
rTMS-induced virtual lesion of the posterior parietal cortex (PPC) alters the control of reflexive...
31
Author's Accepted Manuscript rTMS-induced virtual lesion of the posterior parietal cortex (PPC) alters the control of reflexive shifts of social attention triggered by pointing hands Giuseppina Porciello, Filippo Crostella, Marco Tullio Liuzza, Elia Valentini, Salvatore Maria Aglioti PII: S0028-3932(14)00139-0 DOI: http://dx.doi.org/10.1016/j.neuropsychologia.2014.04.017 Reference: NSY5155 To appear in: Neuropsychologia Received date: 12 July 2013 Revised date: 14 April 2014 Accepted date: 25 April 2014 Cite this article as: Giuseppina Porciello, Filippo Crostella, Marco Tullio Liuzza, Elia Valentini, Salvatore Maria Aglioti, rTMS-induced virtual lesion of the posterior parietal cortex (PPC) alters the control of reflexive shifts of social attention triggered by pointing hands, Neuropsychologia, http://dx.doi.org/ 10.1016/j.neuropsychologia.2014.04.017 This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting galley proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain. www.elsevier.com/locate/neuropsy- chologia
Transcript of rTMS-induced virtual lesion of the posterior parietal cortex (PPC) alters the control of reflexive...

Author's Accepted Manuscript
rTMS-induced virtual lesion of the posteriorparietal cortex (PPC) alters the control ofreflexive shifts of social attention triggered bypointing hands
Giuseppina Porciello, Filippo Crostella, MarcoTullio Liuzza, Elia Valentini, Salvatore MariaAglioti
PII: S0028-3932(14)00139-0DOI: http://dx.doi.org/10.1016/j.neuropsychologia.2014.04.017Reference: NSY5155
To appear in: Neuropsychologia
Received date: 12 July 2013Revised date: 14 April 2014Accepted date: 25 April 2014
Cite this article as: Giuseppina Porciello, Filippo Crostella, Marco Tullio Liuzza,Elia Valentini, Salvatore Maria Aglioti, rTMS-induced virtual lesion of theposterior parietal cortex (PPC) alters the control of reflexive shifts of socialattention triggered by pointing hands, Neuropsychologia, http://dx.doi.org/10.1016/j.neuropsychologia.2014.04.017
This is a PDF file of an unedited manuscript that has been accepted forpublication. As a service to our customers we are providing this early version ofthe manuscript. The manuscript will undergo copyediting, typesetting, andreview of the resulting galley proof before it is published in its final citable form.Please note that during the production process errors may be discovered whichcould affect the content, and all legal disclaimers that apply to the journalpertain.
www.elsevier.com/locate/neuropsy-
chologia

Porciello et al.�� � 1� �
1�
�
rTMS-induced virtual lesion of the posterior parietal cortex (PPC)
alters the control of reflexive shifts of social attention
triggered by pointing hands
Giuseppina Porciello1,2*, Filippo Crostella1, Marco Tullio Liuzza1,2, Elia Valentini1,2,
and Salvatore Maria Aglioti1,2*
1. Department of Psychology, “Sapienza”, University of Rome, Via dei Marsi 78, I-00185 Rome, Italy.
2. IRCCS, Fondazione Santa Lucia, Via Ardeatina 306, I-00179 Rome, Italy.
* Correspondence should be addressed to:
Salvatore Maria Aglioti Department of Psychology, “Sapienza”, University of Rome Via dei Marsi 78 00185 Rome, Italy Email: [email protected]; Phone number: +39 06 44427601 Fac-simile: +39 06 49917635 URL: http://w3.uniroma1.it/aglioti
or
Giuseppina Porciello Department of Psychology, “Sapienza”, University of Rome Via dei Marsi 78 00185 Rome, Italy Email: [email protected] Phone number: +3906 49927635 Fac-simile: +39 06 49917635

Porciello et al.�� � 2� �
2�
�
Abstract
In highly social groups like human and non human primates, gaze and pointing cues are
fundamentally important for directing the attention of conspecifics. Although neuroimaging studies
indicate that shifts of attention triggered by observation of social cues activate the onlookers' fronto-
parietal cortices, information on whether these regions play a causative role in orienting and re-
orienting of social attention is lacking. To advance our understanding of this, we used event-related
repetitive dual pulse transcranial magnetic stimulation to interfere with neural activity in the right
frontal eye field (rFEF) and posterior parietal cortex (rPPC). This procedure allowed us to explore
how inhibiting rFEF and rPPC influences shifts of attention triggered by the observation of body-
related (gaze and hand) and non body-related (arrow) directional distractors. Participants were
asked to perform a leftward or rightward pointing movement according to the color change of a
central imperative signal while ignoring a distractor, which was either a gaze, a pointing hand or an
arrow. Stimulation of rPPC in a region supposedly linked to attentional re-orienting and to planning
and execution of upper limb movements increased the reflexive tendency to follow distracting
pointing hands but not oriented gaze or arrows. These findings suggest that inhibition of cortical
structures that control attentional shifts triggered by social stimuli brings forth an increase of the
cost of attentional re-orienting. Moreover, our results provide the first causative evidence that
reflexive social attention in humans may be coded according to body-part-centered frames of
reference.
Keywords: Posterior Parietal Cortex; Frontal Eye Field; pointing movements; averted gaze;
rdpTMS; attentional shifts.

Porciello et al.�� � 3� �
3�
�
1. Introduction
Humans, like other primates, have evolved in dynamic social contexts (Dunbar & Shultz,
2007) in which gaze and hand movements are powerful channels for communication (Emery, 2000;
Liszkowski, Carpenter, Henning, Striano, & Tomasello, 2004). Averted gaze and pointing hands,
for example, are deictic gestures used to signal to other individuals where or to what one is paying
attention. Most existing studies on social attention have focused on gaze (Kingstone, 2009)
probably because eye contact is an early hallmark of interpersonal interactions and fundamental
social signals arise from the face. Studies in human (Driver et al., 1999; Friesen & Kingstone, 1998;
Langton & Bruce, 2000; Ricciardelli, Bricolo, Aglioti, & Chelazzi, 2002) and non-human primates
(Deaner, Platt, & Carolina, 2003) demonstrate that seeing the averted gaze of a model induces
pervasive and reflexive shifts of attention in an onlooker, independently of task relevance (Klein,
Shepherd, & Platt, 2009; Shepherd, 2010). However, although the eyes are powerful mediators of
social attention, they are not the only ones (Langton & Bruce, 2000). Indeed, seeing a pointing arm
influences directional judgments and triggers reflexive attentional shifts during hand movement
preparation and response, even when these shifts are detrimental to the performance (Ariga &
Watanabe, 2009; Fischer & Szymkowiak, 2004; Forster & Eimer, 2007; Langton & Bruce, 2000).
Note that systematic attentional shifts in response to dynamic pointing gestures are already present
in infants at 4.5 months of age (Rohlfing, Longo, & Bertenthal, 2012) suggesting they are
predisposed to quickly respond to these seminal cues even in earlier development stages. These
forms of joint attention represent the basis for the emergence of more complex social and cognitive
skills (such as perspective-taking, deception, empathy and theory of mind (Baron-Cohen, 1995;
Whiten, 1998; Emery, 2000)) and may index the development of dedicated neural networks that
map reflexive shifts of social attention.
What we know from previous studies is that these body-evoked attentional shifts occur reflexively
and seem to depend on a fronto-parietal network, which according to non-human and human

