
Soil biota and upper soil layer development in two contrasting post-mining chronosequences
10
Ecological Engineering 17 (2001) 275 – 284 Soil biota and upper soil layer development in two contrasting post-mining chronosequences Jan Frouz a, *, Beate Keplin b , Va ´clav Piz ˇl a , Karel Tajovsky ´ a , Josef Stary ´ a , Alena Lukes ˇova ´ a , Alena Nova ´kova ´ a , Vladimı ´r Balı ´k a , Ladislav Ha ´ne ˇl a , Jan Materna a , Christian Du ¨ ker b , Josef Chalupsky ´ a , Josef Rusek a , Thomas Heinkele b a Institute of Soil Biology, Academy of Sciences of the Czech Republic, Na Sa ´dka ´ch 7, 37005 C & eske ´ Bude ˇjo6ice, Czech Republic b Brandenburg Uni6ersity of Technology at Cottbus, Chair of Soil Protection and Reculti6ation, PO Box 101344, 3013 Cottbus, Germany Accepted 19 August 2000 Abstract Density and community composition of a wide spectrum of soil organisms (fungi, algae, testate amoebae, nematodes, enchytraeids, lumbricids, oribatid mites, diplopods, terrestrial isopods, collembolans and dipteran larvae), direct counts of bacteria, rate of cellulose decomposition and microstructure of upper soil layers were studied in two chronosequences of plots reclaimed from open-cast coal mining near Cottbus (Germany) and near Sokolov (Czech Republic). German plots were characterized by acidic sandy soils and afforested with pine, while the Czech plots possessed alkaline clay soils and were afforested with alder. In both chronosequences, density and species richness in most of investigated groups of soil biota gradually increased with increasing succession age. Typical pioneer species were found in initial stands of both chronosequences. Nevertheless, the initial stands supported poorer assemblages of soil biota in the Cottbus area than did those in the Sokolov area. The Cottbus area was characterized by a moor type of humus and by gradual uniform increase of abundances and species numbers of most studied groups of soil biota. On the contrary, moder type of humus and a more variable pattern of soil biota development during the course of succession were found in the Sokolov area. Slower development of soil biota in initial stages of succession in Cottbus is caused by unfavourable physical and chemical conditions of heap substrates, and by lower input and poor quality of litter in pine plantations. © 2001 Elsevier Science B.V. All rights reserved. Keywords: Succession; Community; Ecosystem; Coal mining; Soil formation; Fauna; Microbiology; Zoology; Chemistry; Mi- crostructure www.elsevier.com/locate/ecoleng 1. Introduction For the reconstruction of ecosystems and eco- logical functions in a post-mining landscape, the * Corresponding author. Tel.: +420-38-7775769; Fax: + 420-38-5300133. E-mail address: [email protected] (J. Frouz). 0925-8574/01/$ - see front matter © 2001 Elsevier Science B.V. All rights reserved. PII:S0925-8574(00)00144-0
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Soil biota and upper soil layer development in two contrasting post-mining chronosequences

Ecological Engineering 17 (2001) 275–284
Soil biota and upper soil layer development in twocontrasting post-mining chronosequences
Jan Frouz a,*, Beate Keplin b, Vaclav Pizl a, Karel Tajovsky a, Josef Stary a,Alena Lukesova a, Alena Novakova a, Vladimır Balık a, Ladislav Hanel a,
Jan Materna a, Christian Duker b, Josef Chalupsky a, Josef Rusek a,Thomas Heinkele b
a Institute of Soil Biology, Academy of Sciences of the Czech Republic, Na Sadkach 7, 37005 C& eske Budejo6ice, Czech Republicb Brandenburg Uni6ersity of Technology at Cottbus, Chair of Soil Protection and Reculti6ation, PO Box 101344,
3013 Cottbus, Germany
Accepted 19 August 2000
Abstract
Density and community composition of a wide spectrum of soil organisms (fungi, algae, testate amoebae,nematodes, enchytraeids, lumbricids, oribatid mites, diplopods, terrestrial isopods, collembolans and dipteran larvae),direct counts of bacteria, rate of cellulose decomposition and microstructure of upper soil layers were studied in twochronosequences of plots reclaimed from open-cast coal mining near Cottbus (Germany) and near Sokolov (CzechRepublic). German plots were characterized by acidic sandy soils and afforested with pine, while the Czech plotspossessed alkaline clay soils and were afforested with alder. In both chronosequences, density and species richness inmost of investigated groups of soil biota gradually increased with increasing succession age. Typical pioneer specieswere found in initial stands of both chronosequences. Nevertheless, the initial stands supported poorer assemblagesof soil biota in the Cottbus area than did those in the Sokolov area. The Cottbus area was characterized by a moortype of humus and by gradual uniform increase of abundances and species numbers of most studied groups of soilbiota. On the contrary, moder type of humus and a more variable pattern of soil biota development during the courseof succession were found in the Sokolov area. Slower development of soil biota in initial stages of succession inCottbus is caused by unfavourable physical and chemical conditions of heap substrates, and by lower input and poorquality of litter in pine plantations. © 2001 Elsevier Science B.V. All rights reserved.
