
Sex of the foetus determines the time of weaning of the previous offspring of captive plains zebra (...
13
Sex of the foetus determines the time of weaning of the previous offspring of captive plains zebra (Equus burchelli) Jan Pluha ´c ˇek a,b, * , Lude ˇk Bartos ˇ a , Miroslava Dolez ˇalova ´ c , Jitka Bartos ˇova ´-Vı ´chova ´ a a Ethology Group, Research Institute of Animal Production, Pr ˇa ´telstvı ´ 815, 104 01 Praha-Uhr ˇı ´ne ˇves, Czech Republic b Department of Zoology, Faculty of Science, Charles University, Vinic ˇna ´ 7, 128 44 Praha 2, Czech Republic c Dvu ˚r Kra ´love ´ Zoological Gardens, S ˇ tefa ´nikova 1029, 544 01 Dvu ˚r Kra ´love ´ nad Labem, Czech Republic Accepted 29 May 2006 Available online 7 July 2006 Abstract We examined if captive plains zebra (Equus burchelli) mares change the timing of weaning according to the sex of the suckling foal and of the new impending foetus. We observed 19 captive plains zebra foals, in three different formations, at the Dvu ˚r Kra ´love ´ Zoo. The earliest time for weaning of any foal was recorded at the age of only 243 days. The latest suckling that was ever recorded occurred 83 days before the dam’s next delivery, when the foal was at the age of 355 days. We found that pregnant mares weaned their foals sooner than the non-pregnant mares. Survival analysis revealed that the sex of the foetus was the most influential factor in determining the weaning age of the current foal. Hence, if the foetus was a male, then weaning occurred sooner than if the dam was carrying a female foetus. This finding is the first of this kind among ungulates. The reproductive state of the mare and the sex of her foetus should be considered when interpreting weaning times of equids. # 2006 Elsevier B.V. All rights reserved. Keywords: Equus burchelli; Weaning time; Zebra; Maternal investment; Trivers–Willard model; Zoo www.elsevier.com/locate/applanim Applied Animal Behaviour Science 105 (2007) 192–204 * Corresponding author. Tel.: +420 267 009 765; fax: +420 267 710 797. E-mail address: [email protected] (J. Pluha ´c ˇek). 0168-1591/$ – see front matter # 2006 Elsevier B.V. All rights reserved. doi:10.1016/j.applanim.2006.05.019
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Sex of the foetus determines the time of weaning of the previous offspring of captive plains zebra (...

Sex of the foetus determines the time of weaning
of the previous offspring of captive plains
zebra (Equus burchelli)
Jan Pluhacek a,b,*, Ludek Bartos a, Miroslava Dolezalova c,Jitka Bartosova-Vıchova a
a Ethology Group, Research Institute of Animal Production, Pratelstvı 815,
104 01 Praha-Uhrıneves, Czech Republicb Department of Zoology, Faculty of Science, Charles University, Vinicna 7,
128 44 Praha 2, Czech Republicc Dvur Kralove Zoological Gardens, Stefanikova 1029, 544 01 Dvur Kralove nad Labem,
Czech Republic
Accepted 29 May 2006
Available online 7 July 2006
Abstract
We examined if captive plains zebra (Equus burchelli) mares change the timing of weaning according to
the sex of the suckling foal and of the new impending foetus. We observed 19 captive plains zebra foals, in
three different formations, at the Dvur Kralove Zoo. The earliest time for weaning of any foal was recorded
at the age of only 243 days. The latest suckling that was ever recorded occurred 83 days before the dam’s
next delivery, when the foal was at the age of 355 days. We found that pregnant mares weaned their foals
sooner than the non-pregnant mares. Survival analysis revealed that the sex of the foetus was the most
influential factor in determining the weaning age of the current foal. Hence, if the foetus was a male, then
weaning occurred sooner than if the dam was carrying a female foetus. This finding is the first of this kind
among ungulates. The reproductive state of the mare and the sex of her foetus should be considered when
interpreting weaning times of equids.
# 2006 Elsevier B.V. All rights reserved.
Keywords: Equus burchelli; Weaning time; Zebra; Maternal investment; Trivers–Willard model; Zoo
www.elsevier.com/locate/applanim
Applied Animal Behaviour Science 105 (2007) 192–204
* Corresponding author. Tel.: +420 267 009 765; fax: +420 267 710 797.
E-mail address: [email protected] (J. Pluhacek).
0168-1591/$ – see front matter # 2006 Elsevier B.V. All rights reserved.
doi:10.1016/j.applanim.2006.05.019

1. Introduction
Subsequent pregnancy is one of the main factors that can affect the length of lactation of
mammalian species where the lactating female conceives shortly after birth (Bateson, 1994).
Under most environmental conditions, the optimum weaning time should occur earlier for
pregnant than for non-pregnant females (Bateson, 1994). This has been documented for various
ungulates, such as Saharan arrui (Ammotragus lervia; Cassinello, 1997) and American bison
(Bison bison; Green et al., 1993). In African elephants (Loxodonta africana), the duration of
maternal investment via nursing appeared to be controlled by the inter-birth interval of the
mother (Lee and Moss, 1986).
Differences in time spent suckling between young males and females have been reported for
many sexually dimorphic ungulates, such as red deer (Cervus elaphus; Clutton-Brock et al.,
1981), Saharan arrui (Cassinello, 1996), American bison (Wolff, 1988, but see Green and Berger,
1990), and bighorn sheep (Ovis canadensis; Festa-Bianchet, 1988). Only two reports have
suggested that feral horse (sexually monomorphic species) male foals spent a longer time
suckling than female foals during the first 8 weeks of their life (Duncan et al., 1984; Berger,
1986). On the other hand, however, no sex differences in suckling behaviour have been reported
for other horse populations (Crowell-Davis, 1985; Smith-Funk and Crowell-Davis, 1992; Wolff
and Hausberger, 1994; Cameron et al., 1999b) and sexually monomorphic guanacos (Lama
guanicoe; Sarno and Franklin, 1999) or for many sexually dimorphic ungulates, such as fallow
deer (Dama dama; Gauthier and Barrette, 1985), white-tailed deer (Odocoileus virginianus;
Gauthier and Barrette, 1985), reindeer (Rangifer tarandus; Lavigueur and Barrette, 1992), and
Saharan arrui (Cassinello, 2001). It should be noted, however, that suckling behaviour is not
always a useful predictor of milk intake (Cameron, 1998; Cameron et al., 1999b).
