
Effects and Effectiveness of Cavity Disinfectants in Operative ...

Journal of Applied Bacteriology 1988,65, 329-337 2732/01/88
Sensi t ivi t y of met hicillin-r esis t ant Staphylococcus aureus strains to some antibiotics, antiseptics and disinfectants
S A A D B. AL-MASAUDI, M.J. DAY & A.D. RUssELL*t Department of Applied Biology and *Welsh School of Pharmacy, University of Wales Institute of Science and Technology, PO Box 13, CardiflCF13XF, Wales
Received 27 January 1988, revised 6 June 1988 and accepted 13 June 1988
AL-MASAUDI, S.B., DAY, M.J. & RUSSELL, A.D. 1988. Sensitivity of methicillin-resistant Staphylococcus aureus strains to some antibiotics, antiseptics and disinfectants. Journal of Applied Bacteriology 65,329-337.
The effects of antibiotics, antiseptics and disinfectants against some methicillin- resistant (MRSA) and methicillin-sensitive (MSSA) Staphylococcus aureus strains have been studied. The MRSA and MSSA strains were equally sensitive to phenols, esters of para(4)-hydroxybenzoic acid and chlorhexidine but MRSA strains were slightly more resistant to quaternary ammonium compounds and considerably more so to dibromopropamidine isothionate. Some MRSA strains were also resist- ant to phenylmercuric nitrate (but not another organomercurial, thiomersal), mercu- ric chloride and cadmium chloride. All MRSA strains produced /?-lactamase. Strains from the Royal Free Hospital, London were highly resistant to /?-lactam antibiotics, erythromycin, trimethoprim and tetracyclines but were sensitive to other antibiotics. One strain from the University Hospital of Wales, Cardiff was resistant to gentami- cin but sensitive to tetracycline and trimethoprim.
In recent years there has been a considerable increase in the incidence of methicillin-resistant Staphylococcus aureus (MRSA) strains which are now recognized as being important hospital pathogens. Indeed, some strains show resistance to as many as 20 antimicrobial agents (Lyon & Skurray 1987). Moreover, there is preliminary evidence to show that strains may also exhibit decreased susceptibility to some types of disin- fectants. Some workers (Townsend et al. 1984; Tennent et al. 1985) have pointed out that the most commonly found gentamicin resistance plasmid also encodes resistance to propamidine, quaternary ammonium compounds (QACs) and ethidium bromide, and Brumfitt et al. (1985) found that MRSA strains were approximately four, nine and three times as resistant as methicillin-sensitive Staph. aureus (MSSA) strains to chlorhexidine, propamidine and the QAC, cetrimide. Another possible linkage between antibiotic and disinfectant resistance was demonstrated by Sivaji et al. (1986) who
t Corresponding author.
found that serial passage through disinfectant at subinhibitory concentrations induced antibiotic resistance in about 30% of the derived pheno- types of five strains of Staph. aureus.
We have examined the response of some MRSA and MSSA strains to several antibiotics, antiseptics and disinfectants. In addition, experi- ments have been undertaken to ascertain whether resistance to methicillin can be raised by increasing the content of cellular lipid in an MSSA strain and to determine whether rifampicin-resistant mutants of MRSA and MSSA strains showed the same response to inhibitory agents as rifampicin-sensitive cells.
Materials and Methods
BACTERIAL S T R A I N S
These consisted of Staph. aureus Oxford (NCTC 6571) and of the following strains: Royal Free Hospital, London (RF) strains 2, 3, 10, 11, 12, 13; University Hospital of Wales, Cardiff (UHW) strains 10, 11 and 12; and WG 248 and WF 1320 (WG248 containing plasmid pWG 53)

Saad B. Al-Masaudi et al. from the Western Australian Institute of Tech- nology, Bentley, Western Australia. The organisms were stored in 5% glycerol at -80°C and maintained on nutrient agar. When required, strains were grown in broth at 37°C for 24 h.
Of the RF strains, 2, 12, and 13 were phage type 88A, whereas strains RF3, RF10, and RF11 were also phage type 88A but showed weak additional reactions. All were coagulase- positive.