Porciello et al.�� � 4� �
4�
�
primate studies is involved in a variety of attentional tasks, including covert and overt reorienting of
attention to biological and non-biological stimuli (Corbetta, Patel, & Shulman, 2008, Corbetta &
Shulman, 2002, Tipper, Handy, Giesbrecht, & Kingstone, 2008, Wardak, Olivier, & Duhamel,
2011).
Although laboratory studies do not provide univocal evidence that body-related directional cues
(e.g. gaze, hands or full body) have a special role in capturing our attention with respect to non-
biological directional signals (e.g. regulatory or warning road arrows), it has been suggested that the
former are more relevant than the latter in daily life interpersonal interactions (Birmingham &
Kingstone, 2009; Kingstone, 2009) even because they could activate at the neural level mirror-like
responses (see Shepherd, Klein, Deaner, & Platt, 2009).
Not far from this idea, behavioral studies indicate that specific body-related cues have differential
distracting power depending on the effector participants use to perform the task (see Crostella,
Carducci, & Aglioti, 2009). Indeed, it was found that the type of overt directional response
(saccadic vs pointing movements) was differentially influenced by the type of distracting cue
(averted gaze vs pointing hand). In particular, directional gaze stimuli interfered more with saccadic
movements and directional hand stimuli more with pointing movements. This finding raised the
question of whether the brain codes social attention according to body-centered coordinate systems.
Thus, the influence of three different distractors (gaze, hand or arrows) on directional saccadic or
hand responses triggered by central instruction signals was explored using fMRI (Cazzato,
Macaluso, Crostella, & Aglioti, 2012). Results demonstrated that higher activity in the frontal eye
field (FEF) for distracting gaze was found mainly during the saccadic response task. In the same
vein, higher activity in the left and right inferior parietal sulcus (IPS) regions was found for the
distracting hands mainly in the hand-pointing task (Cazzato et al., 2012). This pattern of results
suggests that the automatic tendency of creating contexts of shared attention between individuals

Porciello et al.�� � 5� �
5�
�
through body-related cues reflects the activity of a fronto-parietal system where attentional mapping
is coded according to body-part specific reference frames.
In the present study, we sought to explore the neural underpinnings of body part-evoked attentional
shifts. To this aim, we combined the hand-pointing paradigm for exploring reflexive social attention
used in our previous studies (Cazzato et al., 2012; Crostella et al., 2009) with repetitive dual-pulse
transcranial magnetic stimulation (rdpTMS), allowing us to interfere with the neural activity of the
frontal eye fields (FEF) and posterior parietal cortex (PPC), two cortical regions involved in the
preparation and execution of saccadic movements (Corbetta, 1998; Paus, 1996) and in the control of
arm and hand movements (Desmurget et al., 1999; Iacoboni, 2006; Rushworth, Paus, & Sipila,
2001) respectively.
Studies report that rdpTMS over FEF and PPC produce facilitatory and inhibitory effects depending
on different stimulation parameters and behavioral contexts (Kapoula, Yang, Sabbah, & Vernet,
2011; O’Shea, Muggleton, Cowey, & Walsh, 2004; Wipfli et al., 2001).�
In the present study we chose the timing parameters of the two pulses to interfere with the initial
programming of pointing movements. Indeed, the first pulse was delivered simultaneously with the
distracting stimulus and the second pulse 25 ms after the onset of the imperative cue. The timing of
TMS stimulation was based on a companion study (Crostella, Porciello, Carducci & Aglioti, 2013)
where early but not late stimulation of FEF and PPC created interferential effects in a task similar to
the one reported in the present study.
Based on the notion that reflexive shifts of social attention are coded according to effector-based
reference frames, we predicted that interference of the neural activity in the posterior parietal cortex
would selectively disrupt participants’ ability to inhibit the reflexive tendency to point in the
direction of the distracting hands, more than in the direction of eyes and arrows. Viceversa, we
predicted that interfering with the frontal eye field should have less specific effects because
participants performed pointing and not saccadic movements.� This pattern of results would be in

Porciello et al.�� � 6� �
6�
�
keeping with our “social somatotopy” model of reflexive social attention developed on the basis of
behavioral (Crostella et al., 2009) and fMRI (Cazzato et al., 2012) data.
Moreover, it is worth noting that according to our model, rdpTMS over FEF when performing the
same task by using saccadic movements should bring about a higher distracting effect of gaze
compared to hands and arrows. Preliminary evidence in support of this prediction is provided in a
series of TMS studies performed in our laboratory (Crostella, Porciello, Carducci & Aglioti, 2013).
Finally, we used a non-body related stimulus (i.e. the arrows) as a control and expected it to be
overall less distracting with respect to the body-related stimuli. Importantly, we did not expect that
interfering with the neural activity of FEF or PPC would change the distracting power triggered by
arrows during the pointing-task.
2. Materials and Methods
2.1 Participants
Fourteen healthy participants (4 males; mean age 24; range 21-29; SD 2.8) were recruited
from a university opportunity sample. All were right-handed according to the Standard Handedness
Inventory (Briggs and Nebes, 1975), had normal or corrected-to-normal visual acuity in both eyes
and no problems with color perception. Participants were naïve about the purposes of the study. At
the beginning of the experimental session they received explanations about the experimental
procedure. Information concerning the experimental hypothesis was provided only at the end. The
procedures were approved by the Local Ethics Committee and were in accordance with the ethical
standards of the 1964 Declaration of Helsinki. All subjects gave their written informed consent.
None had a known history of neurological, psychiatric, other medical problems or any
contraindication to TMS (Rossi, Hallett, Rossini, & Pascual-Leone, 2009). No discomfort or
adverse effects during and after TMS were reported. Two participants reached a very low accuracy
(25% of correct responses) in one or more experimental conditions and for this reason were

Porciello et al.�� � 7� �
7�
�
excluded from the analyses, which were thus performed on twelve subjects (3 males; mean age:
23.08; range: 21-28; SD 3.08).
2.2 Stimuli and Task
Each participant was tested in one experimental session that lasted approximately two hours.
The experiment was created and run with E-Prime software (version 1.1, Psychology Software
Tools, Inc., Pittsburgh, PA) on an IBM compatible computer. We performed the study in an
ordinarily illuminated room. Participants sat quietly on a comfortable chair in front of an LCD
monitor, positioned at about 50 cm from their eyes, with their index finger on the table surface. The
start position of the hand was aligned with the central fixation point but positioned 28 cm below in
the horizontal plane, at about 20 cm from subjects’ torso. Each trial started with the appearance of a
black central fixation square (0.21° x 0.21° in size). Behind the black square, centered on the
monitor, one of three types of distracting stimuli was presented on a light gray background (about
47 cd/m2). The distracting stimuli consisted of digital Photoshop 8.0.1 (Adobe, CA) modified
photographs of a gaze, hand or an arrow (8° x 8° in size). The three distractors were created by
using colored photographs of: 1) a young woman with a directional gaze and an emotionally neutral
expression; 2) a man’s hand pointing; 3) a pointing arrow obtained by digitally scrambling the hand
distractor. To enhance the salience of the distractors and make the attention-capturing effects more
similar to those of daily life, stimuli were animated by presenting two frames in rapid sequence. The
first frame, lasting 500 ms, depicted a straight gaze, an upward pointing fist or a T-like shape. The
second frame depicted a leftward or rightward oriented gaze, extended finger or arrow; it replaced
the first one and lasted until the end of the trial. 75 ms after the appearance of the directional
distractor (a time interval in which the distractors were maximally effective, see Crostella et al,
2009 for details), the black square changed color to orange or blue. The color change was the
imperative instruction for the subjects to make an accurate and rapid pointing movement with their
right index finger toward one of the two lateral black squares (0.43° x 0.43° in size) positioned at

