
Stable isotopes reveal Holocene changes in the diet of Adélie penguins in Northern Victoria Land...
9
Oecologia (2010) 164:911–919 DOI 10.1007/s00442-010-1790-2 123 BEHAVIORAL ECOLOGY - ORIGINAL PAPER Stable isotopes reveal Holocene changes in the diet of Adélie penguins in Northern Victoria Land (Ross Sea, Antarctica) Sandra Lorenzini · Carlo Baroni · Anthony E. Fallick · Ilaria Baneschi · Maria Cristina Salvatore · Giovanni Zanchetta · Luigi Dallai Received: 15 February 2010 / Accepted: 8 September 2010 / Published online: 1 October 2010 © Springer-Verlag 2010 Abstract Adélie penguin (Pygoscelis adeliae) modern and fossil eggshells and guano samples collected from ornithogenic soils in Terra Nova Bay (Victoria Land, Ross Sea) were processed for carbon and nitrogen isotopic ratios with the aim of detecting past penguin dietary changes. A detailed and greatly expanded Adélie penguin dietary record dated back to 7,200 years BP has been reconstructed for the investigated area. Our data indicate a signiWcant die- tary shift between Wsh and krill, with a gradual decrease from past to present time in the proportion of Wsh compared to krill in Adélie penguin diet. From 7,200 to 2,000 years BP, 13 C and 15 N values indicate Wsh as the most eaten prey. The dietary contribution of lower-trophic prey in penguin diet started becoming evident not earlier than 2,000 years BP, when the 13 C values reveal a change in the penguin feeding behavior. Modern eggshell and guano samples reveal a major dietary contribution of krill but not a krill-dominated diet, since 13 C values remain much too high if krill prevail in the diet. According to the Holocene environmental background attested for Victoria Land, Adélie penguin dietary shifts between Wsh and krill seem to reXect penguin paleoecological responses to diVerent paleoenvironmental settings with diVerent conditions of sea-ice extension and persistence. Furthermore, Adélie pen- guin diet appears to be particularly aVected by environmen- tal changes in a very speciWc period within the breeding season, namely the egg-laying period when penguin dietary and feeding habit shifts are clearly documented by the 13 C of eggshell carbonate. Keywords Adélie penguins · Stable isotopes · Paleodiet · Holocene Introduction As a fundamental climatic regulator, the Antarctic conti- nent plays a pivotal role in recording current global climate and environmental changes. Yet Antarctica also holds a long-term record of the Earth’s past climate, locked and preserved in ice and in sediments retaining a rich archive of biological remains (Roberts et al. 2001; Taylor et al. 2001; Ivany et al. 2006). Since the 1990s, valuable new insights into the late Pleistocene and Holocene history of Antarctica have been particularly provided from paleoecological stud- ies of key animal species belonging to the Southern Ocean ecosystems (Baroni and Orombelli 1994a; Lambert et al. 2002; Baroni and Hall 2004; Ainley et al. 2005; Hall et al. Communicated by Scott McWilliams. Electronic supplementary material The online version of this article (doi:10.1007/s00442-010-1790-2) contains supplementary material, which is available to authorized users. S. Lorenzini (&) · C. Baroni · G. Zanchetta Dipartimento di Scienze della Terra, Università degli Studi di Pisa, Via S. Maria 53, 56126 Pisa, Italy e-mail: [email protected] C. Baroni · I. Baneschi · G. Zanchetta · L. Dallai Istituto di Geoscienze e Georisorse, Consiglio Nazionale delle Ricerche, Via G. Moruzzi 1, 56124 Pisa, Italy A. E. Fallick Scottish Universities Environmental Research Centre, East Kilbride , G750QF Scotland, UK M. C. Salvatore Dipartimento di Scienze della Terra, Università di Roma “La Sapienza”, P.le A. Moro, 5, 00185 Rome, Italy
Transcript of Stable isotopes reveal Holocene changes in the diet of Adélie penguins in Northern Victoria Land...

Oecologia (2010) 164:911–919
DOI 10.1007/s00442-010-1790-2BEHAVIORAL ECOLOGY - ORIGINAL PAPER
Stable isotopes reveal Holocene changes in the diet of Adélie penguins in Northern Victoria Land (Ross Sea, Antarctica)
Sandra Lorenzini · Carlo Baroni · Anthony E. Fallick · Ilaria Baneschi · Maria Cristina Salvatore · Giovanni Zanchetta · Luigi Dallai
Received: 15 February 2010 / Accepted: 8 September 2010 / Published online: 1 October 2010© Springer-Verlag 2010
Abstract Adélie penguin (Pygoscelis adeliae) modernand fossil eggshells and guano samples collected fromornithogenic soils in Terra Nova Bay (Victoria Land, RossSea) were processed for carbon and nitrogen isotopic ratioswith the aim of detecting past penguin dietary changes. Adetailed and greatly expanded Adélie penguin dietaryrecord dated back to 7,200 years BP has been reconstructedfor the investigated area. Our data indicate a signiWcant die-tary shift between Wsh and krill, with a gradual decreasefrom past to present time in the proportion of Wsh comparedto krill in Adélie penguin diet. From 7,200 to 2,000 yearsBP, �13C and �15N values indicate Wsh as the most eatenprey. The dietary contribution of lower-trophic prey in
penguin diet started becoming evident not earlier than2,000 years BP, when the �13C values reveal a change inthe penguin feeding behavior. Modern eggshell and guanosamples reveal a major dietary contribution of krill but nota krill-dominated diet, since �13C values remain much toohigh if krill prevail in the diet. According to the Holoceneenvironmental background attested for Victoria Land,Adélie penguin dietary shifts between Wsh and krill seem toreXect penguin paleoecological responses to diVerentpaleoenvironmental settings with diVerent conditions ofsea-ice extension and persistence. Furthermore, Adélie pen-guin diet appears to be particularly aVected by environmen-tal changes in a very speciWc period within the breedingseason, namely the egg-laying period when penguin dietaryand feeding habit shifts are clearly documented by the �13Cof eggshell carbonate.
