
Atlantis, Alien Visitation, and Genetic Manipulation - CiteSeerX
Upload
independentCategory
view
2download
0
Research Article
Seasonal Variation in the Physiologicaland Behavioral Responses to TouristVisitation in Magellanic Penguins
CECILIA VILLANUEVA,1 Centro Nacional Patagonico, CONICET, Boulevard Brown 2915, Puerto Madryn 9120, Chubut, Argentina; andUniversidad Nacional del Comahue, Quintral 1250, Bariloche 8400, Rıo Negro, Argentina
BRIAN GEORGE WALKER, Biology Department, Fairfield University, 1073 North Benson Road, Fairfield, CT 06824, USA
MARCELO BERTELLOTTI, Centro Nacional Patagonico, CONICET, Boulevard Brown 2915, Puerto Madryn 9120, Chubut, Argentina
ABSTRACT Penguin colonies are highly visited worldwide. Although several studies have addressed howpenguins behaviorally respond to tourist visitation at a point in time, nothing is known about their responseacross the entire breeding season. Furthermore, behavioral responses are driven by complex physiologicalprocesses and the basal physiological state of the individual might affect the way it responds to stimuli. To testthe hypothesis that annual changes in corticosterone result from animals having different requirements forexpressing (or not) the glucocorticoid-mediated behaviors at different times of the year in the context oftourist visitation, we examined circulating and stress-induced corticosterone in Magellanic penguins(Spheniscus magellanicus) from non-visited areas from the San Lorenzo colony, Peninsula Valdes, Argentinaacross the breeding season. We also examined the behavioral responses of penguins to a pedestrian approachin tourist-visited and non-visited areas of the colony across the season. Our results showed that circulatinglevels of corticosterone did not vary across the season; however, stress-induced corticosterone was lowestduring molt. Our behavioral results showed that penguins displayed different behavior responses at varyingdistances depending on the area (visited or non-visited) and stage in the season. Penguins in the tourist areawere more tolerant to a human approach than penguins in the non-tourist area. During settlement and molt,penguins showed higher occurrence of behaviors related to self-survival (such as standing, moving fartherinto the nest, and fleeing), whereas during incubation and chick rearing, penguins displayedmostly a behaviorassociated with defense and vigilance (such as alternate head turns). Furthermore, penguins allowed a closerapproach during incubation, but elicited a subsequent behavior quicker than in the rest of the stages,suggesting that they would be particularly sensitive in this stage. Overall, our results suggest thatcorticosterone release across the season is more associated with penguins’ survival in an extreme environmentthan with behavioral regulation. From a conservation perspective, we identified that penguins were moresensitive to human approach during incubation, but also molt should be considered as a vulnerable stagebecause corticosterone secretion is suppressed. � 2014 The Wildlife Society.
KEY WORDS anthropogenic disturbance, behavior, corticosterone, Magellanic penguin, Peninsula Valdes, Spheniscusmagellanicus, tourist management.
Ecotourism is seen as an ecologically friendly activity thatpromotes the preservation of species but also generatesincome for local communities (Bookbinder et al. 1998).Penguins are popular and charismatic birds, and attract theattention of thousands of tourists every year in most oftheir breeding areas (Boersma 2008). Tourist numbers areincreasing in penguin breeding colonies and there is pressureto make more sites available to tourists (Yorio et al. 2001,Seddon and Ellenberg 2008). Therefore, understanding howpenguins respond to human visitation is important to detectpotential negative effects and allow the development ofecotourism in a sustainable way.
One of the most common measures used to assess penguinresponse to visitation has been the study of their behavior(van Heezik and Seddon 1990, Burger and Gochfeld 2007,Holmes 2007). However, behavioral responses in animals aremediated by a series of complex physiological processes(Beale and Monaghan 2004). In addition, the physiologicalstate of an individual might vary within the stages of thebreeding season (Wingfield et al. 1992, Romero et al. 1998).For example, several studies have found that hormoneconcentrations such as corticosterone, a stress hormoneinvolved in preparing an individual to live through stressfulsituations (Sapolsky et al. 2000), vary daily and annually,related to circadian rhythms and life-history cycles (Alberset al. 1985, Atkinson and Waddell 1997). Althoughnumerous avian studies have examined seasonal variabilityin corticosterone levels (reviewed in Romero 2002), relativelylittle is known about how corticosterone concentrations vary
Received: 3 September 2012; Accepted: 28 July 2014Published: 13 October 2014
1E-mail: [email protected], [email protected]
The Journal of Wildlife Management 78(8):1466–1476; 2014; DOI: 10.1002/jwmg.791
1466 The Journal of Wildlife Management � 78(8)

seasonally in penguins (except see Cherel et al. 1988a, b;Otsuka et al. 1998; Vleck et al. 2000) or how hormones affectbehavior (Fowler 1999, Thierry et al. 2014).In attempting to explain the annual variation in cortico-
sterone release, the behavioral hypothesis proposes thatannual changes in corticosterone result from animals havingdifferent requirements for expressing (or not) the glucocor-ticoid-mediated behaviors along the year (Romero 2002).For example, fasting king penguins (Aptenodytes patagonicus)maintain low levels of corticosterone (Cherel et al. 1988a, b)during the start of the breeding stage and molt, when theyneed to stay inland. Corticosterone increases when theenergy reserves are depleted or new plumage is fully grownand penguins are ready to go to the ocean. Also, high levelsof corticosterone have been associated with nest desertion(Groscolas et al. 2008). Furthermore, an experimentalincrease of corticosterone in female macaroni penguins(Eudyptes chrysolophus) during chick rearing increasedforaging effort and chick mass (Crossin et al. 2012). Thesefindings suggest that corticosterone would be involved intriggering the foraging behavior.The corticosterone stress response has been studied
previously in Magellanic penguins (Spheniscus magellanicus;Hood et al. 1998, Fowler 1999, Walker et al. 2006).However, these studies have not considered the wholereproductive season. We were interested in studying thecirculating and stress-induced corticosterone levels inMagellanic penguins under the hypothesis that Magellanicpenguins would show a seasonal variation in corticosteronerelease. We predicted that circulating corticosterone levelswould be high during settlement because in this stage,birds have just arrived from migration and are highly activein rebuilding and defending their nests, and mating. We alsoexpected that circulating corticosterone would decreaseduring incubation to avoid nest abandonment, and increaseduring chick rearing to promote foraging behavior. However,stress-induced corticosterone would be high because birdsduring settlement, incubation, and chick rearing would beable to elicit a robust stress response in times of necessity.During molt, both circulating and stress-induced cortico-sterone levels would be low because high levels of thishormone have been associated with the production of poorquality feathers in other bird species (Romero et al. 2005,DesRochers et al. 2009). Also, penguins lose insulation whenmolting; therefore, if increasing corticosterone triggers theforaging behavior, having low levels of corticosterone duringmolt would be beneficial because they would not be likely togo to sea to forage with insufficient insulation.Our second hypothesis was that the seasonal variation in
corticosterone levels would influence penguin behavior.Therefore, in the context of tourist visitation, penguinswould behave differently depending on the life-history stage.Although several studies have measured the behavioralresponse of penguins to human disturbance at a point in time(van Heezik and Seddon 1990, Fowler 1999, Holmes 2007),only a few studies have addressed how behavioral responseschange over multiple points in a breeding season (Culiket al. 1990, Yorio and Boersma 1992, Cevasco et al. 2001).
