
rone charles maranho - Universidade Estadual de Maringá
54
RONE CHARLES MARANHO DIVERSIDADE GENÉTICA ENTRE E DENTRO DE VARIEDADES DE CANA-DE-AÇÚCAR (Saccharum spp.) UTILIZANDO MARCADORES MICROSSATÉLITES MARINGÁ PARANÁ – BRASIL JUNHO – 2012
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of rone charles maranho - Universidade Estadual de Maringá

RONE CHARLES MARANHO
DIVERSIDADE GENÉTICA ENTRE E DENTRO DE VARIEDADES DE CANA-DE-AÇÚCAR (Saccharum spp.) UTILIZANDO MARCADORES
MICROSSATÉLITES
MARINGÁ PARANÁ – BRASIL
JUNHO – 2012

RONE CHARLES MARANHO
DIVERSIDADE GENÉTICA ENTRE E DENTRO DE VARIEDADES DE CANA-DE-AÇÚCAR (Saccharum spp.) UTILIZANDO MARCADORES
MICROSSATÉLITES
Dissertação apresentada à Universidade Estadual de Maringá, como parte das exigências do Programa de Pós-Graduação em Agronomia, para obtenção do título de Mestre.
MARINGÁ PARANÁ – BRASIL
JUNHO – 2012

Dados Internacionais de Catalogação-na-Publicação (CIP) (Biblioteca Central - UEM, Maringá – PR., Brasil) Maranho, Rone Charles M311d Diversidade genética entre e dentro de variedades
de cana-de-açúcar (Saccharum spp.) utilizando marcadores microssatélites / Rone Charles Maranho. -- Maringá, 2012.
47 f. : il. col., figs., tabs. Orientador: Profª. Drª. Maria de Fátima Pires
Machado da Silva. Dissertação (mestrado) - Universidade Estadual de
Maringá, Centro de Ciências Agrárias, Programa de Pós-Graduação em Agronomia, 2012.
1. Saccharum spp. 2. Marcador SSR. 3. Diversidade
genética. I. Silva, Maria de Fátima Pires Machado da, orient. II. Universidade Estadual de Maringá. Centro de Ciências Agrárias. Programa de Pós-Graduação em Agronomia. III. Título.
CDD 21.ed. 633.6753
AHS-00926

RONE CHARLES MARANHO
DIVERSIDADE GENÉTICA ENTRE E DENTRO DE VARIEDADES DE CANA-DE-AÇÚCAR (Saccharum spp.) UTILIZANDO MARCADORES
MICROSSATÉLITES
Dissertação apresentada à Universidade Estadual de Maringá, como parte das exigências do Programa de Pós-Graduação em Agronomia, para obtenção do título de Mestre.
APROVADA EM JUNHO 2012

ii
A meus pais José Maranho e Inervina Barizon Maranho, meu irmão
Gustavo Barizon Maranho, minha amiga, companheira e namorada Lígia
Adriana Cavalcante Jovino, pela força, amor e carinho.
DEDICO

iii
AGRADECIMENTO
A todas as pessoas que fizeram parte desse trabalho e contribuíram para
que o mesmo fosse concluído:
À professora Dra. Maria de Fátima Pires da Silva Machado pela
oportunidade, orientação, confiança e amizade.
À professora, amiga, Dra. Claudete Aparecida Mangolin, pelo seu
exemplo profissional, pelo aprendizado, crescimento e carinho dedicado.
Ao professor e amigo Dr. Clândio Medeiros da Silva pelo apoio,
aprendizado e exemplo de vida e profissional.
À Usina Alto Alegre S/A pelo fornecimento das variedades de cana-de-
açúcar utilizada neste trabalho.
Aos demais professores do laboratório 17 e 21 pelo aprendizado e
convívio, juntamente com os colegas Raphael Augusto, Liriana Cantagalli,
Carlos Alexandre Marochio, Paulo Henrique Marquiori e os demais
companheiros de bancada que me forneceram os primeiros ensinamentos sobre
as técnicas de trabalho em laboratório e manuseio de equipamentos, auxiliando
no desenvolvimento das pesquisas.
Aos técnicos de laboratório, Sérgio e Leila, pelo auxílio durante o
desenvolvimento deste trabalho assim como os secretários Reinaldo e Érika do
Programa de pós-graduação em Agronomia.
Aos meus pais Inervina e José, meu irmão Gustavo e minha namorada
Lígia Adriana pelo carinho e apoio.

iv
BIOGRAFIA
Rone Charles Maranho, filho de José Maranho e Inervina Barizon
Maranho, nasceu em Jussara, Estado do Paraná, em 06 de maio de 1985.
Realizou o ensino fundamental e médio no Colégio Estadual Igléa
Grollmann na cidade de Cianorte – PR.
Em 30 de janeiro de 2010 diplomou-se licenciado em Ciências
Biológicas pela Universidade Estadual de Maringá (UEM) na cidade de Maringá-
PR.
Em março de 2010 iniciou o curso de mestrado em agronomia pelo
programa de pós-graduação em agronomia na área de produção de plantas na
Universidade Estadual de Maringá.

v
ÍNDICE
RESUMO vi
ABSTRACT viii
1. INTRODUÇÃO 1
2. REVISÃO DE LITERATURA 4
2.1 Classificação Botânica da cana-de-açúcar 4
2.2 Divergência genética em cana-de-açúcar 6
2.3 Uso de marcadores moleculares em Saccharum spp. 7
2.4 Marcadores microssatélites (Simple Sequence Repeats) 9
2.5 Marcadores SSR em Saccharum spp. 12
3 MATERIAIS E MÉTODOS 15
3.1 Variedades de cana-de-açúcar utilizadas 15
3.2 Extração do DNA das folhas de cana-de-açúcar 17
3.3 Seleção dos primers de microssatélites 19
3.5 Análise dos Resultados 21
4 RESULTADOS E DISCUSSÃO 23
5 CONCLUSÃO 38
6 REFERENCIAS 39

vi
RESUMO
MARANHO, Rone Charles, M.Sc. Universidade Estadual de Maringá, junho de 2012. Diversidade Genética Entre e Dentro de Variedades de Cana-de-açúcar (Saccharum spp.) Utilizando Marcadores Microssatélites. Professora Orientadora: Dra. Maria de Fátima Pires Machado da Silva. Professora conselheira: Dra. Claudete Aparecida Mangolin.
O objetivo do presente trabalho foi a caracterização molecular de nove variedades de
cana-de-açúcar cultivadas na região norte do Paraná, e utilizadas por uma das unidades
produtoras de etanol e açúcar, analisando locos microssatélites. A análise do
polimorfismo em locos microssatélites dessas variedades deve informar sobre a
variabilidade presente dentro de cada cultivar e a divergência e similaridade genética
entre as mesmas. Foram testados 40 primers dos quais 17 foram utilizados para análise
das 9 variedades de cana-de-açúcar, por apresentarem melhor definição. Dos 17 primers
utilizados apenas os primers UGSM148 e UGSM59 foram monomórficos sendo úteis
para o estudo de uniformidade genética das variedades. Com os primers UGSM35,
SEGMS240, SEGMS1069, SMC226CG, ESTB130, SMC1237FL, SMC165A,
UGSM117, ESTB41, ESTB145, SMC1047HA, SMC2017FL, ESTC66, mSSCIR56, e
ESTB50, foi encontrado um total de 70 alelos variando de 2 a 6 alelos, sendo uma
média de 4,12 alelos por locos. A análise do polimorfismo nos 15 locos SSR de
Saccharum spp mostrou que os locos Smc226CG, Smc1237FL, Smc1047HA, mSscir56,
e EstB60 apresentaram os valores maiores para as estimativas de heterozigosidade
observada, e no loco Smc1047HA foi encontrado o maior número de alelos. As
variedades RB72454, RB92579 e RB867515 apresentaram os valores mais altos de
heterozigosidade média observado. A diversidade genética calculada mostrou maior
identidade genética entre as variedades RB92579 e RB72454 com 86,31% de
similaridade e a menor similaridade encontrada foi entre as variedades RB845210 e
RB835054. Os locos SSR em variedades de cana-de-açúcar são marcadores promissores
para avaliar a real diversidade genética em nível molecular de variedades que
apresentam características morfoagronômicas uniformes, contrariando o postulado de
que existe uniformidade genética em variedades comerciais de cana-de-açúcar.
Palavras-chave: Saccharum spp. Marcador SSR. Diversidade genética.

