
Seed-based approach for identifying flora at risk from climate warming
13
Seed-based approach for identifying flora at risk from climate warmingANNE COCHRANE, 1 * MATTHEW I. DAWS 2 † AND FIONA R. HAY 3 1 Department of Environment and Conservation, Science Division, Locked Bag 104 Bentley Delivery Centre,Western Australia 6983, Australia (Email: [email protected]); 2 Royal Botanic Gardens, Kew, Seed Conservation Department,Wakehurst Place,Ardingly,West Sussex, UK; and 3 TT Chang Genetic Resources Center, International Rice Research Institute, Metro Manila, Philippines Abstract In obligate seeding species, the germination niche is crucial for colonization and population survival. It is a high-risk phase in a plant’s life cycle, and is directly regulated by temperature. Seeds germinate over a range of temperatures within which there is an optimum temperature, with thresholds above and below which no germi- nation occurs. We suggest that abrupt changes in temperature associated with a warming climate may cause a disconnect between temperatures seeds experience and temperatures over which germination is able to occur, rendering obligate seeding species vulnerable to decline and extinction. Using a bidirectional temperature gradient system, we examined the thermal constraints in the germination niche of some geographically restricted species from the low altitude mountains of the Stirling Range, southernWestern Australia, including seedlots from lowland populations of four of these species. We demonstrated that high temperatures are not a limiting factor for germination in some restricted species, signifying a lack of relationship between geographic range size and breadth of the germination niche. In contrast, we identified other restricted species, in particular Sphenotoma drummondii, as being at risk of recruitment failure as a consequence of warming: seeds of this species showed a strong negative relationship between percentage germination and increasing temperature above a relatively low optimum constant temperature (13°C).We found some ecotypic differences in the temperature profiles between seeds collected from montane or lowland populations of Andersonia echinocephala, and while specific populations may become more restricted, they are perhaps at less risk of extinction from climate warming.This seed-based approach for identifying extinction risk will contribute tangibly to efforts to predict plant responses to environmental change and will assist in prioritizing species for management actions, directing limited resources towards further investigations and can supplement bioclimatic modelling. Key words: climate change, germination niche, seed germination, south-west Australia, temperature gradient plate, vulnerability. INTRODUCTION Climatic variables influence reproductive traits such as seed germination and seedling growth (Fenner & Thompson 2005). Although soil moisture availability is critical for seedling emergence and survival, tem- perature is arguably the most important climatic vari- able as it synchronizes germination to environmental conditions most suitable for seedling establishment, such as high likelihood of adequate water availability (Bell 1994; Fenner & Thompson 2005).Temperature affects seed responses through regulating dormancy loss in both dry and moist dormant seeds, and germi- nation rate in non-dormant seeds (Probert 2000). Germination and seedling establishment are the highest-risk phases in the life cycle of plants (Harper 1977; Silverton & Charlesworth 2001). Consequently, mechanisms that minimize the risk will be under strong selection pressure (Meyer et al. 1997), with natural selection favouring seed germination patterns that increase the probability of successful seedling establishment. For example, although temperature is not considered to be a limiting factor for recruitment in Mediterranean-type ecosystems (Lloret et al. 2004), germination generally coincides with early winter when temperatures are low (typically 10–15°C; Bell 1999) and rainfall is reliable (Mott 1972; Bellairs & Bell 1990; Bell & Bellairs 1992; Bell et al. 1995). This maximizes the period of root development prior to summer drought, thereby increasing seedling survival (Bell 1999). For many species, temperature-mediated germination cues are important to ensure population persistence. Consequently, environmental change, including anthropogenic climate warming, might be *Corresponding author. †Current address: Energy Resources of Australia, Level 3, Energy House, 18–20 Cavenagh Street, Darwin 0800 NT Australia. Accepted for publication October 2010. Austral Ecology (2011) 36, 923–935 © 2010 The Authors doi:10.1111/j.1442-9993.2010.02211.x Journal compilation © 2010 Ecological Society of Australia
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Seed-based approach for identifying flora at risk from climate warming

Seed-based approach for identifying flora at risk fromclimate warmingaec_2211 923..935
ANNE COCHRANE,1* MATTHEW I. DAWS2† AND FIONA R. HAY3
1Department of Environment and Conservation, Science Division, Locked Bag 104 Bentley DeliveryCentre,Western Australia 6983, Australia (Email: [email protected]); 2Royal BotanicGardens, Kew, Seed Conservation Department,Wakehurst Place, Ardingly,West Sussex, UK; and 3TTChang Genetic Resources Center, International Rice Research Institute, Metro Manila, Philippines
Abstract In obligate seeding species, the germination niche is crucial for colonization and population survival. Itis a high-risk phase in a plant’s life cycle, and is directly regulated by temperature. Seeds germinate over a range oftemperatures within which there is an optimum temperature, with thresholds above and below which no germi-nation occurs. We suggest that abrupt changes in temperature associated with a warming climate may cause adisconnect between temperatures seeds experience and temperatures over which germination is able to occur,rendering obligate seeding species vulnerable to decline and extinction. Using a bidirectional temperature gradientsystem, we examined the thermal constraints in the germination niche of some geographically restricted speciesfrom the low altitude mountains of the Stirling Range, southernWestern Australia, including seedlots from lowlandpopulations of four of these species. We demonstrated that high temperatures are not a limiting factor forgermination in some restricted species, signifying a lack of relationship between geographic range size and breadthof the germination niche. In contrast, we identified other restricted species, in particular Sphenotoma drummondii,as being at risk of recruitment failure as a consequence of warming: seeds of this species showed a strong negativerelationship between percentage germination and increasing temperature above a relatively low optimum constanttemperature (13°C).We found some ecotypic differences in the temperature profiles between seeds collected frommontane or lowland populations of Andersonia echinocephala, and while specific populations may become morerestricted, they are perhaps at less risk of extinction from climate warming.This seed-based approach for identifyingextinction risk will contribute tangibly to efforts to predict plant responses to environmental change and will assistin prioritizing species for management actions, directing limited resources towards further investigations and cansupplement bioclimatic modelling.
Key words: climate change, germination niche, seed germination, south-west Australia, temperature gradientplate, vulnerability.
INTRODUCTION
Climatic variables influence reproductive traits suchas seed germination and seedling growth (Fenner &Thompson 2005). Although soil moisture availabilityis critical for seedling emergence and survival, tem-perature is arguably the most important climatic vari-able as it synchronizes germination to environmentalconditions most suitable for seedling establishment,such as high likelihood of adequate water availability(Bell 1994; Fenner & Thompson 2005). Temperatureaffects seed responses through regulating dormancyloss in both dry and moist dormant seeds, and germi-nation rate in non-dormant seeds (Probert 2000).
Germination and seedling establishment are thehighest-risk phases in the life cycle of plants (Harper1977; Silverton & Charlesworth 2001). Consequently,mechanisms that minimize the risk will be understrong selection pressure (Meyer et al. 1997), withnatural selection favouring seed germination patternsthat increase the probability of successful seedlingestablishment. For example, although temperature isnot considered to be a limiting factor for recruitmentin Mediterranean-type ecosystems (Lloret et al. 2004),germination generally coincides with early winterwhen temperatures are low (typically 10–15°C; Bell1999) and rainfall is reliable (Mott 1972; Bellairs &Bell 1990; Bell & Bellairs 1992; Bell et al. 1995). Thismaximizes the period of root development prior tosummer drought, thereby increasing seedling survival(Bell 1999). For many species, temperature-mediatedgermination cues are important to ensure populationpersistence. Consequently, environmental change,including anthropogenic climate warming, might be
*Corresponding author.†Current address: Energy Resources of Australia, Level 3,
Energy House, 18–20 Cavenagh Street, Darwin 0800 NTAustralia.
Accepted for publication October 2010.
