
Soil and litter nematode diversity of Mount Hamiguitan, the Philippines, with description of...
20
Nematology 17 (2015) 325-344 brill.com/nemy Soil and litter nematode diversity of Mount Hamiguitan, the Philippines, with description of Bicirronema hamiguitanense n. sp. (Rhabditida: Bicirronematidae) Xue QING 1 , Wim BERT 1 , Hanne S TEEL 1 , Joeseph QUISADO 2 and Irma TANDINGAN DE L EY 3,∗ 1 Nematology Unit, Department of Biology, Ghent University, K.L. Ledeganckstraat 35, 9000 Ghent, Belgium 2 Department of Entomology, University of Southern Mindanao, the Philippines 3 Department of Nematology, University of California, Riverside, CA, USA Received: 19 October 2014; revised: 31 December 2014 Accepted for publication: 1 January 2015; available online: 18 February 2015 Summary – The nematode diversity in soil and litter was investigated on Mount Hamiguitan, the Philippines, along four eco-habitats from elevations of 75-1600 m a.s.l. A total of 155 and 467 nematodes were identified to 39 and 62 genera from litter and soil, respectively. The nematode assemblages and diversity did not show any relation to eco-habitat or elevation. Bacterivorous nematodes were the most common group (37.5%). Acrobeloides was most abundant from the soil and Aphelenchoides from the litter. Bicirronema hamiguitanense n. sp. is herein described based on morphology, morphometrics and molecular data. The new species has the following diagnostic features: a wide lateral field one-fifth of its body diam. with four incisures forming two ornamented ridges; gubernaculum with proximal thickening; spicules (35-38 μm) and gubernaculum (18-20 μm) longer than B. caledoniense; and with 37 molecular autapomorphies supporting its new species status. The phylogenetic position of the new species within Cephalobomorpha is discussed. Keywords – molecular, morphology, morphometrics, new species, phylogeny, taxonomy. Nematode diversity studies have historically focused on temperate regions and only ten of 134 published ecosystem surveys represented tropical forests (Boag & Yeates, 1998). Also, these studies only considered soil samples, with very little or no attention given to the epiphytic communities (litter or canopy). However, recent studies (Powers et al., 2009; Porazinska et al., 2010, 2012) have taken into account both soil and litter material, with litter showing higher nematode diversity than soil. Nematode diversity was based only on either Molecular Operational Taxonomic Units (MOTUs) or Operational Clustered Taxonomic Units (OCTUs) for parallel tagged ultra sequencing. It is commonly thought that diversity tends to increase as altitude increases (Stevens, 1992; Begon et al., 1996; Rahbek, 1997) and this has been found in vascular plants and birds (Hunter & Yonzon, 1993; Wolf, 1994; Pendry & Proctor, 1997; Rahbek, 1997; Shepherd, 1998). There are very few studies on nematode altitudinal distribution and these were conducted in grassland, temperate or subtropical forest, or desert environments, often with ∗ Corresponding author, e-mail: [email protected] contradictory results. For instance, nematode diversity was found to decline as altitude increases (Weaver & Smolik, 1987; Chen et al., 2009; Hánˇ el & ˇ Cerevková, 2010); however, in other studies, diversity was not related to elevation (Yeates, 1974; Steinberger et al., 2001; Treonis et al., 2012). The Mount Hamiguitan Range Wildlife Sanctuary is lo- cated in the southeast of Mindanao Island, the Philippines. It has been identified by Conservation International as one of the Philippines’ biodiversity hotspots (see Myers et al., 2000) because of its high degree of species endemism in flora and fauna, and the threat to this from human inter- ference (Amoroso & Aspiras, 2011). Recently, this sanc- tuary was included on the UNESCO World Heritage List (http://whc.unesco.org/en/list/1403). Its tropical climate, wide range of altitude from 75-1637 m a.s.l. and varied eco-habitats make it a good candidate for this altitudinal nematode community study. We hypothesised that there exists an elevational pattern of nematode abundance, di- versity and community structure in Mount Hamiguitan. © Koninklijke Brill NV, Leiden, 2015 DOI 10.1163/15685411-00002870
Transcript of Soil and litter nematode diversity of Mount Hamiguitan, the Philippines, with description of...

Nematology 17 (2015) 325-344 brill.com/nemy
Soil and litter nematode diversity of Mount Hamiguitan, thePhilippines, with description of Bicirronema hamiguitanense
n. sp. (Rhabditida: Bicirronematidae)
Xue QING 1, Wim BERT 1, Hanne STEEL 1, Joeseph QUISADO 2 and Irma TANDINGAN DE LEY 3,∗1 Nematology Unit, Department of Biology, Ghent University, K.L. Ledeganckstraat 35, 9000 Ghent, Belgium
2 Department of Entomology, University of Southern Mindanao, the Philippines3 Department of Nematology, University of California, Riverside, CA, USA
Received: 19 October 2014; revised: 31 December 2014Accepted for publication: 1 January 2015; available online: 18 February 2015
Summary – The nematode diversity in soil and litter was investigated on Mount Hamiguitan, the Philippines, along four eco-habitatsfrom elevations of 75-1600 m a.s.l. A total of 155 and 467 nematodes were identified to 39 and 62 genera from litter and soil,respectively. The nematode assemblages and diversity did not show any relation to eco-habitat or elevation. Bacterivorous nematodeswere the most common group (37.5%). Acrobeloides was most abundant from the soil and Aphelenchoides from the litter. Bicirronemahamiguitanense n. sp. is herein described based on morphology, morphometrics and molecular data. The new species has the followingdiagnostic features: a wide lateral field one-fifth of its body diam. with four incisures forming two ornamented ridges; gubernaculumwith proximal thickening; spicules (35-38 μm) and gubernaculum (18-20 μm) longer than B. caledoniense; and with 37 molecularautapomorphies supporting its new species status. The phylogenetic position of the new species within Cephalobomorpha is discussed.
Keywords – molecular, morphology, morphometrics, new species, phylogeny, taxonomy.
Nematode diversity studies have historically focusedon temperate regions and only ten of 134 publishedecosystem surveys represented tropical forests (Boag &Yeates, 1998). Also, these studies only considered soilsamples, with very little or no attention given to theepiphytic communities (litter or canopy). However, recentstudies (Powers et al., 2009; Porazinska et al., 2010,2012) have taken into account both soil and litter material,with litter showing higher nematode diversity than soil.Nematode diversity was based only on either MolecularOperational Taxonomic Units (MOTUs) or OperationalClustered Taxonomic Units (OCTUs) for parallel taggedultra sequencing.
It is commonly thought that diversity tends to increaseas altitude increases (Stevens, 1992; Begon et al., 1996;Rahbek, 1997) and this has been found in vascular plantsand birds (Hunter & Yonzon, 1993; Wolf, 1994; Pendry& Proctor, 1997; Rahbek, 1997; Shepherd, 1998). Thereare very few studies on nematode altitudinal distributionand these were conducted in grassland, temperate orsubtropical forest, or desert environments, often with
∗ Corresponding author, e-mail: [email protected]
contradictory results. For instance, nematode diversitywas found to decline as altitude increases (Weaver &Smolik, 1987; Chen et al., 2009; Hánel & Cerevková,2010); however, in other studies, diversity was not relatedto elevation (Yeates, 1974; Steinberger et al., 2001;Treonis et al., 2012).
The Mount Hamiguitan Range Wildlife Sanctuary is lo-cated in the southeast of Mindanao Island, the Philippines.It has been identified by Conservation International as oneof the Philippines’ biodiversity hotspots (see Myers et al.,2000) because of its high degree of species endemism inflora and fauna, and the threat to this from human inter-ference (Amoroso & Aspiras, 2011). Recently, this sanc-tuary was included on the UNESCO World Heritage List(http://whc.unesco.org/en/list/1403). Its tropical climate,wide range of altitude from 75-1637 m a.s.l. and variedeco-habitats make it a good candidate for this altitudinalnematode community study. We hypothesised that thereexists an elevational pattern of nematode abundance, di-versity and community structure in Mount Hamiguitan.
© Koninklijke Brill NV, Leiden, 2015 DOI 10.1163/15685411-00002870

X. Qing et al.
Among major groups of nematodes, the suborder Ce-phalobina Andrássy, 1974 (Infraorder CephalobomorphaDe Ley & Blaxter, 2002) is considered one of the morpho-logically and ecologically diverse groups of free-livingmicrobivores (Nadler et al., 2006; Holovachov et al.,2009a) that are widely distributed worldwide and areabundant in nutrient-poor soils with some of the mostrare species recovered from the forest floor. One suchrare genus is Bicirronema Andrássy, 1978 and, to date,only two species have been reported: B. caledoniense An-drássy, 1978 from the tropical island of New Caledoniaand B. peruense Andrássy, 1986 from Peru. Bicirronemawas originally classified within the family Chambersiel-lidae Thorne, 1937, subfamily Bicirronematinae, whichshows putative similarities to the other two subfamiliesChambersiellinae Thorne, 1937 and Macrolaiminae San-wal, 1971 in the labial region (presence of cirri), the stoma(cuticularised cheilostom and gymnostom) and nerve ringposition (anterior to the isthmus). However, a more re-cent study (De Ley, 1997) found that the reproductive sys-tem differed from Chambersiellidae (spermatheca not off-set vs offset in Bicirronema). Subsequently, the subfamilywas raised to family rank and transferred to the superfam-ily Cephaloboidea Filipjev, 1934, based on the homologyof the female reproductive system (presence of an off-set spermatheca at the oviduct-uterus junction) and stoma(stegostom subdivided into four subsections) in the familyBicirronematidae, Cephalobomorpha. Available molecu-lar data for this genus and additional related taxa collectedfrom the Mount Hamiguitan study will therefore shedlight on its phylogenetic position in Cephalobomorpha.
Materials and methods
SAMPLE COLLECTION AND PROCESSING
Samples were collected from Mount Hamiguitan RangeWildlife Sanctuary, the Philippines, in August 2012 (Ta-ble 1). Four sampling sites were identified at random fromeach of the four eco-habitats proposed by Amoroso &Aspiras (2011). At each sampling site, four trees (twolarge and two small, under canopy trees) were identified,and from around each tree, one soil (15 cm depth) andone litter (leaves and organic matter overlying the soil)composite sample were each bulked from 2-4 subsamplesfrom 15 × 15 cm areas. There was a total of 22 sam-ples from four eco-habitats as follows: Agro-forestry (75-420 m a.s.l.: 2), Dipterocarp (421-920 m a.s.l.: 6), Mon-tane (921-1160 m a.s.l.: 10), and Mossy/Pygmy (1161-1637 m a.s.l.: 4). Due to difficulty in getting access to
steep slopes and sites, as well as the loss of some sam-ples during transport, there was unequal sample size perhabitat.
