
Cantone, Kupisch, Müller, Schmitz_Rethinking language dominance

Specific responses to nitrogen and phosphorusenrichment in cyanobacteria: Factors influencingchanges in species dominance along eutrophicgradients
Virginia Loza, Elvira Perona, Pilar Mateo*
Departamento de Biologıa, Facultad de Ciencias, Universidad Autonoma de Madrid, Madrid, Spain
a r t i c l e i n f o
Article history:
Received 3 May 2013
Received in revised form
10 September 2013
Accepted 6 October 2013
Available online 23 October 2013
Keywords:
Rivers
Streams
Benthic cyanobacteria
Growth
Nutrient regimes
Nitrate
Ammonium
Phosphate
Competition
a b s t r a c t
Anthropogenic eutrophication is aworldwide problem, causing proliferationof cyanobacterial
masses, some of whichmay be toxic. However, little is known about whether the response to
nutrient enrichment differs among cyanobacterial species. To address this issue, distinct
patterns in growth and competitive response of benthic cyanobacteria under N and P nutrient
regimes were studied. Nine cyanobacterial species, collected from Guadarrama river biofilms
at several locations with different nutrient concentrations, were isolated and used for a series
of N and P enrichment bioassays. In competition experiments with a mixture of all nine spe-
cies, a great predominance of certain cyanobacteria over others was noted at high nutrient
conditions,while under lownutrient conditions someothers dominated. On the basis of these
results four selected strains were subjected to a gradient of different concentrations of phos-
phate, nitrate and ammonium, in independent bioassays, both in monocultures and mixed
cultures. Depending on the concentration of N and P, stimulation or inhibition of growth was
observed. Some species grewbetter, dominating at highnutrient concentrations, while higher
yields were recorded for others under low nutrient regimes, dominating in these conditions.
Results from this study clarify previously published field observations, whereby a group of
species occurred mostly in downstream nutrient-rich locations, while other was typical of
upstream oligotrophic conditions. Our findings concerning differential growth in relation to
nutrient concentrationsmay be useful for environmental management, because they help us
predict which cyanobacteria may be expected to occur under certain conditions.
ª 2013 Elsevier Ltd. All rights reserved.
1. Introduction
Human disturbances give rise to a range of alterations in river
systems, in which nitrogen and phosphorus inputs can have
profound effects upon the quality of receiving waters, leading
to eutrophication, which in turn causes proliferation of algal
masses, some of which may be toxic (Dodds and Welch, 2000;
Anderson et al., 2002). Other symptoms of eutrophication
include deep water anoxia in lakes, taste and odour problems,
and changes in the composition of aquatic communities
* Corresponding author. Departamento de Biologıa, Facultad de Ciencias, Universidad Autonoma de Madrid, C/Darwin n� 2, 28049Madrid, Spain. Tel.: þ34 91 497 8184; fax: þ34 91 497 8344.
E-mail address: [email protected] (P. Mateo).
Available online at www.sciencedirect.com
ScienceDirect
journal homepage: www.elsevier .com/locate/watres
wat e r r e s e a r c h 4 8 ( 2 0 1 4 ) 6 2 2e6 3 1
0043-1354/$ e see front matter ª 2013 Elsevier Ltd. All rights reserved.http://dx.doi.org/10.1016/j.watres.2013.10.014

(Lewis et al., 2011). Recognition of these problems has
prompted a concerted worldwide effort to develop a rational
predictive framework for preventing and managing fresh-
water eutrophication (European Communities, 2000; USEPA,
2000).
Cyanobacteria are among dominant primary producers in
most freshwater environments, providing the principal en-
ergy base for many aquatic food webs (Scott and Marcarelli,
2012). They play important biogeochemical roles in terms of
nutrient fixation and cycling within the ecosystem (Scott and
Marcarelli, 2012). Nutrient enrichment has been closely linked
to the stimulation of growth of masses of cyanobacteria, but it
did not affect all species equally, although an increase of some
harmful species has been reported (Anderson et al., 2002).
Therefore, an improved understanding of the nature of taxon-
specific responses to N and P influx may help to protect
aquatic ecosystems against future pollution, optimise waste-
water treatment procedures, and resolve the continuing
debate about the respective roles of N and P in regulating
human made eutrophication (Donald et al., 2013).
In previous reports, we described shifts in the structure
and composition of benthic cyanobacterial communities in
rivers with increasing eutrophication in the downstream di-
rection (Perona et al., 1998, 1999; Douterelo et al., 2004; Perona
and Mateo, 2006; Rodriguez et al., 2007). In the Guadarrama
river (central Spain), molecular fingerprint analyses using
temperature gradient gel electrophoresis (TGGE) were
consistent with those obtained from microscopic observa-
tions of field-fixed samples, in which the cyanobacterial
community composition at sampling locations differed be-
tween upstream and downstream locations, whereby some
species predominated at downstream sites with high nutrient
content, while others were scarce or absent (Loza et al., 2013a,
b). However, assessing the causes of the presence or absence
of species is difficult from field observations alone. The causal
factors influencing cyanobacterial species composition are
poorly understood. In trying to detect factors that may affect
species distribution, statistical analyses have been carried out
to detect significant correlations between relative abundances
and water quality variables. However, many environmental
factors are usually interrelated so species distributionsmay be
correlatedwith a broad range of characteristics, although only
one may reflect a causal relationship. Therefore, a better un-
derstanding of the factors controlling the relative dominance
and/or coexistence phenomena in the environment is needed.
We hypothesized that specific characteristics of cyano-
bacterial species, such as differential physiological traits
related to changes in N and P concentration, influence how
species respond to changes in environmental conditions over
space and time, leading to the observed shifts in the structure
of the cyanobacterial communities. Such relationships have
often been proposed, but rarely demonstrated experimentally.
Empirical evidence of this nature would be helpful for inter-
preting species distributions in relation to environmental
changes. Therefore, in this study, a series of bioassays were
designed to test individually significant factors that may be
responsible for the observed changes. The effects of different
nutrient regimes (nitrogen and phosphorus) on the growth of
isolated strains from the Guadarrama river were assayed in an
attempt to identify causal relationships.