Austral Ecology (2011) 36, 923–935
© 2010 The Authors doi:10.1111/j.1442-9993.2010.02211.xJournal compilation © 2010 Ecological Society of Australia

expected to have significant effects on plant distribu-tion and survival as the timing of this key life-historytransition of seedling emergence, may no longer beoptimal (Lloret et al. 2004; Gworek et al. 2006).
Sensitivity to climate change for a given species willdepend on its ecological niche and geographic distri-bution (Thuiller et al. 2005; Broennimann et al. 2006),life-history characteristics, and genetic and phenotypicplasticity (Aitken et al. 2008). Germination nichebreadth may influence the ecological breadth and geo-graphic ranges of species (Donohue et al. 2010) andspecies with a broad germination niche might beexpected to have larger geographic ranges than specieswith narrow germination niches, although this remainsunconfirmed (Thompson & Ceriani 2003). Mostspecies can tolerate some short-term variability inclimate through phenotypic plasticity, although pre-dicted responses to global warming generally involveshifts in either seasonality (Jump & Peñuelas 2005;Bradshaw & Holzapfel 2008) or, for alpine species,altitude (Gworek et al. 2006; Parolo & Rossi 2008),rather than the shifts in thermal tolerance that wouldbe necessary for long-term persistence. In addition,those species most vulnerable to a warming climatemay be those either with restricted niches, in regionswith high anomalies in climate, or with physical barri-ers to migration (Broennimann et al. 2006; Malcolmet al. 2006). Plants with limited dispersal ability, lowreproductive rates (Jump & Peñuelas 2005), obligateseeding (Bell 2001; Bond & Midgley 2001), and thosealready restricted to the physical limits of their range,such as to higher latitudes and upper montane loca-tions (Walther et al. 2002; Parmesan & Yohe 2003;Pauli et al. 2003; Root et al. 2003), are presumed to bemost threatened by climate change.
Fire-prone, species-rich Mediterranean regions ofthe world with their cool wet winters and hot drysummers have been recognized as potentially highlyvulnerable to climate change (Allen 2003; Cowlinget al. 2004). Increasing temperatures and changes inamount and variability of rainfall may also alter firefrequency, further endangering this rich plant diversity(Pittock 2003; Pitman et al. 2007). Under projectedclimate change scenarios, there are suggestions that analmost total collapse of flora endemic to the south-westregion of Western Australia will occur (Hughes et al.1996; Pouliquen-Young & Newman 2000; Malcolmet al. 2006; Fitzpatrick et al. 2008). Similar negativeimpacts on native flora have been predicted for speciesin the Cape Floristic region of South Africa (Midgleyet al. 2002; Hannah et al. 2005; Broennimann et al.2006). However, these predictions are based on theresponse of adult plants to edaphic conditions.
Few investigations have considered the effect ofclimate warming on seed traits or the germinationniche, despite the insights that can be provided bystudying the influence of temperature on seed dor-
mancy and germination in the context of environmentalchange (Ooi et al. 2009). Investigation of germinationtemperatures for seeds of species from south-westWestern Australia has been substantial, but mostlybased on germination at a limited range of constanttemperatures (Bell et al. 1995). For these species, tem-perature optima are generally low, indicative of wintertemperatures when seedling establishment and survivalis likely to be optimal (Bell 1999). However, germina-tion and early seedling responses to temperature forspecies from Western Australia’s low southern moun-tains have not previously been investigated. Speciesfrom this region are likely to be at elevated risk fromclimate change as there is no opportunity for furtherupward or southward migration and this region has agreater predicted rise in temperature compared withthe rest of south-west Australia (Hope et al. 2006).Weuse these species for closer examination because of theirphysical limits to distribution spread and their likeli-hood of facing extinction risk.They provide a case studyfor assessing direct physiological constraints on recruit-ment, for example the width of the germination niche,in particular the width of the temperature window forgermination and growth. Recruitment constraints mayprovide a measure of susceptibility to climate change:species that only germinate over a narrow temperaturewindow are potentially most at risk. Temperaturesoutside that window represent no, or limited, opportu-nity for germination.
We report on the thermal limits for germination andearly seedling growth for 10 perennial plant speciesendemic to southern Western Australia, with sixspecies restricted to the summits of the major moun-tain range in the region (the Stirling Range; maximumelevation <1100 m a.s.l.). We determined (i) whethera restricted geographic distribution and mountainhabitat can explain thermal constraints in the germi-nation niche; (ii) whether thermal constraints for earlyseedling root growth follow the same pattern as forgermination; and (iii) whether the niche limits aredifferent between lowland and montane populations.Because niche characteristics can be powerful indica-tors of species sensitivities to climate change, wediscuss our data in the context of developing predic-tors of extinction risk under climate warming.
MATERIALS AND METHODS
Species and site details
Seeds of 10 perennial species (Table 1) were collected fromthe Stirling Range (34°22′ S; 118°15′ E; the major mountainsof southernWestern Australia. Six of the species are restrictedto peaks of the Stirling Range. The mountains are predomi-nantly sandstone and quartzite with thin layers of slate andphyllite (Semeniuk 1993) and the peaks of the range are
924 A. COCHRANE ET AL.
© 2010 The Authorsdoi:10.1111/j.1442-9993.2010.02211.xJournal compilation © 2010 Ecological Society of Australia
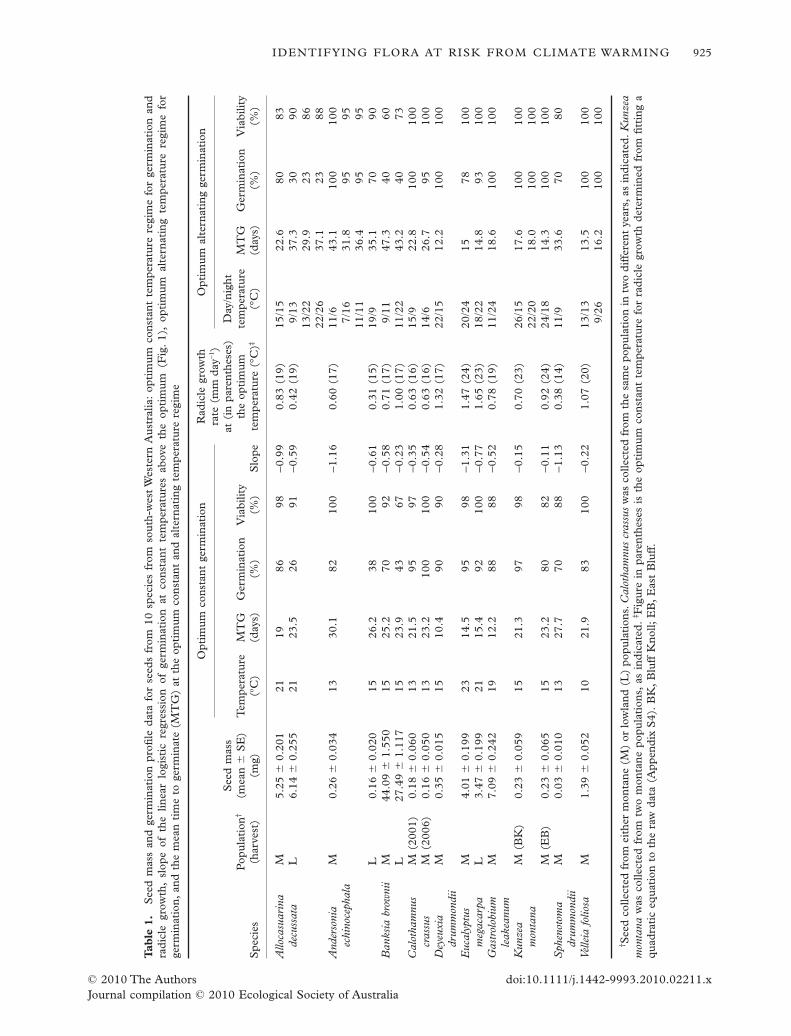
Tab
le1.