The soil samples varied in organic matter and moisturecontent depending on the site and the prevailing rainfall.During the sampling days, we also had to contend withheavy rains, so most samples were soaked. To eliminatevariation associated with sample weight, nematodes wereextracted from soil or finely-chopped litter of 100 ml asmeasured with a beaker. Samples were incubated for 48 hon plastic trays lined with paper towels. Nematodes wereconcentrated using a sieve (25 μm opening), replacing thewater in the process with, and preserving the nematodesin, DESS solution (Yoder et al., 2006). Prior to laboratorystudies, DESS-preserved specimens were rinsed multipletimes with deionised water to remove all traces of thepreservative, transferred to anhydrous glycerin (Seinhorst,1962; as modified by Sohlenius & Sandor, 1987), andmounted on microscope slides.
MORPHOLOGICAL CHARACTERISATION
Measurements and drawings were prepared manuallywith a drawing tube mounted on an Olympus BX51 DICMicroscope equipped with an Olympus C5060Wz cam-era for photography. The holotype was recorded as avideo clip mimicking a multifocal observation througha light microscope (LM) following the Video Captureand Editing (VCE) procedure developed by De Ley &Bert (2002). The resulting virtual specimens are availableonline at http://www.nematodes.ugent.be/vce.html. Illus-trations were prepared using Illustrator® CS 2 software(Adobe Systems) based on LM drawings. For scanningelectron microscopy (SEM), specimens were processedand viewed following the procedure of Steel et al. (2011).
MOLECULAR CHARACTERISATION
Following VCE, genomic DNA was extracted fromidentified glycerin-processed and slide-mounted speci-mens. PCR reaction and sequencing of the D2-D3 ex-pansion segment of the large sub-unit (LSU) rDNA wasdone following the protocol of Múnera Uribe et al.(2010). The LSU rDNA sequence dataset and outgrouptaxa Coslenchus sp. and Aphelenchus sp. (see Nadleret al., 2006; Holovachov et al., 2009b) were alignedwith MUSCLE (Edgar, 2004) using the default settings.Bayesian phylogenetic inference (BI) was performed witha general time reversible model with rate variation acrosssites and a proportion of invariable sites (GTR + I +
326 Nematology
Soil and litter nematode diversity of Mount Hamiguitan
Table 1. Sample source or flora around which soil and litter were collected at different elevations and eco-habitats in Mount Hamiguitan,Mindanao, the Philippines.
No. Flora Eco-habitat Elevation (m a.s.l.) Coordinates
1 Tanguile ((Shorea polysperma) (Blanco) Merr.) Dipterocarp 485 06°43.577′N, 126°09.036′E2 Tanguile Dipterocarp 523 06°43.545′N, 126°09.025′E3 Hambabawod/Hambabalud Dipterocarp 529 06°43.533′N, 126°09.017′E
(Neoaudea formicaria (Elmer) Merr.)4 Yakal (Shorea astylosa Foxw.) Dipterocarp 532 06°43.377′N, 126°09.409′E5 Small bush Dipterocarp 532 06°43.377′N, 126°09.409′E6 Small bush Lower Dipterocarp 730 06°43.457′N, 126°09.609′E9 Cedar (Dacrydium elatum (Roxb.) Wall. ex Hook.) Montane 948 06°43.858′N, 126°10.079′E
11 Ulayan ((Lithocarpus llanosii A. DC.) Rehder) Montane 950 06°43.863′N, 126°10.088′E12 Sagimsim ((Syzygium brevistylum C.B. Rob). Merr.) Montane 952 06°43.870′N, 126°10.134′E14 Tree fern (Cyathea sp.) Montane 972 06°43.939′N, 126°10.066′E15 Bitanghol (Calophyllum blancoi Planchon) Montane 972 06°43.939′N, 126°10.066′E16 Tanguile Upper Montane 993 06°43.978′N, 126°10.060′E17 Lumbay (Metroxylon sivestre Mart.) Upper Montane 999 06°43.978′N, 126°10.040′E18 Mango (Mangifera indica L.) Upper Montane 997 06°43.984′N, 126°10.074′E19 Bayog (Pterospermum diversifolium Blume) Upper Montane 979 06°43.958′N, 126°10.091′E20 Dead tree (unknown) Upper Montane 1008 06°43.972′N, 126°10.077′E21 Pygmy tree (unidentified) Mossy/Pygmy 1220 06°44.172′N, 126°10.377′E22 Grass and bush (unidentified) Pygmy 1600 06°44.373′N, 126°10.941′E23 Pitcher plant (Nepenthes peltata Sh. Kurata) Typical mossy 1423 06°44.345′N, 126°10.950′E25 Mossy tree (unidentified) Typical mossy 1435 06°43.972′N, 126°11.186′E27 Mango (Mangifera indica L.) Agro-forestry 253 06°42.536′N, 126°07.708′E28 Jackfruit (Artocarpus heterophyllus Lam.) Agro-ecosystem 106 06°42.934′N, 126°07.444′E
Flora for which Latin binomials are not known are given common names.
G). The trees were generated using the last 106 gen-erations, well beyond the burn-in value and with aver-age standard deviations of split frequencies approachingzero (<0.005). Molecular character states were traced byMesquite 2.75 (Maddison & Maddison, 2011) and autapo-morphies were mapped on the Bayesian majority consen-sus tree using MUSCLE alignment.
DATA ANALYSES
The terminology of the anterior structures followsDe Ley et al. (1995). Nematode taxa were assignedto trophic groups according to Yeates et al. (1993).If no feeding type was available, their trophic groupwas assigned according to other genera within the samefamily. The ratio of fungivores to bacterivores (F/(F +B)) is calculated based on Neher (1999). The MaturityIndex (MI) was calculated following Bongers (1990), inwhich nematode genera were assigned values of a “1-5coloniser-persister (c-p) scale” according to their r andk life history characteristics (Bongers, 1990; Bongers& Ferris, 1999). To provide an index of diversity, theShannon-Wiener index was calculated (Krebs, 1989). To
test if the differences of MI, Shannon-Wiener index andF/(F + B) ratio between soil and litter were significant,t tests (for parametric data) and Wilcoxon tests (for non-parametric data) were used, and to test differences amongeco-habitats, ANOVAF tests (for parametric data) andKruskal-Wallis (for non-parametric data) tests were used.All statistical analyses were performed with SPSS 19.0.A rarefaction index (Sanders, 1968; Heck et al., 1975)was calculated to correct for differences in number ofidentified taxa caused by different sampling sizes (i.e., 39genera litter samples and 62 in soil samples).
Results
NEMATODE DIVERSITY
A total of 155 nematodes were identified from sevenlitter samples compared to 467 from 21 soil samples.Of these, litter nematodes belonged to 39 genera (withunidentified genera in eight families) whereas soil nema-todes were from 62 genera (with unidentified genera in12 families) (see Tables 2, 3). Acrobeloides was found the
Vol. 17(3), 2015 327
X. Qing et al.
Table 2. List and number of nematode taxa recovered from soil samples, with corresponding trophic group and c-p value, from MountHamiguitan, Mindanao, the Philippines.
Taxon Species Trophic group Ecohabitat Site
Order: EnoplidaFamily: AlaimidaeAlaimus 12 1 DP, MT, MP 3, 5, 11, 12, 14, 19, 21Amphidelus 3 1 MT, AG 12, 16, 27Etamphidelus 2 1 MT 11, 19Paramphidelus 7 1 MT, MP 12, 25Family: RhabdolaimidaeRhabdolaimus 3 1 DP, MT 6, 19Family: IronidaeIronus 15 5 MT, MP 14, 20, 25
Order: TriplonchidaFamily: PrismatolaimidaePrismatolaimus 1 1 MT 16
Order: DorylaimidaFamily: AulolaimidaeAulolaimus 5 1 MT 17Unidentified genera 19 1 MT, MP 9, 12, 17, 18, 19, 25Family: BelondiridaeAxonchium 14 4 DP, MT, MP 4, 17, 18, 25Belondira 4 4 AG 28Dorylaimellus 1 4 MT 20Unidentified genera 5 4 DP, AG 4, 27, 28Family: QudsianematidaeCrassolabium 2 5 MT 12Eudorylaimus 8 5 DP, MT, MP, AG 5, 12, 25, 27, 28Unidentified genera 14 5 DP, MT, MP, AG 1, 2, 4, 5, 6, 12, 16, 25, 28Family: DorylaimidaeDorylaimus 8 4 DP, MT, MP 2, 12, 16, 21Mesodorylaimus 3 4 DP, MP 4, 25Prodorylaimus 3 4 MT, MP 16, 22Unidentified genera 62 4 DP, MT, MP, AG 4, 6, 12, 16, 17, 18, 19, 20, 23, 25, 28Family: LeptonchidaeLeptonchus 1 2 MP 22Unidentified genera 4 2 DP, MT, MP 5, 18, 19, 25Family: NordiidaeOriverutus 3 3 DP, MT 6, 16Pungentus 2 3 MT, AG 12, 28Unidentified genera 3 3 MT, MP 16, 20, 25Family: MydonomidaeTimmus 1 4 DP 4Unidentified genera 3 4 DP, MT 4, 17Family: TylencholaimellidaeTylencholaimellus 3 2 MP 23Unidentified genera 1 2 MP 23Family: TylencholaimidaeTylencholaimus 24 2 DP, MT, MP, AG 2, 4, 11, 16, 19, 20, 25, 27, 28Unidentified genera 38 2 DP, MT, MP, AG 2, 3, 4, 5, 6, 9, 11, 12, 14, 17, 19, 21, 25, 28Family: Actinolaimidae
328 Nematology
Soil and litter nematode diversity of Mount Hamiguitan
Table 2. (Continued.)
