Sistemas celulares de degradación. - U-Cursos
20
Sistemas celulares de degradación. Dr. Alejandro Roth (Biología Celular, 2011) 2 http://www.tmd.ac.jp/med/phy2/E_senmon.html ¿Por qué es importante degradar proteínas? • Reciclaje de proteínas viejas o dañadas. • Control de la función de una proteína o la vía regulada por ella. • Prevención de acumulación de proteínas mal construídas. La síntesis protéica es un proceso metabolicamente “caro”, donde buena parte del costo proviene de los sistemas de control de calidad. Las células presentan dos vías de degradación de proteínas: • Autofágia (degradación “en bulto”) • Proteosoma (selectiva)
-
Upload
khangminh22 -
Category
Documents
-
view
2 -
download
0
Transcript of Sistemas celulares de degradación. - U-Cursos

Sistemas celulares de degradación.
Dr. Alejandro Roth (Biología Celular, 2011)
2http://www.tmd.ac.jp/med/phy2/E_senmon.html
¿Por qué es importante degradar proteínas?
• Reciclaje de proteínas viejas o dañadas.
• Control de la función de una proteína o la vía regulada por ella.
• Prevención de acumulación de proteínas mal construídas.
La síntesis protéica es un proceso metabolicamente “caro”, donde buena parte del costo proviene de los sistemas de control de calidad.
Las células presentan dos vías de degradación de proteínas: • Autofágia (degradación “en bulto”)• Proteosoma (selectiva)

Organelos yProteínas de vida larga
Síntesis proteica
Degradación de proteínasdel Retículo
EndoplasmáticoProteínas anormales
Daño
Adaptación Metabólica
Diferenciación celular
reciclaje
Sistema de Ubiquitinación
Degradación Citosólica
Lisosoma(vacuola)
Degradación Lisosomal • Nutrientes extra-celulares.• Degradación en bulto
Proteínas vida corta
• Organelos con una sola membrana y con forma heterogénea
• Son el principal sistema de degradación de las células:
– Nutrientes incorporados por endocitosis– Componentes celulares para ser reciclados
• pH ácido generado mediante bomba de protones de tipo V (vacuolar) homóloga a F0F1.
• 40 enzimas Hidrolasas: Hidrolasas ácidas (proteasas, lipasas, nucleasas, glicosidasas y sulfatasas)
• La actividad de estas enzimas a pH 7.2 es limitado. ¿Sistema de seguridad?
• Formadas a partir de la maduración del endosoma tardío
• Proteínas residentes tienen señal de destinamiento dependiente de glicosilación.
Lisosomas y Vacuolas
0.2 a 0.5 um
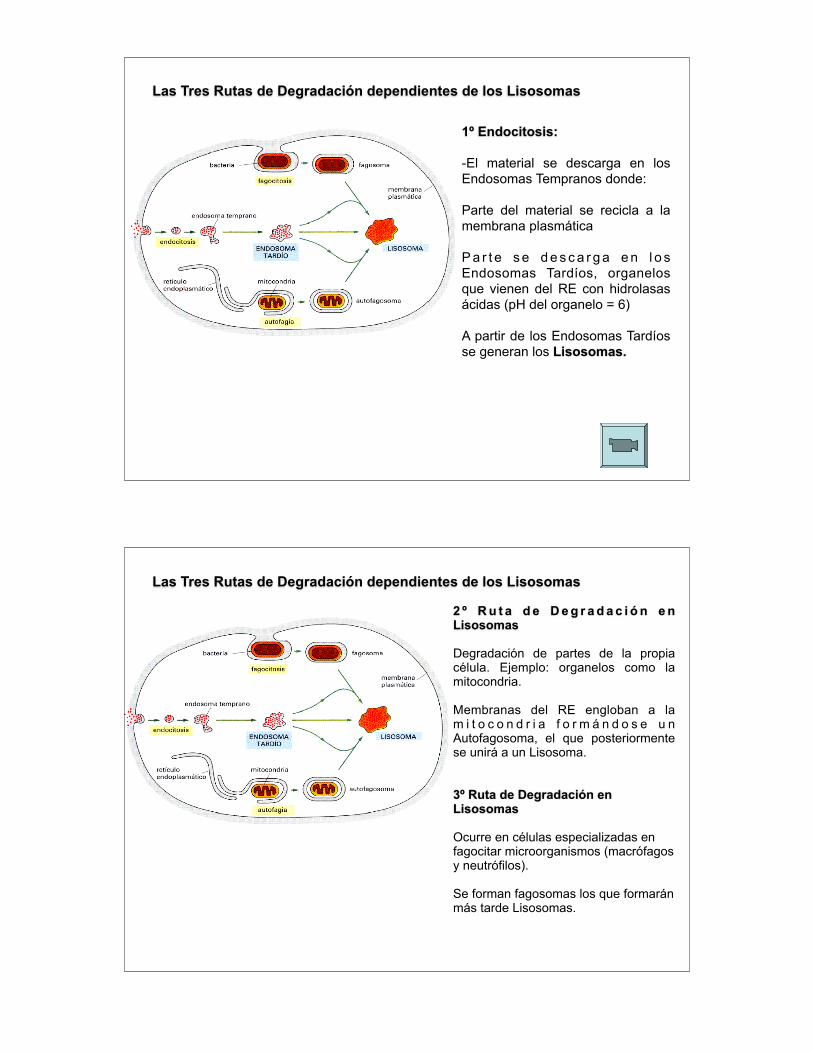
Las Tres Rutas de Degradación dependientes de los Lisosomas
1º Endocitosis:
-El material se descarga en los Endosomas Tempranos donde:
Parte del material se recicla a la membrana plasmática
P a r t e s e d e s c a r g a e n l o s Endosomas Tardíos, organelos que vienen del RE con hidrolasas ácidas (pH del organelo = 6)
A partir de los Endosomas Tardíos se generan los Lisosomas.
2 º R u t a d e D e g r a d a c i ó n e n Lisosomas
Degradación de partes de la propia célula. Ejemplo: organelos como la mitocondria.
Membranas del RE engloban a la m i t o c o n d r i a f o r m á n d o s e u n Autofagosoma, el que posteriormente se unirá a un Lisosoma.
3º Ruta de Degradación en Lisosomas
Ocurre en células especializadas en fagocitar microorganismos (macrófagos y neutrófilos).
Se forman fagosomas los que formarán más tarde Lisosomas.
Las Tres Rutas de Degradación dependientes de los Lisosomas