See
dm
ass
and
germ
inat
ion
profi
leda
tafo
rse
eds
from
10sp
ecie
sfr
omso
uth-
wes
tWes
tern
Aus
tral
ia:
opti
mum
cons
tant
tem
pera
ture
regi
me
for
germ
inat
ion
and
radi
cle
grow
th,
slop
eof
the
linea
rlo
gist
icre
gres
sion
ofge
rmin
atio
nat
cons
tant
tem
pera
ture
sab
ove
the
opti
mum
(Fig
.1)
,op
tim
umal
tern
atin
gte
mpe
ratu
rere
gim
efo
rge
rmin
atio
n,an
dth
em
ean
tim
eto
germ
inat
e(M
TG
)at
the
opti
mum
cons
tant
and
alte
rnat
ing
tem
pera
ture
regi
me
Spe
cies
Pop
ulat
ion†
(har
vest
)
See
dm
ass
(mea
n�
SE
)(m
g)
Opt
imum
cons
tant
germ
inat
ion
Rad
icle
grow
thra
te(m
mda
y-1)
at(i
npa
rent
hese
s)th
eop
tim
umte
mpe
ratu
re(°
C)‡
Opt
imum
alte
rnat
ing
germ
inat
ion
Tem
pera
ture
(o C)
MT
G(d
ays)
Ger
min
atio
n(%
)V
iabi
lity
(%)
Slo
pe
Day
/nig
htte
mpe
ratu
re(°
C)
MT
G(d
ays)
Ger
min
atio
n(%
)V
iabi
lity
(%)
Allo
casu
arin
ade
cuss
ata
M5.
25�
0.20
121
1986
98-0
.99
0.83
(19)
15/1
522
.680
83L
6.14
�0.
255
2123
.526
91-0
.59
0.42
(19)
9/13
13/2
222
/26
37.3
29.9
37.1
30 23 23
90 86 88A
nder
soni
aec
hino
ceph
ala
M0.
26�
0.03
413
30.1
8210
0-1
.16
0.60
(17)
11/6
7/16
11/1
1
43.1
31.8
36.4
100
95 95
100
95 95L
0.16
�0.
020
1526
.238
100
-0.6
10.
31(1
5)19
/935
.170
90B
anks
iabr
owni
iM
44.0
9�
1.55
015
25.2
7092
-0.5
80.
71(1
7)9/
1147
.340
60L
27.4
9�
1.11
715
23.9
4367
-0.2
31.
00(1
7)11
/22
43.2
4073
Cal
otha
mnu
scr
assu
sM
(200
1)0.
18�
0.06
013
21.5
9597
-0.3
50.
63(1
6)15
/922
.810
010
0M
(200
6)0.
16�
0.05
013
23.2
100
100
-0.5
40.
63(1
6)14
/626
.795
100
Dey
euxi
adr
umm
ondi
iM
0.35
�0.
015
1510
.490
90-0
.28
1.32
(17)
22/1
512
.210
010
0
Euc
alyp
tus
meg
acar
paM
4.01
�0.
199
2314
.595
98-1
.31
1.47
(24)
20/2
415
7810
0L
3.47
�0.
199
2115
.492
100
-0.7
71.
65(2
3)18
/22
14.8
9310
0G
astr
olob
ium
leak
eanu
mM
7.09
�0.
242
1912
.288
88-0
.52
0.78
(19)
11/2
418
.610
010
0
Kun
zea
mon
tana
M(B
K)
0.23
�0.
059
1521
.397
98-0
.15
0.70
(23)
26/1
522
/20
17.6
18.0
100
100
100
100
M(E
B)
0.23
�0.
065
1523
.280
82-0
.11
0.92
(24)
24/1
814
.310
010
0S
phen
otom
adr
umm
ondi
iM
0.03
�0.
010
1327
.770
88-1
.13
0.38
(14)
11/9
33.6
7080
Velle
iafo
liosa
M1.
39�
0.05
210
21.9
8310
0-0
.22
1.07
(20)
13/1
39/
2613
.516
.210
010
010
010
0
† See
dco
llect
edfr
omei
ther
mon
tane
(M)
orlo
wla
nd(L
)po
pula
tion
s.C
alot
ham
nus
cras
sus
was
colle
cted
from
the
sam
epo
pula
tion
intw
odi
ffer
ent
year
s,as
indi
cate
d.K
unze
am
onta
naw
asco
llect
edfr
omtw
om
onta
nepo
pula
tion
s,as
indi
cate
d.‡ F
igur
ein
pare
nthe
ses
isth
eop
tim
umco
nsta
ntte
mpe
ratu
refo
rra
dicl
egr
owth
dete
rmin
edfr
omfit
ting
aqu
adra
tic
equa
tion
toth
era
wda
ta(A
ppen
dix
S4)
.B
K,
Blu
ffK
noll;
EB
,E
ast
Blu
ff.
IDENTIFYING FLORA AT RISK FROM CLIMATE WARMING 925
© 2010 The Authors doi:10.1111/j.1442-9993.2010.02211.xJournal compilation © 2010 Ecological Society of Australia

more than 1400 km from the nearest similar mountain peaks.Consequently, these mountains are effectively islands in anotherwise largely flat landscape. The study species are allfound within the threatened thicket vegetation communitywhich grows on the upper levels of all the major peaks. Mostspecies rely on sexual reproduction for population mainte-nance; six of the 10 species rely on formation of a soil-storedseed bank, with the remaining four species holding seedreserves in the plant canopy (Appendix S1). All species gen-erally depend on disturbance (fire) for regeneration, an adap-tation for survival in fire-prone Mediterranean ecosystems.
While the Stirling Range experiences a typically Mediter-ranean climate, the mountains have a strong influence on thelocal environment. Average annual rainfall at the Park RangerStation (34°24′ S; 118°03′ E) is 551 mm. Rainfall averagesvary greatly with elevation, with the highest annual rainfall(about 1000 mm) estimated to be on the eastern peaks(Courtney 1993). The exposed higher peaks exhibit moreextreme temperatures with extended drizzle through summerand even occasional snow during winter. Day temperaturesare usually 5°C cooler on the peaks than on the plains (Tmax
27°C in January;Tmin 15°C in July) and the eastern peaks areoften shrouded with low cloud. Night-time temperatures areusually warmer on the peaks than on the plains (Tmax 13°C inJanuary; Tmin 6°C in July) as cold air drains down the slopes(Courtney 1993).
Three species (B. brownii, A. decussata and E. megacarpa)also occur outside the Stirling Range and seedlots fromcoastal populations, c. 100 km south of the Stirling Range,were included in this investigation. On the coast, the annualrainfall averages 929.8 mm (1877–2007) although for morerecent recording (1971–2007) averages just 875.5 mm. Rain-fall is concentrated mainly between May and September(autumn–winter). February and July are the hottest andcoolest months, respectively, with mean maximum tempera-tures of 22.9 and 15.4°C and mean minimum temperaturesof 15.7 and 8.1°C, respectively (Australian GovernmentBureau of Meteorology: Albany station 35°1′48′′S;117°52′48′′E; elevation 3 m).
Seeds were collected between December 2000 and April2007. Collections made prior to 2007 were dried at 15°C and15% relative humidity and stored at -20°C in sealed lami-nated foil bags until use in April 2007. Bags were allowed towarm to laboratory temperature (about 20°C) beforeopening. More recent collections were similarly dried, butnot stored at -20°C prior to experiment set-up.
Mean seed mass (after drying as above, then equilibratedto laboratory temperature, approximately 20°C) was deter-mined to the nearest mg by weighing 20 individual seeds ofBanksia brownii, Eucalyptus megacarpa, Velleia foliosa, Alloca-suarina decussata, Gastrolobium leakeanum and Deyeuxiadrummondii or five replicates of 10 seeds each, for thesmaller-seeded Calothamnus crassus, Andersonia echinocephala,Kunzea montana and Sphenotoma drummondii.