Taxon Species Trophic group Ecohabitat Site
Unidentified genera 1 5 MT 18Family: AporcelaimidaeUnidentified genera 6 5 DP, MT, AG 6, 12, 17, 28Family: LongidoridaeUnidentified genera 4 3 DP, MP, AG 5, 21, 28
Order: MononchidaFamily: MononchidaeClarkus 4 5 DP, MT 2, 6, 16Family: CobbonchidaeCobbonchus 1 5 MT 14Family: MononchulidaeMononchulus 1 5 AG 28Family: MylonchulidaeMylonchulus 7 5 DP, MP, AG 1, 6, 22, 28
Order: DesmodoridaFamily: DesmodoridaeProdesmodora 3 1 MP 23
Order: MonhysteridaFamily: MonhysteridaeGeomonhystera 1 1 DP 2
Order: AraeolaimidaFamily: DiplopeltidaeCylindrolaimus 2 1 DP, MT 4, 16
Order: PlectidaFamily: PlectidaeHaliplectus 2 1 MT 14Plectus 10 1 DP, MT, AG 3, 6, 17, 27, 28Family: LeptolaimidaeAphanolaimus 8 1 MT, MP, AG 11, 12, 20, 25, 27Chronogaster 12 1 DP, MT, MP 4, 14, 25
Order: AphelenchidaFamily: AphelenchidaeAphelenchus 3 2 DP, AG 6, 28Family: AphelenchoididaeAphelenchoides 17 2 DP, AG 2, 3, 27, 28Seinura 2 5 MT 11, 14
Order: TylenchidaFamily: TylenchidaeBasiria 1 3 AG 28Filenchus 1 3 MT 14Malenchus 3 3 DP 2Tylenchus 1 3 MP 23Family: PratylenchidaePratylenchus 3 3 DP, MP 3, 25Family: HoplolaimidaeRotylenchus 2 3 DP, MP 21, 25
Vol. 17(3), 2015 329
X. Qing et al.
Table 2. (Continued.)
Taxon Species Trophic group Ecohabitat Site
Family TylenchulidaeTylenchulus 1 3 MT 14
Order: RhabditidaFamily: RhabditidaeBuetschlinema 1 1 MT 12Crustorhabditis 1 1 MT 12Mesorhabditis 7 1 DP, MT 2, 17Oscheius 1 1 MT 11Protorhabditis 3 1 MT 16, 20Rhabditis 1 1 MP 21Family: CephalobidaeAcrobeloides 29 1 DP, MT, AG 2, 6, 9, 11, 16, 17, 18, 19, 27Cephalobus 9 1 MP, AG 25, 27, 28Eucephalobus 1 1 AG 27Pseudacrobeles 13 2 DP, MT, MP, AG 1, 2, 16, 19, 23, 27Teratolobus 2 1 DP, AG 2, 28Zeldia 1 1 AG 28Family: DiplogastridaeDiplogaster 2 1 DP 2, 3Diplogasteritus 2 1 DP 4Family: PanagrolaimidaeHalicephalobus 2 1 AG 28Panagrellus 1 1 MT 20Panagrolaimus 3 1 DP, AG 3, 28Family: BicirronematidaeTricirronema 7 1 MT, MP 12, 14, 22
Trophic groups are assigned as: 1 = bacterivorous, 2 = fungivorous, 3 = plant-parasitic, 4 = omnivorous, 5 = predatory. Ecohabitatassigned as: DP = Dipterocarp, MT = Montane, MP = Mossy/Pygmy, AG = Agro-ecosystem.
most abundant from soil while Aphelenchoides was dom-inant from litter. Among identified taxa, Teratolobus sp.(Cephalobidae: Acrolobinae) and T. tentaculatum (Bicir-ronematidae) were selected for sequencing as they are thefirst molecular data in this genus and species, respectively.
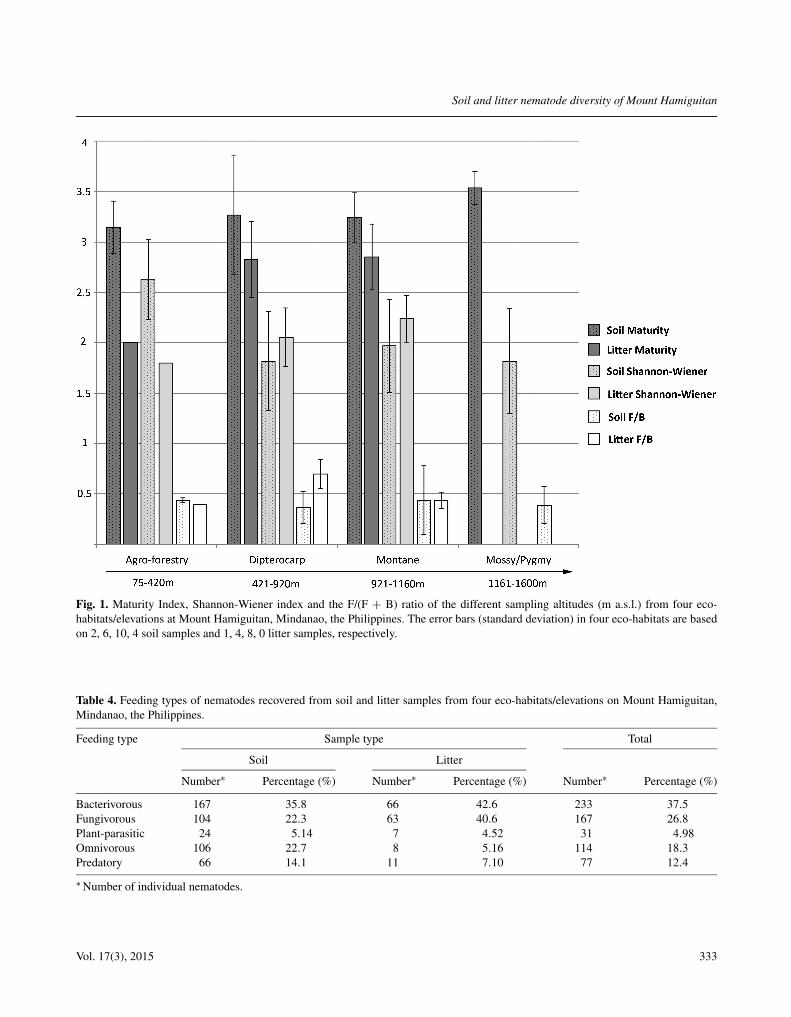
The diversity, F/(F + B) ratio and MI were not signifi-cantly different (P > 0.05) between eco-habitats/eleva-tions, nor between sample type (soil vs litter) (Fig. 1).On average, litter samples had a slightly higher diversity(2.22 ± 0.50) than soil (1.96 ± 0.69); however, follow-ing rarefaction analysis (to compensate for the number ofsample size analysed), the soil is slightly more diversethan litter (39 recorded taxa in litter vs 49.6 correctednumber of taxa in the soil).
Regardless of eco-habitats, bacterivores were the mostabundant trophic group (37.5%), while plant-parasiteswere the least abundant (5%) in both litter and soil(Table 4).
DESCRIPTION OF NEW SPECIES
Bicirronema hamiguitanense* n. sp.(Figs 2-4)
MEASUREMENTS
See Table 5.
DESCRIPTION
Adults
In both sexes, body ventrally curved to C-shapedfollowing DESS fixation. Cuticle loose and thick (2.3-2.8 μm at mid-body). Annulations 1.2-1.4 μm wide.Lateral field areolated, starting at ca 35 μm anterior to
* The new species is named after the Mount Hamiguitan RangeWildlife Sanctuary where the sample was collected.
330 Nematology
Soil and litter nematode diversity of Mount Hamiguitan
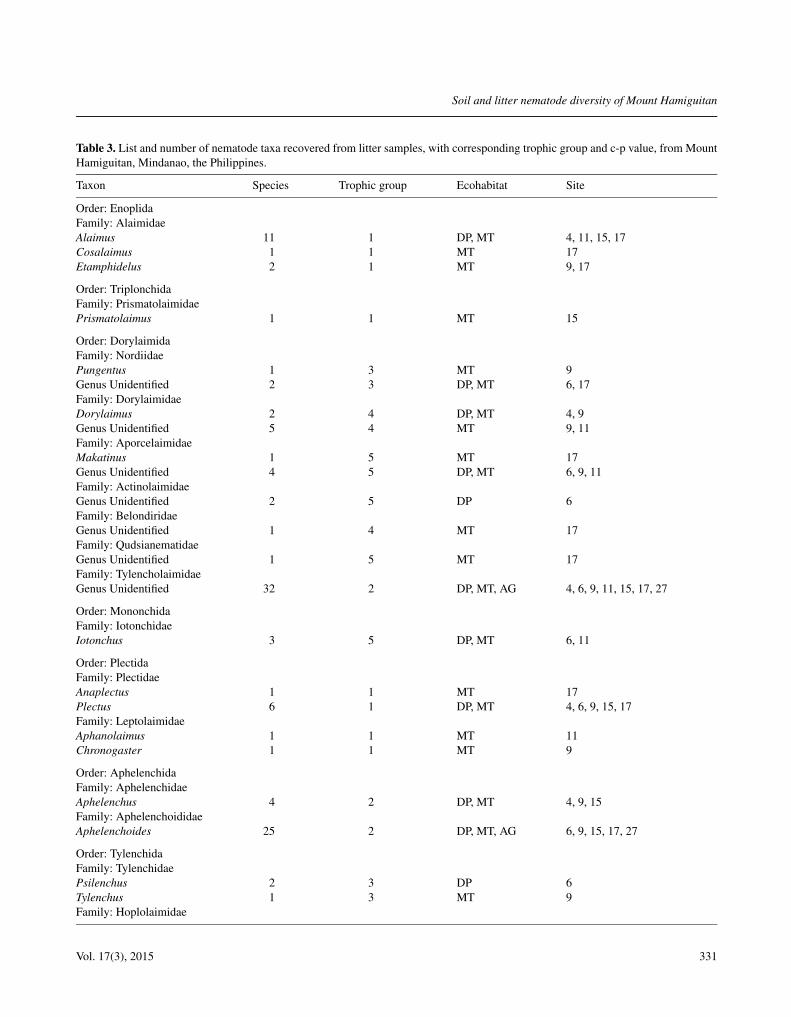
Table 3. List and number of nematode taxa recovered from litter samples, with corresponding trophic group and c-p value, from MountHamiguitan, Mindanao, the Philippines.