Keywords Adélie penguins · Stable isotopes · Paleodiet · Holocene
Introduction
As a fundamental climatic regulator, the Antarctic conti-nent plays a pivotal role in recording current global climateand environmental changes. Yet Antarctica also holds along-term record of the Earth’s past climate, locked andpreserved in ice and in sediments retaining a rich archive ofbiological remains (Roberts et al. 2001; Taylor et al. 2001;Ivany et al. 2006). Since the 1990s, valuable new insightsinto the late Pleistocene and Holocene history of Antarcticahave been particularly provided from paleoecological stud-ies of key animal species belonging to the Southern Oceanecosystems (Baroni and Orombelli 1994a; Lambert et al.2002; Baroni and Hall 2004; Ainley et al. 2005; Hall et al.
Communicated by Scott McWilliams.
Electronic supplementary material The online version of this article (doi:10.1007/s00442-010-1790-2) contains supplementary material, which is available to authorized users.
S. Lorenzini (&) · C. Baroni · G. ZanchettaDipartimento di Scienze della Terra, Università degli Studi di Pisa, Via S. Maria 53, 56126 Pisa, Italye-mail: [email protected]
C. Baroni · I. Baneschi · G. Zanchetta · L. DallaiIstituto di Geoscienze e Georisorse, Consiglio Nazionale delle Ricerche, Via G. Moruzzi 1, 56124 Pisa, Italy
A. E. FallickScottish Universities Environmental Research Centre, East Kilbride , G750QF Scotland, UK
M. C. SalvatoreDipartimento di Scienze della Terra, Università di Roma “La Sapienza”, P.le A. Moro, 5, 00185 Rome, Italy
123

912 Oecologia (2010) 164:911–919
2006; Millar et al. 2008; de Bruyn et al. 2009). AmongAntarctic organisms studied in the context of Antarcticpaleoclimatic and paleoenvironmental research, Adéliepenguins (Pygoscelis adeliae) are considered one of thebest indicators of environmental adaptation, according tothe extreme environmental sensitivity that regulates theirpresence and geographic distribution around the continent(Ainley et al. 1998; Ainley 2002; Arrigo et al. 2002;Croxall et al. 2002; Emmerson and Southwell 2008). In theice-free coastal areas, the discovery in the ornithogenic soils(bird-guano deposits; Ugolini 1972) of high concentrationsof Adélie penguin remains dating back »40,000 years(Baroni and Orombelli 1994a, b; Lambert et al. 2002;Millar et al. 2008) has presented the unique opportunity totrack the colonization history of this species during thisextended period. Along the Victoria Land coast (Ross Sea,Antarctica), radiocarbon dated penguin bones, eggshellsand guano document the presence of the Adélies since theearly Holocene (»8,000 years BP) (Baroni and Orombelli1994a; Lambert et al. 2002). Testifying the past availabilityof ice-free terrains, this record enabled scientists to date theearly phases of coastal deglaciation and emergence, whichoccurred in Victoria Land after the Last Glacial Maximum(»18,000–20,000 years BP) (Baroni and Hall 2004; Hallet al. 2004). The identiWcation of abandoned colonies inareas no longer occupied by Adélie penguins also docu-ments that during the Holocene the local and regional envi-ronmental conditions frequently changed in Antarctica,resulting in situations more or less favorable to Adéliepenguin settlement (Stonehouse 1970; Baroni and Orombelli1994a, b; Hall et al. 2006; Emslie et al. 2007).
In the context of paleoecological research, ornithogenicsoils also provide a unique opportunity to study the diet andthe feeding behavior of a single species as well as its die-tary changes both in diVerent areas and in diVerent periodsof time, since at least the early Holocene (Emslie andPatterson 2007).
By analyzing the C and N isotopic composition of fossiland modern penguin eggshells and guano, this paper aimsto provide a new detailed Adélie penguin paleodietaryrecord dated back to ca. 8,000 years BP for the area ofTerra Nova Bay (Victoria Land, Ross Sea). In fact, analysisof N and C stable isotope ratios in animal tissues representsa powerful technique for determining the trophic level atwhich foraging occurs and the importance of diVerent preyitems in the diet. This approach is based on the fact that sta-ble isotope abundances of carbon and nitrogen in tissuespass from prey to predator increasing approximately 0.8–3.4‰, respectively, with each trophic level (DeNiro andEpstein 1978; Minagawa and Wada 1984; Vander Zandenand Rasmussen 2001), and that they are preserved inexcreta. These isotopic enrichments provide direct informa-tion about the average diet of an individual and its dietary
patterns, especially when the dietary sources consist of preyfrom diVerent trophic levels (i.e., diVerent C and N isotopiccomposition) (Hobson et al. 1994; Koch et al. 1994;Hobson 1995; Fry 2006). In particular, diVerences in nitro-gen isotope ratios are frequently used to predict trophiclevel, while carbon has more often been used to assess for-aging behavior (DeNiro and Epstein 1978; Minagawa andWada 1984).
In this work, we investigated the Adélie penguin paleo-diet and its temporal shift by complementing the isotopicdata obtained from Adélie penguin eggshells by 13C/12Cand 15N/14N measurements on penguin guano. In temporalterms, these data will provide diVerent kinds of informationabout Adélie penguin paleodiet. The isotopic compositionof eggshell is an indication of the diet of the female at thetime of eggshell secretion, whereas the isotopic composi-tion of guano should represent a more integrated signal forboth males and females over a longer time.
Materials and methods
Study area and sample collection
Straddling the boundary between northern and southernVictoria Land at the western side of the Ross Sea, theinvestigated area stretches along the coast between Edmon-son Point (74°20�S, 165°08�E) and Inexpressible Island(74°53�S, 163°45�E; Fig. 1).
The occurrence of series of datable marine depositsrelated to the post-LGM glacial retreat has made Terra
Fig. 1 The Terra Nova Bay region showing the location of the sevenAdélie penguin (Pygoscelis adeliae) colonies sampled in this work.Black circles and white circles indicate abandoned and modern colo-nies, respectively
123

Oecologia (2010) 164:911–919 913
Nova Bay a key area to study the early phase of Ross Seadeglaciation (Baroni and Orombelli 1991, 1994b; Baroniand Hall 2004). The crucial paleoenvironmental value,added to the presence of both modern and abandonedAdélie penguin colonies on a coastal stretch no more than100 km long, make Terra Nova Bay a suitable area for thisstudy.
