion of the bacteria associatedwith foregut fermentation which produces a form of trophic level effectin ruminants, serving to enrich 15N in their body nitrogen pool com-pared to hindgut-fermenting herbivores. However, a controlled feedingexperiment, in which horses and cattle were fed the same diet, did notdemonstrate this same trend (Sponheimer et al., 2003). In fact, resultingδ15N values of hair sampled from cattle were slightly depleted in 15Ncompared to those from horses, although the total range of all valuesfor horses and cattle were within ~1‰. This suggests differences ingut physiologymaynot be sufficient to explain inter-specific differencesin δ15N observed at Neumark-Nord 2 or elsewhere.
The habitual selection of low quality forage by horses has also beenused to explain lower δ15N collagen values observed in archaeologicalexamples of this species compared to bovids, for example at MarillacCave in France (Fizet et al., 1995).Modern feral horses in North Americaand zebra in the Serengeti have been shown to routinely select theherbivorous diet with the highest fibre content and lowest protein con-tent within grazing herbivorous communities (Gwynne and Bell, 1968;Ryden, 1972; Janis, 1976). This may lead to lower levels of urea excre-tion (Schmidt-Nielsen, 1958), which is depleted in 15N compared tothe diet (Steele and Daniel, 1978), and therefore could lead to lower tis-sue δ15N values compared to other species eating higher protein diets asthe 15N composition of waste is elevated compared to the diet. This re-action could also occur in bovids, equids and other herbivore speciessuch as the deer where low protein diets are consumed (Livingston et

Fig. 3. Mean δ15N and δ13C values (±1σ) of Late Pleistocene equid and bovid skeletal collagen from a range of previously published studies of sites in north–west Europe includingCamiac (France, ~35,000 BP; Bocherens et al., 2005), La Berbie (France, ~35,000 BP; Bocherens et al., 2005), Saint Césaire (France, ~35,000 BP; Bocherens et al., 2005), Kent's Cavern(England, ~40,000 BP; Bocherens et al., 1995), Scladina 1A (Belgium, ~40,000 BP; Bocherens et al., 1997), Marillac (France, 40,000–45,000 BP; Fizet et al., 1995), Jonzac (France,40,000–55,000 BP; Richards et al., 2008), and Neumark-Nord 2 (Germany, ~126,000 BP; this study).
Fig. 4.Mean difference between stable isotope values (δ15N and δ13C) of Late Pleistoceneequid and bovid skeletal collagen from a range of sites in north–west Europe includingCamiac (France, ~35,000 BP; Bocherens et al., 2005), La Berbie (France, ~35,000 BP;Bocherens et al., 2005), Saint Césaire (France, ~35,000 BP; Bocherens et al., 2005), Kent'sCavern (England, ~40,000 BP; Bocherens et al., 1995), Scladina 1A (Belgium, ~40,000 BP;Bocherens et al., 1997), Marillac (France, 40,000–45,000 BP; Fizet et al., 1995), Jonzac(France, 40,000–55,000 BP; Richards et al., 2008), and Neumark-Nord 2 (Germany,~126,000 BP; this study).
173K. Britton et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 333-334 (2012) 168–177
al., 1962; Slade et al., 1970), although could consistently cause depletionin 15N in equids compared to bovids were they to habitually select lowquality forage.
6.4. Cross-site comparisons
In order to assess if the physiological factors or habitual forage selec-tion are themost likely explanation for the inter-specific differences ob-served between ruminant and non-ruminant species at Neumark-Nord2 and elsewhere, cross-site comparisons are necessary to determine ifthis is a universal trend (as with the δ13C values). As discussed above,bovids do display higher mean δ15N than equids at a number of LatePleistocene European sites in addition to Neumark-Nord 2, includingCamiac (Bocherens et al., 2005) and Marillac (Fizet et al., 1995). How-ever, horses are not consistently depleted in 15N compared to ruminantBos/Bison at all Pleistocene sites in Europe (see Figs. 3 and 4). For exam-ple, at the Late Pleistocene site of Kent's Cavern in Southern Englandnitrogen isotope values in Equus samples (tooth and bone=6.5‰) arehigher than woolly rhino (Coelodonta; bone=5.2‰, tooth=6.5‰)and aurochs (Bos sp.; tooth=5.6‰) (Bocherens et al., 1995). At SaintCésaire (Bocherens et al., 2005) and Scladina 1A (Bocherens et al.,1997) mean Equus sp. δ15N values are also slightly elevated comparedto Bison/Bos mean values, although inter-species difference are small(b1‰). Carbon and nitrogen stable isotope ratios of equids and bovidsare comparable at the French sites of La Berbie (Bocherens et al.,2005) and Jonzac (Richards et al., 2008).