It has previously been found that the sex ratio of the offspring of captive equids did not differ
significantly from 1:1 (Mlıkovsky, 1988). To date, in four equid species, a few different ways of
sex-biased maternal care has been documented. Firstly, relationships between high maternal rank
and male biased offspring production, and probability of conception and inter-birth intervals
were observed in captive plains zebra (Equus burchelli; Schilder and Boer, 1987; Pluhacek et al.,
2006). Secondly, in contrast to the previous findings, dominant mountain zebra (E. zebra)
mothers produced more daughters than sons (Lloyd and Rasa, 1989). Thirdly, female-biased sex
ratios following a poor year and after the dam had produced a male in the preceding year (Monard
et al., 1997) and male-biased sex ratios for offspring in mares in better condition was observed in
feral horses (E. caballus; Cameron et al., 1999a). Finally, a male-biased sex ratios in offspring
was observed in nonprimiparous and young Asiatic wild asses (E. hemionus; Saltz and
Rubenstein, 1995; Saltz, 2001).
The plains zebra is a polygynous species in which there is relatively high competition among
males compared to females. The gestation period of plains zebra ranges from 361 to 385 days
(Wackernagel, 1965; Klingel, 1969a; Lobanov, 1983). The first fertile oestrus typically occurs
within 7–9 days post-partum (Wackernagel, 1965; Klingel, 1969a; Smuts, 1976b). Conception
rates ranging between 50 and 79% per season have been reported (Klingel, 1969a; Smuts,
1976b). At least half of the zebra mares conceive when they are still lactating. Thus, mares
frequently have to invest in two offspring at one time. It was suggested by Keiper (1979) that
there is an increased probability for feral horse mares to abort when nursing a foal in comparison
with pregnant mares that have no dependent offspring.
Under natural conditions, a plains zebra herd contains just one stallion (Klingel, 1969a,
1969b) who is always dominant over the entire herd (Klingel, 1967; Schilder and Boer, 1987;
J. Pluhacek et al. / Applied Animal Behaviour Science 105 (2007) 192–204 193

Andersen, 1992). His reproductive potential is limited only to the time he spends with the herd
(for an exception see Rubenstein, 1986), while females can reproduce throughout most of their
lifetime (Klingel, 1969a). Plains zebra foals frequently suffer from predation (in wild; Smuts,
1976a) and infanticide (in captivity; Pluhacek and Bartos, 2000, 2005). Stallion keep the herd as a
group by active herding (Andersen, 1992), which can disturb suckling behaviour. Mares that live
in herds without stallions can change their behaviour in other unexpected ways. For example, in
horses, the adult mares allogroomed more in groups without stallion than in groups with a stallion
(Sigurjonsdottir et al., 2003). Therefore, we also consider the presence of the stallion within the
herd as a possible factor influencing weaning time.
Various reports in equids have focused on the age at weaning of the foal (King, 1965; Zwolinski
and Siudzinski, 1966; Tyler, 1972; Rashek, 1976; Smuts, 1976b; Martin-Rosset et al., 1978;
Penzhorn, 1979; Duncan et al., 1984; Berger, 1986; Oom and Reis, 1986; Becker and Ginsberg,
1990). Only very few of these researchers, however, analysed any other factor that could influence
the weaning age. In horses, Asiatic wild asses and mountain zebra, pregnant mares nurse their foals
for a shorter period than the non-pregnant dams (Rashek, 1976; Duncan et al., 1984; Penzhorn,
1984; Berger, 1986; Cameron et al., 2000). However, older (over 9 years of age) horse mothers did
not significantly differ in foal weaning age in relation to their reproductive condition (Cameron
et al., 2000). These mares generally weaned their foals before 1 year of age, regardless of whether or
not they were pregnant, whereas younger mothers continued to suckle their yearling offspring if
they did not have a subsequent pregnancy in the next breeding season (Cameron et al., 2000). It has
been reported that the duration of lactation is longer for primiparous than for multiparous mares
(Duncan et al., 1984). Furthermore, lactation lasted for equal lengths of time for both male and
female foals (Duncan et al., 1984; Berger, 1986; Cameron and Linklater, 2000) and weaning dates
did not vary in relation to the condition of the mare (Berger, 1986; Cameron and Linklater, 2000).
We are aware of only one study dealing with interspecific differences in weaning time among
equids that has been published (Becker and Ginsberg, 1990) and, this study did not reflect any
other factors such as pregnancy or parity of the mare, which were previously documented as
important (Rashek, 1976; Duncan et al., 1984; Berger, 1986).
Investigation of the factors affecting pregnancy and weaning provides an opportunity to test
the Trivers–Willard model (TWM; Trivers and Willard, 1973). The TWM predicts that mothers
in polygynous species, that are in good condition, would be advantaged by investing more in
sons, whereas mothers in poor condition would be advantaged by investing more in daughters.
There are two ways to accomplish this biased investment, either by allocating resources
preferentially to one sex or by adaptive adjustment of the sex ratio at conception (Cassinello,
1996). Ungulates are a useful group of mammals for evaluating different theories of offspring sex
ratio variation, because they (1) give birth to quite few young, (2) parental investment for an
individual varies, and (3) parents may provision in sons and daughters differentially (Kojola,
1999). In ungulates, the second way has been tested more frequently in the past (Rutberg, 1986;
Hewison and Gaillard, 1999; Kruuk et al., 1999; Lindstrom et al., 2002). However, ungulates
typically have a long duration of maternal investment (gestation and lactation; Saltz, 2001) and
the TWM should only be applied when three assumptions are met: (1) the condition of the young
at the end of maternal investment will tend to be correlated with the condition of the mother
during her investment; (2) differences in young at the end of the period of maternal investment
will tend to endure into adulthood; (3) adult males will be differentially helped in reproductive
success compared with adult females by slight advantageous in condition. The TWM was
successfully used in studies of feral horse populations (Cameron et al., 1999a; Cameron and
Linklater, 2000) and we believe that the plains zebra also fit all three of these assumptions.