Taxon Species Trophic group Ecohabitat Site
Order: EnoplidaFamily: AlaimidaeAlaimus 11 1 DP, MT 4, 11, 15, 17Cosalaimus 1 1 MT 17Etamphidelus 2 1 MT 9, 17
Order: TriplonchidaFamily: PrismatolaimidaePrismatolaimus 1 1 MT 15
Order: DorylaimidaFamily: NordiidaePungentus 1 3 MT 9Genus Unidentified 2 3 DP, MT 6, 17Family: DorylaimidaeDorylaimus 2 4 DP, MT 4, 9Genus Unidentified 5 4 MT 9, 11Family: AporcelaimidaeMakatinus 1 5 MT 17Genus Unidentified 4 5 DP, MT 6, 9, 11Family: ActinolaimidaeGenus Unidentified 2 5 DP 6Family: BelondiridaeGenus Unidentified 1 4 MT 17Family: QudsianematidaeGenus Unidentified 1 5 MT 17Family: TylencholaimidaeGenus Unidentified 32 2 DP, MT, AG 4, 6, 9, 11, 15, 17, 27
Order: MononchidaFamily: IotonchidaeIotonchus 3 5 DP, MT 6, 11
Order: PlectidaFamily: PlectidaeAnaplectus 1 1 MT 17Plectus 6 1 DP, MT 4, 6, 9, 15, 17Family: LeptolaimidaeAphanolaimus 1 1 MT 11Chronogaster 1 1 MT 9
Order: AphelenchidaFamily: AphelenchidaeAphelenchus 4 2 DP, MT 4, 9, 15Family: AphelenchoididaeAphelenchoides 25 2 DP, MT, AG 6, 9, 15, 17, 27
Order: TylenchidaFamily: TylenchidaePsilenchus 2 3 DP 6Tylenchus 1 3 MT 9Family: Hoplolaimidae
Vol. 17(3), 2015 331
X. Qing et al.
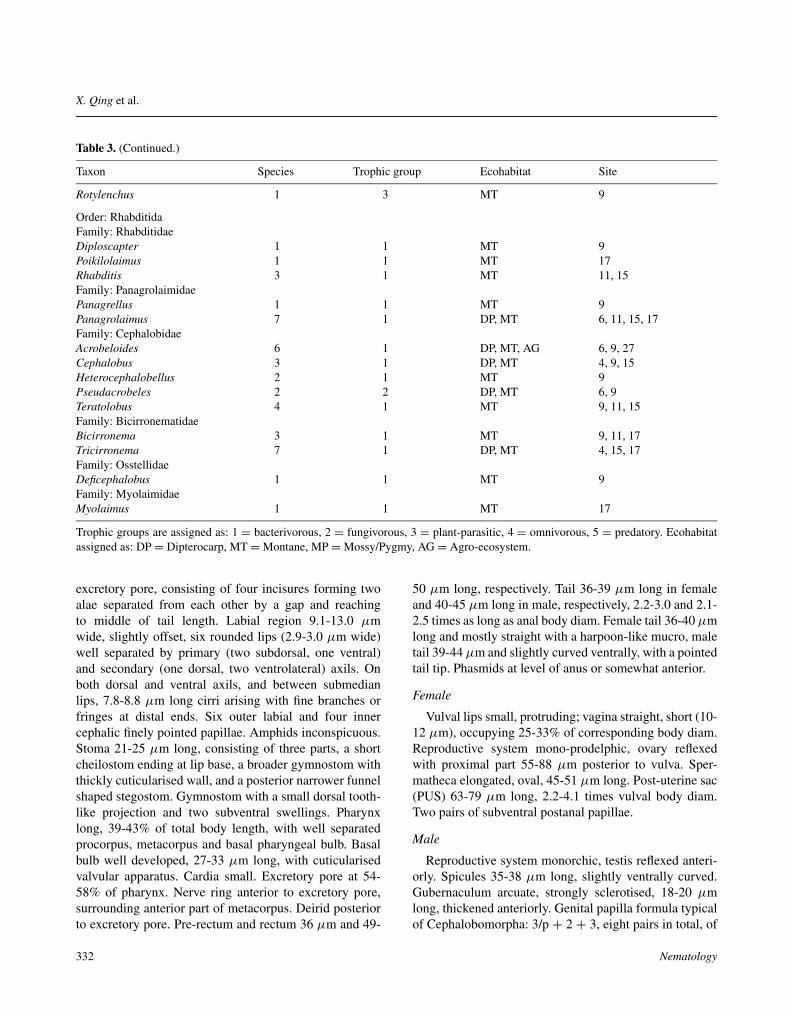
Table 3. (Continued.)
Taxon Species Trophic group Ecohabitat Site
Rotylenchus 1 3 MT 9
Order: RhabditidaFamily: RhabditidaeDiploscapter 1 1 MT 9Poikilolaimus 1 1 MT 17Rhabditis 3 1 MT 11, 15Family: PanagrolaimidaePanagrellus 1 1 MT 9Panagrolaimus 7 1 DP, MT 6, 11, 15, 17Family: CephalobidaeAcrobeloides 6 1 DP, MT, AG 6, 9, 27Cephalobus 3 1 DP, MT 4, 9, 15Heterocephalobellus 2 1 MT 9Pseudacrobeles 2 2 DP, MT 6, 9Teratolobus 4 1 MT 9, 11, 15Family: BicirronematidaeBicirronema 3 1 MT 9, 11, 17Tricirronema 7 1 DP, MT 4, 15, 17Family: OsstellidaeDeficephalobus 1 1 MT 9Family: MyolaimidaeMyolaimus 1 1 MT 17
Trophic groups are assigned as: 1 = bacterivorous, 2 = fungivorous, 3 = plant-parasitic, 4 = omnivorous, 5 = predatory. Ecohabitatassigned as: DP = Dipterocarp, MT = Montane, MP = Mossy/Pygmy, AG = Agro-ecosystem.
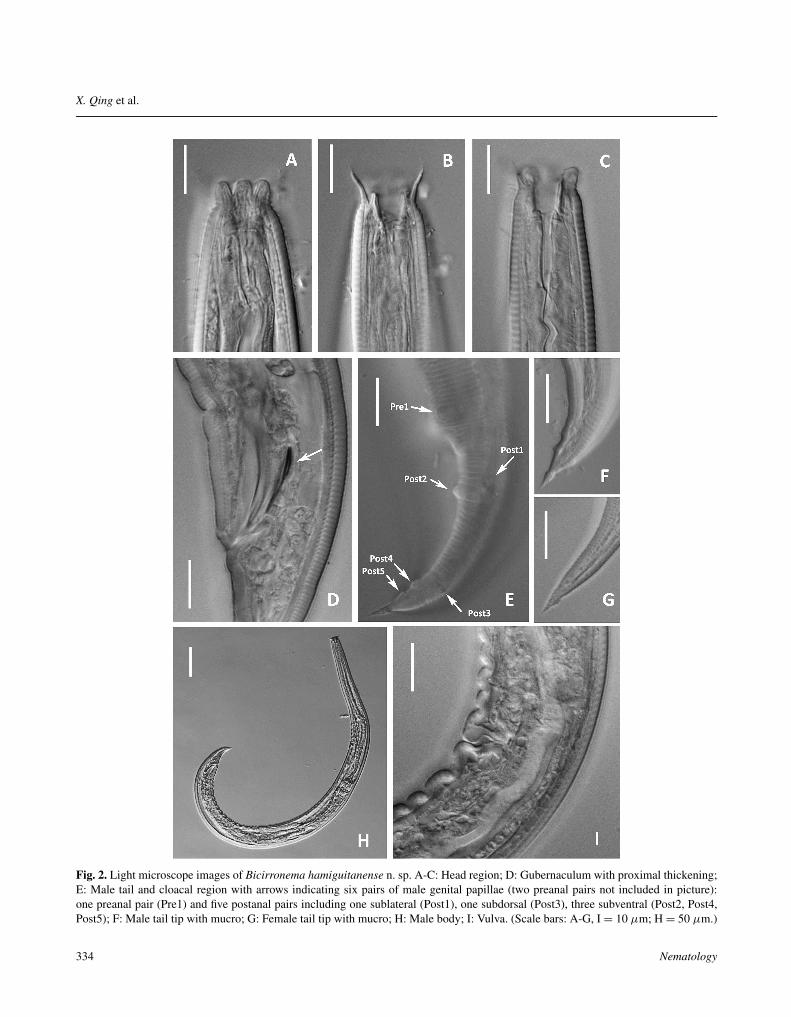
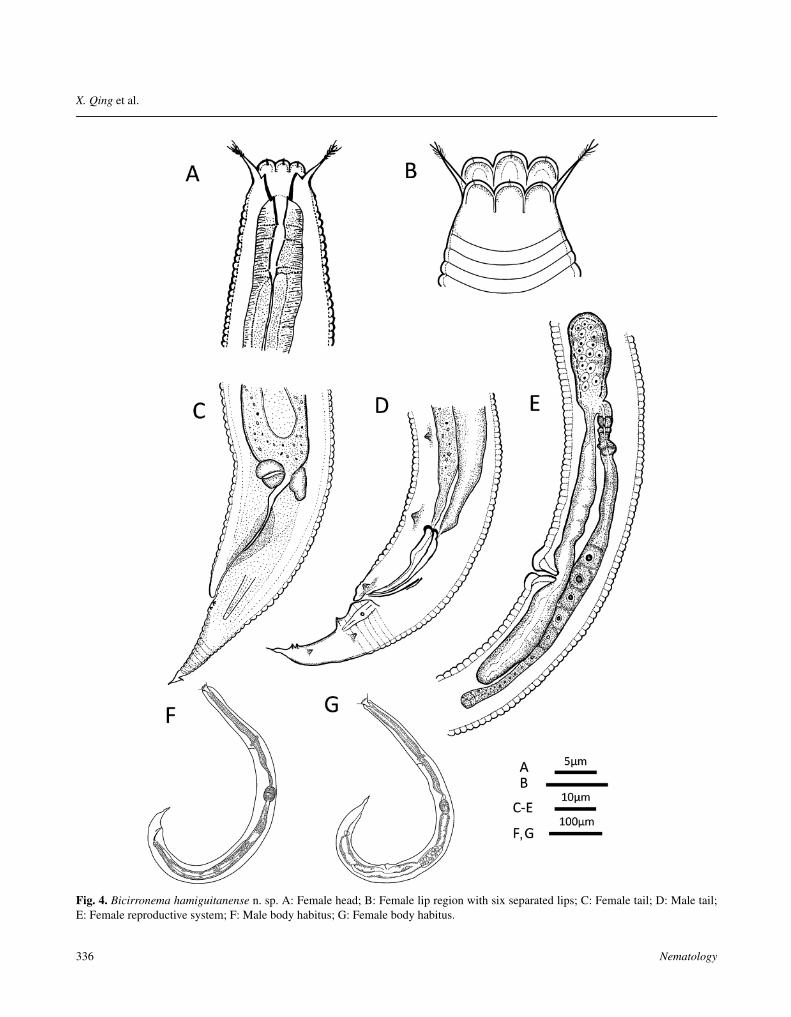
excretory pore, consisting of four incisures forming twoalae separated from each other by a gap and reachingto middle of tail length. Labial region 9.1-13.0 μmwide, slightly offset, six rounded lips (2.9-3.0 μm wide)well separated by primary (two subdorsal, one ventral)and secondary (one dorsal, two ventrolateral) axils. Onboth dorsal and ventral axils, and between submedianlips, 7.8-8.8 μm long cirri arising with fine branches orfringes at distal ends. Six outer labial and four innercephalic finely pointed papillae. Amphids inconspicuous.Stoma 21-25 μm long, consisting of three parts, a shortcheilostom ending at lip base, a broader gymnostom withthickly cuticularised wall, and a posterior narrower funnelshaped stegostom. Gymnostom with a small dorsal tooth-like projection and two subventral swellings. Pharynxlong, 39-43% of total body length, with well separatedprocorpus, metacorpus and basal pharyngeal bulb. Basalbulb well developed, 27-33 μm long, with cuticularisedvalvular apparatus. Cardia small. Excretory pore at 54-58% of pharynx. Nerve ring anterior to excretory pore,surrounding anterior part of metacorpus. Deirid posteriorto excretory pore. Pre-rectum and rectum 36 μm and 49-
50 μm long, respectively. Tail 36-39 μm long in femaleand 40-45 μm long in male, respectively, 2.2-3.0 and 2.1-2.5 times as long as anal body diam. Female tail 36-40 μmlong and mostly straight with a harpoon-like mucro, maletail 39-44 μm and slightly curved ventrally, with a pointedtail tip. Phasmids at level of anus or somewhat anterior.