vii
ABSTRACT
MARANHO, Rone Charles, M.Sc. State University of Maringá, June 2012. Genetic Diversity In and Among Varieties of Sugarcane (Saccharum spp.) Using Microsatellite Markers. Advisor Professor: Dra. Maria de Fátima Pires Machado da Silva. Assistant Professor: Dra. Claudete Aparecida Mangolin.
The purpose of this study was the molecular characterization of nine varieties of
sugarcane grown in the northern region of Paraná, and used by one of the units
producing sugar and ethanol, analyzing microsatellite loci. The analysis of
polymorphism in microsatellite loci of these varieties should provide information on the
variability present within each cultivar and genetic similarity and divergence between
them. Forty primers were tested of which 17 were used for analysis of nine varieties of
sugarcane, because they presented better definition. Among the 17 primers used only
primers were monomorphic UGSM59 UGSM148 and being useful for the study of
genetic uniformity of varieties. With the primers UGSM35, SEGMS240, SEGMS1069,
SMC226CG, ESTB130, SMC1237FL, SMC165A, UGSM117, ESTB41, ESTB145,
SMC1047HA, SMC2017FL, ESTC66, mSSCIR56 and ESTB50, were found a total of
70 alleles ranging from 2 to 6 alleles, a average of 4.12 alleles per locus. The analysis of
polymorphism at 15 SSR loci of Saccharum spp showed that the loci Smc226CG,
Smc1237FL, Smc1047HA, mSscir56 and EstB60 showed higher values for the
estimates of observed heterozygosity, and in situ Smc1047HA found the largest number
of alleles. The varieties RB72454, RB92579 and RB867515 showed the highest values
of heterozygosity was observed. The calculated genetic diversity showed greater genetic
identity among the varieties RB92579 and RB72454 with 86.31% similarity and the
lowest similarity was found between the varieties and RB845210 RB835054. The SSR
loci in varieties of sugarcane are promising markers for assessing the actual genetic
diversity at the molecular level of varieties that have uniform agronomic characteristics,
contradicting the assumption that there is genetic uniformity in commercial varieties of
sugarcane.
Keywords: Saccharum spp. SSR Marker. Genetic Diversity.

1
1. INTRODUÇÃO
A cana-de-açúcar (Saccharum spp.), com provável origem em Nova
Guiné, vem a cada ano gerando riquezas e movimentando a economia nacional e
regional através de seus produtos como o etanol e o açúcar. O setor
sucroenergético é responsável por grande movimentação econômica e social
onde a empregabilidade e as riquezas financeiras são reflexos da produtividade e
demanda pelos derivados da cana-de-açúcar. A safra de 2009/2010 movimentou
56 bilhões de reais na produção de cana, etanol, açúcar e bioeletricidade,
representando cerca de 2% do PIB nacional e gerando 4,5 milhões de empregos
diretos e indiretos. (Cana Web/dados e estatísticas, 2012;
www.jornalcana.com.br).
Para a obtenção de etanol e açúcar foram moídas 624 milhões de
toneladas de cana-de-açúcar na safra de 2010/2011, produzindo 33 milhões de
toneladas de açúcar e 27,6 bilhões de litros de etanol. O setor é composto
atualmente por 437 unidades produtoras sendo 168 produtoras de álcool, 16 de
açúcar e 253 de álcool e açúcar (Ministério do Desenvolvimento, Indústria e
Comércio Exterior, 2012). O total de área plantada equivalente a 7,8 milhões de
hectares, correspondendo a 3,5 % da área agricultável do Brasil (Cana Web/
dados e estatísticas, 2012; www.jornalcana.com.br).
O aumento da demanda de cana-de-açúcar implica num investimento no
valor de R$ 97 bilhões até 2020, para suprir o aumento do consumo interno de
170% (EPE- Empresa de Pesquisa Energética, 2011) junto com o aumento de
exportação do álcool combustível para países como Estados Unidos da América,
Japão, Jamaica e outras nações que compram o etanol do Brasil. No Brasil, a
utilização do bagaço da cana-de-açúcar tem sido proposta como matéria prima
adicional para a produção de etanol, no programa de Bioetanol Brasileiro (Zanin
et al., 2000), e para a obtenção de filme de celulose (Ruzene et al., 2009). As
cinzas de bagaço de cana-de-açúcar também têm sido apontadas como um
material interessante para ser usado na indústria de cimento (Sousa, 2009). O

2
potencial biotecnológico do bagaço da cana-de-açúcar para ser utilizado em
processos de bioconversão usando microorganismos foi destacado como uma
fonte promissora em relação a outras fontes de bagaço, nas investigações de
Pandey et al. (2000).
A crescente utilização dos derivados da cana-de-açúcar e a consequente
necessidade do aumento de produção criam elevada expectativa sobre os
programas de melhoramento genético. Os estudos realizados por esses programas
são de vital relevância para o aumento da produtividade do setor sucroenergético
permitindo suprir a demanda de álcool e açúcar do mercado interno e externo.
Segundo Barbosa et al. (2000) o melhoramento genético da cana-de-açúcar foi
fundamental para o desenvolvimento do setor canavieiro nacional.
Outra característica importante a ser considerada na produção de cana-
de-açúcar, é o estreitamento genético devido à origem comum das variedades
comerciais atualmente plantadas, podendo causar assim um fenômeno genético
conhecido como vulnerabilidade genética (Barbosa, 2001) em que tornam
vulneráveis a diversos fatores como doenças e pragas, os canaviais atualmente
estabelecidos no Brasil. Para obter melhores e mais rápidos resultados em
programas de melhoramento genético vegetal, ferramentas biotecnológicas vem
sendo amplamente utilizadas na caracterização molecular de grandes culturas,
gerando informações que podem ajudar no trabalho de melhoramento genético,
auxiliando na escolha dos progenitores geneticamente mais divergentes para o
desenvolvimento de clones e posterior variedade de uso comercial.
A seleção dos progenitores é uma etapa fundamental nos programas de
melhoramento genético, onde a análise de divergência genética fornece
parâmetros para a identificação de progenitores que após o cruzamento,
possibilitam maior efeito heterótico na progênie e maior possibilidade de
recuperação de genótipos superiores nas gerações segregantes (Cruz et al., 2004).
Desta forma, conhecer a variabilidade genética das variedades cultivadas é
importante para orientar o melhoramento.
Dentre os marcadores moleculares utilizados para estimar a diversidade
genética em cana-de-açúcar estão as sequências simples repetidas de DNA,

3
denominadas de locos SSR ou locos microssatélites, descritas em plantas na
década de 1990 por Lagercrantz et al.(1993) e Wang et al. (1994). O número e a
frequência de alelos diferentes nos locos microssatélites podem ser usados para
expressar a diversidade genética em variedades de plantas e a divergência
genética entre variedades. A análise de locos microssatélites também é útil para
monitorar a estabilidade genética de genótipos selecionados (clones) para
características de interesse morfoagronômico.
A análise do polimorfismo em locos microssatélites destas variedades
deve informar sobre a variabilidade presente dentro de cada cultivar e a
divergência e similaridade genética entre as mesmas. O padrão de diversidade
genética em locos microssatélites de cada variedade pode ser acrescentado à
relação de características morfoagronômicas de cada uma delas, para traçar
estratégias de utilização de cada genótipo em programas de melhoramento, no
cultivo destas em áreas geográficas distintas, no controle de doenças, e como
matéria prima para as indústrias de produção de seus derivados.
Por isso, o objetivo no presente trabalho foi a caracterização molecular
de nove variedades de cana-de-açúcar cultivadas na região norte do Paraná, e
utilizadas por uma das unidades produtoras de etanol e açúcar, analisando locos
microssatélites.