To our knowledge, no studies have examined changes acrossan entire breeding season. Because tourist visitation occursduring the whole season, it is important to know if there areperiods when the birds are more vulnerable. Based on aprevious study (Villanueva et al. 2012), we predicted thatpenguins in an area with a history of tourist visitation wouldshow a reduced behavioral response to a pedestrian approachcompared to penguins in a non-visited area of the colonybecause of habituation of the former to tourist visitation. Wealso predicted that during settlement, penguins would behighly alert of their surroundings and be more prompted tostand, hide in the nest, or flee, because they had not investedin reproduction yet and would display behaviors related toprotecting themselves. During incubation, we expectedpenguins to be less alert to their surroundings (i.e., shorteralert distance and fewer occurrences of alert behavior), wouldallow a closer approach, and would display a defensive orvigilant behavior more frequently (i.e., alternate head turns)but would not flee because they had invested in reproduction(coinciding also with low circulating corticosterone). Duringchick rearing, we expected birds to be more alert to theirsurroundings (i.e., greater alert distance than incubation andgreater occurrence of alert behavior, coinciding with higherlevels of circulating corticosterone) and also be very defensiveof the nest and chicks (i.e., greater occurrence of head turnsand at greater distances) but still would not flee. During moltwhen penguins do not have dependent offspring to protectbut cannot escape to the ocean because they have lostinsulation, we predicted that birds would be less alert to theirsurroundings and would elicit a behavioral response relatedto self-preservation (i.e., standing and running into the nest)but would not flee (also coinciding with low levels ofcirculating and stress-induced corticosterone).In summary, the goals of this study were to determine if any
seasonal change in corticosterone release might be associatedwith changes in the behavioral response to visitation over thebreeding season in Magellanic penguins at Estancia SanLorenzo, Peninsula Valdes, Argentina; and to identify ifthere were periods when penguins were more susceptible todisturbance. We measured corticosterone concentrations(both circulating and stress-induced) in non-tourist visitedpenguins to investigate if corticosterone release varied acrossthe breeding season. We also measured behavioral responsesof penguins to a standardized pedestrian approach in tourist-visited and non-visited areas across the breeding season.
STUDY AREA
We carried out this study in a Magellanic penguin colonylocated in Estancia San Lorenzo (428 050S, 638 490W),Peninsula Valdes, Argentina. The colony was approximately100 ha in area, stretched along 4 km of coastline, andextended 800m inland at its widest point. The predominantvegetation was mainly low shrubs (i.e., Schinus johnstonii,Lycium chilense, Chuquiraga avellanedae; Daciuk 1977).Penguins can nest under shrubs, on scrapes, or in burrows,all of which vary in their amount of cover (Williams andBoersma 1995). For this study, we chose birds using all 3types of nests but made sure each nest allowed us to see the
Villanueva et al. � Seasonal Response to Visitation in Penguins 1467

owner properly, so as to register the behavioral responsesaccurately. The most recent published survey (Schiaviniet al. 2005) estimated approximately 55,000 breeding pairs inthe colony, but it has continued to grow since then, and wasthe largest colony located on Peninsula Valdes. Touristvisitation started in September of 2000, and was wellmonitored and controlled, with tourists restricted to a verysmall area of the colony, on a trail with well-defined,fenced-off edges. Currently, San Lorenzo receives around10,000 tourists seasonally (M. Machinea, Estancia’s owner,personal communication).We studied penguins nesting on or within 2–3m of the fenced
edge of the tourist trail (behavioral study) and penguins nestingin an area of the colony where no tourists visit (physiological andbehavioral studies). The non-tourist area is approximately 600mfrom the tourist trail. Both areas had similar habitat character-istics and were equidistant from the sea.
METHODS
Penguin PhenologyThe reproductive season starts in September when malepenguins arrive to the colony to rebuild their nests. Femalepenguins arrive 2 to 3 weeks later, and courtship and matingcommence. We refer to this initial stage as settlement. In thefirst week of October, females start laying their eggs; theincubation stage then lasts 38–41 days, and both members ofthe pair alternate time on the eggs. Chicks start hatchingaround the second week of November. The chick-broodingstage lasts 90–110 days, and it can be divided into 2 stages:early chick, when 1 parent continually remains in the nestbrooding the thermally dependent chicks, and late chick,when the chicks are thermally independent and can be leftalone in the nest while the parents forage at sea. Both parentsshare parental activities. Chicks start fledging at the end ofJanuary and by mid-February all the chicks have gone to thesea. Adults start molting in March, after a period of timespent re-fattening after their chicks have fledged. The moltcan last up to 3 weeks, in which time birds do not forage andstay on land in or very near their nests. At the end of Marchor early April, all penguins return to the ocean and migratenorth, spending the winter foraging at sea (Williams andBoersma 1995, Falabella et al. 2009).
Corticosterone ResponseCorticosterone is involved in several physiological mecha-nisms related to dealing with a stressor. Increased secretionof this hormone helps redirect behaviors and activatesphysiological survival mechanisms, such as elevation ofcirculating glucose and blood pressure, and stimulation oflipolysis (Wingfield 1994, Sapolsky et al. 2000). Wemeasured circulating levels of corticosterone (baseline) andthe stress response of penguins to a novel stressor (stress-induced corticosterone) during incubation, early chick, latechick, and molt in the breeding season 2008–2009. We didnot take blood samples from birds during settlement becausewe were concerned that they would abandon the nestpermanently because of the disturbance. We sampledcorticosterone from birds in the non-tourist area only
because in a previous study we found no differences in thestress response (both baseline and stress-induced corticoste-rone levels) in tourist-visited versus non-visited birds in thesame colony (Villanueva et al. 2012). Because differencesbetween sexes have been documented in stress physiologyof Magellanic penguins (Hood et al. 1998, Villanuevaet al. 2012), and there are previous data for Magellanicpenguin males throughout the season (albeit from a differentcolony; B.G. Walker, Fairfield University, unpublished data;Walker et al. 2004), we chose to only include females in thisstudy.We followed a standard capture-restraint protocol used in
field studies (Cockrem et al. 2006, Romero et al. 2006). Wecaptured the bird in the nest and took 4 consecutive bloodsamples. We collected the first sample (i.e., baseline) within3 minutes of capture because corticosterone increases after3 minutes in response to the stress of capture (Romero andReed 2005). We collected the remaining samples at 10,30, and 45minutes post-capture. We used venipunture ofthe interdigitary (food webbing) vein to collect the samples(75–100ml) into heparinized microcapillary tubes. Duringthe 45-minute period, the researcher restrained penguins intheir lap, and we covered the penguins’ eyes with a cloth todecrease their startle response to movement around them.During incubation, we removed the eggs from the nest tokeep them warm during the restraint period. We returnedthe eggs after the bird had been released back in the nest. Allanimal handling and experimental procedures were approvedby the Office of Tourism and Protected Areas from Chubutprovince and Fauna and Flora Department, Argentina andFairfield Animal Care Committee (FACC, IACUC),Fairfield University, Fairfield CT, USA, under permit00-014. We obtained export and import permits for bio-logical samples from both Argentine and U.S. governmentagencies.We kept blood samples in a cooler up to 6 hours in the field,
and then we centrifuged them for 10minutes to separatethe plasma back at camp. We froze the resulting plasma at�208C and transferred it frozen to the laboratory foranalysis. We collected all blood samples early in the day(from 0700 to 1230), to minimize any potential natural dailyvariation in corticosterone concentration. After the restraintperiod, we took body measurements to determine the sex ofthe birds (Bertellotti et al. 2002). Finally, we marked birdswith small numbered tags in the webbing of the left foot andreleased them into their nests. We did not sample any birdsmore than once during the study (i.e., within the breedingseason).We determined plasma corticosterone concentrations
following a modified direct radioimmunoassay (RIA) formatwithout chromatography (Wingfield and Farner 1975).Validation of the RIA using penguin plasma was conductedin a previous study (Walker et al. 2004). We usedCorticosterone Antibody B3-163 (Esoterix, Calabasas,CA). The sensitivity of the assay was 0.50 ng/mL. Weassayed each sample in duplicate after extraction with freshlydistilled dichloromethane. We ran all samples in 2 assays.Intra-assay variation was 3.6%, calculated as the average
1468 The Journal of Wildlife Management � 78(8)

coefficient of variation between all of the duplicate samples inthe assay. Inter-assay variation was 12.3%, calculated from aplasma pool sample, which we ran in both assays.