We processed penguin remains from three modern andfour abandoned Adélie penguin colonies (Fig. 1), sampledover several Weld surveys between 1994 and 2007 (Baroniand Orombelli 1994a; Lambert et al. 2002; Baroni and Hall2004; Hall et al. 2006), in the framework of Italian Antarcticexpeditions (PNRA). At all investigated sites ornithogenicsoils were excavated layer-by-layer using the stratigraphicexcavation techniques commonly used in archeologicalresearch (Baroni and Orombelli 1994a; Lambert et al. 2002).This careful Weldwork allowed collection at distinct organiclayers of several well-preserved and age-constrained fossilpenguin remains, such as bones, eggshells and guano.
The accurate stratigraphic control of sampling and themultiple dates obtained from diVerent penguin remains col-lected at the same organic layer show a good chronologicalcorrespondence. Therefore, neither contamination nor mix-ing of sediments from diVerent layers has occurred sincethe ornithogenic layers formed. For this reason, eggshellfragments collected from the same stratigraphic level as thedated fragments are attributed to the same age. A few sam-ples were also dated by association with other penguinremains of known age (i.e., penguin bones and guano) col-lected from the same stratigraphic level or, in only twocases (see Appendix A, samples GX21406 and OS36027),from bracketing ages from bottom layers. Fresh guano andmodern eggshell samples were also collected during theWeldwork at the outskirts of occupied nests.
Laboratory analyses
Fossil eggshells were selected both from each excavatedornithogenic level at the time of Weld excavation andthrough the subsequent sieving of guano samples in the lab-oratory. Penguin eggshells were cleaned in an ultrasonicbath with a 10% vol H2O2, to remove organic matter, rinsedseveral times in deionized water, and then dried in an ovenat low temperature. Eggshells were then crushed with apestle in an agate mortar until the shells were reduced to aWne-grained powder. Carbon stable isotope analyses wereperformed at SUERC (Scottish Universities EnvironmentalResearch Centre, East Kilbride, Scotland) with the AP2003mass spectrometer equipped with a separate acid injectorsystem, after reaction with 105% H3PO4 under He atmo-sphere at 70°C. Totals of 858 fossil and 65 modern penguineggshell samples were analyzed for carbon isotope compo-sition.
Carbon and nitrogen stable isotope analyses were per-formed on 62 fossil and 6 modern penguin guano samplesat the IGG-CNR, Pisa, Italy. These samples had been for-merly sieved and dried at low temperature. For each sam-ple, we selected the <63-�m grain size fraction, whichquantitative analyses reported as the richest in organic car-bon content. Before isotope analyses, samples were acidi-Wed with 10% HCl to remove inorganic carbon and thenwashed several times with deionised water to neutral pHand dried. Subsamples of 2–20 mg, depending on nitrogenand carbon content, were weighed into tin capsules andanalyzed using a Thermo Finnigan Elemental AnalyzerEA-1108 interfaced with an isotope ratio mass spectrome-ter (Finnigan DeltaPlusXL) via the ConXo II interface(Finnigan, Bremen, Germany).
Isotopic abundances are reported using the conventionaldelta (�) notation as parts per thousand (‰) deviation inisotope ratio from international standards, namely V-PDBfor carbon and atmospheric N2 (AIR) for nitrogen. The�13C and �15N data obtained from modern and fossileggshells and guano samples are given in Appendix A.Sixty-six new eggshell and guano samples were radiocar-bon-dated at the NOSAMS laboratory of Woods Hole(USA). All AMS dates (in 14C years BP) were corrected forthe upwelling of old water in the Southern Ocean by sub-tracting a marine-carbon reservoir eVect of 1,300 years. Atthe moment, this represents the best estimate and conven-tional value used for the Ross Sea region, based on dates ofpre-bomb organisms of known ages (Berkman and Forman1996). For the eggshells, large enough fossil samples weredated directly by splitting the eggshell and using one partfor radiocarbon analysis and the other for stable isotopeanalysis, in order to ensure an accurate and absolute datefor the corresponding stable isotope values. In order toexamine whether the original composition of eggshells hasbeen seriously aVected by diagenetic and recrystallizationeVects, random modern and fossil samples were analyzedand compared using X-ray diVraction and scanning electronmicroscope (SEM).
All statistical analyses were performed using the soft-ware Statistica, version 7.1 for Windows (StatSoft 2005).The data and the results of the statistical analyses weregraphed using Statistica and Microsoft Excel.