Keywords: Succession; Community; Ecosystem; Coal mining; Soil formation; Fauna; Microbiology; Zoology; Chemistry; Mi-crostructure
www.elsevier.com/locate/ecoleng
1. Introduction
For the reconstruction of ecosystems and eco-logical functions in a post-mining landscape, the
* Corresponding author. Tel.: +420-38-7775769; Fax: +420-38-5300133.
E-mail address: [email protected] (J. Frouz).
0925-8574/01/$ - see front matter © 2001 Elsevier Science B.V. All rights reserved.
PII: S 0925 -8574 (00 )00144 -0

J. Frouz et al. / Ecological Engineering 17 (2000) 275–284276
reconstruction of soils is necessary. The formationof upper soil layers, namely of organo-mineralhorizons, affected by vegetation cover as a sourceof organic matter and by soil biota, which trans-form organic matter coming to soil, is of particu-lar importance. Despite the principal role of soilbiota in these processes, more attention was tradi-tionally paid to vegetation development on thedumps. A complex approach is desirable in thestudy of soil biota because individual groups mayreflect successional changes in different ways(Tajovsky, 1990; Pizl, 1992; Scheu and Schulz,1996; Frouz, 1997a). Complex studies dealingwith a wide spectrum of soil organisms aremethodologically difficult and thus very scarce(Dunger, 1968, 1991; Topp et al., 1992). Variabil-ity of excavated substrates followed by a highvariability of environmental conditions as well asdifferent reclamation technologies result inmarked heterogeneity in the development of soilbiota (Dunger, 1991; Topp et al., 1992; Schneideret al., 1995). More empirical studies of theseprocesses in variable conditions are therefore ur-gently needed to better understand the factorsaffecting soil biota development in the post-min-ing landscape.
The aim of this study was to describe thesuccession of soil biota in two contrastingchronosequences of reclaimed post-mining : areaswith very adverse environmental conditions, af-forested with conifers, producing hardly decom-posable litter; and, on the other hand, areas withbetter conditions, afforested with deciduous trees,producing easily decomposable litter. Thechronosequences may serve as examples of soilbiota development during the forest reclamationof a post-mining landscape.
2. Material and methods
2.1. The sites
The study was carried out at two chronose-quences of plots reclaimed from open-cast coalmining in two areas, the Lusatian lignite miningdistrict near Cottbus (Germany) and the North-west Bohemian brown coal mining district near
Sokolov (Czech Republic). The heaps in the Cot-tbus area were formed by lignite rich tertiarysand. Pioneer stages had a high salinity and highcontent of fossil organic matter and low pH(Duker et al., 1997) ameliorated with fly ash froma power station (Fig. 1a). The plots were af-forested with Pinus sil6estris in 1964, 1978, 1982,1996 and Pinus nigra in 1982. The annual input ofpine litter was 0, 142, 336 and 563 g m−2 inplantation planted in 1696, 1982, 1978 and 1964,respectively (Pietsch, 1996). The dumps nearSokolov were formed by tertiary clay material.Initial stages had alkaline pH. In this area, plotsafforested with Alnus glutinosa in 1935, 1961,1974, 1978, 1989 and 1995 were selected. Theoldest site was located on a dump from sub-sur-
Fig. 1. Chemical parameters and soil microflora in individualplots of investigated chronosequences near Cottbus (trianglesand thin lines) and near Sokolov (circles and bold lines). Barsrepresent standard errors, plots are ordered according to age,and points unconnected by line represent control plots. Allweights are given on dry-weight basis. (a) pH in H2O; (b)organic carbon content; (c) cellulose decomposition rate; (d)direct counts of bacteria; (e), (f) fungi (CFU, colony formingunits); (g), (h) soil algae.

J. Frouz et al. / Ecological Engineering 17 (2000) 275–284 277
face mining and consisted of tertiary and quater-nary clay material. The amount of litter in litterlayer of Sokolov plots (lined in acceding age) were0, 1376, 512, 560, 512, and 1200 g m−2. Theproportion of woody fragments increased in thelitter of the oldest plot. In both areas, controlplots on natural soil were investigated. They rep-resent forests of similar age as the oldest re-claimed site: a 35-year-old stand of P. sil6estris inthe Cottbus area and a 70-year-old stand of A.glutinosa in the Sokolov area.
2.2. Sampling and analyses
Samplings were made four times at each plot(30 May–1 June 1997, 29–30 September 1997,27–29 April 1998 and 29–30 October 1998 atCottbus; and 14–15 May 1997, 16–17 September1997, 12–13 May 1998 and 13–14 September1998). Exceptions were the study of bacteria, car-ried out in 1998 only, and the study of cellulosedecomposition.