Several issues pose complications for the interpretation of thesedata sets, most notably the small group sample sizes from many ofthe sites, perhaps a product of sample representation in the fossilrecord, the limitations of sampling permissions and also individualsample preservation: all of these are factors which can influence(and potentially bias)mean values. Furthermore, the datasets variablyinclude the analysis of collagen from both bone and tooth dentine,with bone representing time averaged dietary inputs (>5 yr prior todeath) and dentine representing dietary inputs from potentiallymuch short time periods (~1 yr or less, depending on the tooth, thespecies and the extent of sampling/portion of the tooth sampled). Inaddition, deciduous or early forming permanent teeth could be subjectto a breastfeeding or weaning influence, which can serve to elevateδ15N values in the offspring compared to that of the mother (e.g.Balasse et al., 1999; Balasse and Tresset, 2002). This could be especiallypertinent to the study of horses, which do not wean their young until
the age of ~9 months, a time at which portions of the permanent firstand second molars have already began to form (Hoppe et al., 2004).Finally, even in the most favourable circumstances, the nature ofthe archaeological record and the limitations of dating methodsmean that there can be no guarantee of the contemporaneity of thedifferent components of a faunal assemblage. ‘Baseline’ stable isotoperatios have been demonstrated to reflect large-scale differences in cli-mate and environment, which vary through time (van Klinken et al.,2000; Richards and Hedges, 2003). These changes are thought to re-flect large-scale environmental differences and/or climatic processes,which influence soils and plants at the base of the food web. Weresamples not contemporaneous, such baseline changes could result indifferent isotope values for herbivorous species at the same site,even where feeding niche is conserved through time. At Neumark-Nord 2, the Eemian archaeological deposits represent a relativelyshort period of time (Sier et al., 2011), and the fact that the majorityof samples of each species are from the same level (Level B3) helpsto further constrain date range of the faunal remains included in this

174 K. Britton et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 333-334 (2012) 168–177
study.4 However, this may not always be the case at other sites and lo-cations. Therefore, inter-specific variations of less than b1% should betreated with caution when assessing diet, trophic positions and nichefeeding behaviours (Casey and Post, 2011). However, given the largerisotopic differences (specifically in δ15N) between wild equids andbovids at Neumark-Nord 2 and other Late Pleistocene Europeansites, it seems likely these values may reflect real inter-specific differ-ences. Given the inconsistent nature of these variations across a rangeof Pleistocene sites, it seems that neither differences in physiology orhabitual feeding preference may be entirely sufficient to explain theinter-specific differences observed. There are several other explana-tions that could contribute to these variations, which will now be ex-plored, namely the influence of herbivoremovement/migration habits(biogeography) as well as the possibility that the herbivore feedingbehaviours themselves are not consistent through time. Such differ-ences in ranging behaviour or differences in dietary niche and feedingbehaviour (and the extent to which the niches of horses and bovidsare separate at any given time or place) may influence the stable iso-tope values of body tissues.
6.5. Late Pleistocene herbivore biogeography
In light of ‘baseline’ changes known to occur geographically (seediscussion in van Klinken et al., 2000) were a species to undertakeregular long-distance movements or migrations between climatic orenvironmental zones, this would lead to the incorporation of plantsfrom other areas into the diets of the animals, and therefore the incor-poration of non-local carbon and nitrogen isotope signatures into thebone collagen of this species. Therefore, migratory behaviour couldaccount for inter-specific differences in Pleistocene herbivore isotoperatios seen at some sites, particularly in archaeological species wheremigratory behaviour has previously been identified using strontiumisotope analysis such as European reindeer (Britton et al., 2011) andNorth American pachyderms (Koch et al., 1998; Hoppe et al., 1999).There has been very little research into the spatial behaviour of pre-historic wild horses, bison or aurochs in Europe. In North America,strontium isotope analysis has recently been used to identify limitedsmall-scale nomadic movements in prehistoric (pre-contact) GreatPlains bison herds (Widga et al., 2010). Similar evidence has alsobeen obtained from a single sample of Middle Palaeolithic Bison cf.priscus in France, demonstrating these large ungulates may have un-dertaken very limited lifetime movements compared to seasonally-migrating reindeer at the same site (Britton et al., 2011). Similarly,although some modern species of wild Equus undertake seasonalmigrations such as Burchell's zebra (Equus burchelli, Smuts, 1978),others do not (e.g. Equus kiang, see Schaller, 1998), and limited stron-tium isotope evidence from late Upper Palaeolithic Equus hydruntinusindicates this was not the case for at least this species of EuropeanPleistocene horse (Pellegrini et al., 2008). In light of recent and historicanalogues of northern Equus and Bos/Bison, along with the limitedstrontium isotope studies discussed above, there is little evidence tosuggest that Late Pleistocene horses, aurochs or bison undertook con-siderable long-distance seasonal movements. However, the influencethat slightly different geographical range limits could have on light sta-ble isotope bone collagen chemistry is difficult to estimate. There is aclear need for multi-period studies addressing herbivore palaeoecologyand palaeobiogeography through time.
4 It should be noted that amongst the bovids, there is, however, one clear outlier(s-eva-14828), originating fromHorizon A. Although included in themean calculations,this sample has a low δ15N value, compared to both the other bovids and the horses. Itcan be speculated that this may be a result of this sample coming from the uppermost(and slightly more recent) find horizon, reflecting such temporal ‘baseline’ changes.However, given that this is the only sample successfully analysed from Horizon A thereis no opportunity for comparison and it is not possible for any specific sequential (ortemporal) trends to be elucidated.