Results
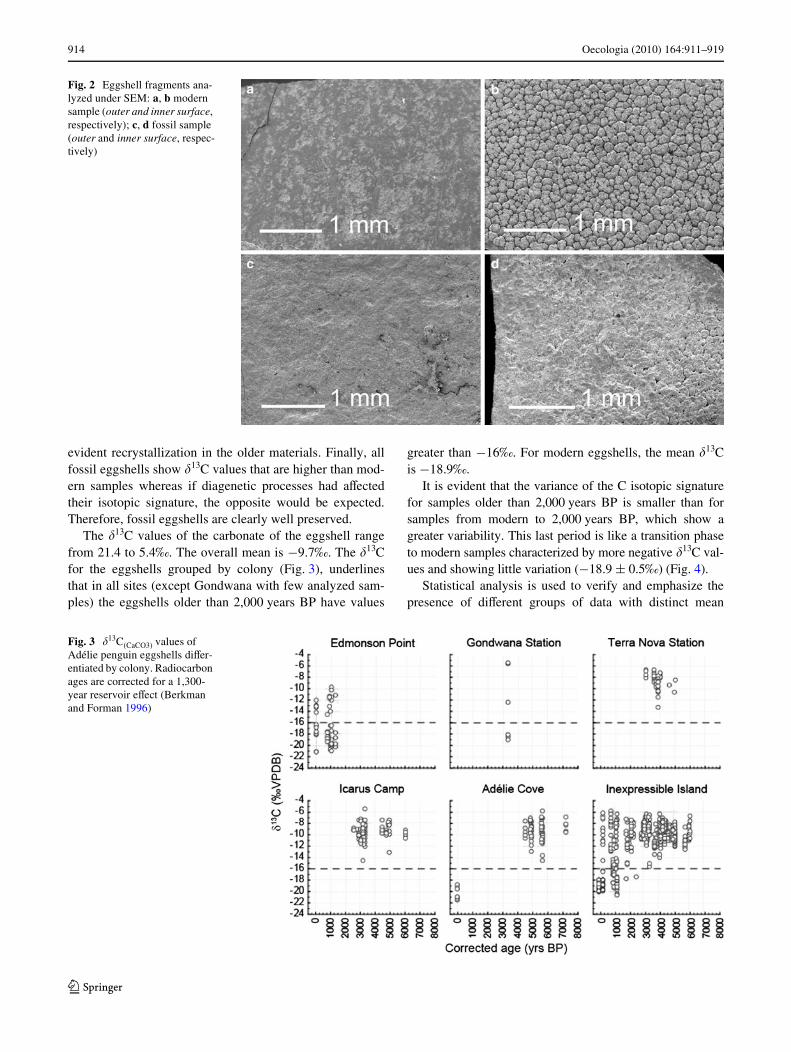
SEM analyses indicate that the inner spherulitic microstruc-tures generated by crystal growth of calcite are easily rec-ognized both in the modern and the archaeological eggshellsamples (Fig. 2). Furthermore, diVractograms obtainedfrom eggshell fragments document an almost perfect over-lap of signals between modern and fossil samples, with no
123
914 Oecologia (2010) 164:911–919
evident recrystallization in the older materials. Finally, allfossil eggshells show �13C values that are higher than mod-ern samples whereas if diagenetic processes had aVectedtheir isotopic signature, the opposite would be expected.Therefore, fossil eggshells are clearly well preserved.
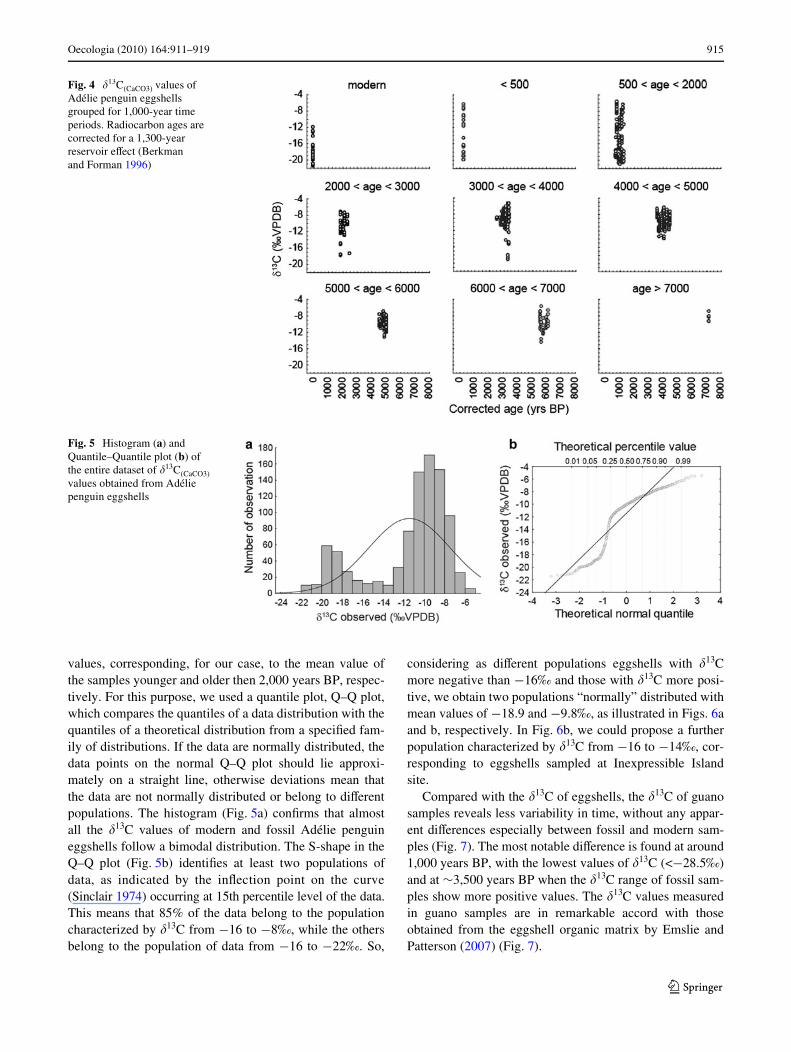
The �13C values of the carbonate of the eggshell rangefrom 21.4 to 5.4‰. The overall mean is ¡9.7‰. The �13Cfor the eggshells grouped by colony (Fig. 3), underlinesthat in all sites (except Gondwana with few analyzed sam-ples) the eggshells older than 2,000 years BP have values
greater than ¡16‰. For modern eggshells, the mean �13Cis ¡18.9‰.
It is evident that the variance of the C isotopic signaturefor samples older than 2,000 years BP is smaller than forsamples from modern to 2,000 years BP, which show agreater variability. This last period is like a transition phaseto modern samples characterized by more negative �13C val-ues and showing little variation (¡18.9 § 0.5‰) (Fig. 4).
