
Semantic and phonological task-set priming and stimulus processing investigated using...
14
Neuropsychologia 45 (2007) 1041–1054 Semantic and phonological task-set priming and stimulus processing investigated using magnetoencephalography (MEG) F. McNab a,b,∗ , G. Rippon a , A. Hillebrand a , K.D. Singh c , S.J. Swithenby b a The Wellcome Trust Laboratory for MEG Studies, Neurosciences Research Institute, Aston University, Birmingham, UK b Department of Physics and Astronomy, Open University, Milton Keynes, UK c School of Psychology, Cardiff University, Cardiff, UK Received 6 December 2005; received in revised form 7 July 2006; accepted 11 September 2006 Available online 23 October 2006 Abstract In this study the neural substrates of semantic and phonological task priming and task performance were investigated using single word task-primes. Magnetoencephalography (MEG) data were analysed using Synthetic Aperture Magnetometry (SAM) to determine the spatiotemporal and spectral characteristics of cortical responses. Comparisons were made between the task-prime conditions for evidence of differential effects as a function of the nature of the task being primed, and between the task-prime and the task performance responses for evidence of parallels in activation associated with preparation for and completion of a specific task. Differential priming effects were found. Left middle temporal and inferior frontal voxels showed a statistically significant power decrease associated with the semantic task-prime, and a power increase associated with the phonological task-prime, within beta and gamma frequency bands respectively. Similarities between the task-related differential effects associated with task-prime presentation and those associated with target stimulus presentation were also found. For example, within the semantic task condition, left superior frontal and middle temporal regions showed a significant power decrease within both task-prime and target epochs; within the phonological task condition there were significant parietal and cerebellar power decreases within both types of epoch. In addition there was evidence within the priming epochs of dissociable patterns of activity which could be interpreted as indices of de-activation of task-irrelevant networks. Following a phonological task-prime, significant power increases were observed in those inferior frontal and middle temporal regions in which significant power decreases were associated with semantic task priming and performance. © 2006 Elsevier Ltd. All rights reserved. Keywords: Synthetic Aperture Magnetometry (SAM); Task operations; Task cue; Language; Neuroimaging 1. Introduction 1.1. Task-sets A “task-set” has been defined as “an effective intention to perform a particular task, regardless of which of the range of task-relevant stimuli will occur” (Rogers & Monsell, 1995, p. 207). Such sets can be established by forewarning or ‘prim- ing’ the subsequent task, the improved performance associated with this taken as reflecting adaptation in preparation for task- performance (Sakai & Passingham, 2003). ∗ Corresponding author at: Karolinska Institutet, MR Centrum, Karolinska Sjukhuset N8:00, 17176 Stockholm, Sweden. Tel.: +46 8 517 76176. E-mail address: [email protected] (F. McNab). Findings of significantly longer reaction times associated with trials involving a task-switch (a “switch cost”) compared to those that do not involve a task-switch (Brass & von Cramon, 2002; Mayr & Kliegl, 2003; Meiran, 1996; Rogers & Monsell, 1995) have been interpreted in terms of a persisting activation or suppression of such “task-sets” (Arbuthnott & Woodward, 2002; Koch, 2001; Meiran, 1996). 1.1.1. Neural correlates of task-setting Although a number of studies have investigated the neural correlates associated with task-set manipulations, such as task- switching (Peri´ a˜ nez et al., 2004; Sohn, Ursu, Anderson, Stenger, & Carter, 2000) and “imposing an attentional set” (Banich et al., 2000), few have directly investigated the process of task-set preparation. Within one such functional Magnetic Resonance Imaging (fMRI) study (Sakai & Passingham, 2003) participants 0028-3932/$ – see front matter © 2006 Elsevier Ltd. All rights reserved. doi:10.1016/j.neuropsychologia.2006.09.008
Transcript of Semantic and phonological task-set priming and stimulus processing investigated using...

A
taaa
absac
ot©
K
1
1
pt2iwp
S
0d
Neuropsychologia 45 (2007) 1041–1054
Semantic and phonological task-set priming and stimulus processinginvestigated using magnetoencephalography (MEG)
F. McNab a,b,∗, G. Rippon a, A. Hillebrand a, K.D. Singh c, S.J. Swithenby b
a The Wellcome Trust Laboratory for MEG Studies, Neurosciences Research Institute, Aston University, Birmingham, UKb Department of Physics and Astronomy, Open University, Milton Keynes, UK
c School of Psychology, Cardiff University, Cardiff, UK
Received 6 December 2005; received in revised form 7 July 2006; accepted 11 September 2006Available online 23 October 2006
bstract
In this study the neural substrates of semantic and phonological task priming and task performance were investigated using single wordask-primes. Magnetoencephalography (MEG) data were analysed using Synthetic Aperture Magnetometry (SAM) to determine the spatiotemporalnd spectral characteristics of cortical responses. Comparisons were made between the task-prime conditions for evidence of differential effectss a function of the nature of the task being primed, and between the task-prime and the task performance responses for evidence of parallels inctivation associated with preparation for and completion of a specific task.
Differential priming effects were found. Left middle temporal and inferior frontal voxels showed a statistically significant power decreasessociated with the semantic task-prime, and a power increase associated with the phonological task-prime, within beta and gamma frequencyands respectively. Similarities between the task-related differential effects associated with task-prime presentation and those associated with targettimulus presentation were also found. For example, within the semantic task condition, left superior frontal and middle temporal regions showedsignificant power decrease within both task-prime and target epochs; within the phonological task condition there were significant parietal and
erebellar power decreases within both types of epoch.
In addition there was evidence within the priming epochs of dissociable patterns of activity which could be interpreted as indices of de-activationf task-irrelevant networks. Following a phonological task-prime, significant power increases were observed in those inferior frontal and middleemporal regions in which significant power decreases were associated with semantic task priming and performance.
2006 Elsevier Ltd. All rights reserved.
e; La
wt21o2
eywords: Synthetic Aperture Magnetometry (SAM); Task operations; Task cu
. Introduction
.1. Task-sets
A “task-set” has been defined as “an effective intention toerform a particular task, regardless of which of the range ofask-relevant stimuli will occur” (Rogers & Monsell, 1995, p.07). Such sets can be established by forewarning or ‘prim-
ng’ the subsequent task, the improved performance associatedith this taken as reflecting adaptation in preparation for task-erformance (Sakai & Passingham, 2003).∗ Corresponding author at: Karolinska Institutet, MR Centrum, Karolinskajukhuset N8:00, 17176 Stockholm, Sweden. Tel.: +46 8 517 76176.
E-mail address: [email protected] (F. McNab).
1
cs&apI
028-3932/$ – see front matter © 2006 Elsevier Ltd. All rights reserved.oi:10.1016/j.neuropsychologia.2006.09.008
nguage; Neuroimaging
Findings of significantly longer reaction times associatedith trials involving a task-switch (a “switch cost”) compared
o those that do not involve a task-switch (Brass & von Cramon,002; Mayr & Kliegl, 2003; Meiran, 1996; Rogers & Monsell,995) have been interpreted in terms of a persisting activationr suppression of such “task-sets” (Arbuthnott & Woodward,002; Koch, 2001; Meiran, 1996).
.1.1. Neural correlates of task-settingAlthough a number of studies have investigated the neural
orrelates associated with task-set manipulations, such as task-witching (Perianez et al., 2004; Sohn, Ursu, Anderson, Stenger,
Carter, 2000) and “imposing an attentional set” (Banich etl., 2000), few have directly investigated the process of task-setreparation. Within one such functional Magnetic Resonancemaging (fMRI) study (Sakai & Passingham, 2003) participants

1 cholo
ws(whrptap
wrtdpr“m(
tpRaa(psaaf(irgp
spafirtt
fiasa2
1
eo
ticGoctpt(csiitnfaon
1
tfigws2aass
1
dtbslasedmd
1p
i
042 F. McNab et al. / Neuropsy
ere required to remember either a sequence of letters, or theequence of spatial positions in which the letters were presentedi.e. verbal/spatial task domains). In certain trials participantsere instructed to recall the items in the order in which theyad been presented, and in others to recall the items in theeverse order (i.e. forward/backwards task operations). Task-rimes informed participants of both the task domain to whichhey should attend and the operation they should perform. It wasssumed that the presentation of such primes would lead to thereparation of the appropriate task-set.
Regardless of the domain or operation performed, activationas observed within the anterior prefrontal cortex (BA10) in
esponse to the task-prime. This activation was time-locked tohe presentation of the instruction, and was significantly strongeruring the instruction delay than during the pre-instructioneriod. These results are in keeping with previous claims ofostral prefrontal cortical regions showing an involvement inprospective memory” studies which require participants toaintain delayed “intentions to act” regardless of the task
Burgess, Scott, & Frith, 2003).Similarly Brass and von Cramon (2002) used fMRI to observe
he neural correlates associated with the presentation of task-rimes during trials in which the target stimulus failed to appear.esults indicate that the presentation of a task-prime was associ-ted with activity within a frontal network, which was describeds being similar to that identified within task-switching studiesBrass & von Cramon, 2002), and included the junction of therecentral sulcus and the inferior frontal sulcus in both hemi-pheres, and bilateral middle frontal regions. Likewise Ruge etl. (2005) reported a network of frontal and parietal areas associ-ted with the process of task preparation (including the inferiorrontal junction-IFJ). In a later study, Brass and von Cramon2004) manipulated the number of times a task was preparedn order to subtract activation related to prime encoding. Theesults indicated that the left IFJ area, the right inferior frontalyrus and the right intraparietal sulcus were involved in taskreparation.
More generally, Derrfuss, Brass, and von Cramon (2004)howed IFJ activation that was common to a task-switchingaradigm, a Stroop task and a verbal n-back task, suggestingn important role for this region in “cognitive control”. Suchndings have lead to the conclusion that the inferior IFJ “isesponsible for implementing a task set, which requires the selec-ion of relevant stimulus-response mappings for the upcomingask” (Brass & von Cramon, 2002, p. 914).
Event-related potential (ERP) studies that have also revealedrontal “general preparation” effects common to task-primesndicating either a task-switch or a task repetition are interpreteds reflecting “cue-triggered anticipatory control processes . . .
uch as top-down preparation of selective stimulus processingnd response mechanisms . . .” (Poulsen, Luu, Davey, & Tucker,005, p. 149).