ANTIBIOTIC SENSITIVITY T E S T I N G
Bacterial sensitivity to antibiotics was deter- mined using sensitivity discs and Diagnostic Sensitivity Agar (DST, Oxoid), with incubation at 30'C and 37°C. In addition, DST containing 5% w h sodium chloride was used when examining sensitivity to methicillin and other 8- lactam antibiotics. In other experiments, minimum inhibitory concentrations (MICs) were determined as described below, usually at 37°C but also at 30°C when required.
SENSITIVITY T O A N T I S E P T I C S A N D
D I S I N F E C T A N T S
All agents were purchased from Sigma except for mercuric chloride (BDH) and chlorhexidine diacetate (ICI Pharmaceuticals). Three phenol- ics (phenol, chlorocresol, cresol), two QACs (cetrimide, cetylpyridinium chloride), four esters (the parabens : methyl, ethyl, propyl, butyl) of para (4)-hydroxybenzoic acid, chlorhexidine diacetate, mercuric chloride and the organomercurials thiomersal and phenylmercu- ric nitrate (PMN) were studied. Cadmium chlo- ride, and dibromopropamidine isothionate (DBPI) were also used. The MICs were deter- mined in DST agar: overdried plates (3 h at 37°C) containing different concentrations of the inhibitor were inoculated using a Denley Multi- point Inoculator (Denley, Billinghurst), with 1 p1 volumes (c. 5-10 x lo5 cfu) of undiluted over- night 37°C cultures. Plates were incubated at 37°C and the presence or absence of growth noted after 24 h. The MIC was the lowest con- centration totally preventing growth.
jl-L A c T A M A S E D E T E C T I O N
Strains were examined for 8-lactamase pro- duction by the nitrocefin method (O'Callaghan et al. 1972). Two drops of a nitrocefin solution
(500 pg/ml) were spread on the surface of 24 h agar plate cultures : 8-lactamase production was indicated by a change in colour from yellow to red.
R I F A M P I C I N - R E S I S T A N T ( R I F ~ ) M U T A N T S
A 0.1 ml volume of an overnight broth culture grown at 37°C was added to 9 ml broth and incubated at 37°C for 2 h, then 1 ml of a pre- warmed rifampicin solution (10 pgjml) was added. After further incubation for 4 h at 37"C, the cells were harvested by centrifugation (5000 rev/min for 15 min), washed and resuspended in 10 ml one-quarter strength Ringer's solution. The Denley multipoint inoculator was used to place 1 p1 volumes on to the surface of over- dried DST agar plates containing &lo0 pg/ml rifampicin. After incubation for 24 h at 37"C, RifR colonies were observed on plates contain- ing rifampicin.
Rifampicin resistant mutants were tested for their stable resistance to rifampicin, for their antibiotic, antiseptic, and disinfectant sensiti- vities and for their cell surface hydrophobicity.
F A T T E N E D CELLS
Fattened cells of Staph. aureus (Oxford) were produced by repeated daily subculturing in 3% glycerol broth for 10 d (Hugo & Stretton 1966). Sensitivity of the fattened cells in 3% glycerol- containing DST agar was determined as described above. In addition, electron micro- graphs of sections of normal (unfattened) and of fattened cells were examined.
CELL S U R F A C E H Y D R O P H O B I C I T Y
Two methods were employed: (a) bacterial adherence to hydrocarbon (BATH) method, using PUM-buffer (Rosenberg et al. 1980), the hydrocarbon xylene and the alcohol n-octanol (octan-1-01); (b) hydrophobic interaction chro- matography (HIC) method (Smyth et al. 1978), using octyl- and phenyl-Sepharose (Sigma).