4
2. REVISÃO DE LITERATURA
2.1 Classificação Botânica da cana-de-açúcar
Em 1758, Linneu descreveu a cana-de-açúcar como Saccharum
officinarum e Saccharum spicatum. O gênero Saccharum a que pertence à cana-
de-açúcar (Saccharum spp.) é apresentado por Matsuoka et al. (1999) como
integrado a família Poaceae, tribo Andropogeneae, sendo uma espécie alógama
de ciclo perene naturalmente e semiperenes quando cultivada. A classificação
taxonômica mais completa e aceita atualmente inclui a cana-de-açúcar na família
Poaceae, subfamília Panicoideae, tribo Andropogoneae, subtribo Saccharinae,
grupo Saccharastrae e gênero Saccharum (Daniels e Roach,1987).
A origem da cana-de-açúcar ainda é discutida nos meios científicos
sendo que sua provável origem é em Nova Guiné, porém a grande variedade de
hibridização encontrada atualmente dificulta a precisão de sua origem. O grupo
“Saccharum” é denominado por alguns estudiosos da cana-de-açúcar como um
complexo composto pelos gêneros Saccharum, Erianthus, Sclerostachya,
Narenga e Miscanthus. O intercruzamento entre os integrantes deste complexo
foi responsável pela domesticação da cana-de-açúcar no continente Asiático no
início de sua domesticação (Matsuoka et al., 2005).
As variedades atuais de cana-de-açúcar surgiram após vários
cruzamentos interespecíficos, e retrocruzamentos com cana nobre (Saccharum
officinarum) obtendo-se números variáveis de cromossomos (Figura 1) que
também ocorre devido à aneuploidia, comum em plantas poliplóides (Barreto e
Simon, 1982). Atualmente encontramos variedades comerciais de cana com
número cromossômico que varia de 100 a 130, sendo que 80% dos cromossomos
derivados de S. officinarum, 10% de S. spontaneum e 10% dos cromossomos
recombinantes entre os dois genomas (D´Hont et al., 1996; Ferreira, 2010).

5
Figura 1 – A,B,C: Esquemas mostrando o processo de obtenção de variedades atuais de cana-de-açúcar através do processo de Nobilização. (Ming et al., 2006; Ferrari, 2010).

6
2.2 Divergência genética em cana-de-açúcar
A diversidade genética disponível atualmente nos clones e variedades de
cana-de-açúcar provém, segundo Ferreira (2010), de cruzamentos realizados no
início do século XX pelos programas de melhoramento genético de cana-de-
açúcar da Índia (Coimbatore) e de Java, onde foram desenvolvidos híbridos
interespecíficos entre S. officinarum e S. spontaneum. Após o processo de
nobilização (cruzamento de S. officinarum com outras espécies de cana-de-
açúcar, seguido de retrocruzamento com S. officinarum ) foram obtidos genótipos
com números variados de cromossomos e características agronômicas também
variadas.
No Brasil as primeiras variedades foram desenvolvidas pelo Instituto
Agronômico de Campinas (IAC) e foram nomeadas de IAC36-25 e IAC47-31
(Figueiredo, 2008; Ferreira, 2010). Vários outros programas de melhoramento
foram criados no Brasil, sendo os mais importantes representados por:
PLANALSUCAR [atualmente denominado RIDESA e detentora das variedades
RB (República do Brasil)]; COPERSUCAR [detentora das variedades SP (São
Paulo) e que atualmente é chamado de Centro de Tecnologia Canavieira
(desenvolvendo variedades com prefixo CTC]; e o mais recente programa de
melhoramento genético de cana-de-açúcar do Brasil é representado pela
CanaVialis S.A (Ferreira, 2010).
Atualmente no Brasil a cultura da cana-de-açúcar é caracterizada pelo
monocultivo de híbridos com uma base genética estreita apresentando
populações com alta similaridade genética como observado por Duarte-Filho et
al. (2010) em seu trabalho com variedades comerciais RB. A vulnerabilidade
genética (Barbosa, 2001; Almeida, 2005) da cultura de cana-de-açúcar no Brasil,
pode ocasionar a redução de produtividade devido ao aumento de ocorrência de
doenças e outras anomalias causadas pela uniformidade genética das plantações.
Durante muitos anos os programas de melhoramento genético de cana-de-açúcar
utilizaram marcadores morfológicos para a obtenção de progenitores com maior
variabilidade genética, visto que, o conhecimento da diversidade genética entre

7
variedades comerciais em programas de melhoramento genético é de vital
importância para os melhoristas na identificação e organização dos recursos
genéticos disponíveis, visando à utilização desses na produção de novas
variedades promissoras (Palomino et al., 2005; Silva et al., 2011). Os marcadores
morfológicos apresentam algumas limitações como instabilidade decorrente da
relação com o ambiente, e a dependência de um estado adulto da planta para
análise dos caracteres agronômicos. Outro fator prejudicial à utilização de
marcadores morfológicos no estudo da divergência genética é o estreitamento da
base genética em grandes culturas como a da cana-de-açúcar, conferindo às
cultivares, características agronômicas em comum, não podendo ser distinguidas
(Schuster et al., 2006; Silva, 2010).
2.3 Uso de marcadores moleculares em Saccharum spp.
Atualmente as pesquisas para o desenvolvimento de novas variedades
não se restringem ao melhoramento convencional, mas sim, do uso de técnicas
moleculares que facilitam e geram informações confiáveis sobre a estrutura
genética da cana-de-açúcar. O polimorfismo do DNA pode ser explorado através
das técnicas de análise molecular, onde, um grande número de marcadores
moleculares pode ser encontrado. Um marcador molecular pode ser definido
como locos cromossômicos, genes ou sequências de DNA que apresentam
segregação mendeliana (Borém et al., 2009) e que sejam úteis no estudo de
polimorfismos na comparação entre indivíduos.
As ferramentas biotecnológicas estão sendo expressamente utilizadas
para caracterização genética, criação de mapas de ligação e uma infinidade de
outras aplicações que favorecem o conhecimento sobre um determinado genótipo
e sua relação intra e interespecífica. Entre as técnicas mais utilizadas em cana-de-
açúcar encontramos trabalhos com o RFLP (Restriction Fragment Lenght
Polymorphic; Botstein et al., 1980) o RAPD (Random Amplified Polymorphic
DNA; Williams et al., 1990) além de outros marcadores como as SSR (Simple
Sequence Repeats; Hamada et al.,1982; Litt e Luty, 1989; Weber e May, 1989;

8
Jacob et al., 1991) também chamados de marcadores microssatélites, e o AFLP
(Amplified Fragment Lenght Polymorphism; Vos et al., 1995).
A utilização de marcadores moleculares para estudos genéticos em cana-
de-açúcar vem se expandindo com a adaptação dos protocolos e desenvolvimento
de novas técnicas mais economicamente viáveis e informativas. Silva (2006a)
utilizou marcadores RAPD em 8 genótipos de cana-de-açúcar proveniente de 2
programas de melhoramento genético, para investigar a indução de variabilidade
genética na micropropagação das variedades. No estudo o autor testou 61
primers, e usou 13 para a avaliação dos genótipos. A análise dos fragmentos
aleatórios demonstrou a diversidade gerada pelo cultivo in vitro e agrupou as
variedades cultivadas tradicionalmente e micropropagadas em grupos distintos.
Os resultados sugeriram a eficiência do uso de marcadores RAPD no estudo de
diversidade em cana-de-açúcar para as variedades estudadas.
Silva (2006b) também fez uso de marcadores tipo RAPD e caracteres
morfoagronômicos para analisar a divergência genética em cana-de-açúcar,
obtendo correlação significativa de média magnitude (r=0,47) entre a divergência
genética e os caracteres morfoagronômicos.
Selvi et al. (2005) utilizaram marcadores AFLP para caracterização
molecular de 28 variedades de cana-de-açúcar cultivadas na Índia utilizando
eficientemente apenas 11 primers para separar individualmente cada cultivar.
Segundo os autores, a técnica utilizada foi eficiente na caracterização de todas as
variedades e permitiu observar a maior diversidade genética entre as variedades
tropicais e subtropicais quando comparadas entre as variedades de cada uma das
regiões.
Estudos utilizando marcadores AFLP em cana-de-açúcar também foram
realizados por Lima et al. (2002), onde os objetivos foram avaliar o nível de
similaridade genética entre 79 cultivares (híbridos interespecíficos) e a correlação
entre similaridade genética e coeficiente de parentesco utilizando fragmentos de
segmentos polimórficos (AFLP) e dados de genealogia. Foram testados 64
primers e destes, 40 combinações foram selecionados por apresentarem alto
número de bandas polimórficas sendo que para a produção do perfil AFLP foram

