
Sorghum (Sorghum bicolor L. Moench) contains a XIP-type xylanase inhibitor but none of the TAXI- and...
10
Journal of Cereal Science 48 (2008) 203–212 Sorghum (Sorghum bicolor L. Moench) contains a XIP-type xylanase inhibitor but none of the TAXI- and TLXI-types Kurt Gebruers a, , Hind Mokrane b,c , Boubekeur Nadjemi b , Johnny Beaugrand a , Katleen Fierens a , Paul Proost d , Christophe M. Courtin a , Jan A. Delcour a a Laboratory of Food Chemistry and Biochemistry, Katholieke Universiteit Leuven, Kasteelpark Arenberg 20, B-3001 Leuven, Belgium b Laboratoire des produits bioactifs et de la valorisation de la biomasse, Ecole Normale Supe´rieure, B.P. 92, 16 050 Vieux-Kouba, Alger, Alge´rie c De´partement Ge´nie de l’environnement, Ecole Nationale Polytechnique, B.P. 182, 16200 El Harrach, Alger, Alge´rie d Laboratory of Molecular Immunology, Katholieke Universiteit Leuven, Minderbroedersstraat 10, B-3000 Leuven, Belgium Received 22 May 2007; received in revised form 23 August 2007; accepted 5 September 2007 Abstract A xylanase inhibitor of the xylanase-inhibiting protein (XIP) type was detected in sorghum (Sorghum bicolor L. Moench) whole meal using Western blot and immunoprobing with polyclonal anti-XIP antibodies. No detectable levels of Triticum aestivum xylanase inhibitor (TAXI) and thaumatin-like xylanase inhibitor (TLXI) type xylanase inhibitors were present. Trichoderma longibrachiatum xylanase affinity chromatography (AC) was used for the purification of sorghum XIP. Biochemical characteristics and protein sequence data show that sorghum XIP strongly corresponds to wheat (T. aestivum L.) XIP-I. Like wheat XIP-I, it inhibits both glycoside hydrolase family (GH) 10 and 11 xylanases, indicating that the XIP-I active site residues are well conserved in sorghum XIP. However, in contrast to wheat XIP-I, the inhibitor is unable to affect Aspergillus niger GH 11 xylanase activity. Its specific inhibition activities towards the other xylanases tested are comparable to those of wheat XIP-I. Based on the available sorghum expressed sequence tag (EST) database, XIP is expressed in sorghum in different tissues and developmental stages. Also expression in the presence of several plant hormones and under biotic as well as abiotic stress conditions is suggested. r 2007 Elsevier Ltd. All rights reserved. Keywords: Sorghum bicolor; Xylanase inhibitor; XIP; Xylanase 1. Introduction Sorghum (Sorghum bicolor L. Moench) is the fifth most important cereal crop worldwide, both in terms of planted area and metric tons harvested, and is used for various applications in food, feed, bio-fuel, paper, building materials, etc. (Berenji and Dahlberg, 2004; Taylor et al., 2006). It is a potentially important source of nutraceuticals such as antioxidant phenolics and cholesterol lowering waxes. As sorghum is a gluten-free cereal, it is suitable for coeliacs. (Awika and Rooney, 2004; Taylor et al., 2006). Glucuronoarabinoxylan (GAX) is a very important component in sorghum grain cell walls and is largely water unextractable. It is mainly composed of a backbone of (1-4)-b-linked D-xylopyranose residues with single arabinofuranose units attached at O-3 or at both O-2 and O-3 of certain xylose units. Glucuronic acid and minor amounts of galacturonic acid and 4-O-methylglucuronic acid residues are attached at O-2 of xyloses (Huisman et al., 2000; Taylor et al., 2006; Verbruggen et al., 1993, 1995). Sorghum GAX also contains acetyl and feruloyl substi- tuents (Duodu et al., 2003; Verbruggen et al., 1993). Because of its high degree of substitution, GAX is less degradable by xylanases than arabinoxylan from wheat (Triticum aestivum L.) or barley (Hordeum vulgare L.) ARTICLE IN PRESS www.elsevier.com/locate/jcs 0733-5210/$ - see front matter r 2007 Elsevier Ltd. All rights reserved. doi:10.1016/j.jcs.2007.09.005 Abbreviations: AC, affinity chromatography; CEC, cation exchange chromatography; BSA, bovine serum albumin; EST, expressed sequence tag; GAX, glucuronoarabinoxylan; GH, glycoside hydrolase family; [I]/ [E] 50 , molar inhibitor over enzyme ratio resulting in 50% inhibition; MM, molecular mass; TAXI, Triticum aestivum xylanase inhibitor; TLP, thaumatin-like protein; TLXI, thaumatin-like xylanase inhibitor; XIP, xylanase-inhibiting protein; XIU, xylanase inhibitor unit Corresponding author. Tel.: +32 16321634; fax: +32 16321997. E-mail address: [email protected] (K. Gebruers).
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Sorghum (Sorghum bicolor L. Moench) contains a XIP-type xylanase inhibitor but none of the TAXI- and...

ARTICLE IN PRESS
0733-5210/$ - se
doi:10.1016/j.jc
Abbreviations
chromatograph
tag; GAX, gluc
[E]50, molar inh
molecular mass
thaumatin-like
xylanase-inhibit�CorrespondE-mail addr
Journal of Cereal Science 48 (2008) 203–212
www.elsevier.com/locate/jcs
Sorghum (Sorghum bicolor L. Moench) contains a XIP-type xylanaseinhibitor but none of the TAXI- and TLXI-types
Kurt Gebruersa,�, Hind Mokraneb,c, Boubekeur Nadjemib, Johnny Beaugranda,Katleen Fierensa, Paul Proostd, Christophe M. Courtina, Jan A. Delcoura
aLaboratory of Food Chemistry and Biochemistry, Katholieke Universiteit Leuven, Kasteelpark Arenberg 20, B-3001 Leuven, BelgiumbLaboratoire des produits bioactifs et de la valorisation de la biomasse, Ecole Normale Superieure, B.P. 92, 16 050 Vieux-Kouba, Alger, Algerie
cDepartement Genie de l’environnement, Ecole Nationale Polytechnique, B.P. 182, 16200 El Harrach, Alger, AlgeriedLaboratory of Molecular Immunology, Katholieke Universiteit Leuven, Minderbroedersstraat 10, B-3000 Leuven, Belgium
Received 22 May 2007; received in revised form 23 August 2007; accepted 5 September 2007
Abstract
A xylanase inhibitor of the xylanase-inhibiting protein (XIP) type was detected in sorghum (Sorghum bicolor L. Moench) whole meal
using Western blot and immunoprobing with polyclonal anti-XIP antibodies. No detectable levels of Triticum aestivum xylanase
inhibitor (TAXI) and thaumatin-like xylanase inhibitor (TLXI) type xylanase inhibitors were present. Trichoderma longibrachiatum
xylanase affinity chromatography (AC) was used for the purification of sorghum XIP. Biochemical characteristics and protein sequence
data show that sorghum XIP strongly corresponds to wheat (T. aestivum L.) XIP-I. Like wheat XIP-I, it inhibits both glycoside
hydrolase family (GH) 10 and 11 xylanases, indicating that the XIP-I active site residues are well conserved in sorghum XIP. However, in
contrast to wheat XIP-I, the inhibitor is unable to affect Aspergillus niger GH 11 xylanase activity. Its specific inhibition activities
towards the other xylanases tested are comparable to those of wheat XIP-I. Based on the available sorghum expressed sequence tag
(EST) database, XIP is expressed in sorghum in different tissues and developmental stages. Also expression in the presence of several
plant hormones and under biotic as well as abiotic stress conditions is suggested.
r 2007 Elsevier Ltd. All rights reserved.