T R A N S M I S S I O N E L E C T R O N M I C R O S C O P Y
Suspensions of fattened and normal (unfattened) cells of an MSSA strain were fixed in 3% vjv glutaraldehyde in 0.1 mol/l phosphate buffer for 1 H at 4°C and post-fixed with 1% w/v osmium tetroxide for 1 h. The fixed pellets were dehy- drated through a graded series of ethanol con- centrations and embedded in LR white resin

Staphylococcus aureus and disinfectants 331 Table 1. Antibiotic sensitivity" of Staphylococcus aureus strains
fi-lactamase MIC @g/ml) oft
Strain production Pen Meth Gent Eryth Vanc Teic Nov Rif
Oxford (NCTC 6571) - 0.1 <5 1 1 1 0.5 1 <0.01 Oxford RifR - ND <5 1 1 1 ND 1 >300 RF2 + >lo0 >800 1 1100 1 1.5 1 <0.01 RF3 + >lo0 >800 1 >lo0 1 1.5 1 <0.01 RFlO + >lo0 >800 2.5 >lo0 1 1.5 1 <0.01 RFll + >lo0 >800 2.5 >lo0 1 1.5 1 <0.01 RF12 + >lo0 >800 2.5 >lo0 1 1.5 1 <0.01 RF13 + >lo0 >800 2.5 >lo0 1 1.5 1 <0.01 RF13 RifR + >lo0 >800 2.5 >lo0 1 1.5 1 >300 WG 248 - 0.1 <5 1 1 1 1.5 1 <0.01 WG 1320 - 0.1 <5 1 1 1 1.5 1 <0.01 UHW 10 + ND ,800 >lo0 >lo0 1 ND 1 ND UHW 1 1 - ND <5 1 1 1 N D 1 ND UHW 12 - ND <5 1 1 1 N D 1 ND
* Disc sensitivity testing confirmed the sensitivity pattern and additionally demonstrated that methicillin- resistant strains were also resistant to other penicillins and to cephalosporins but were sensitive to chlorampheni- col and fucidin. By this method, the RF strains (but not UHW10) were found to be trimethoprim- and tetracycline-resistant.
t Pen, benzylpenicillin; Meth, methicillin; Gent, gentamicin; Eryth, erythromycin; Vanc, vancomycin; Teic, teicoplanin; Nov, novobiocin; Rif, rifampicin; ND, not determined.
(London Resin Co. Ltd, Woking, Surrey). Sec- tions were cut at a minimal 60 nm thickness and stained with 5% uranyl acetate and 2% lead citrate. Ultrastructural examination was per- formed by means of a Philips EM4ODT electron microscope.
Results
ANTIBIOTIC SENSITIVITY A N D /3-LACTAMASE PRODUCTION
The Oxford strain of Staph. aureus was highly sensitive to the entire range of antibiotics tested and did not produce B-lactamase (Table 1). Strains WG248, WG1320 and UHWll were also non-8-lactamase producers but were less sensitive to many antibiotics. B-Lactamase was not detected with strain UHW12, but this organism was resistant to penicillins. The RF strains were considerably more resistant to penicillins and cephalosporins as well as to erythromycin, trimethoprim and tetracyclines but remained sensitive to novobiocin, vancomy- cin, chloramphenicol, rifampicin and fusidic acid. They were rather less sensitive to gentami- cin than the Oxford strain. Strain UHWlO was similar in some respects to the RF strains but was highly gentamicin-resistant (MIC > 100 pg/ml) and tetracycline- and trimethoprim-
sensitive. The MICs of methicillin were >lo0 pg/ml at 30°C and 37°C for the RF strains and UHW10, whereas Oxford, WG 248, WG1320, UHWll and UHW12 were sensitive at both temperatures (MICs c 5 pg/ml).
ANTISEPTIC A N D DISINFECTANT SENSITIVITY
The effects of different antiseptics and disin- fectants on the MSSA and MRSA strains were investigated. Of the three phenolics tested, phenol was the least active and chlorocresol the most active. All strains showed the same order of response to an individual phenol, MICs being 2000 pg/ml for phenol, 200 pg/ml for chlorocre- sol and 750 pg/ml (Oxford) to 1250 pg/ml (other strains) for cresol. Strains were likewise equally sensitive to esters of para(4)-hydroxybenzoic acid, MICs being (pg/ml) 2000 (Me), lo00 (Et), 400 (Pr) and 125 (Bu).
When cationic bactericides were studied, the results depicted in Table 2 were obtained. The strains showed similar sensitivity to chlorhexi- dine, the MRSA strains requiring only slightly higher concentrations for inhibition than the Oxford strain. The MRSA strains tended to be somewhat less sensitive to the two QACs, cetylpyridinum chloride and cetrimide, but gen- erally only by a factor of 2- to 4-fold (Table 2).