Statistical analysis is used to verify and emphasize thepresence of diVerent groups of data with distinct mean
Fig. 2 Eggshell fragments ana-lyzed under SEM: a, b modern sample (outer and inner surface, respectively); c, d fossil sample (outer and inner surface, respec-tively)
Fig. 3 �13C(CaCO3) values of Adélie penguin eggshells diVer-entiated by colony. Radiocarbon ages are corrected for a 1,300-year reservoir eVect (Berkman and Forman 1996)
123
Oecologia (2010) 164:911–919 915
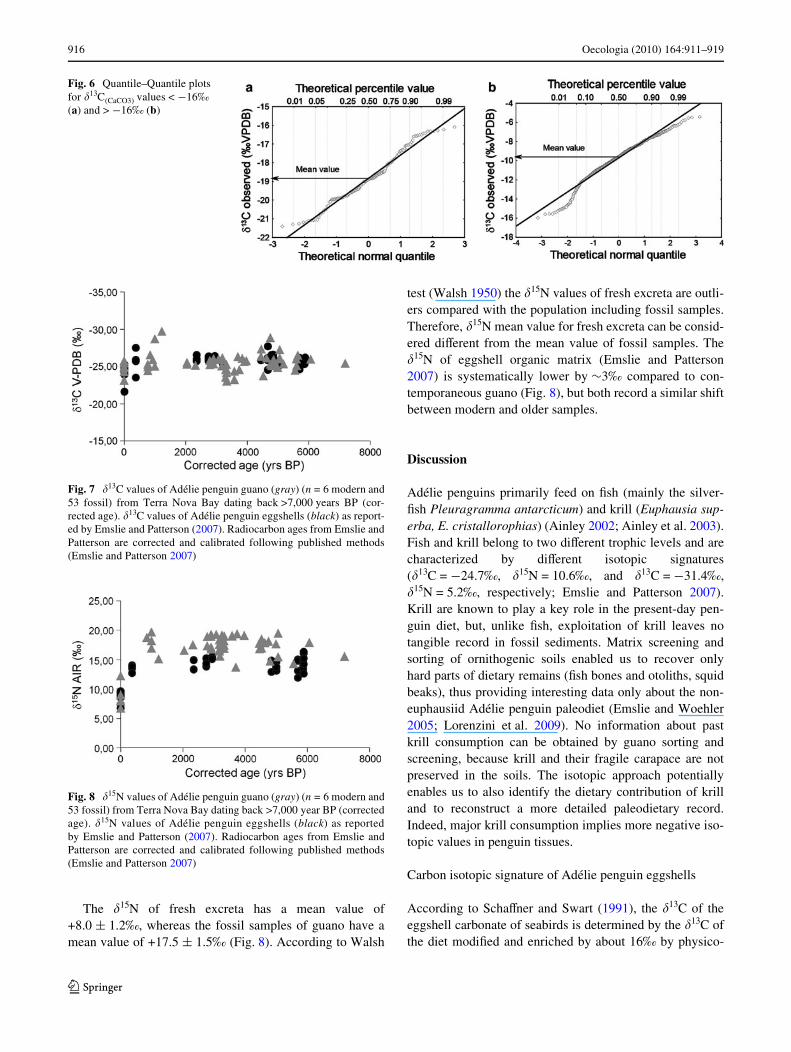
values, corresponding, for our case, to the mean value ofthe samples younger and older then 2,000 years BP, respec-tively. For this purpose, we used a quantile plot, Q–Q plot,which compares the quantiles of a data distribution with thequantiles of a theoretical distribution from a speciWed fam-ily of distributions. If the data are normally distributed, thedata points on the normal Q–Q plot should lie approxi-mately on a straight line, otherwise deviations mean thatthe data are not normally distributed or belong to diVerentpopulations. The histogram (Fig. 5a) conWrms that almostall the �13C values of modern and fossil Adélie penguineggshells follow a bimodal distribution. The S-shape in theQ–Q plot (Fig. 5b) identiWes at least two populations ofdata, as indicated by the inXection point on the curve(Sinclair 1974) occurring at 15th percentile level of the data.This means that 85% of the data belong to the populationcharacterized by �13C from ¡16 to ¡8‰, while the othersbelong to the population of data from ¡16 to ¡22‰. So,
considering as diVerent populations eggshells with �13Cmore negative than ¡16‰ and those with �13C more posi-tive, we obtain two populations “normally” distributed withmean values of ¡18.9 and ¡9.8‰, as illustrated in Figs. 6aand b, respectively. In Fig. 6b, we could propose a furtherpopulation characterized by �13C from ¡16 to ¡14‰, cor-responding to eggshells sampled at Inexpressible Islandsite.
Compared with the �13C of eggshells, the �13C of guanosamples reveals less variability in time, without any appar-ent diVerences especially between fossil and modern sam-ples (Fig. 7). The most notable diVerence is found at around1,000 years BP, with the lowest values of �13C (<¡28.5‰)and at »3,500 years BP when the �13C range of fossil sam-ples show more positive values. The �13C values measuredin guano samples are in remarkable accord with thoseobtained from the eggshell organic matrix by Emslie andPatterson (2007) (Fig. 7).
Fig. 4 �13C(CaCO3) values of Adélie penguin eggshells grouped for 1,000-year time periods. Radiocarbon ages are corrected for a 1,300-year reservoir eVect (Berkman and Forman 1996)
Fig. 5 Histogram (a) and Quantile–Quantile plot (b) of the entire dataset of �13C(CaCO3) values obtained from Adélie penguin eggshells
123
916 Oecologia (2010) 164:911–919
The �15N of fresh excreta has a mean value of+8.0 § 1.2‰, whereas the fossil samples of guano have amean value of +17.5 § 1.5‰ (Fig. 8). According to Walsh
test (Walsh 1950) the �15N values of fresh excreta are outli-ers compared with the population including fossil samples.Therefore, �15N mean value for fresh excreta can be consid-ered diVerent from the mean value of fossil samples. The�15N of eggshell organic matrix (Emslie and Patterson2007) is systematically lower by »3‰ compared to con-temporaneous guano (Fig. 8), but both record a similar shiftbetween modern and older samples.
Discussion
Adélie penguins primarily feed on Wsh (mainly the silver-Wsh Pleuragramma antarcticum) and krill (Euphausia sup-erba, E. cristallorophias) (Ainley 2002; Ainley et al. 2003).Fish and krill belong to two diVerent trophic levels and arecharacterized by diVerent isotopic signatures(�13C = ¡24.7‰, �15N = 10.6‰, and �13C = ¡31.4‰,�15N = 5.2‰, respectively; Emslie and Patterson 2007).Krill are known to play a key role in the present-day pen-guin diet, but, unlike Wsh, exploitation of krill leaves notangible record in fossil sediments. Matrix screening andsorting of ornithogenic soils enabled us to recover onlyhard parts of dietary remains (Wsh bones and otoliths, squidbeaks), thus providing interesting data only about the non-euphausiid Adélie penguin paleodiet (Emslie and Woehler2005; Lorenzini et al. 2009). No information about pastkrill consumption can be obtained by guano sorting andscreening, because krill and their fragile carapace are notpreserved in the soils. The isotopic approach potentiallyenables us to also identify the dietary contribution of krilland to reconstruct a more detailed paleodietary record.Indeed, major krill consumption implies more negative iso-topic values in penguin tissues.