J. Pluhacek et al. / Applied Animal Behaviour Science 105 (2007) 192–204194

We tested the following four hypotheses: (1) pregnant plains zebra mares would wean sooner
than non-pregnant ones, (2) the age of foal at weaning would increase with increasing time to
delivery of the next foal. We assumed that the captive zebra mares in this study enjoyed under
good conditions (due to management, feeding, lack of predation, etc.) and they should invest
more into sons than into daughters due to high future competition among sons. Based on the
TWM and on our assumption concerning the condition we further predicted that, (3) if the mare
was pregnant and the foetus was a male, she should wean her recent offspring sooner than when
the foetus was a female, and (4) male foals should be weaned later than female foals due to
increased investment in male offspring due to their greater competitive potential.
As potential predictors, we also tested other factors, which might affect natural weaning in
captive plains zebra, such as mortality of a new offspring, the presence of a stallion, the age of the
parents, and the experience of mother (i.e. the number of previous foals successfully reared per
mother), etc.
We are aware of three reports to date on the weaning time of plains zebra (King, 1965; Smuts,
1976b; Becker and Ginsberg, 1990) and all these reports focused on from free ranging
populations. Nevertheless, these reports substantially differed from each other. While King
(1965) reported weaning to occur at 7 months (25 weeks) of age in East Africa, Smuts (1976b)
observed that weaning occurred between 9 and 16 (‘‘normally 11’’) months in South Africa. In
the East Africa, Becker and Ginsberg (1990) reported that weaning occurred at a mean age of
15.5 (�2.2) months (n = 7). Kingdon (1997) more recently summarized that ‘‘foals suck milk for
up to 6 months’’. However, no factor influencing weaning time was considered in any of these
reports of free ranging zebra populations. The knowledge from natural maternal weaning
behaviour gives important information about the investment of a mare in her offspring. Hence,
this biological information is deemed necessary in testing for some sociobiological hypotheses
(Ebensperger, 1998; Pluhacek and Bartos, 2000, 2005).
2. Animals, materials and methods
2.1. Animals
We observed 19 foals of 11 individual plains zebra mares, split into three different herds, at the Dvur
Kralove Zoo, Czech Republic. In three cases, the mares were not pregnant and all their foals (two males and
one female) were weaned artificially (by human intervention).
In all other cases (16) the mares (10 individual) were pregnant at the time of weaning. Ten of the foals
were weaned naturally by their mothers (9 individual mothers). Six of these foals were males and four were
females. It was later found that five of the subsequent foetuses were female and five were male. Three of the
mares delivered stillborn foals; of the stillborns, there was one male and two females.
The six foals of the remaining pregnant mares (five individual) were weaned by human intervention.
These foals were presented in each of the observed herds. Four of these foals were males and two were
females. It was also later found that one of the foetuses was male and the five others were females. Two of
these mares delivered stillborn foals, which were both females.
There was at least one female foal in each herd and one female foetus in each herd. The herd sizes ranged
between two and six breeding mares, aged from 4 to 20 years. All of the observed adult females were
multiparous. The size of the enclosures for the three herds were 800, 1350 and 1750 m2, respectively. There
was almost no vegetation present in any of the enclosures and food was provided to the zebras ad libitum.
They were given fresh food (hay and carrot in winter, grass in summer) daily, usually in the morning. The
criterion used to establish the survival status of the following offspring was set at 6 months of age. In one
herd the stallion died, in two other herds he was separated from the herd for several months and after he was
returned to its ‘‘own’’ herd for management purposes. There was at least one case of absence and presence of
J. Pluhacek et al. / Applied Animal Behaviour Science 105 (2007) 192–204 195

the stallion in each herd at the time of weaning, giving us a chance to assess the possible role of the stallion in
the weaning process. All of the stallions involved were fathers of the observed foals and no case of
aggression of the stallion towards his foals was observed. The dominance hierarchy among the adult females
was determined by biting and offensive kicking as per Pluhacek et al. (2006). Based on dyadic interactions
between mares, for each mare during each period, we estimated rank by a simple criterion of the number of
mares who dominated her (further referred to as ‘‘number of dominant mares’’).
Each group was observed at least once a week (on Saturday or Sunday) from January 1999 to January
2000 and from September 2001 to March 2002. We performed the observations in four different sessions
each week. Each session lasted for 180 min, either from 0800 to 1100 h or from 1400 to 1700 h on Saturday
or Sunday. We observed only one herd during each session (i.e. on Saturday morning we observed herd no.
1, in the afternoon herd no. 2, on Sunday morning herd no. 3 and in the afternoon again herd no. 1, the next
Saturday we began with herd no. 2, etc.). Data concerning all suckling events were recorded in each
observation session.
From February 2000 to August 2001, we performed additional observations of weaning. During these
periods we observed the foals only until they started suckling or until 180 min elapsed.
With regard to the many different definitions of natural weaning in mammals (Martin, 1984; Counsilman
and Lim, 1985; Green et al., 1993) we used the term natural weaning to indicate the cessation of suckling
(Counsilman and Lim, 1985). Thus, if the foal was not observed to be suckling during the 180 min observation
period, then the foal was regarded as weaned. Moreover, we checked for suckling during the next 180 min
period and we asked the zoo keepers to record daily any suckling events when working with the animals. We
defined the day of weaning to be the last day on which a suckling event was seen for each individual foal.
All of the other data concerning the animals (i.e. date of birth of the foals and parents, number of previous
foals of the mother, etc.) were obtained from curators of the Dvur Kralove Zoo. The study was approved by
Institutional Animal Care and Use Committee of the Research Institute of Animal Production and was
conducted in accordance with Czech Central Committee for Protection of Animals number 13803/2003-
1020.