.1.2. Neural correlates of differential task-settingThe above studies did not differentiate between the differ-
nt tasks that were being prepared. Based upon the definitionf a task-set as a “neurocognitive state in which an up coming
(Pta
gia 45 (2007) 1041–1054
ask is prospectively configured” (Sakai & Passingham, 2003),t seems reasonable to assume that a task-set should be spe-ific to the anticipated task. This is supported by the study ofiesbrecht, Woldorff, Song, and Mangun (2003) who carriedut direct statistical comparisons of prime related activity asso-iated with tasks that directed attention either to the location orhe color of the stimulus. They report sub-regions of a fronto-arietal network that were significantly more active during spa-ial than non-spatial orienting. Similarly Sakai and Passingham2003) showed that presentation of the task-prime was asso-iated with “domain specific activations” in regions linked topatial and verbal processing, depending on whether the tasknvolved attending to positions or letters. The task-primes alsondicated the nature of the operation to be performed. Althoughhe magnitude of activation in response to the task-prime wasot operation dependent, there were operation-dependent dif-erences in the interactions between anterior prefrontal cortexnd posterior prefrontal areas. This highlights the importancef differentiating between different tasks when investigating theeural correlates of task priming.
.1.3. Temporal correlates of differential task-settingThis is supported by the results of ERP studies. For example,
ask-specific differential effects have been observed within therst 450 ms of task-prime presentation, and again at latenciesreater than 700 ms (slow electric potentials) when participantsere primed to attend to either faces or dot patterns that were
uperimposed onto the face pictures (Tarkka & Mnatsakanian,003). Another ERP study reported that activity predicting eitherverbal or a spatial task started over parietal electrodes 160 msfter prime onset (Miniussi, Marzi, & Nobre, 2005), which againupports the idea that the priming of different tasks should betudied separately.
.1.4. Semantic versus phonological task-set comparisonsIn order then to investigate the potential dissociability of
ifferent task-sets, it is important to choose sufficiently dis-inctive yet appropriately matched tasks. The present study isased around the preparation of semantic and phonological task-ets. It has been claimed that the sub lexical/phonological andexical/semantic reading routes can be separately activated byppropriate priming (Zevin & Balota, 2000). We used primedemantic and phonological tasks but with identical stimuli withinach task condition, thereby reducing the possibility of intro-ucing potential confounds, for example, differences in eyeovements associated with differences in spatial attention (as
iscussed by Giesbrecht et al., 2003).
.2. Neural correlates of semantic versus phonologicalrocessing
Neuroimaging studies have revealed a number of differencesn activity between semantic or phonological task conditions
Abdullaev & Posner, 1998; Roskies, Fiez, Balota, Raichle, &etersen, 2001). Semantic networks have tended to include leftemporal/temporo-parietal cortex, left inferior frontal regions,nd right cerebellum (Noppeney & Price, 2003; Poldrack et al.,

cholo
1e1ePts(
stMeccss
2eiwdlG
loBsapw1etw
1p
hecacohlK
lrDtc
lFswpsradi
cic(2soan(1rieviatasoBIsoirF
lKt1tatd
phtS
F. McNab et al. / Neuropsy
999; Posner, Abdullaev, McCandliss, & Sereno, 1999; Roskiest al., 2001; Vandenberghe, Price, Wise, Josephs, & Frackowiak,996). Phonological networks have tended to consist of pari-tal regions including the supramarginal gyrus (Mummery,atterson, Hodges, & Price, 1998), the temporo-parietal junc-
ion (Petersen, Fox, Posner, Mintun, & Raichle, 1989), the leftuperior temporal lobe (Posner et al., 1999) and Broca’s areaPaulesu, Frith, & Frackowiak, 1993).
Anterior differences between the activity associated withemantic tasks and non-semantic tasks have been reported, par-icularly within BA10 (Demb et al., 1995; Kapur et al., 1994;
acLeod, Buckner, Miezin, Petersen, & Raichle, 1998; Roskiest al., 2001). Using PET, Mummery et al. (1998) specificallyompared a semantic task condition with a phonological taskondition and observed significant BA10/11 activation for theemantic task only, a finding that has been supported by othertudies (McDermott, Petersen, Watson, & Ojemann, 2003).
There have been reports of both bilateral (McDermott et al.,003) and right lateralised (Roskies et al., 2001; Vandenberghet al., 1996) cerebellar effects associated with semantic process-ng. Right cerebellar effects have been linked to semantic taskshen compared to phonological tasks, such as making rhymingecisions (Roskies et al., 2001) and functionally related to theeft prefrontal cortex during semantic processing (Desmond,abrieli, & Glover, 1998; Raichle et al., 1994).Studies designed to identify the neural correlates of phono-
ogical processing have tended to highlight the importancef parietal regions (specifically the supramarginal gyrus andA39/40) in tasks thought to require processing of the soundtructure of words and letters (Paulesu et al., 1993). Bilateralctivation within the supramarginal gyrus (BA40) and posteriorarietal regions has been observed for syllable-counting taskshen compared to various semantic tasks (Mummery et al.,998; Otten & Rugg, 2001). Bilateral effects within BA40 haveven been observed when participants were required to “attendo the relations” between rhyming words, compared to wordshich were semantically linked (McDermott et al., 2003).
.3. Temporal and spectral correlates of semantic versushonological processing
The time course of phonological and/or semantic processingas been tracked using ERPs. The N400 response is a well-stablished index of semantic processing. This negative polarityomponent emerges at about 250 ms and peaks at about 400 msfter word onset (Kutas & Hillyard, 1980). A ‘phonologicalomponent’ has been identified as late as 450 ms after wordnset, but much earlier responses, about 200 ms poststimulusave been reported depending on the task demands and stimu-us context (Khateb et al., 1999; Rodriguez-Fornells, Schmitt,utas, & Munte, 2002).It has been argued that the process of averaging trials col-
apses their rich event-related dynamics into a single evoked
esponse, discarding much of the dynamic information (Makeig,ebener, Onton, & Delorme, 2004). It has also been shownhat for higher level cognitive processes, such as language pro-esses, timing is inherently variable, and therefore poorly phase-
ttat
gia 45 (2007) 1041–1054 1043
ocked to the stimulus across trials (Hillebrand, Singh, Holliday,urlong, & Barnes, 2005; Michalewski, Prasher, & Starr, 1986)uggesting that such processes would be particularly vulnerablehen trials are averaged to obtain an evoked response. Contem-orary techniques allow analysis of trial by trial variations inpectral power, investigating ‘induced’ as opposed to ‘evoked’esponses, providing not only information about the nature andmplitude of such changes, i.e. whether there are increases orecreases in power, but also the spectral or frequency character-stics of the response (Makeig et al., 2004).
Contemporary models of brain activity are based on the con-ept of rapid and transient integration of neuronal clusters, withncreases and decreases in activity taken as a measure of theoupling and uncoupling of local areas from larger networksMakeig et al., 2004; Varela, Lachaux, Rodriguez, & Martinerie,001). The changes have been termed event-related synchroni-ation and desynchronisation (Pfurtscheller & Aranibar, 1977)r event-related spectral perturbations (Makeig et al., 2004)nd have been associated with the activation of task-relevanteuronal assemblies or de-activation of task-irrelevant areasNeuper & Pfurtscheller, 2001; Pfurtscheller & Lopes da Silva,999). Initial interpretations of decreases in power (or event-elated desynchronisations) were that they indexed activationn task-specific cortical areas, whereas increases in power (orvent-related synchronisations) were associated with deacti-ated cortex (Pfurtscheller, Stancak, & Neuper, 1996). However,t is becoming clear that increases in power may also indicatectivation of inhibitory processes where task demands requirehis, for example, in the suppression of potentially distractingctivity in divided attention tasks (Ward, 2003). Even withinpecific frequency bands, both suppression and enhancementf activity can be associated with both increased and decreasedOLD signals, varying as a function of task state (Pineda, 2005).
t is therefore unclear how spectral changes relate to other mea-ures of neural activity, although a coincidence between regionsf power decrease in the alpha and beta range and an increasen the haemodynamic response measured with fMRI has beeneported in two cognitive paradigms (Singh, Barnes, Hillebrand,orde, & Williams, 2002).
There are reports of power increases associated with phono-ogical or auditory task conditions (Krause, Lang, Laine,uusisto, & Porn, 1996; Tiihonen et al., 1991), but not semantic
ask conditions (Karrasch, Krause, Laine, Heikki Lang, & Lehto,998). It has been suggested that this may arise from seman-ic representations involving larger cortical areas (Karrasch etl., 1998), comprising a greater number of regionally and func-ionally distinct clusters, leading to the observation of powerecrease at the macroscopic level.
The emergence of sophisticated models of the spatiotem-oral dynamics of brain oscillations at different frequenciesas generated speculation as to the functional significance ofhese different frequencies (Basar, Basar-Eroglu, Karakas, &churmann, 2001). There have been relatively few studies of
he spectral characteristics of cortical responses during linguis-ic processing and no clear consensus has yet emerged. Ripponnd Brunswick (1998, 2000) describe variations in beta as a func-ion of phonological ability, and Klimesch et al. (2001) suggest

1 cholo
bBfi(aifti
1
bsTbPwt
atsom&1pdsoccch
2
2
fn
2
weW(&(
2
dA
rcctttwpsoysawr
2
SmpsuMa
2
5atd&tccai(tarn2tbbiSa
uibttaitv
044 F. McNab et al. / Neuropsy
eta variations index phonetic decoding processes. Braeutigam,ailey, & Swithenby (2001) describe variations in the gamma
requency range (20–80 Hz) as a function of semantic process-ng demands, whereas Rohm, Klimesch, Haider, & Doppelmayr2001) suggest that increases in the upper alpha range (individu-lly defined, 11–13 Hz approximately) reflect semantic process-ng. Von Stein and Sarnthein (2000) report variations in the betarequency range associated with semantic processing, althoughhe task was not specifically linguistic. The nature of our studyndicates a focus on beta and gamma activity will be appropriate.
.3.1. Task-prime/task-performanceAn additional aspect of task priming studies is that there may
e parallels between the process of establishing a particular task-et and the process of performing that task on a given stimulus.his may comprise not only the activation of task-relevant areasut also the de-activation of irrelevant areas. Although Sakai andassingham (2003) report that areas showing pre-task activityere also active during task performance, the task-specificity of
hese effects is unclear.To generate a detailed picture of the subtlety of the cortical
ctivity associated with task-setting, not only the spatial charac-eristics of the response but also the temporal and spectral aspectshould be considered. Recent developments in the applicationf beamformer techniques to magnetoencephalographic (MEG)easures offer this possibility (Hillebrand et al., 2005; RobinsonVrba, 1999; Van Veen, van Drongelen, Yuchtman, & Suzuki,
997). Furthermore, such analysis of induced activity may bearticularly advantageous to the study of linguistic task setting,ue to poor phase locking in higher level cognitive tasks. Ourtudy aimed to directly compare the spatiotemporal dynamicsf the cortical activity associated with semantic and phonologi-al task-set preparation (to test hypothesised differences) and toompare the evolution of task-set preparation with that of taskompletion within each of the two task conditions (to investigateypothesised similarities).
