
A Novel Autotransporter Adhesin Is Required for Efficient Colonization during Bubonic Plague
Upload
uni-wuerzburgCategory
view
2download
0
ARTICLE IN PRESS
1438-4221/$ - se
doi:10.1016/j.ijm
Abbreviations
cells.�CorrespondE-mail addr
(T.A. Oelschlae
International Journal of Medical Microbiology 299 (2009) 467–478
www.elsevier.de/ijmm
Split immune response after oral vaccination of mice with recombinant
Escherichia coli Nissle 1917 expressing fimbrial adhesin K88
Katharina A. Remera, Marita Bartrowa, Bianca Roegera, Heidrun Molla,Ulrich Sonnenbornb, Tobias A. Oelschlaegera,�
aUniversity of Wurzburg, Institute for Molecular Infection Biology, Rontgenring 11, 97070 Wurzburg, GermanybArdeypharm GmbH, Div. Biological Research, Herdecke, Germany
Received 3 February 2009; received in revised form 6 March 2009; accepted 15 March 2009
Abstract
Enterotoxigenic Escherichia coli (ETEC) are a leading cause of diarrhoea in piglets and newborn calves. Massiveefforts have therefore been made to develop a vaccine for the induction of protective mucosal immunity against ETEC.Since it has been shown that the probiotic strain E. coli Nissle 1917 (EcN) can serve as a safe carrier for targeteddelivery of recombinant molecules to the intestinal mucosa, we constructed the recombinant strain EcN pMut2-kanK88 (EcN-K88) stably expressing the determinant for the K88 fimbrial adhesin of ETEC on the bacterial surface.
After oral application of EcN-K88 to mice for one week, EcN-K88 as well as wild-type EcN and EcN mock-transformed with the plasmid vector only could be detected in faecal samples for a minimum of 7 days after the lastfeeding, indicating that EcN can transiently colonise the murine intestine. Oral application of EcN-K88 resulted insignificant IgG serum titres against K88 as early as 7 days after the initial feeding with EcN-K88, but no significant IgAtitres. In contrast, we failed to detect any specific T cell responses towards the K88 antigen both in spleen andmesenteric lymph nodes. Although dendritic cells readily upregulated maturation and activation markers in responseto K88 stimulation, accompanied by secretion of interleukin (IL)-12, IL-6, IL-10, and tumour necrosis factor,restimulation of T cells from mice having received EcN-K88 with K88-loaded dendritic cells did not result in detectableT cell proliferation and IL-2 secretion, but rather induced an IL-10 bias. While the serum antibody responses clearlydemonstrate that K88 is recognized by the humoral immune system, our findings indicate that oral application ofprobiotic EcN expressing the K88 fimbrial adhesin does not induce a selective T cell response towards the antigen.r 2009 Elsevier GmbH. All rights reserved.
Keywords: Oral vaccination; Mucosal immunity; E. coli Nissle 1917; Split tolerance; T helper cell response
e front matter r 2009 Elsevier GmbH. All rights reserved.
m.2009.03.003
: EcN, Escherichia coli Nissle 1917; DC, Dendritic
ing author. Tel.: +49931 312150; fax: +49931 312578.
ess: [email protected]
ger).
Introduction
Probiotic bacteria are live, non-pathogenic microor-ganisms which are beneficial for the health of the host.One of these, the Escherichia coli strain Nissle 1917(EcN) (serotype O6:K5:H1), has been used since the1920s as a probiotic agent for the treatment of infectiousand chronic inflammatory diseases of the intestine

ARTICLE IN PRESSK.A. Remer et al. / International Journal of Medical Microbiology 299 (2009) 467–478468
(Lodinova-Zadnikova and Sonnenborn, 1997; Nissle,1918, 1919). E. coli Nissle 1917 is a non-pathogenicfaecal isolate which has been characterized extensively(Blum et al., 1995; Westendorf et al., 2005a). Thegenome has recently been sequenced and was shown tocontain many fitness factors that support competitive-ness and a very effective colonisation especially of thelarge intestine of the host (Grozdanov et al., 2004;Lodinova-Zadnikova et al., 1998; Sun et al., 2005). Weand others could show that E. coli Nissle 1917 exerts itsprobiotic effects in various ways by blocking adherenceand invasion of pathogenic strains, regulation of centralT cell functions including gd T cells and modulation ofcytokine responses as well as induction of the anti-microbial peptide b-defensin 2 in human mucosalepithelium cells. The later seems to be a commonmechanism of probiotic bacteria, which EcN shares withseveral other strains (Altenhofer et al., 2004; Boudeauet al., 2003; Guzy et al., 2008; Mondel et al., 2009; Schleeet al., 2007, 2008; Sturm et al., 2005). Furthermore, ithas been demonstrated that genetically modified E. coli
Nissle 1917 can serve as a safe carrier for site-directeddelivery of antigen to the intestine due to its excellentcolonisation properties and apathogenicity even underconditions of active inflammation or in mono-associatedgnotobiotic animals (Gunzer et al., 2002; Schultz et al.,2004, 2005; Waidmann et al., 2003; Westendorf et al.,2005b).
On the other side of the wide spectrum of E. coli
reside the enterotoxigenic E. coli (ETEC) strains. ETEC-associated diarrhoea causes morbidity and mortality inneonatal and weaning piglets, the porcine post-weaningdiarrhoea syndrome, which is one of the economicallymost important diseases in swine husbandry (Chenet al., 2004; Frydendahl, 2002; Gyles, 1994; Sojka et al.,1960). ETEC express on their surface long, hair-likeproteinaceous appendages, the fimbriae, for attachmentto the gut epithelial receptors. In porcine post-weaningdiarrhea F4 (K88) is the most prevalent type of fimbrialadhesins of ETEC (Berberov et al., 2004; Moon, 1978;Smith and Linggood, 1971; Zhang et al., 2007). K88 is akey virulence factor by mediating attachment of thebacteria to the F4 receptor on the brush borders ofenterocytes in the epithelium of the small intestine, thusfacilitating the initial colonisation of the gastrointestinaltract. Subsequently, various enterotoxins are produced,which disrupt the intestinal fluid homoeostasis andcause fluid hypersecretion together with electrolyteimbalance, resulting in massive diarrhoea (Bakkeret al., 1992; Jones and Rutter, 1972; Van den Broecket al., 2000).
Since K88 fimbriae are major colonizing factors inETEC-induced diarrhoea, they constitute a primarytarget for vaccination against ETEC (Dougan andMorrissey, 1985). Fimbriae-based parenteral vaccina-tion of sows has been demonstrated to protect suckling
piglets via passive lactogenic immunity. However, in thepost-weaning period, piglets need to acquire an activeintestinal mucosal immune response with high levels ofK88-specific IgA to be further protected (Bianchi et al.,1996; Verdonck et al., 2004). Several reports havedemonstrated that this cannot be achieved via parent-eral immunisation stimulating mainly a systemic but nota mucosal immune response (Bianchi et al., 1996; Vanden Broeck et al., 1999; Van der Stede et al., 2003).Therefore, several attempts have been made in the lastyears to induce a robust mucosal immunity against K88by various methods of oral vaccination (Attridge et al.,1988; Morona et al., 1994). The most promising resultshave been achieved with attenuated Salmonella strainsgenetically modified to express the whole K88 genecluster or only the FaeG subunit of K88. However, todate no Salmonella-based vaccine against K88 has beengranted licence for use in veterinary medicine (Lee et al.,2001).
In contrast, E. coli Nissle 1917 has been approved as abiotherapeutic agent that is safe for the use in humans aswell as domestic animals even under conditions of acuteinflammation. The aim of this study was, therefore, toexplore the immunogenic potential of recombinantE. coli Nissle 1917 expressing the K88 fimbrial antigenas a safe vaccine for oral vaccination against the majorETEC virulence factor K88 fimbrial adhesin.
Materials and methods
Bacteria and oral vaccination of mice
For oral vaccination of mice, a spontaneous strepto-mycin-resistant mutant of E. coli Nissle 1917 (EcN;DSM6601, serotype O6:K5:H1, originally providedfrom Ardeypharm GmbH, Herdecke, Germany) wascured of its 2 cryptic plasmids pMUT1and pMUT2(EcN-wt) resulting in EcNc. By inserting a kanamycinresistance cassette in the pMUT2 plasmid, the respectivecontrol strain EcNc pMUT2-Kn (EcN-mock) wascreated. In this plasmid vector, the determinant forexpression of the K88 fimbrial adhesin from enterotoxi-genic E. coli was inserted in the singular EcoRIrestriction site to create EcNc pMUT2-KnK88(EcN-K88) with both the kanamycin resistance cassetteand K88 expression. Surface expression of the K88adhesin on EcN-K88 was confirmed by serum aggluti-nation of fresh bacteria suspensions with polyclonalrabbit anti-K88 serum (a generous gift fromDr. W. Schrodl, University of Leipzig, Germany).