9
selecionados apenas as 21 melhores combinações de primers. Os autores
relataram que através das análises dos clusters foi possível sugerir uma
importante relação genética entre as cultivares. Os dados analisados permitiram
observar a existência de correlação (r=0,42, P<0,001) entre similaridade genética
e o coeficiente de parentesco, sugerindo que os dados de AFLP podem ajudar na
quantificação do grau de relação com maior precisão entre cultivares de cana-de-
açúcar.
Palhares (2010) utilizou em seus estudos, marcadores AFLP e
marcadores baseados em retrotransposons para construção de mapa de ligação
em cana-de-açúcar. Foram analisados os marcadores de 188 indivíduos e foram
obtidos 600 marcadores de dose única a partir de 22 combinações de enzimas de
restrição e primers de AFLP. Segundo a autora do trabalho, a grande quantidade
de locos polimórficos possibilitou a geração dos mapas de ligação presente nos
resultados.
2.4 Marcadores microssatélites (Simple Sequence Repeats)
Os microssatélites, também denominados de SSR (Simple Sequence
Repeats) ou STR (Short Tandem Repeats), segundo Borém et al. (2009), são
sequências simples, repetidas e presentes densamente nos genomas eucariotos e
que consistem em um a seis nucleotídeos repetidos em tandem. A conservação
das sequências de DNA que flanqueiam os microssatélites, permitem a seleção
de primers específicos que amplificam, via PCR, fragmentos contendo o DNA
repetitivo em todos os genótipos (Borém e Caixeta, 2009).
A reprodutibilidade e simplicidade técnica, a pequena quantidade de
DNA requerida, o custo baixo em relação ao RFLP e AFLP, o grande poder de
resolução e os altos níveis de polimorfismo são as principais características que
justificam o desenvolvimento de marcadores microssatélites em plantas
cultivadas. Os microssatélites parecem ter distribuição frequente e aleatória,
permitindo uma cobertura completa do genoma (Rallo et al., 2000). Além disso,
para Ferreira e Grattapaglia (1998), os microssatélites constituem uma das

10
classes de marcadores moleculares mais polimórficos. Os microssatélites também
apresentam vantagens sobre os demais marcadores baseado em PCR, como o
RAPD, porque são codominantes e facilmente reproduzíveis (Borém e Caixeta,
2009).
Em cana-de-açúcar, marcadores microssatélites podem ser desenvolvidos
a partir de sequências genômicas ou de sequências depositadas nos banco de
dados de EST (Expressed Sequence Tag; Cordeiro et. al., 2003). Muitas das
caracterizações de microssatélites foram realizadas através de pesquisas em
banco de dados, de sequências publicadas, e da construção de bibliotecas
genômicas (Cordeiro et al., 2000).
O nível alto de diversidade alélica dos marcadores SSR, possibilita a
obtenção de polimorfismo em populações multiparentais e em populações
derivadas de híbridos de genótipos relacionados, além de distinguir acessos de
germosplasma intimamente relacionados (Borém e Caixeta, 2009). O alto nível
de polimorfismo dos marcadores microssatélites os torna efetivos no estudo da
diversidade genética das mais diversas espécies e organismos vegetais. Borba et
al. (2009) utilizou 12 marcadores microssatélites em 417 variedades locais de
arroz (Oriza sativa) coletados em diferentes anos de produção. Os pesquisadores
encontram um aumento na variabilidade genética e relacionaram essa variação a
provável introdução de variedades modernas conforme necessidades de mercado.
Salem et al. (2008) estudou a diversidade genética em variedades de
trigo (Triticum aestivum L.) utilizando marcadores SSR e marcadores
morfológicos, associando ambos os resultados para verificar a eficiências desses
parâmetros. Os autores obtiveram de 2 a 7 alelos por locos e 1 valor de PIC
(conteúdo de informação de polimorfismo) igual a 0,278 para o primer Xgwm95
a 0.816 para o primer Xgwm437, obtendo uma média de 0,548. Para os
pesquisadores ambos os parâmetros analisados geraram informações coerentes
entre as técnicas e julgaram pertinente a utilização de marcadores microssatélites
para distinção dos genótipos e análise da diversidade genética.

11
Silva (2010) utilizou sequenciador automático para estudar locos
microssatélite em trigo, e fazer a caracterização molecular de variedades de trigo
nacional. O autor do trabalho obteve 21 marcadores polimórficos, revelando de 2
a 8 alelos por loco com média de 4,29 alelos por loco. O PIC encontrado teve a
média de 0,593. Para o pesquisador os locos SSR se mostraram bastante
eficientes na caracterização de trigo estando assim disponíveis dados para fins de
registro e proteção de cultivares segundo leis vigentes.
No estudo com soja (Glycine max (L.) Merrill ) Vieira et al. (2009)
analisaram a variabilidade genética de 53 cultivares utilizando 283 marcadores
SSR sendo que destes 53 foram selecionados por serem evidentemente mais
polimórficos. No trabalho os autores obtiveram 124 alelos, com média de 2,34
alelos por loco e PIC (Conteúdo de Informação de Polimorfismo) médio de 0,47.
Foi obtida também a distância genética média de 0,47. Os valores obtidos
indicaram o bom caráter informativo do marcador microssatélite para o estudo e
identificação das variedades comerciais, mesmo as elites, de soja presente no
mercado.
No estudo da diversidade em milho, frequentemente são utilizados
marcadores microssatélites assim como encontrado no trabalho de Meirelles
(2009). A pesquisadora analisou 18 locos SSR em 40 linhagens de milho
oriundas de 2 grupos compostos: Dentado e Flintisa. Foi encontrado
polimorfismo em todos os locos sendo observada diversidade entre os dois
grupos com um total de 117 alelos. Os valores de heterozigosidade encontrados
foram abaixo dos valores esperados para a população em equilíbrio de Hardy-
Weinberg, sendo que o valor médio da heterozigosidade esperada foi de 0,7 e o
valor médio da heterozigosidade observada foi de 0,06. O desvio no equilíbrio de
Hardy-Weinberg, segundo a autora, foi devido alto índice de endogamia o que é
corroborado pelos índices de fixação que variou de 0,83 a 1,00.

12
2.5 Marcadores SSR em Saccharum spp.
A geração de informações genéticas da cana-de-açúcar através de
marcadores microssatélites tem aumentado consideravelmente com o
aprimoramento das técnicas de análises. Os marcadores SSR há alguns anos já
são utilizados para criação de mapas genéticos (McIntyre et al., 2001) e
Fingerprint de clones de cana-de-açúcar (Pan et al., 2002) porém sua utilização
para estudos de diversidade genética tem aumentado consideravelmente com a
caracterização de novos marcadores microssatélites e desenvolvimento de novos
primers para Saccharum spp..
Cordeiro et al. (2000) desenvolveram uma biblioteca genômica com a
cultivar Q124 e desenhara um total de 124 primers SSR onde 71 foram derivados
de repetições dinucleotideos e 29 de repetições trinucleotideos, sendo que dos
124 primers 91% apresentaram-se polimórficos. Pan (2006) avaliou um conjunto
de 221 primers microssatélites desenvolvidos pelo ISMC (Consórcio
Internacional de Microssatélites de cana-de-açúcar). Os primers foram testados
em cinco clones elite americanos incluindo as variedades LCP85-384
(Louisiana), HoCP 96-540 (Louisiana), CP72-1210 (Florida) e CP85-1308
(Florida). No total foram identificados 467 alelos onde 350 eram polimórficos
com média de cinco alelos polimórficos por marcador com tamanho variando de
80 a 460 pb. Parida et al. (2009) desenvolveram duas bibliotecas genômicas com
as variedades indianas Co 7201 e Co 86011, e foram desenhados um total de 567
marcadores moleculares Classe I. O nível de polimorfismo detectado entre
espécies, gênero e variedades de cana-de-açúcar, para os marcadores
desenvolvidos foi de 88,6%. Parida et al. (2009) também analisaram a
transferência cruzada e obtiveram taxa de transferência igual a 93,2% entre o
complexo Saccharum e 25% para cereais.
Oliveira et al. (2009) utilizando o banco de dados de EST (Expressed
Sequence Tags) da SUCEST (Sugar Cane EST Project) para desenvolver 342
marcadores microssatélites (EST-SSRs) em 18 variedades de cana-de-açúcar
brasileiras, encontraram uma média de 7,55 alelos por locos, sendo que 65,5%