. Methods
.1. Participants
Eleven participants gave informed consent to take part in this study (sixemales). All stated themselves to be right handed native English speakers, withormal or corrected to normal vision.
.2. Stimuli
One hundred and twenty-eight nouns were visually presented. Sixty-fourere names of living things and 64 were names of non-living things. Within
ach of these categories 32 words had one syllable and 32 had two syllables.ords from each of these four groups were matched in terms of number of letters
3–9 letters, mean word length = 5.2, σ = 1.6) and word frequency score (KuceraFrancis, 1967) obtained from the MRC Psycholinguistic Database (1987)
maximum word frequency = 114, mean word frequency = 11.5, σ = 21.2).
.3. Procedure
Participants were shown the words on a computer monitor which was viewedirectly through a window in the MEG shielded room. There were two tasks.
semantic judgment task required participants to decide whether the word
fl
f4
gia 45 (2007) 1041–1054
eferred to a living or non-living entity and a phonological task involved syllable-ounting. Each word was presented twice; once within each of the two taskonditions. Trials were presented in pseudo-random order, so there is no reasono believe that one task condition would have involved more repeated wordshan the other. Prior to stimulus onset, participants were informed which tasko perform by the presentation of a task-prime, which took the form of a singleord question (“living?”, “non-living?”, “one?”, and “two?”). All words wereresented in black, on a white background, with a single line frame around thetimuli in order to differentiate them from the task-primes. The different typesf stimuli were evenly assigned to the different task-prime conditions. Primedes/no responses were made via a button press with the left index finger. Thetimulus sequence is represented in Fig. 1. Following a 1.5 s prestimulus period,task-prime appeared for 0.2 s informing participants which task to performhen the stimulus appeared at 1.5 s. Participants were asked to wait until a
esponse cue appeared at 3.0 s before making their response.
.4. MEG recording and coregistration with MRI data
MEG data was collected using a 151-channel CTF Omega system (CTFystems Inc., Port Coquitlam, Canada). The data were collected in third orderode at a sampling rate of 625 Hz. Following data acquisition the shape of the
articipant’s head was digitized using a 3D digitizer (Polhemus Isotrack). Thisurface was matched to that extracted from the participant’s anatomical MRI,sing Align (http://www.ece.drexel.edu/ICVC/Align/align11.html) so that theEG data obtained from each participant could be coregistered with a previously
cquired anatomical MRI scan (Adjamian et al., 2004).
.5. Data analysis
The data were bandpass filtered between 0.7 and 80 Hz. In addition a0 Hz powerline filter was used and D.C. offset was removed. The data werenalysed using Synthetic Aperture Magnetometry (SAM), which is an adap-ive beamformer technique whereby each voxel in the brain is linked to theetection array using an optimal spatial filter (Hillebrand et al., 2005; Vrba
Robinson, 2001). Such analysis enables the production of images of spec-ral power change between predefined active and passive time windows. In thease of this study, both direct and indirect comparisons were performed. Directomparisons involved treating the semantic condition as the “active” phasend the phonological condition as the “passive” phase. Indirect comparisonsnvolved comparing either the semantic condition or phonological condition“active phase”) with a prestimulus baseline (which occurred directly prior toask-prime onset and acted as the “passive phase”). The pseudoT images includedepth weighting (Hillebrand & Barnes, 2005; Vrba & Robinson, 2001). Noise
egularisation was not applied. SAM analysis enabled us to increase the sig-al to noise ratio (with the weights acting as a spatial filter) (Hillebrand et al.,005; Hoogenboom, Schoffelen, Oostenveld, Parkes, & Fries, 2006). Details ofhe sensitivity of our MEG system to signals from different brain regions cane found in Hillebrand and Barnes (2002). In addition, the spatial resolution ofeamformer images can be very high (∼1 mm) (Hillebrand & Barnes, 2005), ands generally in the order of several millimeters (Barnes, Hillebrand, Fawcett, &ingh, 2004) depending on factors such as the source strength, number of sensorsnd MEG-MRI coregistration accuracy (Hillebrand & Barnes, 2003).
Using Statistical Parametric Mapping (SPM99, Friston et al., 1995), individ-al SAM images can be spatially normalised and averaged to produce a groupmage, referred to as group SAM (Singh et al., 2002). Group SAM images cane visualised using mri3dx (http://www.jiscmail.ac.uk/lists/mri3dX.html) andheir statistical significance assessed using statistical non-parametric permuta-ion testing (SnPM) (Singh, Barnes, & Hillebrand, 2003). Only results significantt the p < 0.05 level are reported here. The SnPM procedure employed herencluded a multiple comparison correction that involved using a probability dis-ribution generated by the largest T values in the volume, instead of using the Talue at each voxel (see Singh et al., 2003 for details). No correction was made
or the number of time windows and frequency bands analysed and no clusterevel analysis was performed.SAM analysis was performed using overlapping 200 ms time windowsollowing onset of the task-prime (i.e. 0–200, 100–300, 200–400, 300–500,00–600, 500–700 and 600–800 ms), on the basis of previous literature on task

F. McNab et al. / Neuropsychologia 45 (2007) 1041–1054 1045
F e appa at 3.0
pUpuF3ssb&5
aolMvtl
3
cuht(d(ac
wwtw
wmcrcetoo(
4
woiwwit
4
iaiwccbnomttTwcwt(
4
woiT
ig. 1. The stimulus sequence. Following a 1.5 s prestimulus period, a task-primppeared at 1.5 s. Participants were asked to wait until a response cue appeared
riming (for example, Miniussi et al., 2005; Tarkka & Mnatsakanian, 2003).sing these time windows, comparisons were made between the semantic andhonological conditions, and between each type of task-prime and a prestim-lus baseline of 200 ms (which occurred prior to the onset of the task-prime).requency bands of 8–13 Hz (alpha), 14–20 Hz (beta 1), 20–30 Hz (beta 2),0–40 Hz (gamma 1) and 40–50 Hz (gamma 2) were selected for the analy-is. Data analysis within the target stimulus epoch was carried out using theame frequency bands, but in this case time windows of interest were selectedased upon the literature on semantic and phonological processing (Abdullaev
Posner, 1998; Posner & Pavese, 1998). These were 50–250, 350–550 and00–700 ms following onset of the target stimulus.
Using SAM analysis, it was possible to return to the individual datasetsnd place a so-called virtual electrode within voxels corresponding to regionsf the template brain that showed a statistically significant effect at the groupevel (Barnes & Hillebrand, 2003; Hall et al., 2004; Singh et al., 2002).
ann–Whitney time–frequency representations could then be produced from theirtual electrode output, and peak values of the Mann–Whitney Z statistic iden-ified in order to investigate time/frequency effects at the individual participantevel.
. Behavioural study
In order to validate the assumed distinctiveness of phonologi-al and semantic processing, a behavioural study was conductedsing 50 undergraduate students (46 females, 4 males, 44 rightanded, and 6 left handed), all of whom claimed English to beheir first language. The paradigm used within the MEG studyFig. 1) took the form of an incidental learning phase, the onlyifference being that in this case each word appeared only oncein either the semantic or the phonological task condition). Thellocation of the stimuli to the different task conditions wasounterbalanced across participants.
Immediately following completion of this phase, participantsere randomly presented with the 128 previously processedords, and 128 new words. For each word they were asked
o indicate, via a button press, whether they had encountered itithin the previous phase.The mean percentage of semantically encoded words that
ere correctly recognised was 69% (σ = 14%), whereas theean percentage of phonologically encoded words that were
orrectly recognised was 60% (σ = 16%). The mean false alarmate was 13.3% (σ = 9.6%). A one-way within-subjects ANOVAonducted on the recognition scores revealed a significant mainffect of task type (F(1, 96) = 9.342, p < 0.01). This indicates that
here are different types of processing associated with semanticr phonological task requirements, in keeping with the “levelf processing” memory effect described by Craik and Lockhart1972).4
t
eared for 0.2 s informing participants which task to perform when the stimuluss before making their response.
. MEG results
Significant task-dependent differential effects were identifiedithin the task-prime epoch. Also, a number of similarities werebserved between these task-specific effects and those observedn response to the presentation of the target stimulus (althoughithin the target-epoch significant effects were observed onlyith respect to the prestimulus baseline, semantic v phonolog-
cal differential effects failed to reach statistical significance athe p < 0.05 level).
.1. Priming effects
The group SAM and SnPM analyses identified three regionsn which there were significant differences between semanticnd phonological prime related activity in the epoch follow-ng task-prime presentation (Fig. 2). The first was observedithin the 0–200 ms time window, in left medial/inferior frontal
ortex (talairach coordinates: −54, 51, −15), and was asso-iated with a power increase within the 30–40 Hz frequencyand, in the phonological task-prime condition (T = 2.56), butot in the semantic task-prime condition (T = −0.06). The sec-nd was observed in the 200–400 ms time window, in leftiddle temporal cortex (talairach coordinates: −51, −42, 0)
hat was associated with a power decrease within the semanticask-prime condition (T = −2.05 in the semantic condition and= −0.22 in the phonological condition). The third was seenithin the 400–600 ms time window, in right inferior temporal
ortex (talairach coordinates: 48, −3, −51), and was associatedith a power increase within the phonological task-prime condi-
ion (T = 2.56), but not within the semantic task-prime conditionT = −1.28).
.2. Priming versus task effects
In addition to the task-dependent differential effects observedithin the task-prime epoch, a number of parallels werebserved between effects occurring within this epoch and thosen the epoch associated with target presentation (see Fig. 1).hese are shown in Fig. 3.