For oral application, EcN was grown overnight inLuria-Bertani broth (LB, BD Microbiology, Heidelberg,Germany). To determine the exact EcN titres, colony-forming units (CFU) were determined by colony

ARTICLE IN PRESSK.A. Remer et al. / International Journal of Medical Microbiology 299 (2009) 467–478 469
counting after plating 1:107 dilutions of the respectiveEcN overnight cultures on LB agar with 50 mg/mlstreptomycin. Bacteria were washed in saline and dilutedto a titre of 33� 108CFU/ml in sodium phosphatebuffer, pH 7.0.
Female wild-type BALB/c mice at 6 weeks of age wereobtained from Charles River Breeding Laboratories(Sulzfeld, Germany). Housing and handling of theanimals followed the guidelines of the animal welfarecommittee of the German government. All animalexperiments were approved by the government of LowerFranconia. For oral vaccination, mice were individuallyfed by pipette with 108CFU/day in 30 ml of sodiumphosphate buffer for 7 days. To support the gutcolonisation with EcN by reducing competing gut flora,mice received 5mg/ml streptomycin in the drinkingwater, starting 2 days prior to the beginning of EcNfeeding, for the whole feeding period.
CFU in faeces and tissue samples
To monitor the colonisation of the gut by EcN, freshfaeces from individual animals were collected prior tothe beginning of the EcN feeding and then on days 1 and7 of each feeding period, 2 weeks after the end of thefirst feeding period, and at the end of the experiment,one week after the last feeding period. For thelocalisation of EcN after oral application, mice werefed for 3 days with 108CFU/day, as described above,and mesenteric lymph nodes, Peyer’s patches, ilealmucosa plus adjacent mucus, and faeces from thecaecum were collected one day after the last feeding.
Faeces and tissue samples were immediately weighted,diluted 1:10 in sterile saline, and either plated directlyon LB agar with 50 mg/ml streptomycin or 30 mg/ml ofkanamycin for the lymph nodes and Peyer’s patches, orfurther diluted in saline to a final dilution of 1:104 forthe mucosa samples and 1:107 for the faecal samples.Where Salmonella typhimurium SL1344 smr was used asa control, MacConkey agar (BD Microbiology) with50 mg/ml streptomycin was used. Colonies were countedafter 24 h and CFU calculated per g of faeces or tissue.
Heat extraction of K88 fimbrial adhesin
K88 fimbrial adhesin was prepared from E. coli
HB101 pMF205 ampr. After overnight culture in LBbroth with 75 mg/ml ampicillin, bacteria were collectedby centrifugation. The pellet was resuspended in 0.5mMTris-HCl buffer, pH 7.4, and incubated for 30min at60 1C. Supernatant containing K88 was separated frombacterial debris by ultracentrifugation at 12,000 rpm for30min at 4 1C and the fimbrial adhesin in the super-natant further concentrated with a Centricon YM-10filter (Millipore GmbH, Eschborn, Germany) for 40min
at 5000� g and 15 1C. Protein concentration wasdetermined by the method of Bradford, and the finalconcentration of purified K88 was adjusted to 300 mg/mlin phosphate buffered saline (PBS).
Preparation of primary cells
Naıve T cells were isolated from the spleens ofBALB/c mice one week after the last EcN feeding bymagnetic bead separation according to the manufac-turer’s instructions with a Pan T Cell Isolation Kit(Miltenyi Biotec, Bergisch Gladbach, Germany) andresuspended to a density of 2� 106 cells/ml in RPMI1640 medium (GIBCO Invitrogen, Karlsruhe,Germany), supplemented with 10% heat-inactivatedfoetal calf serum (FCS), 2mM L-glutamine, 10mMHEPES buffer, 60 mg/ml penicillin, and 20 mg/ml genta-mycin for proliferation assays.
Murine dendritic cells (DC) were generated from bonemarrow progenitors following the protocol of Lutz et al.(1999). Briefly, bone marrow cells were cultured inRPMI 1640 medium in the presence of 200U/mlrecombinant murine granulocyte-macrophage colony-stimulating factor (GM-CSF) (PeproTech, London,UK). Fresh GM-CSF was added to the cultures ondays 3 and 6. After 10 days, non-adherent cells werecollected and used for further experimentation. Thesecells were shown to have typical myeloid DC morphol-ogy (Ramırez-Pineda et al., 2004).
T cell proliferation after restimulation with K88
To examine the induction of a specific T cell responseagainst K88, DC generated from the bone marrow ofBALB/c mice were loaded overnight with K88pili (20 mg/ml) and were additionally stimulated with25 mg/ml CpG oligodeoxynucleotides (ODN) 1668(50-TCCATGACGTTCCTGATGCT-30) which wereobtained from Qiagen Operon (Cologne, Germany)and were not phosphorothioate-modified. Seven daysafter the last feeding with EcN-wt, EcN-mock, orEcN-K88, animals were sacrificed, and spleens wereharvested aseptically for T cell isolation by magneticbead depletion. T cells of the respective treatmentgroups were plated at a density of 2� 106 cells/ml inRPMI medium in 96-well microtitre plates and were leftunstimulated or were restimulated with K88-loaded,CpG ODN-stimulated DC at decreasing concentrationsranging from 2� 105 to 2� 102 cells/ml. After 24 h, thedye Alamar Blue (Trinova Biochem, Gießen, Germany)was added, and cell proliferation was measured color-imetrically after a total incubation time of 72 h in anenzyme-linked immunosorbent assay (ELISA) readerwith Multiskan Ascent software (Thermo ElectronCorporation, Dreieich, Germany). Culture supernatants

ARTICLE IN PRESSK.A. Remer et al. / International Journal of Medical Microbiology 299 (2009) 467–478470
were stored at �80 1C until additional analysis of IL-2secretion was performed by ELISA.
Analysis of cytokine production and antibody titres
Cytokine levels in the culture supernatants weremeasured by sandwich ELISA as described previously(Flohe et al., 1998). Briefly, microtitre plates were coatedwith purified rat antibodies directed against murineIL-2, IL-6, IL-10, IL-12, or TNF, followed by overnightincubation with the culture supernatants and serialdilutions of recombinant cytokines for the generation ofstandard curves. After washing, bound cytokines weredetected by incubation with the appropriate biotin-conjugated rat anticytokine antibodies (all antibodypairs and recombinant cytokine standards from BDPharmingen, Heidelberg, Germany), followed by addi-tion of streptavidin-alkaline phosphatase complex (BDPharmingen). After washing, the enzyme activity wasvisualized using the substrate p-nitrophenylphosphate(Sigma-ALDRICH Chemie GmbH, Taufkirchen, Ger-many). Samples were set up in duplicates, and theoptical densities (OD) were measured in an ELISAreader (Thermo Electron Corporation). Cytokine detec-tion thresholds were 39 pg/ml for IL-2 and IL-6,20 pg/ml for IL-10 and IL-12, and 78 pg/ml for TNF.
The levels of K88-specific IgG, IgM, and IgAantibodies in the sera of mice were analyzed by ELISAone day after the last oral application of EcN. Briefly,wells of a 96-well plate (MaxiSorb, Nalgen NuncInternational, Wiesbaden, Germany) were coated withpurified K88 pili (100 ng/well) in PBS at 4 1C overnight.After blocking unspecific binding sites with 1% BSA inPBS for 4 h at room temperature, the antigen-coatedwells were probed for 2 h at 4 1C with serum samplesdiluted 1:500 for IgG and IgM or 1:100 for IgA in PBSwith 1% bovine serum albumin (BSA). Bound K88-specific antibodies were detected with biotinylatedmonoclonal antibodies against mouse IgG (1:5000, fromSigma-ALDRICH Chemie GmbH), mouse IgM ormouse IgA (both 1:250, BD Pharmingen) dilutedin PBS with 1% BSA overnight at 4 1C, followed byalkaline phosphatase-conjugated streptavidin (1:1000 inPBS, BD Pharmingen). For the detection of IgA in thefaeces of mice 7 days after the last oral EcN application,96-well plates (MaxiSorb, Nalgen Nunc International)were coated with 100 ng/well purified K88 pili diluted in0.1M NaHCO3-buffer, pH 8.3, overnight at 4 1C. Toblock unspecific binding, wells were incubated with 5%BSA in PBS for 4 h at room temperature, followed byprobing with the faeces samples diluted 1:50 in PBS with1% BSA for 1 h at room temperature. After binding ofalkaline phosphatase-conjugated streptavidin (1:1000 inPBS, BD Pharmingen) to the secondary antibodies,
substrate was added, and the enzyme activity wasmeasured using an ELISA reader as described above.