2. Materials and Methods
2.1. Cyanobacterial cultures
Cyanobacterial strains used for bioassays were isolated from
epilithic biofilms from the riverbed of the Guadarrama river
(central Spain, Madrid), as previously described (Loza et al.,
2013b). Three culture media were used for the isolation in
order to provide a wide range of nutrient concentrations, since
we had previously found different patterns of cyanobacterial
growth to be associated with the particular culture media used
(Berrendero et al., 2008). CHU No. 10modified (CHU10) medium
(Chu, 1942; Gomez et al., 2009) was selected as a low nutrient
content medium (0.99 mg P L�1, 3.5 mg N L�1), BG110 (Rippka
et al., 1979) was selected as an intermediate nutrient level me-
dium (5.5 mg P L�1, 247 mg N L�1), and Allen & Arnon medium
(AAN) (Allen and Arnon, 1955) was chosen as a high nutrient
concentration medium (61.9 mg P L�1, 175 mg N L�1). The nine
isolated cyanobacteria used for the experiments were the
coccoid and non-heterocystous filamentous species Cyanobium
sp., Aphanocapsa muscicola, Pleurocapsa minor, Pseudanabaena
catenata, Leptolyngbya boryana, Leptolyngbya nostocorum, Phormi-
dium sp. and the two heterocyst-forming speciesNostoc carneum
and Tolypothrix tenuis. A detailed description of all isolates used
in this study can be found in Loza et al. (2013b, c).
2.2. Experimental setup
2.2.1. Competition experiments with a mixture of all ninespeciesThedifferential growthofcyanobacterial strainswasanalyzedby
measuring the percentage of the surface of 14-cm diameter agar
plates colonized in the low nutrient CHU10 medium and the
nutrient-rich AAN, described above, simulating the oligo-
mesotrophic and hypertrophic conditions previously found in
the river (Loza et al., 2013a,b). Isolates were inoculated on 2%
agar-platesandwerekept for24h ina16:8h light:darkregimeata
temperature of 18 �C, with an irradiance of 20 mmol photon
m�2 s�1, without shaking, in order to fix inocula on the agar
surface.After that,agarplateswerecoveredby5mLofboth liquid
media (CHU10 and AAN medium) on each, and then put in the
shaker under the same conditions. Media were replaced on day
10 of the experiment. Relative abundance was analyzed using a
dissecting microscope (Leica; Leica Microsystems) and the per-
centageof the area colonizedwas calculated for each isolate. The
experiment was replicated two times, yielding similar results.
Values from a representative experiment are shown.
2.2.2. Nutrient gradient bioassaysOn the basis of the results obtained from the competition exper-
iments with all strains, a series of bioassays under a gradient of
different concentrations of derived nutrients from nitrogen (ni-
trate ðNO�3 Þ and ammonium ðNHþ
4 Þ and phosphorus ðPO3�4 Þwere
carried out. T. tenuis, N. carneum, Phormidium sp., and L. boryana.
were selected for these bioassays. Independent experiments for
each nutrient were performed over 20 days in CHU10medium in
which only the concentration of the ammonium, nitrate or
phosphatewasmodified to final concentrations of 0.2, 3, 6, 10, 50
and 100 mg L�1 (ranges: 0.04e22.58 mg L�1 Ne NO�3 ;
wat e r r e s e a r c h 4 8 ( 2 0 1 4 ) 6 2 2e6 3 1 623

0.16e82.35 mg L�1 NeNHþ4 ; 0.06e32.63 mg L�1 PePO3�
4 ). The ex-
periments were carried out with triplicate samples for each
cyanobacterium in monoculture, as well as in competition, in
which two species were placed together. Media at different
nutrient concentrationswere distributed in the different wells of
sterile polystyrene 24-well microtitre plates (IWAKI Microplate).
For each nutrient experiment, isolated cultures were washed
twice in the appropriate medium and 100 mL were transferred to
the sterile plates with 2 mL of each medium at the correct con-
centration. Finally, plates weremaintained for 24 h with a 16:8 h
light:dark period at a temperature of 18 �C, with an irradiance of
20 mmol photonm�2 s�1,without shaking. Theywere thenplaced
on a rotating shaker (200 rpm) to obtain a homogeneous suspen-
sionof theculture.Themediumwaspartially replaced (50%)once
during the 10 day experiment. For each species, the starting
inoculum for all experiments was taken from the same stock
culture, which was grown under low nutrient conditions for one
week.
2.3. Sample counting
Growth of monocultures and mixed cultures was measured as
the totalnumberof cellsmL�1. Sincea thinbiofilmwasdeveloped
at the bottom of the wells at the end of the experiment, due to a
great tendency of epilithic cyanobacteria to attach to the sub-
strata, samples were firmly scraped with a sterile scalpel. Sam-
ples were then preserved adding 10 mL of formaldehyde to each
well (4% final concentration), and kept in dark at 4 �C, taken into
consideration this dilution in further cell counting. Total number
of cells was determinedmicroscopically by counting at least 400
cells in each sample in aNeubauer chamber (BRANDRef.718605).
2.4. Statistical analysis
All groups of data were analysed for normality by the
ShapiroeWilk test and for homoscedasticity by the Levene’s
test. Significant differences in growth were identified using
the non-parametric KruskaleWallis one-way analysis of
variance as several of our data did not follow a normal dis-
tribution and/or did not present homogeneity of variances.
Further post hoc comparisons were performed using the
Student-Newman-Keuls method. Statistics were carried out
using SigmaPlot version 9.0 with SigmaStat integration
version 3.1.
3. Results
3.1. Competition experiments with a mixture of all ninespecies
Representation of differential growth trends under nutrient
regimes in cyanobacterial isolates is shown in Fig. 1. The dis-
tribution of the species on day 10 at high nutrient concentra-
tions (AANmedium) showed a dominance of N. carneum (37%)
and Phormidium sp. (27%), followed by A. muscicola, P. catenata
and L. nostocorum (Fig. 1A). However, on day 20 (Fig. 1B) a clear
dominance of Phormidium sp. over the rest of cyanobacteria
was noticed, with almost 59% of the area colonized, and with
N. carneum accounting for only 7.6% of the coverage. T. tenuis
and P. minor were represented by very low percentages of the
colonized surface: 0.4 and 0.7%, respectively (Fig. 1B). On the
other hand, under the low nutrient regime (CHU10medium) T.
tenuiswas dominant on day 10 (Fig. 1A) occupying 41.5% of the
total area colonized, followed by N. carneum (37.5%) and L.
nostocorum (5%). Similar results were obtained after day 20 of
the treatment (Fig. 1B), in which T. tenuis comprised 49% of the
colonized surface, closely followed by N. carneum (40%).