2.2. Statistics
We used Kruskal–Wallis test for determining whether pregnancy influenced weaning time or not in terms
of the duration of lactation. Then we analysed weaning time in pregnant mares, only. The numeric variables
that were analysed are shown in Table 1. The categorical predictors that were tested were the sex of the
weaned foal, the sex of the foetus, the presence or absence of a stallion in the herd at time of weaning and the
survival status of the next offspring. To utilize the information of the incomplete data and to shed light on the
possible interactions among the predictors, we applied a survival analysis using the Cox proportional
hazards model (Cox, 1972) using PROC PHREG (Patteta, 2001). An intrinsic characteristic of survival data
is the possibility for censoring of observations, that is, the actual time until the event is not observed. Such
J. Pluhacek et al. / Applied Animal Behaviour Science 105 (2007) 192–204196
Table 1
The numerical variables used in the model (n = 16 foals weaned by a pregnant mother)
Variable Unit Mean � S.D. Range
Foal’s age Days 313.3 � 62.9 176–461
Mother’s age Years 13.7 � 5.1 5.2–18.3
Time to next parturition Days 162 � 53 83–285
Father’s age Years 16.9 � 2.5 12.2–19.3
Number of dominant mares Animals 1.88 � 1.45 0–4
Herd size Animals 7.8 � 1.8 3–11
Number of other suckling foals Animals 1.7 � 1.1 0–3
Number of previous foals successfully reared per mother Animals 6.0 � 3.5 0–10
Number of previous foals per mother Animals 7.7 � 4.4 1–13
censoring can arise from withdrawal from the experiment or termination of the experiment. Since the
response was duration, some of the possible events may not yet have occurred when the period for data
collection had terminated. The dependent variable was the time which elapsed from the birth of a foal to its
weaning (age at weaning). Weaning occurred either naturally, being induced by the mare (an event weaning
time), or it was induced earlier by humans (a censored weaning time). For the age at weaning, a full model
containing all of the explanatory variables and the first-order interaction terms was initially fitted. Each term
was then sequentially dropped from the full model unless doing so had a significant effect on the fit of the
model. All of the data were analysed using the SAS System, version 8.2.
In a study with horses, Cameron et al. (1999a) found a tendency for female offspring to be conceived
after more cycles than for male offspring. To check this in captive plains zebra we collected data from the
herds studied (in years 1990–2004) and applied multivariate general linear mixed model (GLMM) with
interval duration between introduction of a male into the herd (including non-pregnant mares) and date of
delivery as the dependent variable and sex of the foal as independent variable. To account for the repeated
measures on the same individuals across different years, the analysis was performed using mixed model
analysis with individual mare as a random factor, using PROC MIXED (SAS, version 9.1).
For obvious technical reasons our data set was limited. Therefore, we analysed a statistical test’s power
(the probability that the test procedure will result in statistical significance) using The SAS/STAT power and
sample size (PSS) application comparing two survival curves. The power was related to the sample size, the
size of the type 1 (alpha) error, the actual size of the effect and the size of experimental error.
3. Results
In total, 859 h (during 166 days) of observations of the animals were collected and 3301 suckling
bouts were recorded. In addition, 314 suckling bouts were recorded during the additional
observations (191 days; see Section 2.1) aimed at recording the end of the suckling period.
The earliest weaning that was recorded occurred when a (male) foal was only 243 days old.
His mother gave birth to the next offspring 160 days after. The earliest weaning of the female foal
was recorded when the foal was 284 days old. The latest suckling that was ever recorded occurred
J. Pluhacek et al. / Applied Animal Behaviour Science 105 (2007) 192–204 197
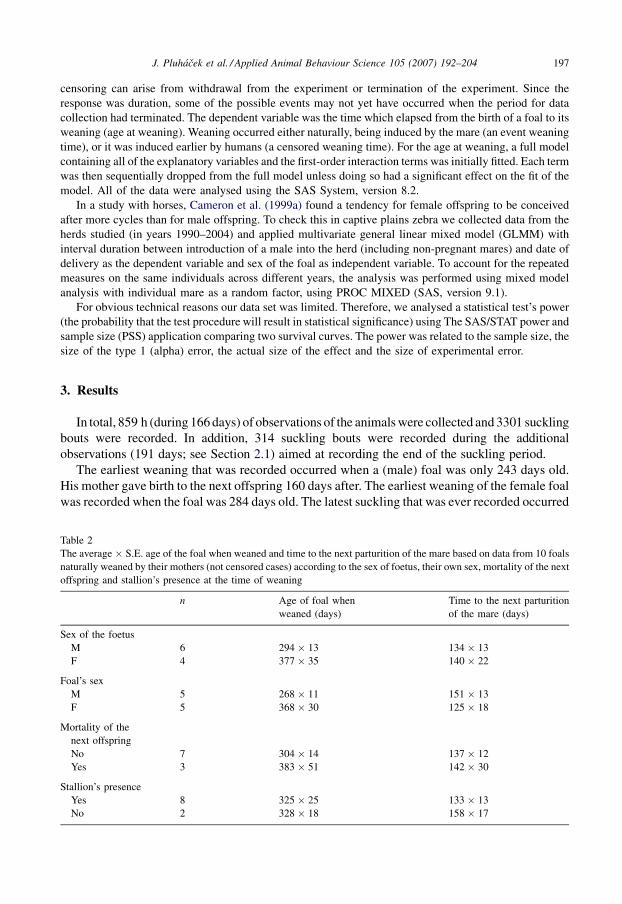
Table 2
The average � S.E. age of the foal when weaned and time to the next parturition of the mare based on data from 10 foals
naturally weaned by their mothers (not censored cases) according to the sex of foetus, their own sex, mortality of the next
offspring and stallion’s presence at the time of weaning
n Age of foal when
weaned (days)
Time to the next parturition
of the mare (days)
Sex of the foetus
M 6 294 � 13 134 � 13
F 4 377 � 35 140 � 22
Foal’s sex
M 5 268 � 11 151 � 13
F 5 368 � 30 125 � 18
Mortality of the
next offspring
No 7 304 � 14 137 � 12
Yes 3 383 � 51 142 � 30
Stallion’s presence
Yes 8 325 � 25 133 � 13
No 2 328 � 18 158 � 17

83 days before the next delivery, when the (female) foal was at the age of 355 days. The average
time of weaning of 10 foals who were weaned naturally is showed in Table 2.
The age of a foal at the natural weaning by pregnant mares was lower (327.4 � 64.0 days,
n = 10) than that at the artificial weaning of non-pregnant ones (444.0 � 26.9 days, n = 3;
d.f. = 1; x2 = 4.48; P < 0.05).