13
dos fragmentos encontrados eram polimórficos. Para as análises os referidos
autores desenvolveram uma matriz binária e utilizaram o coeficiente de Jaccard
para determinar a similaridade genética (GS), utilizando também a matriz de
similaridade para desenho do dendrograma seguindo o método UPGMA
(Unweighted Pair Group Method using Arithimetic Average).
Singh et al (2008) no trabalho analisaram o potencial polimórfico de
marcadores microssatélites em duas variedades comerciais indianas de cana-de-
açúcar (Co86011; CoH70) utilizando primers desenhados pela National Reserch
Centre on Plant Biotechnology, Indian Agricultural Institute. Foram testados um
total de 702 primers SSR sendo destes: 176 marcadores microssatélite Class I
derivados de unigenes de cana-de-açúcar (UGSM), 272 marcadores
microssatélites classe I de genoma enriquecido de cana-de-açúcar (SEGM), 50
marcadores microssatélites genômico classe I de cana-de-açúcar (SGM), 32
marcadores microssatélites classe I derivados de cDNA de cana-de-açúcar
(SCM) e 172 marcadores microssatélites classe 2 derivados de unigenes de cana-
de-açúcar (UGSM). Do total de primers utilizados 494 (74,6%) detectaram locos
monomorficos e 168 (26%) detectaram locos polimórficos. Os primers que
detectaram polimorfismo foram: oitenta e sete (51,7%) UGSM, quarenta e cinco
(26,7%) SEGM, dezessete (10,1%) SGM, dezenove (11,3%) SCM.
Com o objetivo de determinar a diversidade genética entre o complexo
Saccharum, Cordeiro et al. (2003) utilizaram seis marcadores microssatélites em
sessenta e seis acessos do gênero Saccharum e Erianthus. Os autores
encontraram um total de 187 alelos para os seis marcadores utilizados, sendo que
um mínimo de dois marcadores foi suficiente para distinguir todo material.
Foram encontradas as médias de 3,8 alelos para espécies de Sorghun, 14,3 alelos
para espécies Erianthus e 17,8 alelos para as espécies de Saccharum incluindo as
variedades de cana-de-açúcar. Para as espécies que originam as variedades
comerciais brasileiras os autores encontraram uma maior variabilidade genética
em Saccharum spontaneum (GS= 0.30) que em Saccharum officinarum (GS=
0.50), segundo os autores a maior variabilidade de S. spontaneum é
provavelmente devido a sua maior distribuição geográfica, maior variabilidade

14
cromossômica e maior variedade de fenótipos. Já a espécie de S. officinarum
apresentou menor variabilidade; segundo os autores, devido à pressão seletiva
para a alta quantidade de sacarose requerida no processo de domesticação da
cana-de-açúcar.
Duarte-Filho et al. (2010) analisaram a similaridade genética entre
cultivares comerciais (RB) de cana-de-açúcar brasileiras utilizando marcadores
SSR e coeficiente de parentesco (COP). Foram utilizadas 30 variedades de cana-
de-açúcar dos cultivares República do Brasil (RB) onde do total de 19
marcadores gerados, 10 apresentaram-se polimórficos. Foi encontrado um total
de 57 alelos com média de 3,2 alelos por locos. Utilizando o coeficiente de
similaridade de Jaccard foi encontrado GS variável de 0,95 entre os genótipos
RB93509 e RB931011, e 0,56 entre os genótipos RB928064 e RB813804, com
uma similaridade média de 0,78. Para o coeficiente de parentesco encontrado
entre 28 acessos de cana-de-açúcar foi encontrado um COP de 0 a 0,76 com
média de 0,14, demonstrando que os genótipos estudados apresentaram alto
COP.

15
3 MATERIAIS E MÉTODOS
3.1 Variedades de cana-de-açúcar utilizadas
Foram utilizadas oito variedades comerciais de cana-de-açúcar dentre as
mais plantadas e com melhores características agronômico-econômicas,
amplamente cultivadas na região norte do Paraná, e um clone (RB036152) em
fase de teste para ser lançado como variedade. As variedades foram
desenvolvidas pelos programas de melhoramento RIDESA (Rede
Interuniversitária para o desenvolvimento do setor sucroenergético; variedades
RB) e COPERSUCAR. Foram analisadas oito plantas de cada variedade
SP813250, RB845210, RB935744, RB867515, RB036152, RB835054,
RB855453, RB92579, e 10 plantas da variedade RB72454. As amostras estavam
disponíveis nos canaviais localizados na Fazenda Junqueira no distrito de Alto
Alegre em Colorado no Paraná, e são usadas pela indústria para a fabricação de
álcool e açúcar.
Folhas jovens do cartucho foram coletadas diretamente dos canaviais,
armazenadas em sacos de sombrite e devidamente etiquetadas para serem então
acondicionadas numa caixa térmica contendo gelo e separadas em camadas com
folhas de jornal para evitar que as amostras fossem molhadas. A identificação foi
feita com a nomenclatura das respectivas variedades e numeradas conforme
coletadas. No laboratório as amostras foram devidamente congeladas em
nitrogênio líquido e guardadas em ultrafreezer (-80 °C). Na Tabela 1 estão
relacionadas as variedades com suas principais características agronômicas e
genealogia.

16
Tabela 1 - Principais características agronômicas e genealogia das variedades de cana-de-açúcar estudadas
VARIEDADES
PRINCIPAIS CARACTERÍSTICAS AGRONÔMICAS
GENEALOGIA
RB72454 - Alta produtividade
- Ampla adaptabilidade CP53-76 X ?
RB845210 - Alta produtividade
- Ótimo teor de sacarose
RB72454 X SP70-1143
RB935744 - Alta produtividade
- Excelente sanidade RB835089 X RB765418
RB867515 -Alta produtividade
- Alto teor de sacarose RB72454 X ?
RB036152 - Precocidade SP835073 X RB867515
RB835054 - Alta produtividade
- Ampla adaptabilidade RB72454 X NA56-79
RB 855453 - Alta produtividade
- Alto teor de sacarose TUC71-7 X ?
RB92579 - Resistente a pragas
- Alta produtividade RB75126 X RB72199
SP813250 - Alta produtividade
-Alto teor de sacarose CP70–1547 X SP71–127
Fonte: RIDESA; COPERSUCAR.

17
3.2 Extração do DNA das folhas de cana-de-açúcar
Para a extração do DNA das folhas de cana-de-açúcar, foi utilizado o
protocolo de extração descrito por Hoisington et al. (1994), amplamente utilizado
em pesquisas com cana-de-açúcar e outras culturas. O material vegetal
corretamente identificado foi macerado em nitrogênio líquido com auxílio de
almofariz e pistilo até a obtenção de um pó fino e com poucas fibras visíveis.
Foram colocados 300 mg de folha macerada em tubo eppendorf (2 mL) e 800 µL
de solução de extração CTAB (NaCl 5M; Tris-HCl 1M pH 7,5; EDTA 0,5 M pH
8,0; CTAB 1% (Cationic Hexadecyl trimethyl ammonium bromide) e 140 mM de
β-mercaptoetanol). Todos os tubos foram homogeneizados com agitação manual
até completo contado da superfície das partículas vegetais com a solução de
extração. Os tubos foram mantidos em banho-maria a 65°C por 1 hora e com
suave agitação manual a cada 5 minutos. Depois de incubados, os tubos foram
mantidos à temperatura ambiente por 5 minutos. Na sequência foram acrescidos
800 µL de clorofórmio-álcool isoamílico (24:1) e agitados lentamente por 5
minutos. Os tubos foram centrifugados em rotação de 15.000 RPM e à
temperatura ambiente por 15 minutos sendo posteriormente recuperada a fase
líquida e transferida para eppendorf limpo. Foram adicionados 800 µL de
clorofórmio-álcool isoamílico (24:1) à fase recuperada, posteriormente, a mistura
foi centrifugada a 15.000 RPM por 10 minutos.
O sobrenadante foi transferido para tubo eppendorf limpo. Para a
precipitação do DNA foram adicionados 300 µL de isopropanol gelado com
agitação manual por inversão durante 1 minuto. Após a visualização do
precipitado os tubos foram armazenados em freezer (-20 ºC) por 12 horas para
melhor precipitação do DNA. Após esse período de precipitação os tubos foram
centrifugados com 10.000 RPM por 10 minutos. O sobrenadante de cada tubo foi
descartado e foram acrescentados 800 µL de álcool etílico 70% com posterior
agitação por inversão durante 1 minuto. Os tubos foram então centrifugados com
10.000 RPM em temperatura de 4°C por 10 minutos. O sobrenadante foi