Decomposition of cellulose was studied usingthe litterbag method (Dunger and Fiedler, 1997).The litterbags (10 cm×10 cm; mesh size, 1.5 mm)were buried in the humus layer or, if the humuslayer was not developed, at 5 cm depth. At Cot-tbus, at each study site, a set of 40 litterbags filledwith 5.0 g oven-dried strips of filter paper wasused in 1995 and 1996. Eight litterbags from eachstudy site were sampled at 12-week intervals. Inthe laboratory, the remaining mass of cellulosewas determined after incineration (Malkomes,1980). Daily decomposition rate was calculated asmilligrams per day applying the single exponentialdecay function (Wieder and Lang, 1982). AtSokolov, four litter bags, each with 1 g filterpaper, were exposed for 6 weeks three times dur-ing May–June in 1996–1998. Decomposition ratewas calculated as loss of cellulose per days ofexposition. Mixed samples from the depth of 0–5cm were used for the determination of pH (H2O,1:5) and organic carbon content (by wetcombustion).
For the study of microflora, mixed samplesconsisting of at least five particular samples wereused. Sampling depth for bacteria and fungi was 5cm. Bacteria were estimated by direct counts us-
ing the DAPI method (Bloem, 1995). Fungi werequantified using dilution plate method on soilextract agar. Various media were used for theidentification of fungi (Garrett, 1991). Samplingdepth for algae was 1 cm. Algae were directlycounted by epifluorescence microscopy, and iden-tified by microscopy and cultivation (Lukesova,1993).
For the study of microfauna, five samples (area,10 cm2; depth, 5 cm) were taken. Nematodes wereextracted with Baermann funnels. Testate amoe-bae were studied using direct microscopy of sus-pended samples (Foissner, 1993). Oribatid mitesand collembolans were sampled by the same wayas microfauna; high gradient extractor was usedfor extraction of animals. For the study of Enchy-traeids, five samples were taken in each plot (10cm2 area and 10 cm depth in the Sokolov area;and 20 cm2 area and 12 cm depth in the Cottbus)and animals were extracted by wet funnel. Soilmacrofauna was extracted (by Tullgren type ex-tractor) from three soil samples (625 cm2×10 cm)from Sokolov and five soil samples (125 cm2×10cm) from the Cottbus areas, respectively.
The collected material, except for bacteria, wasidentified to species level or distinguishable ‘mor-phospecies’. Average values of abundance andnumbers of species recorded during the observa-tion period are presented; standard error coversvariability among individual sampling terms.
In addition, one undisturbed soil sample (5×5cm2; depth, 10 cm) was taken from each reclaimedplot to study soil microstructure. The sampleswere dried and thin soil sections were preparedaccording to Rusek (1978). The thickness of litter,humus and fermentation layers was measured onfour replicate blocks used for preparation of soilthin sections.
3. Results
3.1. Chemical and microbiological parameters
The differences in pH of the Cottbus plots aredue to different intensity of amelioration (Fig.1a). In the Sokolov area, the pH gradually de-creased with the age of the plots (Fig. 1a). Con-

J. Frouz et al. / Ecological Engineering 17 (2000) 275–284278
tent of organic carbon increased gradually withthe age of the plots in the Cottbus chronose-quence; a lower value in control forest in com-parison with the oldest reclaimed site is caused bythe surplus of fossil organic matter in reclaimedplots. The content of organic carbon in theSokolov area increased steeply from pioneer plotsto 20- 30-year-old plots than in older plots (Fig.1b). The cellulose decomposition rate and directcounts of bacteria gradually increased fromyounger plots to the older ones in the Cottbusarea. In the Sokolov area, the values were higherand increased steeply during the first 20 to 30years of succession. Counts of bacteria slowlyincreased in older plots, whereas cellulose decom-position slightly decreased (Fig. 1c,d). Counts ofsoil micro-fungi were low in pioneer plots in bothchronosequences and higher on older plots. Thecounts were higher in the control pine forest thanthose in the alder control (Fig. 1e). The number offungal species increased with the age of plots inthe Cottbus chronosequence, whereas high num-bers of species (20–35) were found in all plots inthe Sokolov area. (Fig. 1f). The proportion ofMucoraceae, utilizing easily decomposable sub-strates, increased with succession age in reclaimedplots near Cottbus, whereas representatives of thisfamily were frequent along the whole chronose-quence in the Sokolov area. The abundance ofsoil algae in the Cottbus chronosequence followedan increasing trend with the age of plots. In theSokolov area, however, the highest abundance ofalgae was found in the youngest plot not fullycovered with trees and it was substantially lowerin older plots shaded by trees (Fig. 1g). Thespecies number displayed a similar pattern andfollowed a more or less increasing trend along theage gradient at Cottbus and a decreasing trend atSokolov (Fig. 1h). In both areas, green algae(Chlorophyceae) dominated. Nevertheless,cyanobacteria (Cyanophyceae) and diatoms(Bacillariophyceae) were quite important in alka-line pioneer stages at Sokolov.