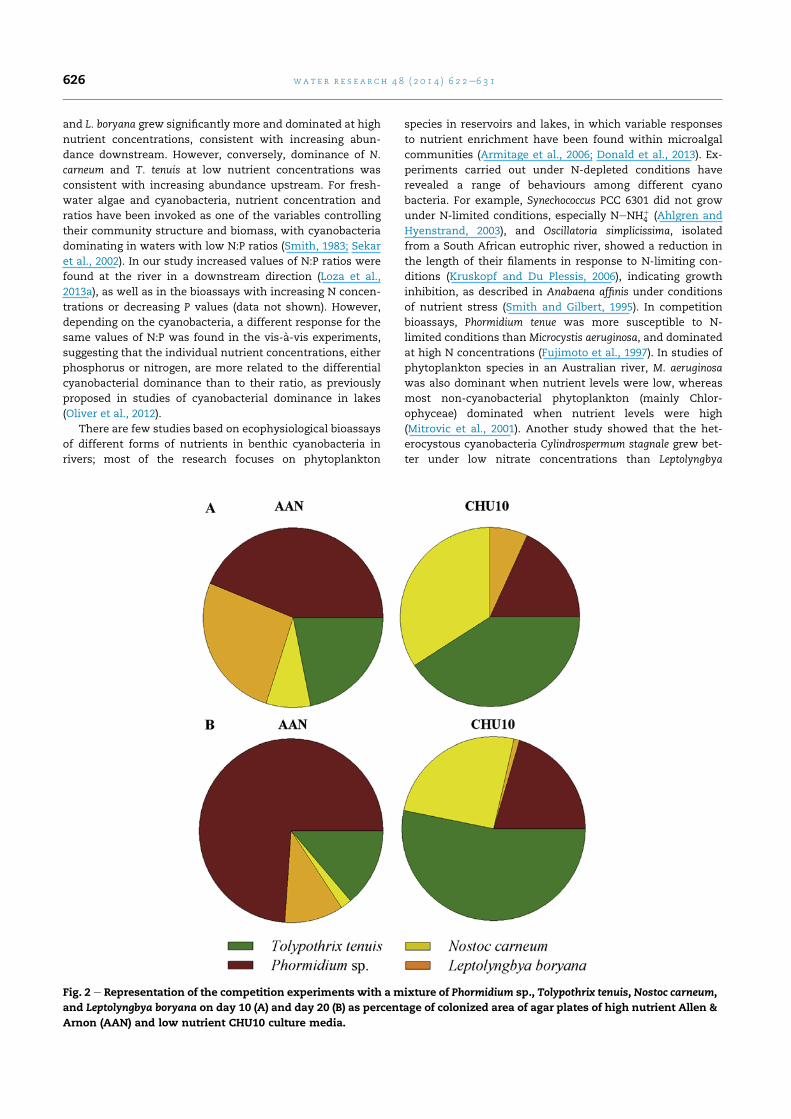
On the basis of these results, a further experiment (Fig. 2)
was carried out in the different culture media with the two
dominant cyanobacteria, Phormidium sp. and T. tenuis, select-
ing N. carneum as the other N-fixing cyanobacteria whose
response varied with the medium and time of culture, and L.
boryana as the non-heterocystous cyanobacteria that grew
similarly in both nutrient regimes (Fig. 1). Similar results were
found concerning the dominant species, whereby for all
exposure durations, Phormidium sp. greatly predominated
under high nutrient conditions, while T. tenuis dominated
under low nutrient conditions. N. carneum also exhibited
greater growth at lower nutrient concentrations, while L.
boryana grewmore under the higher nutrient regime. Based on
these results, these four cyanobacterial species were selected
for subsequent bioassays in order to identify the patterns
among these isolates more clearly.
3.2. Nutrient gradient bioassays
Selected species were subjected to a gradient of different
concentrations of ammonium, nitrate and phosphate. Cya-
nobacterial growthwasmeasured inmonocultures andmixed
cultures. On the basis of previous results, competition ex-
periments were performed mixing T. tenuis with Phormidium
sp., and N. carneum with L. boryana.
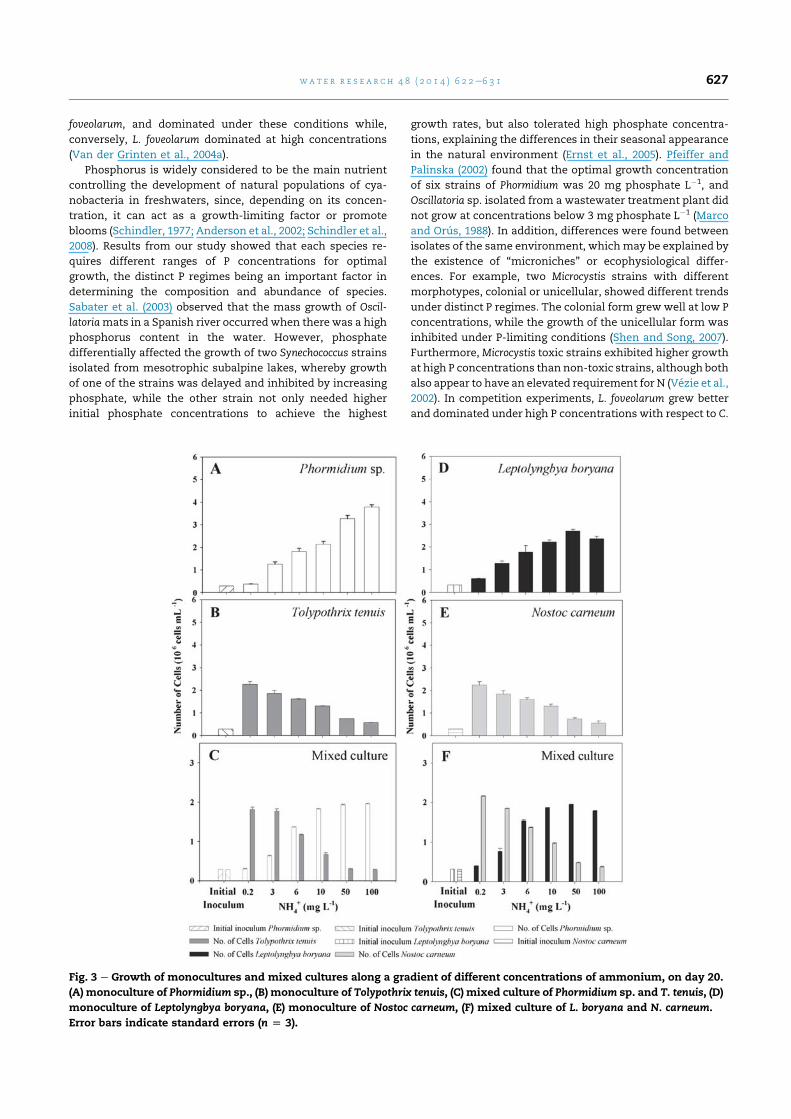
3.2.1. Ammonium responsesIn monoculture bioassays, Phormidium sp. and L. boryana grew
significantly more at higher ammonium concentrations
(KruskaleWallis, p < 0.01) (Fig. 3A, D), except for a slight
decrease in L. boryana biomass at 100 mg L�1. At the lowest
concentration (0.2 mg L�1) the number of cells was similar to
that of the initial inoculum, indicating that these conditions
were adverse for both species. Conversely, growth of mono-
cultures of T. tenuis and N. carneum decreased as the concen-
tration of ammonium increased, both with similar growth
patterns (Fig. 3B, E). The same trend was noticed for each
cyanobacterium in competition experiments, leading to a
clear and significant dominance of Phormidium sp. and L. bor-
yana at higher ammonium concentrations, while T. tenuis and
N. carneum dominated significantly at low ammonium con-
centrations (Fig. 3C, F) (KruskaleWallis, p < 0.01).