18
novamente descartado e o pellet de DNA obtido foi seco por 1 hora na
temperatura ambiente.
Após a secagem do pellet foram adicionados 400 µL de TE (1 M
TRIS/HCl e 0,5 M EDTA). Decorrido 1 hora de suspensão do DNA em TE
foram adicionados 2µL de RNAse 20 ng/µL, deixando em temperatura ambiente
por 2 horas. Em seguida foram adicionados 200 µL de fenol equilibrado (pH 8,0)
e 200 µL de clorofórmio-álcool isoamílico (24:1). Os tubos foram então agitados
por 5 minutos e centrifugados com 10.000 RPM em temperatura ambiente por 10
minutos. A fase aquosa foi transferida para eppendorf limpo e acrescido 400 µL
de cloroformio-álcool isoamilico (24:1). Em seguida os tubos foram agitados por
5 minutos e então centrifugados com 10.000 RPM por 10 minutos à temperatura
ambiente. O líquido da fase superior foi retirado e transferido para tubo
eppendorf limpo onde foram adicionados 250 µL de isopropanol gelado e 25 µL
de NaCl 5M com posterior agitação suave por 1 minuto. Após visualização do
precipitado os tubos foram condicionados em freezer (-20 °C) por 12 horas para
maior precipitação.
Decorrido esse período os tubos foram novamente centrifugados a
10.000 RPM por 10 minutos em temperatura de 4ºC. O sobrenadante foi
descartado e o precipitado foi lavado com 800 µL de etanol 70% gelado sendo
misturado por inversão durante 1 minuto. Foi realizada uma centrifugação com
10.000 RPM durante 10 minutos à temperatura de 4°C. O sobrenadante foi
descartado e o pellet de DNA foi seco por 1 hora sendo então ressuspendido em
50 µL de solução de TE (1 M TRIS/HCl e 0,5 M EDTA). Os tubos contendo a
suspensão de DNA foram armazenados em geladeira a 4°C. As amostras de DNA
obtidas foram quantificadas utilizando amostras de DNA padrão (Fago λ) em gel
de agarose 0,8% com tampão TAE 1x (TRIS, EDTA, Acetato de Sódio, pH 8,0)
(Hoisington et al.,1994). Com a quantificação em gel de agarose a 0,8% foi
possível observar a integridade do DNA extraído e com o auxílio do fago λ,
verificar a concentração de DNA disponível para a reação de amplificação via
PCR. Para o processo de quantificação foram utilizados fago λ de concentração
do DNA extraído.

19
3.3 Seleção dos primers de microssatélites
Os primers foram selecionados a partir de 40 primers descritos na
literatura para utilização em reações de microssatélite em Saccharum spp. e que
apresentavam maior polimorfismo e maior número de alelos segundo os autores
dos trabalhos consultados. Ao final dos testes foram selecionados 17 primers
para analisar o DNA das nove variedades de cana-de-açúcar (Tabela 2). Os
primers selecionados foram descritos nos trabalhos de Cordeiro et al. (2000),
Singh et al. (2008) e Oliveira et al. (2010), sendo a maioria desenvolvidos pela
International Sugarcane Microssatelite Consortiun (ISMC) e todos os primers
foram sintetizados pela Invitrogen, USA.
3.4 Reação de amplificação do DNA
O DNA amostrado das variedades de cana-de-açúcar foi amplificado
com utilização de um termociclador marca Techne TC-512. Para as reações de
PCR foi utilizado um volume final de 20 µL, sendo composto por 13,1 µL de
água ultrapura (Millipore Corporation), 1,5 µL de DNA genômico (10 ng ∙ µL-1),
0,4 µL de primer forward e reverse (Invitrogen, Life Technologies Corporation),
0,8 µL de dNTPs, 1,6 µL de MgCl2 (2 mM), 0,2 µL (1 U) de Platinum® Taq
DNA polimerase (Invitrogen), 2,0 µL de buffer 1x (Invitrogen). Além da
concentração de DNA supracitada, foram testadas as concentrações de 25 e 10 ng
para padronização das reações.
Duas configurações do termociclador foram utilizadas para o presente
trabalho, sendo uma PCR-TD (Touchdown) e outra com a Temperatura de
anelamento (T °A) específica para cada primer. A PCR-TD foi composta por

20
Tabela 2. Relação dos primers e temperaturas utilizadas para a amplificação do DNA das nove variedades de Saccharum spp. analisadas.
Primer
Repetição
SSR
Tamanho
(pares de Base)
°C
Sequência Forward do Primer Sequência Reverse do Primer
SMC1237FL (CA)6 100-140 58 TTCACGAACACCCCACCTA GCGCCAGGTAACCTACTGAA
SMC1047HA (GA)26 110-170 62 TGAGCCTAAGCCAGAAAGAAG GGAACTAATTTCCTACGAGAA CAC
SMC2017FL (CTT)13 210-260 62
CACAAGTGAAGATAATAG TGT CCCT GAT CCC AAA TCC CTT GAT CTC
SMC165A (SGA)11 90-130 58 GCAACCGCGTCCTCATAC GAGCTTCTGATCGGCCTTC
SMC226CG (CA)10 136 TD GAGGCTCAGAAGCTGGCAT ACCCTCTATTTCCGAGTTGGT
UGSM148 (GAA)5GGGCA..(GAG)5 42-647 TD GATGACTCCTTGTGGTGG CTTGACGACCCTGCTGCT
UGSM59 (GGC))7 71-500 55 GTGAACGACTCCATCGCC TTGAAACCACCGTAAATAAGA
UGSM38 (ACG)7 50-1185 55 CCGAGTGATGATGTGATGT GGGACAACTAATGTAACTGATT
UGSM117 (GCT)5GCCAT..(TG) 93-867 TD GAGGAGGTGACGGTGGAG GACGCTACGGTTCATTCA
SEGMS240 (GTT)12 100-900 53 CAACAAGAAAGAACACAATGAG AATGTGAATAACTAGGTGGTCC
SEGMS1069 (AGA)10 100-600 53 CGGGAATTCGATTTCATGGGTTC GACTACTCTAAGCATCGTCCTC
ESTB130 (CAA)13 124 50 GCCCAGGTAATTATCCAGACTC GCTGTTGCTCACTGGTTCC
ESTB41 (CGA)8 163 63 CATGGAGAGCTGGGCGACCTG GGCGGCGGCGAGGATGA
ESTB145 (CCT)5 234 68 GGGAAGCAAGCGAGAGCAGCAGAG GAGCGCGAGGCCGTTGTTGAG
ESTB60 (TTG)10 198 53 AGCCGCAATGAATCCAACTG CTCTAGCTCCGACGATGATACCTC
ESTC66 (CCGC)3 152 61 AGTACAGGCTGCTCTCAATCAA TCTGTCATCTGTGTTCGTTCTG
mSSCIR56 (GT)24 125-170 58 ATTTGACGCTACGATGGTG ATCCGTTTTTCAGCAGAGC
Fonte: Cordeiro et al. (2000), Singh et al. (2008) e Oliveira et al. (2010).

21
desnaturação inicial de 1 minuto a 94°C seguido de 10 ciclos, com 1 minuto a
94°C, 1 minuto com temperatura inicial de 65 °C ocorrendo a redução de 1 °C a
cada ciclo, 2 minutos a 72 °C e mais 20 ciclos de 1 minuto a 94 °C, 1 minuto a
55 °C e 2 minutos a 72 °C, sendo finalizado com extensão final de 5 minutos a
72 °C. A reação de PCR em que se utilizaram as temperaturas específicas de
anelamento do primer (Tabela 2) foi constituída de temperatura de 94 °C por 5
minutos para a desnaturação da dupla fita de DNA, uma sequência de 30 ciclos
foi realizada após a desnaturação do DNA onde cada ciclo era composto por uma
etapa inicial a 94 °C por 1 minuto; a segunda etapa com a respectiva temperatura
de anelamento de cada primer por 1 minuto, e a última etapa do ciclo a 72 °C por
1 minuto. Finalizando a reação, foi realizada uma extensão final por mais 15
minutos a 72 °C.
As amostras após a reação de PCR foram armazenadas em freezer com -
20 °C até sua utilização. O DNA amplificado obtido pós-reação foi preparado
para aplicação em gel de agarose e posterior análise dos produtos. A separação
dos segmentos amplificados foi feita em gel de agarose 4% (50% agarose
ultrapure e 50% agarose Metaphor™) utilizando tampão TBE 0,5X (89 mmol/L
TRIS, 89 mmol/L, Ácido Bórico e 2 mmol/L EDTA) com 60 Volts por 4 horas.
Foi utilizado ladder (Invitrogen) de 100pb como marcador de peso molecular.
Após a eletroforese o gel foi corado em banho de brometo de etídio a 0,5 µg/mL
e fotografado utilizando transiluminador com luz UV da Loccus Biotecnologia e
software L-Pix image 1.21.
3.5 Análise dos Resultados
As bandas geradas pelas reações de PCR foram analisadas como caráter
único e matrizes binárias foram geradas a partir do perfil de alelos apresentados
em cada gel, sendo atribuído o valor 1 para a presença de banda e 0 para a
ausência.