3.2. Soil zoological parameters
The abundances of all investigated groups ofsoil invertebrates increased gradually from
Fig. 2. Soil microfauna and mesofauna in individual plots ofinvestigated chronosequences near Cottbus (triangles and thinlines) and near Sokolov (circles and bold lines). (a), (b) Tes-tacea; (c), (d) Nematoda; (e), (f) Collembola; (g), (h) Orib-atida. For further explanation, see Fig. 1.
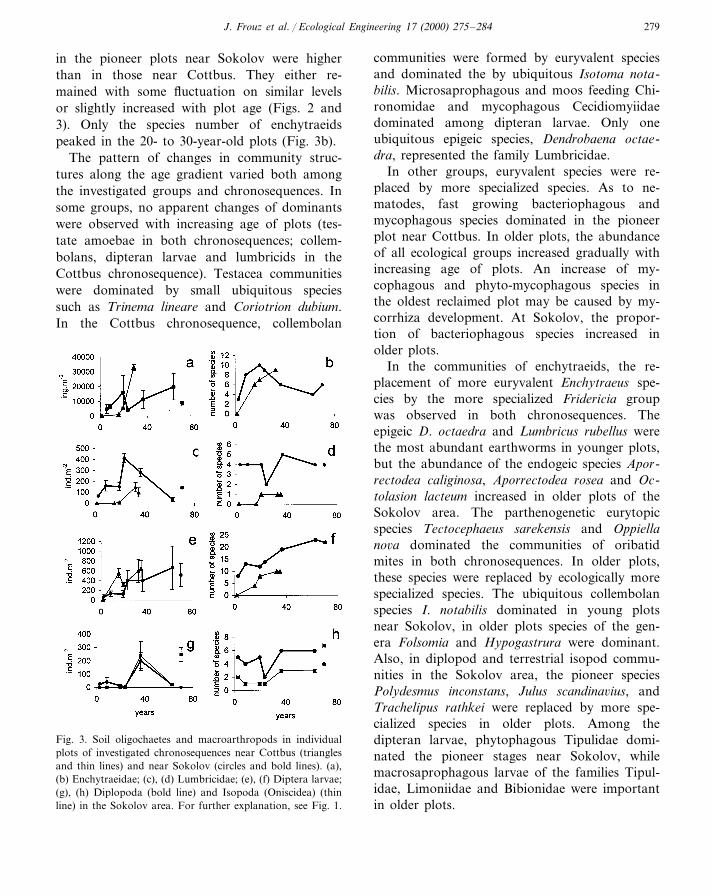
younger plots to the older ones in the Cottbuschronosequence, except for nematodes. The abun-dance of nematodes was relatively high in youngplots, and maintained on a similar level in olderones. A more variable pattern of abundancechanges was observed in the Sokolov area, where,except for nematodes, the abundances of investi-gated groups increased in young plots and peakedin 20 to 30-year-old plots. After this peak, theabundance either decreased (oribatid mites, lumb-ricids, diplopods, terrestrial isopods) or increasedagain, but this increase was either slower(dipteran larvae) or followed a depression(collembolans, testate amoebae, enchytraeids)(Figs. 2 and 3).
The species numbers of all investigated groupsof soil invertebrates were low in the pioneer plotof the Cottbus chronosequence and gradually in-creased with the age of plots. The species numbers
J. Frouz et al. / Ecological Engineering 17 (2000) 275–284 279
in the pioneer plots near Sokolov were higherthan in those near Cottbus. They either re-mained with some fluctuation on similar levelsor slightly increased with plot age (Figs. 2 and3). Only the species number of enchytraeidspeaked in the 20- to 30-year-old plots (Fig. 3b).
The pattern of changes in community struc-tures along the age gradient varied both amongthe investigated groups and chronosequences. Insome groups, no apparent changes of dominantswere observed with increasing age of plots (tes-tate amoebae in both chronosequences; collem-bolans, dipteran larvae and lumbricids in theCottbus chronosequence). Testacea communitieswere dominated by small ubiquitous speciessuch as Trinema lineare and Coriotrion dubium.In the Cottbus chronosequence, collembolan
communities were formed by euryvalent speciesand dominated the by ubiquitous Isotoma nota-bilis. Microsaprophagous and moos feeding Chi-ronomidae and mycophagous Cecidiomyiidaedominated among dipteran larvae. Only oneubiquitous epigeic species, Dendrobaena octae-dra, represented the family Lumbricidae.
In other groups, euryvalent species were re-placed by more specialized species. As to ne-matodes, fast growing bacteriophagous andmycophagous species dominated in the pioneerplot near Cottbus. In older plots, the abundanceof all ecological groups increased gradually withincreasing age of plots. An increase of my-cophagous and phyto-mycophagous species inthe oldest reclaimed plot may be caused by my-corrhiza development. At Sokolov, the propor-tion of bacteriophagous species increased inolder plots.