3.2.2. Nitrate responsesGrowth responses of Phormidium sp. and L. boryana under ni-
trate gradient bioassays also had a significantly higher yield
(KruskaleWallis, p< 0.01) at higher concentrations (Fig. 4A, D).
On the other hand, growth of N. carneum decreased signifi-
cantly (KruskaleWallis, p < 0.05) as the nitrate concentration
in the culture medium increased, whereas in the T. tenuis
bioassay, growth varied little along the nitrate gradient
(Fig. 4B, E). In mixed cultures, again, T. tenuis and N. carneum
wat e r r e s e a r c h 4 8 ( 2 0 1 4 ) 6 2 2e6 3 1624

grew more under low nitrate concentrations, while Phormi-
dium sp. and L. boryana dominated significantly at higher
concentrations (KruskaleWallis, p < 0.01) (Fig. 4C, F).
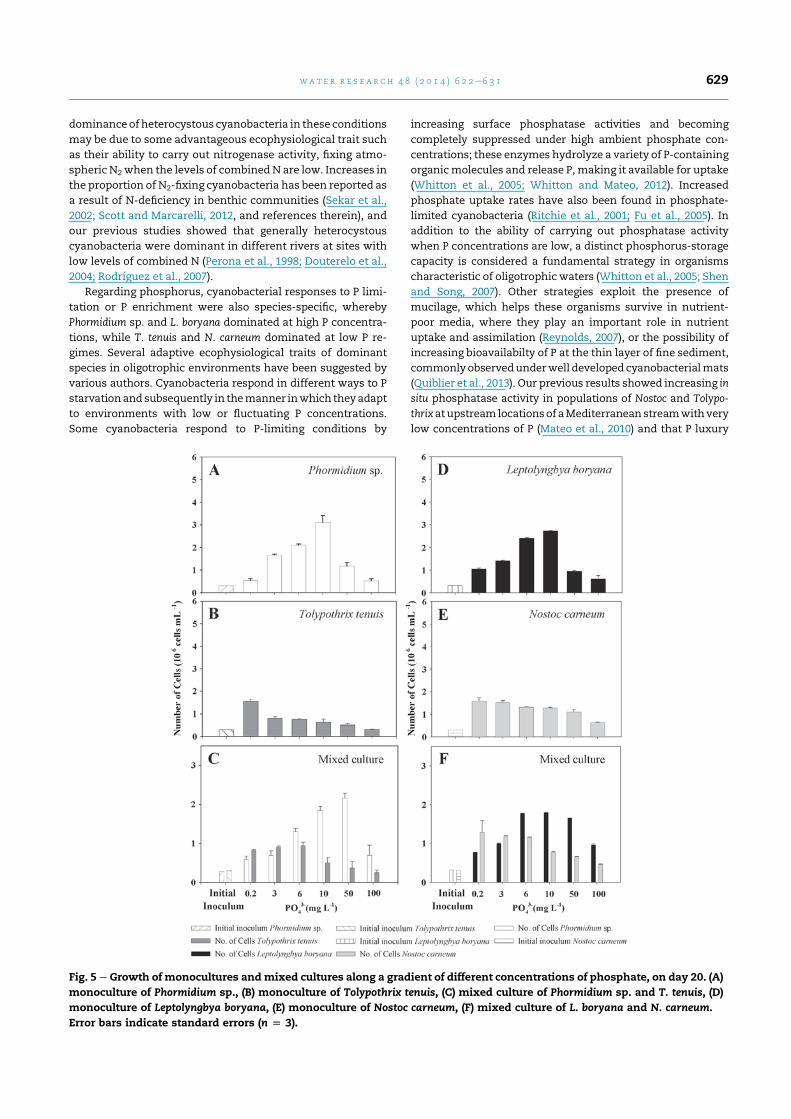
3.2.3. Phosphate responsesIn monocultures, under increased phosphate regimes, Phormi-
dium sp. and L. boryana showed a progressive increase in
biomass, peaking at 10mgL�1, and thendecreasing significantly
atevenhigher concentrations (KruskaleWallis,p< 0.01) (Fig. 5A,
D). T. tenuis and N. carneum showed a significant reduction in
biomass along the gradient (Fig. 5B, E) (KruskaleWallis, p< 0.01,
both species). The trend in competition experiments (Fig. 5C, F)
was the same as that in previous nutrients, whereby T. tenuis
and N. carneum dominated under low phosphate regimes, but
Phormidium sp. and L. boryana dominated at higher phosphate
concentrations (KruskaleWallis, p < 0.01).
4. Discussion
Patterns of growth and competitive response under different
nutrient regimes differed between cyanobacteria. Depending
on the concentration of phosphorus and nitrogen, growthwas
either stimulated or inhibited. Some species grew better at
high nutrient concentrations, while higher yields were
recorded for others under low nutrient regimes, leading to
significant shifts in dominance depending on the N and P,
enrichments, as well as on the redox distinction between ni-
trogen nutrients. These results are consistent with the dif-
ferential distribution of these cyanobacteria in the
Guadarrama river, according to downstream increasing
eutrophication; whereby a group of species occurredmostly in
downstreamnutrient-rich locations, while otherswere typical
of upstream oligotrophic conditions (Loza et al., 2013a,b c).
Therefore, the studied cyanobacteria responded differently in
terms of their nutrient requirements. Those cyanobacteria
which dominated in the nutrient-rich culture medium (AAN)
appeared in great abundance in the river under hypertrophic
conditions, while those with a higher abundance in the sam-
pling site with low nutrient concentrations, in turn, grew
better in the culture medium that simulated oligotrophic
conditions (CHU10). In nutrient gradient bioassays, where
ranges of concentration were representative of downstream
increasing eutrophication (Loza et al., 2013b), Phormidium sp.