Multifactorial survival analysis was employed and the 10 cases of event weaning time and six
cases of censored weaning time were entered into the model. The final model, containing two
predictors (sex of the foetus and the time to delivery), rejected the null hypothesis that all of the
regression coefficients of the model were equal to 0 (likelihood ratio, d.f. = 2; x2 = 12.43;
P < 0.001). The sex of the foetus appeared to be the most important factor for weaning time
(x2 = 5.48; P = 0.019, hazard ratio = 0.064). The time to delivery almost reached a level of
significance (x2 = 3.31; P = 0.069, hazard ratio = 0.98) and hence we kept it in the model. The
Cox proportional ‘‘hazard ratio’’ is the ‘‘hazard’’ for one group of predictor variable (male
foetus) divided by the ‘‘hazard’’ for another group of predictor variable (female foetus; Patteta,
2001). The estimate is 0.064, implying that the ‘‘hazard function’’ for male foetus is smaller than
the ‘‘hazard function’’ for female foetus. In other words, mares bearing a male foetus would wean
her foal earlier than a mare bearing a female foetus. Fig. 1 shows the pattern of the survivor
function estimate of the age of the foal at weaning (corrected for the time to the next delivery)
according to sex of the mother’s foetus. At the age of about 1 year, the foals of the mares bearing a
male foetus were likely to be weaned over 100 days earlier than those of the mares bearing a
female foetus. Except the time to delivery and the sex of the foetus, none of the factors shown in
Tables 1 and 2 affected weaning time significantly.
Then we calculated the power analysis for two survival curves. For a log-rank test comparing
two survival curves with a two-sided significance level of 0.05, a sample size of 18 per group is
required to obtain a power of at least 0.8 for the two piecewise linear curves, ‘‘Curve 1’’ and
‘‘Curve 2’’. The actual power was 0.820.
J. Pluhacek et al. / Applied Animal Behaviour Science 105 (2007) 192–204198
Fig. 1. Survivor function estimate plotted against the weaning age of the current foal according to the sex of the foetus.

For checking possible difference between the sex of the foals in interval between introduction
of a male into the herd and date of delivery, we collected data on 29 foals delivered by 15 mares.
GLMM revealed no difference in this interval between the sex of the foal (mean � S.E., for males
411.14 � 10.36 days, for females 393.27 � 13.45 days, F = 1.23; d.f. = 1, 28.9; n.s.).
4. Discussion
Pregnant captive plains zebra mares weaned their foals sooner than the non-pregnant ones.
Two individual mares were observed in both situations as pregnant and non-pregnant and one of
them having male foals only and the other mare produced female foals. When pregnant, they
weaned their foals more than 50 days earlier than when they were non-pregnant and all weaning
cases of non-pregnant mares were artificial. Thus, the variation in weaning time between
pregnant and non-pregnant mares is in fact much grater. Moreover, keepers in the zoo reported
observation of several other non-pregnant mares suckling much older foals than those of the
pregnant dams included in our analyses (Bella, personal communication). The significance of
pregnancy in the weaning process of older feral horse mares was doubted in one recent report on
weaning in feral horses (Cameron et al., 2000). The zebra mares included in our analyses were
aged from 5 to 18 years and their age did not influence the timing of weaning in our study. Two of
three of the non-pregnant mares were 14 and 15 years old (classified as old in Cameron et al.,
2000), and all of these mares nursed foals for more than one and a half year.
The time to delivery ranged from 83 to 185 days and is comparable with that in feral horses
(Duncan et al., 1984; Berger, 1986) and wild mountain zebra (Joubert, 1972). We did not record
any indications of prematurity or dysmaturity among the neonates. The power analysis revealed
that the optimal data set required to obtain a power of at least 0.8 for the two piecewise, linear
curves should be twice as much as the one available. This means that time to delivery with
significance of only P = 0.069 is in fact potentially of high importance in weaning in plains zebra,
as reported for horses, asses and mountain zebra (Tyler, 1972; Rashek, 1976; Duncan et al., 1984;
Penzhorn, 1984) and suggested for plains zebra (Smuts, 1976b). This factor has been neglected in
earlier reports on plains zebra (King, 1965; Becker and Ginsberg, 1990).
In the present study, the sex of the foetus appeared to be the most significant factor influencing
weaning time (P = 0.019). Our available data set was only 50% of the optimal number revealed
by the power analysis. This could mean that the sex of the foetus would be a very strong factor
affecting the weaning time of the suckling foal. That is, if the next offspring was to be male, then
the mare was more likely to wean her currently suckling offspring earlier than if the next
offspring was to be a female. This in part may be likely due to preferential investment in male
offspring compared to female offspring during the time when the offspring (impending foetus) is
completely dependent on maternal investment. As far as we know, this is the first record of this
kind in ungulates (compare with review by Hewison and Gaillard, 1999). In our study of captive
plains zebra, we did not find any tendency for male offspring to be conceived later (longer after
foaling) than for female offspring as suggested for horses (Cameron et al., 1999a). Thus, captive
plains zebra mares appear to be changing the length of time between weaning and, delivery of the
next foal, suggesting they might be trading off a value of current and future offspring.
Our study did not reveal any influence of the sex of the weaned foal on the timing of the
weaning. Similarly, no differences were reported in the weaning ages of male and female foals in
studies on feral horses (Duncan et al., 1984; Cameron and Linklater, 2000). Since the suckling
foals were also consuming solid food with increasing age, the pregnant mare could invest
preferentially into the foetus, and this action would bear little risk for the survival of the suckling
J. Pluhacek et al. / Applied Animal Behaviour Science 105 (2007) 192–204 199

foal. The mare could then wean her suckling foal when the demand of the suckling foal is too
high.
In ungulates, the TWM has been tested predominantly on sexually dimorphic species
(Hewison and Gaillard, 1999; Sheldon and West, 2004). In these species the male offspring are
already bigger and grow faster than the female offspring irrespective of the maternal investment
(Cameron and Linklater, 2000; Cameron, 2004). Consequently it is much more difficult to
measure the differential investment by mothers in daughters when they are in poor condition
(Cameron and Linklater, 2000). Thus, the more sexually dimorphic a species is, the more difficult
it is to test the TWM. Equid species, including the plains zebra may be a more appropriate species
than sexually dimorphic cervids or bovids to test the TWM (Cameron et al., 1999a; Cameron and
Linklater, 2000). We assumed that all of the mothers in captivity were in good condition (due to
management, feeding, lack of predation, etc.) and would follow the TWM model for good
condition mothers (Trivers and Willard, 1973). On the other hand, the behavioural dominance
may give a better index of maternal ability than morphological or physiological measuring of
condition due to the following arguments (Sheldon and West, 2004). Firstly, resources available
for reproduction are likely to be determined more by continued future access than by prior access.