Keywords: Sorghum bicolor; Xylanase inhibitor; XIP; Xylanase
1. Introduction
Sorghum (Sorghum bicolor L. Moench) is the fifth mostimportant cereal crop worldwide, both in terms of plantedarea and metric tons harvested, and is used for variousapplications in food, feed, bio-fuel, paper, buildingmaterials, etc. (Berenji and Dahlberg, 2004; Taylor et al.,2006). It is a potentially important source of nutraceuticals
e front matter r 2007 Elsevier Ltd. All rights reserved.
s.2007.09.005
: AC, affinity chromatography; CEC, cation exchange
y; BSA, bovine serum albumin; EST, expressed sequence
uronoarabinoxylan; GH, glycoside hydrolase family; [I]/
ibitor over enzyme ratio resulting in 50% inhibition; MM,
; TAXI, Triticum aestivum xylanase inhibitor; TLP,
protein; TLXI, thaumatin-like xylanase inhibitor; XIP,
ing protein; XIU, xylanase inhibitor unit
ing author. Tel.: +3216321634; fax: +3216321997.
ess: [email protected] (K. Gebruers).
such as antioxidant phenolics and cholesterol loweringwaxes. As sorghum is a gluten-free cereal, it is suitable forcoeliacs. (Awika and Rooney, 2004; Taylor et al., 2006).Glucuronoarabinoxylan (GAX) is a very important
component in sorghum grain cell walls and is largelywater unextractable. It is mainly composed of a backboneof (1-4)-b-linked D-xylopyranose residues with singlearabinofuranose units attached at O-3 or at both O-2 andO-3 of certain xylose units. Glucuronic acid and minoramounts of galacturonic acid and 4-O-methylglucuronicacid residues are attached at O-2 of xyloses (Huisman et al.,2000; Taylor et al., 2006; Verbruggen et al., 1993, 1995).Sorghum GAX also contains acetyl and feruloyl substi-tuents (Duodu et al., 2003; Verbruggen et al., 1993).Because of its high degree of substitution, GAX is lessdegradable by xylanases than arabinoxylan from wheat(Triticum aestivum L.) or barley (Hordeum vulgare L.)

ARTICLE IN PRESSK. Gebruers et al. / Journal of Cereal Science 48 (2008) 203–212204
endosperms (Taylor et al., 2006; Verbruggen et al., 1995).Hence, extensive degradation of GAX requires the useof proper xylanases in combination with accessoryenzymes such as arabinofuranosidases, glucuronidasesand esterases.
Xylanases are already routinely used in cereal proces-sing, e.g. in bread making (Courtin and Delcour, 2002) andin gluten–starch separation (Frederix et al., 2004). How-ever, their efficiency is often lowered by the presence ofproteinaceous xylanase inhibitors in cereals (Sørensenet al., 2004). So far, three types of xylanase inhibitors havebeen studied, i.e. Triticum aestivum xylanase inhibitor(TAXI), xylanase-inhibiting protein (XIP) and thaumatin-like xylanase inhibitor (TLXI)-type proteins. TAXI-typexylanase inhibitors have a molecular mass (MM) of ca.40 kDa and occur in bread wheat, durum wheat (Triticum
durum Desf.), barley and rye (Secale cereale L.). They areactive towards glycoside hydrolase family (GH) 11xylanases (Gebruers et al., 2001, 2002a; Goesaert et al.,2003a). XIP proteins (ca. 30 kDa) of these cereals as well asof rice (Oryza sativa L.) and maize (Zea mays L.) have alsobeen studied. They possess two separate active sites forinhibiting GH 10 and GH 11 xylanases (Durand et al.,2005; Gebruers et al., 2002b; Goesaert et al., 2003b;McLauchlan et al., 1999; Payan et al., 2004). To date, taxi
and xip genes have already been identified and characteri-zed in several of the above cited cereals (Durand et al.,2005; Elliott et al., 2002; Fierens et al., 2004; Raedschelderset al., 2004, 2005). TAXI- and XIP-type xylanase inhibitorsare known to be competitive inhibitors. Crystallographicstudies of inhibitor–enzyme complexes (Payan et al., 2004;Sansen et al., 2004) and the analysis of inhibitor andenzyme mutants (Fierens et al., 2005; Tahir et al., 2002,2004) demonstrated that both types of inhibitors directlyinteract with the active sites of their target enzymes. So far,TLXI (ca. 18 kDa) has only been studied in bread wheat.Like TAXI-type inhibitors, TLXI only inhibits GH 11xylanases. However, it has a different xylanase specificitythan TAXI (Fierens et al., 2007). Based on immunoprobingexperiments, TLXI homologs have been detected in durumwheat and rye (Beaugrand et al., 2006).
The present report describes for the first time thedetection, purification and characterization of XIP proteinsfrom sorghum whole meal. Indeed, while Elliott et al.(2003) already suggested the presence of three putative XIPhomologous nucleotide sequences in sorghum, they wereunable to detect XIP-type inhibitors in the sorghum wholemeal sample they examined using Western blot analysisand immunoprobing.
2. Experimental
2.1. Materials
A commercial sorghum sample (Beyers Granen,Schoten, Belgium) was ground into whole meal using aCyclotec 1093 sample mill (Tecator, Hoganas, Sweden).
All electrophoresis and chromatography media, and MMand pI markers were from GE Healthcare (Uppsala,Sweden), unless specified otherwise. Bacillus subtilis
GH 11 xylanase was supplied by Eng. Filip Arnaut(NV Puratos, Groot-Bijgaarden, Belgium). Two GH 11xylanases from Trichoderma longibrachiatum (also knownas T. reesei), one with a pI value of 5.5 and the other with apI value of 9.0, GH 11 xylanases from Aspergillus niger andTrichoderma viride, and Xylazyme-AX tablets were fromMegazyme (Bray, Ireland). GH 10 xylanases from Asper-
gillus oryzae and Penicillium purpurogenum were kindlymade available by Prof. Maija Tenkanen (VTT Biotech-nology and the University of Helsinki, Finland) andProf. Jaime Eyzaguirre (Laboratorio de Bioquimıca,Facultad de Ciencias Biologicas, Pontificia UniversidadCatolica de Chile, Chile), respectively. A. niger GH 10xylanase was purified from an A. niger CBS 110.42culture filtrate (Gebruers et al., 2005). Goat anti-rabbitantibodies conjugated to horseradish peroxidase, substrate(3,30,5,50-tetramethylbenzidine) for the horseradish peroxi-dase and all other chemicals were from Sigma-Aldrich(Bornem, Belgium).
2.2. Affinity matrix preparation
Before immobilization, the Megazyme T. longibrachiatum
GH 11 xylanase (pI 5.5) was dialysed overnight againstdeionized water. N-hydroxysuccinimide activated Sepharose4 Fast Flow matrix was used for the preparation of affinitymatrix (5.0ml) according to Gebruers et al. (2002a, b).