332 Saad B. Al-Masaudi et al. Table 2. Sensitivity of Staphylococcus aureus strains to cationic type bacte- riocides
Sensitive (S) or resistant (R) to
MIC (pgjml) o r
Strain methicillin CHA CPC Cet DBPI
Oxford S 1.5 2.5 2.5 1
RF2 R 2.5 2.5 5 7.5 RF3 R 2 2.5 5 7.5 RFlO R 2 5 10 35 RFI 1 R 2 5 5 7.5 RF12 R 2 2.5 1.5 7.5 RF13 R 2 5 5 35 RF13 (RifR) R 2.25 N D ND ND WG248 S 1.5 2.5 5 4 WG1320 S 2.25 2.5 5 4 UHWlO R 2.25 2.5 10 10 UHWll S 2 2.5 5 7.5 UHWl2 S 2.25 2.5 5 7.5
cetrimide; DBPI, dibromopropamidine isothionate; ND, not done.
Oxford (Rif") S 2 2.5 2.5 1
* CHA, chlorhexidine diacetate; CPC, cetylpyridinium chloride; Cet,
Results with dibromopropamidine isothionate showed a greater degree of fluctuation than with any other compound. All MRSA strains were considerably more resistant to this antibacterial agent than the Oxford strain, however, in some instances by > 3O-fold (Table 2).
S E N S I T I V I T Y TO M E T A L S
Organic mercury compounds are important pharmaceutical and medical preservatives (Hugo & Russell 1982). Two such compounds were studied, thiomersal and phenylmercuric
nitrate. For comparison, mercuric chloride and cadmium chloride were also examined because Staph. aureus strains frequently show a linked antibiotic-metal resistance pattern (Hall 1970; Dyke et al. 1970).
Results are presented in Table 3. All strains showed the same order of sensitivity to thiomer- sal; all MSSA strains and one MRSA strain (surprisingly UHW10) were highly sensitive to phenylmercuric nitrate and to mercuric chloride but those MRSA strains highly resistant to the phenylmercuric compound were also resistant to mercuric chloride. Some strains were also
Table 3. Sensitivity of Staphylococcus aureus strains to metals
Sensitive (S) or resistant (R) to
MIC (pgjrnl) of*
Strain methicillin Thio PMN HgCI, CdCI,
Oxford Oxford (RifR) RF2 RF3 RFlO RF11 RF12 RF13 RF13 (Rif") WG248 WG1320 UHWlO UHWll UHWl2
S S R R R R R R R S S R S S
2.5 0.5 3 2.5 0.5 3 2.5 7.5 > 25 2.5 7.5 20 2.5 7.5 > 25 2.5 7.5 > 25 2.5 7.5 > 25 2.5 7.5 > 25 2.5 7.5 > 25 2.5 0.5 3 2.5 0.5 3 2.5 1.5 3 2.5 1.5 3 2.5 1.5 3
15 lo00 100 100 100 100 100 100 45 15 15
1500 20
1500
Thio, thiomersal; PMN, phenylmercuric nitrate.

Staphylococcus aureus and disinfectants 333 resistant to cadmium chloride, including all of the MRSA ones and UHW12.
CELL SURFACE HYDROPHOBICITY
Cell surface hydrophobicity was determined by two methods, BATH and HIC. In the BATH procedure, the O.D. of the aqueous phase after cell exposure to xylene was measured: the greater the decrease in O.D. the greater the hydrophobicity. Microscopical examination of the solvent phase established that cell lysis did not occur. The clinically isolated MSSA strains showed a high surface hydrophobicity (Figs l a & lb), but the Oxford strain was less hydropho- bic. With 0.3 ml xylene, for example, the per- centage fall in O.D. of the aqueous phase was 80 (WG248), >90 (WG1320), 90 (UHWll), >95 (UHW12) and 28 (Oxford). With the MRSA strains and 0.3 ml xylene, values of 50 (RF2), 10 (RF10) and 5 or less (UHWIO, RF3, RF11, RF12, RF13) were recorded (Figs l c and Id). With higher xylene volumes, greater partition- ing occurred into the solvent phase and differ- ences in response were noted with the UHW and individual RF strains.