Behavioral Response to VisitationWe carried out a standardized human-disturbance experi-ment on penguins nesting in both tourist-visited and non-visited areas of the colony at 5 stages during the reproductiveseason: settlement, incubation, early chick, late chick, andmolt, during 2007–2008. The experiment consisted of asingle person approaching a nesting penguin in a straight lineat a constant and slow pace until the person was 0.2m awayfrom the bird, simulating a responsible tourist. We recordedwhat behaviors the bird elicited in response to the approach,as well as the distance from the visitor that the bird initiatedthe particular behavior. We classified the behaviors as inYorio and Boersma (1992), with the addition of a categorywe called into the nest. The behaviors are defined as 1) alert:the bird noticed and watched the approaching person; 2)alternate head turns: the bird rotated its head from side toside, a behavior that has been described as defense (Eggletonand Siegfried 1977) and vigilance (Holmes et al. 2006); 3)standing: a lying bird stood up; 4) into the nest: a bird on theboundary of the nest entered the nest or if already on the nestwent farther in for more protection; and 5) flee: the birdleft the nest. The approaching person signaled changingbehaviors by dropping sticks on the ground during theapproach, and measured distances from the nest to the sticksafter the completion of the entire experimental approach.The greater the distance of a particular response indicated amore rapid, or less tolerable response by the focal bird. Allpenguins had not seen the approaching person before theapproach started and were lying on the nest or its boundary.The same person performed all approaches to maintainconsistency. We marked the nests with tape after theexperience to avoid resampling the same birds.
AnalysisWe tested normality with Shapiro–Wilks tests, andhomoscedasticity with Bartlett’s tests. We carried outTukey’s post hoc tests for comparisons among stagesconsidering the areas together. We performed all statisticalanalyzes in R (Version 2.13.0., http://www.r-project.org,accessed 9 Aug 2011). We considered P� 0.05 to bestatistically significant.Physiological responses.—We compared the seasonal varia-
tion in baseline (first sample after capture) and totalcorticosterone levels over the restraint period. We calculatedtotal corticosterone levels using the arithmetic trapezoid rule,which calculates the area under the corticosterone stressresponse curve for each penguin (Walker et al. 2005).We logtransformed baseline corticosterone levels because of lack ofnormality, and ran an analysis of variance (ANOVA) forbaseline and total corticosterone levels with stage (incuba-tion, early chick, late chick, or molt) as the main factor.Behavioral responses.—We analyzed behavior data using
general linear models (GLM), and assessed the significanceof the terms in the models by likelihood ratio tests (LRT;Hilborn and Mangel 1997) following a backward stepwise
procedure from a complex model to simpler ones. Thesignificance of the terms in the models was assessed bylooking at the changes in deviance; if after removing a termthe increase in the deviance was statistically significant(assessed by a chi-square test), we considered the termimportant and added it back into the model.First, we compared the frequency of occurrence of
displayed behaviors between areas and stages. We fit aGLM with a binomial distribution and a logit link functionfor proportional data (Crawley 2007). The response variablein the full model was a matched pair of counts of the numberof birds that elicited (successes) and the number of birds thatdid not elicit (failures) a certain behavior from the totalnumber of birds approached in each stage (in practice theresponse variable is a matrix comprising the number ofsuccesses and failures). The explanatory variables in themodel were type of behavior, area, stages, and the 2-wayinteraction terms.We next wanted to compare the distance at which penguins
elicited the behaviors between areas and stages. Becausesome behaviors had low occurrence, we were able to completethis analysis for only the following behaviors: alert and headturns. The start distance varied because we chose the targetbird in an opportunistic way to ensure the bird did not see theresearcher prior to initiation of the approach. Start distancewas also affected by local topography (e.g., bushes, fences).Therefore, we included start distance as another term in themodel to control for overall start distance variation. We fit 2different full GLMs for the response variables of alertdistance and head turns distance, applying a gammadistribution and the inverse as the link function. Theexplanatory variables were area, stage, start distance, and the2-way interaction terms.Finally, we calculated the difference between the initial
alert distance and distance at which the bird presented thesubsequent behavior to measure how tolerant a bird was to anapproaching person. We defined this distance as reactiondistance and it was the response variable in the models. Alonger reaction distance (which resulted in the approachingperson being closer to the bird) represented greater toleranceto the approaching person; in contrast, a short reactiondistance represented low tolerance. We modeled reactiondistances as functions of area, stage, start distance, andthe 2-way interaction terms using GLM with a Gammadistribution and inverse link function.
RESULTS
Physiological ResponsesCirculating corticosterone levels in female Magellanicpenguins were similar across stages (F3, 44¼ 2.65,P¼ 0.06; Fig. 1). Corticosterone concentrations increasedsignificantly over the time of restraint during all stages(Fig. 1). Corticosterone release reached its maximum around30minutes after first extraction during incubation and chickrearing; however, during molt it did not saturate in blood,even after 45minutes. Stress-induced corticosterone levels(the area under the stress-series curve) were different across
Villanueva et al. � Seasonal Response to Visitation in Penguins 1469
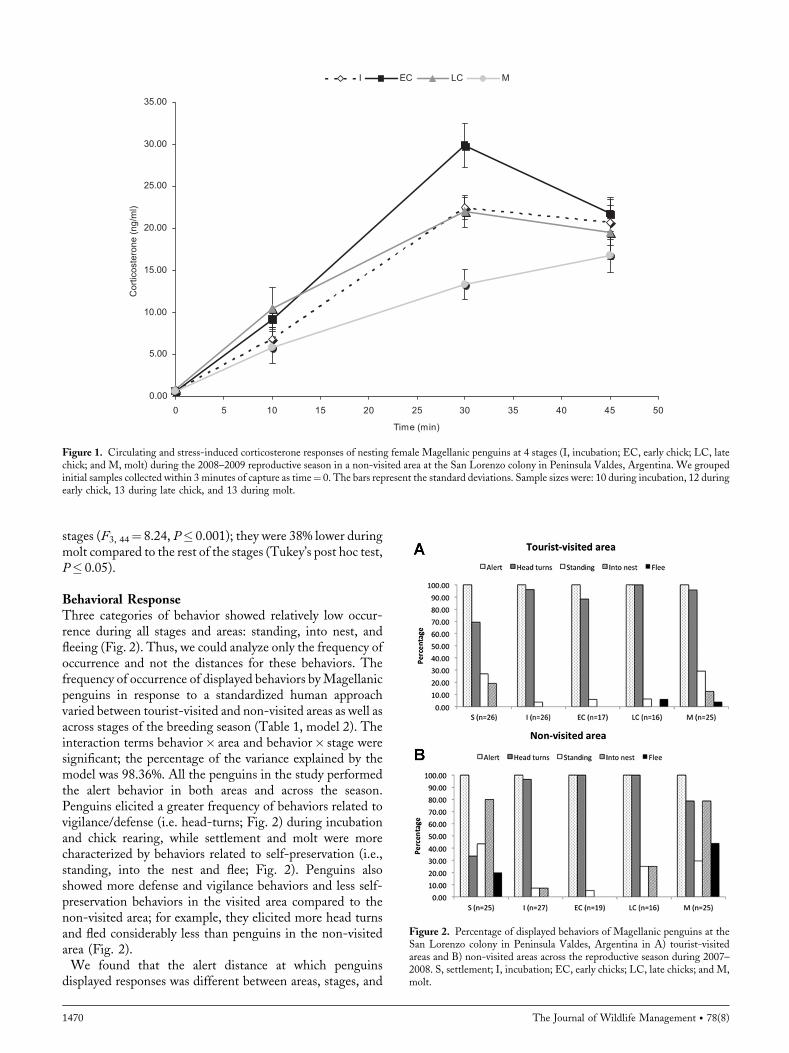
stages (F3, 44¼ 8.24, P� 0.001); they were 38% lower duringmolt compared to the rest of the stages (Tukey’s post hoc test,P� 0.05).