2.3. Purification of xylanase inhibitor
Sorghum whole meal (1.0 kg) was suspended in aqueous0.1% (w/v) L-ascorbic acid solution (5.0 l), stirred overnightat 7 1C and centrifuged (10 000g, 30min, 7 1C). L-Ascorbicacid was used to reduce the oxidation of phenoliccompounds during the extraction. The pH of the super-natant was adjusted to 4.5 by adding 2.0M HCl.Proteins with xylanase inhibition activity in the super-
natant were retained by cation exchange chromatography(CEC) on a SP Sepharose Big Beads column[180� 130mm2, equilibrated with sodium acetate buffer(25mM, pH 4.5)]. The bound protein fraction was eluted inone step with NaCl solution (1.0M, 1.0 l) resulting in theCEC protein fraction. Portions of the CEC fraction(50.0ml) were diluted 1:2 in sodium acetate buffer(25mM, pH 5.0) and loaded on the T. longibrachiatum
xylanase affinity column [10� 65mm, equilibrated withsodium acetate buffer (25mM, pH 5.0, containing 0.2MNaCl), flow rate of 0.33ml/min]. XIP proteins were elutedfrom the column with Tris solution (250mM, pH 11.0,5.0ml, flow rate of 1.0ml/min) and immediately neutra-lized with acetic acid solution (1.0M). The protein fractioneluted during affinity chromatography (AC) is furtherreferred to as the AC protein fraction or purified sorghumXIP (vide infra).

ARTICLE IN PRESSK. Gebruers et al. / Journal of Cereal Science 48 (2008) 203–212 205
2.4. Xylanase inhibition assay
Inhibition activities were determined with the colori-metric Xylazyme-AX method (Gebruers et al., 2001). Allxylanase solutions were prepared in sodium acetatebuffer (25mM, pH 5.0) with bovine serum albumin(BSA, 0.5mg/ml) and contained 2.0 xylanase units per1.0ml. Xylanase units were as defined by Gebruers et al.(2002a, b) and, under the conditions of the assay, thexylanase concentrations corresponding to 1.0 unit wereapproximately 5.1 nM for the A. niger, 8.9 nM for theT. longibrachiatum (pI 5.5), 2.1 nM for the T. long-
ibrachiatum (pI 9.0), 11.6 nM for the T. viride and 5.8 nMfor the B. subtilis GH 11 xylanases. The correspondingconcentrations for the GH 10 xylanases were 36.6 nM forthe A. niger, 2.9 nM for the A. oryzae and 17.7 nM for theP. purpurogenum enzymes. All measurements were per-formed in triplicate. The inhibition activity towards axylanase is often expressed in xylanase inhibitor units(XIUs), corresponding to the amount of inhibitor neededto inhibit that xylanase for 50% in the Xylazyme-AXmethod. The [I]/[E]50, i.e. the molar inhibitor over enzymeratio resulting in 50% inhibition, is a measure of theefficiency of an inhibitor.
To determine the temperature and pH conditions formaximal inhibition activity, the Xylazyme-AX assay wasperformed at different temperatures at pH 5.0 and differentpH conditions at 40 1C, respectively. For the latter,universal buffers of pH 3.0–8.0 instead of the above citedsodium acetate buffer were used. The buffers wereprepared from a stock solution of 30mM citric acid,30mM KH2PO4, 30mM H3BO3 and 20mM diethylbarbituric acid by adding HCl or NaOH (2.0M) solutions.
2.5. Electrophoresis
SDS-PAGE under reducing conditions was performed on20% polyacrylamide gels with a PhastSystem unit, asdescribed in the GE Healthcare separation technique file110. 2-Mercaptoethanol (5%) was used as reducing agent.The GE Healthcare low MM markers were used. The gelswere Coomassie stained as described in the GE Healthcaredevelopment technique file 200. The pI value of the inhibitorprotein was determined by isoelectric focusing as describedin the GE Healthcare separation technique file 100, using thePhastSystem unit with polyacrylamide gels containingampholytes (pH 3–9). The broad range pI markers(3.5–9.3) were used. The gels were silver stained as describedin the GE Healthcare development technique file 210.
2.6. Protein content determination
Protein concentrations were determined by the BradfordCoomassie brilliant blue method with BSA as standard(Bradford, 1976). For pure sorghum XIP samples, proteinconcentrations were determined spectrophotometrically at280 nm using a specific absorbance value of 2.348AU for
1mg/ml XIP (1.000 cm UV-cell path length), calculatedfrom the complete sorghum XIP amino acid sequence (videinfra) using the ProtParam tool (Gasteiger et al., 2005).
2.7. Western blot and immunoprobing
Rabbit polyclonal antibodies (PAbs) against nativewheat TAXI, XIP and TLXI were prepared and Westernblot analysis was performed as described by Beaugrandet al. (2006). After SDS-PAGE and Western blotting,sorghum whole meal extract proteins (40 mg loaded) wereprobed with anti-TAXI, anti-TLXI and anti-XIP PAbs.CEC fraction proteins (50 mg loaded) were probed withanti-TLXI and anti-XIP PAbs. Finally, AC fractionprotein ( ¼ purified sorghum XIP, 2.5 mg loaded) wasprobed with anti-TLXI and anti-XIP PAbs. Purified wheatTAXI, XIP and TLXI (for all 0.25 mg loaded) were used aspositive controls.
2.8. Protein sequencing
To determine the N-terminal amino acid sequence,inhibitor protein (ca. 50 mg) was separated under reducingconditions (5% 2-mercaptoethanol) with SDS-PAGE in a12% polyacrylamide gel using the Hoeffer Mighty Smallunit (GE Healthcare) (electric potential difference of 160V,1 h, room temperature), electro blotted onto a PVDFmembrane with the Trans-Blot Semi-Dry ElectrophoreticTransfer Cell (Bio-Rad, Hercules, CA, USA) (electricpotential difference of 10V, 1 h, room temperature) andsubjected to Edman degradation after protein visualizationby Coomassie staining. Sequence analysis was performedon a Procise cLC 491 Sequencer (Applied Biosystems,Foster City, CA, USA).
2.9. DNA and protein sequence analysis
The experimentally determined N-terminal amino acidsequence was analyzed against the available S. bicolor
expressed sequence tag (EST) database using the tBLASTnsearch engine (Altschul et al., 1997). Prediction of thesignal sequence cleavage site in the complete sorghum XIPpreprotein amino acid sequence and the cellular localiza-tion of the inhibitor was performed with the SignalP 3.0(Nielsen and Krogh, 1998) and Psort (Horton et al., 2006)tools, respectively. N- and O-glycosylation sites werepredicted with the NetNGlyc 1.0 (Blom et al., 2004) andNetOGlyc 3.1 (Julenius et al., 2005) servers, respectively.Sequence alignments were performed with the ClustalWprogram (Higgins et al., 1994).