Fig. 1 e Representation of the competition experiments with a mixture of nine cyanobacterial species on day 10 (A) and day
20 (B) as percentage of colonized area of agar plates of high nutrient Allen & Arnon (AAN) and low nutrient CHU10 culture
media (see Materials and Methods).
wat e r r e s e a r c h 4 8 ( 2 0 1 4 ) 6 2 2e6 3 1 625
and L. boryana grew significantly more and dominated at high
nutrient concentrations, consistent with increasing abun-
dance downstream. However, conversely, dominance of N.
carneum and T. tenuis at low nutrient concentrations was
consistent with increasing abundance upstream. For fresh-
water algae and cyanobacteria, nutrient concentration and
ratios have been invoked as one of the variables controlling
their community structure and biomass, with cyanobacteria
dominating in waters with low N:P ratios (Smith, 1983; Sekar
et al., 2002). In our study increased values of N:P ratios were
found at the river in a downstream direction (Loza et al.,
2013a), as well as in the bioassays with increasing N concen-
trations or decreasing P values (data not shown). However,
depending on the cyanobacteria, a different response for the
same values of N:P was found in the vis-a-vis experiments,
suggesting that the individual nutrient concentrations, either
phosphorus or nitrogen, are more related to the differential
cyanobacterial dominance than to their ratio, as previously
proposed in studies of cyanobacterial dominance in lakes
(Oliver et al., 2012).
There are few studies based on ecophysiological bioassays
of different forms of nutrients in benthic cyanobacteria in
rivers; most of the research focuses on phytoplankton
species in reservoirs and lakes, in which variable responses
to nutrient enrichment have been found within microalgal
communities (Armitage et al., 2006; Donald et al., 2013). Ex-
periments carried out under N-depleted conditions have
revealed a range of behaviours among different cyano
bacteria. For example, Synechococcus PCC 6301 did not grow
under N-limited conditions, especially NeNHþ4 (Ahlgren and
Hyenstrand, 2003), and Oscillatoria simplicissima, isolated
from a South African eutrophic river, showed a reduction in
the length of their filaments in response to N-limiting con-
ditions (Kruskopf and Du Plessis, 2006), indicating growth
inhibition, as described in Anabaena affinis under conditions
of nutrient stress (Smith and Gilbert, 1995). In competition
bioassays, Phormidium tenue was more susceptible to N-
limited conditions than Microcystis aeruginosa, and dominated
at high N concentrations (Fujimoto et al., 1997). In studies of
phytoplankton species in an Australian river, M. aeruginosa
was also dominant when nutrient levels were low, whereas
most non-cyanobacterial phytoplankton (mainly Chlor-
ophyceae) dominated when nutrient levels were high
(Mitrovic et al., 2001). Another study showed that the het-
erocystous cyanobacteria Cylindrospermum stagnale grew bet-
ter under low nitrate concentrations than Leptolyngbya
Fig. 2 e Representation of the competition experiments with a mixture of Phormidium sp., Tolypothrix tenuis, Nostoc carneum,
and Leptolyngbya boryana on day 10 (A) and day 20 (B) as percentage of colonized area of agar plates of high nutrient Allen &
Arnon (AAN) and low nutrient CHU10 culture media.
wat e r r e s e a r c h 4 8 ( 2 0 1 4 ) 6 2 2e6 3 1626
foveolarum, and dominated under these conditions while,
conversely, L. foveolarum dominated at high concentrations
(Van der Grinten et al., 2004a).
Phosphorus is widely considered to be the main nutrient
controlling the development of natural populations of cya-
nobacteria in freshwaters, since, depending on its concen-
tration, it can act as a growth-limiting factor or promote
blooms (Schindler, 1977; Anderson et al., 2002; Schindler et al.,
2008). Results from our study showed that each species re-
quires different ranges of P concentrations for optimal
growth, the distinct P regimes being an important factor in
determining the composition and abundance of species.
Sabater et al. (2003) observed that the mass growth of Oscil-
latoriamats in a Spanish river occurred when there was a high
phosphorus content in the water. However, phosphate
differentially affected the growth of two Synechococcus strains
isolated from mesotrophic subalpine lakes, whereby growth
of one of the strains was delayed and inhibited by increasing
phosphate, while the other strain not only needed higher
initial phosphate concentrations to achieve the highest
growth rates, but also tolerated high phosphate concentra-
tions, explaining the differences in their seasonal appearance
in the natural environment (Ernst et al., 2005). Pfeiffer and
Palinska (2002) found that the optimal growth concentration
of six strains of Phormidium was 20 mg phosphate L�1, and
Oscillatoria sp. isolated from a wastewater treatment plant did
not grow at concentrations below 3 mg phosphate L�1 (Marco
and Orus, 1988). In addition, differences were found between
isolates of the same environment, whichmay be explained by
the existence of “microniches” or ecophysiological differ-
ences. For example, two Microcystis strains with different
morphotypes, colonial or unicellular, showed different trends
under distinct P regimes. The colonial form grew well at low P
concentrations, while the growth of the unicellular form was
inhibited under P-limiting conditions (Shen and Song, 2007).
Furthermore,Microcystis toxic strains exhibited higher growth
at high P concentrations than non-toxic strains, although both
also appear to have an elevated requirement for N (Vezie et al.,
2002). In competition experiments, L. foveolarum grew better
and dominated under high P concentrations with respect to C.
Fig. 3 e Growth of monocultures and mixed cultures along a gradient of different concentrations of ammonium, on day 20.
(A) monoculture of Phormidium sp., (B) monoculture of Tolypothrix tenuis, (C) mixed culture of Phormidium sp. and T. tenuis, (D)
monoculture of Leptolyngbya boryana, (E) monoculture of Nostoc carneum, (F) mixed culture of L. boryana and N. carneum.
Error bars indicate standard errors (n [ 3).
wat e r r e s e a r c h 4 8 ( 2 0 1 4 ) 6 2 2e6 3 1 627

stagnale (Van der Grinten et al., 2004b). Planktothrix agardii
(formerly Oscillatoria agardii), which is highly abundant in
many eutrophic lakes in the Netherlands, was not competitive
under P-limiting conditions, wherein it is displaced by the
prochlorophyta Prochlorothrix hollandice in competition exper-
iments (Ducobu et al., 1998).
There is renewed interest in understanding patterns in the
distribution not only of taxa but also of their traits. Trait-based
approaches to microbial biodiversity and biogeography offer
the possibility of advancing ecological theory and predicting
responses to environmental change (Green et al., 2008).