In the communities of enchytraeids, the re-placement of more euryvalent Enchytraeus spe-cies by the more specialized Fridericia groupwas observed in both chronosequences. Theepigeic D. octaedra and Lumbricus rubellus werethe most abundant earthworms in younger plots,but the abundance of the endogeic species Apor-rectodea caliginosa, Aporrectodea rosea and Oc-tolasion lacteum increased in older plots of theSokolov area. The parthenogenetic eurytopicspecies Tectocephaeus sarekensis and Oppiellano6a dominated the communities of oribatidmites in both chronosequences. In older plots,these species were replaced by ecologically morespecialized species. The ubiquitous collembolanspecies I. notabilis dominated in young plotsnear Sokolov, in older plots species of the gen-era Folsomia and Hypogastrura were dominant.Also, in diplopod and terrestrial isopod commu-nities in the Sokolov area, the pioneer speciesPolydesmus inconstans, Julus scandina6ius, andTrachelipus rathkei were replaced by more spe-cialized species in older plots. Among thedipteran larvae, phytophagous Tipulidae domi-nated the pioneer stages near Sokolov, whilemacrosaprophagous larvae of the families Tipul-idae, Limoniidae and Bibionidae were importantin older plots.
Fig. 3. Soil oligochaetes and macroarthropods in individualplots of investigated chronosequences near Cottbus (trianglesand thin lines) and near Sokolov (circles and bold lines). (a),(b) Enchytraeidae; (c), (d) Lumbricidae; (e), (f) Diptera larvae;(g), (h) Diplopoda (bold line) and Isopoda (Oniscidea) (thinline) in the Sokolov area. For further explanation, see Fig. 1.

J. Frouz et al. / Ecological Engineering 17 (2000) 275–284280
Fig. 4. Micromorphology of upper soil layers. Soil thin sections from 20-year-old plots from the Cottbus and Sokolov areas.Cottbus: (a) general view, (b) pine needles partly consumed by dipteran larvae (white arrow) and enchytraeids (black arrow), (c)compressed layer of partly decomposed needles and excrements; Sokolov: (d) general view, (e) partly broken excrement ofmacrosaprophagous animals above the layer of broken and compressed excrements, (f) excrement of macrosaprophagous animalsinside earthworm cast. Scale bars represent 2 mm in (a) and (b), and 0.2 mm in others; broken lines represent borders betweenmineral and organic horizons.

J. Frouz et al. / Ecological Engineering 17 (2000) 275–284 281
3.3. Microstructure of upper soil layers
Fermentation and humus layers were hardlydistinguishable from each other in the Cottbusarea (Fig. 4a). They form one layer developinggradually with the age of plot. On the contrary, atSokolov, the thickness of the fermentation layerincreased in the first 20–30 years, but it decreasedagain later. The thickness of the humus layerincreased gradually (Fig. 5). The most importantdecomposers of pine needless in Cottbus plotswere fungi, enchytraeids, dipteran larvae andcollembolans (Fig. 4b). Partly decomposed litterwas concentrated on the interface between organicand mineral horizons and after further com-paction and decomposition, it formed fermenta-tion and humus layers (Fig. 4a,c). In Sokolov,litter was intensively broken down by macros-aprophagous invertebrates, mostly dipteran lar-vae, diplopods and terrestrial isopods. Partlydecayed and compressed faecal pellets were con-centrated on the interface between organic andmineral layers. Faecal pellets in various stages ofdecay and rests of decomposed leaves were trans-ported in the soil profile by earthworms (Fig. 4f).Due to earthworm activity, the borders betweenindividual layers are very irregular (Fig. 4d). In
the oldest reclaimed plot, the transport of faecalpellets by earthworms was less pronounced, butthe mechanical movement was enhanced by sub-strates, which contained more macropores.
4. Discussion
Density and species richness consequently in-creased during succession in most investigatedgroups of soil biota in both choronosequences.The differences were found in patterns of thedevelopment of soil biota. The comparison of soilbiota communities and humus layer profiles indi-cates that a hypothetical final equilibrium stage inboth chronosequences is different. Moor type ofhumus, common in acidic soils with hardly de-composable litter (Dickinson and Pugh, 1977;Davis et al., 1992; Fons and Klinka, 1998) wasfound in pine stands near Cottbus. On the con-trary, the formation of moder type of humus,which shifted to mull in older plots, was observedin the Sokolov area, and the soil biota found heredisplays features typical for this humus type,namely high abundance of litter breaking macro-fauna and higher counts of bacteria.
Both abundances and species numbers of soilorganisms were lower in pioneer stands near Cot-tbus than in those near Sokolov. The low abun-dances seem to be connected with adversephysico-chemical conditions of substrate, mainlyhigh acidity, salinity and low ability of sandysubstrate to retain water. Laboratory experimentswith the survival of Enchytraeus crypticus indi-cated that this substrate may be toxic for soilanimals (Frouz, unpublished work). Adverse con-ditions of sandy substrates may take part in aslower development of soil biota near Cottbusthan near Sokolov. A lower input of litter inconiferous forest may also reduce the develop-ment of soil biota in the Cottbus plots. Thedevelopment of most groups of soil biota in pinestands near Cottbus showed a uniform pattern.Both abundances and species numbers increasedgradually with the age of plots. This pattern ofsuccession corresponds with the data from conif-erous reclamation near Gorlitz (Dunger, 1991).The development of soil biota in the Cottbus area
Fig. 5. Thickness of litter, fermentation and humus layers inreclaimed plots of various ages in the Cottbus (a) and Sokolov(b) areas.