3. Results and discussion
3.1. Xylanase inhibitor detection in sorghum grains
Western blot analysis and immunoprobing revealed thepresence of XIP-type xylanase inhibitors in sorghum whole
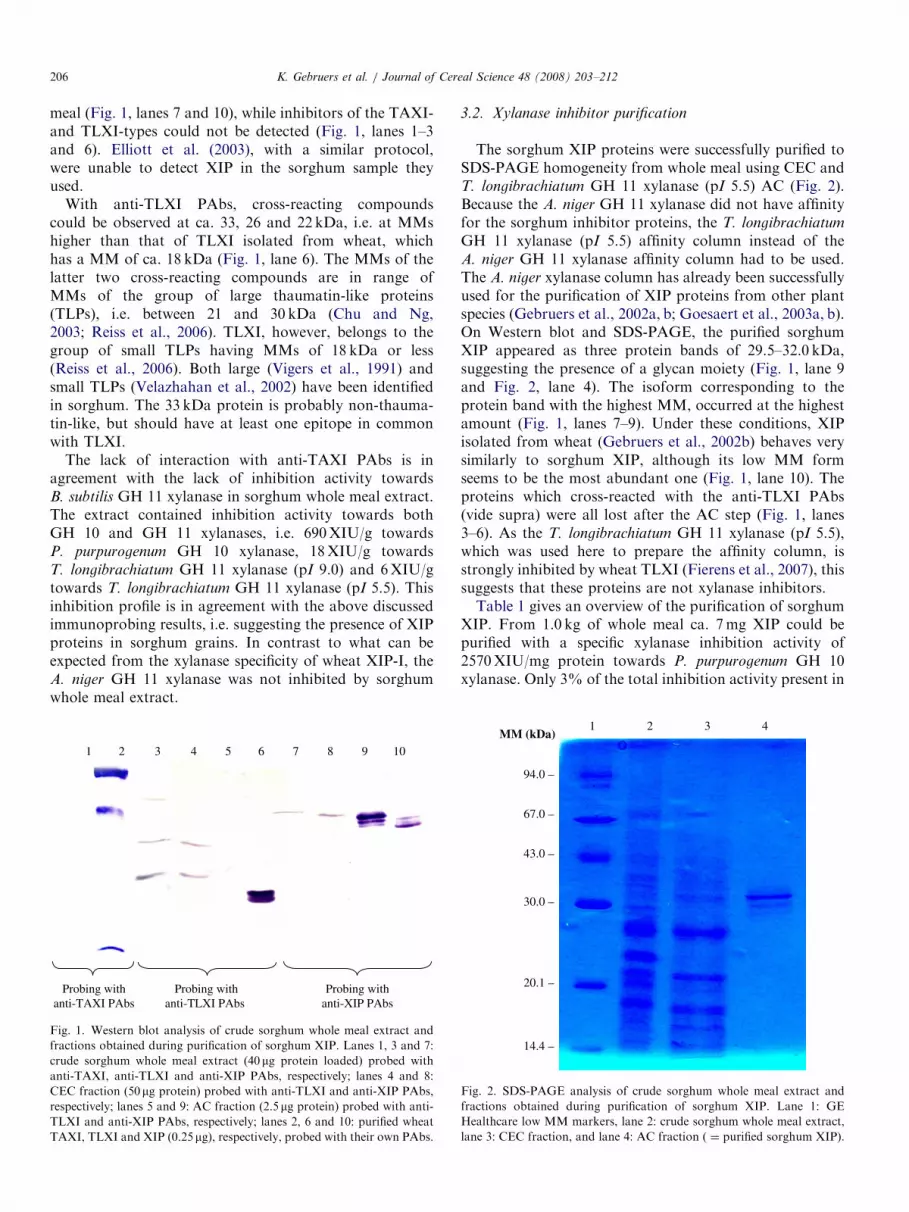
ARTICLE IN PRESSK. Gebruers et al. / Journal of Cereal Science 48 (2008) 203–212206
meal (Fig. 1, lanes 7 and 10), while inhibitors of the TAXI-and TLXI-types could not be detected (Fig. 1, lanes 1–3and 6). Elliott et al. (2003), with a similar protocol,were unable to detect XIP in the sorghum sample theyused.
With anti-TLXI PAbs, cross-reacting compoundscould be observed at ca. 33, 26 and 22 kDa, i.e. at MMshigher than that of TLXI isolated from wheat, whichhas a MM of ca. 18 kDa (Fig. 1, lane 6). The MMs of thelatter two cross-reacting compounds are in range ofMMs of the group of large thaumatin-like proteins(TLPs), i.e. between 21 and 30 kDa (Chu and Ng,2003; Reiss et al., 2006). TLXI, however, belongs to thegroup of small TLPs having MMs of 18 kDa or less(Reiss et al., 2006). Both large (Vigers et al., 1991) andsmall TLPs (Velazhahan et al., 2002) have been identifiedin sorghum. The 33 kDa protein is probably non-thauma-tin-like, but should have at least one epitope in commonwith TLXI.
The lack of interaction with anti-TAXI PAbs is inagreement with the lack of inhibition activity towardsB. subtilis GH 11 xylanase in sorghum whole meal extract.The extract contained inhibition activity towards bothGH 10 and GH 11 xylanases, i.e. 690XIU/g towardsP. purpurogenum GH 10 xylanase, 18XIU/g towardsT. longibrachiatum GH 11 xylanase (pI 9.0) and 6XIU/gtowards T. longibrachiatum GH 11 xylanase (pI 5.5). Thisinhibition profile is in agreement with the above discussedimmunoprobing results, i.e. suggesting the presence of XIPproteins in sorghum grains. In contrast to what can beexpected from the xylanase specificity of wheat XIP-I, theA. niger GH 11 xylanase was not inhibited by sorghumwhole meal extract.
1
Probing with anti-TAXI PAbs
Probing with anti-TLXI PAbs
2 3 5 4 6 7 8 9 10
Probing with anti-XIP PAbs
Fig. 1. Western blot analysis of crude sorghum whole meal extract and
fractions obtained during purification of sorghum XIP. Lanes 1, 3 and 7:
crude sorghum whole meal extract (40mg protein loaded) probed with
anti-TAXI, anti-TLXI and anti-XIP PAbs, respectively; lanes 4 and 8:
CEC fraction (50mg protein) probed with anti-TLXI and anti-XIP PAbs,
respectively; lanes 5 and 9: AC fraction (2.5mg protein) probed with anti-
TLXI and anti-XIP PAbs, respectively; lanes 2, 6 and 10: purified wheat
TAXI, TLXI and XIP (0.25mg), respectively, probed with their own PAbs.