6.6. Niche separation and resource partitioning amongst herbivores inLate Pleistocene Europe
As explored above, the differences in Equus and Bos/Bison stableisotope values often seen at Late Pleistocene sites are not consistent.Physiological explanations for these differences (i.e. foregut vs. hind-gut digestion) are undermined by these inconsistencies, and also bya controlled feeding experiment involving modern horses and cattlefed identical diets (Sponheimer et al., 2003). These inconsistenciesalso suggest that habitual feeding behaviours (due to feeding prefer-ence or dental physiology) are not sufficient to explain the inter-specific differences observed at Neumark-Nord 2 or at other sites,although the caveats of archaeological datasets, such as potential sam-ple discontemporaneity, are recognized. Another possible explanationthat could account for the inter-specific differences at Neumark-Nord2 and elsewhere is that feeding behaviours in these species varythrough time, and across different environmental, climatic and eco-logical conditions, inducing different degrees of niche separationamongst the different herbivore species. As opposed to habitual feed-ing preference, this explanation suggests that the degree to whichequid and bovid feeding niches can or do overlap is flexible and con-tingent on the local environmental suite and biota, and the densityand variety of herbivore species in a particular area. Differences inresource partitioning can therefore account for the inconsistent dif-ferences between equids and bovids observed at the different LatePleistocene sites. Modern ecosystem studies support this idea, thatinter-specific competition between these two mid-sized grazers willcreate differences in feeding ecology and the degree of their niche sep-aration (Menard et al., 2002). The degree of this niche separation mayvary, but is inevitably greater ‘in times of scarcity’, resulting in comple-mentary and not overlapping niches (Janis, 1976). Therefore, suchresource partitioning (and plant species diversity) allows herbivorespecies that have similar ecologies to coexist within the same ecosys-tems. In the case of ‘wild’ modern European horses and cattle, it hasbeen proposed that natural coexistence in European grazing systemsis only possible if there are ‘adequate amounts of broadleaved plantsunavailable to horses’ (Menard et al., 2002: 131). Resource partition-ing implies a degree of fluidity in herbivore communities and dietaryniche across space and time. The Late Pleistocene was marked byclimatic change, which would have affected biotic productivity andinfluenced vegetation. It is not unreasonable to assume these changeswould have affected herbivore communities; triggering changes inlocal faunal suite, species types and density, and possibly also in thedegree of niche separation between different grazing herbivoresthrough resource partitioning. It may therefore be that the varyingisotopic spacing observed between different herbivores at differentsites throughout the Late Pleistocene is a reflection of the range oflocal plant δ15N values and the diversity of the floral suite (perhapsreflecting the localized affects of fluctuating gross climatic condi-tions), along with the density and diversity of grazing herbivorespecies and the niche separation/resource partitioning that wouldnecessitate that. This may indeed account for the high diversity ofherbivore taxa in the northern European steppe in the Late Pleistocene(Guthrie, 1982; Bocherens, 2003), with a high floral biodiversityallowing resource partitioning and the avoidance of direct inter-specific competition (Drucker et al., 2003).
Local contemporary pollen records at Neumark-Nord 2 can be usedto indicate the biodiversity of flora at this site, and indeed indicatemany plant species including aquatic and riparian species, and grassesfrom open-land andwoodland species (Bakels in Sier et al., 2011: SM).The range of δ15N values exhibited in plants in modern high-latitudeecosystems is larger than in warmer areas (Nadelhoffer et al., 1996),and aquatic and riparian species have been demonstrated to varyisotopically, compared to both open-grassland environments andwooded areas (Cloern et al., 2002, e.g. Nadelhoffer and Fry, 1994). Itmay be that the nitrogen isotope spacing between bovids and equids

175K. Britton et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 333-334 (2012) 168–177
at Neumark-Nord 2 may reflect the range of δ15N exhibited by plantcommunities at the site, and feeding strategies of the different speciessurrounding this unique ‘water hole’ site. The environmental mosaicand diverse plant species likely helped to support the numerousspecies living in this area, including bovids (Bos/Bison), horse (Equussp.), cervids (such as red deer, Cervus elaphus) and straight-tuskedelephant (Elephas antiquus). Stable isotope ecosystem studies ofmodern northern European wild/feral herbivore populations sharingthe same environments and local plants would help to better charac-terize how environmental suit, plant/soil δ15N values and resourcepartitioning can influence herbivore stable isotope values.
6.7. Implications for hominin dietary studies
The results of this study demonstrate the importance of selectingfaunal samples from the full range of species available at a site wheninvestigating human diet. Where herbivores are often used in stableisotope palaeodietary studies to establish a local (and contemporary)isotopic ‘baseline’, biases in sample selection or sample preservationcould lead to a species bias that could lead tomisinterpretation concern-ing the origin of human dietary protein. In the case of the Neumark-Nord 2 herbivores, the difference between two species of the same tro-phic level (i.e. horses and aurochs/bison) is 2‰, approaching a trophiclevel difference in magnitude (i.e. ~3‰). These data, and in comparisonto other published data, highlight potential stable isotope differencesthat can occur amongst contemporary herbivore populations. Althoughsuch temporal, spatial, and inter-specific effects have been previouslyexplored (van Klinken et al., 2000; Bocherens, 2003; Richards andHedges, 2003; Hedges et al., 2004; Stevens and Hedges, 2004), herewe stress the importance of considering niche feeding behaviours ofparticular species and highlight that niche conservation amongst herbi-vores in different geographic, climatic or environmental zones may notbe predictable through time, should not be assumed and can influencebone collagen stable isotope ratios even in purely C3 environments.
7. Conclusions
This study represents one of the largest bone collagen stable iso-tope data sets from an early Last Interglacial (Eemian) site, likelydue to the favourable local preservation conditions at Neumark-Nord 2. Carbon and nitrogen isotope analysis of extracted herbivorebone collagen revealed inter-specific differences in δ15N and δ13C be-tween equids, bovids and a single cervid at the site and the potentialexplanations for these differences have been explored, includingphysiology, herbivore feeding ecology, biogeography and resourcepartitioning through niche separation. Comparison with other LatePleistocene herbivore data sets, and a single modern controlled feed-ing experiment, reveal inconsistencies in the nitrogen isotope spacingbetween Equus and Bos/Bison, indicating that physiological differencesor differences in habitual forage selection between these different taxaalone may not be sufficient explanations for observed differences inbody tissue stable isotope chemistry. While acknowledging that farmore research is required into herbivore isotope ecology in both thepast and the present, we suggest that differences in biogeographyand/or the degree of niche separation in Late Pleistocene herbivorecommunities may contribute to observed intra-specific isotope differ-ences and that consideration of this should play an important role inhuman palaeodietary isotope studies. Biodiversity of local flora mayhave allowed resource partitioning in the steppic biomes of Europein the Late Pleistocene (Drucker et al., 2003), facilitating biodiversityand density of herbivore guilds (Guthrie, 1982; Bocherens, 2003);the constrained and well-dated sedimentary sequence at Neumark-Nord 2, including a local pollen sequence, offers a unique opportunityto compare herbivore isotope data to the contemporary floral suite.The diverse local floral suite may have facilitated niche separationbetween competing grazers, which may be evidenced in the bone
collagen stable isotope chemistry of equids and bovids from the site.We suggest that changes in species biogeography or feeding behav-iour could be contingent on contemporary climatic conditions and,while modern ecosystem studies cannot provide time depth requiredto explore the influence of broad-scale climate change on local bioticproductivity and biodiversity could potentially have on niche separa-tion (or geographical separation) between grazing species, archaeo-logical and palaeoecological studies of well-dated material have thepotential to explore this.