Female
Vulval lips small, protruding; vagina straight, short (10-12 μm), occupying 25-33% of corresponding body diam.Reproductive system mono-prodelphic, ovary reflexedwith proximal part 55-88 μm posterior to vulva. Sper-matheca elongated, oval, 45-51 μm long. Post-uterine sac(PUS) 63-79 μm long, 2.2-4.1 times vulval body diam.Two pairs of subventral postanal papillae.
Male
Reproductive system monorchic, testis reflexed anteri-orly. Spicules 35-38 μm long, slightly ventrally curved.Gubernaculum arcuate, strongly sclerotised, 18-20 μmlong, thickened anteriorly. Genital papilla formula typicalof Cephalobomorpha: 3/p + 2 + 3, eight pairs in total, of
332 Nematology
Soil and litter nematode diversity of Mount Hamiguitan
Fig. 1. Maturity Index, Shannon-Wiener index and the F/(F + B) ratio of the different sampling altitudes (m a.s.l.) from four eco-habitats/elevations at Mount Hamiguitan, Mindanao, the Philippines. The error bars (standard deviation) in four eco-habitats are basedon 2, 6, 10, 4 soil samples and 1, 4, 8, 0 litter samples, respectively.
Table 4. Feeding types of nematodes recovered from soil and litter samples from four eco-habitats/elevations on Mount Hamiguitan,Mindanao, the Philippines.
Feeding type Sample type Total
Soil Litter
Number∗ Percentage (%) Number∗ Percentage (%) Number∗ Percentage (%)
Bacterivorous 167 35.8 66 42.6 233 37.5Fungivorous 104 22.3 63 40.6 167 26.8Plant-parasitic 24 5.14 7 4.52 31 4.98Omnivorous 106 22.7 8 5.16 114 18.3Predatory 66 14.1 11 7.10 77 12.4
∗ Number of individual nematodes.
Vol. 17(3), 2015 333
X. Qing et al.
Fig. 2. Light microscope images of Bicirronema hamiguitanense n. sp. A-C: Head region; D: Gubernaculum with proximal thickening;E: Male tail and cloacal region with arrows indicating six pairs of male genital papillae (two preanal pairs not included in picture):one preanal pair (Pre1) and five postanal pairs including one sublateral (Post1), one subdorsal (Post3), three subventral (Post2, Post4,Post5); F: Male tail tip with mucro; G: Female tail tip with mucro; H: Male body; I: Vulva. (Scale bars: A-G, I = 10 μm; H = 50 μm.)
334 Nematology

Soil and litter nematode diversity of Mount Hamiguitan
Fig. 3. Scanning electron micrographs (SEM) of female Bicirronema hamiguitanense n. sp. A: Head region with six separated lips; B:Lateral field with two ornamented ridges; C: Ventral view of vulva; D: Tail; E: Tail terminus with postanal papillae (arrows indicate thesubventral papillae 1 and 2). (Scale bar = 10 μm.)
Vol. 17(3), 2015 335
X. Qing et al.
Fig. 4. Bicirronema hamiguitanense n. sp. A: Female head; B: Female lip region with six separated lips; C: Female tail; D: Male tail;E: Female reproductive system; F: Male body habitus; G: Female body habitus.
336 Nematology
Soil and litter nematode diversity of Mount Hamiguitan
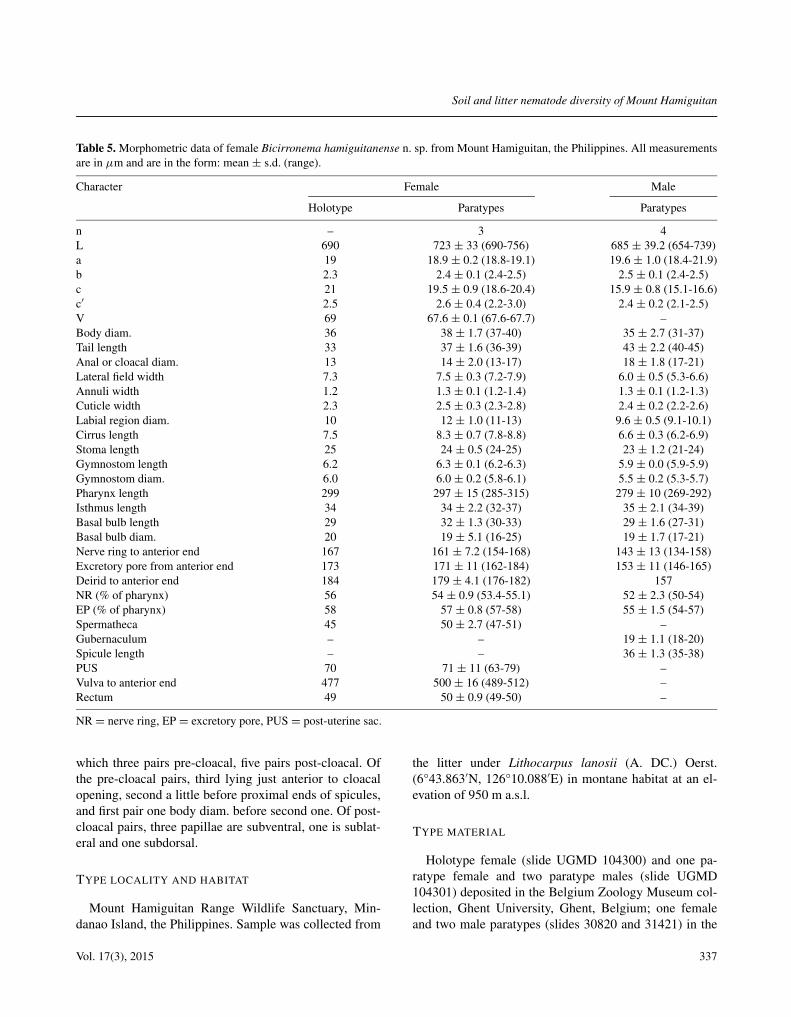
Table 5. Morphometric data of female Bicirronema hamiguitanense n. sp. from Mount Hamiguitan, the Philippines. All measurementsare in μm and are in the form: mean ± s.d. (range).
Character Female Male
Holotype Paratypes Paratypes
n – 3 4L 690 723 ± 33 (690-756) 685 ± 39.2 (654-739)a 19 18.9 ± 0.2 (18.8-19.1) 19.6 ± 1.0 (18.4-21.9)b 2.3 2.4 ± 0.1 (2.4-2.5) 2.5 ± 0.1 (2.4-2.5)c 21 19.5 ± 0.9 (18.6-20.4) 15.9 ± 0.8 (15.1-16.6)c′ 2.5 2.6 ± 0.4 (2.2-3.0) 2.4 ± 0.2 (2.1-2.5)V 69 67.6 ± 0.1 (67.6-67.7) –Body diam. 36 38 ± 1.7 (37-40) 35 ± 2.7 (31-37)Tail length 33 37 ± 1.6 (36-39) 43 ± 2.2 (40-45)Anal or cloacal diam. 13 14 ± 2.0 (13-17) 18 ± 1.8 (17-21)Lateral field width 7.3 7.5 ± 0.3 (7.2-7.9) 6.0 ± 0.5 (5.3-6.6)Annuli width 1.2 1.3 ± 0.1 (1.2-1.4) 1.3 ± 0.1 (1.2-1.3)Cuticle width 2.3 2.5 ± 0.3 (2.3-2.8) 2.4 ± 0.2 (2.2-2.6)Labial region diam. 10 12 ± 1.0 (11-13) 9.6 ± 0.5 (9.1-10.1)Cirrus length 7.5 8.3 ± 0.7 (7.8-8.8) 6.6 ± 0.3 (6.2-6.9)Stoma length 25 24 ± 0.5 (24-25) 23 ± 1.2 (21-24)Gymnostom length 6.2 6.3 ± 0.1 (6.2-6.3) 5.9 ± 0.0 (5.9-5.9)Gymnostom diam. 6.0 6.0 ± 0.2 (5.8-6.1) 5.5 ± 0.2 (5.3-5.7)Pharynx length 299 297 ± 15 (285-315) 279 ± 10 (269-292)Isthmus length 34 34 ± 2.2 (32-37) 35 ± 2.1 (34-39)Basal bulb length 29 32 ± 1.3 (30-33) 29 ± 1.6 (27-31)Basal bulb diam. 20 19 ± 5.1 (16-25) 19 ± 1.7 (17-21)Nerve ring to anterior end 167 161 ± 7.2 (154-168) 143 ± 13 (134-158)Excretory pore from anterior end 173 171 ± 11 (162-184) 153 ± 11 (146-165)Deirid to anterior end 184 179 ± 4.1 (176-182) 157NR (% of pharynx) 56 54 ± 0.9 (53.4-55.1) 52 ± 2.3 (50-54)EP (% of pharynx) 58 57 ± 0.8 (57-58) 55 ± 1.5 (54-57)Spermatheca 45 50 ± 2.7 (47-51) –Gubernaculum – – 19 ± 1.1 (18-20)Spicule length – – 36 ± 1.3 (35-38)PUS 70 71 ± 11 (63-79) –Vulva to anterior end 477 500 ± 16 (489-512) –Rectum 49 50 ± 0.9 (49-50) –
NR = nerve ring, EP = excretory pore, PUS = post-uterine sac.
which three pairs pre-cloacal, five pairs post-cloacal. Ofthe pre-cloacal pairs, third lying just anterior to cloacalopening, second a little before proximal ends of spicules,and first pair one body diam. before second one. Of post-cloacal pairs, three papillae are subventral, one is sublat-eral and one subdorsal.