3.2. Xylanase inhibitor purification
The sorghum XIP proteins were successfully purified toSDS-PAGE homogeneity from whole meal using CEC andT. longibrachiatum GH 11 xylanase (pI 5.5) AC (Fig. 2).Because the A. niger GH 11 xylanase did not have affinityfor the sorghum inhibitor proteins, the T. longibrachiatum
GH 11 xylanase (pI 5.5) affinity column instead of theA. niger GH 11 xylanase affinity column had to be used.The A. niger xylanase column has already been successfullyused for the purification of XIP proteins from other plantspecies (Gebruers et al., 2002a, b; Goesaert et al., 2003a, b).On Western blot and SDS-PAGE, the purified sorghumXIP appeared as three protein bands of 29.5–32.0 kDa,suggesting the presence of a glycan moiety (Fig. 1, lane 9and Fig. 2, lane 4). The isoform corresponding to theprotein band with the highest MM, occurred at the highestamount (Fig. 1, lanes 7–9). Under these conditions, XIPisolated from wheat (Gebruers et al., 2002b) behaves verysimilarly to sorghum XIP, although its low MM formseems to be the most abundant one (Fig. 1, lane 10). Theproteins which cross-reacted with the anti-TLXI PAbs(vide supra) were all lost after the AC step (Fig. 1, lanes3–6). As the T. longibrachiatum GH 11 xylanase (pI 5.5),which was used here to prepare the affinity column, isstrongly inhibited by wheat TLXI (Fierens et al., 2007), thissuggests that these proteins are not xylanase inhibitors.Table 1 gives an overview of the purification of sorghum
XIP. From 1.0 kg of whole meal ca. 7mg XIP could bepurified with a specific xylanase inhibition activity of2570XIU/mg protein towards P. purpurogenum GH 10xylanase. Only 3% of the total inhibition activity present in
1 2 4MM (kDa)
94.0 –
67.0 –
43.0 –
30.0 –
20.1 –
14.4 –
3
Fig. 2. SDS-PAGE analysis of crude sorghum whole meal extract and
fractions obtained during purification of sorghum XIP. Lane 1: GE
Healthcare low MM markers, lane 2: crude sorghum whole meal extract,
lane 3: CEC fraction, and lane 4: AC fraction ( ¼ purified sorghum XIP).

ARTICLE IN PRESS
Table 1
Overview of the purification of XIP from sorghum whole meal monitored by assessment of total protein and inhibition activity towards P. purpurogenum
GH 10 xylanase. Specific inhibition activity and recovery are also presented
Total protein (mg/kg) Total inhibition activity (� 103XIU/kg) Specific inhibition activity (XIU/mg) Recovery (%)
Crude extract 800 690 860 100
CECa fraction 470 550 1170 80
CEC-RTb fraction 170 50 290 7
ACc fraction 7 18 2570 3
AC-RT fraction 390 12 30 2
aCEC ¼ cation exchange chromatography.bRT ¼ run-through.cAC ¼ affinity chromatography.
Table 2
Sorghum ESTsa used to derive the complete nucleotide and amino acid sequence of sorghum XIP
GenBank id Size (nt) Stage/tissue Growth conditions Variantb
N-terminal
CX614867 572 Seedlings GA- or brassinolide-treated V
CD207904 558 Seedlings Heat-shocked A
CD423209 540 Seedlings Salicylic acid-treated V
CD227733 483 Callus culture/cell suspension n.s.c A
CB925367 531 Seedlings Abscissic acid-treated A
CD462578 453 Seedlings Ethylene-treated V
AW747051 450 n.s. Water-stressed V
BE595815 372 n.s. Pathogen-induced V
CN148778 600 Leaves Wounded V
CN141616 573 Roots Oxidative-stressed V
CN127196 651 Roots Acid/alkaline-treated V
Internal
AW746639 582 n.s. Water-stressed V
BE594070 540 n.s. Water-stressed V
C-terminal
CF755785 480 Flowering sorghum Drought-stressed A
BG356033 471 Embryo n.s. V
BE593758 474 n.s Water-stressed V
CD423096 438 Seedlings Salicylic acid-treated n.a.d
CD223474 438 Callus culture/cell suspension n.s. n.a.
CD427895 411 Seedlings Ethylene-treated n.a.
CN129911 633 Roots Acid/alkaline-treated V
aESTs from Genbank release 158.bThe amino acid residue at position 118 is either valine or alanine. In the overlapping regions between different ESTs all other residues are identical.cn.s. ¼ not specified.dn.a. ¼ not available.
K. Gebruers et al. / Journal of Cereal Science 48 (2008) 203–212 207
sorghum grains was recovered. The loss in inhibitionactivity is probably attributed to inactivation duringintermediate freezing and storage of the CEC fraction.After thawing, part of the protein precipitated.
3.3. Sequence determination
N-terminal amino acid sequence analysis of the purifiedsorghum XIP isoforms corresponding to the three bandson SDS-PAGE, each time yielded the same sequence,i.e. AGXKTGQVTVFWGRNKAEGTLREAXDSGLYT(X is an unidentified amino acid). A tBLASTn search
against the available S. bicolor EST database using thissequence yielded several matching nucleotide sequences.Assembly of these sequences yielded the complete signalsequence in addition to approximately two-thirds of thecomplete amino acid sequence of the mature inhibitorprotein. A second tBLASTn search using the assembledprotein sequence yielded additional EST sequences corre-sponding to internal regions and the C-terminus of thesorghum XIP protein. Representative ESTs with 100%identity in their overlapping regions are listed in Table 2.Their assembly allowed construction of the completeamino acid sequence and corresponding nucleotide

ARTICLE IN PRESS
Fig. 3. Nucleotide and amino acid sequences of the valine variant of sorghum XIP, including the signal sequence. The shaded sequence denotes the
experimentally confirmed N-terminal amino acid sequence. The potential N-glycosylation sites are presented in bold. The signal sequence is underlined and
the stop codon is marked by an asterisk.
K. Gebruers et al. / Journal of Cereal Science 48 (2008) 203–212208
sequences (Fig. 3). The mature sorghum XIP proteinconsists of 272 amino acids and is preceded by a signalsequence of 35 amino acids. The start of the mature proteinwas predicted with the SignalP 3.0 program (Nielsen andKrogh, 1998) and corresponded to the experimentallyobtained first amino acid (vide supra). According to thePsort tool (Horton et al., 2006), sorghum XIP is most likelylocated extracellularly. The complete preprotein is codedby a 924 bp nucleotide sequence. Some ESTs suggest theoccurrence of two variants, i.e. one having a valine residueat position 118 and another having an alanine residue atthis position. The calculated MM is 30,269Da for thevaline variant and 30,241Da for the alanine variant. Bothvariants have a calculated pI value of 6.11, while, inisoelectric focusing, a pI value of ca. 6.7 was observed(results not shown). The sorghum EST library suggests thatsorghum XIP is expressed at different developmentalstages, in various tissues, after treatment of seedlingswith plant hormones and under abiotic stress conditions(Table 2).