Climate change scenarios predict that rivers and other fresh-
water ecosystems will experience higher temperatures and
altered nutrient loading (Carey et al., 2012; Elliot, 2012). Elliot
(2012) indicated that most of the modelling studies that
included a change innutrients showedanenhancementunder
thehigher nutrient scenarios of the cyanobacterial response to
the climate drivers. Paerl and Paul (2012) pointed out that the
expansion of harmful bloom-forming cyanobacteria, linked to
climate change, has serious consequences for humandrinking
and irrigation water supplies, fisheries and recreational re-
sources, and this has ramifications for water management. In
addition, temperature, light, and nutrients may have interac-
tive effects on cyanobacterial growth and toxin production,
with consequent effects on aquatic food webs (Paerl and Paul,
2012; Elliot, 2012). These environmental drivers will have
substantial effects on species composition, and it has been
predicted that spatial variation in climate change will interact
with physiological variation in cyanobacteria to alter the
dominant cyanobacterial taxa from region to region (Carey
et al., 2012). The results of our research show that ecological
rangesof individual cyanobacterial populations candifferwith
nutrient concentration, and the persistence of several species
under low or high nutrient concentrations in the river can be
explained on the basis of their different ecophysiological
properties. Increased levels of ammonium and/or nitrate-
nitrogen favour the development of the non-heterocystous
cyanobacteria Phormidium sp. and L. boryana, whereas com-
bined nitrogen scarcity facilitates the development of the
heterocystous cyanobacteria N. carneum and T. tenuis. The
Fig. 4 e Growth of monocultures and mixed cultures along a gradient of different concentrations of nitrate, on day 20. (A)
monoculture of Phormidium sp., (B) monoculture of Tolypothrix tenuis, (C) mixed culture of Phormidium sp. and T. tenuis, (D)
monoculture of Leptolyngbya boryana, (E) monoculture of Nostoc carneum, (F) mixed culture of L. boryana and N. carneum.
Error bars indicate standard errors (n [ 3).
wat e r r e s e a r c h 4 8 ( 2 0 1 4 ) 6 2 2e6 3 1628
dominance ofheterocystous cyanobacteria in these conditions
may be due to some advantageous ecophysiological trait such
as their ability to carry out nitrogenase activity, fixing atmo-
spheric N2when the levels of combinedN are low. Increases in
the proportion of N2-fixing cyanobacteria has been reported as
a result of N-deficiency in benthic communities (Sekar et al.,
2002; Scott and Marcarelli, 2012, and references therein), and
our previous studies showed that generally heterocystous
cyanobacteria were dominant in different rivers at sites with
low levels of combined N (Perona et al., 1998; Douterelo et al.,
2004; Rodrıguez et al., 2007).
Regarding phosphorus, cyanobacterial responses to P limi-
tation or P enrichment were also species-specific, whereby
Phormidium sp. and L. boryana dominated at high P concentra-
tions, while T. tenuis and N. carneum dominated at low P re-
gimes. Several adaptive ecophysiological traits of dominant
species in oligotrophic environments have been suggested by
various authors. Cyanobacteria respond in different ways to P
starvationandsubsequently in themanner inwhich theyadapt
to environments with low or fluctuating P concentrations.
Some cyanobacteria respond to P-limiting conditions by
increasing surface phosphatase activities and becoming
completely suppressed under high ambient phosphate con-
centrations; these enzymes hydrolyze a variety of P-containing
organicmolecules and release P, making it available for uptake
(Whitton et al., 2005; Whitton and Mateo, 2012). Increased
phosphate uptake rates have also been found in phosphate-
limited cyanobacteria (Ritchie et al., 2001; Fu et al., 2005). In
addition to the ability of carrying out phosphatase activity
when P concentrations are low, a distinct phosphorus-storage
capacity is considered a fundamental strategy in organisms
characteristic of oligotrophic waters (Whitton et al., 2005; Shen
and Song, 2007). Other strategies exploit the presence of
mucilage, which helps these organisms survive in nutrient-
poor media, where they play an important role in nutrient
uptake and assimilation (Reynolds, 2007), or the possibility of
increasing bioavailabilty of P at the thin layer of fine sediment,
commonlyobservedunderwell developed cyanobacterialmats
(Quiblier et al., 2013). Our previous results showed increasing in
situ phosphatase activity in populations of Nostoc and Tolypo-
thrixatupstream locationsofaMediterraneanstreamwithvery
low concentrations of P (Mateo et al., 2010) and that P luxury
Fig. 5 e Growth of monocultures andmixed cultures along a gradient of different concentrations of phosphate, on day 20. (A)
monoculture of Phormidium sp., (B) monoculture of Tolypothrix tenuis, (C) mixed culture of Phormidium sp. and T. tenuis, (D)
monoculture of Leptolyngbya boryana, (E) monoculture of Nostoc carneum, (F) mixed culture of L. boryana and N. carneum.
Error bars indicate standard errors (n [ 3).
wat e r r e s e a r c h 4 8 ( 2 0 1 4 ) 6 2 2e6 3 1 629

consumption, by which cells can be taken up in excess of im-
mediate requirement and stored in form the polyphosphate
granules, could assist to overcome P deficient conditions
(Mateo et al., 2006;Munoz-Martın et al., 2013). Furthermore, the
phosphatase activity of Nostoc and Tolypothrix was induced
under P deficiency, and there was also a negative correlation
between this activity and external inorganic P concentration
(Mateo et al., 2006; Munoz-Martın et al., 2013). Therefore, in an
environment with variable and intermittent P inputs, some
cyanobacteria can overcome changes in nutrient loading by
regulatingabsorptionmechanismsandPutilization for optimal
growth. In addition, different cyanobacterial taxa can also vary
in their relative capacity and efficiency in relation to these
processes, which can also explain the differential response
found between the studied cyanobacteria.
5. Conclusions
- Taxa-specific variability in cyanobacterial growth responses
to nutrient enrichment demonstrates that these primary
producer components of fluvial systems do not respond
uniformly to changes in nutrient input, leading to shifts in
the structure and composition of communities.
- Although it is generally assumed that nutrient-rich condi-
tions favour cyanobacterial growth, results from this study
indicate that cyanobacteria should be treated individually
rather than as a homogeneous group. Some species prefer
low nutrient conditions, while others prefer eutrophic to
hypertrophic conditions.
- Specific adaptive ecophysiological traits offer the possibility
for cyanobacteria to occupy distinct niches, which would
explain their environmental divergence, and therefore their
differential distribution.
- Our study provides further information about species-
specific autecologies and interactions between species,
and in turn, highlights the importance of understanding the
dominance of each one, according to their different
ecological preferences, which may provide more informa-
tion about the trophic state in which they occur.
- Information about species-specific autecologies and in-
teractions between speciesmay be useful for environmental
management, because it helps to predict which cyanobac-
teria may be expected to occur under certain conditions.