Future access to resources would seem more likely to be related to behavioural dominance than to
condition itself. Secondly, the data on behavioural dominance are measures that require repeated
sampling events and of higher quality than data relating to purely physical condition (Sheldon
and West, 2004). In our case, this might mean that dominant mares would prefer sons whereas
submissive mares would invest mainly in daughters. However, we did not find any influence
among dominant mares (behavioural dominance) on weaning time. Indeed, our results appear to
suggest that all of the mares preferred investment into male offspring. Thus, our findings do not
seem to be consistent with the TWM.
The weaning process did not influence the mortality rate of the newborn foals (Table 2). This
result suggests that mortality of the newborn zebra foals at the zoo is probably not caused by
nutritional stress, since plentiful resources were available.
Evidence of an effect on the weaning process by the stallion had not been detected in our
study. Under natural conditions, just one stallion is present in each herd of plains zebra (Klingel,
1964, 1967, 1969b; Smuts, 1976a). Therefore, the stallion uninterruptedly present within his
harem can be certain about the paternity of the foals (but see Rubenstein, 1986). This is quite
different from the situation of horses, where more than one stallion can live within a group at the
same time (Linklater et al., 1999; Linklater, 2000). It must be noted, however, that the number of
stallions in a group did not affect the timing of weaning in horses either (Cameron et al., 2003).
We did not find an influence on the weaning process by any of the following other factors that
were included in the statistical model: age of the parents, herd size, number of other suckling
foals in the herd or experience of the mother (measured as the number of parturitions as well as
the number of successful rearings per mother).
The earliest weaning age recorded in our study (243 days of age) was higher than in King’s
report (King, 1965), but lower than in two other reports (Smuts, 1976b; Becker and Ginsberg,
1990). Those reports were based on observations of wild populations, and, they neglected other
possible factors such as pregnancy, sex of foals, foetuses, etc. Thus, the weaning time of captive
plains zebra ranges within that reported for the mares in wild population. Our weaning age
finding was higher than what has been reported for feral or domestic horses, where the youngest
weaned foal was 196 (Cameron et al., 2003), 140 (Duncan et al., 1984) or 143 (Zwolinski and
Siudzinski, 1966) days old. The earliest weaning age reported for Asiatic wild ass mares (236
days; Rashek, 1976) is also lower than that in our study. It has been previously reported that, the
J. Pluhacek et al. / Applied Animal Behaviour Science 105 (2007) 192–204200

age of the 10 months has been recorded as the time of weaning in mountain zebra (Joubert, 1972;
Penzhorn, 1984). Therefore, the plains and mountain zebra mares may wean their young at a later
age than other equids. Still, as our study shows, in comparing weaning times, other factors such as
the mare’s pregnancy, the sex of her foetus and probably also the sex of her suckling foal, should
be considered. It is very likely that interspecific differences at least among horses, plains and
mountain zebras are based on differences in their gestation periods (Wackernagel, 1965; Joubert,
1974; Jones, 1976; Grubb, 1981; Lobanov, 1983; Penzhorn, 1988; Churcher, 1993). In horses,
plains and mountain zebras, the time between weaning and the next foal delivery seems to be the
same. Under optimal conditions, mares are mated shortly post-partum and the inter-birth
intervals differ due to different gestation period lengths. This might be the main reason why
horses and Asiatic wild asses wean their offspring sooner than zebra do.
Even though many interspecies differences in weaning age among equid species have been
previously discussed, factors such as sex of the foetus and time to next delivery, at least, affecting
weaning have been neglected (Becker and Ginsberg, 1990). Based on results of this study, we do
not agree with the possibility of interspecific differences in terms of weaning time among equids
as proposed by Becker and Ginsberg (1990). In captive conditions, we are able more easily to
identify some factors that affect the weaning process, which we are typically unable to measure in
the wild (e.g. age of parents, experience of the mother, etc.). In a recent meta-analysis, no
evidence was found that wild and captive ungulate populations differ in respect to effect of
maternal condition on sex ratio in relation to maternal condition (Sheldon and West, 2004).
However, we cannot fully reject the possibility that the captive condition affected our results.
5. Conclusion
In conclusion, our results suggest that there are at least two main factors: (a) pregnancy of the
mare and (b) the sex of the foetus if she is pregnant, which appears to influence the timing of
weaning in captive plains zebra. The reproductive state of the mare and the sex of her foetus
should be considered when interpreting weaning times of equids.
Acknowledgments
The authors gratefully acknowledge the assistance of the staff of Zoological Garden at Dvur
Kralove, in particular to Ludek Culık, Ales Kopecky, Dagmar Jankova, Barbara Rakova, Martina
Trohorova, Kristina Tomasova, Miroslav Sprachal, Pavel Moucha, Dana Holeckova, Ludek Bella
and Jan Parık. We thank Wayne L. Linklater, Pavel Stopka and two anonymous referees for their
helpful comments on the earlier draft of the manuscript. We are grateful to Trevor de Vries and
Jana Kalnova for improving English. The project was partly supported by grants from the Grant
Agency of the Czech Republic (523/03/H076) and from the Ministry of Agriculture of the Czech
Republic (MZE 0002701402).
References
Andersen, K.F., 1992. Size, design and interspecific interactions as restrictions of natural behaviour in multi-species
exhibits. 1. Activity and intraspecific interactions of plains zebra (Equus burchelli). Appl. Anim. Behav. Sci. 34, 157–
174.
Bateson, P., 1994. The dynamics of parent–offspring relationships in mammals. Trends Ecol. Evol. 9, 399–403.
Becker, C.D., Ginsberg, J.R., 1990. Mother-infant behaviour of wild Grevy’s zebra: adaptations for survival in semidesert
East Africa. Anim. Behav. 40, 1111–1118.
J. Pluhacek et al. / Applied Animal Behaviour Science 105 (2007) 192–204 201

Berger, J., 1986. Wild horses of the Great Basin. University of Chicago Press, Chicago.
Cameron, E.Z., 1998. Is suckling behaviour a useful predictor of milk intake? A review. Anim. Behav. 56, 521–532.
Cameron, E.Z., 2004. Facultative adjustment of mammalian sex ratios in support of the Trivers–Willard hypothesis:
evidence for a mechanism. Proc. R. Soc. London Ser. B: Biol. Sci. 271, 1723–1728.