Activation of DC by K88 and EcN
To examine the activation of DC by K88 in vitro,immature DC at a density of 1� 106 cells/ml in RPMI1640 medium were cultured in the presence of purifiedK88 pili (20 mg/ml). CpG ODN (25 mg/ml) was used as apositive control, mock-treated cultures received PBSonly. For stimulation of DC with whole EcN-wt,EcN-mock and EcN-K88 after overnight culture in LBbroth, bacteria were washed in PBS, resuspended to adensity of 100mg wet weight/ml in PBS, and heat-inactivated for 1h at 65 1C. DC were then cultured with20mg/ml of the respective EcN preparations. After 48h,cells were harvested for FACS analysis and supernatantswere collected and stored at �80 1C for analysis ofcytokine secretion by ELISA. To rule out unspecificactivation of DC by LPS contamination of the K88preparation, DC were generated from the bone marrow ofTLR4�/� C3H/HeJ mice as described above.
To examine the activation und upregulation ofcostimulatory molecules on DC by EcN and K88, DCwere cultured together with purified K88 pili (20 mg/ml)or heat-inactivated whole EcN wt, EcN mock or EcN-K88 (20 mg/ml). CpG ODN (25 mg/ml) and LPS(1 mg/ml, from E. coli 026:B26, Sigma-ALDRICHChemie GmbH) were used for positive controls. After24 h of culture, cells were washed and fixed with 4%paraformaldehyde in PBS, pH 7.4, followed by labellingwith fluorochrome-conjugated antibodies against majorhistocompatibility complex (MHC) class II molecules,CD80, and CD86 (all from BD Pharmingen). For dataanalysis, CellQuest software (BD Biosciences) was used.
Statistical analysis
Data were analyzed using the GraphPad Prism 4.00software. For determining statistical significance of theELISA experiments, multiple group comparisons wereperformed by non-parametric one-way analysis ofvariance (Kruskal-Wallis test).
Results
Gut colonisation after oral application of EcN
Oral application of 108CFU of EcN-wt, EcN-mock,or EcN-K88 per day to BALB/c mice pretreated withstreptomycin via the drinking water resulted in pro-longed colonisation of the gut in all treatment groups.Already one day after the first application, up to107CFU EcN were present in the faeces, increasing to
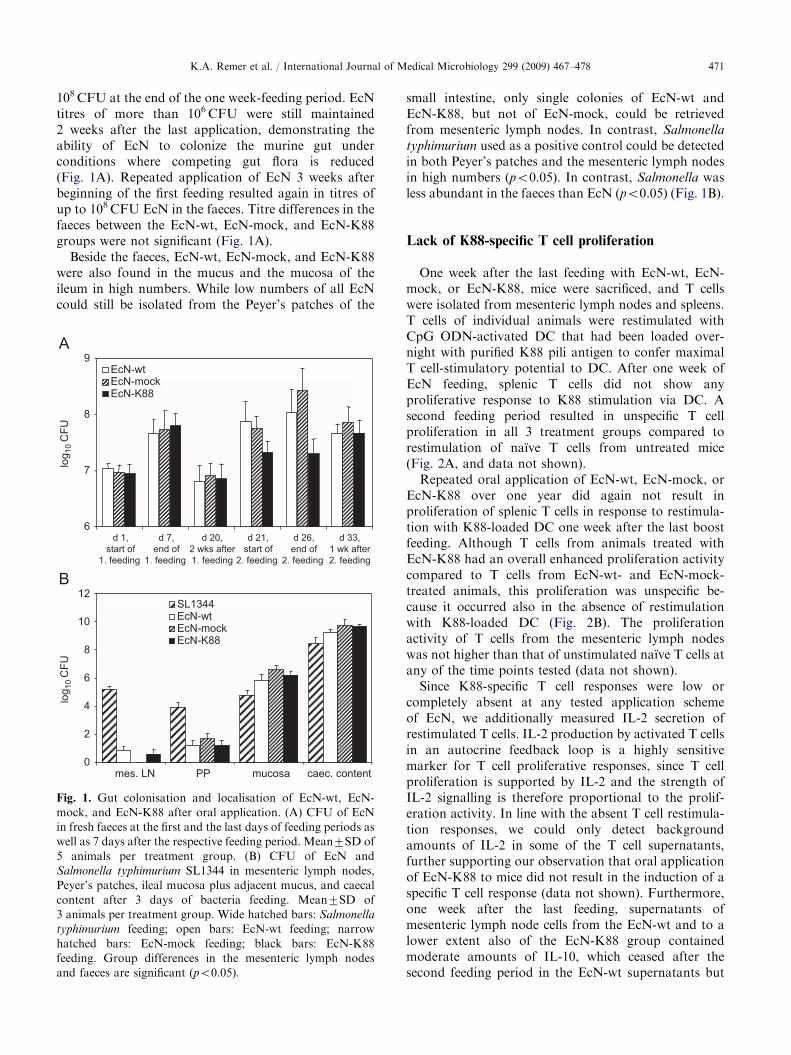
ARTICLE IN PRESSK.A. Remer et al. / International Journal of Medical Microbiology 299 (2009) 467–478 471
108CFU at the end of the one week-feeding period. EcNtitres of more than 106CFU were still maintained2 weeks after the last application, demonstrating theability of EcN to colonize the murine gut underconditions where competing gut flora is reduced(Fig. 1A). Repeated application of EcN 3 weeks afterbeginning of the first feeding resulted again in titres ofup to 108CFU EcN in the faeces. Titre differences in thefaeces between the EcN-wt, EcN-mock, and EcN-K88groups were not significant (Fig. 1A).
Beside the faeces, EcN-wt, EcN-mock, and EcN-K88were also found in the mucus and the mucosa of theileum in high numbers. While low numbers of all EcNcould still be isolated from the Peyer’s patches of the
6
7
8
9
d 1,start of
1. feeding
d 7,end of
1. feeding
d 20,2 wks after1. feeding
d 21,start of
2. feeding
d 26,end of
2. feeding
d 33,1 wk after2. feeding
log 1
0 C
FU
EcN-wtEcN-mockEcN-K88
mes. LN
SL1344EcN-wtEcN-mockEcN-K88
log 1
0 C
FU
0
2
4
6
8
10
12
PP mucosa caec. content
Fig. 1. Gut colonisation and localisation of EcN-wt, EcN-
mock, and EcN-K88 after oral application. (A) CFU of EcN
in fresh faeces at the first and the last days of feeding periods as
well as 7 days after the respective feeding period. Mean7SD of
5 animals per treatment group. (B) CFU of EcN and
Salmonella typhimurium SL1344 in mesenteric lymph nodes,
Peyer’s patches, ileal mucosa plus adjacent mucus, and caecal
content after 3 days of bacteria feeding. Mean7SD of
3 animals per treatment group. Wide hatched bars: Salmonella
typhimurium feeding; open bars: EcN-wt feeding; narrow
hatched bars: EcN-mock feeding; black bars: EcN-K88
feeding. Group differences in the mesenteric lymph nodes
and faeces are significant (po0.05).
small intestine, only single colonies of EcN-wt andEcN-K88, but not of EcN-mock, could be retrievedfrom mesenteric lymph nodes. In contrast, Salmonella
typhimurium used as a positive control could be detectedin both Peyer’s patches and the mesenteric lymph nodesin high numbers (po0.05). In contrast, Salmonella wasless abundant in the faeces than EcN (po0.05) (Fig. 1B).