22
Marcadores SSR são considerados codominantes, porém a cana-de-
açúcar por possuir genoma altamente poliplóide, pode apresentar vários
cromossomos com mais de dois alelos diferentes, simultaneamente para um
único loco. Isto torna complexa a estimativa das frequências dos alelos, e
consequentemente, os cálculos dos valores de heterozigosidade média esperada, e
do déficit de heterozigotos na amostra de cada variedade. Somente os valores de
heterozigosidade média observada podem ser obtidos por contagem direta de
plantas heterozigotas para dois ou mais alelos.
Após a construção da matriz binária, foram utilizados os softwares
estatísticos Popgene 1.32 (Yeh et al., 1999) e NTSYSpc versão 2.1 (Rohlf,
2000). Para o cálculo da distância genética foi utilizado o índice de similaridade
de Jaccard, sendo a distância genética representada nos dendrogramas
empregando-se o método de agrupamento UPGMA (Unweighted Pair_Group
Method Using Arithmetic Avarage).

23
4 RESULTADOS E DISCUSSÃO
O protocolo de extração descrito por Hoisington et al. (1994), foi
eficiente para a extração do DNA de cana-de-açúcar, onde foi obtido
concentrações variáveis de 10 a 100 ng ∙ µL-1 (Figura 2). Silva (2006), utilizando
o mesmo protocolo para cana-de-açúcar obteve concentrações variáveis de DNA
entre 60 e 200 ng ∙ µL-1, sendo que essas diferenças de concentrações podem
estar relacionadas a fatores externos como a idade do material coletado e na
habilidade de desenvolvimento da técnica.
A temperatura de anelamento para cada primer está registrada na Tabela
2; foram realizadas várias reações para padronização dos primers cuja
temperatura efetiva não foi coincidente com a temperatura registrada na
literatura. Os primers que tiveram suas respectivas temperaturas alteradas para as
reações foram: SMC226CG (T °A = 50 °C); UGSM148 (T °A = 52 °C);
UGSM117 (T °A = 57 °C); SEGMS240 (T °A = 55 °C); SEGMS1069 (T °A =
55 °C); ESTB60 (T °A = 55 °C); mSSCIR56 (T °A = 56 °C). A metodologia de
PCR-Touchdown (PCR-TD) foi eficiente (Figura 3), evitando leituras
equivocadas de bandas inconsistentes. A PCR-TD possui o potencial de superar
problemas associados com altas temperaturas de anelamento requeridas por
algumas combinações de modelos de primers e, é particularmente útil para
modelos com dificuldades de amplificação, tais como aqueles com estruturas
extensíveis secundárias, com alto percentual de guanina e citosina no genoma e
com valor maior que 60% de G+C (Korbie e Mattick, 2008). A presença de
bandas não representativas pode interferir na leitura dos alelos, gerando
informações equivocadas e diminuindo o poder informativo do marcador SSR.
Portanto a padronização das temperaturas de anelamento foi fundamental para a
real caracterização dos genótipos de cana-de-açúcar estudados, assim como pode
ser observado na Figura 3A, onde bandas não alélicas de diversos pesos
moleculares apresentaram-se visíveis após a eletroforese durante 4 horas com 60
V. Após padronização da temperatura de anelamento é possível ver o mesmo

24
primer da Figura 3A, porém com os reais alelos característicos do genótipo em
questão.
Foi encontrado um total de 70 alelos para os dezessete pares de primers.
Dos dezessete primers utilizados para o estudo da diversidade genética entre e
dentro das variedades de cana-de-açúcar, apenas os primers UGSM148 e
UGSM59 foram monomórficos para as variedades estudadas. Portanto, o
polimorfismo nos locos SSR das nove variedades de cana-de-açúcar foi 88,2%.
Valores altos de polimorfismo nos locos SSR de cana-de-açúcar também foram
descritos por Cordeiro et al. (2000) que encontraram 91% e por Oliveira et al.
(2009) que encontraram 93% de polimorfismo trabalhando com EST-SSR em
Saccharum sp., evidenciando assim, o potencial desses marcadores para o estudo
da diversidade genética de cana-de-açúcar.
O número de alelos nos locos SSR polimórficos variou de dois alelos
(primers SGMS240, ESTB145) a oito alelos (primer mSSCIR56), sendo uma
média de 4,12 alelos por locos. Duarte-Filho et al. (2010) encontrou uma
quantidade média de 3,2 alelos por locos usando 18 marcadores microssatélite
em 30 variedades República do Brasil (RB).
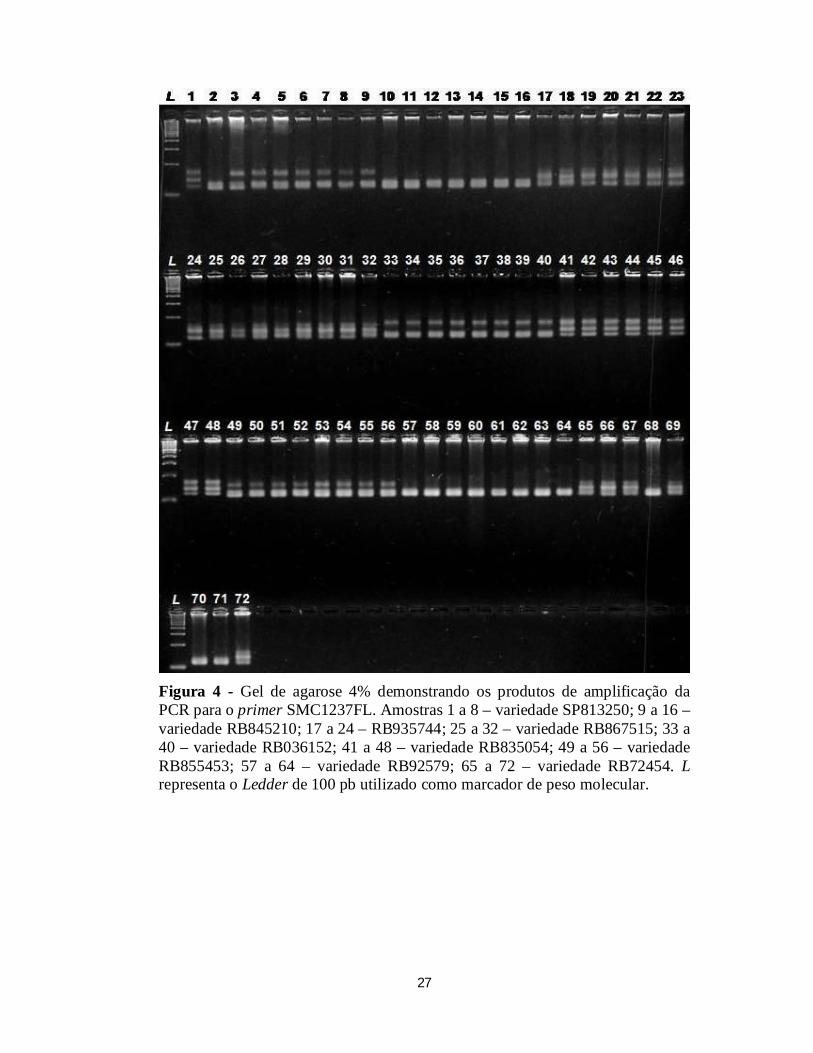
A Figura 4 apresenta os produtos de amplificação do primer
SMC1237FL altamente polimórfico, sendo evidente o polimorfismo entre e
dentro das variedades. As variedades RB72454 e RB845210 foram as que
apresentaram maior número de locos polimórficos, apresentando 100% de
polimorfismo para os primers utilizados no presente estudo. As variedades
RB835054 e RB855453 apresentaram menor número de locos polimórficos
(33,3% para as duas variedades).