Cameron, E.Z., Linklater, W.L., Stafford, K.J., Veltman, C.J., 1999a. Birth sex ratios relate to mare condition at
conception in Kaimanawa horses. Behav. Ecol. 10, 472–475.
Cameron, E.Z., Stafford, K.J., Linklater, W.L., Veltman, C.J., 1999b. Suckling behaviour does not measure milk intake in
horses, Equus caballus. Anim. Behav. 57, 673–678.
Cameron, E.Z., Linklater, W.L., 2000. Individual mares bias investment in sons and daughters in relation to their
condition. Anim. Behav. 60, 359–367.
Cameron, E.Z., Linklater, W.L., Stafford, K.J., Minot, E.O., 2000. Aging and improving reproductive success in horses:
declining residual reproductive value or just older and wiser? Behav. Ecol. Sociobiol. 47, 243–249.
Cameron, E.Z., Linklater, W.L., Stafford, K., 2003. Social grouping and maternal behaviour in feral horses (Equus
caballus): the influence of males on maternal protectiveness. Behav. Ecol. Sociobiol. 53, 92–101.
Cassinello, J., 1996. High ranking females bias their investment in favour of male calves in captive Ammotragus lervia.
Behav. Ecol. Sociobiol. 38, 417–424.
Cassinello, J., 1997. Mother–offspring conflict in Saharan arrui, Ammotragus lervia sahariensis. Ethology 103, 127–137.
Cassinello, J., 2001. Offspring grazing and suckling rates in a sexually dimorphic ungulate with biased maternal
investment (Ammotragus lervia). Ethology 107, 173–182.
Churcher, C.S., 1993. Equus grevyi. Mammalian Species 453, 1–9.
Clutton-Brock, T.H., Albon, S.D., Guinness, F.E., 1981. Parental investment in male and female offspring in polygynous
mammals. Nature 289, 487–489.
Counsilman, J.J., Lim, L.M., 1985. The definition of weaning. Anim. Behav. 32, 1023–1024.
Cox, D.R., 1972. Regression models and life tables. J. R. Stat. Soc. B34, 187–220.
Crowell-Davis, S.L., 1985. Nursing behaviour and maternal aggression among Welsh ponies (Equus caballus). Appl.
Anim. Behav. Sci. 14, 11–25.
Duncan, P., Harvey, P.H., Wells, S.M., 1984. On lactation and associated behaviour in a natural herd of horses. Anim.
Behav. 32, 255–263.
Ebensperger, L.A., 1998. Strategies and counterstrategies to infanticide in mammals. Biol. Rev. 73, 321–346.
Festa-Bianchet, M., 1988. Nursing behaviour of bighorn sheep: correlates of ewe age, parasitism, lamb age, birthdate and
sex. Anim. Behav. 36, 1445–1454.
Gauthier, D., Barrette, C., 1985. Suckling and weaning in captive white tailed and fallow deer. Behaviour 94, 128–149.
Green, W.C.H., Berger, J., 1990. Maternal investment in sons and daughters: problems of methodology. Behav. Ecol.
Sociobiol. 27, 99–102.
Green, W.H.C., Rothstein, A., Griswold, J.G., 1993. Weaning and parent–offspring conflict: variation relative to interbirth
interval in bison. Ethology 95, 105–125.
Grubb, P., 1981. Equus burchelli. Mammalian Species 157, 1–9.
Hewison, A.J.M., Gaillard, J.M., 1999. Successful sons or advantages daughters? The Trivers–Willard model and sex-
biased maternal investment in ungulates. Trends Ecol. Evol. 14, 229–234.
Jones, D.M., 1976. The husbandry and veterinary care of wild horses in captivity. Equine Vet. J. 8, 140–146.
Joubert, E., 1972. The social organisation and associated behaviour in the Hartmann zebra Equus zebra hartmannae.
Madoqua Ser. I. 6, 17–56.
Joubert, E., 1974. Notes on the reproduction in Hartmann zebra Equus zebra hartmannae in South West Africa. Madoqua
Ser. I. 8, 31–35.
Keiper, R., 1979. Population dynamics of feral ponies. In: Dennis, R.H. (Ed.), Symposium on the Ecology and Behavior
of Wild and Feral Equids, September 6–8. University of Wyoming, Laramie, pp. 175–183.
King, J.S., 1965. A field guide to the reproduction of Grant’s zebra and Grevy’s zebra. East Afr. Wildlife J. 3, 99–117.
Kingdon, J., 1997. The Kingdon Field Guide to African Mammals. Academic Press, London.
Klingel, H., 1964. Zur Socialstruktur des Steppenzebras (Equus quagga boehmi Matschie). Sonderdruck aus die
Naturwissenschaften 51, 347 (in German).
Klingel, H., 1967. Soziale Organisation und Verhalten freilebender Steppenzebras. Zeitschrift fur Tierpsychologie 24,
580–624 (in German, with English summary).
Klingel, H., 1969a. Reproduction in the plains zebra, Equus burchelli boehmi: behaviour and ecological factors. J. Reprod.
Fertil. Suppl. 6, 339–345.
Klingel, H., 1969b. The social organization and population ecology of the plains zebra (Equus quagga). Zool. Africana 4,
249–263.
J. Pluhacek et al. / Applied Animal Behaviour Science 105 (2007) 192–204202

Kojola, I., 1999. Sex ratio and maternal investment in ungulates. Oikos 83, 567–573.
Kruuk, L.E.B., Clutton-Brock, T.H., Albon, S.D., Pemberton, J.M., Guinness, F.E., 1999. Population density affects sex
ratio variation in red deer. Nature 399, 459–461.
Lavigueur, L., Barrette, C., 1992. Suckling, weaning and growth in captive woodland caribou. Can. J. Zool. 70, 1753–1766.
Lee, P.C., Moss, C.J., 1986. Early maternal investment in male and female African elephant calves. Behav. Ecol.
Sociobiol. 18, 353–361.
Lindstrom, J., Coulson, T., Kruuk, L., Forchhammer, M.C., Coltman, D.W., Clutton-Brock, T., 2002. Sex-ratio variation in
Soay sheep. Behav. Ecol. Sociobiol. 53, 25–30.