Lack of K88-specific T cell proliferation
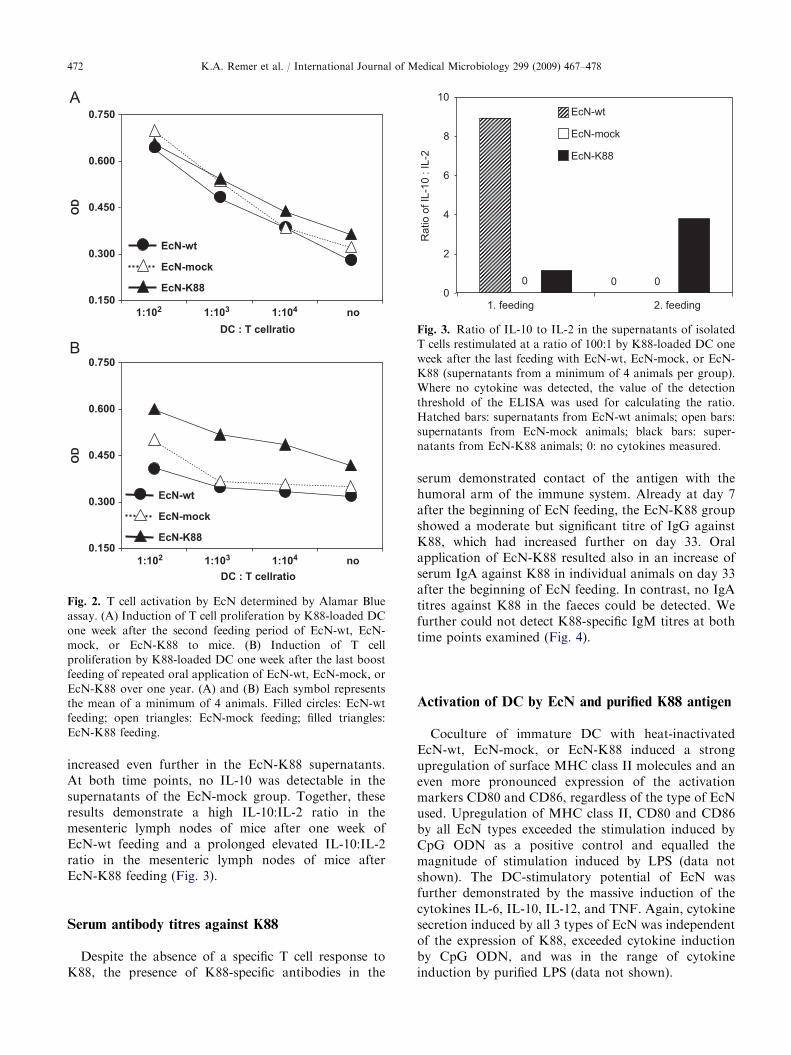
One week after the last feeding with EcN-wt, EcN-mock, or EcN-K88, mice were sacrificed, and T cellswere isolated from mesenteric lymph nodes and spleens.T cells of individual animals were restimulated withCpG ODN-activated DC that had been loaded over-night with purified K88 pili antigen to confer maximalT cell-stimulatory potential to DC. After one week ofEcN feeding, splenic T cells did not show anyproliferative response to K88 stimulation via DC. Asecond feeding period resulted in unspecific T cellproliferation in all 3 treatment groups compared torestimulation of naıve T cells from untreated mice(Fig. 2A, and data not shown).
Repeated oral application of EcN-wt, EcN-mock, orEcN-K88 over one year did again not result inproliferation of splenic T cells in response to restimula-tion with K88-loaded DC one week after the last boostfeeding. Although T cells from animals treated withEcN-K88 had an overall enhanced proliferation activitycompared to T cells from EcN-wt- and EcN-mock-treated animals, this proliferation was unspecific be-cause it occurred also in the absence of restimulationwith K88-loaded DC (Fig. 2B). The proliferationactivity of T cells from the mesenteric lymph nodeswas not higher than that of unstimulated naıve T cells atany of the time points tested (data not shown).
Since K88-specific T cell responses were low orcompletely absent at any tested application schemeof EcN, we additionally measured IL-2 secretion ofrestimulated T cells. IL-2 production by activated T cellsin an autocrine feedback loop is a highly sensitivemarker for T cell proliferative responses, since T cellproliferation is supported by IL-2 and the strength ofIL-2 signalling is therefore proportional to the prolif-eration activity. In line with the absent T cell restimula-tion responses, we could only detect backgroundamounts of IL-2 in some of the T cell supernatants,further supporting our observation that oral applicationof EcN-K88 to mice did not result in the induction of aspecific T cell response (data not shown). Furthermore,one week after the last feeding, supernatants ofmesenteric lymph node cells from the EcN-wt and to alower extent also of the EcN-K88 group containedmoderate amounts of IL-10, which ceased after thesecond feeding period in the EcN-wt supernatants but
ARTICLE IN PRESS
0.150
0.300
0.450
0.600
0.750
1:102 1:103 1:104 noDC : T cellratio
OD
EcN-wt
EcN-mock
EcN-K88
1:102 1:103 1:104 noDC : T cellratio
0.150
0.300
0.450
0.600
0.750
OD
EcN-wt
EcN-mock
EcN-K88
Fig. 2. T cell activation by EcN determined by Alamar Blue
assay. (A) Induction of T cell proliferation by K88-loaded DC
one week after the second feeding period of EcN-wt, EcN-
mock, or EcN-K88 to mice. (B) Induction of T cell
proliferation by K88-loaded DC one week after the last boost
feeding of repeated oral application of EcN-wt, EcN-mock, or
EcN-K88 over one year. (A) and (B) Each symbol represents
the mean of a minimum of 4 animals. Filled circles: EcN-wt
feeding; open triangles: EcN-mock feeding; filled triangles:
EcN-K88 feeding.
0
2
4
6
8
10
1. feeding
Rat
io o
f IL-
10 :
IL-2
EcN-wt
EcN-mock
EcN-K88
0 00
2. feeding
Fig. 3. Ratio of IL-10 to IL-2 in the supernatants of isolated
T cells restimulated at a ratio of 100:1 by K88-loaded DC one
week after the last feeding with EcN-wt, EcN-mock, or EcN-
K88 (supernatants from a minimum of 4 animals per group).
Where no cytokine was detected, the value of the detection
threshold of the ELISA was used for calculating the ratio.
Hatched bars: supernatants from EcN-wt animals; open bars:
supernatants from EcN-mock animals; black bars: super-
natants from EcN-K88 animals; 0: no cytokines measured.
K.A. Remer et al. / International Journal of Medical Microbiology 299 (2009) 467–478472
increased even further in the EcN-K88 supernatants.At both time points, no IL-10 was detectable in thesupernatants of the EcN-mock group. Together, theseresults demonstrate a high IL-10:IL-2 ratio in themesenteric lymph nodes of mice after one week ofEcN-wt feeding and a prolonged elevated IL-10:IL-2ratio in the mesenteric lymph nodes of mice afterEcN-K88 feeding (Fig. 3).
Serum antibody titres against K88
Despite the absence of a specific T cell response toK88, the presence of K88-specific antibodies in the
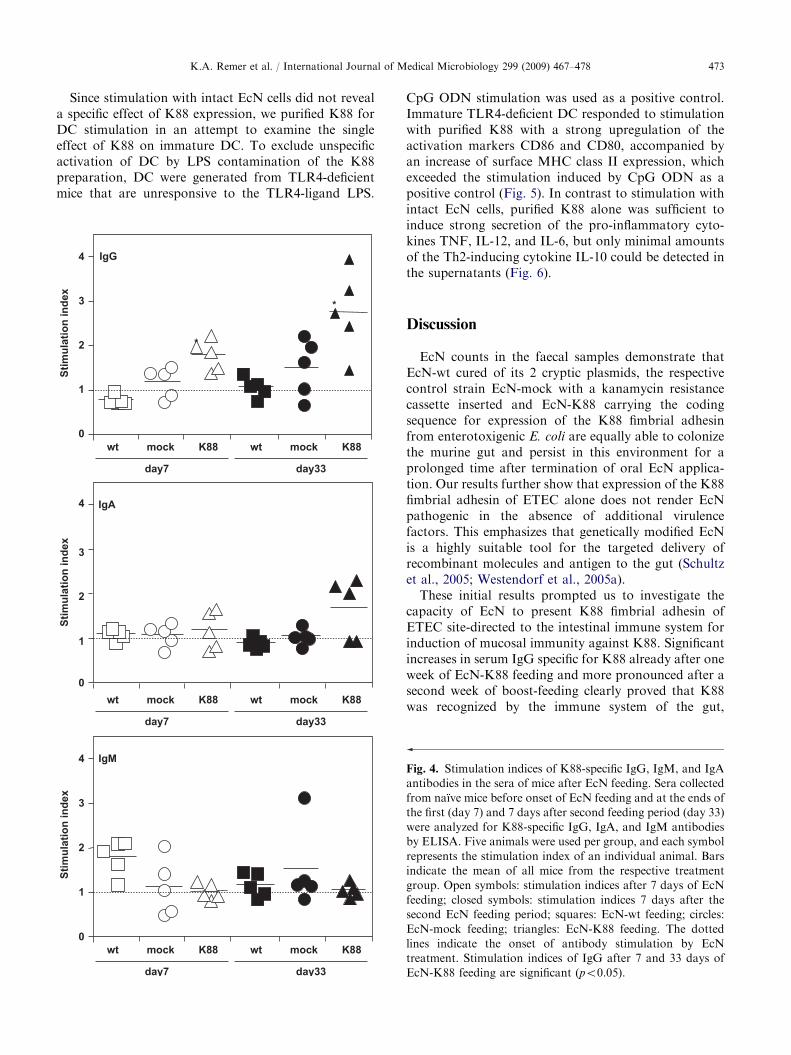
serum demonstrated contact of the antigen with thehumoral arm of the immune system. Already at day 7after the beginning of EcN feeding, the EcN-K88 groupshowed a moderate but significant titre of IgG againstK88, which had increased further on day 33. Oralapplication of EcN-K88 resulted also in an increase ofserum IgA against K88 in individual animals on day 33after the beginning of EcN feeding. In contrast, no IgAtitres against K88 in the faeces could be detected. Wefurther could not detect K88-specific IgM titres at bothtime points examined (Fig. 4).