Acknowledgements
With thanks to Stefanie Bösel, Annabell Reiner and Olaf Nehlich(MPI-EVA) for technical and practical support with isotope analysis;and to the Landesamt für Denkmalpflege und Archäologie, Saxony-Anhalt, and Landesmuseum Sachsen-Anhalt in Halle for providingsamples; to Olaf Nehlich and Sandi Copeland (MPI-EVA, UC-Boulder)for comments on early versions of this manuscript; to Corrie Bakels(Leiden) for her perspectives on the pollen record at Neumark-Nord2; and to Jane Degabriel (Aberdeen) for useful discussions on modernherbivore community ecology. Financial support for the Neumark-Nord 2 excavations was provided by the Lausitzer MitteldeutscheBraunkohlengesellschaft mbH, the Landesamt für Denkmalpflegeund Archäologie Sachsen-Anhalt (Harald Meller, Susanne Friederich),the Römisch-Germanisches Zentralmuseum Mainz, the Leids Univer-siteits Fonds “Campagne voor Leiden” program and the NetherlandsOrganization for Scientific Research (N.W.O.). The isotope researchwas funded by Max Planck Institute and a Deutscher AkademischerAustausch Dienst Junior Research Grant to KB (ref: A0970923). Thanksalso to the University of Aberdeen for financial and professional sup-port during this project and preparation of the manuscript.
References
Ambrose, S.H., 1990. Preparation and characterization of bone and tooth collagen forisotopic analysis. Journal of Archaeological Science 17, 431–451.
Balasse, M., Tresset, A., 2002. Early weaning of Neolithic domestic cattle (Bercy, France)revealed by intra-tooth variation in nitrogen isotope ratios. Journal of Archaeolog-ical Science 29, 853–859.
Balasse, M., Bocherens, H., Mariotti, A., 1999. Intra-bone variability of collagen and ap-atite isotopic composition used as evidence of a change of diet. Journal of Archae-ological Science 26, 593–598.
Ben-David, M., Shochat, E., Adams, L.G., 2001. Utility of stable isotope analysis in study-ing foraging ecology of herbivores: examples from moose and caribou. Alces 37,421–434.
Bocherens, H., 2003. Isotopic biogeochemistry and the palaeoecology of the mam-moth steppe fauna. DEINSEA—Annual of the Natural History Museum Rotterdam,9, pp. 57–76.
Bocherens, H., Drucker, D., 2003. Trophic level isotopic enrichment of carbon and nitro-gen in bone collagen: case studies from recent and ancient terrestrial ecosystems.International Journal of Osteoarchaeology 13, 46–53.
Bocherens, H., Fogel, M.L., Tuross, N., Zeder, M., 1995. Trophic structure and climatic in-formation from isotopic signatures in Pleistocene cave fauna of Southern England.Journal of Archaeological Science 22, 327–340.
Bocherens, H., Billiou, D., Patou-Mathis, M., Bonjean, D., Otte, M., Mariotti, A., 1997. Pa-leobiological implications of the isotopic signatures (C-13, N-15) of fossil mammalcollagen in Scladina cave (Sclayn, Belgium). Quaternary Research 48, 370–380.
Bocherens, H., Billiou, D., Mariotti, A., Patou-Mathis, M., Otte, M., Bonjean, D., Toussaint,M., 1999. Palaeonvironmental and palaeodietary implications of isotopic biogeo-chemistry of last interglacial Neanderthal and mammal bones in Scladina Cave(Belgium). Journal of Archaeological Science 26, 599–607.
Bocherens, H., Billiou, D., Mariotti, A., Toussaint, M., Patou-Mathis, M., Bonjean, D., Otte,M., 2001. New isotopic evidence for dietary habits of Neandertals from Belgium.Journal of Human Evolution 40, 497–505.
Bocherens, H., Drucker, D.G., Billiou, D., Patou-Mathis, M., Vandermeersch, B., 2005. Iso-topic evidence for diet and subsistence pattern of the Saint-Césaire I Neanderthal:review and use of a multi-source mixing model. Journal of Human Evolution 49,71–87.
Bogaard, A., Heaton, T.H.E., Poulton, P., Merbach, I., 2007. The impact of manuring onnitrogen isotope ratios in cereals: archaeological implications for reconstructionof diet and crop management practices. Journal of Archaeological Science 34,335–343.
Britton, K., Grimes, V., Niven, L., Steele, T., Mcpherron, S., Soressi, M., Kelly, T.E., Jaubert,J., Hublin, J.-J., Richards, M.P., 2011. Strontium isotope evidence for migration inlate Pleistocene Rangifer: implications for Neanderthal hunting strategies at theMiddle Palaeolithic site of Jonzac, France. Journal of Human Evolution 61, 176–185.