Acknowledgements
This work was supported by grants from the Ministerio de
Educacion y Ciencia, Spain (CGL2008-02397/BOS), and from the
Comunidad Autonoma de Madrid, Spain (S2009/AMB-1511).
r e f e r e n c e s
Ahlgren, G., Hyenstrand, P., 2003. Nitrogen limitation effects ofdifferent nitrogen sources on nutritional quality of two
freshwater organisms, Scenedesmus quadricauda(Chlorophyceae) and Synechococcus sp. (Cyanophyceae). J.Phycology 39 (5), 906e917.
Allen, M.B., Arnon, D.I., 1955. Studies on nitrogen-fixing blue-green algae. I. Growth and nitrogen fixation by Anabaenacylindrica Lemm. Plant Physiol. 30 (4), 366e372.
Anderson, D.M., Glibert, P.M., Burkholder, J.M., 2002. Harmfulalgal blooms and eutrophication: nutrient sources,composition, and consequences. Estuaries 25 (4b), 704e726.
Armitage, A.R., Frankovich, T.A., Fourqurean, J.W., 2006. Variableresponseswithin epiphytic andbenthicmicroalgal communitiesto nutrient enrichment. Hydrobiologia 569, 423e435.
Berrendero, E., Perona, E., Mateo, P., 2008. Genetic andmorphological characterization of Rivularia and Calothrix(Nostocales, Cyanobacteria) from running water. Int. J. Syst.Evol. Microbiol. 58, 447e460.
Carey, C.C., Ibelings, B.W., Hoffmann, E.P., Hamilton, D.P.,Brookes, J.D., 2012. Eco-physiological adaptations that favourfreshwater cyanobacteria in a changing climate. Water Res. 46(5), 1394e1407.
Chu, S.P., 1942. The influence of the mineral composition of themedium on the growth of planktonic algae. Part 1. Methodsand culture media. J. Ecol. 30, 284e325.
Dodds, W.K., Welch, E.B., 2000. Establishing nutrient criteria instreams. J. North Am. Benthological Soc. 19 (1), 186e196.
Donald, D.B., Bogard, M.J., Finlay, K., Bunting, L., Leavitt, P.R.,2013. Phytoplankton-specific response to enrichment ofphosphorus-rich surface waters with ammonium, nitrate, andurea. PLoS ONE 8 (1), e53277. http://dx.doi.org/10.1371/journal.pone.0053277.
Douterelo, I., Perona, E., Mateo, P., 2004. Use of cyanobacteria toassess water quality in running waters. Environ. Pollut. 127 (3),377e384.
Ducobu, H., Huisman, J., Jonker, R.R., Mur, L.R., 1998. Competitionbetween a prochlorophyte and a cyanobacterium undervarious phosphorus regimes: comparison with the droopmodel. J. Phycology 34 (3), 467e476.
Elliot, J.A., 2012. Is the future blue-green? A review of the currentmodel predictions of how climate change could affect pelagicfreshwater cyanobacteria. Water Res. 46, 1364e1371.
Ernst, A., Deicher, M., Herman, P.M.J., Wollenzien, U.I.A., 2005.Nitrate and phosphate affect cultivability of cyanobacteriafrom environments with low nutrient levels. Appl. Environ.Microbiol. 71 (6), 3379e3383.
European Communities, 22/12/2000. Directive 2000/60/EC of theEuropean Parliament and of the Council of the 23 October 2000establishing a framework for community action in the field ofwater policy. Official J. Eur. Communities L 327.
Fu, F.X., Zhang, Y., Bell, P.R.F., Hutchins, D.A., 2005. Phosphateuptake and growth kinetics of Trichodesmium (Cyanobacteria)isolates from the North Atlantic Ocean and the Great BarrierReef, Australia. J. Phycology 41 (1), 62e73.
Fujimoto, N., Sudo, R., Sugiura, N., Inamori, Y., 1997. Nutrient-limited growth of Microcystis aeruginosa and Phormidium tenueand competition under various N: P supply ratios andtemperatures. Limnol. Oceanogr. 42 (2), 250e256.
Gomez, N., Donato, J.C., Giorgi, A., Guasch, H., Mateo, P.,Sabater, S., 2009. La biota de los rıos: los microorganismosautotrofos. In: Elosegi, A., Sabater, S. (Eds.), Conceptos ytecnicas en ecologıa fluvial. Fundacion BBVA., Bilbao, Spain,pp. 219e242.
Green, J.L., Bohannan, B.J.M., Whitaker, R.J., 2008. Microbialbiogeography: from taxonomy to traits. Science 320,1039e1043.
Kruskopf, M., Du Plessis, S., 2006. Growth and filament length ofthe bloom forming Oscillatoria simplicissima (Oscillatoriales,Cyanophyta) in varying N and P concentrations. Hydrobiologia556, 357e362.
wat e r r e s e a r c h 4 8 ( 2 0 1 4 ) 6 2 2e6 3 1630

Lewis Jr., W.M., Wurtsbaugh, W.A., Paerl, H.W., 2011. Rationale forcontrol of anthropogenic nitrogen and phosphorus to reduceeutrophication of inland waters. Environ. Sci. Technol. 45 (24),10300e10305.
Loza, V., Perona, E., Mateo, P., 2013a. Molecular fingerprinting ofcyanobacteria from river biofilms as a water qualitymonitoring tool. Appl. Environ. Microbiol. 79 (5), 1459e1472.
Loza, V., Berrendero, E., Perona, E., Mateo, P., 2013b. Polyphasiccharacterization of benthic cyanobacterial diversity frombiofilms of the Guadarrama (Spain): morphological, molecularand ecological approaches. J. Phycology 49 (2), 282e297.
Loza, V., Perona, E., Carmona, J., Mateo, P., 2013c. Phenotypic andgenotypic characteristics of Phormidium-like cyanobacteriainhabiting microbial mats are correlated with the trophicstatus in running waters. Eur. J. Phycology 48 (2), 235e252.
Marco, E., Orus, M.I., 1988. Variation in growth and metabolismwith phosphorus nutrition in two cyanobacteria. J. PlantPhysiol. 132 (3), 339e344.
Mateo, P., Douterelo, I., Berrendero, E., Perona, E., 2006.Physiological differences between two species ofcyanobacteria in relation to phosphorus limitation. J.Phycology 42 (1), 61e66.
Mateo, P., Berrendero, E., Perona, E., Loza, V., Whitton, B.A., 2010.Phosphatase activities of cyanobacteria as indicators ofnutrient status in a Pyrenees river. Hydrobiologia 652 (1),255e268.
Mitrovic, S.M., Bowling, L.C., Buckney, R.T., 2001. Responses ofphytoplankton to in-situ nutrient enrichment; potentialinfluences on species dominance in a river. Int. Rev.Hydrobiology 86 (3), 285e298.