J. Frouz et al. / Ecological Engineering 17 (2000) 275–284282
is connected with gradually growing fermentationlayer. The abundance and species number ofgroups dwelling mainly in fermentation layer suchas testate amoebae (Laminger, 1980; Laminger etal., 1982) increased with time faster in the Cottbuschronosequence, than in the Sokolov one. Litteraccumulation and growth of fermentation layerincrease nutrient supply and improve physico-chemical conditions of adverse sandy substratesand thus act as key ecological factors affectingsoil organisms.
On the contrary, the pattern of development ofindividual groups of soil biota in alder reclama-tion near Sokolov is more complex. Dunger(1991) observed similar differences in the patternof soil biota succession between reclaimed areasafforested with coniferous and deciduous trees.The community of pioneer stages in the Sokolovarea is more diverse and abundant in comparisonwith Cottbus. Dominant pioneer species atSokolov were in many cases similar to thosefound at Cottbus, they were typical of data strate-gists also recorded from other disturbed areas(Fujikawa, 1988). Nevertheless, more suitable en-vironmental conditions and a rapid developmentof herbaceous vegetation in pioneer plots atSokolov supported the species that were absent inpioneer plot in the Cottbus area, namely speciesfixed on litter or living tissue of herbaceous vege-tation (some phytophagous nematodes anddipteran larvae). The development of similar spe-cies communities fixed on herbaceous vegetationwas recorded also from other coal mining areaswhere the dump environment is not so extreme(Dunger, 1968, 1991; Topp et al., 1992). In youngalder plantations, the communities of soil organ-isms develop rapidly and some groups reach, in20–30-year-old plots, abundances comparablewith similar forests on natural soil undisturbed bymining (Tajovsky, 1990; Pizl, 1992; Frouz, 1997a).A rapid development of the soil biota communitywas observed also in plantations of deciduoustrees near Gorlitz (Dunger, 1968, 1991). It seemsto be connected with a better quality of leaf litter(Dickinson and Pugh, 1977) and a suitable sub-strate condition. Nevertheless, in terms of domi-nant species, these communities differed in manyinvestigated groups from stabilized forest commu-
nities. In older plantations, the abundance ofmost of the investigated groups grows slower oreven decreases. Similarly, Dunger (1968, 1991)also observed a decrease in number of somegroups in older deciduous plantations. These de-pressions may have several reasons. As usual indeciduous forests, the litter production per soilsurface area increases in young plantations withthe growth, and this increase is higher in youngplantations. In older plantations, when a highcover is reached, the litter production stagnatedor even decreased (Landsberg, 1986; Bartelink,1997). Abundant decomposer community, inolder plantations, consume a substantial part ofleaf litter, and thus it can limit their own develop-ment (Hovemeyer, 1992; Ponge et al., 1993).Moreover, increasing shading during the growthof alder plantation may reduce the cover of herblayer and thus enhance the effect already de-scribed. In addition, changes in upper soil layersrealized by soil biota can bring some feedback.Macrosaprophagous animals such as litter feedingdipteran larvae, diplopods and terrestrial isopodstransform a substantial part of litter fall intofaecal pellets that accumulate in the fermentationlayer (Szabo, 1974; Tajovsky, 1992). Fast develop-ment of the fermentation layer and accumulationof organic carbon in the upper soil layer wasobserved at Sokolov during the first 20–30 yearsof succession (Fig. 1b and Fig. 4b). Both of theseparameters stagnated in older plots. Several rea-sons may be responsible for this process; one ofthem can be the activity of Lumbricidae, whichbind organic matter in minerals similarly to thatdescribed by Bernier and Ponge (1994) in alpineforests. Dunger (1991) described a similar shift inthe structure of humus layer. This shift in thedevelopment of fermentation and humus layerscan be affected by soil dwelling organisms. An-other factor that may effect the development ofsoil biota in older plots is a gradual decrease ofpH, which is slightly alkaline in initial stages ofsuccession and, with accumulation of organicmatter, shifts to neutral and acidic. It can bespeculated that neutral or slightly acidic pH,which is reached in 20–30-year-old plots, is morefavourable for most soil biota than a more acidicpH in the older ones (Nordstrom and Rundgren,1974).