.3. Parallels between semantic prime and semantic task
Fig. 3a shows a statistically significant power decrease withinhe left superior frontal gyrus for the semantic task-prime con-
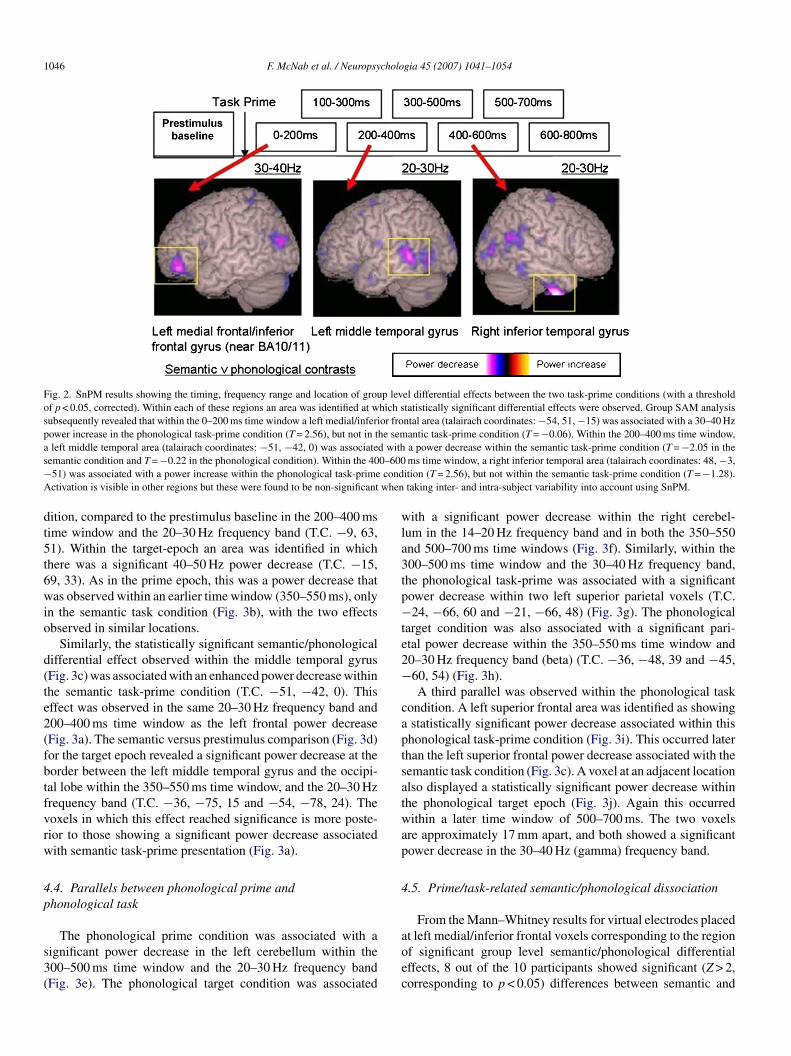
1046 F. McNab et al. / Neuropsychologia 45 (2007) 1041–1054
Fig. 2. SnPM results showing the timing, frequency range and location of group level differential effects between the two task-prime conditions (with a thresholdof p < 0.05, corrected). Within each of these regions an area was identified at which statistically significant differential effects were observed. Group SAM analysissubsequently revealed that within the 0–200 ms time window a left medial/inferior frontal area (talairach coordinates: −54, 51, −15) was associated with a 30–40 Hzpower increase in the phonological task-prime condition (T = 2.56), but not in the semantic task-prime condition (T = −0.06). Within the 200–400 ms time window,a left middle temporal area (talairach coordinates: −51, −42, 0) was associated with a power decrease within the semantic task-prime condition (T = −2.05 in thes 0–600− condA when
dt5t6wio
d(te2(fbtfvrw
4p
s3(
wla3tp−te2−
captsatwap
4
emantic condition and T = −0.22 in the phonological condition). Within the 4051) was associated with a power increase within the phonological task-primectivation is visible in other regions but these were found to be non-significant
ition, compared to the prestimulus baseline in the 200–400 msime window and the 20–30 Hz frequency band (T.C. −9, 63,1). Within the target-epoch an area was identified in whichhere was a significant 40–50 Hz power decrease (T.C. −15,9, 33). As in the prime epoch, this was a power decrease thatas observed within an earlier time window (350–550 ms), only
n the semantic task condition (Fig. 3b), with the two effectsbserved in similar locations.
Similarly, the statistically significant semantic/phonologicalifferential effect observed within the middle temporal gyrusFig. 3c) was associated with an enhanced power decrease withinhe semantic task-prime condition (T.C. −51, −42, 0). Thisffect was observed in the same 20–30 Hz frequency band and00–400 ms time window as the left frontal power decreaseFig. 3a). The semantic versus prestimulus comparison (Fig. 3d)or the target epoch revealed a significant power decrease at theorder between the left middle temporal gyrus and the occipi-al lobe within the 350–550 ms time window, and the 20–30 Hzrequency band (T.C. −36, −75, 15 and −54, −78, 24). Theoxels in which this effect reached significance is more poste-ior to those showing a significant power decrease associatedith semantic task-prime presentation (Fig. 3a).
.4. Parallels between phonological prime andhonological task
The phonological prime condition was associated with aignificant power decrease in the left cerebellum within the00–500 ms time window and the 20–30 Hz frequency bandFig. 3e). The phonological target condition was associated
aoec
ms time window, a right inferior temporal area (talairach coordinates: 48, −3,ition (T = 2.56), but not within the semantic task-prime condition (T = −1.28).taking inter- and intra-subject variability into account using SnPM.
ith a significant power decrease within the right cerebel-um in the 14–20 Hz frequency band and in both the 350–550nd 500–700 ms time windows (Fig. 3f). Similarly, within the00–500 ms time window and the 30–40 Hz frequency band,he phonological task-prime was associated with a significantower decrease within two left superior parietal voxels (T.C.24, −66, 60 and −21, −66, 48) (Fig. 3g). The phonological
arget condition was also associated with a significant pari-tal power decrease within the 350–550 ms time window and0–30 Hz frequency band (beta) (T.C. −36, −48, 39 and −45,60, 54) (Fig. 3h).A third parallel was observed within the phonological task
ondition. A left superior frontal area was identified as showingstatistically significant power decrease associated within thishonological task-prime condition (Fig. 3i). This occurred laterhan the left superior frontal power decrease associated with theemantic task condition (Fig. 3c). A voxel at an adjacent locationlso displayed a statistically significant power decrease withinhe phonological target epoch (Fig. 3j). Again this occurredithin a later time window of 500–700 ms. The two voxels
re approximately 17 mm apart, and both showed a significantower decrease in the 30–40 Hz (gamma) frequency band.
.5. Prime/task-related semantic/phonological dissociation
From the Mann–Whitney results for virtual electrodes placed
t left medial/inferior frontal voxels corresponding to the regionf significant group level semantic/phonological differentialffects, 8 out of the 10 participants showed significant (Z > 2,orresponding to p < 0.05) differences between semantic and
F. McNab et al. / Neuropsychologia 45 (2007) 1041–1054 1047
Fig. 3. Group SAM images showing a statistically significant (a) superior frontal power decrease associated with the semantic task prime (talairach co-ordinates:−9, 63, 51), (b) superior frontal power decrease in the semantic target-epoch (talairach co-ordinates: −15, 69, 33), (c) left middle temporal semantic/phonologicaldifferential effects in the task-prime epoch (this was associated with an enhanced power decrease within the semantic task-prime condition) (talairach coordinates:−51, −42, 0), (d) left middle temporal power decrease within the semantic target epoch (talairach coordinates: −36, −75, 15 and −54, −78, 24), (e) left cerebellarpower decrease associated with the phonological task prime epoch (talairach co-ordinates: −60, −69, −27), (f) right cerebellar power decrease associated with thephonological target epoch (talairach co-ordinates: 30, −87, −48 and 48, −54, −30, respectively), and left inferior frontal power decrease (talairach coordinates: −57,24, 12), (g) left superior parietal power decrease associated with the phonological task-prime epoch (talairach coordinates: −24, −66, 60 and −21, −66, 48), (h) leftinferior parietal power decrease associated with the phonological target epoch (talairach coordinates: −36, −48, 39 and −45, −60, 54), (i) power decrease within theleft middle/superior frontal gyrus, BA9/10 (talairach coordinates: −36, 51, 42) in the phonological prime epoch and (j) superior frontal power decrease associatedwith the phonological target epoch (talairach co-ordinates: −39, 39, 30) and right parietal postcentral gyrus (talairach coordinates: 60, −30, 51). 1Some of thes al sura normat late b
potpiv
ef
Fppfs
ignificant effects identified with group SAM were placed just outside the corticssociated with the process of MEG-MRI coregistration, and with the spatialherefore described with respect to the nearest cortical structure within the temp
honological activity within both the first second of targetnset and the first second of prime onset. In all of these cases
he semantic task was associated with power decrease and thehonological task associated with power increase (an examples shown in Fig. 4a and b). There was a certain degree of indi-idual variability regarding the frequency band in which thesemtt
ig. 4. An example from one representative participant of the Mann–Whitney Z stathonological task conditions, the semantic vs. prestimulus baseline comparison, andlaced within the left inferior frontal gyrus for (a) the task-prime epoch and (b) theor the task-prime epoch, and 27–29 Hz for the target-epoch). In both the task-primeeemed to arise from a power increase associated with the phonological task conditio
face. There is inevitably a certain degree of experimental error and uncertainlylisation and averaging of SAM images. The position of these activations arerain.
ffects were observed, but all occurred within the beta/gammarequency range (20–50 Hz).
A similar dissociation was observed within the leftiddle temporal gyrus. A statistically significant seman-
ic/phonological differential effect was observed within theask-prime epoch with an enhanced power decrease within the
istic as a function of time for differential effects between the semantic and thethe phonological vs. prestimulus baseline comparison for a virtual electrode
target epoch. In both cases 2 Hz wide frequency bands were used (33–35 Hzand target epochs significant (Z > 2) differential effects were observed which
n and a power decreases associated with the semantic task condition.