176 K. Britton et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 333-334 (2012) 168–177
Brown, T.A., Nelson, D.E., Vogel, J.S., Southon, J.R., 1988. Improved collagen extractionby modified Longin method. Radiocarbon 30, 171–177.
Casey, M.M., Post, D.M., 2011. The problem of isotopic baseline: reconstructing the dietand trophic position of fossil animals. Earth-Science Reviews 106, 131–148.
Cerling, T.E., Harris, J.M., 1999. Carbon isotope fractionation between diet and bioapatitein ungulate mammals and implications for ecological and paleoecological studies.Oecologia 120, 337–363.
Cerling, T., Harris, J.M., Leakey, M.G., 1999. Browsing and grazing in elephants: theisotope record of modern and fossil proboscideans. Oecologia 120, 364–374.
Cerling, T.E., Hart, J.A., Hart, T.B., 2004. Stable isotope ecology in the Ituri Forest. Oeco-logia 138, 5–12.
Choi, W.-J., Ro, H.-M., Hobbie, E.A., 2003. Patterns of natural 15N in soils and plants fromchemically and organically fertilized uplands. Soil Biology and Biochemistry 35,1493–1500.
Cleveringa, P., Meijer, T., Van Leeuwen, R.J.W., De Wolf, H., Pouwer, R., Lissenberg, T.,Burger, A.W., 2000. The Eemian type locality at Amersfoort in the central Netherlands:redeployment of old and new data. In: Van Kolfschoten, T., Gibbard, P.L. (Eds.), TheEemian—local sequences, global perspectives: Geologie en Mijnbouw / NetherlandsJournal of Geosciences, 79 (2/3), pp. 197–216.
Cloern, J.E., Canuel, E.A., Harris, D., 2002. Stable carbon and nitrogen isotope composi-tion of aquatic and terrestrial plants of the San Francisco Bay estuarine system.Limnology and Oceanography 47, 713–729.
Collins, M.J., Galley, P., 1998. Towards an optimal method of archaeological collagenextraction: the influence of pH and grinding. Ancient Biomolecules 2, 209–222.
Collins, M.J., Nielsen-Marsh, C.M., Hiller, J., Smith, C.I., Roberts, J.P., Prigodich, R.V.,Wess, T.J., Csapò, J., Millard, A.R., Turner-Walker, G., 2002. The survival of organicmatter in bone: a review. Archaeometry 44, 383–394.
Coltrain, J.B., Harris, J.M., Cerling, T.E., Ehleringer, J.R., Dearing, M.-D., Ward, J., Allen, J.,2004. Rancho La Brea stable isotope biogeochemistry and its implications for thepalaeoecology of late Pleistocene, coastal southern California. Palaeogeography,Palaeoclimatology, Palaeoecology 205, 199–219.
Crutzen, P.J., Aslemann, I., Seiler, W., 1986. Methane production by domestic animals,wild ruminants, and other herbivorous fauna, and humans. Tellus 38B, 271–284.
Deniro, M.J., 1985. Postmortem preservation and alteration of in vivo bone collagenisotope ratios in relation to palaeodietary reconstruction. Nature 317, 806–809.
Deniro, M.J., Epstein, S., 1978. Influence of diet on the distribution of carbon isotopes inanimals. Geochimica et Cosmochimica Acta 42, 495–506.
Dobberstein, R.C., Collins, M.J., Craig, O.E., Taylor, G., Penkman, K.E.H., Ritz-Timme, S.,2009. Archaeological collagen: why worry about collagen diagenesis? Archaeolog-ical and Anthropological Sciences 1, 31–42.
Drucker, D., Bocherens, H., Pike-Tay, A., Mariotti, A., 2001. Isotopic tracking of seasonaldietary change in dentine collagen: preliminary data from modern caribou. Earthand Planetary Sciences 333, 303–309.
Drucker, D.G., Bocherens, H., Billiou, D., 2003. Evidence for shifting environmental con-ditions in Southwestern France from 33 000 to 15 000 years ago derived fromcarbon-13 and nitrogen-15 natural abundances in collagen of large herbivores.Earth and Planetary Science Letters 216, 163–173.
Drucker, D.G., Hobson, K.A., Münzel, S.C., Pike-Tay, A., 2010. Intra-individual variationin stable carbon (δ13C) and nitrogen (δ15N) isotopes in mandibles of modern car-ibou of Qamanirjuaq (Rangifer tarandus groenlandicus) and Banks Island (Rangifertarandus pearyi): Implications for tracing seasonal and temporal changes in diet.International Journal of Osteoarchaeology. doi:10.1002/oa.1220.
Eissman, L., 2002. Quaternary geology of the eastern Germany (Saxony, Saxon-Anhalt,South Brandenburg, Thüringia), type area of the Elsterian and Saalian Stages inEurope. Quaternary Science Reviews 21, 1275–1346.
Evans, R.D., 2001. Physiological mechanisms influencing plant nitrogen isotope compo-sition. Trends in Plant Science 6, 121–126.
Feranec, R.S., 2007. Stable carbon isotope values reveal evidence of resource partitioningamong ungulates frommodemC-3-dominated ecosystems in North America. Palaeo-geography, Palaeoclimatology, Palaeoecology 252, 575–585.
Feranec, R.S., Hadly, E.A., Paytan, A., 2007. Determining landscape use of Holocenemammals using strontium isotopes. Oecologia 153, 943–950.
Feranec, R., Garcia, N., Diez, J.C., Arsuaga, J.L., 2010. Understanding the ecology ofmamma-lian carnivorans and herbivores from Valdegoba cave (Burgos, northern Spain)through stable isotope analysis. Palaeogeography, Palaeoclimatology, Palaeoecology297, 263–272.