J. Frouz et al. / Ecological Engineering 17 (2000) 275–284 283
In many groups, not only abundances but alsospecies numbers increased slower in older plotsthan in the younger ones. The competition ofresident species with new invaders could be re-sponsible for this, as well as the reduction ofinvader flow. This may be caused by the fact thatsoil invertebrates specialized in old forests arefrequently very poor migrants even in groupswhich normally migrate fast (Neumann, 1971;Moldenke and Latin, 1990). Because of the largedistance from natural forests, migration of thesespecies can be very slow. The wind transport isimportant in colonization of post-mining dumps(Wanner et al., 1998); particularly for micro- andmesofauna, it can be expected that increasing treecover may reduce the number of wind transportedinvaders in older plantations. This factor is en-hanced by the fact that larger areas are frequentlyplanted at the same time. In addition, the intro-duction of new species as a by-product of humanactivity is more likely in younger plots, namelyduring the planting and care for young trees.Finally, some species can be lost during the courseof succession because pioneer stages are fre-quently more heterogeneous than the older stagesof succession (Brown, 1991) and because somespecies specialized in early stages of successionmay disappear later on (Frouz, 1997b, 1998).
In comparison with the Cottbus area, the devel-opment of soil biota in the Sokolov area is morevariable and the development of individual groupsvaries along the aforementioned main pattern.This seems to be caused by the fact that theslightly alkaline clay substrate is more suitablethan the sandy acidic one and may potentiallyhost a more diverse community of soil biota(Nordstrom and Rundgren, 1974). Due to bettersubstrate conditions, singularities of particularplots can also be more pronounced. This variabil-ity may be enhanced by the fact that the decom-poser community of deciduous forests is verysensitive to fluctuation in litter input (Hovemeyer,1992; Ponge et al., 1993).
In conclusion, gradual development of soilbiota was observed in both chronosequences. Pat-terns of soil biota differ, in detail, in dependenceon both substrate qualities as well as tree speciesused for afforestation. The former seems to be
more important in initial stages of succession. Theresponse differs also among individual group ofsoil biota, which underline the importance of amulti-approach comparison for better under-standing of the succession process in post-miningareas.
Acknowledgements
The authors would like to thank the DeutscheForschungsgemeinschaft, Internationales Burodes BMBF, the KONTAKT program (grant no.ME 076), the Grant Agency of the Czech Repub-lic (grant no. 526/98/P156), the ENKI v.p.s., andthe Sokolovska uhelna a.s for financial support.The authors thank three anonymous referees fortheir helpful comments.
References
Bartelink, H.H., 1997. Allometric relationships for biomassand leaf area of beech (Fagus syl6atica L.). Amm. Sci. For.54, 39–50.
Bernier, N., Ponge, J.F., 1994. Humus form dynamics duringthe sylvogenetic cycle in mountain spruce forest. Soil Biol.Biochem. 26, 183–220.
Bloem, J., 1995. Fluorescent staining of microbes for totaldirect counts. In: Akkermans, A.D.L., Van Elsas, D.J., DeBruijn, F.J. (Eds.), Molecular Microbial Ecology Manual,vol. 4.1.8. Kluwer Academic, Dordrecht, pp. 1–12.
Brown, K.H., 1991. The effect of changes in habitat structureduring succession in terrestrial communities. In: Bell, S.S.,McCoy, E.D., Mushinsky, H.R. (Eds.), Habitat Structure:The Physical Arrangement of Object in Space. Chapmanand Hall, London, pp. 141–168.
Davis, B.N.K., Walker, N., Ball, D.F., Fitter, A.H., 1992. TheSoil. Harper Collins, London, 192 pp.
Dickinson, C.H., Pugh, G.J., 1977. Biology of Plant LitterDecomposition. Academic Press, London, p. 775.
Duker, C., Rumpel, C., Keplin, B., Kogel-Knabner, I., Huttl,R.F., 1997. Enchytraeidenabundanz, artenspektrum undvertikalverteilung in unterschiedlich alten forstlich rekul-tivierten Kippkohlesanden. Mitt. Dtsch. Bodenkund.Gesellsch. 85, 481–484.
Dunger, W., 1968. Die entwicklung der bodenfauna auf rekul-tivierten kippen und halden des braunkohlentagebaues.Abh. Ber. Naturkundemus. Gorlitz 43, 1–256.
Dunger, W., 1991. Zur primarsukzession humiphager tiergrup-pen auf bergbauflachen. Zool. Jahrb. Syst. 118, 423–447.
Dunger, W., Fiedler, H.J., 1997. Methoden der Bodenbiologie.Gustav Fischer, Jena, 537 pp.

J. Frouz et al. / Ecological Engineering 17 (2000) 275–284284
Foissner, W., 1993. Microfauna. In: Schinner, F., O8 hlinger,R., Kandeler, R., Margesin, R. (Eds.), BodenbiologischeArbeitsmethoden, 2nd ed. Springer, Berlin, pp. 289–311.
Fons, J., Klinka, K., 1998. Chemical and biotic properties andtemporal variation of humus forms in the rain forest nearVancouver, British Columbia. Geoderma 86, 83–98.
Frouz, J., 1997a. Changes in communities of soil dwellingdipteran larvae during secondary succession in abandonedfields. Eur. J. Soil Biol. 33, 57–65.
Frouz, J., 1997b. The effect of vegetation patterns on oviposi-tion habitat preference: a driving mechanisms in terrestrialChironomid (Diptera: Chironomidae) succession? Res.Popul. Ecol. 39, 207–213.