Carbon isotopic signature of Adélie penguin eggshells
According to SchaVner and Swart (1991), the �13C of theeggshell carbonate of seabirds is determined by the �13C ofthe diet modiWed and enriched by about 16‰ by physico-
Fig. 6 Quantile–Quantile plots for �13C(CaCO3) values < ¡16‰ (a) and > ¡16‰ (b)
Fig. 7 �13C values of Adélie penguin guano (gray) (n = 6 modern and53 fossil) from Terra Nova Bay dating back >7,000 years BP (cor-rected age). �13C values of Adélie penguin eggshells (black) as report-ed by Emslie and Patterson (2007). Radiocarbon ages from Emslie andPatterson are corrected and calibrated following published methods(Emslie and Patterson 2007)
Fig. 8 �15N values of Adélie penguin guano (gray) (n = 6 modern and53 fossil) from Terra Nova Bay dating back >7,000 year BP (correctedage). �15N values of Adélie penguin eggshells (black) as reportedby Emslie and Patterson (2007). Radiocarbon ages from Emslie andPatterson are corrected and calibrated following published methods(Emslie and Patterson 2007)
123

Oecologia (2010) 164:911–919 917
chemical fractionation due to metabolism. So, the end-member values for Adélie penguin diet are expected to bearound ¡8 or ¡15‰ depending on the prey (Wsh or krill)mainly eaten. These values approximately correspond tothe mean values calculated for samples older and youngerthan 2,000 years BP, respectively (Figs. 6a, b). Moreover,between 7,000 and 2,000 years BP, consistently high �13Cvalues suggest that the diet was primarily based on high�13C food, implying signiWcant Wsh (mainly Pleuragrammaantarcticum) exploitation. This is also supported by thehighest number of Wsh remains recovered in guano samplesbeing collected from the same localities and dated between4,000 and 2,000 years BP (Lorenzini et al. 2009). Between1,300 and 230 years BP, the greater spread in �13C valuesand preponderance of negative and low values, especiallynoticeable at Inexpressible Island, clearly reveals a changein the feeding behavior of Adélie penguins with an increas-ing contribution of 13C-depleted prey (i.e., krill). Through-out this phase, however, the range of variability remainsvery high, likely reXecting a mixed diet of Wsh and krill.The shift toward a diet based on the consumption of a lower�13C prey (i.e., krill) seems to occur mostly in the modernsamples (lowest values). The 13C/12C ratios measured forour eggshell carbonate samples indicate a diet with increas-ing amount of krill with time.
Besides being useful to predict food source (and informabout trophic level), carbon isotope ratios are more fre-quently used to assess foraging behavior. In fact, theseratios diVer more between terrestrial versus marine, inshoreversus oVshore, and pelagic versus benthic food webs thanby trophic level (DeNiro and Epstein 1978; Minagawa andWada 1984). From this point of view, a signiWcative changein the foraging habit of Adélie penguins seem tooccur »2,000 years BP, when our data indicate a noticeableincreasing preponderance in lower �13C values.
Carbon and nitrogen isotopic signature of Adélie penguin guano
This dietary shift seems not to be recorded in the �13C val-ues of penguin guano samples. The lack of any particularevidence for Xuctuation in �13C values for this record isunlikely to be related to the decomposition of organic car-bon in the soil; in a similar study, Mizutani and Wada(1988) reported no signiWcant �13C variability betweenfresh guano and ornithogenic soils.
The isotopic ratios of eggshell and guano samples pro-vide diVerent kinds of information on Adélie penguin diet,corresponding to short and long periods, respectively. Inparticular, the �13C values of eggshell likely reXect a sig-niWcant moment of the Adélie penguin breeding season, theegg-laying event, that normally starts in the early Novem-ber and ends in December with egg-hatching (Ainley
2002). Since it takes on average 24 h to complete the for-mation of the egg and its organic tissue (Astheimer andGrau 1985), its isotopic signature records the most recentdietary signal of a single female individual. Conversely, theisotopic signature of guano samples is an integrated sea-sonal signal and a result of diVerent individuals, with diVer-ent age and sex and possibly diet. Similarly to �13C valuesof penguin guano, the �13C values measured for fossil andmodern eggshell organic matrices by Emslie and Patterson(2007) do not reveal any signiWcant variation over all theinvestigated time period. Thus, it is reasonable to concludethat organic matter carbon isotope data do not register astrong and compelling dietary shift similar to that recordedin inorganic carbon.
Fossil samples have �15N values consistently higherthan those of modern excreta. However, the gap betweenold and modern nitrogen isotope ratios is too high(�15Nold¡new + 10‰) to be interpreted simply as a result ofnutrient uptake. A change in the diet would likely imply anisotopic increase of about 3.4‰ with each trophic level(Minagawa and Wada 1984; Vander Zanden and Rasmus-sen 2001). By comparing the N isotopic composition offresh guano and ornithogenic soils, Mizutani and Wada(1988) deduced that nitrogen isotope fractionation associ-ated with the volatilization of ammonia after excreta depo-sition caused an increase in �15N of residual nitrogen, up to10‰. The ammonia volatilization eVect could reasonablyexplain the very high �15N values of ornithogenic samplescompared to modern guano and the shift observed betweenfossil and modern guano samples. Thus, compared to thewell-deWned dietary shift recorded in the �13C of eggshellcarbonate, neither �13C nor �15N adjusted for the volatiliza-tion eVect of Adélie penguin guano seems to detect any sig-niWcant change, directly related to a shift in the penguindiet.