Porciello et al.�� � 8� �
8�
�
10.2° of eccentricity in the right and left visual field. The lateral squares were mounted on a
transparent Plexiglas board, which was placed vertically on the desk between the subjects and the
computer screen. Subjects were instructed to point toward the right target if the color of the
instruction cue changed to orange and, viceversa, toward the left target if the color of the instruction
cue changed to blue. At the end of the movement, they had to return to the resting position. The
colored cue remained visible until the end of the trial. Note that, subjects were continuously
instructed to ignore the distracting stimulus and focus their attention on the central square
throughout the trial. They were explicitly informed that the direction of the distractors did not
predict the imperative cue. This allowed us to measure attentional shifts independent of eye
movements. In fact, throughout the experiment eye position was constantly monitored using a
rearview mirror. We considered pointing movements erroneous if, before reaching the target, the
participant moved at least 2 cm in the wrong direction (i.e., the one indicated by the distractor)
instead of following the imperative cue.
2.3 Transcranial Magnetic Stimulation Procedure
To interfere with the neural activity of our sites of interest during the above-described task,
we employed an rdpTMS paradigm. Specifically, at the beginning of the experimental session,
before starting the hand-pointing task, subjects sat in a comfortable chair with their hands placed on
a pillow. They wore a tightly fitting bathing cap (on which the scalp stimulation points were
marked) and earplugs to guard against potential hearing damage during the stimulation. Motor-
evoked potentials (MEPs) were recorded from the first dorsal interosseous (FDI) muscle of the left
hand. An active surface Ag/AgCl electrode was placed over the muscle belly and the reference over
the interphalangeal joint. A ground electrode was placed on the dorsal surface of the left wrist.
Responses were amplified at a gain of 1000 by a Digitimer D360 amplifier (Digitimer,
Hertfordshire, UK). They were bandpass-filtered (20 Hz – 2.5 kHz) and digitized (sampling rate: 5
kHz) by means of a CED Power 1401 controlled with Spike 2 software (Cambridge Electronic

Porciello et al.�� � 9� �
9�
�
design, Cambridge, UK). The resting motor threshold (rMT), defined as the lowest intensity able to
evoke five out of ten MEPs with an amplitude of at least 50 mV, was determined by holding the
stimulation coil over the optimal controlateral scalp position (i.e., the motor cortex area
controlateral to the left hand producing the largest MEPs) for the FDI muscle. Stimulation sites
were identified on each participant’s scalp with the SofTaxic Navigator System (EMS, Bologna,
Italy), as in our previous TMS research (Avenanti, Annella, Candidi, Urgesi, & Aglioti, 2013;
Avenanti, Bolognini, Maravita, & Aglioti, 2007; Urgesi, Candidi, Ionta, & Aglioti, 2007). Skull
landmarks (nasion, inion and two preauricular points) and about 60 points providing a uniform
representation of the scalp were digitized using a Fastrak Polhemus digitizer (Polhemus, Colchester,
VT). Coordinates in Talairach space (Talairach & Tournoux, 1988) were automatically estimated by
the SofTaxic Navigator from an MRI-constructed stereotaxic template (see Fig.1). The scalp
locations that corresponded best to right FEF (Paus, 1996) and right PPC (see for a review Iacoboni,
2006) were identified and marked on the bathing cap with a pen.
---------------------------
Please insert Fig.1 near here
--------------------------
rdpTMS was applied by connecting two Magstim Mode 200 stimulators with a Bistim module (The
Magstim Company, Carmarthenshire, UK), producing a maximum output of 1.75T at the coil
surface (stimulus attenuation, 22%; duration, 1 ms; rise time, 110 ms). Two pulses were delivered
with an interstimulus interval (ISI) of 100 ms by means of a 70 mm figure-of-eight stimulation coil
(Magstim polyurethane-coated coil). In keeping with our hypothesis, the first TMS pulse was
delivered at 500 ms, simultaneously with the appearance of the directional distractor; the second
was delivered at 600 ms, 25 ms after the appearance of the imperative cue. Stimulation intensity

Porciello et al.�� � 10� �
10�
�
was supra-threshold and set at 120% of the rMT for both pulses and both stimulation sites; it ranged
from 37% to 65% (mean 51.3%) of the maximum stimulator output. These parameters were
consistent with published safety guidelines for TMS stimulation (Rossi et al., 2009). During
stimulation, the intersection of the coil was held tangentially to the scalp with the handle pointing
backward and laterally at a 45° angle from the middle sagittal axis of the participant’s head. Figure
2 shows a schematic representation of a typical event trial.
---------------------------
Please Insert Fig.2 near here
--------------------------
2.4 Experimental Design
We used an experimental block design consisting of 144 trials, with 48 trials per block.
Subjects were tested in three separate blocks in counterbalanced order; each block was associated
with a different type of distractor. To engage automatic responses and minimize expectations, inside
the block the directions of the distractors were equiprobable (50% congruent and 50% incongruent
with the imperative cue) and presented in a randomized sequence. In each block, Congruency
(congruent and incongruent trials) and Stimulation site (FEF and PPC) conditions were repeated 12
times. The stimulation coil was positioned on the rFEF or the rPPC according to a
pseudorandomized order. Before starting each experimental block, participants completed eight
practice trials, which were not included in the analysis. To avoid possible anticipations of the
stimuli, we inserted a random inter-trial interval (ITI) ranging from 5000 to 6000 ms.
2.5 Data Acquisition and Statistical Analysis
During the experiment, we recorded participants’ pointing movements using an
electromagnetic Pohlemus Fastrack (Pohlemus, USA) marker taped over the nail of their right index

Porciello et al.�� � 11� �
11�
�
finger. Hand trajectories were sampled at 100 Hz. A calibration procedure to set the spatial points of
reference was performed at the beginning of each experimental block. Data were collected and
analyzed off-line using specifically developed softwares.
For each experimental condition, we calculated individual mean RTs, velocity, acceleration and
accuracy (as percentage of correct responses) of pointing movements.
Pointing RTs (i.e., the latency between the appearance of the colored imperative cue and the
beginning of the movement toward the target), measured in ms, were calculated starting from a 1
cm increase on the vertical axis. We discarded from the analysis: trials in which participants started
moving with a latency lower than 100 ms (anticipations, < 1% ); trials in which participants did not
start moving within 600 ms from the imperative cue (delays, < 1%; see Sheliga, Craighero, Riggio
& Rizzolatti, 1997, for a similar approach); and trials in which participants touched the wrong target
(1.04%), probably due to attentional loss.
To simplify our description of the data, we computed a Cuing Effect (CE) index by subtracting
percentages of correct pointing responses obtained in incongruent conditions from the ones obtained
in the congruent conditions. For pointing RTs, velocity and acceleration the index was computed by
subtracting the congruent from the incongruent conditions. Thus, for all the analyzed variables, the
magnitude of the CE indicates the magnitude of the reflexive attentional capture/re-orienting cost
(i.e., high CE values indicate high attentional capture/re-orienting cost) mediated by distracting
stimuli.
Statistical differences between mean RTs, velocity, acceleration and accuracy responses for
pointing movements were tested using repeated measures ANOVA. We performed four separate 2 x
3 ANOVAs with Stimulation site (rFEF, rPPC) and Distractor (gaze, hand and arrow) as within-
subjects factors. Post hoc multiple comparisons were performed using the Duncan test. The
threshold for statistical significance was set at 0.05. All statistical tests were performed with
STATISTICA 7 software (StatSoft, Tulsa, OK, USA).