Frouz, J., 1998. Changes of craneflies (Tipulidae) larvae popu-lation during secondary succession. In: Pizl, V., Tajovsky,K. (Eds.), Soil Zoological Problems in Central Europe.ISB, Ceske Budejovice, pp. 31–36.
Fujikawa, T., 1988. Fluctuation of oribatid mites in Nayoronature farming field during ten years. Edaphologia 39,29–37.
Garrett, S.D., 1991. Soil Fungi and Soil Fertility. PergamonPress, Oxford, 150 pp.
Hovemeyer, K., 1992. Response of Diptera populations toexperimentally modified leaf litter input in beech forest onlimestone. Pedobiologia 36, 35–49.
Laminger, H., Kainz, G., Horvath, G., 1982. Populationsdy-namik terrestrischer Protozoen (Testacea, Rhizopoda) inzentralalpinen Lagen Tirols. IV Distribution der Tes-taceenzonosen in Hochgebirgsboden. Zool. Anz. 208, 339–351.
Laminger, H., 1980. Bodenprotozoologie. Mikrobios 1, 1–142.Landsberg, J.J., 1986. Physiological Ecology of Forest Produc-
tion. Academic Press, Harcourt Brace Jovanovich, Lon-don, 193 pp.
Lukesova, A., 1993. Soil algae in four secondary successionalstages on abandoned fields. Algol. Stud. 71, 81–102.
Malkomes, H.P., 1980. Strohrotteversuche zur erfassung vonHerbizid-Nebenwirkungen auf den strohumsatz im Boden.Pedobiologia 20, 417–427.
Moldenke, A.P., Latin, D.J., 1990. Dispersal characteristics ofold-growth arthropods: the potential for loss of diversityand biological function. Northwest Environ. J. 6, 408–409.
Neumann, U., 1971. Die sukzession der bodenfauna (Cara-bidae [Coleoptera], Diplopoda und Isopoda) in denforstlich rekultivierten Gebieten des RheinischenBraunkohlenreviers. Pedobiologia 11, 193–226.
Nordstrom, S., Rundgren, S., 1974. Environmental factors andlumbricid associations in southern Sweden. Pedobiologia14, 1–27.
Pietsch, W., 1996. Pflanzliche Sukzession auf Kippenboden–Entwicklung und Wechselwirkungen–wechserlwirkungenzwischen Pflazenbewuchs und Bodeneigenschaften auf Kip-pen-standorten. In: O8 kologisches Entwicklungspotentialder Bergbaufolgelanschaften im Lausitzer Braunkohlere-vier. BTU Centre of Excelence, Cottbus 98–103.
Pizl, V., 1992. Succession of earthworm populations in aban-doned fields. Soil. Biol. Biochem. 24, 1623–1628.
Ponge, J.F., Aprin, P., Vanier, G., 1993. Colembola responseto experimental perturbations of litter supply in a temper-ate forest ecosystem. Eur. J. Soil. Biol. 29, 141–153.
Rusek, J., 1978. Pedozootische sukzession wahrend der en-twicklung von okosystemen. Pedobiologia 18, 426–433.
Scheu, S., Schulz, E., 1996. Secondary succession, soil forma-tion and development of a diverse community of oribatidsand saprophagous soil macro-invertebrates. BiodiversityConserv. 5, 235–250.
Schneider, R., Schneider, H., Schroder, D., 1995. Beziehungzwischen bodenorganismen und gefugeentwicklung aufLoss-Neuland unter acker- und waldnutzung. Z. Pflanzen-ernahr. Bodenk. 158, 197–204.
Szabo, I.M., 1974. Microbial Communities in Forest RendzinaEcosystem. Akademia Kiado, Budapest, 415 pp.
Tajovsky, K., 1990. Diplopoda in secondary soil successionalrow. In: Minelli, A. (Ed.), Proceedings of the 7th Interna-tional Congress of Myriapodology. Brill, Leiden, pp. 229–234.
Tajovsky, K., 1992. Feeding biology of the millipede Glomerishexasticha. Ber. Nat.-Med. Verein Innsbruck Suppl. 10,305–311.
Topp, W., Gemesi, O., Gruning, C., Tach, P., Zhou, H., 1992.Forstlische rekultivierung mit altwaldboden in RheinischenBraunkohlenrevier. Die sukzession der bodenfauna. Zool.Jahrb. Syst. 119, 505–533.
Wanner, M., Dunger, W., Schulz, H.J., Voigtlander, K., 1998.Primary immigration of soil organisms on coal mined areasin Eastern Germany. In: Pizl, V., Tajovsky, K. (Eds.), SoilZoological Problems in Central Europe. ISB, Ceske Bude-jovice, pp. 267–275.
Wieder, R.K., Lang, G.E., 1982. A critique of the analyticalmethod used in examining decomposition data obtainedfrom litter bags. Ecology 63, 1636–1642.
.




