Activation of DC by EcN and purified K88 antigen
Coculture of immature DC with heat-inactivatedEcN-wt, EcN-mock, or EcN-K88 induced a strongupregulation of surface MHC class II molecules and aneven more pronounced expression of the activationmarkers CD80 and CD86, regardless of the type of EcNused. Upregulation of MHC class II, CD80 and CD86by all EcN types exceeded the stimulation induced byCpG ODN as a positive control and equalled themagnitude of stimulation induced by LPS (data notshown). The DC-stimulatory potential of EcN wasfurther demonstrated by the massive induction of thecytokines IL-6, IL-10, IL-12, and TNF. Again, cytokinesecretion induced by all 3 types of EcN was independentof the expression of K88, exceeded cytokine inductionby CpG ODN, and was in the range of cytokineinduction by purified LPS (data not shown).
ARTICLE IN PRESSK.A. Remer et al. / International Journal of Medical Microbiology 299 (2009) 467–478 473
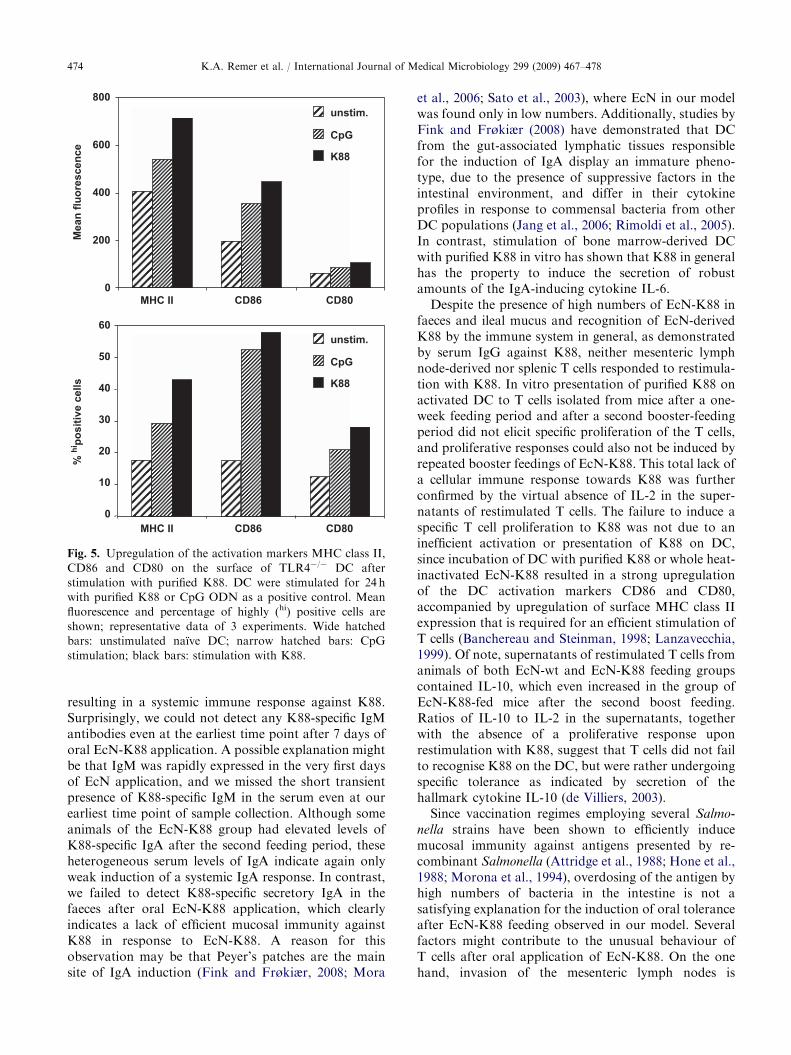
Since stimulation with intact EcN cells did not reveala specific effect of K88 expression, we purified K88 forDC stimulation in an attempt to examine the singleeffect of K88 on immature DC. To exclude unspecificactivation of DC by LPS contamination of the K88preparation, DC were generated from TLR4-deficientmice that are unresponsive to the TLR4-ligand LPS.
IgM
0
1
2
3
4
Stim
ulat
ion
inde
x
wt K88 wt mock K880
1
2
3
4
Stim
ulat
ion
inde
x
IgG
*
*
day7
0
1
2
3
4
Stim
ulat
ion
inde
x
IgA
mock
day33
wt K88 wt mock K88
day7
mock
day33
wt K88 wt mock K88
day7
mock
day33
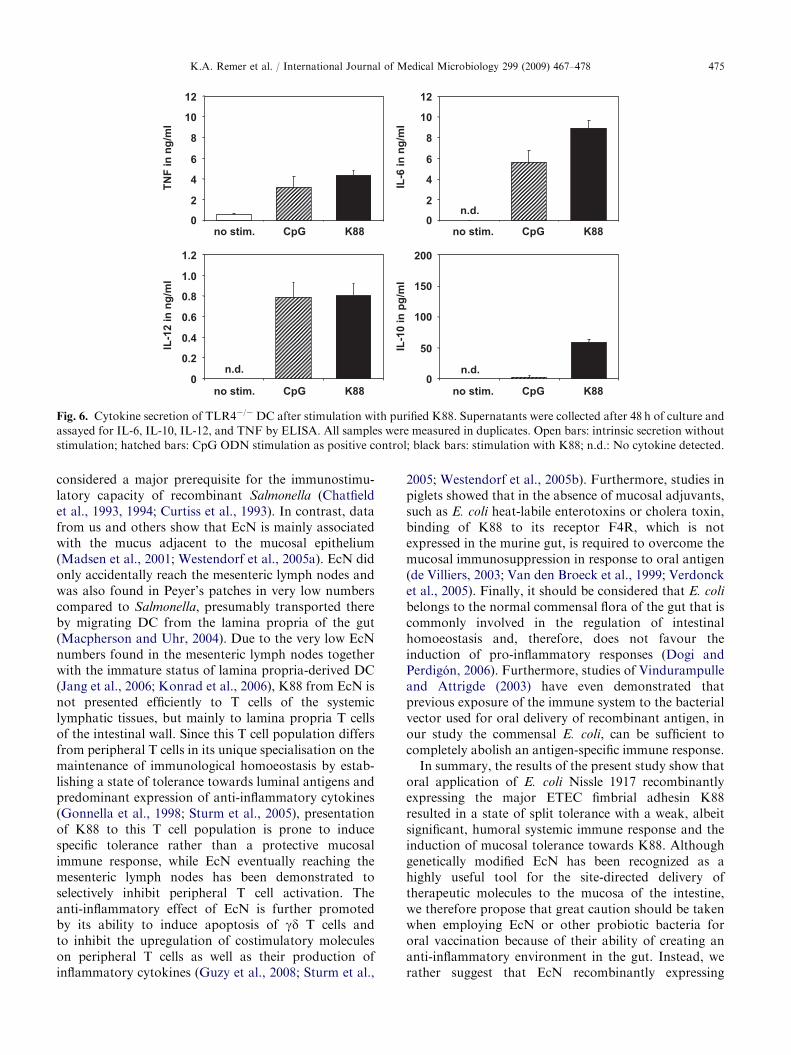
CpG ODN stimulation was used as a positive control.Immature TLR4-deficient DC responded to stimulationwith purified K88 with a strong upregulation of theactivation markers CD86 and CD80, accompanied byan increase of surface MHC class II expression, whichexceeded the stimulation induced by CpG ODN as apositive control (Fig. 5). In contrast to stimulation withintact EcN cells, purified K88 alone was sufficient toinduce strong secretion of the pro-inflammatory cyto-kines TNF, IL-12, and IL-6, but only minimal amountsof the Th2-inducing cytokine IL-10 could be detected inthe supernatants (Fig. 6).
Discussion
EcN counts in the faecal samples demonstrate thatEcN-wt cured of its 2 cryptic plasmids, the respectivecontrol strain EcN-mock with a kanamycin resistancecassette inserted and EcN-K88 carrying the codingsequence for expression of the K88 fimbrial adhesinfrom enterotoxigenic E. coli are equally able to colonizethe murine gut and persist in this environment for aprolonged time after termination of oral EcN applica-tion. Our results further show that expression of the K88fimbrial adhesin of ETEC alone does not render EcNpathogenic in the absence of additional virulencefactors. This emphasizes that genetically modified EcNis a highly suitable tool for the targeted delivery ofrecombinant molecules and antigen to the gut (Schultzet al., 2005; Westendorf et al., 2005a).