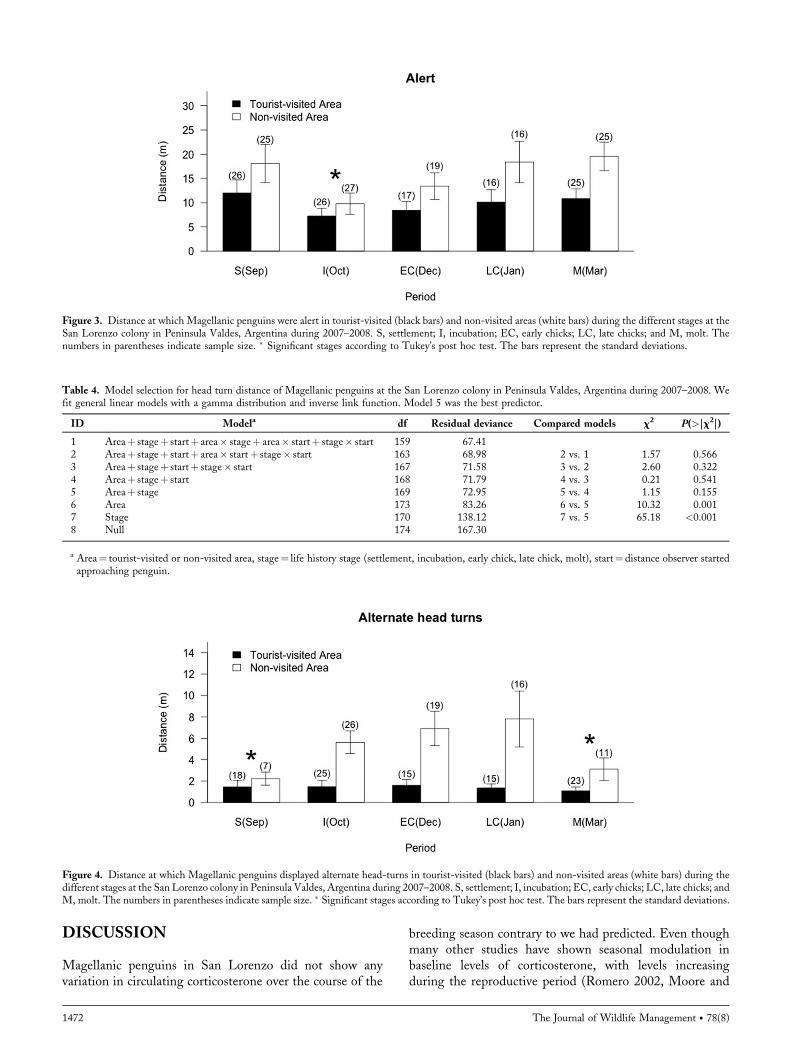
Behavioral ResponseThree categories of behavior showed relatively low occur-rence during all stages and areas: standing, into nest, andfleeing (Fig. 2). Thus, we could analyze only the frequency ofoccurrence and not the distances for these behaviors. Thefrequency of occurrence of displayed behaviors byMagellanicpenguins in response to a standardized human approachvaried between tourist-visited and non-visited areas as well asacross stages of the breeding season (Table 1, model 2). Theinteraction terms behavior� area and behavior� stage weresignificant; the percentage of the variance explained by themodel was 98.36%. All the penguins in the study performedthe alert behavior in both areas and across the season.Penguins elicited a greater frequency of behaviors related tovigilance/defense (i.e. head-turns; Fig. 2) during incubationand chick rearing, while settlement and molt were morecharacterized by behaviors related to self-preservation (i.e.,standing, into the nest and flee; Fig. 2). Penguins alsoshowed more defense and vigilance behaviors and less self-preservation behaviors in the visited area compared to thenon-visited area; for example, they elicited more head turnsand fled considerably less than penguins in the non-visitedarea (Fig. 2).We found that the alert distance at which penguins
displayed responses was different between areas, stages, and
0.00
5.00
10.00
15.00
20.00
25.00
30.00
35.00
0 5 10 15 20 25 30 35 40 45 50
Time (min)
Cor
ticos
tero
ne (n
g/m
l)
I EC LC M
Figure 1. Circulating and stress-induced corticosterone responses of nesting female Magellanic penguins at 4 stages (I, incubation; EC, early chick; LC, latechick; and M, molt) during the 2008–2009 reproductive season in a non-visited area at the San Lorenzo colony in Peninsula Valdes, Argentina. We groupedinitial samples collected within 3 minutes of capture as time¼ 0. The bars represent the standard deviations. Sample sizes were: 10 during incubation, 12 duringearly chick, 13 during late chick, and 13 during molt.
Figure 2. Percentage of displayed behaviors of Magellanic penguins at theSan Lorenzo colony in Peninsula Valdes, Argentina in A) tourist-visitedareas and B) non-visited areas across the reproductive season during 2007–2008. S, settlement; I, incubation; EC, early chicks; LC, late chicks; and M,molt.
1470 The Journal of Wildlife Management � 78(8)

start distances of the researcher (Table 2, model 5). Thepercentage of variance explained by the model was 44%.Penguins in the non-visited area became alert on average of5.85m farther from the researcher than penguins in thetourist-visited area during all stages (Table 3). The alertdistance was on average 5.32m shorter (Table 3) duringincubation than during any of the other 4 stages (Tukey’spost hoc test for the 2 areas combined, P� 0.05 for allcomparisons; Fig. 3). Also, penguins became alert to theapproaching researcher farther away at greater startdistances.The distance at which penguins started doing alternating
head turns was different between areas and stages; the startdistance of the researcher did not affect the head turnsdistance (Table 4, model 5). The percentage of varianceexplained by the model was 56%. Penguins in the non-visitedarea elicited head turns on average 4.34m farther than birds
in the tourist-visited area (Table 3, Fig. 4). The meandistance at which the head turned was 2.52m shorter duringthe settlement andmolt stages compared to the other 3 stages(Tukey’s post hoc test for the 2 areas combined, P� 0.05,Fig. 4).A greater reaction distance represented a longer period of
tolerance to an approaching person. The reaction distancewas different between areas, stages, and the start distance ofthe researcher (Table 5, model 4). The variance explained bythe model was 15%. The reaction distance was 1.13m greaterin the tourist-visited area than in the non-visited area(Table 3), indicating that penguins in the tourist-visited areatook longer to begin their defensive response following theirinitial alert as compared to penguins in the non-visited area.Penguins presented a shorter reaction distance duringincubation than in the other stages (3.59m shorter; Tukey’spost hoc for the 2 areas combined, P� 0.05; Fig. 5).