According to the NetNGlyc 1.0 tool (Blom et al., 2004),the sorghum XIP amino acid sequence contains threepotential N-glycosylation sites, i.e. at positions 3, 132 and263 (Fig. 3). This is in line with the occurrence of differentinhibitor isoforms on SDS-PAGE and Western blot, andmight explain why the third amino acid residue, i.e. Asn,could not be detected in the Edman N-terminal amino acidanalysis (vide supra). As suggested by the NetOGlyc 3.1
tool (Julenius et al., 2005), the inhibitor probably does notcarry O-linked glycan moieties. On the protein level,sorghum XIP shows homology with a putative Z. mays
XIP (DQ245902, 92% similarity), a putative Oryza sativa
XIP (CT836240, 92% similarity), T. aestivum XIP-I(AJ422119, 85% similarity) and XIP-III (AB204556, 84%similarity) (Elliott et al., 2003), T. durum XIP-II(AJ318884, 66% similarity) (Elliott et al., 2003), RIXI[an O. sativa XIP protein, AB027415, 65% similarity(Durand et al., 2005)], and another O. sativa XIP(AK064356, 66% similarity) (Goesaert et al., 2005).Fig. 4 shows the phylogenetic tree giving the relationshipsbetween these sequences.Alignment of the amino acid sequence of these XIP
proteins in ClustalW (Higgins et al., 1994) shows that insorghum XIP the regions responsible for interaction withGH 10 and GH 11 xylanases can be recognized, similar tothose identified in wheat XIP-I by Payan et al. (2004)(Fig. 5). In particular, the key amino acid residues (Lys232and Arg150, respectively), which interact with the catalyticresidues of the enzymes and the residues surroundingthem are identical. The largest differences between wheatXIP-I and sorghum XIP are present in region 4 and thefirst part of region 5 (Fig. 5), which participate in theinteraction with GH 10 xylanases. However, becausesorghum XIP has a comparable inhibition activity towardsA. oryzae GH 10 xylanase (vide infra), these differences donot dramatically affect XIP activity. Larger amino acid

ARTICLE IN PRESS
S_bicolor_XIPZ_mays_XIP_putative
O_sativa_XIP_putative O_sativa_XIP
O_sativa_RIXI T_durum_XIP_II
T_aestivum_XIP_I T_aestivum_XIP_III
Fig. 4. Phylogenetic tree showing the relationship between different XIP sequences. The phylogenetic tree [produced with the ClustalW program (Higgins
et al., 1994)] shows the relationship between sequences of S. bicolor XIP, a putative XIP of Z. mays (DQ245902), O. sativa XIP proteins [RIXI
(AB027415), another rice XIP (AK064356) and a putative XIP (CT836240)], and XIPs from T. aestivum [XIP-I (AJ422119) and XIP-III (AB204556)] and
T. durum [XIP-II (AJ318884)].
Fig. 5. Alignment of the amino acid sequence of S. bicolor XIP with XIP sequences of other cereals. The S. bicolor XIP amino acid sequence is aligned
with sequences of a putative XIP of Z. mays (DQ245902), O. sativa XIP proteins [RIXI (AB027415), another rice XIP (AK064356) and a putative XIP
(CT836240)], and XIPs from T. aestivum [XIP-I (AJ422119) and XIP-III (AB204556)] and T. durum [XIP-II (AJ318884)]. The regions involved in the
interaction with GH 10 and GH 11 xylanases are shaded in dark (regions 4–6) and light gray (regions 1–3), respectively. The key residues which interact
with the active site residues of the xylanases are underlined [based on crystallographical data available for XIP-I (Payan et al., 2004)]. Clustering was
performed using the ClustalW program (Higgins et al., 1994). For XIP-II, XIP-III and the putative Z. mays and O. sativa XIP proteins the start of the
mature protein was predicted using the SignalP 3.0 server (Nielsen and Krogh, 1998).
K. Gebruers et al. / Journal of Cereal Science 48 (2008) 203–212 209
sequence differences can be observed between sorghumXIP on the one hand and RIXI, another rice XIP anddurum wheat XIP-II on the other hand. The latterconstitute a subfamily of xylanase inhibitors within GH18 (Durand et al., 2005).
3.4. Biochemical characterization
As already mentioned above, sorghum XIP inhibits bothGH 10 and GH 11 xylanases, just like wheat XIP-I(Flatman et al., 2002; Payan et al., 2004). This is in

ARTICLE IN PRESS
0
20
40
60
80
100
40 50 60
Rela
tive x
yla
nase in
hib
itio
n
acti
vit
y (
%)
0
20
40
60
80
100
0
20
40
60
80
100
3 5 6 7
Rela
tive x
yla
nase in
hib
itio
n
acti
vit
y (
%)
0
20
40
60
80
pH
Rela
tive x
yla
nase
acti
vit
y (
%)
Rela
tive x
yla
nase a
cti
vit
y (
%)
20 30
T (°C)
70
4 8
100
(A)
(B)
Fig. 6. Relative xylanase and xylanase inhibition activity at varying
temperature and pH conditions. Inhibition activity of sorghum XIP
towards P. purpurogenum GH 10 xylanase (~) and activity of the enzyme
(’) were determined at different temperature (A) and pH conditions (B)
K. Gebruers et al. / Journal of Cereal Science 48 (2008) 203–212210
agreement with the conservation of the two active sites ofthe inhibitor responsible for the inhibition of thesexylanase families (vide supra). Based on biochemicalstudies (Flatman et al., 2002; McLauchlan et al., 1999)and crystal structures (Payan et al, 2004), both sites act in acompetitive mode.
The [I]/[E]50-values measured with the GH 10 xylanasesvaried between 0.9 and 2.2 (Table 3). These low values areindicative for tight-binding inhibition (Morrison, 1982)and are in the order of magnitude also observed byFlatman et al. (2002) for wheat XIP-I. They reported [I]/[E]50-values of 0.6 and 1.5 for the GH 10 xylanases ofAspergillus nidulans and A. oryzae, respectively. Also forthe GH 11 xylanases comparable [I]/[E]50-values wereobserved (Table 3). However, for the A. niger GH 11xylanase no inhibition could be detected under theexperimental conditions, whereas Flatman et al. (2002)describe a [I]/[E]50-value for this xylanase and wheat XIP-Iof 3.8 (Table 3). A double amino acid substitution inregion 1 (Fig. 5), involved in the interaction with GH 11xylanases, might be the cause for this different behaviortowards the A. niger enzyme compared to wheat XIP-I.However, additional experimental work needs to bedone to substantiate this hypothesis. In case of TAXI, [I]/[E]50-values of 0.6–1.1 have been reported. For theTAXI-II isoform and xylanases with low pH optimum,much higher values, or even no inhibition, are expected(Gebruers et al, 2004; Raedschelders et al., 2005). [I]/[E]50-values of 4.2 up to 289.6 are determined for TLXI (Fierenset al., 2007).
Table 3
[I]/[E]50 values for the inhibition of different GH 10 and GH 11 xylanases
by sorghum XIP compared with values reported for wheat XIP-I
NCBI accession no. [I]/[E]50
Sorghum
XIP
Wheat
XIP-Ia
GH 10 xylanases
Aspergillus niger CAA03655 0.9 n.a.b
Penicillium
purpurogenum
AAF71268 1.3 n.a.
Aspergillus oryzae BAA75475 2.2 1.5
GH 11 xylanases
Aspergillus niger CAA01470 n.i.c 3.8
Trichoderma
longibrachiatum (pI
5.5)
CAA46294 104.0 n.a.
Trichoderma
longibrachiatum (pI
9.0)
CAA49293 6.1 5.3
Trichoderma viride CAB60757 37.0 63.0
Bacillus subtilis AAA22897 n.i. n.i.
Penicillium funiculosum CAC15487 n.d.d 1.0
aData described by Flatman et al. (2002).bn.a. ¼ not available.cn.i. ¼ not inhibited.dn.d. ¼ not determined.
using the Xylazyme-AX method. Relative activities are presented.
The GH 10 P. purpurogenum xylanase is maximallyinhibited by sorghum XIP at 35–45 1C and pH 5.7–6.7,whereas the enzyme is optimally active at 50–55 1C and pHca. 5.7 (Fig. 6). As for wheat XIP-I and A. niger GH 11xylanase, the inhibition activity increased from pH 4.5 to6.5 (Flatman et al., 2002). TAXI proteins are optimallyactive at ca. pH 5.0, and at 30–40 1C for A. niger GH 11xylanase and 20–30 1C for B. subtilis GH 11 xylanase(Gebruers et al., 2004).In conclusion, sorghum grains contain XIP proteins with
protein sequence data and biochemical properties similarto those of wheat XIP-I. No TAXI and TLXI proteinscould be detected in the grains. Insight into the presenceand properties of xylanase inhibitors in sorghum may helpto understand the functionality of xylanases in theproduction of sorghum-based products.