Experimental design
A bidirectional temperature gradient plate (Model GRD1,Grant Instruments, Cambridge, UK) was used to provideeither 12 constant temperatures or 30 alternating and sixconstant temperatures ranging from 6°C to 32°C. When
using the temperature gradient plate in one direction (12constant temperatures), two replicates of 30 seeds each wereused per collection per temperature, with the exception ofS. drummondii, G. leakeanum and A. echinocephala (lowlandpopulation; L) that had two replicates of 20 seeds each. Anincubator was used to obtain the lower limit of 5°C. Whenseed availability was limiting (because of the rare and threat-ened nature of some species), two replicates of 30 seeds (B.brownii), or three replicates of 10 seeds each (V. foliosa and D.drummondii), were placed in germination incubators at sixconstant temperatures (5, 10, 15, 20, 25 and 30°C). Subse-quently, the temperature gradient plate was used to obtain arange of diurnally alternating temperatures ranging fromapproximately 5°C to 35°C. Because of space constraints onthe temperature gradient plate, or seed availability, one50-mm Petri dish per temperature per collection contained10 (S. drummondii, A. echinocephala), 15 (B. brownii), 20 (K.montana, E. megacarpa, C. crassus, D. drummondii, G.leakeanum and A. echinocephala) or 30 seeds (V. foliosa and A.decussata).
Seeds were sown on the surface of 1% w/v water agar on50-mm Petri dishes except for seeds of K. montana that wereincubated on two layers of Whatman No. 2 filter paper overthick germination paper. The filter papers were moistenedwith 2 mL of 10-7 M butenolide solution (3-methyl-2Hfuro[2,3-c]pyran-2-one), the major germination stimulantin smoke; previous studies on K. montana germination haveindicated a requirement for smoke stimulation (Cochraneunpubl. data, May 2002). The butenolide used was isolated,purified and identified from smoke-saturated water derivedfrom burned Passerina vulgaris Thod. and Themeda triandraL. as described by van Staden et al. (2004). Other dormancy-breaking treatments used included the addition of gibberellicacid to the agar for S. drummondii (as GA3 at 25 mg L-1) andGA3 and butenolide (at 250 mL L-1) for A. echinocephala;these treatments have been found to produce optimal resultsfor those species (Cochrane unpubl. data, December 1998).Seeds of G. leakeanum have a water-impermeable seed coat(physical dormancy sensu Baskin & Baskin 2001) andrequired scarification of the seed coat prior to imbibition andradicle emergence. This was achieved by removing a smallportion of the seed coat with a scalpel along the longitudinalaxis of each seed. Actual temperatures experienced by seedson the temperature gradient plate were determined byplacing five bead thermistors connected to a data logger(Grant Instruments Squirrel 1000 Series) on the surface ofthe germination medium in the Petri dishes in each cornerand in the middle of the temperature gradient plate.
All environments received irradiance for 12 h day-1. Ger-mination was scored every other day and defined as visibleradicle emergence. Germination tests ran for up to 40 (con-stant temperatures) or 55 days (alternating temperatures).Non-germinated seeds were subjected to a cut-test to deter-mine potential viability; percentage germination was calcu-lated after subtracting the number of empty seeds from thenumber of seeds sown.
Radicle growth rates
To determine the temperature at which radicle growth exten-sion rate (RGER) was maximal, 12 germinated seedlings from
926 A. COCHRANE ET AL.
© 2010 The Authorsdoi:10.1111/j.1442-9993.2010.02211.xJournal compilation © 2010 Ecological Society of Australia

each collection germinated under constant temperature con-ditions were transferred to an agar plate held at the sametemperature and under identical light conditions either onthe temperature gradient plate or in incubators. Root lengthwas measured to the nearest mm with an eye piece reticulecalibrated with a stage micrometer under the microscope(small seedlings) or with vernier callipers (large seedlings)every 1–2 days for 5 days (at least three times per 5-dayperiod) and the mean radicle growth rate RGER calculated foreach temperature.
Statistical analysis and data presentation
Statistical analysis was carried out using GenStat 11.1 (VSNInternational Ltd., UK). T-tests of log10 transformed data (fornormality, confirmed by the Shapiro–Wilk test) were used tocompare the mass of seeds of the same species collected fromdifferent populations, or from the same population in differentyears. Mean time to germinate (MTG) was calculated for theoptimum constant and optimum alternating temperatureregime (highest percentage germination or most rapid if morethan one regime resulted in the same maximum percentagegermination) according to the equation:
MTG = ×( )∑ n d N
where: n = number of seeds germinated between scoringintervals, d = the incubation period in days at that time pointand N = total number of seeds germinated.
Binary logistic regression analysis (logit link function) wasused to assess the effect of increasing temperature above theoptimum (constant temperatures only) on the proportion ofseeds that germinated (linear model). Origin 6.1 (Microcal)was used to draw contour plots showing germination on thebidirectional temperature gradient plate. c2 tests (maximumlikelihood) were used for pairwise comparison of the propor-tion of seeds that germinated under the optimum constant oroptimum alternating temperature regimes within eachseedlot, or of the proportion of seeds that germinated fromseedlots collected from different populations within species.For each seedlot, the optimum and limiting temperatures forradicle growth were determined by fitting a quadratic rela-tionship to the raw data.
RESULTS
Seed mass
Seed mass ranged between 0.03 mg for S. drummondiiand 44.09 mg for the montane population of B.brownii(Table 1). Mean seed mass for A. decussata was signifi-cantly greater for seeds from the lowland populationscompared with seeds from the montane populations(P < 0.01). Conversely, for B. brownii, E. megacarpaand A. echinocephala, seeds from the montane popula-tions had significantly greater mean seed mass thanseeds from the lowland populations (P < 0.05). In thecase of C. crassus, mean seed mass was significantlygreater in 2001 compared with 2006 (P < 0.01).There
was no difference in the mean mass of seeds collectedfrom two different montane populations of K.montana.
Germination at constant temperatures
The optimum constant germination temperature (i.e.temperature where the proportion of seeds that germi-nated was greatest) ranged from 10°C to 23°C for the16 seedlots studied (Table 1; Appendix S2).There waslittle or no difference in the optimum germinationtemperature determined for seeds collected frommontane or lowland populations of A. decussata(21°C), A. echinocephala (13–15°C), B. brownii (15°C)and E. megacarpa (21–23°C), or from differentmontane populations of K. montana (15°C). Similarly,the optimum germination temperature for the older(2001) and more recent (2006) seedlots of C. crassusdid not differ (13°C).
The proportion of seeds that germinated at theoptimum germination temperature was generally high(>80%; Table 1; Appendix S2). However, there wassignificantly lower germination of seeds from thelowland populations of A. decussata, A. echinocephalaand B. brownii (�43%) compared with seeds of thesame species from the montane populations over thesame time period (P � 0.005).
For some seedlots, for example those of K. montana,there was a relatively wide range of constant tempera-tures where high levels of germination were attained(13–29°C; Appendix S2). However, for other seedlots,for example those of A. echinocephala and S. drummon-dii, germination was confined to a narrow rangeof temperatures (11–15°C; Appendix S2). Logisticregression analysis indicated significant relationshipsbetween percentage germination and temperature as itincreased above the optimum (P < 0.001; Fig. 1).Theslope of this relationship varied between species,ranging from -0.11 (K. montana; relatively small effectof increasing temperature above the optimum on per-centage germination) to -1.13 (S. drummondii; largeeffect of increasing temperature; Table 1).
Seeds placed at suboptimal temperatures wereslower to germinate. They often had a longer delaybefore germination commenced and a higher MTG,and generally exhibited lower total percentage germi-nation at the time when the tests were terminated(Appendix S2).