TYPE LOCALITY AND HABITAT
Mount Hamiguitan Range Wildlife Sanctuary, Min-danao Island, the Philippines. Sample was collected from
the litter under Lithocarpus lanosii (A. DC.) Oerst.(6°43.863′N, 126°10.088′E) in montane habitat at an el-evation of 950 m a.s.l.
TYPE MATERIAL
Holotype female (slide UGMD 104300) and one pa-ratype female and two paratype males (slide UGMD104301) deposited in the Belgium Zoology Museum col-lection, Ghent University, Ghent, Belgium; one femaleand two male paratypes (slides 30820 and 31421) in the
Vol. 17(3), 2015 337

X. Qing et al.
Department of Nematology Nematode Collection, Uni-versity of California, Riverside, CA, USA. The remainingslides are with the first author.
DIAGNOSIS AND RELATIONSHIPS
The presence of two branches of cirri, wide cuticu-larised stoma and unusually long pharynx are charactersdiagnostic to Bicirronema. Bicirronema hamiguitanensen. sp. is characterised by three unique features: i) a widelateral field (20% of body diam.) with four incisuresforming two ornamented ridges; ii) gubernaculum withthickened anterior end; and iii) spicules 36 ± 1.3 (35-38) μm long and gubernaculum 19 ± 1.1 (18-20) μmlong.
Bicirronema hamiguitanense n. sp. differs from B.caledoniense and B. peruense, the only other nominalmembers of the genus, in a number of characters (seeTable 6).
MOLECULAR CHARACTERISATION
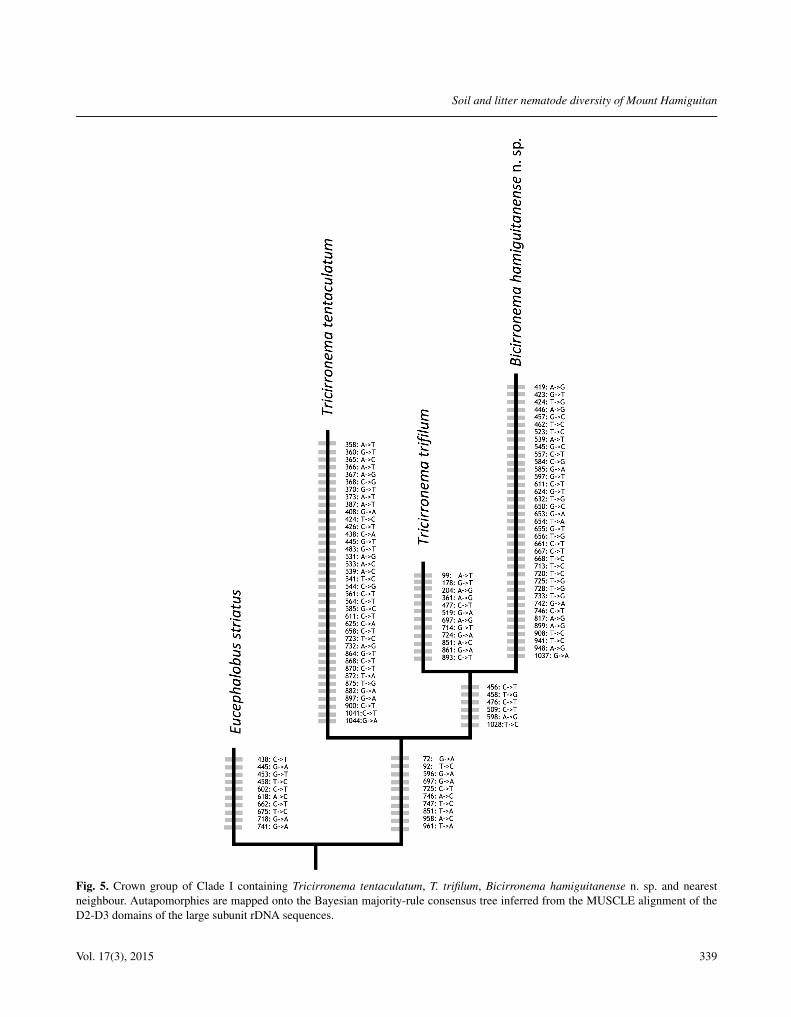
De novo sequences were deposited in GenBank un-der the accession numbers KJ652550 (Tricirronema ten-taculatum), KJ652551 (B. hamiguitanense n. sp.) andKJ652552 (Teratolobus sp.). The 41-taxa D2-D3 LSUrDNA dataset contained 1121 positions, 281 of whichwere variable and 208 parsimony informative. Furthercharacter analysis of MUSCLE-aligned dataset showedthat B. hamiguitanense n. sp. and its close relative, T.trifilum, have 37 and 12 autapomorphies, respectively(Fig. 5) and thus they are separated from each other basedon the phylogenetic species concept (Adams, 1998, 2001).
Discussion
NEMATODE DIVERSITY
Global nematode diversity patterns are equivocal. Sev-eral studies show that tropical ecosystems have a lowernematode diversity than temperate habitats (Procter, 1984;Groombridge, 1992; Giller, 1996; Maraun et al., 2007),while other studies have reported relatively higher trop-ical nematode diversity, e.g., tropical forest in India andCameroon (Pradhan & Dash, 1987; Bloemers et al.,1997). Zhang et al. (2012) identified 62 genera (from soilsamples of 100 g) in their study of four types of temper-ate forests in Changbai Mountain, China. Yeates (1973)listed 6-24 species from different tropical forests in theNew Hebrides (now Vanuatu). Our samples showed a rel-atively high diversity on genus level (39 genera and eightfamilies out of 153 individuals in litter and 62 genera and12 families out of 467 individuals in soil). Siemann et al.(1996) suggested species richness (S) and the number ofindividuals (I) within a size class scale as S = I0.5. For ourstudy, this calculation underestimates the diversity of oursamples around three-fold (i.e., 22 in soil and 12 in littervs the actual 62 in soil and 39 in litter), given that our ne-matodes were only identified to genus level (cf., Tables 2,3). Although a comparison with molecular defined unitsis not straightforward, two recent studies also reported ahigh diversity in tropical rain forest based on OCTU (Po-razinska et al., 2010) and MOTU (Powers et al., 2009).Thus, our study confirms the presence of a high nematodediversity in this tropical forest.
Our study is the first comparison of litter and soil ne-matode diversity in tropical forest based on morphology-based identification. After the rarefaction index was usedto eliminate difference in sample size, soil shows slightly
Table 6. Comparative diagnostic characters of Bicirronema hamiguitanense n. sp., B. caledoniense and B. peruense.
Character B. hamiguitanense n. sp. B. caledoniense B. peruense
Lateral field Wide (1/5 body diam.), 4incisures forming twoornamented ridges
Narrow, 2 incisures Narrow, 2 incisures
Spicule 36 (35-38) μm 28-32 μm Male not knownGubernaculum With proximal thickening, 19
(18-20) μmSimple, no proximal thickening, 16-18 μm Male not known
Post-uterine sac 2 vulval body diam. long 2 vulval body diam. long 0.3 vulval body diam. longBody length Female: 723 (690-756) μm Female: 760-880 μm Female: 470-490 μm
Male: 685 (654-739) μm Male: 670-780 μm Male: not known
338 Nematology
Soil and litter nematode diversity of Mount Hamiguitan
Fig. 5. Crown group of Clade I containing Tricirronema tentaculatum, T. trifilum, Bicirronema hamiguitanense n. sp. and nearestneighbour. Autapomorphies are mapped onto the Bayesian majority-rule consensus tree inferred from the MUSCLE alignment of theD2-D3 domains of the large subunit rDNA sequences.
Vol. 17(3), 2015 339
X. Qing et al.
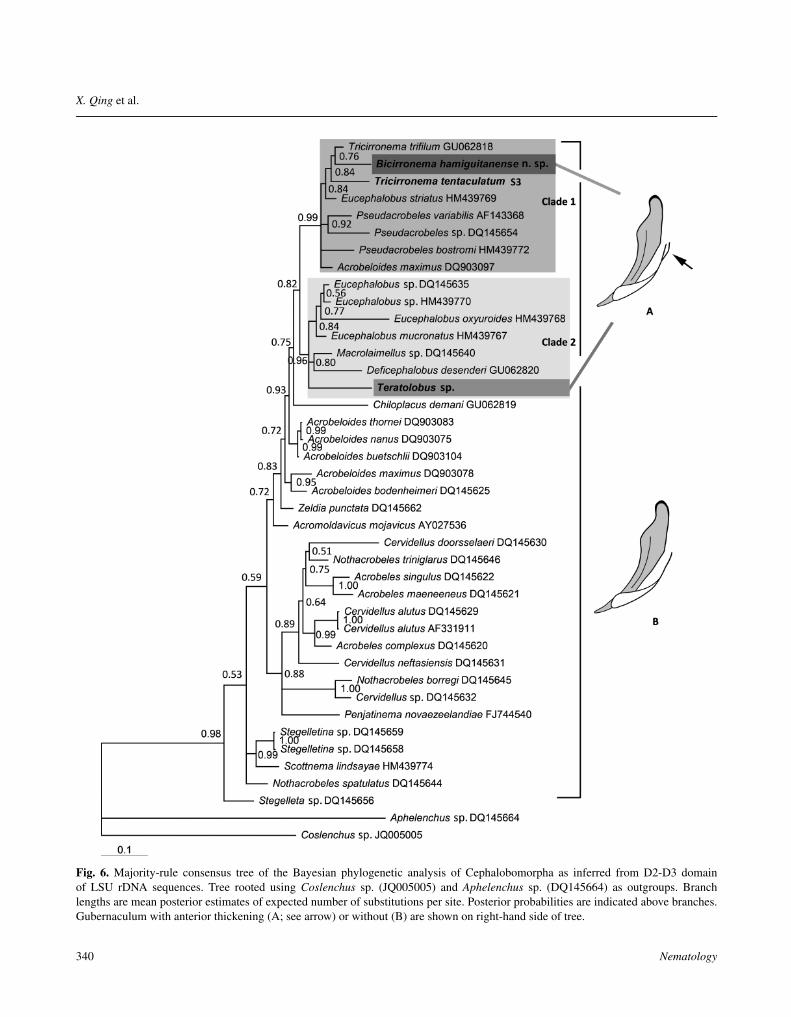
Fig. 6. Majority-rule consensus tree of the Bayesian phylogenetic analysis of Cephalobomorpha as inferred from D2-D3 domainof LSU rDNA sequences. Tree rooted using Coslenchus sp. (JQ005005) and Aphelenchus sp. (DQ145664) as outgroups. Branchlengths are mean posterior estimates of expected number of substitutions per site. Posterior probabilities are indicated above branches.Gubernaculum with anterior thickening (A; see arrow) or without (B) are shown on right-hand side of tree.