Different solvents produced different responses. Strains RF2, WG248 and WG1320 showed strong activity towards an alcohol (n- octanol) traditionally widely used in partition- ing studies. Strains UHWll and UHW12 showed moderate activity and UHWlO none. Little correlation was found, however, between the BATH method and the HIC procedure (results not shown).
RIFAMPICIN-RESISTANT MUTANTS
MRSA and MSSA stains were found to be highly sensitive to rifampicin (MIC range 0.005- 0.01 pg/ml, Table 1). Mutants of Staph. aureus which are highly resistant to this antibiotic can readily be isolated both in vitro and in vivo.
Stable RifR mutants of Staph. aureus (Oxford) and Staph. aureus RF13 were isolated. The Oxford RifR strain showed a slight decrease in sensitivity to piperacillin, ceftriaxone and eryth- romycin and the RF13 RifR strain a slight increase in sensitivity to piperacillin, gentamicin and trimethoprim. No other marked differences in response to antibiotics were noted.
The Oxford RifR and RF13 RifR strains showed the same response as the original
(rifampicin-sensitive) strains to phenolics, para- bens, chlorhexidine, QACs, dibromopropami- dine, mercuric chloride, and organomercurials. The Oxford RifR strain was surprisingly more resistant to cadmium chloride than the Oxford strain itself (Table 3), but this has not been investigated further.
Surface hydrophobicity determinations, whilst not conclusive, demonstrated that the RifR cells had similar surface properties to the parent, Rifs, cells.
FATTENED CELLS
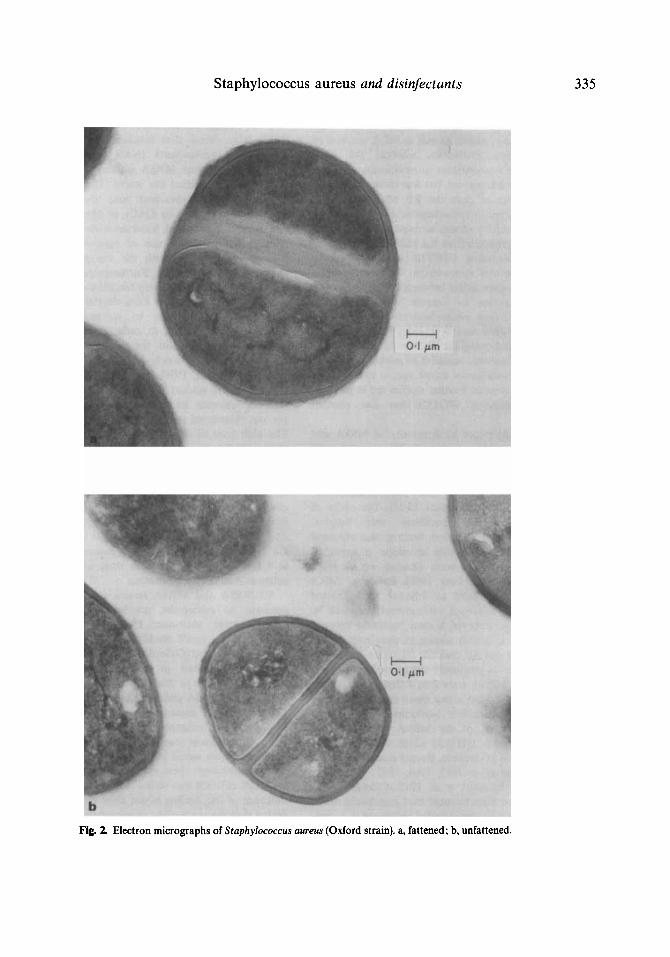
By growing Staph. aureus (Oxford) cultures repeatedly in glycerol broth the cells acquired a distinct thickening of the cell wall (Fig. 2a) as compared with normal unfattened cells (Fig. 2b). Sensitivity of fattened cell cultures was deter- mined in glycerol-containing agar and com- pared with that of unfattened cell cultures in this medium and in normal, unsupplemented agar. The MIC values (pg/ml) of different agents were as follows, the first value representing normal cells and the second fattened ones: chlorhexidine 1.5, 1.5; novobiocin 1, 1 ; methicil- lin 2, 4; cetylpyridinium chloride 2.5, 2.5; tetra- cycline 1, 1; and rifampicin 0.01, 0.1. Thus, the only marked difference occurred with rifampi- cin. Disc sensitivity testing methods revealed that the fattened culture was less sensitive than the normal strain to mezlocillin, piperacillin, cephaloridine, and cefamandole but not to the other antibiotics. The result with rifampicin (by disc testing) is surprising in view of the MIC value for rifampicin quoted above against the fattened cells.