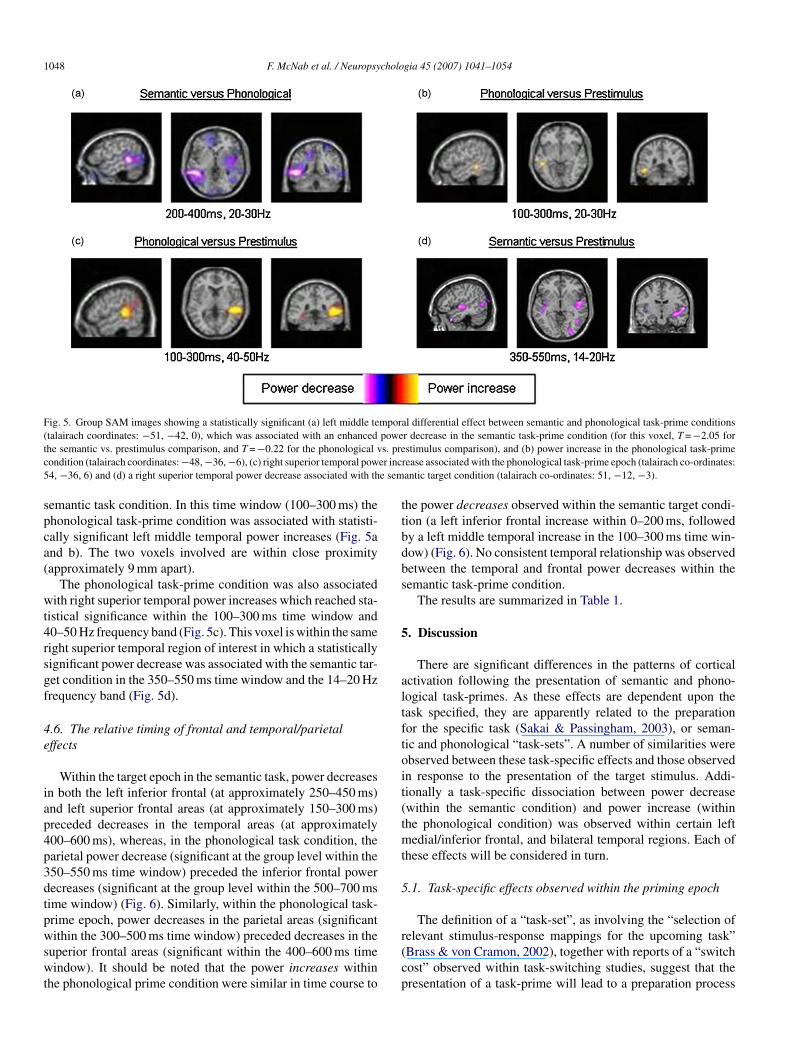
1048 F. McNab et al. / Neuropsychologia 45 (2007) 1041–1054
Fig. 5. Group SAM images showing a statistically significant (a) left middle temporal differential effect between semantic and phonological task-prime conditions(talairach coordinates: −51, −42, 0), which was associated with an enhanced power decrease in the semantic task-prime condition (for this voxel, T = −2.05 fort s. prec er inc5 e sem
spca(
wt4rsgf
4e
iap4p3dtpwswt
ttbdbs
5
altftoit(tmt
5
he semantic vs. prestimulus comparison, and T = −0.22 for the phonological vondition (talairach coordinates: −48, −36, −6), (c) right superior temporal pow4, −36, 6) and (d) a right superior temporal power decrease associated with th
emantic task condition. In this time window (100–300 ms) thehonological task-prime condition was associated with statisti-ally significant left middle temporal power increases (Fig. 5and b). The two voxels involved are within close proximityapproximately 9 mm apart).
The phonological task-prime condition was also associatedith right superior temporal power increases which reached sta-
istical significance within the 100–300 ms time window and0–50 Hz frequency band (Fig. 5c). This voxel is within the sameight superior temporal region of interest in which a statisticallyignificant power decrease was associated with the semantic tar-et condition in the 350–550 ms time window and the 14–20 Hzrequency band (Fig. 5d).
.6. The relative timing of frontal and temporal/parietalffects
Within the target epoch in the semantic task, power decreasesn both the left inferior frontal (at approximately 250–450 ms)nd left superior frontal areas (at approximately 150–300 ms)receded decreases in the temporal areas (at approximately00–600 ms), whereas, in the phonological task condition, thearietal power decrease (significant at the group level within the50–550 ms time window) preceded the inferior frontal powerecreases (significant at the group level within the 500–700 msime window) (Fig. 6). Similarly, within the phonological task-rime epoch, power decreases in the parietal areas (significant
ithin the 300–500 ms time window) preceded decreases in theuperior frontal areas (significant within the 400–600 ms timeindow). It should be noted that the power increases within
he phonological prime condition were similar in time course to
r(cp
stimulus comparison), and (b) power increase in the phonological task-primerease associated with the phonological task-prime epoch (talairach co-ordinates:antic target condition (talairach co-ordinates: 51, −12, −3).
he power decreases observed within the semantic target condi-ion (a left inferior frontal increase within 0–200 ms, followedy a left middle temporal increase in the 100–300 ms time win-ow) (Fig. 6). No consistent temporal relationship was observedetween the temporal and frontal power decreases within theemantic task-prime condition.
The results are summarized in Table 1.
. Discussion
There are significant differences in the patterns of corticalctivation following the presentation of semantic and phono-ogical task-primes. As these effects are dependent upon theask specified, they are apparently related to the preparationor the specific task (Sakai & Passingham, 2003), or seman-ic and phonological “task-sets”. A number of similarities werebserved between these task-specific effects and those observedn response to the presentation of the target stimulus. Addi-ionally a task-specific dissociation between power decreasewithin the semantic condition) and power increase (withinhe phonological condition) was observed within certain left
edial/inferior frontal, and bilateral temporal regions. Each ofhese effects will be considered in turn.
.1. Task-specific effects observed within the priming epoch
The definition of a “task-set”, as involving the “selection of
elevant stimulus-response mappings for the upcoming task”Brass & von Cramon, 2002), together with reports of a “switchost” observed within task-switching studies, suggest that theresentation of a task-prime will lead to a preparation process
F. McNab et al. / Neuropsychologia 45 (2007) 1041–1054 1049
Fig. 6. An illustration of the spatial temporal dynamics associated with frontal and temporo/parietal power changes within the two task conditions, and both thetask-prime and target epochs. In the case of the semantic task target epoch, a significant power decrease was observed in frontal and temporal regions within the sametime window used for the beamformer analysis. However, further analysis of the time–frequency Mann–Whitney representations for these regions revealed that forf left ifi iatedt d witd
tbtrttit(tbftw
2worr&ipmt
ettescSb
ttcatoStst(T
our of the five participants with a peak T value larger than 2 in both regions, thegure therefore shows that whereas frontal and parietal power decreases assoc
he task-prime and target epochs, frontal and temporal power increases observeecreases observed within the semantic target word condition.
hat is specific to the task that is being primed, as supportedy results from spatial orienting (Giesbrecht et al., 2003). Onhe basis of their fMRI results, Sakai and Passingham (2003)eported no significant task-dependant amplitude changes withinhe task-prime epoch, although they did observe a dissocia-ion between the two task conditions in terms of inter-regionalnteractions. In contrast, the results presented here show spec-ral power differences between preparation for a phonologicalsyllable counting) or a semantic (living/non-living decision)ask. Such differential effects were observed within left frontal,ilateral temporal, left parietal and left cerebellar regions. Dif-erential effects were evident in the early (0–200 ms) as well ashe later (400–600 ms) time window in the high beta/low gammaaveband (20–40 Hz).In line with previous studies (for example, Miniussi et al.,
005) task-dependent differential effects were observed earlyithin the task-prime epoch (within 0–200 ms of stimulusnset). These were seen within the left medial/inferior frontalegion (BA10/11). Although there have been reports of frontalegions being “responsible for implementing a task set (Brass
von Cramon, 2002, p. 914), and showing enhanced activation
n response to a task-prime, irrespective of the task that is beingrimed, the frontal region identified within the present study isore in line with the BA10 regions identified by studies designedo investigate the neural correlates of semantic processing (for
oips
nferior frontal power decrease preceded the left temporal power decrease. Thiswith the phonological task condition follow the same time course within bothhin the phonological task-prime condition follow the time course of the power
xample, MacLeod et al., 1998). This is also consistent withhe finding of similar task-dependent BA10 effects within thearget epoch. Within both the task and target epochs, the differ-ntial effects were associated with power decreases within theemantic condition and power increases within the phonologi-al condition (the possible significance of which is discussed inection 1). In both cases, these effects were confined to the higheta frequency band.