Table 1. Model selection for the occurrence of behavioral responses of Magellanic penguins at the San Lorenzo colony in Peninsula Valdes, Argentinaduring 2007–2008. We fit general linear models with a binomial distribution for proportional data and logit link functions. We removed the terms in themodels 1 at a time and compared pairs of models with maximum likelihood ratio tests (LRT). Model 2 was the best descriptor.
ID Modela df Residual deviance Compared models x2 P(>|x2|)
1 Behaviorþ areaþ stageþ behavior� areaþ behavior� stageþ area� stage 16 13.622 Behaviorþ areaþ stageþ behavior� areaþ behavior� stage 20 16.33 2 vs. 1 2.71 0.6073 Behaviorþ areaþ stageþ behavior� area 36 147.51 3 vs.2 131.18 <0.0014 Behaviorþ areaþ stageþ behavior� stage 24 58.69 4 vs. 2 42.35 <0.0015 Null 49 998.27
a Behavior¼ type of behavior displayed (alert, head turn, stand, into the nest, flee), area¼ tourist-visited or non-visited area, stage¼ life history stage(settlement, incubation, early chick, late chick, molt).
Table 2. Model selection for alert distance of Magellanic penguins at the San Lorenzo colony in Peninsula Valdes, Argentina during 2007–2008. We fitgeneral linear models with a gamma distribution and inverse link function. Model 4 was the best predictor.
ID Modela df Residual deviance Compared models x2 P(>|x2|)
1 Areaþ stageþ startþ area� stageþ area� startþ stage� start 206 45.682 Areaþ stageþ startþ area� startþ stage� start 210 46.13 2 vs. 1 0.44 0.6553 Areaþ stageþ startþ stage� start 211 46.50 3 vs. 2 0.37 0.1494 Areaþ stageþ start 215 47.40 4 vs. 3 0.90 0.2875 Areaþ stage 216 55.74 5 vs. 4 8.34 <0.0016 Areaþ start 219 51.89 6 vs. 4 4.49 <0.0017 Stageþ start 216 51.50 7 vs. 5 4.10 <0.0018 Null 221 79.19
a Area¼ tourist-visited or non-visited area, stage¼ life history stage (settlement, incubation, early chick, late chick, molt), start¼ distance observer startedapproaching penguin.
Table 3. Mean distances and standard deviations (inm) for the displayed behaviors and reaction distance of Magellanic penguins in the San Lorenzo colonyin Peninsula Valdes, Argentina during the 2007–2008 breeding season. TV¼ tourist-visited, NV¼ non-visited.
Distancea Area Settlement Incubation Early chicks Late chicks Molt
Start TV 25.60 (7.17) 20.21 (7.28) 16.8 (4.78) 21.77 (5.32) 24.34 (7.76)NV 30.37 (10.02) 22.34 (8.85) 23.53 (5.76) 21.78 (5.28) 32.32 (7.87)
Alert TV 12.04 (6.61) 7.27 (3.84) 8.43 (3.48) 10.14 (4.78) 10.87 (4.87)NV 18.08 (9.64) 9.80 (5.56) 13.41 (5.69) 18.38 (8.03) 19.56 (7.14)
Alternate head-turns TV 1.45 (1.54) 1.48 (1.43) 1.61 (1.08) 1.37 (0.70) 1.10 (0.84)NV 2.23 (1.50) 5.63 (2.67) 6.92 (3.33) 7.80 (4.94) 3.12 (2.56)
Reaction distance TV 10.64 (6.99) 5.67 (3.67) 6.77 (3.21) 8.80 (4.73) 9.26 (5.28)NV 7.51 (6.85) 4.15 (4.84) 6.49 (3.39) 9.78 (6.55) 8.72 (6.76)
a Start¼ distance observer started approaching penguin, alert¼ distance penguin detected the approaching person, alternate head-turns¼ distance penguinrotated its head from side to side, reaction distance¼ alert distance – distance penguin elicited a subsequent behavior.
Villanueva et al. � Seasonal Response to Visitation in Penguins 1471
DISCUSSION
Magellanic penguins in San Lorenzo did not show anyvariation in circulating corticosterone over the course of the
breeding season contrary to we had predicted. Even thoughmany other studies have shown seasonal modulation inbaseline levels of corticosterone, with levels increasingduring the reproductive period (Romero 2002, Moore and
Figure 3. Distance at which Magellanic penguins were alert in tourist-visited (black bars) and non-visited areas (white bars) during the different stages at theSan Lorenzo colony in Peninsula Valdes, Argentina during 2007–2008. S, settlement; I, incubation; EC, early chicks; LC, late chicks; and M, molt. Thenumbers in parentheses indicate sample size. � Significant stages according to Tukey’s post hoc test. The bars represent the standard deviations.
Table 4. Model selection for head turn distance of Magellanic penguins at the San Lorenzo colony in Peninsula Valdes, Argentina during 2007–2008. Wefit general linear models with a gamma distribution and inverse link function. Model 5 was the best predictor.
ID Modela df Residual deviance Compared models x2 P(>|x2|)
1 Areaþ stageþ startþ area� stageþ area� startþ stage� start 159 67.412 Areaþ stageþ startþ area� startþ stage� start 163 68.98 2 vs. 1 1.57 0.5663 Areaþ stageþ startþ stage� start 167 71.58 3 vs. 2 2.60 0.3224 Areaþ stageþ start 168 71.79 4 vs. 3 0.21 0.5415 Areaþ stage 169 72.95 5 vs. 4 1.15 0.1556 Area 173 83.26 6 vs. 5 10.32 0.0017 Stage 170 138.12 7 vs. 5 65.18 <0.0018 Null 174 167.30
a Area¼ tourist-visited or non-visited area, stage¼ life history stage (settlement, incubation, early chick, late chick, molt), start¼ distance observer startedapproaching penguin.
Figure 4. Distance at which Magellanic penguins displayed alternate head-turns in tourist-visited (black bars) and non-visited areas (white bars) during thedifferent stages at the San Lorenzo colony in Peninsula Valdes, Argentina during 2007–2008. S, settlement; I, incubation; EC, early chicks; LC, late chicks; andM, molt. The numbers in parentheses indicate sample size. � Significant stages according to Tukey’s post hoc test. The bars represent the standard deviations.