Seedling radicle growth rates and temperature
The quadratic relationship between radicle growthrate and temperature for each seedlot clearly showedhow radicle growth occurred over a broad range oftemperatures for some species, and a narrow range for
IDENTIFYING FLORA AT RISK FROM CLIMATE WARMING 927
© 2010 The Authors doi:10.1111/j.1442-9993.2010.02211.xJournal compilation © 2010 Ecological Society of Australia
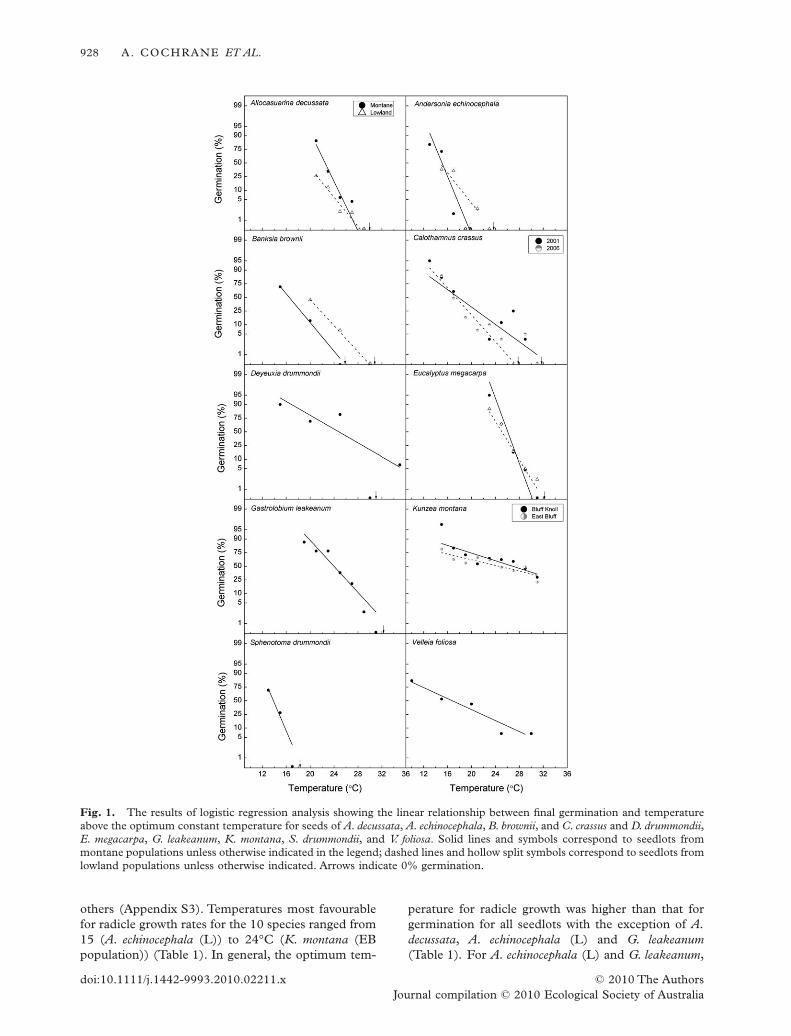
others (Appendix S3). Temperatures most favourablefor radicle growth rates for the 10 species ranged from15 (A. echinocephala (L)) to 24°C (K. montana (EBpopulation)) (Table 1). In general, the optimum tem-
perature for radicle growth was higher than that forgermination for all seedlots with the exception of A.decussata, A. echinocephala (L) and G. leakeanum(Table 1). For A. echinocephala (L) and G. leakeanum,
Fig. 1. The results of logistic regression analysis showing the linear relationship between final germination and temperatureabove the optimum constant temperature for seeds of A. decussata, A. echinocephala, B. brownii, and C. crassus and D. drummondii,E. megacarpa, G. leakeanum, K. montana, S. drummondii, and V. foliosa. Solid lines and symbols correspond to seedlots frommontane populations unless otherwise indicated in the legend; dashed lines and hollow split symbols correspond to seedlots fromlowland populations unless otherwise indicated. Arrows indicate 0% germination.
928 A. COCHRANE ET AL.
© 2010 The Authorsdoi:10.1111/j.1442-9993.2010.02211.xJournal compilation © 2010 Ecological Society of Australia

the temperature most favourable for radicle growth didnot differ from that for germination. For montanepopulations of both A. decussata and A. echinocephalaradicle growth rate was twice that of seedlings fromlowland populations at the optimum temperature. Formost species, the temperature at which radicle growthrate peaked showed little difference to that foroptimum germination. The seedlot with the greatestdifference between the optimum temperaturefor germination and that for radicle growth was V.foliosa. Radicle growth rate was fastest at 20°C(1.07 mm day-1) and relatively slow (0.49 mm day-1)at the temperature which produced optimum germi-nation (10°C).
Germination at alternating temperatures
The contour plots (Figs 2,3) show the germinationresponse of each seedlot across a range of both con-stant and alternating temperature regimes.The prefer-ence for low germination temperatures as seen in theconstant germination experiments for A.echinocephala,C. crassus, S. drummondii and V. foliosa is also apparentas ‘high’ isopleths (light filling) towards the bottom-left corner of the contour plots (Figs 2,3). In contrast,E. megacarpa seeds showed a preference for highergermination temperatures (Fig. 3) while D. drummon-dii, G. leakeanum, K. montana and V. foliosa germinatedacross a wide range of both the constant and alternat-ing bidirectional temperature gradient plate regimes(Fig. 3).
In general, the maximum proportion of seeds thatgerminated under a constant temperature regime(using incubators or the temperature gradient plateunidirectionally) was similar to the maximum propor-tion observed when the seeds were placed on the bidi-rectional temperature gradient plate (Figs 2,3), with afew exceptions (Table 1).There was greater maximumgermination of seeds from both populations of A. echi-nocephala when they were sown on the bidirectionaltemperature gradient plate. For seeds from themontane population, there was 100% germination at11/6°C compared with a maximum of 82% germina-tion at a constant 13°C (c2 = 6.89, P < 0.01). Forseeds from the lowland population, there was 70%germination at 19/9°C compared with 38% germina-tion at 15°C, although this difference was not statisti-cally significant (because of low seed numbers for thebidirectional temperature gradient plate experiment;P > 0.05). Seeds from both populations of K. montanaalso showed a preference for alternating temperatures,with a higher proportion of germinating seeds and afaster rate of germination (lower MTG; Fig. 3,Table 1). For seeds from the East Bluff population,the difference in germination between the optimumalternating temperature regime (24/18°C) and the
optimum constant temperature (15°C) when seedswere sown on the unidirectional temperature gradientplate, was significant (c2 = 7.59, P < 0.01). In contrastthere was significantly lower maximum germination ofB. brownii (M) seeds (c2 = 4.52, P < 0.05) and E.megacarpa (M) seeds (c2 = 6.81, P < 0.01) on the bidi-rectional temperature gradient plate compared withthe constant temperature regimes (incubators or uni-directional temperature gradient plate for B. browniiand E. megacarpa, respectively) (Table 1).
DISCUSSION
Predictions of changes in species geographic distribu-tion as a consequence of climate change assume thatplants survive within their current range because theexisting soil and climate conditions are optimal, andthat shifts in distribution may be predicted undervarious climate change scenarios when conditionschange (Thomas et al. 2004; Malcolm et al. 2006; Fitz-patrick et al. 2008). However, such modelling does notconsider vulnerability (likelihood of a decrease inrange) as a consequence of climate change at particu-lar stages in a plant life cycle, or the implications ofsuboptimal conditions on life stage transitions. As ger-mination and seedling establishment are the highest-risk phases for plants (Harper 1977; Silverton &Charlesworth 2001), germination temperature data,such as presented here, may help to identify speciesmost at risk in a changing climate.