These initial results prompted us to investigate thecapacity of EcN to present K88 fimbrial adhesin ofETEC site-directed to the intestinal immune system forinduction of mucosal immunity against K88. Significantincreases in serum IgG specific for K88 already after oneweek of EcN-K88 feeding and more pronounced after asecond week of boost-feeding clearly proved that K88was recognized by the immune system of the gut,
Fig. 4. Stimulation indices of K88-specific IgG, IgM, and IgA
antibodies in the sera of mice after EcN feeding. Sera collected
from naıve mice before onset of EcN feeding and at the ends of
the first (day 7) and 7 days after second feeding period (day 33)
were analyzed for K88-specific IgG, IgA, and IgM antibodies
by ELISA. Five animals were used per group, and each symbol
represents the stimulation index of an individual animal. Bars
indicate the mean of all mice from the respective treatment
group. Open symbols: stimulation indices after 7 days of EcN
feeding; closed symbols: stimulation indices 7 days after the
second EcN feeding period; squares: EcN-wt feeding; circles:
EcN-mock feeding; triangles: EcN-K88 feeding. The dotted
lines indicate the onset of antibody stimulation by EcN
treatment. Stimulation indices of IgG after 7 and 33 days of
EcN-K88 feeding are significant (po0.05).
ARTICLE IN PRESS
0
200
400
600
800
MHC II
Mea
n flu
ores
cenc
e
unstim.
CpG
K88
0
10
20
30
40
50
60
% hi
posi
tive
cells
unstim.
CpG
K88
CD86 CD80
MHC II CD86 CD80
Fig. 5. Upregulation of the activation markers MHC class II,
CD86 and CD80 on the surface of TLR4�/� DC after
stimulation with purified K88. DC were stimulated for 24 h
with purified K88 or CpG ODN as a positive control. Mean
fluorescence and percentage of highly (hi) positive cells are
shown; representative data of 3 experiments. Wide hatched
bars: unstimulated naıve DC; narrow hatched bars: CpG
stimulation; black bars: stimulation with K88.
K.A. Remer et al. / International Journal of Medical Microbiology 299 (2009) 467–478474
resulting in a systemic immune response against K88.Surprisingly, we could not detect any K88-specific IgMantibodies even at the earliest time point after 7 days oforal EcN-K88 application. A possible explanation mightbe that IgM was rapidly expressed in the very first daysof EcN application, and we missed the short transientpresence of K88-specific IgM in the serum even at ourearliest time point of sample collection. Although someanimals of the EcN-K88 group had elevated levels ofK88-specific IgA after the second feeding period, theseheterogeneous serum levels of IgA indicate again onlyweak induction of a systemic IgA response. In contrast,we failed to detect K88-specific secretory IgA in thefaeces after oral EcN-K88 application, which clearlyindicates a lack of efficient mucosal immunity againstK88 in response to EcN-K88. A reason for thisobservation may be that Peyer’s patches are the mainsite of IgA induction (Fink and Frøkiær, 2008; Mora
et al., 2006; Sato et al., 2003), where EcN in our modelwas found only in low numbers. Additionally, studies byFink and Frøkiær (2008) have demonstrated that DCfrom the gut-associated lymphatic tissues responsiblefor the induction of IgA display an immature pheno-type, due to the presence of suppressive factors in theintestinal environment, and differ in their cytokineprofiles in response to commensal bacteria from otherDC populations (Jang et al., 2006; Rimoldi et al., 2005).In contrast, stimulation of bone marrow-derived DCwith purified K88 in vitro has shown that K88 in generalhas the property to induce the secretion of robustamounts of the IgA-inducing cytokine IL-6.
Despite the presence of high numbers of EcN-K88 infaeces and ileal mucus and recognition of EcN-derivedK88 by the immune system in general, as demonstratedby serum IgG against K88, neither mesenteric lymphnode-derived nor splenic T cells responded to restimula-tion with K88. In vitro presentation of purified K88 onactivated DC to T cells isolated from mice after a one-week feeding period and after a second booster-feedingperiod did not elicit specific proliferation of the T cells,and proliferative responses could also not be induced byrepeated booster feedings of EcN-K88. This total lack ofa cellular immune response towards K88 was furtherconfirmed by the virtual absence of IL-2 in the super-natants of restimulated T cells. The failure to induce aspecific T cell proliferation to K88 was not due to aninefficient activation or presentation of K88 on DC,since incubation of DC with purified K88 or whole heat-inactivated EcN-K88 resulted in a strong upregulationof the DC activation markers CD86 and CD80,accompanied by upregulation of surface MHC class IIexpression that is required for an efficient stimulation ofT cells (Banchereau and Steinman, 1998; Lanzavecchia,1999). Of note, supernatants of restimulated T cells fromanimals of both EcN-wt and EcN-K88 feeding groupscontained IL-10, which even increased in the group ofEcN-K88-fed mice after the second boost feeding.Ratios of IL-10 to IL-2 in the supernatants, togetherwith the absence of a proliferative response uponrestimulation with K88, suggest that T cells did not failto recognise K88 on the DC, but were rather undergoingspecific tolerance as indicated by secretion of thehallmark cytokine IL-10 (de Villiers, 2003).
Since vaccination regimes employing several Salmo-
nella strains have been shown to efficiently inducemucosal immunity against antigens presented by re-combinant Salmonella (Attridge et al., 1988; Hone et al.,1988; Morona et al., 1994), overdosing of the antigen byhigh numbers of bacteria in the intestine is not asatisfying explanation for the induction of oral toleranceafter EcN-K88 feeding observed in our model. Severalfactors might contribute to the unusual behaviour ofT cells after oral application of EcN-K88. On the onehand, invasion of the mesenteric lymph nodes is
ARTICLE IN PRESS
0
2
4
6
8
10
12
IL-6
in n
g/m
l
TNF
in n
g/m
l
no stim.
IL-1
2 in
ng/
ml
0
2
4
6
8
10
12
0
0.2
0.4
0.6
0.8
1.0
1.2
0
50
100
150
200
IL-1
0 in
pg/
ml
n.d.n.d.
n.d.
CpG K88 no stim. CpG K88
no stim. CpG K88 no stim. CpG K88
Fig. 6. Cytokine secretion of TLR4�/�DC after stimulation with purified K88. Supernatants were collected after 48 h of culture and
assayed for IL-6, IL-10, IL-12, and TNF by ELISA. All samples were measured in duplicates. Open bars: intrinsic secretion without
stimulation; hatched bars: CpG ODN stimulation as positive control; black bars: stimulation with K88; n.d.: No cytokine detected.
K.A. Remer et al. / International Journal of Medical Microbiology 299 (2009) 467–478 475
considered a major prerequisite for the immunostimu-latory capacity of recombinant Salmonella (Chatfieldet al., 1993, 1994; Curtiss et al., 1993). In contrast, datafrom us and others show that EcN is mainly associatedwith the mucus adjacent to the mucosal epithelium(Madsen et al., 2001; Westendorf et al., 2005a). EcN didonly accidentally reach the mesenteric lymph nodes andwas also found in Peyer’s patches in very low numberscompared to Salmonella, presumably transported thereby migrating DC from the lamina propria of the gut(Macpherson and Uhr, 2004). Due to the very low EcNnumbers found in the mesenteric lymph nodes togetherwith the immature status of lamina propria-derived DC(Jang et al., 2006; Konrad et al., 2006), K88 from EcN isnot presented efficiently to T cells of the systemiclymphatic tissues, but mainly to lamina propria T cellsof the intestinal wall. Since this T cell population differsfrom peripheral T cells in its unique specialisation on themaintenance of immunological homoeostasis by estab-lishing a state of tolerance towards luminal antigens andpredominant expression of anti-inflammatory cytokines(Gonnella et al., 1998; Sturm et al., 2005), presentationof K88 to this T cell population is prone to inducespecific tolerance rather than a protective mucosalimmune response, while EcN eventually reaching themesenteric lymph nodes has been demonstrated toselectively inhibit peripheral T cell activation. Theanti-inflammatory effect of EcN is further promotedby its ability to induce apoptosis of gd T cells andto inhibit the upregulation of costimulatory moleculeson peripheral T cells as well as their production ofinflammatory cytokines (Guzy et al., 2008; Sturm et al.,
2005; Westendorf et al., 2005b). Furthermore, studies inpiglets showed that in the absence of mucosal adjuvants,such as E. coli heat-labile enterotoxins or cholera toxin,binding of K88 to its receptor F4R, which is notexpressed in the murine gut, is required to overcome themucosal immunosuppression in response to oral antigen(de Villiers, 2003; Van den Broeck et al., 1999; Verdoncket al., 2005). Finally, it should be considered that E. coli
belongs to the normal commensal flora of the gut that iscommonly involved in the regulation of intestinalhomoeostasis and, therefore, does not favour theinduction of pro-inflammatory responses (Dogi andPerdigon, 2006). Furthermore, studies of Vindurampulleand Attrigde (2003) have even demonstrated thatprevious exposure of the immune system to the bacterialvector used for oral delivery of recombinant antigen, inour study the commensal E. coli, can be sufficient tocompletely abolish an antigen-specific immune response.