Task-specific differential effects were also observed withinemporal regions, during the task-prime epoch. The left middleemporal region showing this differential effect approximatelyoincides with that identified by Sakai and Passingham (2003)s showing stronger activation in response to a verbal task-primehan a spatial task-prime. The peak of the differential effect webserve is approximately 2 mm away from that identified byakai and Passingham (2003). The results presented here suggest
hat this left middle temporal region also responds differently toemantic task-primes (associated with power decreases withinhe high beta band) compared with phonological task-primesassociated with power increases within the low beta band).hese differential effects are observed within the first 400 ms
f task-prime onset. The middle temporal regions identified aren close proximity to middle temporal regions identified as sup-orting semantic processing. The voxels identified within thistudy lie within 1cm of those identified using fMRI as showing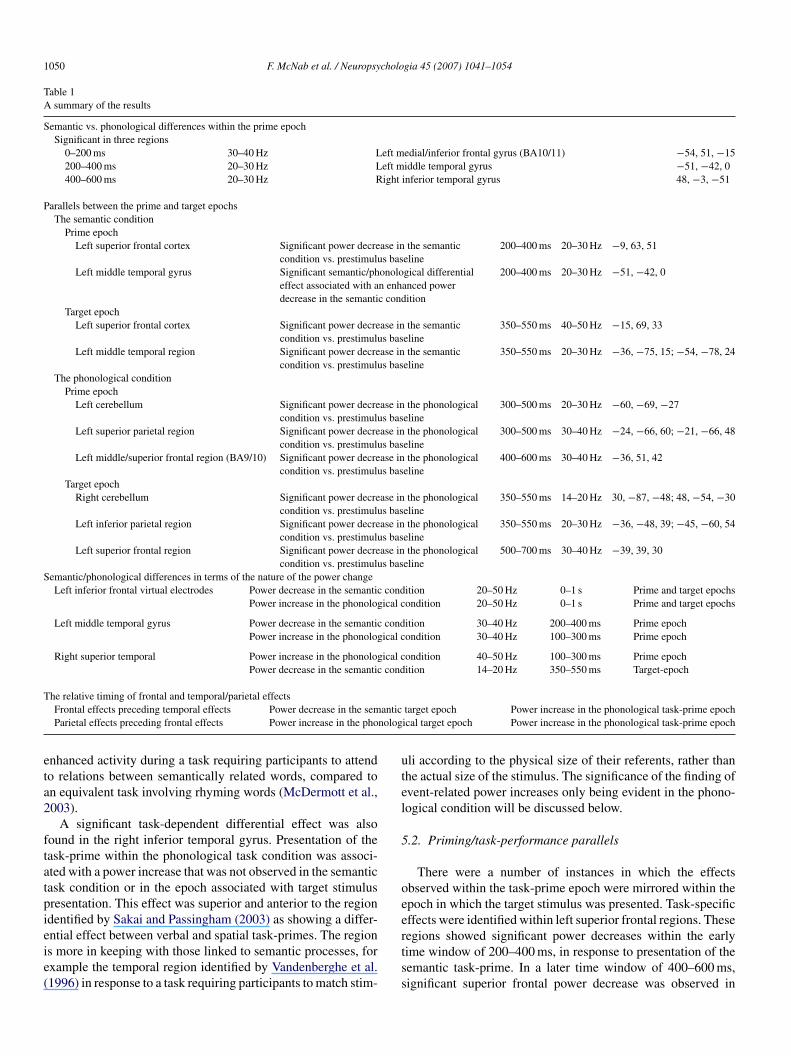
1050 F. McNab et al. / Neuropsychologia 45 (2007) 1041–1054
Table 1A summary of the results
Semantic vs. phonological differences within the prime epochSignificant in three regions
0–200 ms 30–40 Hz Left medial/inferior frontal gyrus (BA10/11) −54, 51, −15200–400 ms 20–30 Hz Left middle temporal gyrus −51, −42, 0400–600 ms 20–30 Hz Right inferior temporal gyrus 48, −3, −51
Parallels between the prime and target epochsThe semantic condition
Prime epochLeft superior frontal cortex Significant power decrease in the semantic
condition vs. prestimulus baseline200–400 ms 20–30 Hz −9, 63, 51
Left middle temporal gyrus Significant semantic/phonological differentialeffect associated with an enhanced powerdecrease in the semantic condition
200–400 ms 20–30 Hz −51, −42, 0
Target epochLeft superior frontal cortex Significant power decrease in the semantic
condition vs. prestimulus baseline350–550 ms 40–50 Hz −15, 69, 33
Left middle temporal region Significant power decrease in the semanticcondition vs. prestimulus baseline
350–550 ms 20–30 Hz −36, −75, 15; −54, −78, 24
The phonological conditionPrime epoch
Left cerebellum Significant power decrease in the phonologicalcondition vs. prestimulus baseline
300–500 ms 20–30 Hz −60, −69, −27
Left superior parietal region Significant power decrease in the phonologicalcondition vs. prestimulus baseline
300–500 ms 30–40 Hz −24, −66, 60; −21, −66, 48
Left middle/superior frontal region (BA9/10) Significant power decrease in the phonologicalcondition vs. prestimulus baseline
400–600 ms 30–40 Hz −36, 51, 42
Target epochRight cerebellum Significant power decrease in the phonological
condition vs. prestimulus baseline350–550 ms 14–20 Hz 30, −87, −48; 48, −54, −30
Left inferior parietal region Significant power decrease in the phonologicalcondition vs. prestimulus baseline
350–550 ms 20–30 Hz −36, −48, 39; −45, −60, 54
Left superior frontal region Significant power decrease in the phonologicalcondition vs. prestimulus baseline
500–700 ms 30–40 Hz −39, 39, 30
Semantic/phonological differences in terms of the nature of the power changeLeft inferior frontal virtual electrodes Power decrease in the semantic condition 20–50 Hz 0–1 s Prime and target epochs
Power increase in the phonological condition 20–50 Hz 0–1 s Prime and target epochs
Left middle temporal gyrus Power decrease in the semantic condition 30–40 Hz 200–400 ms Prime epochPower increase in the phonological condition 30–40 Hz 100–300 ms Prime epoch
Right superior temporal Power increase in the phonological condition 40–50 Hz 100–300 ms Prime epochPower decrease in the semantic condition 14–20 Hz 350–550 ms Target-epoch
Tantic
nolog
eta2
ftatpieie(
utel
5
oee
he relative timing of frontal and temporal/parietal effectsFrontal effects preceding temporal effects Power decrease in the semParietal effects preceding frontal effects Power increase in the pho
nhanced activity during a task requiring participants to attendo relations between semantically related words, compared ton equivalent task involving rhyming words (McDermott et al.,003).
A significant task-dependent differential effect was alsoound in the right inferior temporal gyrus. Presentation of theask-prime within the phonological task condition was associ-ted with a power increase that was not observed in the semanticask condition or in the epoch associated with target stimulusresentation. This effect was superior and anterior to the regiondentified by Sakai and Passingham (2003) as showing a differ-
ntial effect between verbal and spatial task-primes. The regions more in keeping with those linked to semantic processes, forxample the temporal region identified by Vandenberghe et al.1996) in response to a task requiring participants to match stim-rtss
target epoch Power increase in the phonological task-prime epochical target epoch Power increase in the phonological task-prime epoch
li according to the physical size of their referents, rather thanhe actual size of the stimulus. The significance of the finding ofvent-related power increases only being evident in the phono-ogical condition will be discussed below.
.2. Priming/task-performance parallels
There were a number of instances in which the effectsbserved within the task-prime epoch were mirrored within thepoch in which the target stimulus was presented. Task-specificffects were identified within left superior frontal regions. These
egions showed significant power decreases within the earlyime window of 200–400 ms, in response to presentation of theemantic task-prime. In a later time window of 400–600 ms,ignificant superior frontal power decrease was observed in
cholo
tttAf
oWrftept
socpBbscprtct
wwdieaosBetbf
lrppbp
ttstdZs
at
5o
oecilrmduadtis
rpicMfttwaowiif
ictfwpaoitpitao
F. McNab et al. / Neuropsy
he phonological task-prime condition. This mirrors the rela-ive latencies of significant power decreases observed withinhe target epoch within the two task conditions in this region.gain, the differences were evident in the high beta/low gamma
requency range.Assuming that this region plays a role in the executive control
f processes such as selection and retrieval (Davachi, Maril, &agner, 2001; Otten & Rugg, 2001), these results suggest the
egion is recruited earlier for the semantic executive task thanor the phonological executive task. The results also suggesthat task processing per se is not necessary for these differentialffects to become apparent, and that the presentation of a task-rime priming the appropriate task gives rise to similar effectso those observed during the process of task completion.
Further parallels were observed. For example, statisticallyignificant power decreases in the left cerebellar region werebserved within the task-prime epoch only for the phonologi-al condition, with right cerebellar power decreases within thehonological target epoch at a similar latency (300–500 ms).oth of these effects were observed within low beta frequencyands. These results agree with Mummery et al. (1998) whohow the involvement of cerebellar regions within syllable-ounting tasks, and support the interpretation that these regionslay a role in sub-vocalisation processes. They are contrary toeports of cerebellar effects being specifically linked to semanticask conditions (Roskies et al., 2001). The results suggest thaterebellar sub-vocal rehearsal processes may also be primed byhe presentation of an appropriate task-prime.
Similarly, a significant left superior parietal power decreaseas observed within the phonological task-prime epoch. Thisas in close proximity to the left superior parietal (BA40) powerecrease observed within the phonological target-epoch at a sim-lar latency (between 300 and 550 ms). Again, both of theseffects were observed within beta frequency bands. The resultsre in keeping with studies investigating the neural correlatesf phonological processing which highlight the involvement ofuch parietal regions, specifically the supramarginal gyrus andA39/40 (Mummery et al., 1998; Otten & Rugg, 2001; Paulesut al., 1993). As with the cerebellar results, this also suggestshat the task-specific involvement of these regions can be primedy the presentation of a task-prime informing participants of aorthcoming phonological task.
There is also evidence to suggest that these effects may fol-ow a task-specific time course, which can be seen both inesponse to presentation of the target stimulus and the task-rime. Whereas within the semantic target epoch the frontalower decreases preceded the temporal power decrease, withinoth the phonological target and prime epochs parietal decreasesreceded frontal decreases.
The similarities between the effects observed in response tohe presentation of a task-prime and those observed in responseo presentation of a target stimulus support the notion of a task-pecific “task-set” being established prior to presentation of the
arget stimulus. Specifically, these results support the notion ofistinct phonological and semantic “task-sets”, as suggested byevin and Balota (2000). These results extend the notion byhowing similar task-specific macroscopic neural effects, withlp
d
gia 45 (2007) 1041–1054 1051
task-specific time course, in response to presentation of theask-prime and during task completion.
.3. Dissociation between the nature of the power changebserved in the two task conditions
Additionally an apparently task-specific dissociation wasbserved in terms of the nature of the power changes. Whereasach of the significant effects observed within the semanticondition took the form of power decreases, significant powerncreases were observed in the phonological condition withineft inferior frontal, left middle temporal, right inferior tempo-al and right superior temporal regions. The underlying neuralechanisms that lead to observations of power increases and
ecreases at the macroscopic level, and might enable a greaternderstanding of this dissociation are still a matter of debates indicated in Section 1 (Pineda, 2005; Ward, 2003) Whereasecreases in activity are reliably associated with activated cor-ical areas, increases have been associated both with ‘corticaldling’ (Pfurtscheller et al., 1996) and with active inhibition oruppression of irrelevant networks (Ward, 2003).
This latter interpretation can be applied to the findingseported here. Firstly, the left inferior frontal, left middle tem-oral and right inferior temporal regions, in which powerncreases were observed within the phonological task, are typi-ally associated with semantic processing (MacLeod et al., 1998;cDermott et al., 2003). Furthermore, within the left inferior
rontal gyrus, a significant power decrease was observed withinhe semantic condition at very similar latencies and frequencieso those at which significant power increases were observedithin the phonological task condition (within both task-prime
nd target epochs). Similarly, a significant power decrease wasbserved in the target-epoch of the semantic task conditionithin the right superior temporal region, and a significant power
ncrease was observed within this region during the phonolog-cal task-prime epoch (although in this case the latencies andrequency bands differed).
Finally, the relative timing of power decrease and powerncrease observed within frontal and temporal “stores” (in thease of the semantic task) and parietal “stores” (in the case ofhe phonological task) also support this interpretation. Whereasrontal power decreases preceded temporal power decreasesithin the semantic task-prime epoch, parietal power decreasesreceded frontal power decreases within both the task-primend the target stimulus epochs. However, the power increasesbserved within temporal and left inferior frontal regions dur-ng the phonological task-prime epoch followed the relativeime course of the power decreases within the semantic task-rime epoch. This also seems to support the notion that powerncreases within the phonological condition may reflect inhibi-ion of semantic processes, both in terms of the regions involved,nd the time course of the effects. Similarly, power increasesbserved within typically ‘semantic’ regions during the phono-
ogical task condition may represent the inhibition of semanticrocesses.This interpretation is consistent with descriptions of a powerecrease as representing a correlate of an activated cortical

1 cholo
awGbnpn
indbdim2iode
wScwt
pTeigasftb
tspstna
5
ossscstt
apltoiwrgcmtpci
5
orsbslsiatt
R
A
A
A
B
B
B
B
B
052 F. McNab et al. / Neuropsy
rea (Pfurtscheller, 2001), observations of power decreasesithin a range of cognitive paradigms (Dujardin, Bourriez, &uieu, 1995; Singh et al., 2002) and the reported coincidenceetween regions of power decrease and an evoked haemody-amic response observed using fMRI (Singh et al., 2002), sup-orting the notion that a power decrease represents increasedeural activation in a cortical area.