Fizet, M., Mariotti, A., Bocherens, H., Langebadre, B., Vandermeersch, B., Borel, J.P.,Bellon, G., 1995. Effect of diet, physiology and climate on carbon and nitrogen sta-ble isotopes of collagen in a late Pleistocene anthropic palaeoecosytem—Marillac,Charente, France. Journal of Archaeological Science 22, 67–79.
Gadbury, C., Todd, L., Jahren, A.H., Amundson, R., 2000. Spatial and temporal variationsin the isotopic composition of bison tooth enamel from the Early HoloceneHudson-Meng Bone Bed, Nebraska. Palaeogeography, Palaeoclimatology, Palaeoe-cology 157, 79–93.
Grogan, P., Bruns, T.D., Chapin, F.S., 2000. Fire effects on ecosystem nitrogen cycling in aCalifornian bishop pine forest. Oecologia 122, 537–544.
Guthrie, R.D., 1982. Mammals of the mammoth steppe as paleoenvironmental indica-tors. In: Hopkins, D.M., Matthews, J.V.J., Schweger, C.E., Young, S.B. (Eds.), Paleo-ecology of Beringia. Academic Press, New York, pp. 307–326.
Gwynne, M.D., Bell, R.H.V., 1968. Selection of vegetation components by grazing ungu-lates in the Serengeti National Park. Nature 220, 390–393.
Heaton, T.H.E., 1987. The 15N/14N ratios of plants in South Africa and Namibia: relation-ship to climate and coastal/saline environments. Oecologia 74, 236–246.
Heaton, T.H.E., 1999. Spatial, species and temporal variations in the 13C/12C ratios of C3plants: implications for palaeodiet studies. Journal of Archaeological Science 26,637–649.
Heaton, T.H.E., Vogel, J.C., Von La Chevallerie, G., Collett, G., 1986. Climatic influence onthe isotopic composition of bone nitrogen. Nature 322, 822–823.
Hedges, R.E.M., Millard, A.R., 1995. Bones and groundwater: towards the modelling ofdiagenetic processes. Journal of Archaeological Science 22, 155–164.
Hedges, R.E.M., Stevens, R.E., Richards, M.P., 2004. Bone as a stable isotope archive forlocal climatic information. Quaternary Science Reviews 23, 959–965.
Hicks, B.J., Wipfli, M.S., Lang, D.W., Lang, M.E., 2005. Marine-derived nitrogen andcarbon in freshwater–riparian food webs of the Copper River Delta, southcentralAlaska. Oecologia 144, 558–569.
Hobbie, E.A., Macko, S.A.,Williams,M., 2000. Correlations between foliar δ15N andnitrogenconcentrationsmay indicate plant–mycorrhizial interactions. Oecologia 122, 273–283.
Hoppe, K.A., Koch, P.L., Carlson, R.W., Webb, D.S., 1999. Tracking mammoths andmastodons: reconstruction of migratory behaviour using strontium isotope ratios.Geology 27, 439–442.
Hoppe, K.A., Stover, S.M., Pascoe, J.R., Amundson, R., 2004. Tooth enamel biomineraliza-tion in extant horses: implications for isotopic microsampling. Palaeogeography,Palaeoclimatology, Palaeoecology 206, 355–365.
Janis, C., 1976. The evolutionary strategy of the equidae and the origins of rumen andcecal digestion. Evolution 30, 757–774.
Jans, M.M.E., Nielsen-Marsh, C.M., Smith, C.I., Collins, M.J., Kars, H., 2004. Characterisa-tion of microbial attack on archaeological bone. Journal of Archaeological Science31, 87–95.
Jones, A.M., O'connell, T.C., Young, E.D., Scott, K., Buckingham, C.M., Iacumin, P., Brasier,M.D., 2001. Biogeochemical data from well preserved 200 ka collagen and skeletalremains. Earth and Planetary Science Letters 193, 143–149.
Keppel, G., Wickens, T.D., 2004. Design and Analysis: A Researcher's Handbook. PearsonPrentice Hall, Upper Saddle River.
Koch, P.L., Heisinger, J., Moss, C., Carlson, R.W., Fogel, M.L., Behrensmeyer, A.K., 1995.Isotopic tracking of change in diet and habitat use in African elephants. Science267, 1340–1343.
Koch, P.L., Hoppe, K.A., Webb, D.S., 1998. The isotopic ecology of late Pleistocene mam-mals in North America. Part 1: Florida. Chemical Geology 152, 119–138.
Kohn, M.J., 1999. You are what you eat. Science 283, 335–336.Krysl, L.J., Hubbert, M.E., Sowell, B.F., Plumb, G.E., Jewett, T.K., Smith, M.A., Waggoner,
J.W., 1984. Horses and cattle grazing in the Wyoming Red Desert, I. Food habitsand dietary overlap. Journal of Range Management 31, 72–76.
Laurat, T., Brühl, E., 2006. Zum Stand der archäologischen Untersuchungen im TagebauNeumark-Nord, Ldkr. Merseberg-Querfurt (Sachsen-Anhalt) - Vorbericht zu denAusgrabungen 2003–2005: Jahresschrift für Mitteldeutsche Vorgeschichte, 90,pp. 9–69.
Lee-Thorp, J., Van Der Merwe, N.J., 1987. Carbon isotope analysis of fossil bone apatite.South African Journal of Science 83, 712–715.
Livingston, H.G., Payne, W.J.A., Friends, M.T., 1962. Urea extretion in ruminants. Nature194, 1057–1058.
Longin, R., 1971. New method of collagen extraction for radiocarbon dating. Nature230, 241–242.
Macfadden, B.J., 2008. Geographic variation in diets of ancient populations of 5-million-year-old (early Pliocene) horses from southern North America. Palaeogeo-graphy, Palaeoclimatology, Palaeoecology 266, 83–94.