Ruta Biosintética secretora: proteínas van desde RE a membrana plasmática, o mediante endosomas tardíos a los lisosomas.
Ruta Endocítica, las moléculas son ingeridas y trasladadas a endosomas tempranos y vía endosomas tardíos van a los lisosomas
Rutas de Recuperación (de retorno)
Los lisosomas son producidos mediante maduración de los endosomas
Transporte desde la Red Trans Golgi hacia los Lisosomas
Todas las proteínas llegan hasta la zona trans para ser destinadas (con excepción de las proteínas residentes permanentes del A. de Golgi)
ADN (Núcleo) en azul (Hoechst). Mitocondrias en verde (MitoTracker) Lisosomas, en rojo (LysoTracker).

Formación de Lisosomas:
Maduración
Más que “formar” un lisosoma, las células “maduran” los endosomas tardíos. pH: 5
H+ATP ADP + Pi
pH: 5Aminoácidos
azúcares
nucleótidos
Autofagia• “Auto-canibalismo celular”
– Proceso mediante el que membranas internas de la célula realizan una captura de elementos citoplasmáticos (organelos, acúmulos de proteínas, membranas, etc..) dentro de una vesícula de doble membrana, denominada autofagosoma, la cual es destinada a lisosomas, donde el contenido es degradado.
• Mecanismo de las células para:– Reciclaje de organelos dañados o superfluos– Requerimientos energéticos (“hambruna”)– Evitar la acumulación de agregados proteicos
• Presente en células de plantas, hongos y animales.• Altamente regulado por una cascada de señales fosforilativas
Control Tamoxifen
Células de cáncer de mama, tratadas con Tamoxifen (b) presentan fagosomas (flechas)

• Inducción:• Formación de autofagosoma• Unión a lisosoma• Degradación del contenido y
eliminación del autofagosoma.• Enfermedades asociadas a
autofagia.• Autofagia en: cancer vs apoptosis
Autofagia: etapas
Lumenautofagosoma
No hay consenso
• Inducción: Autofagia: etapas
S6SubU
ribosomal
PP76S6K (quinasa)
mTor
(-)Autofagia:
• No dilucidado 100%
• mTor actúa como un sensor de los niveles de ATP (¿qué procesos fallan en caso de falta de ATP?), nutrientes y estado Redox celular.
• Si existe suficiente ATP, se inhibe la autofagia.
mTor: mammalian target of rapamycin
ATP
nutrientes
[H+]