Porciello et al.�� � 12� �
12�
�
3. Results
Table 1 shows mean (± SEM) values of the different parameters (percent accuracy; RTs;
velocity, and acceleration) acquired during the experimental hand-pointing task. Each parameter is
reported as functions of Congruency (congruent vs incongruent), Stimulation site (rFEF vs rPPC)
and Distractor (eyes, hand, arrow).
3.1 Accuracy of hand-pointing responses
The ANOVA performed on accuracy showed no significant main effect of Stimulation site
[F(1, 11) = 0.69, p = .42] or Distractor [F(2, 22) = 1.64, p = .22]; and the interaction between
Distractor and Stimulation site was not significant [F(2, 22) = 0.30, p = .74], which indicates there
was no overall difference in the accuracy of pointing responses in our experimental conditions.
3.2 Hand-pointing RTs
The ANOVA performed on RTs revealed a significant main effect of the Distractor
[F(2,22)= 7.39; p = 0.00]. This was explained by the fact that during the hand-pointing task, hand
cuing interfered more than arrow cuing (p = 0.01) and gaze cuing marginally more than arrow cuing
(p = 0.055). No main effect of the Stimulation site was found in the ANOVA [F(1,11)= 0.90; p =
0.36].
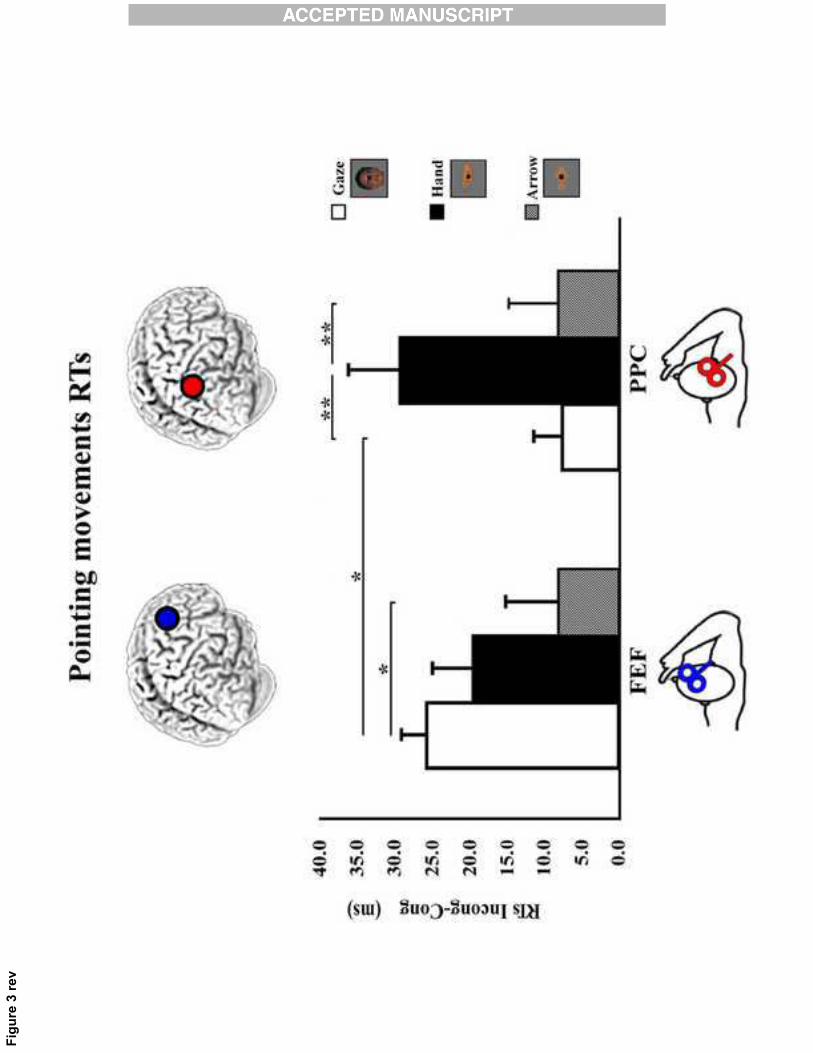
Crucial for testing the study's hypotheses was the significant interaction between Stimulation site
and Distractor [F(2,22)= 3.65; p = 0.04], (see Fig.3).
Post hoc analysis revealed that the significance of the interaction was explained by the higher CE
triggered by the pointing hand with respect to the other two distractors (all ps = 0.01) during the on-
line rPPC stimulation. Furthermore, during rFEF stimulation gaze-CE was significantly higher than
arrow-CE (p = 0.03) but not significantly different from the hand-CE (p = 0.41).
Finally, the effect of distracting gaze was higher for rFEF than rPPC stimulation (p = 0.04). No
differential effects of rFEF vs rPPC stimulation were found for distracting hands and arrows (all ps

Porciello et al.�� � 13� �
13�
�
> 0.22). In summary, the present results indicate that interfering with rPPC (but not with rFEF),
induced higher distracting effect for hands compared to both gaze and arrow directional stimuli.
---------------------------
Please Insert Fig.3 near here
--------------------------
3.3 Hand-pointing velocity and acceleration
The raw data for these two parameters are reported in Table 1. The ANOVA performed on
the velocity of pointing movements calculated by subtracting the congruent trials from the
incongruent ones showed no significant main effect for either Stimulation site [F(1,11) = 0.02, p =
.88] or Distractor [F(2,22) = 1.61, p = .22]; furthermore, the interaction between Distractor and
Stimulation site was not significant [F(2,22) = 1.19, p = .32], indicating no overall difference in the
velocity of pointing responses in our experimental conditions. Moreover, no significant differences
were found in the acceleration of pointing movements between the two Stimulation sites [F(1,11) =
0.02, p = .88] and the different types of Distractors [F(2,22) = 1.56, p = .23]. The interaction was
not significant as well [F(2,22) = 0.97, p = .39].
4. Discussion�
Several studies have investigated the neural substrates of social attention in human and non
human primates with particular emphasis on the role of the gaze (Callejas, Shulman, & Corbetta,
2014; Deaner et al., 2003; Engell et al., 2010; Greene, Mooshagian, Kaplan, Zaidel, & Iacoboni,
2009; Sato, Kochiyama, Uono, & Yoshikawa, 2009; Shepherd et al., 2009; Tipper, Handy,
Giesbrecht, & Kingstone, 2008). Yet, the functional and neural bases underlying these processes are
not completely understood. For example, whether body-evoked attentional shifts recruit the same

Porciello et al.�� � 14� �
14�
�
brain areas as the symbolic cues, or involve specialized systems or wider networks is still an open
and hotly debated question. It has been suggested, mostly by gaze cuing paradigms, that attentional
shifts triggered by body-related cues recruit different brain regions compared to symbolic cues since
they activate cortical and subcortical areas involved in the: orienting and re-orienting of attention
(i.e., intra-parietal sulcus/superior parietal lobule (IPS/SPL), frontal eye field (FEF)), face
perception (i.e., fusiform gyrus and superior temporal sulcus (STS)), gaze perception (i.e.., STS and
occipito-temporal regions), imitation (the action observation network (AON)), emotion and mental
state attribution (i.e., amygdala, medial prefrontal cortex (mPFC) and temporal-parietal junction
(TPJ)). Coherent with this hypothesis, seminal studies by Perrett et al. (1985) found that a defined
population of neurons present in the monkey STS is responsive to the gaze direction, a result
confirmed also in humans (Allison, Puce, & McCarthy, 2000; Hoffman & Haxby, 2000). Thus, the
difference between attentional shifts triggered by gaze cues compared to symbolic cues is also
explained by the fact that face/gaze processing regions are not only part of a wider socio-attentional
system but also modulate the supramodal attentional networks consisting of dorsal and ventral
fronto-parietal nodes (see Callejas et al., 2014; Nummenmaa, Passamonti, Rowe, Engell, & Calder,
2010). Relevant to this issue, studies in human and non human primates (Grosbras, Laird, & Paus,
2005; Shepherd et al., 2009) support the notion that “mirroring networks” are also involved in
social attention. Gaze following, for example, may lead to the sharing of attention between
individuals thanks to mirroring mechanisms. Strong evidence supporting this theory comes from the
single cell recording study in the monkey lateral intraparietal cortex a structure that is heavily
connected to frontal eye field and is involved in the organization of eye movements (Shepherd et
al., 2009). By requiring monkeys to perform a modified version of a Posner cueing task using
saccadic responses, the authors demonstrate that neurons in LIP map shifts of attention in both self
and others (Shepherd et al., 2009). Specifically, 30 out of 153 neurons, recorded in LIP, responded
not only when monkeys oriented attention towards the receptive field of a given neuron, but also