Under projected scenarios, by 2030 there will bea 0.5–2.1°C increase in summer temperatures, a0.5–2.0°C increase in winter temperature and a2–20% decrease in rainfall across south-west WesternAustralia (IOCI 2009). A 2°C rise in temperature hasalready been shown to impact on seed responsesincluding seed production, seed mass, seedling emer-gence and establishment and soil seed bank dynamics(Williams et al. 2007; Hovenden et al. 2008; Hedhlyet al. 2009; Ooi et al. 2009). Changes in recruitmentcan lead to population decline and alter vegetationcommunity patterns (species composition and abun-dance), paving the way for species invasions (Parolo &Rossi 2008). Levels of recruitment may be affected bytotal germination and/or the rate of germination. Somespecies may increase these parameters as temperaturesrise, while in others they may be reduced (Williamset al. 2007; Milbau et al. 2009). Rising temperaturesmay reduce the rate of dormancy loss in seeds oftemperate or alpine species in the soil seed bank, par-ticularly those needing cold stratification before ger-mination occurs (Steadman & Pritchard 2003), or inarid zone species (Ooi et al. 2009). Perhaps species atmost risk will be those with complex dormancy whichneed to go through seasonal cycles (Lolium rigidum:Steadman et al. 2004).
IDENTIFYING FLORA AT RISK FROM CLIMATE WARMING 929
© 2010 The Authors doi:10.1111/j.1442-9993.2010.02211.xJournal compilation © 2010 Ecological Society of Australia

Fig. 2. Contour plots, with points of equal percentage germination connected by germination isopleths, for seeds of A.decussata, A. echinocephala, B. brownii and C. crassus.The shading calibration on the right hand side of the figure is for percentagegermination: light filling isopleths represents high germination, dark filling low or no germination. Seeds were collected fromeither montane or lowland populations, as indicated (Appendix S1). Constant temperatures are presented on the diagonal linefrom the bottom-left corner of the diagrams (lowest temperature approximately 6°C) to the top-right corner (maximumtemperature approximately 34°C). All points above and below the diagonal line represent alternating temperature regimes, withgreatest amplitude at the top-left and bottom-right corners of each graph.
930 A. COCHRANE ET AL.
© 2010 The Authorsdoi:10.1111/j.1442-9993.2010.02211.xJournal compilation © 2010 Ecological Society of Australia

Fig. 3. Contour plots, with points of equal percentage germination connected by germination isopleths, for seeds of D.drummondii, G. leakeanum, E. megacarpa, K. montana, S. drummondii and V. foliosa. Presentation is as in Figure 2.
IDENTIFYING FLORA AT RISK FROM CLIMATE WARMING 931
© 2010 The Authors doi:10.1111/j.1442-9993.2010.02211.xJournal compilation © 2010 Ecological Society of Australia

Plant traits, thermal constraints and thegermination niche
Here, we have considered first, whether or not thespecies relies on its seed for propagation and second,the temperature constraints for seed germination.First, our data show a relationship between seed massand altitude for three of the seedlot pairs investigated(B. brownii, A. echinocephala and E. megacarpa). Seedmass is considered a key influencing factor for seedsurvival and dispersal (Harper 1977). Altitude hasbeen shown to have an effect on seed mass with a trendfor lowland species to have smaller seeds than uplandspecies (Murray et al. 2003). As temperature andmoisture decrease with altitude, seed size increases.Three of the four small-seeded species (A. echi-nocephala, C. crassus and S. drummondii with <0.3 mgseed mass) exhibited a ‘typical’ Mediterranean germi-nation syndrome with low temperature requirementsfor germination (Bell & Bellairs 1992). Species withsmall seeds are reported to have narrower temperatureoptima for germination than larger seeded species(Bell et al. 1995) and often have more precise cues forgermination (Keeley 1991) possibly because they areespecially susceptible to drought (Daws et al. 2007;Jankowska-Blaszczuk & Daws 2007); high mean tem-perature or wide temperature alternations may coin-cide with risky periods for germination, when there islimited water availability. Of these three species, S.drummondii, displayed an extreme sensitivity to hightemperature. Seeds of this obligate seeding speciesonly germinated over a narrow range of cooler tem-peratures; increasing temperature above the optimumconstant temperature had a pronounced negativeeffect on germination, with an absence of germinationevident across a wide range of warmer temperatureconditions. Current winter temperatures on theStirling Range peaks with mean temperatures of 13(day) and 6°C (night) correspond to the observedgermination temperature optima. This already rarespecies may be at greatest risk of climate warmingcompared with the other species studied.
Similar to S. drummondii, the slope of the relation-ship between temperature above the optimum con-stant temperature and germination was high foranother obligate seeder, A. echinocephala and thisspecies also showed a preference for cooler tempera-tures for germination. However, there were someecotypic differences in the temperature profilesbetween seeds collected from montane and lowlandpopulations, with seeds from the lowland population,while more dormant, tending to germinate at highertemperatures (Fig. 2). Dormancy regulating chemicalssuch as the GA3 used to stimulate germination in bothS. drummondii and A. echinocephala may have inducedgermination over a wider temperature range thanwould have occurred in its absence, but temperature
ranges for germination in these species were nonethe-less narrow for these seedlots.
We found no indication that smaller seeds oflowland populations of A. decussata, A. echinocephalaand B. brownii had particularly different temperaturerequirements for germination than the larger seeds oftheir montane counterparts. Seeds from the lowlandpopulations of these species did show evidence ofslower germination, with �43% maximum germina-tion at constant temperatures after 28, 35 and 37 days,respectively. However, an alternating temperatureregime increased germination in A. echinocephala andthe results for seeds from the montane populationsuggest that higher maximum germination may havebeen observed with lower night temperatures thanachieved during this experiment. Ecotypic differencesin temperature optima have been observed for otherspecies. For example, Thurling (1966) reported thatseeds of Cardamine showed considerable difference ingermination response across an altitudinal gradientwith high temperature (>25°C) inhibition of germina-tion in lowland populations and low temperature inhi-bition (<10°C) in montane populations. Such speciesare clearly at less immediate risk, although specificpopulations (ecotypes) may decline. Species that haveflexibility in their regeneration strategy tend to havehigher germination temperature optima than obligateseeding species (Bell et al. 1995). For the speciesincluded in our study, the highest temperature optimafor germination were observed for seedlots from two ofthe species which are able to regenerate by resproutingas well as by seed, A. decussata (21°C) and E. megac-arpa (21–23°C).
The width of the germination niche for seeds of D.drummondii, G. leakeanum, K. montana and V. foliosawas greater than for either S. drummondii or A.echinocephala. The temperature tolerances of thesespecies may be wider than the climatic envelope theycurrently occupy, or recruitment events may occurwell into the warmer months when extended drizzleand precipitation from cloud cover may provide seedwith sufficient moisture for successful germination andseedling growth. While these species may have thephenotypic plasticity to survive some climatewarming, the fact that they already have a conservationlisting and/or are restricted to higher altitudes withinthe Stirling Range, suggests that despite the broadtolerance with respect to germination temperature,their niche breadth is determined by other matureplant traits like dispersal. Furthermore, these, andother species that are already restricted to higher eleva-tions, may face greater competition from species thatmigrate from lower altitudes as a consequence ofclimate warming (Gworek et al. 2006; Parolo & Rossi2008).
Our data suggest that germination may be a moresensitive life stage to climate warming than seedling
932 A. COCHRANE ET AL.
© 2010 The Authorsdoi:10.1111/j.1442-9993.2010.02211.xJournal compilation © 2010 Ecological Society of Australia

root growth, with temperatures for seedling growth atleast as high as for germination in most species. Inaddition, as soil temperature is a primary driver fordifferential selection of plant growth rates with coldsoils determining the growth limits of plants by inhib-iting root initiation (Kaspar & Bland 1992), warmingsoils may alter the competitive balance between seed-lings of different species.While there is some certaintyregarding increasing temperature and our ability topredict direct physiological changes on individualspecies, changes in other variables and their interac-tions that affect seed germination and establishmentsuch as rainfall, seasonality, fire and soil moistureremain uncertain (Williams et al. 2007; Berg et al.2010) and further studies that integrate these factorsare required.