However, there are a number of problems associated withnterpreting power decrease and power increase in such phe-omenological terms. Although, power increases have beenescribed as resulting from “the cooperative or synchronisedehaviour of a large number of neurons” (Pfurtscheller & Lopesa Silva, 1999, p. 1852), and power decreases as represent-ng desynchronised behaviour, it has been argued that such a
odel relies on a number of assumptions (Hadjipapas & Barnes,005). In particular, the “cortical idling” interpretation of powerncreases is criticised based on findings such as those showingccipital alpha power increases to covary with increasing taskemands (Jensen, Gelfand, Kounious, & Lisman, 2002; Krauset al., 1996).
The significant task-related differences reported here wereithin the high beta/low gamma frequency bands. Von Stein andarnthein (2000) suggest that beta activity is associated withonnectivity between neighbouring cortical sites and gammaith activity within localised areas, which would accord with
he spatial characteristics of these response differences.It has been suggested that reports of power increases within
honological or auditory task conditions (Krause et al., 1996;iihonen et al., 1991), but not semantic task conditions (Karrascht al., 1998), may arise from semantic representations involv-ng larger cortical areas (Karrasch et al., 1998), comprising areater number of regionally and functionally distinct clusters,nd leading to the observation of power decrease at the macro-copic level. Conversely, phonological processing may involveewer of these clusters, and therefore show power increases athe macroscopic level, even when phonological processes areeing primed.
Therefore, although a number of observations fit an interpre-ation of power increases representing inhibitory processes, iteems likely that the apparent task-specific dissociation betweenower decrease and power increase, as observed within thistudy, can only be fully understood once more is known abouthe functional organisation and the dynamics of the underlyingeuronal networks at different spatial scales. This is an excitingrea for future research.
.4. Future considerations
As anticipated, there was no significant difference in the levelf accuracy with which the two task conditions were completed,uggesting that the tasks were similar in terms of difficulty. Ithould be noted that reaction times were not collected within thistudy as a primed response was used, in an attempt to reduce
onfounds associated with response initiation occurring at theame time as stimulus processing. Syllable counting tasks haveended to be associated with longer reaction times comparedo various semantic decision tasks (for example Mummery etB
B
gia 45 (2007) 1041–1054
l., 1998). It therefore seems reasonable to assume that had thearadigm allowed the measurement of reaction times, the phono-ogical task would have been associated with longer reactionimes than the semantic task. Although this difference may havebscured the results of semantic versus phonological compar-sons (which in this study failed to reach statistical significanceithin the target epoch), by considering each condition with
espect to the prestimulus baseline, it was possible to investi-ate semantic and phonological effects separately. Future studiesould address these potential confounds by using more closelyatched task conditions. Furthermore, while this study concen-
rates on power increases and decreases associated with taskreparation, it would also be interesting to investigate neuralooperativity between brain regions in semantic and phonolog-cal task preparation.
.5. Summary
In keeping with the idea of “task-set” preparation, the resultsf this study show spectral power change differences betweenesponses to task cues that prime either a phonological or aemantic task prior to stimulus onset. Parallels were observedetween these differential effects and those observed duringtimulus processing (in terms of location, frequency band andatency). This suggests that the preparation of a phonological oremantic “task-set” involves processes that are similar to thosenvolved during phonological or semantic task completion. Inddition, there is some evidence that preparation for one kind ofask may not only involve priming for the task itself but inhibi-ion of task-irrelevant or competing areas.
eferences
bdullaev, Y. G., & Posner, M. I. (1998). Event-related brain potential imagingof semantic encoding during processing single words. NeuroImage, 7, 1–3.
djamian, P., Barnes, G. R., Hillebrand, A., Holliday, I. E., Singh, K. D.,Furlong, P. L., et al. (2004). Co-registration of MEG with MRI using bite-bar-based fiducials and surface-matching. Clinical Neurophysiology, 115(3),691–698.
rbuthnott, K. D., & Woodward, T. S. (2002). The influence of cue-task asso-ciation and location on switch cost and alternating-switch cost. CanadianJournal of Experimental Psychology, 56(1), 18–29.
anich, M. T., Milham, M. P., Atchley, R. A., Cohen, N. J., Webb, A., Wszalek,T., et al. (2000). Prefrontal regions play a predominant role in imposing anattentional “set”: Evidence from fMRI. Cognitive Brain Research, 10, 1–9.
arnes, G. R., & Hillebrand, A. (2003). Statistical flattening of MEG beam-former images. Human Brain Mapping, 18, 1–12.
arnes, G. R., Hillebrand, A., Fawcett, I. P., & Singh, K. D. (2004). Realis-tic spatial sampling for MEG beamformer images. Human Brain Mapping,23(2), 120–127.
asar, E., Basar-Eroglu, C., Karakas, S., & Schurmann, M. (2001). Gamma,alpha, delta, and theta oscillations govern cognitive processes. InternationalJournal of Psychophysiology, 39, 241–249.
raeutigam, S., Bailey, A., & Swithenby, S. (2001). Phase-locked gammaband responses to semantic violation stimuli. Cognitive Brain Research,10, 365–377.
rass, M., & von Cramon, D. Y. (2002). The role of the frontal cortex in taskpreparation. Cerebral Cortex, 12, 908–914.
rass, M., & von Cramon, D. Y. (2004). Decomposing components of taskpreparation with functional magnetic resonance imaging. Journal of Cogni-tive Neuroscience, 16(4), 609–620.

cholo
B
C
D
D
D
D
D
F
G
H
H
H
H
H
H
H
J
K
K
K
K
K
K
K
K
M
M
M
M
M
M
M
M
M
N
N
O
P
P
P
P
P
P
F. McNab et al. / Neuropsy
urgess, P. W., Scott, S. K., & Frith, C. D. (2003). The role of the rostral frontalcortex (area 10) in progressive memory: A lateral versus medial dissociation.Neuropsychologia, 41, 906–918.
raik, F. I. M., & Lockhart, R. S. (1972). Levels of processing: A frameworkfor memory research. Journal of Verbal Learning and Verbal Behavior, 11,671–684.
avachi, L., Maril, A., & Wagner, A. D. (2001). When keeping in mind sup-ports later bringing to mind: Neural markers of phonological rehearsalpredict subsequent remembering. Journal of Cognitive Neuroscience, 13(8),1059–1070.
emb, J. B., Desmond, J. E., Wagner, A. D., Vaidya, C. J., Glover, G.H., & Gabrieli, J. D. E. (1995). Semantic encoding and retrieval inthe left inferior prefrontal cortex: A functional MRI study of task diffi-culty and process specificity. The Journal of Neuroscience, 15(9), 5870–5878.
errfuss, J., Brass, M., & von Cramon, D. Y. (2004). Cognitive control in theposterior frontolateral cortex: Evidence from common activations in taskcoordination, interference control, and working memory. NeuroImage, 23,604–612.
esmond, J. E., Gabrieli, J. D. E., & Glover, G. H. (1998). Dissociation of frontaland cerebellar activity in a cognitive task: Evidence for a distinction betweenselection and search. NeuroImage, 7, 368–376.
ujardin, K., Bourriez, J. L., & Guieu, J. D. (1995). Event-related desynchro-nization (ERD) patterns during memory processes: Effects of aging and taskdifficulty. Electroencephalography and Clinical Neurophysiology/EvokedPotentials Section, 96(2), 169–182.
riston, K. J., Holmes, A. P., Worsley, K. J., Poline, J. B., Frith, C. D., & Frack-owiak, R. S. J. (1995). Statistical parametric maps in functional imaging: Ageneral linear approach. Human Brain Mapping, 2, 189–210.
iesbrecht, B., Woldorff, M. G., Song, A. W., & Mangun, G. R. (2003). Neu-ral mechanisms of top-down control during spatial and feature attention.NeuroImage, 19(3), 496–512.
adjipapas, A., & Barnes, G. R. (2005). Event related synchronisa-tion/desynchronisation in spatially extended nonlinear systems. Journal ofPsychphysiology, 19(1), 62.
all, S. D., Barnes, G. R., Hillebrand, A., Furlong, P. L., Singh, K. D., & Hol-liday, I. E. (2004). Spatio-temporal imaging of cortical desynchronisationin migraine visual aura: A magnetoencephalography case study. Headache,44(3), 204–208.
illebrand, A., & Barnes, G. R. (2002). A quantitative assessment of the sensi-tivity of whole-head MEG to activity in the adult human cortex. NeuroImage,16(3), 638–650.
illebrand, A., & Barnes, G. R. (2003). The use of anatomical constraints withMEG beamformers. NeuroImage, 20(4), 2302–2313.
illebrand, A., & Barnes, G. R. (2005). Beamformer analysis of MEG data.International Review of Neurobiology (Special Volume on Magnetoen-cephalography), 68, 149–171.
illebrand, A., Singh, K. D., Holliday, I. E., Furlong, P. L., & Barnes, G. R.(2005). A new approach to neuroimaging with magnetoencephalography.Human Brain Mapping, 5(2), 199–211.
oogenboom, N., Schoffelen, J. M., Oostenveld, R., Parkes, L. M., & Fries, P.(2006). Localizing human visual gamma-band activity in frequency, timeand space. NeuroImage, 29(3), 764–773.
ensen, O., Gelfand, J., Kounious, J., & Lisman, J. E. (2002). Oscillations inthe alpha band (9–12 Hz) increase with memory load during retention in ashort-term memory task. Cerebral Cortex, 12, 877–882.
apur, S., Craik, F. I. M., Tulving, E., Wilson, A. A., Houle, S., & Brown, G. M.(1994). Neuroanatomical correlates of encoding in episodic memory: Levelsof processing effect. Proceedings of the National Academy of Sciences, USA,91, 2008–2011.
arrasch, M., Krause, C. M., Laine, M., Heikki Lang, A., & Lehto, M. (1998).Event-related desynchronization and synchronization during an auditory lex-ical matching task. Electroencephalography and Clinical Neurophysiology,
107, 112–121.hateb, A., Annoni, J.-M., Landis, T., Pegna, A. J., Custodi, M.-C., Fonteneau,E., et al. (1999). Spatio-temporal analysis of electric brain activity duringsemantic and phonological word processing. International Journal of Psy-chophysiology, 32, 215–231.