Munoz-Martın, M.A., Martınez-Rosell, A., Perona, E., Fernandez-Pinas, F., Mateo, P., 2013. Monitoring bioavailable phosphorusin lotic systems: a polyphasic approach based oncyanobacteria. Sci. Total Environ.. http://dx.doi.org/10.1016/j.scitotenv.2013.06.076.
Oliver, R.L., Hamilton, D.P., Brookes, J.D., Ganf, G.G., 2012.Physiology, blooms and prediction of planktoniccyanobacteria. In: Whitton, B.A. (Ed.), Ecology ofCyanobacteria II. Their Diversity in Space and Time. Springer,Dordrecht, pp. 155e194.
Paerl, H.W., Paul, V.J., 2012. Climate change: links to globalexpansion of harmful Cyanobacteria. Water Res. 46,1349e1363.
Perona, E., Bonilla, I., Mateo, P., 1998. Epilithic cyanobacteriacommunities and water quality: an alternative tool formonitoring eutrophication in the Alberche River (Spain). J.Appl. Phycology 10 (2), 183e191.
Perona, E., Bonilla, I., Mateo, P., 1999. Spatial and temporalchanges in water quality in a Spanish river. Sci. Total Environ.241 (1e3), 75e90.
Perona, E., Mateo, P., 2006. Benthic cyanobacterial assemblages asindicators of nutrient enrichment regimes in a Spanish river.Acta Hydrochimica et Hydrobiologica 34 (1e2), 67e72.
Pfeiffer, C., Palinska, K.A., 2002. Characterization of marinePhormidium isolates-conformity between molecular andecophysiological results. J. Plant Physiol. 159 (6), 591e598.
Quiblier, C., Wood, S., Echenique-Subiabre, I., Heath, M.,Villeneuve, A., Humbert, J.F., 2013. A review of currentknowledge on toxic benthic freshwater cyanobacteria e
ecology, toxin production and risk management. Water Res..http://dx.doi.org/10.1016/j.watres.2013.06.042.
Reynolds, C.S., 2007. Variability in the provision and function ofmucilage in phytoplankton: facultative responses to theenvironment. Hydrobiologia 578 (1), 37e45.
Rippka, R., Deruelles, J., Waterbury, J.B., Herdman, M.,Stanier, R.Y., 1979. Generic assignments, strain histories, andproperties of pure cultures of cyanobacteria. J. Gen. Microbiol.111, 1e61.
Ritchie, R.J., Trautman, D.A., Larkum, A.W.D., 2001. Phosphatelimited cultures of the cyanobacterium Synechococcus arecapable of very rapid, opportunistic uptake of phosphate. NewPhytol. 152 (2), 189e201.
Rodrıguez, V., Aguirre de Carcer, D., Loza, V., Perona, E., Mateo, P.,2007. A molecular fingerprint technique to detect pollution-related changes in river cyanobacterial diversity. J. Environ.Qual. 36 (2), 464e468.
Sabater, S., Vilalta, E., Gaudes, A., Guasch, H., Munoz, I.,Romanı, A., 2003. Ecological implications of mass growth ofbenthic cyanobacteria in rivers. Aquat. Microb. Ecol. 32 (2),175e184.
Schindler, D.W., 1977. Evolution of phosphorus limitation inlakes. Science 195, 260e262.
Schindler, D.W., Hecky, R.E., Findlay, D.L., Stainton, M.P.,Parker, B.R., Paterson, M.J., Beaty, K.G., Lyng, M.,Kasian, S.E.M., 2008. Eutrophication of lakes cannot becontrolled by reducing nitrogen input: results of a 37-yearwhole-ecosystem experiment. Proc. Natl. Acad. Sci. UnitedStates America 105 (32), 11254e11258.
Scott, J.T., Marcarelli, A.M., 2012. Cyanobacteria in freshwaterbenthic environments. In: Whitton, B.A. (Ed.), Ecology ofCyanobacteria II. Their Diversity in Space and Time. Springer,Dordrecht, pp. 271e289.
Sekar, R., Nair, K.V.K., Rao, V.N.R., Venugopalan, V.P., 2002.Nutrient dynamics and successional changes in a lenticfreshwater biofilm. Freshw. Biol. 47, 1893e1907.
Shen, H., Song, L.R., 2007. Comparative studies on physiologicalresponses to phosphorus in two phenotypes of bloom-formingMicrocystis. Hydrobiologia 592, 475e486.
Smith, V.H., 1983. Low nitrogen to phosphorus ratios favordominance by blue-green-algae in lake phytoplankton.Science 221, 669e671.
Smith, A.D., Gilbert, J.J., 1995. Spatial and temporal variability infilament length of a toxic cyanobacterium (Anabaena affinis).Freshw. Biol. 33 (1), 1e11.
USEPA (United States Environmental Protection Agency), 2000.Nutrient Criteria. Technical Guidance Manual. Rivers andStreams. EPA-822-B-00-002. United States EnvironmentalProtection Agency, Washington, pp. 17e28.
Van der Grinten, E., Simis, S.G.H., Barranguet, C., Admiraal, W.,2004a. Dominance of diatoms over cyanobacterial species innitrogen-limited biofilms. Archiv fur Hydrobiologie 161 (1),99e112.
Van der Grinten, E., Janssen, M., Simis, S.G.H., Barranguet, C.,Admiraal, W., 2004b. Phosphate regime structures speciescomposition in cultured phototrophic biofilms. Freshw. Biol.49 (4), 369e381.
Vezie, C., Rapala, J., Vaitomaa, J., Seitsonen, J., Sivonen, K., 2002.Effect of nitrogen and phosphorous on growth of toxic andnontoxic Microcystis strains on intracelular microcystinconcentrations. Microb. Ecol. 43 (4), 443e454.
Whitton, B.A., Mateo, P., 2012. Rivulariaceae. In: Whitton, B.A.(Ed.), Ecology of Cyanobacteria II. Their Diversity in Space andTime. Springer, Dordrecht, pp. 561e592.
Whitton, B.A., Al-Shehri, A.M., Ellwood, N.T.W., Turner, B.L., 2005.Ecological aspects of phosphatase activity in cyanobacteria,eukaryotic algae and bryophytes. In: Turner, B.L., Frossard, E.,Baldwin, D.S. (Eds.), Organic Phosphatase in the Environment.CABI, Wallingford, pp. 205e242.
wat e r r e s e a r c h 4 8 ( 2 0 1 4 ) 6 2 2e6 3 1 631
Copyright © 2022 FDOKUMEN




