Porciello et al.�� � 15� �
15�
�
when they observed a conspecific gazing towards the same direction. Interestingly a second
population of neurons recorded in the same area suppressed activity in response to the observed
gaze cues, possibly to suppress task-irrelevant behavioral responses triggered by the observed gaze,
thereby maintaining the fixation to successfully complete the task.
To better investigate the causal role of fronto-parietal regions (i.e., rFEF and rPPC) in the reflexive
shifts of social attention triggered by gaze, pointing hands or arrows,�we used an online TMS event-
related design while participants performed directional pointing movements, instructed by
imperative cues. This approach allowed us to provide direct causal evidence about the differential
influence of the frontal and parietal cortices in modulating reflexive shifts of social attention.
Accordingly, results show that interfering with the normal activity of the rPPC differentially
affected hand-pointing responses depending on whether the directional distractor was an oriented
hand, gaze or arrow. In particular, increased manual RTs were recorded when subjects had to inhibit
the automatic tendency to follow distracting hands with respect to the distracting gaze and arrows
conditions. Thus, a main point of novelty in the present study is that the rPPC has an active role in
controlling the ability to ignore specific stimuli (i.e., hands) signaling a direction different from that
indicated by a non-directional central cue in the condition of distractor-effector directional
mismatch. The interfering effect induced by distracting hands on pointing gesture programming
preceded the imperative instruction to move (the SOA was -75 ms and corresponded with the
delivery of the first TMS pulse) and is coherent with the idea that the PPC plays an early role in
action control (i.e., Della-Maggiore, Malfait, Ostry, & Paus, 2004; Glover, Miall, & Rushworth,
2005).
In contrast, rdpTMS over the FEF resulted in less clear effects, eliciting an increased
interference of incongruent gaze only compared to a non-body related distractor (directional arrow),
which was used as control stimulus. In keeping with our predictions, rFEF and rPPC did not exert a
specific modulation of attentional shifts triggered by directional arrows.

Porciello et al.�� � 16� �
16�
�
This pattern of results fits with the hypothesis that social attention relies on a fronto-parietal system
and that, according to our social somatotopy hypothesis, activity of different nodes in this network
is modulated by the correspondence between the observed distractor and the effector used for
responding.
This result goes in parallel with the BOLD signal recorded in our previous fMRI data. Here, higher
activity in the left and right IPS regions was associated with distracting hands in the hand-pointing
task and, viceversa, higher bilateral activation in the FEFs was associated with distracting gaze
during the saccadic response task (Cazzato et al., 2012). It is relevant to note that the increased
BOLD signal in the incongruent vs the congruent condition may reflect the cost of inhibiting the
preponderant tendency to follow distracting cues. This is in keeping with influential models of
attentional re-orienting that hint at a crucial the role of a dorsal frontoparietal network specialized
for selecting and linking stimuli and responses and crucially activated during both endogenous and
exogenous orienting of attention (Corbetta, Patel, & Shulman, 2008). Thus, our study suggests that
the transitory disruption of the PPC activity reduces the control function that this structure normally
exerts on distracting stimuli following body-centered rules.
The fact that in the fMRI study we found higher activity specifically in the IPS and not the PPC can
be explained by considering that the fMRI pointing-task performed by participants was defined as
an index finger extension towards the right or the left button key with the shoulder and the arm
immobilized instead of a free hand-arm movement in space.
Concerning our target areas, studies indicate that fronto-parietal circuits are involved in mirroring
others' actions (Rizzolatti and Sinigaglia, 2010), as well as in attentional re-orienting (Corbetta et
al., 2008; Corbetta & Shulman, 2002). More specifically, mirroring observed actions seems to be
linked to inferior parietal lobule, inferior frontal gyrus and the adjacent ventral premotor cortex, as
well as primary visual cortex, cerebellum and parts of the limbic system, (see Molenberghs,
Cunnington, & Mattingley, 2012, for an extensive review) while attentional re-orienting relies upon

Porciello et al.�� � 17� �
17�
�
a dorsal (i.e., intra-parietal sulcus/superior parietal lobule (IPS/SPL) and frontal eye field (FEF))
and a ventral (i.e., temporal-parietal junction (TPJ) and ventral frontal cortex/insula) fronto-parietal
circuit. Therefore, while action mirroring and attentional re-orienting are not underpinned by the
very same areas, overlapping nodes of fronto-parietal systems seem to mediate fundamental aspects
of social attention.
4.1 Hand-centered coordinate systems for controlling reflexive shifts of social attention in the
posterior parietal cortex
The posterior parietal cortex is a complex and extended area that integrates multiple sensory
inputs with motor outputs and is crucial for the spatial and temporal processes underlying a variety
of different functions including planning movements, guiding attention, mirroring and
understanding actions and even making decisions (Andersen, Snyder, Bradley, & Xing, 1997;
Galati et al., 2011; Iacoboni, 2006; Rushworth, Johansen-Berg, Göbel, & Devlin, 2003). Relevant to
the present research are non-human and human primate studies showing that distinct sectors of the
posterior parietal cortex around the IPS have visuo-motor control over specific effectors, depending
on the type of movements an individual plans to implement (Andersen & Buneo, 2002; Caminiti et
al., 2010; Culham & Valyear, 2006; Grefkes & Fink, 2005; Konen, Mruczek, Montora, & Kastner,
2013). For example, studies in monkeys show that saccadic movements are preferentially linked to
the neural activity in the lateral intraparietal area, whereas grasp and reach movements are linked to
the activity in the anterior intraparietal area and parietal reach region, respectively (Andersen &
Buneo, 2002; Fattori et al., 2009; Snyder, Batista, & Andersen, 1997; Platt & Glimcher, 1998). For
a more direct evidence, a recent monkey study shows that injections of muscimol, a GABAA
agonist that suppresses local neuronal activity, in the monkey parietal reach area led to temporary
impairment of reaching movements with preservation of saccadic movements (Hwang, Hauschild,
Wilke, & Andersen, 2012). A pattern of results that is reminiscent of the deficits found in patients
with optic ataxia following lesions of the parietal cortex, who showed impairments during arm-

Porciello et al.�� � 18� �
18�
�
movements, but not saccades, towards visual objects (D’Esposito, 2003; Mesulam, 2000; Perenin,
& Vighetto, 1988). Analogously, fMRI studies in humans suggest the presence of pointing/reach-
selective (more than saccade-selective) areas, in specific clusters of parietal cortices (Cavina-Pratesi
et al., 2010; Connolly, Andersen, & Goodale, 2003; Filimon, Nelson, Huang, & Sereno, 2009;
Prado et al., 2005). Although there is abundant correlational evidence for this differentiation in
humans, causative evidence is comparatively meager. Stimulation studies indicate that the posterior
parietal area is causally involved in updating reaching movements to compensate for jumps in target
location (Desmurget et al., 1999), during the learning of new movement trajectories (Della-
Maggiore et al., 2004) and in hand-related attention-switching tasks (Rushworth et al., 2001).
Moreover, selective interference on specific movements was demonstrated in a recent online
interferential rTMS study in which three high-frequency pulses were delivered to different parietal
regions while healthy subjects planned either a saccadic or a reaching task (Vesia, Prime, Yan,
Sergio, & Crawford, 2010). Interference with the midposterior intraparietal sulcus and angular
gyrus affected both reaches and saccades, whereas interference with a superior parieto-occipital
region affected reaches but not saccades, thus providing clear causative evidence for body-part
effector-based action planning in PPC (Vesia et al., 2010).
In the present study, we demonstrate that stimulation of a posterior parietal region, supposedly
linked to control of planning and execution of upper limb movements, interferes with directional
pointing movements instructed by a central cue by increasing the effect of the conflict between the
central cue and the direction of a distracting hand.
Specifically, these data hint that the PPC plays a crucial role in controlling the reorienting of
attention when people have to plan hand movements in the opposite direction respect to the
direction of signaling hands. It is worth noting that, based on our behavioral (Crostella et al, 2009)
and fMRI (Cazzato et al, 2012) data, the so-called “social somatotopy” model of reflexive social
attention predicts that in tasks where the hand/arm movements are used to respond (like in the