P
gia 45 (2007) 1041–1054 1053
limesch, W., Doppelmayr, M., Wimmer, H., Gruber, W., Rohm, D., Schwaiger,J., et al. (2001). Alpha and beta band power changes in normal and dyslexicchildren. Clinical Neurophysiology, 112, 1186–1195.
och, I. (2001). Automatic and intentional activation of task sets. Journalof Experimental Psychology: Learning, Memory, and Cognition, 27(6),1474–1486.
rause, C. M., Lang, A. H., Laine, M., Kuusisto, M., & Porn, B. (1996).Event-related EEG desynchronization and synchronization during an audi-tory memory task. Electroencephalography and Clinical Neurophysiology,98, 319–326.
ucera, H., & Francis, W. (1967). Computational analysis of present-day Amer-ican English. Providence: Brown University Press.
utas, M., & Hillyard, S. A. (1980). Reading senseless sentences: Brain poten-tials reflect semantic incongruity. Science, 207, 203–205.
acLeod, A. K., Buckner, R. L., Miezin, F. M., Petersen, S. E., & Raichle,M. E. (1998). Right anterior prefrontal cortex activation during semanticmonitoring and working memory. NeuroImage, 7, 41–48.
akeig, S., Debener, S., Onton, J., & Delorme, A. (2004). Mining event-relatedbrain dynamics. Trends in Cognitive Sciences, 8(5), 204–210.
ayr, U., & Kliegl, R. (2003). Differential effects of cue changes and taskchanges on task-set selection costs. Journal of Experimental Psychology:Learning, Memory, and Cognition, 29(3), 362–372.
cDermott, K. B., Petersen, S. E., Watson, J. M., & Ojemann, J. G. (2003).A procedure for identifying regions preferentially activated by attention tosemantic and phonological relations using functional magnetic resonanceimaging. Neuropsychologia, 41, 293–303.
eiran, N. (1996). Reconfiguration of processing mode prior to task perfor-mance. Journal of Experimental Psychology: Learning, Memory, and Cog-nition, 22(6), 1423–1442.
ichalewski, H. J., Prasher, D. K., & Starr, A. (1986). Latency variability andtemporal interrelationships of the auditory event-related potentials (N1, P2,N2, and P3) in normal subjects. Electroencephalography and Clinical Neu-rophysiology/Evoked Potentials Section, 65(1), 59–71.
iniussi, C., Marzi, C. A., & Nobre, A. C. (2005). Modulation of brain activityby selective task sets observed using event-related potentials. Neuropsy-chologia, 43, 1514–1528.
RC Psycholinguistic Database (1987). http://www.psych.rl.ac.uk/MRCPsych Db.html.
ummery, C. J., Patterson, K., Hodges, J. R., & Price, C. J. (1998). Functionalneuroanatomy of the semantic system: Divisible by what? Journal of Cog-nitive Neuroscience, 10(6), 766–777.
euper, C., & Pfurtscheller, G. (2001). Event-related dynamics of corticalrhythms: Frequency-specific features and functional correlates. Interna-tional Journal of Psychophysiology, 43(1), 41–58.
oppeney, U., & Price, C. J. (2003). Functional imaging of the semantic system:Retrieval of sensory-experienced and verbally learned knowledge. Brain andLanguage, 84, 120–133.
tten, L. J., & Rugg, M. D. (2001). Task-dependency of the neural correlates ofepisodic encoding as measured by fMRI. Cerebral Cortex, 11, 1150–1160.
aulesu, E., Frith, C. D., & Frackowiak, R. S. J. (1993). The neural correlatesof the verbal component of working memory. Nature, 362, 342–345.
erianez, J. A., Maestu, F., Barcelo, F., Fernandez, A., Amo, C., & Alonso, T.O. (2004). Spatiotemporal brain dynamics during preparatory set shifting:MEG evidence. NeuroImage, 21(2), 687–695.
etersen, S. E., Fox, P. T., Posner, M. I., Mintun, M., & Raichle, M. E. (1989).Positron emission tomographic studies of the processing of single words.Journal of Cognitive Neuroscience, 1(2), 153–170.
furtscheller, G. (2001). Functional brain imaging based on powerdecrease/powers increases. Vision Research, 41, 1257–1260.
furtscheller, G., & Aranibar, A. (1977). Event-related cortical desynchronisa-tion detected by power measurement of scalp EEG. Electroencephalographyand Clinical Neurophysiology, 42(6), 817–826.
furtscheller, G., & Lopes da Silva, F. H. (1999). Event-related EEG/MEG
synchronization and desynchronization: Basic principles. Clinical Neuro-physiology, 110, 1842–1857.furtscheller, G., Stancak, A., & Neuper, C. (1996). Event-related synchronisa-tion (ERS) in the alpha band—An electrophysiological correlate of corticalidling: A review. International Journal of Psychophysiology, 24, 39–46.

1 cholo
P
P
P
P
P
R
R
R
R
R
R
R
R
R
S
S
S
S
T
T
V
V
V
V
V
054 F. McNab et al. / Neuropsy
ineda, J. A. (2005). The functional significance of mu rhythms: Translating“seeing” and “hearing” into “doing”. Brain Research Reviews, 50, 57–68.
oldrack, R. A., Wagner, A. D., Prull, M. W., Desmond, J. E., Glover, G. H., &Gabrieli, J. D. E. (1999). Functional specialization for semantic and phono-logical processing in the left inferior prefrontal cortex. NeuroImage, 10,15–35.
osner, M. I., Abdullaev, Y. G., McCandliss, B. D., & Sereno, S. C. (1999).Anatomy, circuitry and plasticity of word reading. NeuroReport, 10,R12–R23.
osner, M. I., & Pavese, A. (1998). Anatomy of word and sentence meaning.Proceedings of the National Academy of Sciences, USA, 95, 899–905.
oulsen, C., Luu, P., Davey, C., & Tucker, D. M. (2005). Dynamics of tasksets: Evidence from dense-array event-related potentials. Cognitive BrainResearch, 24, 133–154.
aichle, M. E., Fiez, J. A., Videen, T. O., MacLeod, A. M., Pardo, J. V., Fox, P.T., et al. (1994). Practice-related changes in human brain functional anatomyduring nonmotor learning. Cerebral Cortex, 4, 8–26.
ippon, G. M. J., & Brunswick, N. (1998). EEG correlates of phonologicalprocessing in dyslexic children. Journal of Psychophysiology, 12, 261–274.
ippon, G., & Brunswick, N. (2000). State and trait EEG indices of informationprocessing in developmental dyslexia. International Journal of Psychophys-iology, 36(3), 251–265.
obinson, S. E., & Vrba, J. (1999). Functional neuroimaging by synthetic aper-ture magnetometry (SAM). In Recent advances in biomagnetism. Sendai:Tohoku University Press, pp. 302–305.
odriguez-Fornells, A., Schmitt, B. M., Kutas, M., & Munte, T. F. (2002). Elec-trophysiological estimates of the time course of semantic and phonologicalencoding during listening and naming. Neuropsychologia, 40(7), 778–787.
ogers, R. D., & Monsell, S. (1995). Costs of a predictable switch between sim-ple cognitive tasks. Journal of Experimental Psychology: General, 124(2),207–231.
ohm, D., Klimesch, W., Haider, H., & Doppelmayr, M. (2001). The role oftheta and alpha oscillations for language comprehension in the human elec-troencephalogram. Neuroscience Letters, 310, 137–140.
oskies, A. L., Fiez, J. A., Balota, D. A., Raichle, M. E., & Petersen, S. E.
(2001). Task-dependent modulation of regions in the left inferior frontal cor-tex during semantic processing. Journal of Cognitive Neuroscience, 13(6),829–943.uge, H., Brass, M., Koch, I., Rubin, O., Meiran, N., & von Cramon, D. Y.(2005). Advance preparation and stimulus-induced interference in cued task
W
Z
gia 45 (2007) 1041–1054
switching: Further insights from BOLD fMRI. Neuropsychologia, 43(3),340–355.
akai, K., & Passingham, R. E. (2003). Prefrontal interactions reflect future taskoperations. Nature Neuroscience, 6(1), 75–81.
ingh, K. D., Barnes, G. R., & Hillebrand, A. (2003). Group imaging oftask-related changes in cortical synchronisation using non-parametric per-mutation testing. NeuroImage, 19(4), 1589–1601.
ingh, K. D., Barnes, G. R., Hillebrand, A., Forde, E. M. E., & Williams, A. L.(2002). Task-related changes in cortical synchronization are spatially coin-cident with the hemodynamic response. NeuroImage, 16, 103–114.
ohn, M., Ursu, S., Anderson, J. R., Stenger, A., & Carter, C. S. (2000). Therole of prefrontal cortex and posterior parietal cortex in task switching.Proceedings of the National Academy of Sciences, USA, 97(24), 13448–13453.
arkka, I. M., & Mnatsakanian, E. V. (2003). Functional specialization in thehuman frontal cortex observed during task anticipation. Neuropsychobiol-ogy, 48(2), 102–110.
iihonen, J., Hari, R., Kajola, M., Karhu, J., Ahlfors, S., & Tissari, S. (1991).Magnetoencephalographic 10-Hz rhythm from the human auditory cortex.Neuroscience Letters, 129(2), 303–305.
andenberghe, R., Price, C., Wise, R., Josephs, O., & Frackowiak, R. S. J. (1996).Functional anatomy of a common semantic system for words and pictures.Nature, 383, 254–256.
an Veen, B. D., van Drongelen, W., Yuchtman, M., & Suzuki, A. (1997).Localization of brain electrical activity via linearly constrained minimumvariance spatial filtering. IEEE Transactions on Biomedical Engineering,44(9), 867–880.
arela, F., Lachaux, J., Rodriguez, E., & Martinerie, J. (2001). The brainweb:Phase synchronisation and large-scale integration. Nature Reviews: Neuro-science, 2, 229–239.
on Stein, A., & Sarnthein, J. (2000). Different frequencies for different scalesof cortical integration: From local gamma to long range alpha/theta synchro-nisation. International Journal of Psychophysiology, 38, 301–313.
rba, J., & Robinson, S. E. (2001). Signal processing in magnetoencephalogra-phy. Methods, 25(2), 249–271.
ard, L. M. (2003). Synchronous neural oscillations and cognitive processes.Trends in Cognitive Sciences, 7, 553–559.
evin, J. D., & Balota, D. A. (2000). Priming and attentional control of lexicaland sublexical pathways during naming. Journal of Experimental Psychol-ogy: Learning, Memory, and Cognition, 26(1), 121–135.




