Discussion
The Oxford strain of Staph. aureus was, as expected, highly sensitive to 8-lactam and other antibiotics and to antiseptics and preservatives. Several, but not all, of the isolates produced a fl-lactamase. Strain UHW12, however, was a non-producer but showed reduced sensitivity to some 8-lactam antibiotics. This could be the result of intrinsic resistance or of tolerance to the lethal action of penicillins and cephalo- sporins (Sabath 1982). R F strains showed the same order of response to all of the antibiotics tested, viz. resistance to methicillin and to the other fl-lactam antibiotics as well as to erythro-

3 34 Saad B. Al-Masaudi et al.
0' 0 3 0 6 0 9 1 2 I 8 0.3 0.6 0.9 1.2 I .8
Solvent volume (ml)
Fig. 1. Surface hydrophobicity of some Staphylococcus aureus strains by means of the BATH technique using xylene as test solvent. a, A, Oxford; a, WG248; 0, WG1320. b, 0, UHWl1; 0, UHW12. c, 0, RF2; 0, RF3; 0, RF10; A, RF11. d, a. RF12; O,RF13; fJ,UHW10.
Staphylococcus aureus and disinfectants
Fig. 2. Electron micrographs of Staphylococcus aureus (Oxford strain). a, fattened; b, unfattened.
335

Saad B. Al-Masaudi et al. mycin, trimethoprim and tetracycline. Of three UHW Staph. aureus strains two were methicillin-sensitive, as well as being sensitive to most other antibiotics, whereas the third (UHWlO) was resistant to methicillin, gentami- cin and erythromycin, but was more sensitive to cephalosporins than the RF strains and was also sensitive to trimethoprim and tetracycline. The R F MRSA strains were rather more resist- ant to gentamicin than the MSSA strains but all strains, including UHW 10, were sensitive to rifampicin and vancomycin. Thus, a consider- able difference exists between MRSA strains in their response to various antibiotics. Two strains of Staph. aureus were received from Aus- tralia, WG248 and WG1320 (WG248 contain- ing plasmid pWG53). In our hands, however, both strains were sensitive to methicillin and to all other antibiotics tested and to cationic and other antiseptics. Further studies are in progress to determine if WG1320 has lost plasmid pWG53.
Little difference in sensitivity of MSSA and MRSA strains to phenolic disinfectants and parabens was observed. Of the phenols, chloro- cresol was more active than cresol or phenol (ratio of MICs, molar basis: phenol : cresol 1.8- 3.1 ; phenol : chlorocresol 15.15). The order of activity of the parabens was butyl > propyl > ethyl > methyl; findings also obtained with studies involving envelope mutants of Gram-negative bacteria (Russell et al. 1985, 1987; Russell & Furr 1986). Ratios of MICs (molar basis) were as follows: methyl : ethyl 2.18; methyl : propyl 5.92; methyl : butyl 12.76. There was, however, a clear variation between MRSA and MSSA strains in their response to some but not all cationic bacteriocides. Small differences in sensitivity were noted with chlor- hexidine, MRSA were 2 to 4 times more resist- ant to QACs but many times more resistant to dibromopropamidine isothionate. One of the most resistant of the MRSA strains to this diamidine was UHWlO which was also highly resistant to gentamicin. Recent studies elsewhere (Townsend et al. 1983, 1984, 1985; Tennent et al. 1985; Brumfitt et al. 1985; Coleman et al. 1985) have demonstrated that gentamicin resist- ance plasmids also encoded resistance to propa- midine, QACs and ethidium bromide. It was suggested (Townsend et al. 1984; Tennent et al. 1985) that the extensive use of cationic disin- fectants in the hospital environment was
responsible for the emergence of antibiotic- resistant strains. Furthermore, Townsend et al. (1985) have found that resistance to nucleic acid-binding components (NAB compounds) among many recent MRSA isolates is widely distributed throughout the world. The lack of resistance to chlorhexidine and the slight increase in resistance to QACs in our MRSA strains are not in accord, however, with the sug- gestion that extensive use of cationic disin- fectants is associated with the emergence of antibiotic-resistant strains. Furthermore, dibro- mopropamidine has only very limited use.