In summary, the results of the present study show thatoral application of E. coli Nissle 1917 recombinantlyexpressing the major ETEC fimbrial adhesin K88resulted in a state of split tolerance with a weak, albeitsignificant, humoral systemic immune response and theinduction of mucosal tolerance towards K88. Althoughgenetically modified EcN has been recognized as ahighly useful tool for the site-directed delivery oftherapeutic molecules to the mucosa of the intestine,we therefore propose that great caution should be takenwhen employing EcN or other probiotic bacteria fororal vaccination because of their ability of creating ananti-inflammatory environment in the gut. Instead, werather suggest that EcN recombinantly expressing

ARTICLE IN PRESSK.A. Remer et al. / International Journal of Medical Microbiology 299 (2009) 467–478476
selected antigens might provide a highly interesting toolfor ‘negative vaccination’, the selective induction of oraltolerance in cases of food allergy.
Acknowledgements
This work was supported by grant no. MO418/9-3by the Deutsche Forschungsgemeinschaft (DFG),Germany, to H.M. T.A.O. was in part supportedby Pharma-Zentrale GmbH, Herdecke, Germany. Theauthors thank C. de Witt for her excellent care of theanimals.
References
Altenhofer, A., Oswald, S., Sonnenborn, U., Enders, C.,
Schulze, J., Hacker, J., Oelschlaeger, T.A., 2004. The
probiotic Escherichia coli strain Nissle 1917 interferes with
invasion of human intestinal epithelial cells by different
enteroinvasive bacterial pathogens. FEMS Immunol. Med.
Microbiol. 40, 223–229.
Attridge, S., Hackett, J., Morona, R., Whyte, P., 1988.
Towards a live oral vaccine against enterotoxigenic
Escherichia coli of swine. Vaccine 6, 387–389.
Bakker, D., Willemsen, P.T., Simons, L.H., van Zijderveld,
F.G., de Graaf, F.K., 1992. Characterization of the
antigenic and adhesive properties of FaeG, the major
subunit of K88 fimbriae. Mol. Microbiol. 6, 247–255.
Banchereau, J., Steinman, R.M., 1998. Dendritic cells and the
control of immunity. Nature 392, 245–252.
Berberov, E.M., Zhou, Y., Francis, D.H., Scott, M.A.,
Kachman, S.D., Moxley, R.A., 2004. Relative importance
of heat-labile enterotoxin in the causation of severe
diarrheal disease in the gnotobiotic piglet model by a strain
of enterotoxigenic Escherichia coli that produces multiple
enterotoxins. Infect. Immun. 72, 3914–3924.
Bianchi, A.T., Scholten, J.W., van Zijderveld, A.M., van
Zijderveld, F.G., Bokhout, B.A., 1996. Parenteral vaccina-
tion of mice and piglets with F4+ Escherichia coli
suppresses the enteric anti-F4 response upon oral infection.
Vaccine 14, 199–206.
Blum, G., Marre, R., Hacker, J., 1995. Properties of
Escherichia coli strains of serotype O6. Infection 23,
234–236.
Boudeau, J., Glasser, A.L., Julien, S., Colombel, J.F.,
Darfeuille-Michaud, A., 2003. Inhibitory effect of probiotic
Escherichia coli strain Nissle 1917 on adhesion to and
invasion of intestinal epithelial cells by adherent-invasive
E. coli strains isolated from patients with Crohn’s disease.
Aliment. Pharmacol. Ther. 18, 45–56.
Chatfield, S., Roberts, M., Li, J., Starns, A., Dougan, G.,
1994. The use of live attenuated Salmonella for oral
vaccination. Dev. Biol. Stand. 82, 35–42.
Chatfield, S., Roberts, M., Londono, P., Cropley, I., Douce,
G., Dougan, G., 1993. The development of oral vaccines
based on live attenuated Salmonella strains. FEMS
Immunol. Med. Microbiol. 7, 1–7.
Chen, X., Gao, S., Jiao, X., Liu, X.F., 2004. Prevalence of
serogroups and virulence factors of Escherichia coli strains
isolated from pigs with postweaning diarrhoea in eastern
China. Vet. Microbiol. 103, 13–20.
Curtiss III, R., Kelly, S.M., Hassan, J.O., 1993. Live oral
avirulent Salmonella vaccines. Vet. Microbiol. 37, 397–405.
de Villiers, W.J., 2003. Crohn’s disease and IL-10 therapy:
promise regained. Inflamm. Bowel Dis. 9, 210–211.
Dogi, C.A., Perdigon, G., 2006. Importance of the host
specificity in the selection of probiotic bacteria. J. Dairy
Res. 73, 357–366.
Dougan, G., Morrissey, P., 1985. Molecular analysis of the
virulence determinants of enterotoxigenic Escherichia coli
isolated from domestic animals: applications for vaccine
development. Vet. Microbiol. 10, 241–257.
Fink, L.N., Frøkiær, H., 2008. Dendritic cells from Peyer’s
patches and mesenteric lymph nodes differ from spleen
dendritic cells in their response to commensal gut bacteria.
Scand. J. Immunol. 68, 270–279.
Flohe, S.B., Bauer, C., Flohe, S., Moll, H., 1998. Antigen-
pulsed epidermal Langerhans cells protect susceptible mice
from infection with the intracellular parasite Leishmania
major. Eur. J. Immunol. 28, 3800–3811.
Frydendahl, K., 2002. Prevalence of serogroups and virulence
genes in Escherichia coli associated with postweaning
diarrhoea and edema disease in pigs and a comparison of
diagnostic approaches. Vet. Microbiol. 85, 169–182.
Gonnella, P.A., Chen, Y., Inobe, J., Komagata, Y., Quartulli,
M., Weiner, H.L., 1998. In situ immune response in gut-
associated lymphoid tissue (GALT) following oral antigen
in TCR-transgenic mice. J. Immunol. 160, 4708–4718.
Grozdanov, L., Raasch, C., Schulze, J., Sonnenborn, U.,
Gottschalk, G., Hacker, J., Dobrindt, U., 2004. Analysis of
the genome structure of the nonpathogenic probiotic
Escherichia coli strain Nissle 1917. J. Bacteriol. 186,
5432–5441.
Gunzer, F., Hennig-Pauka, I., Waldmann, K.H., Sandhoff, R.,
Grone, H.J., Kreipe, H.H., Matussek, A., Mengel, M.,
2002. Gnotobiotic piglets develop thrombotic microangio-
pathy after oral infection with enterohemorrhagic Esche-
richia coli. Am. J. Clin. Pathol. 118, 364–375.
Guzy, C., Paclik, D., Schirbel, A., Sonnenborn, U., Wieden-
mann, B., Sturm, A., 2008. The probiotic Escherichia coli
strain Nissle 1917 induces gd T cell apoptosis via caspase-
and FasL-dependent pathways. Int. Immunol. 20, 829–840.
Gyles, C.L., 1994. Escherichia coli in domestic animals and
humans. CAB International. Wallingford, Oxford, UK.
Hone, D., Attridge, S., van den Bosch, L., Hackett, J., 1988.
A chromosomal integration system for stabilization of
heterologous genes in Salmonella based vaccine strains.
Microb. Pathog. 5, 407–418.
Jang, M.H., Sougawa, N., Tanaka, T., Hirata, T., Hiroi, T.,
Tohya, K., Guo, Z., Umemoto, E., Ebisuno, Y., Yang,
B.G., Seoh, J.Y., Lipp, M., Kiyono, H., Miyasaka, M.,
2006. CCR7 is critically important for migration of
dendritic cells in intestinal lamina propria to mesenteric
lymph nodes. J. Immunol. 176, 803–810.
Jones, G.W., Rutter, J.M., 1972. Role of the K88 antigen in
the pathogenesis of neonatal diarrhea caused by Escherichia
coli in piglets. Infect. Immun. 6, 918–927.

ARTICLE IN PRESSK.A. Remer et al. / International Journal of Medical Microbiology 299 (2009) 467–478 477
Konrad, A., Cong, Y., Duck, W., Borlaza, R., Elson, C.O.,
2006. Tight mucosal compartmentation of the murine
immune response to antigens of the enteric microbiota.
Gastroenterology 130, 2050–2059.
Lanzavecchia, A., 1999. Dendritic cell maturation and
generation of immune responses. Haematologica 84 (Suppl.
EHA-4), 23–25.
Lee, E., Platt, R., Kang, S., Roth, J.A., Phillips, G.J., 2001.
Chromosomal integration and expression of the Escherichia
coli K88 gene cluster in Salmonella enterica ser. Chole-
raesuis strain 54 (SC54). Vet. Microbiol. 83, 177–183.
Lodinova-Zadnikova, R., Sonnenborn, U., 1997. Effect of
preventive administration of a nonpathogenic Escherichia
coli strain on the colonization of the intestine with
microbial pathogens in newborn infants. Biol. Neonate
71, 224–232.
Lodinova-Zadnikova, R., Sonnenborn, U., Tlaskalova, H.,
1998. Probiotics and E. coli infections in man. Vet. Q. 20
(Suppl. 3), 78–81.