Porciello et al.�� � 19� �
19�
�
present study), the disruption of neural processing in PPC mainly alters the control of attentional
shifts triggered by hand distractors.
4.2 Distracting gaze and temporal disruption of the frontal eye field
Although fMRI studies indicate that the human frontal eye field may be similarly involved
in attending, looking and pointing (Astafiev et al., 2003; Hagler, Riecke, & Sereno, 2007) and may
lack effector specificity (Levy, Schluppeck, Heeger, & Glimcher, 2007), lesions of this region
induce defects in saccadic generation (Latto & Cowey, 1971; Rizzolatti, 1983) and electric
stimulation evokes saccadic movements (Godoy, Luders, Dinner, Morris, & Wyllie, 1990) in the
absence of influences on hand movement preparation. In the present study, in which participants
performed the task using their hands, rdpTMS over rFEF increased the interference induced by a
distracting gaze with respect to that triggered by non-biological cues. Therefore, although there is
an attentional capture of the oriented gaze, stimulation of FEF activity led to less clear effects
compared of stimulation of PPC. This difference may be due to the fact that participants responded
using their hands instead of their eyes. This interpretation is supported by a companion study which
used saccadic instead of hand movements and showed that TMS interference with FEF (not with
rPPC) activity selectively increased the distracting power triggered by gaze compared to that
triggered by hands and arrows (Crostella, et al. 2013). All together, our results provide the first
causative evidence in humans that the attentional fronto-parietal network responsible of the
reorienting of visuo-spatial attention works according to body-part centered frames of reference and
expand our previous behavioral and fMRI findings (Cazzato et al., 2012; Crostella et al., 2009).
Because in this type of task participants had to ignore distracting body-related stimuli that
could be congruent or incongruent with the motor response instructed by the imperative cue, the
fronto-parietal re-orienting system could be modulated by the activation or the suppression of
mirror mechanisms (e.g. Shepherd et al., 2009). Indeed, although overlapping, these two systems

Porciello et al.�� � 20� �
20�
�
are not isomorphic and further studies are necessary to explore their role in coding attentional and
motor aspects of social inter-actions.
Conclusions
To sum up, our results provide evidence in humans of the pivotal and causal role of rPPC in
controlling reflexive shifts of visuo-spatial attention triggered by directional pointing hands but not
by oriented gaze or arrows. The finding that interference with FEF activity leads to a less specific
effect (e.g. gaze and hand distractors were not significantly different) may be related to the fact that
participants always used pointing responses in this study. In line with this, evidence from our
laboratory suggests that disrupting the activity of FEF may have a specific effect on the attentional
reorienting triggered by gaze cuing when saccadic responses are requested (Crostella et.al., 2013).
Thus, according to our “somatotopy” model of reflexive social attention, the preferential neural
sensitivity of the rPPC in controlling attentional re-orienting mediated by social cues reflects the
relation between specific body-related distractors (hands) and responding body parts (arm/hand
pointing movements).
Acknowledgments
This research was supported by EU Information and Communication Technologies Grant (VERE
project, FP7-ICT-2009-5, Prot. Num. 257695); Istituto Italiano di Tecnologia (IIT) SEED (Prot.
Num. 21538) and the Italian Ministry of Health (RF-2010-2312912). We are thankful to Brittany
Serra Holmes for her help in manuscript editing.

Porciello et al.�� � 21� �
21�
�
References
Allison, T., Puce, A., & McCarthy, G. (2000). Social perception from visual cues : role of the STS region. Trends in Cognitive Sciences, 4 (7), 267-278.
Andersen, R. A., Snyder, L. H., Bradley, D. C., & Xing, J. (1997). Multimodal Representation of Space in the Posterior Parietal Cortex and Its Use in Planning Movements. Annual Review of Neuroscience, 20(1), 303–330.
Andersen, R. A., & Buneo, C. A. (2002). Intentional maps in posterior parietal cortex. Annual review of neuroscience, 25, 189–220.
Ariga, A., & Watanabe, K. (2009). What is special about the index finger?: The index finger advantage in manipulating reflexive attentional shift. Japanese Psychological Research, 51(4), 258–265.
Astafiev, S. V, Shulman, G. L., Stanley, C. M., Snyder, A. Z., Essen, D. C. Van, & Corbetta, M. (2003). Functional Organization of Human Intraparietal and Frontal Cortex for Attending , Looking , and Pointing. The Journal of Neuroscience, 23(11), 4689–4699.
Avenanti, A., Bolognini, N., Maravita, A., & Aglioti, S. M. (2007). Somatic and motor components of action simulation. Current Biology, 17(24), 2129–35.
Avenanti, A., Annella, L., Candidi, M., Urgesi, C., & Aglioti, S. M. (2013). Compensatory plasticity in the action observation network: virtual lesions of STS enhance anticipatory simulation of seen actions. Cerebral Cortex, 23(3), 570–80.
Baron-Cohen, S. (1995). The eye direction detector (EDD) and the shared attention mechanism (SAM): two cases for evolutionary psychology. In: Moore C, Dunham P (eds) Joint attention: its origins and role in development. Lawrence Erlbaum Associates, Hillsdale, 41-59.
Birmingham, E., & Kingstone, A. (2009). Human social attention: A new look at past, present, and future investigations. Annals of the New York Academy of Sciences, 1156, 118–40.
Briggs, G.G. & Nebes, R.D. (1975). Patterns of hand preference in a student population. Cortex, 11, 230–238.
Callejas, A., Shulman, G. L., & Corbetta, M. (2014). Dorsal and ventral attention systems underlie social and symbolic cueing. Journal of Cognitive Neuroscience, 26(1), 1–25.
Caminiti, R., Chafee, M. V, Battaglia-Mayer, A., Averbeck, B. B., Crowe, D. A., & Georgopoulos, A. P. (2010). Understanding the parietal lobe syndrome from a neurophysiological and evolutionary perspective. The European Journal of Neuroscience, 31(12), 2320–40.
Cavina-Pratesi, C., Monaco, S., Fattori, P., Galletti, C., McAdam, T. D., Quinlan, D. J., Goodale, M. A., et al. (2010). Functional magnetic resonance imaging reveals the neural substrates of arm transport and grip formation in reach-to-grasp actions in humans. The Journal of Neuroscience, 30(31), 10306–23.
Cazzato, V., Macaluso, E., Crostella, F., & Aglioti, S. M. (2012). Mapping reflexive shifts of attention in eye-centered and hand-centered coordinate systems. Human Brain Mapping, 33(1), 165–78.

Porciello et al.�� � 22� �
22�
�
Connolly, J. D., Andersen, R. A., & Goodale, M. A. (2003). FMRI evidence for a “parietal reach region” in the human brain. Experimental Brain Research, 153(2), 140–5.
Corbetta, M. (1998). Frontoparietal cortical networks for directing attention and the eye to visual locations: identical, independent, or overlapping neural systems? Proceedings of the National Academy of Sciences of the United States of America, 95(3), 831–8.
Corbetta, M., & Shulman, G. L. (2002). Control of goal-directed and stimulus-driven attention in the brain. Nature reviews. Neuroscience, 3(3), 201–15.
Corbetta, M., Patel, G., & Shulman, G. L. (2008). The reorienting system of the human brain: from environment to theory of mind. Neuron, 58(3), 306–24.
Crostella, F., Carducci, F., & Aglioti, S. M. (2009). Reflexive social attention is mapped according to effector-specific reference systems. Experimental Brain Research, 197(2), 143–51.
Crostella, F., Porciello, G., Carducci, F. & Aglioti, S.M. (unpublished results). Transcranial Magnetic Stimulation over Frontal Eye Field interferes with reflexive shifts of attention induced by averted gaze. Magstim Neuroenhancement Conference & Workshop 2013, Oxford, UK, 3-5, 2013.
Culham, J. C., & Valyear, K. F. (2006). Human parietal cortex in action. Current Opinion in Neurobiology, 16(2), 205–212.
D’Esposito, M. (2003). Neurological Foundations of Cognitive Neuroscience (Cambridge, MA: MIT Press).
Deaner, R. O., Platt, M. L., & Carolina, N. (2003). Reflexive Social Attention in Monkeys and Humans. Current Biology, 13, 1609–1613.
Della-Maggiore, V., Malfait, N., Ostry, D. J., & Paus, T. (2004). Stimulation of the posterior parietal cortex interferes with arm trajectory adjustments during the learning of new dynamics. The Journal of Neuroscience, 24(44), 9971–6.
Desmurget, M., Epstein, C. M., Turner, R. S., Prablanc, C., Alexander, G. E., & Grafton, S. T. (1999). Role of the posterior parietal cortex in updating reaching movements to a visual target. Nature Neuroscience, 563–567.
Driver, J., Davis, G., Ricciardelli, P., Kidd, P., Maxwell, E., & Baron-Cohen, S. (1999). Gaze Perception Triggers Reflexive Visuospatial Orienting. Visual Cognition, 6, 509–540.
Dunbar, R. I. M., & Shultz, S. (2007). Evolution in the social brain. Science, 317(5843), 1344–1347.
Emery, N. J. (2000). The eyes have it : the neuroethology, function and evolution of social gaze. Biobehavioral Reviews, 24, 581-604.
Engell, A. D., Nummenmaa, L., Oosterhof, N. N., Henson, R. N., Haxby, J. V, & Calder, A. J. (2010). Differential activation of frontoparietal attention networks by social and symbolic spatial cues. Soc Cogn Affect Neurosci, 5(4), 432–440.
Fattori, P., Breveglieri, R., Marzocchi, N., Filippini, D., Bosco, A., & Galletti, C. (2009). Hand orientation during reach-to-grasp movements modulates neuronal activity in the medial posterior parietal area V6A. The Journal of Neuroscience, 29(6), 1928–36.