Acknowledgements
This study was in part carried out in framework ofresearch project GOA/03/10 financed by the ResearchFund K.U. Leuven. The authors gratefully acknowledgefinancial support from the ‘Fonds voor WetenschappelijkOnderzoek-Vlaanderen’ (F.W.O.-Vlaanderen, Brussels,Belgium) (Post-doctoral fellowship of K. Gebruers), the

ARTICLE IN PRESSK. Gebruers et al. / Journal of Cereal Science 48 (2008) 203–212 211
Research Fund K.U. Leuven (Leuven, Belgium) (Post-doctoral fellowship of J. Beaugrand) and the BelgiumTechnical Cooperation (BTC, Brussels, Belgium) (Ph.D.scholarship of H. Mokrane in Belgium). E. Peeters (BeyersGranen, Schoten, Belgium) is thanked for supplyingsorghum.
References
Altschul, S.F., Madden, T.L., Schaffer, A.A., Zhang, J., Zhang, Z., Miller,
W., Lipman, D.J., 1997. Gapped BLAST and PSI-BLAST: a new
generation of protein database search programs. Nucleic Acids
Research 25, 3389–3402.
Awika, J.A., Rooney, L.W., 2004. Sorghum phytochemicals and their
potential impact on human health. Phytochemistry 65, 1199–1221.
Beaugrand, J., Gebruers, K., Ververken, C., Fierens, E., Croes, E.,
Goddeeris, B., Courtin, C.M., Delcour, J.A., 2006. Antibodies against
wheat xylanase inhibitors provide a tool for the selective probing and
quantification of xylanase inhibitors in different cereals. Journal of
Cereal Science 44, 59–67.
Berenji, J., Dahlberg, J., 2004. Perspectives of sorghum in Europe. Journal
of Agronomy and Crop Science 190, 332–338.
Blom, N., Sicheritz-Ponten, T., Gupta, R., Gammeltoft, S., Brunak, S.,
2004. Prediction of posttranslational glycosylation and phosphoryla-
tion of proteins from the amino acid sequence. Proteomics 4,
1633–1649.
Bradford, M.M., 1976. A rapid and sensitive method for the quantifica-
tion of microgram quantities of protein utilizing the principle of
protein–dye binding. Analytical Biochemistry 72, 248–254.
Chu, K.T., Ng, T.B., 2003. Isolation of a large thaumatin-like antifungal
protein from seeds of the Kweilin chestnut Castanopsis chinensis.
Biochemical and Biophysical Research Communications 301, 364–370.
Courtin, C.M., Delcour, J.A., 2002. Arabinoxylans and endoxylanases in
wheat flour bread-making. Journal of Cereal Science 35, 225–243.
Duodu, K.G., Taylor, J.N.R., Belton, P.S., Hamaker, B.R., 2003. Factors
affecting sorghum protein digestibility. Journal of Cereal Science 38,
117–131.
Durand, A., Hugher, R., Roussel, A., Flatman, R., Henrissat, B., Juge,
N., 2005. Emergence of a subfamily of xylanase inhibitors within
glycoside hydrolase family 18. FEBS Journal 272, 1745–1755.
Elliott, G.O., Hughes, R.K., Juge, N., Kroon, P.A., Williamson, G., 2002.
Functional identification of the cDNA coding for a wheat endo-1,4-
L-D-xylanase inhibitor. FEBS Letters 519, 66–70.
Elliott, G.O., McLauchlan, W.R., Williamson, G., Kroon, P.A., 2003. A
wheat xylanase inhibitor protein (XIP-I) accumulates in the grain and
has homologues in other cereals. Journal of Cereal Science 37,
187–194.
Fierens, K., Geudens, N., Brijs, K., Courtin, C.M., Gebruers, K., Robben,
J., Van Campenhout, S., Volckaert, G., Delcour, J.A., 2004. High-level
expression, purification, and characterization of recombinant wheat
xylanase inhibitor TAXI-I secreted by the yeast Pichia pastoris. Protein
Expression and Purification 37, 39–46.
Fierens, K., Gils, A., Sansen, S., Brijs, K., Courtin, C.M., Declerck, P.J.,
De Ranter, C.J., Gebruers, K., Rabijns, A., Robben, J., Van
Campenhout, S., Volckaert, G., Delcour, J.A., 2005. His374 of wheat
endoxylanase inhibitor TAXI-I stabilizes complex formation with
glycoside hydrolase family 11 endoxylanases. FEBS Journal 272,
5872–5882.
Fierens, E., Rombouts, S., Gebruers, K., Courtin, C.M., Goesaert, H.,
Brijs, K., Volckaert, G., Van Campenhout, S., Delcour, J.A., 2007.
TLXI, a novel type of xylanase inhibitor from wheat (Triticum
aestivum) belonging to the thaumatin family. Biochemical Journal 403,
583–591.
Flatman, R., McLauchlan, W.R., Juge, N., Furniss, C., Berrin, J.-G.,
Hughes, R.K., Manzanares, P., Ladbury, J.E., O’Brien, R., William-
son, G., 2002. Interaction defining the specificity between fungal
xylanases and the xylanase-inhibiting protein XIP-I from wheat.
Biochemical Journal 365, 773–781.
Frederix, S.A., Courtin, C.M., Delcour, J.A., 2004. Substrate selectivity
and inhibitor sensitivity affect xylanase functionality in wheat flour
gluten-starch separation. Journal of Cereal Science 40, 41–49.
Gasteiger, E., Hoogland, C., Gattiker, A., Duvaud, S., Wilkins, M.R.,
Appel, R.D., Bairoch, A., 2005. Protein identification and
analysis tools on the ExPASy server. In: Walker, J.M. (Ed.), The
Proteomics Protocols Handbook. Humana Press, Totowe, NJ, USA,
pp. 571–607.
Gebruers, K., Debyser, W., Goesaert, H., Proost, P., Van Damme, J.,
Delcour, J.A., 2001. Triticum aestivum L. endoxylanase inhibitor
(TAXI) consists of two inhibitors, TAXI I and TAXI II, with different
specificities. Biochemical Journal 353, 239–244.
Gebruers, K., Goesaert, H., Brijs, K., Courtin, C.M., Delcour, J.A.,
2002a. Purification of TAXI-like endoxylanase inhibitors from wheat
(Triticum aestivum L.) whole meal reveals a family of iso-forms.
Journal of Enzyme Inhibition and Medicinal Chemistry 17, 61–68.
Gebruers, K., Brijs, K., Courtin, C.M., Goesaert, H., Proost, P., Van
Damme, J., Delcour, J.A., 2002b. Affinity chromatography with
immobilised endoxylanases separates TAXI- and XIP-type endoxyla-
nase inhibitors from wheat (Triticum aestivum L.). Journal of Cereal
Science 36, 367–375.