Linklater, W.L., 2000. Adaptive explanation in socio-ecology: lessons from the Equidae. Biol. Rev. 75, 1–20.
Linklater, W.L., Cameron, E.Z., Minot, E.O., Stafford, K.J., 1999. Stallion harassment and the mating system of horses.
Anim. Behav. 58, 295–306.
Lloyd, P.H., Rasa, O.A.E., 1989. Status, reproductive success and fitness in Cape mountain zebra (Equus zebra zebra).
Behav. Ecol. Sociobiol. 25, 411–420.
Lobanov, H.B., 1983. Predstaviteli semeistva equidae v Askanii-Nova. Vestnik zoologii 52, 55–58 (in Russian).
Martin, P., 1984. The meaning of weaning. Anim. Behav. 32, 1257–1259.
Martin-Rosset, W., Doreau, M., Cloix, J., 1978. Etude des activites d’un troupeau de poulinieres de trait et de leurs
poulains au paturage. Ann. Zootechniques 27, 33–45 (in French).
Mlıkovsky, J., 1988. Secondary sex ratio in the Przewalski horse Equus przewalskii (Mammalia: Equidae). Zeitschrift fur
Saugertierkunde 53, 92–101.
Monard, A.M., Duncan, P., Fritz, H., Feh, C., 1997. Variations in the birth sex ratio and neonatal mortality in a natural herd
of horses. Behav. Ecol. Sociobiol. 41, 243–249.
Oom, M.d.M., Reis, M.S., 1986. Ecology, social organization and behaviour of the feral horses in the Peneda-Geres
national park. Arquivos do Museu Bocage serie A3, 169–195.
Patteta, M., 2001. Survival Analysis using the Proportional Hazards Model Course Notes. SAS Institute Inc., Carry, NC,
USA.
Penzhorn, B.L., 1979. Social organisation of the Cape mountain zebra Equus z. zebra in the Mountain Zebra National
Park. Koedoe 22, 115–156.
Penzhorn, B.L., 1984. A long-term study of social organization and behaviour of Cape mountain zebra (Equus zebra
zebra). Zeitschrift fur Tierpsychologie 64, 97–146.
Penzhorn, B.L., 1988. Equus zebra. Mammalian Species 314, 1–7.
Pluhacek, J., Bartos, L., 2000. Male infanticide in captive plains zebra (Equus burchelli). Anim. Behav. 59, 689–694.
Pluhacek, J., Bartos, L., 2005. Further evidence for male infanticide and feticide in captive plains zebra, Equus burchelli.
Folia Zool. 54, 258–262.
Pluhacek, J., Bartos, L., Culık, L., 2006. High-ranking mares of captive plains zebra Equus burchelli have greater
reproductive success than low-ranking ones. Appl. Anim. Behav. Sci. 99, 315–329.
Rashek, V.A., 1976. Details of feeding and suckling behaviour in young wild asses on Barsa-Kelmes island (Aral Sea).
Zoologicheshskii Zhurnal 55, 784–787 (in Russian).
Rubenstein, D.I., 1986. Ecology and sociality in horses and zebras. In: Rubenstein, D.I., Wrangham, R.V. (Eds.), E-
cological Aspects of Social Evolution. Princeton University Press, Princeton, pp. 282–302.
Rutberg, A.T., 1986. Lactation and fetal sex ratios in American bison. Am. Nat. 127, 89–94.
Sarno, R.J., Franklin, W.L., 1999. Maternal expenditure in the polygynous and monomorphic guanaco: suckling behavior,
reproductive effort, yearly variation, and influence on juvenile survival. Behav. Ecol. 10, 41–47.
Saltz, D., 2001. Progeny sex ratio variation in ungulates: maternal age meets environmental perturbation of demography.
Oikos 94, 377–384.
Saltz, D., Rubenstein, D.I., 1995. Population dynamics of a reintroduced Asiatic wild ass (Equus hemionus) herd. Ecol.
Appl. 5, 327–335.
Schilder, M.B.H., Boer, L.P., 1987. Ethological investigation on a herd of plains zebra in a safari park: time-budgets,
reproduction and food competition. Appl. Anim. Behav. Sci. 18, 45–56.
Sheldon, B.C., West, S.A., 2004. Maternal dominance, maternal condition, and offspring sex ratio in ungulate mammals.
Am. Nat. 163, 40–54.
Sigurjonsdottir, H., Van Dierendonck, M.C., Snorrason, S., Thorhallsdottir, A.G., 2003. Social relationships in a group of
horses without a mature stallion. Behaviour 140, 783–804.
Smith-Funk, E., Crowell-Davis, S.L., 1992. Maternal behavior of draft mares (Equus caballus) with mule foals (Equus
asinus � Equus caballus). Appl. Anim. Behav. Sci. 33, 93–119.
Smuts, G.L., 1976a. Population characteristics of Burchell’s zebra (Equus burchelli antiquorum, H. Smith, 1841) in the
Kruger National Park. South Afr. J. Wildlife Res. 6, 99–112.
J. Pluhacek et al. / Applied Animal Behaviour Science 105 (2007) 192–204 203

Smuts, G.L., 1976b. Reproduction in the zebra mare Equus burchelli antiquorum from the Kruger National Park. Koedoe
19, 89–132.
Trivers, R.L., Willard, D.E., 1973. Natural selection of parental ability to vary the sex ratio. Science 179, 90–92.
Tyler, S.J., 1972. The behaviour and social organisation of the New Forest ponies. Anim. Behav. Monogr. 5, 85–196.
Wackernagel, H., 1965. Grant’s zebra, Equus burchelli boehmi, at Basle ZOO—a contribution to breeding biology. Int.
Zoo Yearbook 5, 38–41.
Wolff, A., Hausberger, M., 1994. Behaviour of foals before weaning may have some genetic basis. Ethology 96, 1–10.
Wolff, J.O., 1988. Maternal investment and sex ratio adjustment in American bison calves. Behav. Ecol. Sociobiol. 23,
127–133.
Zwolinski, J., Siudzinski, S., 1966. Obserwacje nad długoscia okresu utrzymywania zrebiat hodowlanych przy matkach.
Medycyna Weterynaryjna 22, 282–285 (in Polish).
J. Pluhacek et al. / Applied Animal Behaviour Science 105 (2007) 192–204204




