340 Nematology

Soil and litter nematode diversity of Mount Hamiguitan
higher diversity (39 in litter vs 49.6 in soil for 155 nema-todes identified) although this is likely an underestimate.Powers et al. (2009) and Porazinska et al. (2012) foundthat in a tropical rainforest of Costa Rica, 62 and 80% ofthe overall genetic diversity, existed in litter and under-story habitats (based on MOTU and OCTU, respectively).Accurate estimates of species richness should thereforeinclude nematodes not only from the soil and plant lit-ter above it, but canopy samples (e.g., epiphytes); terres-trial or arboreal pitcher plants (Bert et al., 2003); andother invertebrates like beetles (Kanzaki et al., 2011),earthworms (Ivanova et al., 2014), fig wasps (Powers etal., 2009), millipedes (Hunt, 1999) and termites (Kan-zaki et al., 2014). Invertebrate sampling was not includedin our current study; however, preliminary unpublisheddata from random collection of millipedes and also wa-ter from pitcher plants (with presumably insect-associatednematodes) have already yielded nematodes and DNA se-quences new to science (Quisado, 2014; Tandingan DeLey, unpubl. data).
In conclusion, the importance of appropriate samplingstrategy, combined with more efficient, accurate and sen-sitive diagnostic tools cannot be overemphasised. A two-pronged morphology and DNA sequence-based approachsuch as MOTU, OCTU, DNA barcodes or, when avail-able, DNA metabarcodes (using next generation sequenc-ing platforms for high-throughput taxon identification),will improve the efficiency and accuracy of biodiversityanalyses.
PHYLOGENETIC POSITION AND SPECIES
CHARACTERISATION
Bicirronematidae consists of three genera: BicirronemaAndrássy, 1978, Tricirronema Siddiqi, 1993 and Tru-alaimus Siddiqi, 1993. Holovachov et al. (2009b) con-ducted the first molecular analysis of this family based onLSU of Tricirronema trifilum Siddiqi, 1993. Their studyalready indicated that Tricirronema is sister of the twoPseudacrobeles Steiner, 1938 species and placed insidea crown group of genera with simple labial features andlong, acute tails. However, the family Bicirronematidaewas represented by only one out of nine known species.Our new sequences provide the first molecular data in thegenus Bicirronema making the first preliminary analysisat family level possible. The results indicate that three Bi-cirronematidae species together with Pseudacrobeles, Eu-cephalobus Steiner, 1936 and Acrobeloides (Cobb, 1924)Thorne, 1937 form a well-supported clade (Fig. 6; Clade1), while Bicirronema and Tricirronema clustered in one
clade but with relatively weak support (Posterior probabil-ity 0.84). Bicirronema appeared to be nested within Tricir-ronema, but also with weak support.
Holovachov et al. (2009b) noted that Pseudacrobelesbostromi Holovachov & De Ley, 2001 is morphologicallyintermediate between Bicirronematidae and Pseudacro-beles, as P. bostromi has a shorter mucronated tail and lowrounded lips which resemble Bicirronematidae. Our treeconfirms that Bicirronematidae is closely related to Pseu-dacrobeles, but the relationships within the latter genusremain unresolved.
The genus Teratolobus Andrássy, 1968 belongs to fam-ily Cephalobidae, subfamily Acrolobinae. Phylogeneticanalysis indicated its position within a well-supportedclade with Eucephalobus, Macrolaimellus Andrássy, 1966and Deficephalobus De Ley & Coomans, 1990. Thepresence of prominent cornua crurum at the guberna-culum was generally considered an apomorphy for sub-family Acrolobinae (contains Teratolobus Andrássy, 1968(= Panagroteratus Andrássy, 1986); Acrolobus Boström,1985; and Panagrolobus (Holovachov & Boström, 2006)).The presence of a thickened anterior of the gubernaculumreminiscent of a cornua crurum in B. hamiguitanense n.sp. seems to suggest its presence in Bicirronematidae and,if so, this thickened anterior gubernaculum has evolved atleast twice in different families (Cephalobidae and Bicir-ronematidae, Fig. 6). However, additional observation ofthis taxonomic character and more taxa sampling will beneeded to confirm this.
Although information of or knowledge on rare taxalike Bicirronematidae is improving, molecular samplingof known diversity is still relatively limited in Cephalobo-morpha. The para/polyphyly of several genera presentedhere implies that there is yet insufficient morphologicaldata and sequence information to propose any rearrange-ments in the current classification of families within thesuperfamily and underscores the importance of a morecomprehensive taxa sampling for Cephalobomorpha.
Acknowledgements
We thank Mr Ruel Colong, Protected Area Superinten-dent for Mount Hamiguitan and San Isidro Mayor EdgarSaulon for granting the sampling permit with very shortnotice; Ms Nic Smol for helping make the expedition pos-sible; Dr Lourdes Generalao (University of SoutheasternPhilippines), Dr Irvin Generalao (Southern PhilippinesAgri-Business and Marine and Aquatic School of Tech-nology) and Dr Lea Angsinco Jimenez (Davao Oriental
Vol. 17(3), 2015 341

X. Qing et al.
State College of Science and Technology) for generouslyproviding logistical support for the team, Dr Emily Fabre-gar (Lapanday Foods Corp.) for kindly allowing us the useof their laboratory facilities and for staff support in nema-tode extraction, Marjolein Couvreur for the SEM images,and Toon Janssen for assistance in phylogenetic analysis.The first author thanks the Flemish Interuniversity Coun-cil (VLIR) and China Scholarship Council (CSC) for pro-viding a grant. Research and teaching of the last author ismade possible through a grant from the Erasmus-MundusEuropean Master of Science in Nematology (EUMAINEat the University of Ghent, Ghent, Belgium).
References
Adams, B.J. (1998). Species concepts and the evolutionaryparadigm in modem nematology. Journal of Nematology 30,1-21.
Adams, B.J. (2001). The species delimitation uncertainty princi-ple. Journal of Nematology 33, 153-160.
Amoroso, V.B. & Aspiras, R.A. (2011). Hamiguitan Range:a sanctuary for native flora. Saudi Journal of BiologicalSciences 18, 7-15.
Andrássy, I. (1966). Erd- und Süsswasser-Nematoden ausGhana. Klasse Secernentea (Phasmidia). Annales Uni-versitatis Scientiarum Budapestensis de Rolando Eötvösnominatae-Sectio Biologica 8, 5-24.
Andrássy, I. (1968). Fauna Paraguayensis. 2. Nematoden ausden Galeriewäldern des Acaray-Flusses. Opuscula ZoologicaBudapestinensis 8, 167-315.
Andrássy, I. (1974). A nematodak evolucioja es rendszerezese.Magyar Tudomanyos Akademia Biologiai Tudmanyok Osz-talyanak Kozlemenyei 17, 13-58.
Andrássy, I. (1978). Bicirronema caledoniense n. gen., n. sp. andAmphidirhabditis longipapillata n. gen., n. sp. (Secernentia:Rhabditida), two remarkable soil-nematodes from New Cale-donia. Revue de Nématologie 1, 257-263.
Andrássy, I. (1986). Fifteen new nematode species from theSouthern Hemisphere. Acta Zoologica Hungarica 32, 1-33.
Begon, M., Harper, J. & Townsend, C.R. (1996). Ecology: in-dividuals, populations and communities. Oxford, UK, Black-well Scientific Publications.
Bert, W., Tandingan De Ley, I., van Driessche, R., Segers,H. & De Ley, P. (2003). Baujardia mirabilis gen. n., sp.n. from pitcher plants and its phylogenetic position withinPanagrolaimidae (Nematoda: Rhabditida). Nematology 5,405-420.
Bloemers, G.F., Hodda, M., Lambshead, P.J.D., Lawton, J.H.& Wanless, F.R. (1997). The effects of forest disturbance ondiversity of tropical soil nematodes. Oecologia 111, 575-582.
Boag, B. & Yeates, G.W. (1998). Soil nematode biodiversity interrestrial ecosystems. Biodiversity and Conservation 7, 617-630.
Bongers, T. (1990). The maturity index: an ecological measureof environmental disturbance based on nematode speciescomposition. Oecologia 83, 14-19.
Bongers, T. & Ferris, H. (1999). Nematode community structureas a bioindicator in environmental monitoring. Trends inEcology & Evolution 14, 224-228.
Boström, S. (1985). Description of Acrobeloides emarginatus(de Man, 1880) Thorne, 1937 and proposal of Acrolobus n.gen. (Nematoda: Cephalobidae). Revue de Nématologie 8,335-340.
Chen, X., You, W. & Yi, L. (2009). Community structure ofsoil fauna along an altitudinal gradient in Taibai Mountainof Tiantong Region, Zhejiang Province. Chinese Journal ofEcology 2, 10-17.
Cobb, N.A. (1924). Observations on nemas. II. Amended char-acterization of the nemic genera Cephalobus and Acrobeles.Journal of Parasitology 11, 108.
De Ley, P. (1997). The current state of affairs in identifica-tion and diagnosis of the genera of the family Cephalo-bidae (Nematoda: Rhabditida). Mededelingen Faculteit Land-bouwkundige en Toegepaste Biologische Wetenschappen Uni-versiteit Gent 62, 657-673.
De Ley, P. & Bert, W. (2002). Video capture and editing as a toolfor the storage, distribution, and illustration of morphologicalcharacters of nematodes. Journal of Nematology 34, 296-302.
De Ley, P. & Blaxter, M.L. (2002). Systematic position andphylogeny. In: Lee, D.L. (Ed.). The biology of nematodes.London, UK, Taylor & Francis, pp. 1-30.
De Ley, P. & Coomans, A. (1990). Terrestrial nematodes ofthe Galapagos Archipelago. I: three Rhabditida from IslaFernandina. Bulletin van het Koninklijk Belgisch Instituutvoor Natuurwetenschappen 60, 5-22.
De Ley, P., Van De Velde, M.C., Mounport, D., Baujard, P. &Coomans, A. (1995). Ultrastructure of the stoma in Cephalo-bidae, Panagrolaimidae and Rhabditidae, with proposal for arevised stoma terminology in Rhabditida (Nematoda). Nema-tologica 41, 153-182.
Edgar, R.C. (2004). MUSCLE: multiple sequence alignmentwith high accuracy and high throughput. Nucleic AcidsResearch 32, 1792-1797.
Filipjev, I.N. (1934). The classification of freeliving nematodesand their relation to the parasitic nematodes. SmithsonianMiscellaneous Collection, Washington 89, 1-63.