Maguas, C., Brugnoli, E., 1996. Spatial variation in carbon isotope discrimintation acrossthe thalli of several lichen species. Plant, Cell & Environment 19, 437–446.
Mania, D., Mania, U., 2008. La stratigraphie et le Paléolithique du complexe saalien dansla région de la Saale et de l'Elbe. L'Anthropologie 112, 15–47.
Mania, D., Thomae, M., Litt, T., Weber, T., 1990. Neumark—Gröbern: Beiträge zur Jagddes mittelpaläolithischen Menschen. Veröffentlichungen des Landesmuseum fürVorgeschichte in Halle 43.
Menard, C., Duncan, P., Fleurance, G., Georges, J.Y., Lila, M., 2002. Comparative foragingand nutrition of horses and cattle in European wetlands. Journal of Applied Ecology39, 120–133.
Menke, B., Tynni, R., 1984. Das Eeminterglazial unddasWeichselfrühglazial von Rederstall/Dittmarschen und ihre Bedeutung für die mitteleuropäische Jungpleistozänglieder-ung. Geoogisches Jahrbuch A76, 3.
Nadelhoffer, K.J., Fry, B., 1994. Nitrogen isotope studies in forest ecosystems. In: Lajtha,K., Michener, R.H. (Eds.), Stable isotopes in ecology and environmental science.Blackwell, Oxford. pp.
Nadelhoffer, K., Shaver, G., Fry, B., Giblin, A., Johnson, L., Mckane, R., 1996. 15N naturalabundances and N use by tundra plants. Oecologia 107, 386–394.
Nielsen-Marsh, C.M., Hedges, R.E.M., 2000. Patterns of diagenesis in bone I: The effectsof site environments. Journal of Archaeological Science 27, 1139–1150.
Nielsen-Marsh, C.M., Gernaey, A., Turner-Walker, G., Hedges, R.E.M., Pike, A., Collins,M., 2000. The chemical degradation of bone. In: Cox, M., Mays, S. (Eds.), HumanOsteology in Archaology and Forensic Science. GMM, London, pp. 439–454.
Nielsen-Marsh, C.M., Smith, C.I., Jans, M., Nord, A., Kars, H., Collins, M.J., 2007. Bonediagenesis in the European Holocene II: taphonomic and environmental consider-ations. Journal of Archaeological Science 34, 1523–1531.
Noe-Nygaard, N., Price, T.D., Hede, S.U., 2005. Diet of aurochs and early cattle in southernScandinavia: evidence from 15N and 13C stable isotopes. Journal of ArchaeologicalScience 32, 855–871.
Park, R., Epstein, S., 1960. Carbon isotope fractionation during photosynthesis. Geochi-mica et Cosmochimica Acta 21, 110–126.
Pellegrini, M., Donahue, R.E., Chenery, C., Evans, J., Lee-Thorp, J., Montgomery, J., Mussi,M., 2008. Faunal migration in late-glacial central Italy: implications for human re-source exploitation. Rapid Communications in Mass Spectrometry 22, 1714–1726.
Richards, M.P., Hedges, R.E.M., 2003. Variations in bone collagen delta C-13 and deltaN-15 values of fauna from Northwest Europe over the last 40 000 years. Palaeogeo-graphy, Palaeoclimatology, Palaeoecology 193, 261–267.

177K. Britton et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 333-334 (2012) 168–177
Richards, M.P., Pettitt, P.B., Trinkaus, E., Smith, F.H., Paunovic, M., Karavanic, I., 2000.Neanderthal diet at Vindija and Neanderthal predation: the evidence from stableisotopes. Proceedings of the National Academy of Sciences of the United States ofAmerica 97, 7663–7666.
Richards, M.P., Taylor, G., Steele, T., Mcpherron, S.P., Soressi, M., Jaubert, J., Orschiedt, J.,Mallye, J.B., Rendu, W., Hublin, J.J., 2008. Isotopic dietary analysis of a Neanderthaland associated fauna from the site of Jonzac (Charente-Maritime), France. Journalof Human Evolution 55, 179–185.
Ryden, H., 1972. America's Last Wild Horses. Ballantine Books, New York.Schaller, G.B., 1998. Wildlife of the Tibetan Steppe. University of Chicago Press, Chicago.Schmidt-Nielsen, B., 1958. Urea excretion in mammals. Physiological Reviews 38,
139–168.Schoeninger, M.J., Deniro, M.J., 1984. Nitrogen and carbon isotopic composition of bone
collagen from marine and terrestrial animals. Geochimica et Cosmochimica Acta48, 625–639.
Schwarcz, H.P., Dupras, T.L., Fairgrieve, S.I., 1999. 15N enrichment in the Sahara: insearch of a global relationship. Journal of Archaeological Science 26, 629–636.
Schwertl, M., Auerswald, K., Schäufele, R., Schnyder, H., 2005. Carbon and nitrogenstable isotope composition of cattle hair: ecological fingerprints of productionsystems? Agriculture, Ecosystems and Environment 109, 153–165.
Shearer, G., Kohl, D.H., 1986. N2-fixation in field settings: estimations based on natural15N abundance. Australian Journal of Plant Physiology 13, 699–757.
Sier, M.J., Roebroecks, W., Bakels, C.C., Dekkers, M.J., Bruhl, E., De Loecker, D.,Gaudzinski-Windheuser, S., Hesse, N., Jagich, A., Kindler, L., Kuijper, W.J., Laurat,T., Mücher, H.J., Penkman, K.E., Richter, D., Van Hinsbergen, D.J., 2011. Directterrestrial-marine correlation demonstrates suprisingly late onset of the last inter-glacial in Central and Northwestern Europe. Quaternary Research 75, 213–218.