Porciello et al.�� � 23� �
23�
�
Filimon, F., Nelson, J. D., Huang, R.-S., & Sereno, M. I. (2009). Multiple parietal reach regions in humans: cortical representations for visual and proprioceptive feedback during on-line reaching. The Journal of Neuroscience, 29(9), 2961–71.
Fischer, M. H., & Szymkowiak, A. (2004). Joint attention for pointing but not grasping postures. Cortex, 40(1), 168–70.
Forster, B., & Eimer, M. (2007). Covert unimanual response preparation triggers attention shifts to effectors rather than goal locations. Neuroscience letters, 419(2), 142–6.
Friesen, C. K., & Kingstone, A. (1998). The eyes have it ! Reflexive orienting is triggered by nonpredictive gaze. Psychonomic Bulletin & Review, 5(3), 490–495.
Galati, G., Committeri, G., Pitzalis, S., Pelle, G., Patria, F., Fattori, P., & Galletti, C. (2011). Intentional signals during saccadic and reaching delays in the human posterior parietal cortex. The European Journal of Neuroscience, 34(11), 1871–85.
Glover, S., Miall, R. C., & Rushworth, M. F. S. (2005). Parietal rTMS disrupts the initiation but not the execution of on-line adjustments to a perturbation of object size. Journal of Cognitive Neuroscience, 17(1), 124–36.
Godoy, J., Luders, H., Dinner, D.S., Morris, H.H. & Wyllie, E., (1990). Versive eye movements elicited by cortical stimulation of the human brain. Neurology, 40, 296-299.
Greene, D. J., Mooshagian, E., Kaplan, J. T., Zaidel, E., & Iacoboni, M. (2009). The neural correlates of social attention: automatic orienting to social and nonsocial cues. Psychological Research, 73(4), 499–511.
Grefkes, C., & Fink, G. R. (2005). The functional organization of the intraparietal sulcus in humans and monkeys. Journal of anatomy, 207(1), 3–17.
Grosbras, M.-H., Laird, A. R., & Paus, T. (2005). Cortical regions involved in eye movements, shifts of attention, and gaze perception. Human Brain Mapping, 25(1), 140–54.
Hagler, D. J., Riecke, L., & Sereno, M. I. (2007). Parietal and superior frontal visuospatial maps activated by pointing and saccades. NeuroImage, 35(4), 1562–77.
Hoffman, E. A., & Haxby, J. V. (2000). Distinct representations of eye gaze and identity in the distributed human neural system for face perception. Nature Neuroscience, 3(1), 80-84.
Hwang, E. J., Hauschild, M., Wilke, M., & Andersen, R. A. (2012). Inactivation of the Parietal Reach Region Causes Optic Ataxia, Impairing Reaches but Not Saccades. Neuron, 76(5), 1021–1029.
Iacoboni, M. (2006). Visuo-motor integration and control in the human posterior parietal cortex: evidence from TMS and fMRI. Neuropsychologia, 44(13), 2691–9.
Kapoula, Z., Yang, Q., Sabbah, N., & Vernet, M. (2011). Different effects of double-pulse TMS of the posterior parietal cortex on reflexive and voluntary saccades. Frontiers in Human Neuroscience, 114(5), 1-9.
Kingstone, A. (2009). Taking a real look at social attention. Current opinion in neurobiology, 19(1), 52–6.

Porciello et al.�� � 24� �
24�
�
Klein, J. T., Shepherd, S. V, & Platt, M. L. (2009). Social attention and the brain. Current Biology, 19(20), R958–62.
Konen, C. S., Mruczek, R. E. B., Montora, J. L., & Kastner, S. (2013). Functional organization of human posterior parietal cortex: grasping- and reaching-related activations relative to topographically organized cortex. Journal of Neurophysiology. 109(12):2897-908.
Langton, S.R.H., & Bruce, V. (2000) You must see the point: automatic pro- cessing of cues to the direction of social attention. J Exp Psychol Hum Percept Perform 26:747–757.
Latto, R., & Cowey, A. (1971). Visual field defects after frontal eye-field lesions in monkeys. Brain Res, 30, 1-24.
Levy, I., Schluppeck, D., Heeger, D. J., & Glimcher, P. W. (2007). Specificity of human cortical areas for reaches and saccades. The Journal of Neuroscience, 27(17), 4687–96.
Liszkowski, U., Carpenter, M., Henning, A., Striano, T., & Tomasello, M. (2004). Twelve-month-olds point to share attention and interest. Developmental science, 7(3), 297–307.
Mesulam, M.M. (2000). Principles of Behavioral and Cognitive Neurology (Oxford: Oxford University Press).
Molenberghs, P., Cunnington, R., & Mattingley, J. B. (2012). Brain regions with mirror properties: a meta-analysis of 125 human fMRI studies. Neuroscience and Biobehavioral Reviews, 36(1), 341–9.
Nummenmaa, L., Passamonti, L., Rowe, J., Engell, A. D., & Calder, A. J. (2010). Connectivity analysis reveals a cortical network for eye gaze perception. Cerebral Cortex, 20(8), 1780–7.
O’Shea, J., Muggleton, N. G., Cowey, A., & Walsh, V. (2004). Timing of target discrimination in human frontal eye fields. Journal of Cognitive Neuroscience, 16(6), 1060–1067.
Paus, T. (1996). Location and function of the human frontal eye-field: a selective review. Neuropsychologia, 34(6).
Perenin, M.T., & Vighetto, A. (1988). Optic ataxia: a specific disruption in visuomotor mechanisms. I. Different aspects of the deficit in reaching for objects. Brain, 111, 643-674.
Perrett, D. I., Smith, P. A. J., Potter, D. D., Mistlin, A. J., Head, A. S., Milner, A. D., & Jeeves, M. A. (1985). Visual Cells in the Temporal Cortex Sensitive to Face View and Gaze Direction. Proceedings of the Royal Society of London Series B, Biological Sciences, 223, 293–317.
Platt M.L. & Glimcher P.W. (1998). Response fileds of intraparietal neurons quantified with multiple saccadic targets. Experimental Brain Research, 121, 65-75.
Prado, J., Clavagnier, S., Otzenberger, H., Scheiber, C., Kennedy, H., & Perenin, M.-T. (2005). Two cortical systems for reaching in central and peripheral vision. Neuron, 48(5), 849–58.
Ricciardelli, P., Bricolo, E., Aglioti, S. M., & Chelazzi, L. (2002). My eyes want to look where your eyes are looking : Exploring the tendency to imitate another individual’ s gaze. Neuroreport, 13(17), 2259–2264.