1472 The Journal of Wildlife Management � 78(8)

Jessop 2003), there are exceptions to this pattern. Forexample, birds that breed in extreme conditions such as aridor polar regions, have been shown to maintain low baselinecorticosterone levels throughout reproduction (Wingfieldet al. 1992, 1995; O’Reilly and Wingfield 2003; Wilson andHolberton 2004). Magellanic penguins similarly face a widerange of weather conditions across the breeding season.Temperature can be quite variable; during arrival andsettlement often below 08C, and up to 408C in January toFebruary during midsummer. Severe rainstorms can alsocause flooding and thermal stress, in some instances causingsignificant reproductive failure in the colonies (Stokes andBoersma 1998). Furthermore, Magellanic penguins aremigrants and reproduction is restricted to a specific periodin the year; it is synchronous among individuals in aparticular colony. Because high baseline levels of circulatingcorticosterone can cause birds to avoid reproduction(Sapolsky et al. 2000), keeping baseline corticosterone levelslow would be an adaptive strategy for animals with a smallreproductive window.In contrast to baseline levels, stress-induced corticosterone
levels varied throughout the season, with lowest levels beingexhibited during molt. Variation in the annual stress-induced
response has been documented in many birds (Romero2002). Although, no clear patterns of variability during thereproductive period have emerged, most show a consistentattenuated stress response during molt (Romero 2002).Having the capacity to modify the reaction to stressors via
the glucocorticoid stress response can be adaptive in times ofnecessity (i.e., heavy storms, predator attacks). In penguins,molt is energetically demanding (Adams and Brown 1990)and, because of the complete change of feathers, the plumageis not waterproof during molt. As such, penguins stay on landuntil molt finishes (Davis and Renner 2003, Otsukaet al. 2004). A robust stress response, intended to allowan animal to escape from a stressor, might not be particularlyadaptive if the birds cannot flee to the ocean. In addition,studies have also shown that higher corticosterone levelsduring molt produce feathers of poorer quality which, inflighted species, would have serious implications in flightefficiency and thermoregulation (Romero et al. 2005, Des-Rochers et al. 2009, Lattin et al. 2011). While flightless, thequality of feathers is equally important in penguins, asfeathers are imperative for thermoregulation while birds areat sea. The down regulation of corticosterone secretionduring molt could be useful in avoiding the degradation of
Table 5. Model selection for reaction distance of Magellanic penguins at the San Lorenzo colony in Peninsula Valdes, Argentina during 2007–2008. We fitgeneral linear models with a gamma distribution and inverse link function. Model 4 was the best predictor.
ID Modela df Residual deviance Compared models x2 P(>|x2|)
1 Areaþ stageþ startþ area� stageþ area� startþ stage� start 199 153.342 Areaþ stageþ startþ area� stageþ area� start 203 154.87 2 vs. 1 1.53 0.5353 Areaþ stageþ startþ area� stage 204 155.21 3 vs. 2 0.34 0.4054 Areaþ stageþ start 208 156.10 4 vs. 3 0.89 0.7675 Areaþ stage 209 168.93 5 vs. 4 12.83 <0.0016 Areaþ start 212 162.4 6 vs. 4 6.30 0.0087 Stageþ start 209 161.38 7 vs. 4 5.28 <0.0018 Null 214 183.61
a Area¼ tourist-visited or non-visited area, stage¼ life history stage (settlement, incubation, early chick, late chick, molt), start¼ distance observer startedapproaching penguin.
Figure 5. Reaction distance (alert distance – next behavior distance) of Magellanic penguins in tourist-visited (black bars) and non-visited areas (white bars)across the season at the San Lorenzo colony in Peninsula Valdes, Argentina during 2007–2008. S, settlement; I, incubation; EC, early chicks; LC, late chicks;and M, molt. The numbers in parentheses indicate sample size. � Significant stages according to Tukey’s post hoc test. The bars represent the standarddeviations.
Villanueva et al. � Seasonal Response to Visitation in Penguins 1473

proteins that are necessary for feather production and quality(Romero et al. 2005, DesRochers et al. 2009, Lattinet al. 2011).All birds living in both tourist-visited and non-visited areas
in the colony approached by a pedestrian during ourbehavioral study, displayed the alert behavior across stagesindicating that penguins perceive humans as intruders in thecolony and might assess if they represent a potential threat.Our results also showed that penguins nesting in tourist-visited areas were more tolerant to visitation than penguins inthe non-tourist area because standing, into the nest, and fleebehaviors were all less common in tourist-visited penguinsand alert and alternate head turn distances were shorter. Thisindicates that tourist-visited penguins have habituated tohaving tourists in the area and no longer perceive them as athreat. These results complement those found in a previousstudy at San Lorenzo where penguins showed no differencesin terms of breeding success, growth, and corticosteroneconcentrations between tourist-visited and non-tourist areas(Villanueva et al. 2012). Such habituation of Magellanicpenguins to tourist visitation have also been reported at thePunta Tombo colony (Yorio and Boersma 1992, Walkeret al. 2006).Our work goes further to show that the responses to human
visitation depend on the time of the breeding seasonin Magellanic penguins. During settlement, penguinsresponded to the pedestrian approach by displaying morebehaviors associated to self-preservation, such as standing,going farther into the nest, and fleeing, as we had predicted.Incubation seems to be a sensitive stage because penguinsshowed a lower tolerance to the approaching person.Although incubating penguins showed a shorter alertdistance (i.e., the pedestrian could get close to the nest),the reaction distance was also short indicating that penguinswere not very tolerant during incubation. A shorter alertdistance might be associated with penguins being in theirnests and not outside in the open during incubation. As such,birds incubating eggs in their nests may have less potential ofbeing alerted by neighbors or more protected sight lines thanbirds in other stages. Also, incubating birds seemed to bemore lethargic (they lie on the eggs with their eyes closed)and paid less attention to their surroundings. Similarfindings were observed in another tourist-visited Magellanicpenguin colony at Puerto Deseado, Argentina; the distanceat which an approaching person was detected was shorter inincubating penguins than when penguins were raising chicks(Cevasco et al. 2001). One important outcome of ourapproach was that by integrating the different behavioralmeasures in the reaction distance, we could identify thatincubation is a particularly critical stage. Without thisanalysis, we would have arrived to completely misleadingconclusions, specifically, incubation would have been theleast vulnerable stage because penguins allowed a closerapproach and stayed in their nests.During chick rearing (both early and late chick stages),
penguins showed a greater alert distance than duringincubation but also displayed a greater proportion ofhead-turns. During late chick stage, however, some birds
showed some self-preservation behaviors (i.e., standing,hiding, or fleeing). This may be because grownup chicks canmove and hide and do not require the full protection of anadult, whereas they are less mobile when they have justhatched.During incubation and chick rearing, when an investment
in parenting had been established, penguins elicited headturns more often than in other stages and displayed no or alow proportion of behaviors related to their own survival suchas standing, hiding in the nest, or fleeing. Head turns mightrepresent a defense behavior (Eggleton and Siegfried 1977)and it has also been associated with vigilance in otherpenguin species (Holmes et al. 2005). Thus, during timeswhen adults are protecting their nests or their contents, theyare more prone to show their defensive response.During molt, penguins showed fewer head turns and more
frequent behaviors related to self-survival such as standing,going farther into the nest, and fleeing. Because molting doesnot involve parental investment, penguins might be lesslikely to elicit defensive responses as compared to incubationor chick rearing, because they are not defending a particularresource, in this case, their eggs or chicks. Reaction distancewas also greatest during molt, suggesting that penguinstook time to perform a secondary response either becausethey were assessing the potential threat or because internalphysiological mechanisms (e.g., corticosterone release)might be suppressed.When we integrated our physiological and behavioral
results, we found there is both variation in stress-inducedcorticosterone and in behavioral responses to visitation acrossthe breeding season, as we had hypothesized. However, wedid not find a seasonal variation in circulating corticosteronelevels, or a relationship with penguin behavioral responses tovisitation. Therefore, we cannot say that the behavioralresponse was directly related to the physiological state of anindividual. For example, during molt, birds showed a lowercorticosterone release, but showed a high frequency of fleeingresponses. Thus, even though birds had a reduced tendencyto release corticosterone, they were capable of expressing arobust behavioral response. Thus, the behavioral hypothesis(Romero 2002) might not well explain the variation inbehavior and physiology we found in the study and thephysiological regulation in Magellanic penguins might bemore associated to survival in an extreme environment.The underlying processes determining behavioral responsesseem to be much more complex than a direct controlby glucocorticoid secretion. More research is needed tounderstand the complex mechanisms relating physiologyto behavior. Ultimately, the physiological state will drivethe behavioral costs and decisions in individuals, but, aswe see here, these relationships are more complex thanglucocorticoid stress physiology alone.