New paleoecological and paleoenvironmental insights
Some speculations can be oVered about the paleoecologicaland paleoenvironmental signiWcance of the Adélie penguinpaleodietary trend as inferred from eggshell inorganic car-bon and guano organic nitrogen. The dietary changesobserved in the last 2,000 years, as suggested by the isoto-pic values of penguin eggshells, probably represent theecological responses of Adélie penguins to habitat modiW-cation. Geomorphological, glaciological and paleoeco-logical evidences have demonstrated that in Victoria Landclimatic and environmental conditions have changed duringthe Holocene, with glacial advances and retreats andchanges in sea-ice extent (Baroni and Orombelli 1991,1994a, 1994b; Baroni and Hall 2004; Hall et al. 2004,2006; Hall 2009). These environmental reconstructionsseem to be also documented in the Adélie penguin paleodi-
123

918 Oecologia (2010) 164:911–919
etary record whose variability seems to be aVected bychanges in the persistence of sea-ice. In fact, a signiWcantchange in the foraging habitat of Adélie penguins occurs atleast »2,000 years BP, when our data indicate a noticeableincreasing preponderance in lower �13C values. Thischange could well be due to a change in the sea-ice coverthat, bringing a change in sunlight conditions, would havereasonably altered the productivity of the waters in whichthe penguins were feeding.
The dietary contribution of prey of lower trophic statusstarts becoming important from 2,000 years BP, at the sametime as a reduction in sea-ice cover, as also suggested byHall et al. (2006). Conversely, the Wsh-rich Adélie penguindiet, occurring between 7,000 and 2,000 years BP, could berelated to the presence of permanent sea ice as the greatspread of Adélie penguin colonies (penguin optimum)would document (Baroni and Orombelli 1994a, b). Thisenvironmental condition would also represent a suitablesetting for a demographic expansion of Pleuragramma ant-arcticum (Zane et al. 2006).
Our data document an important decrease in eggshellcarbonate �13C values for modern samples, likely reXectingan increase in the proportion of krill in Adélie penguin dietonly as a relatively recent attainment. The large number ofprocessed samples and the temporal continuity of the inves-tigated dataset also enabled us to chronologically deWnepast Adélie penguin dietary shifts with more accuracy.According to our results, the nutritional contribution oflower-trophic preys starts to become evident t1,800 yearsearlier than the time pointed out by Emslie and Patterson(2007), when the �13C values revealed a mixed diet basedon Wsh and krill consumption. However, modern eggshellcarbonate reveals a major dietary contribution of krill butnot a krill-dominated diet, since �13C values, which areconsistent at around ¡25 to ¡24‰, are much too high ifkrill prevail in the diet. Finally, isotopic analyses of pen-guin guano indicate that Adélie penguin dietary shifts seemnot to occur throughout during the breeding season, but tobe mostly related to a very short, limited and particularperiod of time, that roughly corresponds to the egg-layingevent.
Conclusion
The stable isotope geochemistry of carbon and nitrogenprovides a powerful and intriguing tool for investigating indetail the Adélie penguin dietary patterns and shifts duringthe past.
Carbon and nitrogen isotope geochemistry of eggshellsand guano allowed us to reconstruct the so far longest andmost detailed Adélie penguin dietary record dating back toca. 8,000 years BP in the Terra Nova Bay region. Our data
indicate that Adélie penguin paleodiet seems to beinXuenced by the Holocene environmental conditions ofVictoria Land coast in terms of sea-ice extension andreduction. In fact, krill consumption became manifest since2,000 years BP, when paleoenvironmental reconstructionsuggested less sea-ice coverage. The endurance of suchenvironmental conditions probably consolidated up to thepresent day a penguin feeding habit mainly based on krillexploitation. Stable isotopic analyses also suggest thatAdélie penguins appear to be particularly aVected by envi-ronmental changes at a very speciWc period within thebreeding season, that is the egg-laying period when penguindietary shifts are clearly documented by the �13C of eggshellcarbonate.
In the context of paleocological research, paleodietarystudies are conWrmed to be a valuable tool to examine bio-diversity history and its evolution through the time. Alsoconsidering their environmental implication, these studiesassume a particular relevance in Antarctica, where the pres-ervation of long-term paleoecological records give newdata about the paleoenvironmental history of the continentsince at least the late Pleistocene.
Acknowledgments This work was executed under the Wnancial sup-port of the Italian National Program on Antarctic Research (PNRA)and of the PhD School in Earth Sciences of the University of Pisa.Isotopic analyses were performed at the SUERC of East Kilbride(Scotland) and at the IGG-CNR of Pisa (Italy). We particularly thankJulie Dougans and Andrew Tait for their irreplaceable technicalsupport and supervision in stable isotope analyses at SUERC. Newradiocarbon dates on penguin remains reported here were performed atthe National Ocean Sciences Accelerator Mass Spectrometry Facility(NOSAMS) at the Woods Hole Oceanographic Institution (USA). WeWnally thank S. McWilliams, D. Ainley and a third anonymous reviewerfor their careful readings and constructive suggestions.