Porciello et al.�� � 25� �
25�
�
Rizzolatti, G., Matelli, M. & Pavesi, G. (1983). Deficits in attention and movement following the removal of postarcuate (area 6) and prearcuate (area 8) cortex in macaque monkeys. Brain, 106, 655-673.
Rohlfing, K. J., Longo, M. R., & Bertenthal, B. I. (2012). Dynamic pointing triggers shifts of visual attention in young infants. Developmental Science, 3(15), 426–435.
Rossi, S., Hallett, M., Rossini, P. M., & Pascual-Leone, A. (2009). Safety, ethical considerations, and application guidelines for the use of transcranial magnetic stimulation in clinical practice and research. Clinical Neurophysiology, 120(12), 2008–39.
Rushworth, M. F. S., Paus, T., & Sipila, P. K. (2001). Attention Systems and the Organization of the Human Parietal Cortex. The Journal of Neuroscience, 21(14), 5262–5271.
Rushworth, M., Johansen-Berg, H., Göbel, S., & Devlin, J. (2003). The left parietal and premotor cortices: motor attention and selection. NeuroImage, 20, S89–S100.
Sato, W., Kochiyama, T., Uono, S., & Yoshikawa, S. (2009). Commonalities in the neural mechanisms underlying automatic attentional shifts by gaze, gestures, and symbols. NeuroImage, 45(3), 984–92.
Sheliga, B. M., Craighero, L., Riggio, L., & Rizzolatti, G. (1997). Effects of spatial attention on directional manual and ocular responses. Experimental Brain Research, 114(2), 339–51.
Shepherd, S. V, Klein, J. T., Deaner, R. O., & Platt, M. L. (2009). Mirroring of attention by neurons in macaque parietal cortex. Proceedings of the National Academy of Sciences of the United States of America, 106(23), 9489–94.
Shepherd, S. V. (2010). Following gaze: gaze-following behavior as a window into social cognition. Frontiers in integrative neuroscience, 4(5) 1-13.
Snyder, L. H., Batista, A. P., & & Andersen, R. H. (1997). Coding of intention in the posterior parietal cortex. Nature, 368.
Talairach, J. & Tournoux, P. (1988). Co-planar stereotaxic atlas of the human brain: 3-dimensional proportional system-an approach to cerebral imaging. Thieme, New York.
Tipper, C. M., Handy, T. C., Giesbrecht, B., & Kingstone, A. (2008). Brain Responses to Biological Relevance. Journal of Cognitive Neuroscience, 879–891.
Urgesi, C., Candidi, M., Ionta, S., & Aglioti, S. M. (2007). Representation of body identity and body actions in extrastriate body area and ventral premotor cortex. Nature Neuroscience, 10(1), 30–1.
Vesia, M., Prime, S. L., Yan, X., Sergio, L. E., & Crawford, J. D. (2010). Specificity of human parietal saccade and reach regions during transcranial magnetic stimulation. The Journal of Neuroscience, 30(39), 13053–65.
Wardak, C., Olivier, E., & Duhamel, J.-R. (2011). The relationship between spatial attention and saccades in the frontoparietal network of the monkey. The European Journal of Neuroscience, 33(11), 1973–81.
Whiten, A. (1998). Evolutionary and developmental origins of the mindreading system. In J. Langer & M. Killen (Eds.), Piaget, evolution, and development (pp. 73–99). Mahwah, NJ: Erlbaum.

Porciello et al.�� � 26� �
26�
�
Wipfli, M., Felblinger, J., Mosimann, U. P., Hess, C. W., Schlaepfer, T. E., & Muri, R. M. (2001). Double-pulse transcranial magnetic stimulation over the frontal eye field facilitates triggering of memory-guided saccades. The European Journal of Neuroscience, 14(3), 571–575.
Table B.1: Table shows mean (±SEM) values of the different parameters: (a.) accuracy, (b.) RTs, (c.) velocity and (d.) acceleration of pointing responses. Each parameter is reported as a function of Stimulation Site, Congruency and Distractor. The last two columns report the Cuing Effect, as a function of Stimulation Site and Distractor, obtained by subtracting congruent trials from incongruent ones, with regard of the accuracy in which was obtained by subtracting the incongruent trials from the congruent ones to avoid negative numbers.
a. FEF PPC FEF PPC % Correct Responses
Congruent Incongruent Congruent Incongruent C-I C-I
Gaze 93.75 (1.5) 75.69 (5.4) 94.44 (1.6) 75.69 (5.8) 18.06 (5.6) 18.75 (5.3) Hand 90.28 (4.1) 77.78 (5.2) 90.97 (1.9) 77.78 (6.0) 12.50 (4.8) 13.19 (5.8) Arrow 92.36 (2.6) 72.92 (5.8) 95.14 (2.2) 69.44 (7.0) 19.44 (6.1) 25.69 (6.6)
b. RTs Congruent Incongruent Congruent Incongruent I-C I-C Gaze 281.88 (15.3) 307.50 (17.4) 287.85 (14.1) 295.54 (17.0) 25.63 (3.6) 7.69 (3.8) Hand 293.16 (16.4) 312.68 (18.7) 295.21 (16.2) 324.41 (19.5) 19.52 (5.5) 29.20 (7.0) Arrow 304.19 (14.3) 312.28 (17.4) 300.03 (13.1) 308.02 (15.4) 8.09 (7.1) 7.99 (6.8)
c. Velocity Congruent Incongruent Congruent Incongruent I-C I-C Gaze 118.59 (5.8) 120.49 (5.1) 122.26 (5.0) 120.84 (4.7) 1.90 (1.3) -1.42 (1.6) Hand 122.58 (5.0) 125.43 (4.9) 123.61 (4.9) 126.45 (5.1) 2.84 (2.2) 2.84 (2.4) Arrow 122.54 (6.3) 119.83 (5.6) 121.51 (6.8) 122.84 (5.8) -2.71 (2.1) 1.33 (2.8)
d. Acceleration Congruent Incongruent Congruent Incongruent I-C I-C Gaze 303.99 (23.9) 307.66 (19.5) 319.35 (22.0) 304.84 (18.2) 3.67 (9.4) -14.51 (11.0) Hand 321.62 (20.5) 325.87 (23.1) 322.92 (19.2) 338.77 (23.8) 4.25 (10.8) 15.84 (12.5) Arrow 324.42 (26.9) 306.27 (22.7) 318.35 (31.2) 310.59 (24.5) -18.15 (13.3) -7.77 (15.1)

Porciello et al.�� � 27� �
27�
�
Appendices:
A. Figure captions:
Fig.A.1 Talairach coordinates of stimulated sites corresponding to right FEF and PPC.
For each participant, stereotaxic coordinates corresponding to the targeted areas were obtained with
the SofTaxic neuronavigation system.
Fig.A.2 Structure of a typical trial.
Example of a typical trial, with the three possible distractors (gaze, hand, arrow). Repetitive dual-
pulse transcranial magnetic stimulation (dpTMS) with an interstimulus interval (ISI) of 100 ms, was
applied on one of the two cortical sites in each trial at 500 ms, simultaneously with onset of the
directional distractor (congruent or incongruent with the direction of the central imperative cue).
Subjects had to point toward the right or the left target according to the color change, i.e., orange or
blue, of the central cue.
Fig.A.3 RTs (mean ±SEM) of the interaction Type of Distractor x Stimulation site.
* p � .05; ** p � .01
Highlights:
� We used TMS to explore the causative role of FEF and PPC in social attention
� TMS was delivered during a pointing-task instructed by a central imperative cue
� TMS over PPC selectively increases attentional shifts triggered by distracting hands�
� Shifts of social attention are somatocentrically mapped in the frontoparietal network
�
Figu
re 1

Figu
re 2
Figu
re 3
rev



