Giller, P.S. (1996). The diversity of soil communities, the “poorman’s tropical rainforest”. Biodiversity and Conservation 5,135-168.
Groombridge, B. (1992). Global biodiversity: status of theEarth’s living resources. London, UK, Chapman & Hall.
342 Nematology

Soil and litter nematode diversity of Mount Hamiguitan
Hánel, L. & Cerevková, A. (2010). Species and genera of soilnematodes in forest ecosystems of the Vihorlat ProtectedLandscape Area, Slovakia. Helminthologia 47, 123-135.
Heck, K.L., Van Belle, G. & Simberloff, D. (1975). Explicitcalculation of the rarefaction diversity measurement and thedetermination of sufficient sample size. Ecology 56, 1459-1461.
Holovachov, O. & Boström, S. (2006). Panagrolobus vanmege-nae, a new genus and species from The Netherlands andUkraine, with taxonomic revision and two new species ofthe genus Teratolobus Andrássy, 1968 (Nematoda: Cephalo-bidae). Nematology 8, 377-395.
Holovachov, O. & De Ley, P. (2001). Description of Pseudacro-beles (Bunobus) bostromi sp. n. (Rhabditida: Cephalobidae)from rotting wood in Roztochya, Ukraine. Journal of Nema-tode Morphology and Systematics 4, 21-29.
Holovachov, O., Tandingan De Ley, I., Mundo-Ocampo, M.& De Ley, P. (2009a). Identification of Cephaloboidea (Ne-matoda). EUMAINE, Gent and Nematology, UC Riverside,pp. 1-86.
Holovachov, O., Boström, S., Nadler, S. & De Ley, P. (2009b).Systematics and phylogenetic position of the genus Tricir-ronema Siddiqi, 1993 (Cephalobomorpha). Journal of Nema-tode Morphology and Systematics 12, 133-143.
Hunt, D.J. (1999). Five new species of Rhigonema Cobb, 1898(Rhigonematida: Rhigonematidae) from the Philippines andVietnam with observations on the morphology of the Indo-Malayan forms and nematophagy by R. voratum sp. n.Nematology 1, 337-361.
Hunter, M.L. & Yonzon, P. (1993). Altitudinal distributionsof birds, mammals, people, forests, and parks in Nepal.Conservation Biology 7, 420-423.
Ivanova, E.S., Sumaya, N.H.N., Mohagan, A.B. & Spiridonov,S.E. (2014). On the diversity of nematodes parasitic in inver-tebrates of Mindanao Island, Philippines. In: Proceedings ofthe 6th International Congress of Nematology, Cape Town,South Africa, May 4-9, 2014, pp. 189-190.
Kanzaki, N., Taki, H., Masuya, H., Okabe, K., Tanaka, R. &Abe, F. (2011). Diversity of stag beetle-associated nematodesin Japan. Environmental Entomology 40, 281-288.
Kanzaki, N., Li, H.-F., Lan, Y.-C. & Giblin-Davis, R.M. (2014).Description of two Pseudaphelenchus species (Tylenchomor-pha: Aphelenchoididae) associated with Asian termites andproposal of new subfamily Tylaphelenchinae n. subfam. Ne-matology 16, 963-978.
Krebs, C.J. (1989). Ecological methodology. New York, NY,USA, Harper Collins.
Maddison, W. & Maddison, D. (2011). Mesquite: a modu-lar system for evolutionary analysis. Version 2.75. http://mesquiteproject.org.
Maraun, M., Schatz, H. & Scheu, S. (2007). Awesome or ordi-nary? Global diversity patterns of oribatid mites. Ecography30, 209-216.
Múnera Uribe, G.E., Bert, W., Vierstraete, A.R., De La Pena, E.,Moens, M. & Decraemer, W. (2010). Burrowing nematodesfrom Colombia and their relationship with Radopholus similispopulations, R. arabocoffeae and R. duriophilus. Nematology12, 619-629.
Myers, N., Mittermeier, R.A., Mittermeier, C.G., da Fonseca,G.A.B. & Kent, J. (2000). Biodiversity hotspots for conser-vation priorities. Nature 403, 853-858.
Nadler, S.A., De Ley, P., Mundo-Ocampo, M., Smythe, A.B.,Stock, S.P., Bumbarger, D., Adams, B.J., Tandingan De Ley,I., Holovachov, O. & Baldwin, J.G. (2006). Phylogeny ofCephalobina (Nematoda): molecular evidence for recurrentevolution of probolae and incongruence with traditionalclassifications. Molecular Phylogenetics and Evolution 40,696-711.
Neher, D.A. (1999). Nematode communities in organicallyand conventionally managed agricultural soils. Journal ofNematology 31, 142-154.
Pendry, C.A. & Proctor, J. (1997). Altitudinal zonation of rainforest on Bukit Belalong, Brunei: soils, forest structure andfloristics. Journal of Tropical Ecology 13, 221-241.
Porazinska, D.L., Giblin-Davis, R.M., Esquivel, A., Powers,T.O., Sung, W. & Thomas, W.K. (2010). Ecometageneticsconfirm high tropical rainforest nematode diversity. Molec-ular Ecology 19, 5521-5530.
Porazinska, D.L., Giblin-Davis, R.M., Powers, T.O. & Thomas,W.K. (2012). Nematode spatial and ecological patterns fromtropical and temperate rainforests. PloS ONE 7, e44641.
Powers, T.O., Neher, D.A., Mullin, P., Esquivel, A., Giblin-Davis, R.M., Kanzaki, N., Stock, S.P., Mora, M.M. &Uribe-Lorio, L. (2009). Tropical nematode diversity: verticalstratification of nematode communities in a Costa Ricanhumid lowland rainforest. Molecular Ecology 18, 985-996.
Pradhan, G. & Dash, M. (1987). Distribution and populationdynamics of soil nematodes in a tropical forest ecosystemfrom Sambalpur, India. Proceedings of the Indian Academyof Sciences (Animal Sciences) 96, 395-402.
Procter, D.L.C. (1984). Towards a biogeography of free-livingsoil nematodes. I. Changing species richness, diversity anddensities with changing latitude. Journal of Biogeography 11,103-117.
Quisado, J. (2013). Nematode diversity of phytotelmata of Ne-penthes spp. in Mount Hamiguitan Range Wildlife Sanctuary,Philippines. M.Sc. Thesis, University of Ghent, Belgium.
Rahbek, C. (1997). The relationship among area, elevation, andregional species richness in neotropical birds. The AmericanNaturalist 149, 875-902.
Sanders, H.L. (1968). Marine benthic diversity: a comparativestudy. American Naturalist 102, 243-282.
Sanwal, K.C. (1971). Geraldius n. gen., Macrolaiminae n.subfam., with a revision of the subfamilies and genera ofChambersiellidae (Nematoda). Canadian Journal of Zoology49, 965-967.
Vol. 17(3), 2015 343

X. Qing et al.
Seinhorst, J.W. (1962). On the killing, fixation and transferringto glycerin of nematodes. Nematologica 8, 29-32.
Shepherd, U.L. (1998). A comparison of species diversityand morphological diversity across the North Americanlatitudinal gradient. Journal of Biogeography 25, 19-29.
Siddiqi, M.R. (1993). Nematodes of tropical rainforests: 2. Fivenew genera and eight new species of cephalobs. Afro-AsianJournal of Nematology 3, 212-225.
Siemann, E., Tillman, D. & Haarstad, J. (1996). Insect speciesdiversity, abundance and body size relationships. Nature 380,704-706.
Sohlenius, B. & Sandor, A. (1987). Vertical distribution ofnematodes in arable soil under grass (Festuca pratensis) andbarley (Hordeum distichum). Biology and Fertility of Soils 3,19-25.
Steel, H., Moens, T., Scholaert, A., Boshoff, M., Houthoofd, W.& Bert, W. (2011). Mononchoides composticola n. sp. (Nema-toda: Diplogastridae) associated with composting processes:morphological, molecular and autecological characterisation.Nematology 13, 347-363.
Steinberger, Y., Liang, W., Savinka, E., Meshi, T. & Barness,G. (2001). Nematode community composition and diversityassociated with a topoclimatic transect in a rain shadowdesert. European Journal of Soil Biology 37, 315-320.
Steiner, G. (1936). Opuscula miscellanea nematologica, III.Proceedings of the Helminthological Society of Washington3, 16-22.
Steiner, G. (1938). Opuscula miscellanea nematologica, VII.Proceedings of the Helminthological Society of Washington5, 35-40.
Stevens, G.C. (1992). The elevational gradient in altitudinalrange: an extension of Rapoport’s latitudinal rule to altitude.American Naturalist 140, 893-911.
Thorne, G. (1937). Revision of the nematode family Cephalo-bidae Chitwood & Chitwood, 1934. Proceedings of theHelminthological Society of Washington 4, 1-16.
Treonis, A., Sutton, K., Kavanaugh, B., Narla, A., McLlarky,T., Felder, J., O’Leary, C., Riley, M., Pikus, A. & Thomas,S. (2012). Soil nematodes and their prokaryotic prey alongan elevation gradient in the Mojave Desert (Death ValleyNational Park, California, USA). Diversity 4, 363-374.
Weaver, T. & Smolik, J. (1987). Soil nematodes of northernRocky Mountain ecosystems: genera and biomasses. GreatBasin Naturalist 47, 473-479.
Wolf, J.H.D. (1994). Factors controlling the distribution ofvascular and non-vascular epiphytes in the northern Andes.Vegetation 112, 15-28.
Yeates, G.W. (1973). Abundance and distribution of soil nema-todes in samples from the New Hebrides. New Zealand Jour-nal of Science 16, 727-736.
Yeates, G.W. (1974). Studies on a climosequence of soils intussock grasslands. 2. Nematodes. New Zealand Journal ofZoology 1, 171-177.
Yeates, G.W., Bongers, T., de Goede, R.G.M., Freckman, D.W.& Georgieva, S.S. (1993). Feeding habits in soil nematodefamilies and genera-an outline for soil ecologists. Journal ofNematology 25, 315-331.
Yoder, M., Tandingan De Ley, I., King, I.W., Mundo-Ocampo,M., Mann, J., Blaxter, M., Poiras, L. & De Ley, P. (2006).DESS: a versatile solution for preserving morphology andextractable DNA of nematodes. Nematology 8, 367-376.
Zhang, M., Liang, W.-J. & Zhang, X.-K. (2012). Soil nematodeabundance and diversity in different forest types at ChangbaiMountain, China. Zoological Studies 51, 619-626.
344 Nematology




