References
Ainley DG (2002) The Adélie penguin: bellwether of climate change.Columbia University Press, New York
Ainley DG, Wilson PR, BartonKJ Ballard G, Nur N, Karl B (1998)Diet and foraging eVort of Adélie penguins in relation to pack-iceconditions in the southern Ross Sea. Polar Biol 20:311–319
Ainley DG, Ballard G, Barton KJ, Karl BJ, Rau GH, Ribic CA, WilsonPR (2003) Spatial and temporal variation of diet within a pre-sumed metapopulation of Adélie penguins. Condor 105:95–106
Ainley DG, Clarke ED, Arrigo K, Fraser WR, Kato A, Barton KJ,Wilson PR (2005) Decadal-scale changes in the climate and biotaof the PaciWc sector of the Southern Ocean, 1950s to the 1990s.Antarct Sci 17:171–182
Arrigo KR, van Dijken GL, Ainley DG, Fahnestock MA, Markus T(2002) Ecological impact of a large Antarctic iceberg. GeophysRes Lett 29:1104
Astheimer LB, Grau CR (1985) The timing and energetic conse-quences of egg formation in the Adélie penguin. Condor 87:256–268
Baroni C, Hall BL (2004) A new Holocene relative sea-level curve forTerra Nova Bay, Victoria Land, Antarctica. J Quat Sci 19:377–396
123

Oecologia (2010) 164:911–919 919
Baroni C, Orombelli G (1991) Holocene raised beaches at Terra NovaBay, Victoria Land, Antartica. Quat Res 36:157–177
Baroni C, Orombelli G (1994a) Abandoned penguin rookeries as Holo-cene paleoclimatic indicators in Antarctica. Geology 22:23–26
Baroni C, Orombelli G (1994b) Holocene glacier variations in TerraNova bay area (Victoria Land, Antartica). Antarct Sci 6:497–505
Berkman PA, Forman SL (1996) Pre-bomb radiocarbon and the reser-voir correction for calcareous marine species in the SouthernOcean. Geophys Res Lett 23:363–366
Croxall JP, Trathan PN, Murphy EJ (2002) Environmental change andAntarctic seabird populations. Science 297:1510–1514
de Bruyn M, Hall BL, Chauke LF, Baroni C, Koch PL, Rus Hoelzel A(2009) Rapid response of a marine mammal species to holoceneclimate and habitat change. PLoS Genet 5(7):e1000554.doi:10.1371/journal.pgen.1000554
DeNiro MJ, Epstein S (1978) InXuences of diet on the distribution ofcarbon isotopes in animals. Geochim Cosmochim Acta42(5):495–506
Emmerson L, Southwell C (2008) Sea ice cover and its inXuence onAdélie enguin reproductive performance. Ecology 89:2096–2102
Emslie SD, Patterson WP (2007) Abrupt shift in �13C and �15N valuesin Adélie penguin eggshell in Antarctica. Proc Natl Acad Sci USA104:11666–11669
Emslie SD, Woehler EJ (2005) A 9000-year record of Adélie penguinoccupation and diet in the Windmill Islands, East Antarctica.Antarct Sci 17:57–66
Emslie SD, Coats L, Licht K (2007) A 45, 000 year record of Adéliepenguins and climate change in the Ross Sea, Antarctica. Geol-ogy 35:61–64
Fry B (2006) Stable isotope ecology. Springer, New YorkHall BL (2009) Holocene glacial history of Antarctica and the sub-
Antarctic islands. Quat Sci Rev 28:2213–2230Hall BL, Baroni C, Denton GH (2004) Holocene relative sea-level his-
tory of the Southern Victoria Land Coast, Antarctica. Glob PlanetChang 42:241–263
Hall BL, Hoelzel AR, Baroni C, Denton GH, Le Boeuf BJ, Overturf B,Töpf AL (2006) Holocene elephant seal distribution implieswarmer-than-present climate in the Ross Sea. Proc Natl Acad SciUSA 103:10213–10217
Hobson KA (1995) Reconstructing avian diets using stable-carbon andnitrogen isotope analysis of egg components: patterns of isotopicfractionation and turnover. Condor 97:752–762
Hobson KA, Piatt JF, Pitocchelli J (1994) Using stable isotopes todetermine seabird trophic relationships. J Anim Ecol 63:786–798
Ivany LC, Van Simaeys S, Domaci EW, Samson SD (2006) Evidencefor an earliest Oligocene ice sheet on the Antarctic Peninsula.Geology 34:377–380
Koch PL, Fogel ML, Tuross N (1994) Tracing the diet of fossil animalsusing stable isotopes. In: Lajtha K, Michener RH (eds) Stable iso-
topes in ecology and environmental science. Blackwell, NewYork, pp 63–92
Lambert DM, Ritchie PA, Millar CD, Holland B, Drummond AJ,Baroni C (2002) Rates of evolution in ancient DNA from Adéliepenguins. Science 295:2270–2273
Lorenzini S, Olmastroni S, Pezzo F, Salvatore MC, Baroni C (2009)Holocene Adélie penguin diet in Victoria Land, Antarctica. PolarBiol 32:1077–1086
Millar CD, Dodd A, Anderson J, Gibb GC, Ritchie PA, Baroni C,Woodhams MD, Hendy MD, Lambert DM (2008) Mutation andevolutionary rates in Adélie penguins from the Antarctic. PLoSGenetics 4:1–9
Minagawa M, Wada E (1984) Stepwise enrichment of �15N along foodchains: further evidence and the relation between �15N and animalage. Geochim Cosmochim Acta 48:1135–1140
Mizutani H, Wada E (1988) Nitrogen and carbon isotope ratios in seabird rookeries and their ecological implications. Ecology 69:340–349
Roberts D, van Ommen TD, McMinn A, Morgan V, Roberts JL (2001)Late-Holocene East Antarctic climate trends from ice-core andlake-sediment proxies. The Holocene 11:117–120
SchaVner FC, Swart PK (1991) InXuence of diet and environmentalwater on the carbon and oxygen isotopic signatures of seabirdeggshell carbonate. Bull Mar Sci 48:23–38
Sinclair AJ (1974) Selection of threshold values in geochemical datausing probability graphs. J Geochem Explor 3:129–149
StatSoft inc (2005) STATISTICA (Data analysis software system).Version 7.1. 2005. Available in: http://www.statsoft.com
Stonehouse B (1970) Recent climatic change in Antarctica suggestedfrom 14C dating of penguin remains. Palaeogeogr PalaeoclimatolPalaeoecol 7:341–343
Taylor F, Whitehead J, Domack E (2001) Holocene paleoclimatechange in the Antarctic Peninsula: evidence from the diatom, sed-imentary and geochemical record. Mar Micropaleontol 41:25–43
Ugolini FC (1972) Ornithogenic soils of Antarctica. In: Llano GA (ed)Antarctic terrestrial biology. American Geophysical Union,Antarct Res Ser, vol 20, pp 181–193
Vander Zanden MJ, Rasmussen JB (2001) Variation in �15N and �13Ctrophic fractionation: implications for aquatic food web studies.Limnol Oceanogr 46:2061–2066
Walsh JE (1950) Some nonparametric tests of whether the largestobservation of a set are too large or too small. Ann Math Statist21:583–592
Zane L, Marcato S, Bargelloni L, Bortolotto E, Papetti C, Simonato M,Varotto V, Patarnello T (2006) Demographic history and popula-tion structure of the Antarctic silverWsh Pleuragramma antarcti-cum. Mol Ecol 15:4499–4511
123




