Strains of Staph. aureus from clinical sources often present resistance to inorganic and organic mercurials and to cadmium chloride, this being associated with the presence of peni- cillinase plasmids (Novick & Roth 1968; Dyke et at. 1970; Hall, 1970). It is likely that the MRSA strains studied were able to inactivate mercuric chloride and phenylmercuric nitrate but not thiomersal (see also Weiss et al. 1977). The high level of cadmium resistance amongst all the MRSA strains may result from a specific efilux system which prevents internal accumula- tion. Recently, Coleman et al. (1985) found that methicillin- and gentamicin-resistant Staph. aureus (MGRSA) strains isolated from Dublin hospitals carried a 21 x lo6 Md plasmid confer- ring resistance to penicillin, ethidium bromide, Cd2+ and mercury. Resistance to cadmium seen in UHW12 suggests that this does not occur solely in B-lactamase producers.
All MSSA and MRSA strains studies were sensitive to rifampicin, an important anti- staphylococcal antibiotic. However, resistant (RifR) mutants could readily be isolated from two strains studied (Oxford and RF13) and were examined in detail in order to determine whether resistance to this antibiotic affected the sensitivity of MSSA and MRSA strains to other agents. The RifR mutants were stably resistant, MIC values of rifampicin being >350 pg/ml. Some differences in response to other, unrelated antibiotics were nated but these were not very marked, although there were differences when cadmium chloride was tested (Table 3). The sig- nificance of this finding is not clear but it sug- gested that the cell surface characteristics of the RifR mutants had been altered in some way, although the primary effect of this rifampicin is on RNA polymerase (Wehrli 1983). Hydropho- bicity studies with both the BATH and HIC

Staphylococcus aureus and disinfectants 337 methods revealed no differences between the parent and RifR cells and there was no increased resistance of the RifR cells to a range of antisep- tics and disinfectants.
Hydrophobicity studies demonstrated that MSSA strains were typically hydrophobic, in agreement with previous reports (Rosenberg et al. 1980; Reifsteck et al. 1987). This property may be determined by a protein or protein- associated molecule located at the cell surface (Reifsteck et al. 1987).
References BRUMFITT, W., DIXSON, S. & HAMILTON-MILLER,
J.M.T. 1985 Resistance to antiseptics in methicillin- and gentamicin-resistant Staphylococcus aureus. Lancet i, 1442-1443.
COLEMAN, D.C., POMEROY, H. & ESTERU)GE, J.K. 1985 Susceptibility to antimicrobial agents and analysis of plasmids in gentamicin and methicillin-resistant Staphylococcus aureus from Dublin hospitals. Journal of Medical Microbiology 20,157-167.
DYKE, K.G.H., PARKER, M.T. & RICHMOND, M.H. 1970 Penicillinase production and metal ion resist- ance in Staphylococcus aureus cultures isolated from hospital patients. Journal of Medical Microbiology 3, 125-136.
HALL, B.M. 1970 Mercury resistance of Staphylo- coccus aureus. Journal of Hygiene, Cambridge 68,
HUGO, W.B. & RUSSELL, A.D. 1982 Types of anti- microbial agents. In Principles and Practice of Disin- fection, Preservation and Sterilisation ed. Russell, A.D., Hugo, W.B. & Ayliffe, G.A.J. pp. 8-106. Oxford : Blackwell Scientific Publications.
HUGO, W.B. & STRE~TON, R.J. 1966 The role of cellu- lar lipid in resistance of Gram-positive bacteria to penicillin. Journal of General Microbiology 42, 133- 138.