• Inducción: Autofagia: etapas
S6ribosomal
P
(+)
P76S6K (quinasa)
mTOR
Autofagia
•Disminución en [nutrientes]>> mTOR (FRAP) controla la síntesis de ribosomas y los niveles de crecimiento.
Rampamicina
(-)(-)
ATP
P
Klionsky et al.(2000) Science 290: 1717
Rampamicina: agente anticancerígeno utilizado contra tumores sólidos.Aumento de masa tumoral depende del reclutamiento de mitógenos y nutrientes.
mTor: mammalian target of rapamycin
Autofagia: Resumen
Levine (2007) Nature 446: 745

Fig. 5 In vivo analysis of autophagy using GFP-LC3 transgenic mice(Upper) Gastrocnemius muscle samples were prepared from GFP-LC3 transgenic mice before (left) or after 24-h starvation (right). Small dots represent autophagosomes.(Lower) Embryonic fibroblasts from GFP-LC3 mice were cultured in Hanks' solution for 2 hours. Bar, 10 um.
No existe claridad de la función homeoestatica de la Autofagia.
El desarrollo de animales transgénicos (GFP-LC3, donde LC3 es un marcador de la membrana autofágica) ha permitido detectar los eventos autofágicos a los que se someten diversos tejidos durante el desarrollo (particlarmente postembrionario)
Laboratorio de Noboru Mizushimahttp://www.tmd.ac.jp/med/phy2/english/index.html
16
La AUTOFAGIA es una respuesta a la falta de nutrientes, sin embargo, también permite a las células destruir proteínas anormales que se acumulan (por ejemplo en el SNC). Al bloquear la autofagia se acumulan proteínas mal formadas y se lleva a neurodegeneración. Laboratorio de Noboru Mizushima
http://www.tmd.ac.jp/med/phy2/english/index.htmlNature 441, 885-889 (2006).

Figure 6-95a,b Molecular Biology of the Cell (© Garland Science 2008)
¿destruir proteínas anormales?PROTEINOPATÍAS:Enfermedad de Alzheimer (y múltiples amiloidosis), Priones, Enfermedad de Huntington, Enfermedad de Parkinson, Cataratas, Fibrosis Quística, Anemia Falsiforme, etc...
Enfermedad de Alzheimer: Daño Cortical
La enfermedad de Alhzeimer se caracteriza por la pérdida de tamaño de las cortezas cerebrales y el aumento del volumen de los ventrículos.
Esta atrofia cerebral es en parte producida por la muerte neuronal y la pérdida de conexiones entre las neuronas.
Esta enfermedad se considerada una proteínopatía, una enfermedad producida por proteínas anormales, las cuales forman acúmulos tóxicos para las neuronas:
• Las Placas Amiloides (péptido ABeta)
• Ovillos Neurofibrilares (Prot. Tau)
http://www.exploralasalud.com/tag/alzheimer/

Basic Neurochemistry, Agranoff et al. 1998
Lesiones histopatológicas de la EA. Tinción de plata de Bielschowsky de una sección de la amígdala de un hombre de 69 años con historia de EA (6 años). Placa senil o Placa Amieloide en el centro (A), rodeada de ovillos neurofibrilares oscuros (cuerpos oscuros, compara con flechas) y un halo de neurtas distróficas (cabeza de flecha)
Imágenes: National Institute on Aging. NIH. USA
Placa Amieloide
Neuronas en degeneración
20

Imágenes: National Institute on Aging. NIH. USA
1.La proteína APP (ABeta precursor protein) se encuentra en la superficie de las células del Cerebro.
Imágenes: National Institute on Aging. NIH. USA
2.La APP es cortada por α-secretasa, cortando el Abeta en mitad y generando un fragmento soluble que promueve la sobrevivencia neuronal
4.La γ-secretasa libera el Abeta de la membrana, dejándolo disponible para formar acúmulos
3. El corte por una β-secretasa deja el fragmento ABeta completo.
sAPPβ
γ-secretasa
β-secretasa
sAPPβAβ
γ-secretasa
β-secretasa β-secretasa
APP
γ-secretasa α-secretasa
sAPPα
γ-secretasa α-secretasa

Imágenes: National Institute on Aging. NIH. USA
Imágenes: National Institute on Aging. NIH. USA
Neurona Muriendo
Ovillos Neurofibrilares

Figure 6-85 Molecular Biology of the Cell (© Garland Science 2008)
Durante la síntesis de proteínas se exponen dominio hidrofóbicos y señales de degradación. Existe una competencia entre las proteínas que ayudan a encontrar el punto correcto y las que intentan degradar las proteínas.
Origen de proteínas anormales
26Figure 6-88 Molecular Biology of the Cell (© Garland Science 2008)