MANAGEMENT IMPLICATIONS
Magellanic penguins nesting in tourist-visited areas in theSan Lorenzo colony have shown to be tolerant to visitation.However, our study showed that incubation is a sensitivestage, regardless of the nesting area. We also found that birds
1474 The Journal of Wildlife Management � 78(8)

were physiologically sensitive during molt because thecorticosterone release is suppressed during this stage. Thesedistinctions are important because tourist visitation occursthroughout the breeding season. We recommend thattourists be informed and have respectful and responsiblebehavior throughout the season, but particularly during thesevulnerable times (incubation and molt). Although it is likelythat, for penguins as a whole, incubation and molt areuniversally sensitive times, other species and or touristvisitation locations might have additional periods of highersensitivity to disturbances. For example, non-colonialnesting penguins might be more susceptible to disturbancesduring their solitary settlement period (Ellenberget al. 2007). Thus, our results should be considered in theirown context and as reference only, and should not necessarilybe applied to all penguins universally. Each species and,perhaps, even individual tourist visitation location shouldbe carefully examined when assessing the impact of touristvisitation.
ACKNOWLEDGMENTS
We thank Estancia San Lorenzo and Argentina Vision forthe support in the field logistics and all the volunteers whohelped during the execution of the project: J. Zanni,M. Roriguez, G. Casasnovas, F. Spizzoucco, J. Haskins,M. Faber, A. Gatto, C. Giese, S. Villanueva, D. Ahumada,P. Villanueva, J. Lemus, M. Frixione, A. Nunez, A.Carranza, and A. Marino. We thank L. M. Romero atTufts University for allowing the access to his laboratory forhormone analyzes. Conservation, Research and EducationOpportunities (CREO) funded the hormone analyzes andC.V while in the United States. Fairfield Universitysponsored the time in Argentina for B.G.W. CONICETfunded the PhD scholarship of C.V. The authors of thispaper have no conflict of interests to declare.
LITERATURE CITEDAdams, N. J., and C. R. Brown. 1990. Energetics of molt in penguins. Pages297–315 in L. S., Davis, and J. T., Darby, editors. Penguin biology.Academic Press, San Diego, California, USA.
Albers, H. E., L. Yogev, R. B. Todd, and B. D. Goldman. 1985. Adrenalcorticoids in hamsters: role in circadian timing. American Journal ofPhysiology – Regulatory. Integrative and Comparative Physiology 248:434–438.
Atkinson, H. C., and B. J. Waddell. 1997. Circadian variation in basalplasma corticosterone and adrenocorticotropin in the rat: sexualdimorphism and changes across the estrous cycle. Endocrinology 138:3842–3848.
Beale, C. M., and P. Monaghan. 2004. Behavioural responses to humandisturbance: a matter of choice? Animal Behaviour 68:1065–1069.
Bertellotti, M., J. L. Tella, J. A. Godoy, G. Blanco, M. Forero, J. A.Donazar, and O. Ceballos. 2002. Determining sex of Magellanic penguinsusing molecular procedures and discriminant functions. Waterbirds 25:479–484.
Boersma, P. D. 2008. Penguins as marine sentinels. BioScience 58:597–607.Bookbinder, P. M., E. Dinerstein, A. Rijal, H. Cauley, and A. Rajouria.1998. Ecotourism’s support of biodiversity conservation. ConservationBiology 12:1399–1404.
Burger, J., and M. Gochfeld. 2007. Responses of emperor penguins(Aptenodytes forsteri) to encounters with ecotourists while commuting toand from their breeding colony. Polar Biology 30:1303–1313.
Cevasco, C. M., E. Frere, and P. A. Gandini. 2001. Intensidad de visitascomo condicionante de la respuesta del pinguino deMagallanes (Spheniscus
magellanicus) al disturbio humano. Ornitologıa Neotropical 12:75–81.[In Spanish.].
Cherel, Y., J. Leloup, and Y. Le Maho. 1988a. Fasting in king penguin. II.Hormonal and metabolic changes during molt. American Journal ofPhysiology – Regulatory. Integrative and Comparative Physiology254:178–184.
Cherel, Y., J.-P. Robin, O. Walch, H. Karmann, P. Netchitailo, and Y. LeMaho. 1988b. Fasting in king penguins. I. Hormonal and metabolicchanges during breeding. American Journal of Physiology – Regulatory.Integrative and Comparative Physiology 254:170–177.
Cockrem, J. F., M. A. Potter, and E. J. Candy. 2006. Corticosterone inrelation to body mass in Adelie penguins (Pygoscelis adeliae) affected byunusual sea ice conditions at Ross Island, Antarctica. General andComparative Endocrinology 149:244–252.
Crawley, M. J. 2007. The R book. John Wiley & Sons, Ltd, West Sussex,England.
Crossin, G. T., P. N. Trathan, R. A. Phillips, K. B. Gorman, A. Dawson,K. Q. Sakamoto, and T. D. Williams. 2012. Corticosterone predictsforaging behavior and parental care in macaroni penguins. AmericanNaturalist 180:31–41.
Culik, B., D. Adelung, and A. J. Woakes. 1990. The effect of disturbanceon the heart rate and behaviour of Adelie penguins (Pygoscelis adeliae)during the breeding season. Pages 177–182 in K. R., Kerry and G.,Hempel, editors. Antarctic ecosystems: ecological change and conserva-tion. Springer-Verlag, Triltsch, Wurzburg, Germany.
Daciuk, J. 1977. Notas faunısticas y bioecologicas de Penınsula Valdes yPatagonia. VI. Observaciones sobre areas de nidificacion de la avifauna dellitoral marıtimo patagonico (Provincias de Chubut y Santa Cruz, Rep.Argetina). El Hornero 11:361–376. [In Spanish.].
Davis, L. S., and M. Renner. 2003. Penguins. Yale University Press,London, United Kingdom.
DesRochers, D.W., J. M. Reed, J. Awerman, J. A. Kluge, J.Wilkinson, L. I.van Griethuijsen, J. Aman, and L. M. Romero. 2009. Exogenous andendogenous corticosterone alter feather quality. Comparative Biochemis-try and Physiology - Part A: Molecular & Integrative Physiology 152:46–52.
Eggleton, P., and W. R. Siegfried. 1977. Displays of the jackass penguin.Ostrich 50:139–167.
Ellenberg, U., A. N. Setiawan, A. Cree, D. M. Houston, and P. J. Seddon.2007. Elevated hormonal stress response and reduced reproductive outputin yellow-eyed penguins exposed to unregulated tourism. General andComparative Endocrinology 152:54–63.
Falabella, V., C. Campagna, and J. Croxall. 2009. Atlas del Mar Patagonico.Especies y espacios. Wildlife Conservation Society and Birdlife,International Buenos Aires, Argentina: [In Spanish.].
Fowler, G. S. 1999. Behavioral and hormonal responses of Magellanicpenguins (Spheniscus magellanicus) to tourism and nest site visitation.Biological Conservation 90:143–149.
Groscolas, R., A. Lacroix, and J.-P. Robin. 2008. Spontaneous egg or chickabandonment in energy-depleted king penguins: a role for corticosteroneand prolactin? Hormones and Behavior 53:51–60.
Hilborn, R., and M. Mangel. 1997. The ecological detective confrontingmodels with data. Princeton University Press, Princeton, New Jersey,USA.