Lutz, M.B., Kukutsch, N., Ogilvie, A.L., Rossner, S., Koch,
F., Romani, N., Schuler, G., 1999. An advanced culture
method for generating large quantities of highly pure
dendritic cells from mouse bone marrow. J. Immunol.
Methods 223, 77–92.
Macpherson, A.J., Uhr, T., 2004. Induction of protective IgA
by intestinal dendritic cells carrying commensal bacteria.
Science 303, 1662–1665.
Madsen, K., Cornish, A., Soper, P., McKaigney, C., Jijon, H.,
Yachimec, C., Doyle, J., Jewell, L., De Simone, C.,
2001. Probiotic bacteria enhance murine and human
intestinal epithelial barrier function. Gastroenterology 121,
580–591.
Mondel, M., Schroeder, B.O., Zimmermann, K., Huber, H.,
Nuding, S., Beisner, J., Fellermann, K., Stange, E.F.,
Wehkamp, J., 2009. Probiotic E. coli treatment mediates
antimicrobial human beta-defensin synthesis and fecal
excretion in humans. Mucosal Immunol. 2, 166–172.
Moon, H.W., 1978. Mechanisms in the pathogenesis of
diarrhea: a review. J. Am. Vet. Med. Assoc. 172, 443–448.
Mora, J.R., Iwata, M., Eksteen, B., Song, S.Y., Junt, T.,
Senman, B., Otipoby, K.L., Yokota, A., Takeuchi, H.,
Ricciardi-Castagnoli, P., Rajewsky, K., Adams, D.H., von
Andrian, U.H., 2006. Generation of gut-homing IgA-
secreting B cells by intestinal dendritic cells. Science 314,
1157–1160.
Morona, R., Morona, J.K., Considine, A., Hackett, J.A., van
den Bosch, L., Beyer, L., Attridge, S.R., 1994. Construction
of K88- and K99-expressing clones of Salmonella typhimu-
rium G30: immunogenicity following oral administration to
pigs. Vaccine 12, 513–517.
Nissle, A., 1918. Die antagonistische Behandlung chronischer
Darmstorungen mit Colibakterien. Med. Klin. 2, 29–30.
Nissle, A., 1919. Weiteres uber die Mutaflorbehandlung unter
besonderer Berucksichtigung der chronischen Ruhr.
Munch. Med. Wochenschr. 25, 678–681.
Ramırez-Pineda, J.R., Frohlich, A., Berberich, C., Moll, H.,
2004. Dendritic cells (DC) activated by CpG DNA ex vivo
are potent inducers of host resistance to an intracellular
pathogen that is independent of IL-12 derived from the
immunizing DC. J. Immunol. 172, 6281–6289.
Rimoldi, M., Chieppa, M., Salucci, V., Avogadri, F.,
Sonzogni, A., Sampietro, G.M., Nespoli, A., Viale, G.,
Allavena, P., Rescigno, M., 2005. Intestinal immune
homeostasis is regulated by the crosstalk between epithelial
cells and dendritic cells. Nat. Immunol. 6, 507–514.
Sato, A., Hashiguchi, M., Toda, E., Iwasaki, A., Hachimura,
S., Kaminogawa, S., 2003. CD11b+ Peyer’s patch dendritic
cells secrete IL-6 and induce IgA secretion from naive B
cells. J. Immunol. 171, 3684–3690.
Schlee, M., Wehkamp, J., Altenhofer, A., Oelschlaeger, T.A.,
Stange, E.F., Fellermann, K., 2007. Induction of human
beta-defensin 2 by the probiotic Escherichia coli Nissle 1917
is mediated through flagellin. Infect. Immun. 75,
2399–2407.
Schlee, M., Harder, J., Koten, B., Stange, E.F., Wehkamp, J.,
Fellermann, K., 2008. Probiotic lactobacilli and VSL#3
induce enterocyte beta-defensin 2. Clin. Exp. Immunol.
151, 528–535.
Schultz, M., Strauch, U.G., Linde, H.J., Watzl, S., Obermeier,
F., Gottl, C., Dunger, N., Grunwald, N., Scholmerich, J.,
Rath, H.C., 2004. Preventive effects of Escherichia coli
strain Nissle 1917 on acute and chronic intestinal inflam-
mation in two different murine models of colitis. Clin.
Diagn. Lab. Immunol. 11, 372–378.
Schultz, M., Watzl, S., Oelschlaeger, T.A., Rath, H.C., Gottl,C., Lehn, N., Scholmerich, J., Linde, H.J., 2005. Green
fluorescent protein for detection of the probiotic micro-
organism Escherichia coli strain Nissle 1917 (EcN) in vivo.
J. Microbiol. Methods 61, 389–398.
Smith, H.W., Linggood, M.A., 1971. Observations on the
pathogenic properties of the K88, Hly and Ent plasmids of
Escherichia coli with particular reference to porcine
diarrhoea. J. Med. Microbiol. 4, 467–485.
Sojka, W.J., Lloyd, M.K., Sweeney, E.J., 1960. Escherichia coli
serotypes associated with certain pig diseases. Res. Vet. Sci.
1, 17–27.
Sturm, A., Rilling, K., Baumgart, D.C., Gargas, K., Abou-
Ghazale, T., Raupach, B., Eckert, J., Schumann, R.R.,
Enders, C., Sonnenborn, U., Wiedenmann, B., Dignass,
A.U., 2005. Escherichia coli Nissle 1917 distinctively
modulates T-cell cycling and expansion via toll-like
receptor 2 signaling. Infect. Immun. 73, 1452–1465.
Sun, J., Gunzer, F., Westendorf, A.M., Buer, J., Scharfe, M.,
Jarek, M., Gossling, F., Blocker, H., Zeng, A.P., 2005.
Genomic peculiarity of coding sequences and metabolic
potential of probiotic Escherichia coli strain Nissle 1917
inferred from raw genome data. J. Biotechnol. 117,
147–161.
Van den Broeck, W., Cox, E., Goddeeris, B.M., 1999.
Receptor-dependent immune responses in pigs after oral
immunization with F4 fimbriae. Infect. Immun. 67,
520–526.
Van den Broeck, W., Cox, E., Oudega, B., Goddeeris, B.M.,
2000. The F4 fimbrial antigen of Escherichia coli and its
receptors. Vet. Microbiol. 71, 223–244.
Van der Stede, Y., Cox, E., Verdonck, F., Vancaeneghem, S.,
Goddeeris, B.M., 2003. Reduced faecal excretion of F4+
-E. coli by the intramuscular immunisation of suckling
piglets by the addition of 1alpha, 25-dihydroxyvitamin D3
or CpG-oligodeoxynucleotides. Vaccine 21, 1023–1032.

ARTICLE IN PRESSK.A. Remer et al. / International Journal of Medical Microbiology 299 (2009) 467–478478
Verdonck, F., Cox, E., Van der Stede, Y., Goddeeris, B.M.,
2004. Oral immunization of piglets with recombinant F4
fimbrial adhesin FaeG monomers induces a mucosal and
systemic F4-specific immune response. Vaccine 22,
4291–4299.
Verdonck, F., Snoeck, V., Goddeeris, B.M., Cox, E., 2005.
Cholera toxin improves the F4(K88)-specific immune
response following oral immunization of pigs with
recombinant FaeG. Vet. Immunol. Immunopathol. 103,
21–29.
Vindurampulle, C.J., Attrigde, S.R., 2003. Impact of vector
priming on the immunogenicity of recombinant Salmonella
vaccines. Infect. Immun. 71, 287–297.
Waidmann, M., Bechtold, O., Frick, J.S., Lehr, H.A.,
Schubert, S., Dobrindt, U., Loeffler, J., Bohn, E.,
Autenrieth, I.B., 2003. Bacteroides vulgatus protects against
Escherichia coli-induced colitis in gnotobiotic interleukin-
2-deficient mice. Gastroenterology 125, 162–177.
Westendorf, A.M., Gunzer, F., Deppenmeier, S., Tapadar, D.,
Hunger, J.K., Schmidt, M.A., Buer, J., Bruder, D., 2005a.
Intestinal immunity of Escherichia coli Nissle 1917: a safe
carrier for therapeutic molecules. FEMS Immunol. Med.
Microbiol. 43, 373–384.
Westendorf, A.M., Templin, M., Geffers, R., Deppenmeier, S.,
Gruber, A.D., Probst-Kepper, M., Hansen, W., Liblau,
R.S., Gunzer, F., Bruder, D., Buer, J., 2005b. CD4+ T cell
mediated intestinal immunity: chronic inflammation versus
immune regulation. Gut 54, 60–69.
Zhang, W., Zhao, M., Ruesch, L., Omot, A., Francis, D.,
2007. Prevalence of virulence genes in Escherichia coli
strains recently isolated from young pigs with diarrhea in
the US. Vet. Microbiol. 123, 145–152.
Copyright © 2022 FDOKUMEN




