• La función de cada proteína dependen de que adquieran su estrucutra tridimensional adecuada.
– La hidrofobia de los aa apolares determina la formación de los dominios internos de las proteínas. Sin embargo, pueden asociarse de manera equivocada en la medida que la proteína es sintetizada.
– Las proteínas Heat shock son de las más abundantes en las células. Su expresión es inducida en respuesta a estrés celular (por ejemplo, por aumento de temperatura). Corresponden a 1–2% de la proteína total en células en condiciones normales
Chaperonas
Durante la traducción de proteínas citosólicas, las chaperonas se unen al amino (N) terminal del polipéptido creciente, estabilizándolo en una configuración no plegada hasta que la síntesis se ha completado. Completada la síntesis, la proteína se libera del ribosoma y adquiere su correcta conformación tridimensional.
ADP+Pi
ADP+Pi
Hsp70 o Hsp90
El complejo de Hsp60 genera un espacio con dominios de unión hidrofóbicos que ayudan a las proteínas mal formadas a ser re-estrucutradas. Actúa post-traduccionalmente
• Hsp100: Desestructurasas. Obligan a las proteínas mal formadas a pasar por un poro de 20 A. Pueden estar asociadas a serina proteasas y degradar automáticamente
Chaperonas•Hsp60/TCP-1 o GroESbacterias: Chaperonina (14 SubU. 1000 KDa)

Vía de Ubiquitinación y destinamiento a Proteosomas
Ubiquitina: Ubicua
Los sistemas proteolíticos intracelulares reconocen y destruyen proteínas dañadas, mal ensambladas y proteínas regulatorias y/o de vida corta (por ejemplo, las ciclinas, receptores, sistemas de presentación de antígenos).
Síntesis proteica
Proteínas anormales
Proteínas vida corta
Sistema de Ubiquitinación
Cascada de ubiquitinación Se genera una cascada de ubiquitinaciones múltiples:
unión covalente de ubiquitina (76aa) a cadenas de lisina de proteínas blanco.
1. Activación: E1 + Ubiquitina + ATP
E1ADP+Pi
E3

Cascada de ubiquitinación Se genera una cascada de ubiquitinaciones múltiples:
unión covalente de ubiquitina (76aa) a cadenas de lisina de proteínas blanco.
1. Activación: E1 + Ubiquitina + ATP
2. Transferencia: E1 transfiere Ubiquitina activada a complejo E2-E3 (ubiquitina ligasa)E1
ADP+Pi
E3
Cascada de ubiquitinación Se genera una cascada de ubiquitinaciones múltiples:
unión covalente de ubiquitina (76aa) a cadenas de lisina de proteínas blanco.
1. Activación: E1 + Ubiquitina + ATP
3. Reconocimiento de señal de degradación en una proteínas y traspaso de ubiquitina:
Fosforilación (de E3 o sustrato)
Desenmascaramiento (sustrato) Des-estabilización N-terminal (sustrato) Ligando (de E3)
2. Transferencia: E1 transfiere Ubiquitina activada a complejo E2-E3 (ubiquitina ligasa)E1
ADP+Pi
E3