Gebruers, K., Brijs, K., Courtin, C.M., Fierens, K., Goesaert, H.,
Raedschelders, G., Robben, J., Sørensen, J.F., Van Campenhout, S.,
Delcour, J.A., 2004. Properties of TAXI-type endoxylanase inhibitors.
Biochimica et Biophysica Acta 1696, 213–221.
Gebruers, K., Courtin, C.M., Moers, K., Noots, I., Trogh, I., Delcour,
J.A., 2005. The bread-making functionalities of two Aspergillus niger
endoxylanases are strongly dictated by their inhibitor sensitivities.
Enzyme and Microbial Technology 36, 417–425.
Goesaert, H., Gebruers, K., Brijs, K., Courtin, C.M., Delcour, J.A.,
2003a. ‘TAXI’-type endoxylanase inhibitors in different cereals.
Journal of Agricultural and Food Chemistry 51, 3770–3775.
Goesaert, H., Gebruers, K., Brijs, K., Courtin, C.M., Delcour, J.A.,
2003b. XIP-type endoxylanase inhibitors in different cereals. Journal
of Cereal Science 38, 317–324.
Goesaert, H., Gebruers, K., Courtin, C.M., Delcour, J.A., 2005.
Purification and characterization of a XIP-type endoxylanase inhibitor
from rice (Oryza sativa). Journal of Enzyme Inhibition and Medicinal
Chemistry 20, 95–101.
Higgins, D., Thompson, J., Gibson, T., Thompson, J.D., Higgins, D.G.,
Gibson, T.J., 1994. CLUSTAL W: improving the sensitivity of
progressive multiple sequence alignment through sequence weighting,
position-specific gap penalties and weight matrix choice. Nucleic Acids
Research 22, 4673–4680.
Horton, P., Park, K.-J., Obayashi, T., Nakai, K., 2006. Protein subcellular
localization prediction with WoLF PSORT. In: Proceedings of the
Fourth Annual Asia Pacific Bioinformatics Conference APBC06,
Taipei, Taiwan, pp. 39–48.
Huisman, M.M.H., Schols, H.A., Voragen, A.G.J., 2000. Glucuronoar-
abinoxylans from maize kernel cell walls are more complex than those
from sorghum kernel cell walls. Carbohydrate Polymers 43, 269–279.
Julenius, K., Mølgaard, A., Gupta, R., Brunak, S., 2005. Prediction,
conservation analysis and structural characterization of mammalian
mucin-type O-glycosylation sites. Glycobiology 15, 153–164.
McLauchlan, W.R., Garcia-Conesa, M.T., Williamson, G., Roza, M.,
Ravestein, P., Maat, J., 1999. A novel class of protein from wheat
which inhibits xylanases. Biochemical Journal 338, 441–446.
Morrison, J.F., 1982. The slow-binding and slow, tight-binding inhibition
of enzyme-catalysed reactions. Trends in Biochemical Sciences 7,
102–105.
Nielsen, H., Krogh, A., 1998. Prediction of signal peptides and signal
anchors by a hidden Markov model. In: Glasgow, J., Littlejohn, T.,
Major, F., Lathrop, R., Sankoff, D., Sensen, C. (Eds.), Proceedings of
the Sixth International Conference on Intelligent Systems for
Molecular Biology (ISMB 6). AAAI Press, Menlo Park, CA, USA,
pp. 122–130.

ARTICLE IN PRESSK. Gebruers et al. / Journal of Cereal Science 48 (2008) 203–212212
Payan, F., Leone, P., Porciero, S., Furniss, C., Tahir, T., Williamson, G.,
Durand, A., Manzanares, P., Gilbert, H.J., Juge, N., Roussel, A.,
2004. The dual nature of the wheat xylanase protein inhibitor XIP-I:
structural basis for the inhibition of family 10 and family 11 xylanases.
Journal of Biological Chemistry 279, 36029–36037.
Raedschelders, G., Debefve, C., Goesaert, H., Delcour, J.A., Volckaert,
G., Van Campenhout, S., 2004. Molecular identification and
chromosomal localization of genes encoding Triticum aestivum
xylanase inhibitor I-like proteins in cereals. Theoretical and Applied
Genetics 109, 112–121.
Raedschelders, G., Fierens, K., Sansen, S., Rombouts, S., Gebruers, K.,
Robben, J., Rabijns, A., Courtin, C.M., Delcour, J.A., Van
Campenhout, S., Volckaert, G., 2005. Molecular identification of
wheat endoxylanase inhibitor TAXI-II and the determinants of its
inhibition specificity. Biochemical and Biophysical Research Commu-
nications 335, 512–522.
Reiss, E., Schlesier, B., Brandt, W., 2006. cDNA sequences, MALDI-TOF
analyses, and molecular modelling of barley PR-5 proteins. Phyto-
chemistry 67, 1856–1864.
Sansen, S., De Ranter, C.J., Gebruers, K., Brijs, K., Courtin, C.M.,
Delcour, J.A., Rabijns, A., 2004. Structural basis for inhibition of
Aspergillus niger xylanase by Triticum aestivum xylanase inhibitor-I.
Journal of Biological Chemistry 279, 36022–36028.
Sørensen, J.F., Kragh, K.M., Sibbesen, O., Delcour, J.A., Goesaert, H.,
Svensson, B., Tahir, T.A., Brufau, J., Perez-Vendrell, A.M., Bellin-
campi, D., D’Ovidio, R., Camardella, L., Giovane, A., Bonnin, E.,
Juge, N., 2004. Potential role of glycosidase inhibitors in industrial
biotechnological applications. Biochimica et Biophysica Acta 1696,
275–287.
Tahir, T.A., Berrin, J.-G., Flatman, R., Roussel, A., Roepstorff, P.,
Williamsson, G., Juge, N., 2002. Specific characterization of
substrate and inhibitor binding sites of a glycosyl hydrolase family
11 xylanase from Aspergillus niger. Journal of Biological Chemistry 46,
44035–44043.
Tahir, T.A., Durand, A., Gebruers, K., Roussel, A., Williamson, G., Juge,
N., 2004. Functional importance of Asp37 from a family 11 xylanase
in the binding of two proteinaceous xylanase inhibitors from wheat.
FEMS Microbiology Letters 239, 9–15.
Taylor, J.R.N., Schober, T.J., Bean, S.R., 2006. Novel food and non-food
uses for sorghum and millets. Journal of Cereal Science 44, 252–271.
Velazhahan, R., Jayaraj, J., Jeoung, J.M., Liang, G.H., Muthukrishnan,
S., 2002. Purification and characterization of an antifungal thaumatin-
like protein from sorghum (Sorghum bicolor) leaves. Journal of Plant
Disease and Protection 109, 452–461.
Verbruggen, M.A., Beldman, G., Voragen, A.G.J., Hollemans, M., 1993.
Water-unextractable cell wall material from sorghum: isolation and
characterization. Journal of Cereal Science 17, 71–82.
Verbruggen, M.A., Beldman, G., Voragen, A.G.J., 1995. The selective
extraction of glucuronoarabinoxylans from sorghum endosperm cell
walls using barium and potassium hydroxide solutions. Journal of
Cereal Science 21, 271–282.
Vigers, A.J., Roberts, W.K., Selitrennikoff, C.P., 1991. A new family of
plant antifungal proteins. Molecular Plant – Microbe Interactions 4,
315–323.




