Slade, L.M., Robinson, D.W., Casey, K.E., 1970. Nitrogen metabolism in non ruminantherbivores. 1—The influence of non protein nitrogen and protein quality on the ni-trogen retention of adult mares. Journal of Animal Science 30, 753–760.
Smith, B.N., Epstein, S., 1970. Biogeochemistry of the stable isotopes of hydrogen andcarbon in salt marsh biota. Plant Physiology 46, 738–742.
Smith, C.I., Nielsen-Marsh, C.M., Jans, M.M.E., Collins, M.J., 2007. Bone diagenesis in theEuropean Holocene I: patterns and mechanisms. Journal of Archaeological Science34, 1485–1493.
Smuts, G.L., 1978. Interrelations between predators, prey, and their environment. Bio-science 28, 316–320.
Sponheimer, M., Robinson, T., Ayliffe, L., Roeder, B., Hammer, J., Passey, B., West, A.,Cerling, T., Dearing, D., Ehleringer, J., 2003. Nitrogen isotopes in mammalian herbi-vores: hair δ15N values from a controlled feeding study. International Journal ofOsteoarchaeology 13, 80–87.
Steele, K.W., Daniel, R.M., 1978. Fractionation of nitrogen isotopes by animals: a furthercomplication to the use of variations in the natural abundance of 15N for tracerstudies. Journal of Agricultural Science 90, 7–9.
Stevens, R.E., Hedges, R.E.M., 2004. Carbon and nitrogen stable isotope analysis ofnorthwest European horse bone and tooth collagen, 40,000 BP-present: Palaeocli-matic interpretations. Quaternary Science Reviews 23, 977–991.
Stevens, R.E., Jacobi, R., Street, M., Germonpre, M., Conard, N.J., Munzel, S.C., Hedges,R.E.M., 2008. Nitrogen isotope analyses of reindeer (Rangifer tarandus), 45,000 BPto 900 BP: palaeoenvironmental reconstructions. Palaeogeography, Palaeoclima-tology, Palaeoecology 262, 32–45.
Strahl, J., Krbetschek, M.R., Luckert, J., Machalett, B., Meng, S., Oches, E.A., Rappsilber, I.,Wansa, S., Zöller, L., 2010. Geologie, Paläontologie und Geochronologie des Eem-Beckens Neumark-Nord 2 und Vergleichmit dem BeckenNeumark-Nord 1 (Geiseltal,Sachsen-Anhalt). E&G Quaternary Science Journal 59, 120–167.
Szpak, P., Gröcke, D.R., Debruyne, R., Macphee, R.D.E., Guthrie, R.D., Froese, D., Zazula,G.D., Patterson, W.P., Poinar, H.N., 2010. Regional differences in bone collagenδ13C and δ15N of Pleistocene mammoths: implications for paleoecology of themammoth steppe. Palaeogeography, Palaeoclimatology, Palaeoecology 286, 88–96.
Teeri, J.A., 1981. Stable carbon isotope analysis of mosses and lichens growing in xericand moist habitats. The Bryologist 84, 82–84.
Van Der Merwe, N.J., Medina, E., 1991. The canopy effect, carbon isotope ratios andfoodwebs in Amazonia. Journal of Archaeological Science 18, 249–259.
Van Groenigen, J.-W., Van Kessel, C., 2002. Salinity-induced patterns of natural abun-dance carbon-13 and nitrogen-15 in plant and soil. Soil Science Society of AmericaJournal 66, 489–498.
Van Klinken, G.J., 1999. Bone collagen quality indicators for palaeodietary and radiocar-bon measurements. Journal of Archaeological Science 26, 687–695.
Van Klinken, G.J., Richards, M.P., Hedges, R.E.M., 2000. An overview of causes for stableisotopic variations in past European human populations: environmental, ecophys-iological, and cultural effects. In: Ambrose, S., Katzenberg, A. (Eds.), Biogeochemi-cal Approaches to Palaeodietary Analysis. Kluwer Academic/Plenum publishers,New York, pp. 39–63.
Van Leeuwen, R.J., Beets, D., Bosch, J.H.A., Burger, A.W., Cleveringa, P., Van Harten, D.,Herngreen, G.F.W., Langereis, C.G., Meijer, T., Pouwer, R., De Wolf, H., 2000. Stratig-raphy and integrated facies analysis of the Eemian in the Amsterdam-Terminalborehole, the Netherlands. In: Van Kolfschoten, T., Gibbard, P.L. (Eds.), TheEemian—local sequences, global perspectives: Geologie en Mijnbouw/NetherlandsJournal of Geosciences, 79 (2/3), pp. 161–196.
Virginia, R.A., Delwiche, C.C., 1982. Natural 15N abundance of presumed N2-fixing andnon-N2-fixing plants from selected ecosystems. Oecologia (Berlin) 54, 317–325.
Widga, C., Walker, J.D., Stockli, L.D., 2010. Middle Holocene Bison diet and mobility inthe eastern Great Plains (USA) based on δ13C, δ18O, and 87Sr/86Sr analyses oftooth enamel carbonate. Quaternary Research 73, 449–463.
Zagwijn, W.H., 1961. Vegetation, climate and radiocarbon datings in the Late Pleisto-cene of The Netherlands, Part I: Eemian and Early Weichselian. Mededelingen Geo-logische Stichting 14, 15–45.





![Palaeolithic and Mesolithic research in the central Balkans [2014]](https://static.fdokumen.com/doc/165x107/63333a85ce61be0ae50e8b63/palaeolithic-and-mesolithic-research-in-the-central-balkans-2014.jpg)