Cascada de ubiquitinación Se genera una cascada de ubiquitinaciones múltiples:
unión covalente de ubiquitina (76aa) a cadenas de lisina de proteínas blanco.
1. Activación: E1 + Ubiquitina + ATP
3. Reconocimiento de señal de degradación en una proteínas y traspaso de ubiquitina:
Fosforilación (de E3 o sustrato)
Desenmascaramiento (sustrato) Des-estabilización N-terminal (sustrato) Ligando (de E3)
2. Transferencia: E1 transfiere Ubiquitina activada a complejo E2-E3 (ubiquitina ligasa)E1
ADP+Pi
E3
4. Repetición del ciclo y elongación de lacadena
De ubiquitinación a proteosoma
Vía de immunoproteosoma
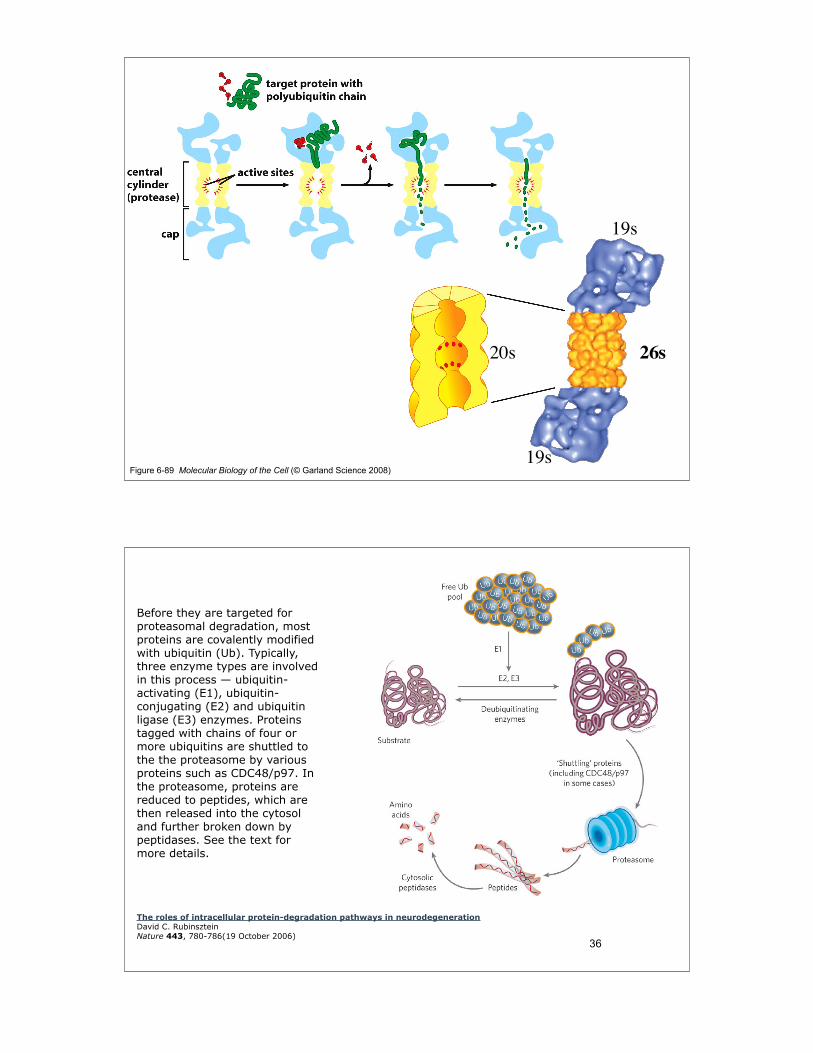
Figure 6-89 Molecular Biology of the Cell (© Garland Science 2008)
20s
19s
19s
26s
36
Before they are targeted for proteasomal degradation, most proteins are covalently modified with ubiquitin (Ub). Typically, three enzyme types are involved in this process — ubiquitin-activating (E1), ubiquitin-conjugating (E2) and ubiquitin ligase (E3) enzymes. Proteins tagged with chains of four or more ubiquitins are shuttled to the the proteasome by various proteins such as CDC48/p97. In the proteasome, proteins are reduced to peptides, which are then released into the cytosol and further broken down by peptidases. See the text for more details.
The roles of intracellular protein-degradation pathways in neurodegenerationDavid C. RubinszteinNature 443, 780-786(19 October 2006)

37
When proteins are accessible to both the ubiquitin (Ub)–proteasome and autophagy pathways, the greater efficiency of the ubiquitin–proteasome system makes it the favoured and dominant clearance route. When a cytosolic protein is aggregate prone and a poor proteasome substrate, then autophagy becomes the main clearance route by default — under these circumstances, the autophagy route becomes more effective than the proteasome.
The roles of intracellular protein-degradation pathways in neurodegenerationDavid C. RubinszteinNature 443, 780-786(19 October 2006)
38
Under normal conditions of substrate synthesis and basal autophagy, the direction of the equilibrium of an aggregate-prone protein is towards aggregate formation (grey arrows). If substrate synthesis is stopped (for example, in a conditional transgenic mouse) or clearance of the soluble/oligomeric forms is enhanced (for instance, by autophagy) then this equilibrium can be reversed and aggregates are indirectly cleared (red arrows).
The roles of intracellular protein-degradation pathways in neurodegenerationDavid C. RubinszteinNature 443, 780-786(19 October 2006)

Tareaopcional (para aprovechar el tiempo y practicar
inglés)
• Podcast:
– This week in Science (www.twis.org)– The Nature PodCast (http://www.nature.com/
podcast/index.html)– The Science Podcast (http://www.sciencemag.org/
multimedia/ )– Cell Podcast (http://www.cellpress.com/misc/page?
page=podcast