Holmes, N. 2007. Comparing king, gentoo, and royal penguin responsesto pedestrian visitation. Journal of Wildlife Management 71:2575–2582.
Holmes, N., M. Giese, H. Achurch, S. Robinson, and L. K. Kriwoken.2006. Behaviour and breeding success of gentoo penguins Pygoscelis papuain areas of low and high human activity. Polar Biology 29:399–412.
Holmes, N., M. Giese, and L. K. Kriwoken. 2005. Testing the minimumapproach distance guidelines for incubating royal penguins Eudyptesschlegeli. Biological Conservation 126:339–350.
Hood, L. C., P. D. Boersma, and J. C. Wingfield. 1998. The adrenocorticalresponse to stress in incubating Magellanic penguins (Spheniscusmagellanicus). Auk 115:76–84.
Lattin, C. R., J. M. Reed, D. W. DesRochers, and L. M. Romero. 2011.Elevated corticosterone in feathers correlates with corticosterone-induceddecreased feather quality: a validation study. Journal of Avian Biology42:247–252.
Moore, I. T., and T. S. Jessop. 2003. Stress, reproduction, and adrenocorticalmodulation in amphibians and reptiles. Hormones and Behavior 43:39–47.
Villanueva et al. � Seasonal Response to Visitation in Penguins 1475

O’Reilly, K.M., and J. C.Wingfield. 2003. Seasonal, age, and sex differencesin weight, fat reserves, and plasma corticosterone in western sandpipers.Condor 105:13–26.
Otsuka, R., K. Aoki, H. Hori, and M. Wada. 1998. Changes in circulatingLH, sex steroid hormones, thyroid hormones and corticosterone inrelation to breeding and molting in captive Humboldt penguins(Spheniscus humboldti) kept in an outdoor open display. ZoologicalScience 15:103–109.
Otsuka, R., T. Machida, and M. Wada. 2004. Hormonal correlations attransition from reproduction to molting in an annual life cycle ofHumboldt penguins (Spheniscus humboldti). General and ComparativeEndocrinology 135:175–185.
Romero, L. M. 2002. Seasonal changes in plasma glucocorticoidconcentrations in free-living vertebrates. General and ComparativeEndocrinology 128:1–24.
Romero, L. M., N. E. Cyr, and R. C. Romero. 2006. Corticosteroneresponses change seasonally in free-living house sparrows (Passerdomesticus). General and Comparative Endocrinology 149:58–65.
Romero, L. M., and J. M. Reed. 2005. Collecting baseline corticosteronesamples in the field: is under 3min good enough? ComparativeBiochemistry and Physiology, Part A:Molecular & Integrative Physiology140:73–79.
Romero, L. M., K. K. Soma, and J. C. Wingfield. 1998. Hypothalamic-pituitary-adrenal axis changes allow seasonal modulation of corticosteronein a bird. American Journal of Physiology 274:R1338–R1344.
Romero, L. M., D. Strochlic, and J. C. Wingfield. 2005. Corticosteroneinhibits feather growth: Potential mechanism explaining seasonal downregulation of corticosterone during molt. Comparative Biochemistry andPhysiology - Part A: Molecular & Integrative Physiology 142:65–73.
Sapolsky, R. M., L. M. Romero, and A. U. Munck. 2000. How doglucocorticoids influence stress responses? Integrating permissive,suppressive, stimulatory, and preparative actions. Endocrine reviews21:55–89.
Schiavini, A., P. Yorio, P. Gandini, A. Raya Rey, and P. D. Boersma. 2005.Los pinguinos de las costas argentinas: estado poblacional y conservacion.Hornero 20:5–23. [In Spanish.].
Seddon, P. J., and U. Ellenberg. 2008. Effects of human disturbance onpenguins: the need for site- and species-specific visitor managementguidelines. Pages 163–180 in J., Higham and M., Luck, editors. Marinewildlife and tourism management: insights from the natural and socialsciences. CAB International, Oxfordshire, United Kingdom.
Stokes, D., and P. D. Boersma. 1998. Nest-site characteristics andreproductive success inMagellanic penguins (Spheniscus magellanicus). Auk151:34–49.
Thierry, A., S. Brajon, M. Spee, and T. Raclot. 2014. Differential effects ofincreased corticosterone on behavior at the nest and reproductive output ofchick-rearing Adelie penguins. Behavioral Ecology and Sociobiology68:721–732.
van Heezik, Y., and P. J. Seddon 1990. Effect of human disturbance onbeach groups of jackass penguins. South African Journal of WildlifeResearch 20:89–93.
Villanueva, C., B. G. Walker, and M. Bertellotti. 2012. A matter of history:effects of tourism on physiology, behaviour and breeding parametersin Magellanic penguins (Spheniscus magellanicus) at two colonies inArgentina. Journal of Ornithology 153:219–228.
Vleck, C. M., N. Vertalino, D. Vleck, and T. L. Bucher. 2000. Stress,corticosterone, and heterophil to lymphocyte ratios in free-living adeliepenguins. Condor 102:392–400.
Walker, B. G., P. D. Boersma, and J. C. Wingfield. 2004. Physiologicalcondition inMagellanic penguins: does it matter if you have to walk a longway to your nest? Condor 106:696–701.
Walker, B. G., P. D. Boersma, and J. C. Wingfield. 2005. Physiological andbehavioral differences in Magellanic penguin chicks in undisturbed andtourist-visited locations of a colony. Conservation Biology 19:1571–1577.
Walker, B. G., P. D. Boersma, and J. C. Wingfield. 2006. Habituation ofadult Magellanic penguins to human visitation as expressed throughbehavior and corticosterone secretion. Conservation Biology 20:146–154.
Williams, T. D., and P. D. Boersma. 1995. Magellanic penguin. Pages 249–258 in The penguins: Spheniscidae. Oxford University Press, Oxford,United Kingdom.
Wilson, C.M., and R. L. Holberton. 2004. Individual risk versus immediatereproductive success: a basis for latitudinal differences in the adrenocorticalresponse to stress in yellow warblers (Dendroica petechia). Auk 121:1238–1249.
Wingfield, I. J., andD. S. Farner. 1975. The determination of five steroids inavian plasma by radioimmunoassay and competitive protein-binding.Steroids 26:311–327.
Wingfield, J. C. 1994. Modulation of the adrenocortical response to stressin birds. Pages. 520–528 in K. G., Davey R. E., Peter and S. S., Tobe,editors. Perspectives in comparative endocrinology. National ResearchCouncil of Canada, Ottawa, Canada.
Wingfield, J. C., K. M. O’Reilly, and L. B. Astheimer. 1995. Modulation ofthe adrenocortical responses to acute stress in arctic birds: a possibleecological basis. American Zoologist 35:285–294.
Wingfield, J. C., C. M. Vleck, and M. C. Moore. 1992. Seasonal changes ofthe adrenocortical response to stress in birds of the Sonoran desert. Journalof Experimental Zoology 264:419–428.
Yorio, P., and P. D. Boersma. 1992. The effects of human disturbance onMagellanic penguin Spheniscus magellanicus behaviour and breedingsuccess. Bird Conservation International 2:161–173.
Yorio, P., E. Frere, P. Gandini, and A. Schiavini. 2001. Tourism andrecreation at seabird breeding sites in Patagonia, Argentina: currentconcerns and future prospects. Bird Conservation International 11:231–245.
Associate Editor: Heather Mathewson.
1476 The Journal of Wildlife Management � 78(8)
Copyright © 2022 FDOKUMEN




