LYON, B.R. & SKURRAY, R. 1987 Antimicrobial resist- ance of Staphylococcus aureus: Genetic basis. Microbiological Reviews 51, 88-137.
NOVICK, R.P. & ROTH, C. 1968 Plasmid-linked resist- ance to inorganic salts in Staphylococcus aureus. Journal of Bacteriology M, 1335-1342.
OCALLAGHAN, C.H., MORRIS, A., KIRBY, S.M. & SHINGLER, A.H. 1972 Novel method for detection of 8-lactamase using a chromogenic cephalosporia substrate. Antimicrobial Agents and Chemotherapy 1,283288.
REIFSTECK, F., WEE, S. & WILKINSON, B.J. 1987 Hydrophobicity - hydrophilicity of staphylococci. Journal of Medical Microbiology 24,6573.
121-129.
ROSENBERG, M., GUTNICK, D. & ROSENBERG, E. 1980 Adherence of bacteria to hydrocarbons: a simple method for measuring cell surface hydrophobicity. FEMS Microbiology Letters 9, 29-33.
RUSSELL, A.D. & FURR, J.R. 1986 Effects of antisep- tics, disinfectants and preservatives on smooth, rough and deep rough strains of Salmonella typhi- murium. International Journal of Pharmaceutics 34, 115-123.
RUSSELL, A.D., FURR, J.R. & PUGH, W.J. 1985 Suscep- tibility of porin and lipopolysaccharide-deficient mutants of Escherichia coli to a homologous series of esters of para-hydroxybenzoic acid. International Journal of Pharmaceutics 21,163-173.
RUSSELL, A.D., FURR, J.R. & PUGH, W.J. 1987 Sequen- tial loss of outer membrane lipopolysaccharide and sensitivity of Escherichia coli to antibacterial agents. International Journal of Pharmaceutics 35,227-233.
SABATH, L.D. 1982 Mechanisms of resistance to 8- lactam antibiotics in strains of Staphylococcus aureus. Annals of Internal Medicine 97,339-344.
SIVAJI, Y., MONDAL, A. & AGASWAL D.S. 1986 Disin- fectant induced changes in the antibiotic sensitivity and phage-typing pattern in Staphylococcus aureus. Journal of Hospital Infection I, 236-243.
SMYTH, C.J., JONSSON, P., OLSON, E., SODERLIND, o., ROSENGREN, J., HJERTEN, S. & WADSTROM, T. 1978 Differences in hydrophobic surface characteristics of porcine enteropathogenic Escherichia coli with or without K88 antigen as revealed by hydrophobic interaction chromatography. Infection and Immu- nity 22,462-472.
TENNENT, J.M., LYON, B.R., GILLESPIE, M.T., MAY, J.W. & SKURRAY, R.A. 1985 Cloning and expression of Staphylococcus aureus plasmid-mediated quatern- ary ammonium resistance in Escherichia coli. Anti- microbial Agents and Chemotherapy 21,7943.
TOWNSEND, D.E., ASHDOWN, N. & GRUBB, W.B. 1983 Plasmid-mediated resistance to quaternary ammon- ium compounds in methicillin-resistant Staphylo- coccus aureus. Medical Journal of Australia 2, 3 1&3 17.
TOWNSEND, D.E., ASHWWN, N., GREED, L.C. & GRUBB, W.B. 1984 Transposition of gentamicin resistance to staphylococcal plasmids encoding resistance to cationic agents. Journal of Antimicro- bial Chemotherapy 14,115-124.
TOWNSEND, D.E., ASHDOWN, N., MOMOH, M. & GRUBB, W.B. 1985 Distribution of plasmid-borne resistance to nucleic acid-binding compounds in methicillin-resistant Staphylococcus aureus. Journal of Antimicrobial Chemotherapy 15, 417434.
WEISS, A.A., MURPHY, S.D. & SILVER, S. 1977 Mercury and organomercurial resistances determined by plasmids in Staphylococcus aureus. Journal of Bacte- riology 132, 197-208.
WEHLRI, W. 1983. Rifampicin: mechanism of action and resistance. Review of Infectious Diseases 5, Sup- plement 3, 5407-541 1.
Copyright © 2022 FDOKUMEN




