Geographic range and germination niche
Our results did not support the hypothesis that germi-nation niche breadth corresponded to breadth of geo-graphic range: species with restricted ranges wereneither more, nor less, sensitive to increased tempera-tures associated with climate change than widespreadones. A lack of consistency in geographic range sizeand germination niche was also reported by Thomp-son et al. (1999) and Thompson and Ceriani (2003)for the British flora, and suggests that factors (bioticand/or abiotic) other than thermal tolerance mayconfine species like V. foliosa and G. leakeanum to theirmontane habitat. Their wide germination niche mayindicate significant phenotypic plasticity which mayimpact on their future persistence in a changingenvironment.
Climate manipulation experiments have demon-strated that some plant populations may be able toadapt to higher temperatures (Jump et al. 2008),although whether this is sufficient to ensure popula-tion survival is unknown. Adaptive capacity is likely tobe low in unique and isolated locations such as in theStirling Range (Pittock 2003).While gene flow will aidadaptation to warming climates in relatively contigu-ous populations, for isolated populations this would bemore limited and may lead to reduced levels ofclimate-related genetic variation rendering them lessable to adapt to any further changes (Jump & Peñuelas2005). Rare or restricted species may already sufferfrom depressed levels of genetic diversity that canreduce a populations’ ability to persist in a changingenvironment (Jump & Peñuelas 2005). Consequently,species lacking phenotypic (e.g. S. drummondii) orgenotypic (e.g. B. brownii: Sampson et al. 1994) plas-ticity in seed traits may experience stress or evenextinction during extended climate change. Delayedreproductive maturity will reduce the number of gen-erations that can establish from seed in any given time
period therefore long-lived woody perennials with longprimary juvenile periods such as B. brownii and specieswith low levels of fecundity are not expected toundergo rapid evolutionary adaptation (Jump &Peñuelas 2005). The importance of phenotypic plas-ticity and/or genetic variation will undoubtedly holdimportant insights into germination niche theory.
Plant life-cycle transition stages that link one genera-tion to the next, such as the germination-emergencetransition stage, are likely to be highly vulnerable toenvironmental change.Where basic knowledge of ger-mination limiting factors is lacking, tipping points orthresholds require identification in order to predict riskof species decline. In a broad sense, our study hashelped to define the bounds of the germination niche,particularly for geographically restricted species forwhich there is already some conservation concern.Wehave used a novel approach to identify species thatpossess conservative germination niches and hence, arepotentially susceptible to environmental change: with aslight upward shift in temperature, seeds of thesespecies simply will not germinate.
ACKNOWLEDGEMENTS
Funding for a research visit to the Millennium SeedBank, Royal Botanic Gardens, Kew, to conduct thisstudy was provided through the international seedconservation partnership between Kew and theWestern Australian Department of Environment andConservation. Financial support for the Kew seedbank was provided by the Millennium Commission,The Wellcome Trust and Orange plc. The RoyalBotanic Gardens, Kew is supported by grant-in-aidfrom Defra. We are grateful to a number of people forassistance with seed collection, including Todd Erick-son, Tony Friend and Sarah Barrett. We acknowledgethe comments of the editor and two reviewers whosesuggestions greatly improved this manuscript.
REFERENCES
Aitken S. N., Yeaman S., Holliday J. A., Wang T. & Curtis-McLane S. (2008) Adaptation, migration or extirpation:climate change outcomes for tree populations. Evol.Appl. 1,95–111.
Allen H. D. (2003) Response of past and present Mediterraneanecosystems to environmental change. Prog. Phys. Geogr. 27,359–77.
Baskin C. C. & Baskin J. M. (2001) Seeds: Ecology, Biogeography,and Evolution of Dormancy and Germination. AcademicPress, San Diego.
Bell D.T. (1994) Interaction of fire, temperature and light in thegermination response of 16 species from the Eucalyptus mar-ginata forest of south-westernWestern Australia. Aust. J.Bot.42, 501–9.
IDENTIFYING FLORA AT RISK FROM CLIMATE WARMING 933
© 2010 The Authors doi:10.1111/j.1442-9993.2010.02211.xJournal compilation © 2010 Ecological Society of Australia

Bell D. T. (1999) The process of germination in Australianspecies. Aust. J. Bot. 47, 475–517.
Bell D. T. (2001) Ecological response syndromes in the flora ofsouthwestern Western Australia: fire resprouters versusreseeders. Bot. Rev. 67, 417–40.
Bell D. T. & Bellairs S. M. (1992) Effects of temperature on thegermination of selected Australian native species used in therehabilitation of bauxite mining disturbance in WesternAustralia. Seed Sci.Technol. 20, 47–55.
Bell D. T., Rokich D. P., McChesney C. J. & Plummer J. A.(1995) Effects of temperature, light and gibberellic acid onthe germination of seeds of 43 species native to WesternAustralia. J.Veg. Sci. 6, 797–806.
Bellairs S. M. & Bell D. T. (1990) Temperature effects on theseed germination of ten kwongan species from Eneabba,Western Australia. Aust. J. Bot. 38, 451–8.
Berg M. P., Kiers E. T., Driessen G. et al. (2010) Adapt ordisperse: understanding species persistence in a changingworld. Glob. Change Biol. 16, 587–98.
Bond W. J. & Midgley J. J. (2001) Ecology of sprouting inwoody plants: the persistence niche. Trends Ecol. Evol. 16,45–51.
Bradshaw W. E. & Holzapfel C. M. (2008) Genetic response torapid climate change: it’s seasonal timing that matters. Mol.Ecol. 17, 157–66.
Broennimann O., Thuiller W., Hughes G., Midgley G. F., Alke-mades J. M. R. & Guisan A. (2006) Do geographic distribu-tion,niche property and life form explain plants’ vulnerabilityto global change? Glob. Change Biol. 12, 1079–93.
Courtney J. (1993) Climate. In: Mountains of Mystery. A NaturalHistory of the Stirling Range (eds C. Thompson, G. Hall & G.Friend) pp. 5–11. Department of Conservation and LandManagement, Perth.
Cowling R. M., Ojeda F., Lamont B. B. & Rundel P. W. (2004)Climate stability in Mediterranean-type ecosystems: impli-cations for the evolution and conservation of biodiversity.MEDECOS 10th Conference. Rhodes, Greece.
Daws M. I., Ballard C., Mullins C. E. et al. (2007) Allometricrelationships between seed mass and seedling characteristicsreveal trade-offs for neotropical gap-dependent species.Oecologia 154, 445–54.
Donohue K., Rubio de Casas R., Burghardt L., Kovach K. &Willis C. (2010) Germination, postgermination adaptation,and species ecological ranges. Annu. Rev. Ecol. Evol. Syst. 41,in press.
Fenner M. & Thompson K. (2005) The Ecology of Seeds. Cam-bridge University Press, Cambridge.
Fitzpatrick M., Gove A. D., Sanders N. J. & Dunn R. R. (2008)Climate change, plant migration and range collapse in aglobal biodiversity hotspot: the Banksia (Proteaceae) ofWestern Australia. Glob. Change Biol. 4, 1337–52.
Gworek J. R., VanderWall S. B. & Brussard P. F. (2006) Changesin biotic interactions and climate determine recruitment ofJeffrey pine along an elevation gradient. For. Ecol. Manage.239, 57–68.
Hannah L., Midgley G., Hughes G. & Bomhard B. (2005) Theview from the Cape: extinction risk, protected areas andclimate change. Bioscience 55, 231–42.
Harper J. L. (1977) Population Biology of Plants. Academic Press,New York.
Hedhly A., Hormaza J. I. & Herrero M. (2009) Global warmingand sexual plant reproduction. Trends Plant Sci. 14, 30–6.
Hope P. K., Drosdowsky W. & Nicholls N. (2006) Shifts in thesynoptic systems influencing southwest Western Australia.Clim. Dyn. 26, 751–64.
Hovenden M. J., Wills K. E., Chaplin R. E. et al. (2008)Warming and elevated CO2 affect the relationship betweenseed mass, germinability and seedling growth in Austrodan-thonia caespitosa, a dominant Australian grass. Glob. ChangeBiol. 14, 1633–41.
Hughes L., Cawsey E. M. & Westoby M. (1996) Climatic rangesizes of eucalyptus species in relation to future climatechange. Glob. Ecol. Biogeogr. Lett. 5, 23–9.
IOCI (2009) Indian ocean climate change initiative. [CitedJanuary 2010.] Available from URL: http://www.ioci.org.au
Jankowska-Blaszczuk M. & Daws M. I. (2007) Impact of red : farred ratios on germination of temperate forest herbs in rela-tion to shade tolerance, seed mass and persistence in thesoil. Funct. Ecol. 21, 1055–62.
Jump A. S. & Peñuelas J. (2005) Running to stand still: adapta-tion and the response of plants to rapid climate change. Ecol.Lett. 8, 1010–20.
Jump A. S., Peñuelas J., Rico L. et al. (2008) Simulated climatechange provokes rapid genetic change in the Mediterraneanshrub Fumana thymifolia. Glob. Change Biol. 14, 637–43.
Kaspar T. C. & Bland W. L. (1992) Soil temperature and rootgrowth. Soil Science. 154, 290- 9.
Keeley J. E. (1991) Seed germination and life history syndromesin the California chaparral. Bot. Rev. 57, 81–116.
Lloret F., Peñuelas J. & Estiarte M. (2004) Experimental evi-dence of reduced diversity of seedlings due to climate modi-fication in a Mediterranean-type community. Glob. ChangeBiol. 10, 248–58.
Malcolm J. R., Liu C., Neilson R. P., Hansen L. & Hannah L.(2006) Global warming and extinctions of endemic speciesfrom Biodiversity Hotspots. Conserv. Biol. 20, 538–48.
Meyer S. E., Allen P. S. & Beckstead J. (1997) Seed germinationin Bromus tectorum and its ecological significance. Oikos 78,475–85.
Midgley G. F., Hannah L., Millar D., Rutherford M. C. & PowrieL. W. (2002) Assessing the vulnerability of species richnessto anthropogenic climate change in a biodiversity hotspot.Glob. Ecol. Biogeogr. 11, 445–51.
Milbau A., Graae B. J., Shevtsova A. & Nijs I. (2009) Effects ofa warmer climate on seed germination in the subarctic. Ann.Bot. 104, 287–96.
Mott J. J. (1972) Germination studies on some annual speciesfrom an arid region of Western Australia. J. Ecol. 60, 293–304.
Murray B. R., Brown A. H. D. & Grace J. P. (2003) Geographicgradients in seed size among and within perennial AustralianGlycine species. Aust. J. Bot. 51, 47–56.
Ooi M. K. J., Auld T. D. & Denham A. J. (2009) Climate changeand bet-hedging: interactions between increased soil tem-peratures and seed bank persistence. Glob. Change Biol. 15,2375–86.
Paczkowska G. & Chapman A. (2000) The Western AustralianFlora: A Descriptive Catalogue. Wildflower Society WesternAustralia, Perth.
Parmesan C. & Yohe G. (2003) A globally coherent fingerprintof climate change impacts across natural systems. Nature421, 37–42.
Parolo G. & Rossi G. (2008) Upward migration of vascularplants following a climate warming trend in the Alps. BasicAppl. Ecol. 9, 100–7.
Pauli H., Gottfried M., Dirnbock T. Dullinger S. & Grabherr G.(2003) Assessing the long-term dynamics of endemic plantsat summit habitats. In: Alpine Biodiversity in Europe (eds L.Nagy, G. Grabherr, C. Horner & D. B. A. Thompson)pp. 195–207. Springer-Verlag, Berlin.
934 A. COCHRANE ET AL.
© 2010 The Authorsdoi:10.1111/j.1442-9993.2010.02211.xJournal compilation © 2010 Ecological Society of Australia

Pitman A. J., Narisma G. T. & McAneney J. (2007) The impactof climate change on the risk of forest and grassland fires inAustralia. Clim. Change 84, 383–401.
Pittock B., ed. (2003) Climate Change: An Australian Guide to theScience and Potential Impacts. Australian Greenhouse Office,Canberra.
Pouliquen-Young O. & Newman P. (2000) The Implications ofClimate Change for Landbased Nature Conservation Strategies.Final Report 96/1306. Australian Greenhouse Office, Envi-ronment Australia & Murdoch University, Canberra &Perth.
Probert R. J. (2000) The role of temperature in the regulation ofseed dormancy and germination. In: Seeds. The Ecology ofRegeneration in Plant Communities, 2nd edn (ed. M. Fenner)pp. 261–92. CAB International, Wallington, Oxon UK.
Root T. L., Price J. T., Hall K. R., Schneider S. H., RosenzweigC. & Pounds J. A. (2003) Fingerprints of global warming onwild animals and plants. Nature 421, 57–60.
Sampson J. F., Collins B. G. & Coates D. J. (1994) Mixed matingin Banksia brownii Baxter ex R. Br. (Proteaceae). Aust. J.Bot.42, 103–11.
Semeniuk V. (1993) Geology, landforms, soils and hydrology. In:Mountains of Mystery. A Natural History of the Stirling Range(eds C. Thompson, G. Hall & G. Friend) pp. 13–26.Department of Conservation and Land Management,Perth.
Silverton J. & Charlesworth D. (2001) In: Introduction to PlantPopulation Biology, 4th edn. Blackwell Science, Oxford.
van Staden J., Jäger A., Light M. & Burger B. (2004) Isolation ofthe major germination cue from plant-derived smoke. S.Afr.J. Bot. 70, 654–9.
Steadman K. J., Ellery A. J., Chapman R., Moore A. & Turner N.C. (2004) Maturation temperature and rainfall influenceseed dormancy characteristics of annual ryegrass (Loliumrigidum). Aust. J. Agric. Res. 55, 1047–57.
Steadman K. J. & Pritchard H. W. (2003) Germination ofAesculus hippocastanum seeds following cold-induced dor-mancy loss can be described in relation to a temperature-
dependent reduction in base temperature (Tb) and thermaltime. New Phytol. 161, 415–25.
Thomas C. D., Cameron A., Green R. E. et al. (2004) Extinctionrisk from climate change. Nature 427, 145–8.
Thompson K. & Ceriani R. M. (2003) No relationship betweenrange size and germination niche width in the UK herba-ceous flora. Funct. Ecol. 17, 335–9.
Thompson K., Gaston K. J. & Band S. R. (1999) Range size,dispersal and niche breadth in the herbaceous flora ofcentral England. J. Ecol. 87, 150–5.
Thuiller W., Lavorel S. & Araújo M. B. (2005) Niche propertiesand geographical extent as predictors of species sensitivity toclimate change. Glob. Ecol. Biogeogr. 14, 347–57.
Thurling N. (1966) Population differentiation in AustralianCardamine. III. Variation in germination response. Aust. J.Bot. 14, 189–94.
Walther G.-R., Post E., Convey P. et al. (2002) Ecologicalresponses to recent climate change. Nature 416, 389–95.
Williams A. L., Wills K. E., Janes J. K., Vander Schoor J. K.,Newton P. C. D. & Hovenden M. J. (2007) Warming andfree-air CO2 enrichment alter demographics in fourco-occurring grassland species. New Phytol. 176, 365–74.
SUPPORTING INFORMATION
Additional Supporting Information may be found inthe online version of this article:
Appendix S1. Collection data for 10 south-westWestern Australian species used in the study.Appendix S2. Germination progress curves at con-stant temperatures for seeds of different seedlots.Appendix S3. The quadratic relationship betweenradicle growth rate and temperature at constant tem-peratures for each seedlot.
IDENTIFYING FLORA AT RISK FROM CLIMATE WARMING 935
© 2010 The Authors doi:10.1111/j.1442-9993.2010.02211.xJournal compilation © 2010 Ecological Society of Australia




