25
Figura 2 - Gel de agarose 0,8% utilizado na quantificação das amostras de DNA das variedades de cana-de-açúcar. As amostras 3 e 4 correspondem à variedade RB867515. As amostras 5 a 13 correspondem à variedade RB036152. O DNA padrão (fago λ) é representado nas amostras 1 e 2 sendo de concentração respectiva de 50 e 100 ng ∙ µL-1 .
A análise do polimorfismo nos 15 locos SSR de Saccharum spp. mostrou
que os locos Smc226CG, Smc1237FL, Smc1047HA, mSscir56, e EstB60
apresentaram os valores maiores (Ho = 1,000) para as estimativas de
heterozigosidade observada (Tabela 3), e no loco Smc1047HA foi encontrado o
maior número de alelos indicando que estes locos podem ser recomendados para
analisar divergências genéticas entre variedades de cana-de-açúcar. Nos locos
Segms1069, EstB130, Smc165A, EstB41, Ugsm38, e EstC66 as estimativas de
heterozigosidade observada também foram altas (Ho > 0,50) e os primers para
estes locos podem ser indicados para avaliar a diversidade genética entre
variedades de cana-de-açúcar.

26
Figura 3 - Gel de agarose 4% demonstrando os produtos de amplificação da PCR para primers e temperaturas específicas pré e pós-padronização. A- Primer SMC226CG com T°A de 50°C (pré padronização). B- Primer SMC226CG pelo método PCR-TD (pós padronização). C- Primer SEGMS2405 com T°A de 55°C (pré-padronização). D- Primer SEGMS2405 pelo método PCR-TD (pós-padronização). A amostra 1 é representado pela variedades RB867515, 2 e 3 pela variedade RB036152.
27
Figura 4 - Gel de agarose 4% demonstrando os produtos de amplificação da PCR para o primer SMC1237FL. Amostras 1 a 8 – variedade SP813250; 9 a 16 – variedade RB845210; 17 a 24 – RB935744; 25 a 32 – variedade RB867515; 33 a 40 – variedade RB036152; 41 a 48 – variedade RB835054; 49 a 56 – variedade RB855453; 57 a 64 – variedade RB92579; 65 a 72 – variedade RB72454. L representa o Ledder de 100 pb utilizado como marcador de peso molecular.

28
Tabela 3 - Parâmetros de diversidade genética avaliados para as nove variedades de cana-de-açúcar obtidos na análise dos dezessete locos SSR Locos N Na Ne Ĥo Ugsm38 74 3,0 2,7552 0,2838 Segms240 74 2,0 1,6172 0,4865 Segms1069 74 3,0 2,9104 0,7973 Smc226CG 74 6,0 2,9221 1,0000 EstB130 74 5,0 3,4473 0,9865 Smc1237FL 74 6,0 3,9283 1,0000 Smc165A 74 5,0 2,9883 0,5676 Ugsm117 74 3,0 2,5715 0,2838 EstB41 74 5,0 2,9640 0,6486 EstB145 74 2,0 1,7951 0,2838 Smc1047HA 74 7,0 4,7411 1,0000 Smc2017FL 74 4,0 3,1336 0,8919 EstC66 74 3,0 2,5679 0,6892 mSsir56 74 8,0 3,0212 1,0000 EstB60 74 6,0 2,5904 1,0000 Média 4,12 2,9302 0,7279
Na=número de alelos observados; Ne=número efetivo de alelos; Ho=heterozigosidade observada.
Os valores altos para heterozigosidade observada em 73,3% dos locos
SSR e nas nove variedades comercias de Saccharum ssp. contrariam as
descrições de baixa variabilidade genética e de uniformidade genética nas
variedades de cana-de-açúcar (Barbosa, 2000; Almeida, 2005). Somente nos
locos Ugsm148 e Ugsm59 foi verificada uniformidade genética (100% de plantas
homozigotas) indicando que estes locos são adequados para monitorar a
estabilidade de genótipos quando for necessário. Os primers UGSM148 e
UGSM59 podem ser usados, por exemplo, como marcadores moleculares para
monitorar a estabilidade de determinada variedade durante os ciclos de rebrota,
para indicar em qual ciclo se verifica a ocorrência de variação no referido loco.
Desta forma, a uniformidade genética descrita para as diversas
características morfológicas e agronômicas das variedades de cana-de-açúcar não
podem ser estendidas para os 88,8% dos locos SSR analisados nas variedades
RB72454, RB845210, RB935744, RB867515, RB036152, RB835054,

29
RB855453, RB92579, e SP813250. O significado funcional dos locos SSR em
cana-de-açúcar é desconhecido. Em outras espécies o significado funcional dos
locos SSR tem sido motivo de especulações por grupos de investigadores.
Enquanto os locos SSR têm sido considerados como marcadores moleculares
evolutivamente neutros (Schlötterer and Wiehe, 1999; Schlötterer, 2000), alguns
pesquisadores tem indicado que os microssatélites com motivos repetidos
diferentes podem estar envolvidos na determinação da estrutura de cromossomos
(Subirana and Messeguer, 2007), ou estarem contidos em genes que apresentam
expressão diferencial durante diferentes estágios de desenvolvimento (Ince et al.,
2010). Portanto, os valores altos para a heterozigosidade observada nos locos
SSR de cana-de-açúcar pode ser decorrente do caráter neutro e independente dos
alelos que conferem características selecionadas para o melhoramento de cana-
de-açúcar (seleção artificial), ou resultado dos cruzamentos interespecíficos e/ou
de heterose. Valores de heterozigosidade média observada altos (1,000 e 0,9865),
intermediários (0,6892 e 0,6486), e baixo (0,2838) foram observados nos locos
microssatélites de sequências expressas (Expressed Sequence Tag) EstB60,
EstB130, EstC66, EstB41, e EstB145, respectivamente (Tabela 3), de Saccharum
spp.
Apesar dos marcadores microssatélites apresentarem característica
codominante, a frequência dos alelos nos locos SSR de cana-de-açúcar não pode
ser estimada porque por ser um híbrido com genoma poliplóide, em alguns locos
foram evidentes de dois a oito alelos. Por isso os marcadores microssatélites
(sequências amplificadas) foram considerados como marcadores dominantes
nesse trabalho. Não foi possível calcular os valores de heterozigosidade média
esperada para cada variedade, mas foi possível analisar a heterozigosidade média
observada para dois ou mais alelos (Ĥo = 1-Pii, em que Pii é a frequência de
genótipos homozigotos) diretamente através da leitura dos géis com as
amplificações.
As variedades RB72454 (Ĥo=0,833), RB92579 (Ĥo=0,808) e RB867515
(Ĥo=0,792) apresentaram os valores mais altos de heterozigosidade média
observada (Tabela 4). Para um marcador genético, um loco com heterozigosidade

30
maior do que 70% é comumente considerado um marcador altamente polimórfico
(Ott, 1992).
Tabela 4. Heterozigosidade média observada (Ĥo), número total de alelos encontrados e porcentagem de locos em heterozigose (%P) para cada variedade de cana-de-açúcar.
Variedade Na/loco Ne Hom(o) Ĥo (%P)
SP813250 2,823 1,744 0,4250 0,5750 87,50 RB845210 2,823 1,897 0,3500 0,6500 87,50 RB935744 2,94 2,05 0,2580 0,7420 100 RB867515 2,47 2,01 0,2080 0,7920 93,75 RB036152 2,35 1,92 0,2670 0,7330 81,25 RB835054 2,59 1,76 0,3420 0,6580 75 RB855453 2,29 1,83 0,2670 0,7330 87,50 RB92579 2,59 2,03 0,1920 0,8080 93,75 RB72454 3,12 2,31 0,1670 0,8330 100 Total 4,12 2,93 0,2720 0,7280
Na: número de alelos; Ne: Número efetivo de alelos; Hom(o): Homozigosidade
média observada; Ĥo: heterozigosidade média observada
Das três variedades que apresentaram a maior heterozigosidade
observada, duas são as variedades mais cultivadas no Brasil (RB867515 e RB
72454). É possível que o alto polimorfismo encontrado nessas variedades seja o
fator que favorece o estabelecimento das culturas, tornando-as menos suscetíveis
a problemas como doenças e clima.
A variedade RB72454 segundo censo varietal de 2011 (RIDESA, 2011;
www.ridesa.com.br) foi a sexta variedade mais cultivada no Brasil, e a sua ampla
adaptabilidade justifica sua utilização em várias regiões do país. Essa variedade é
amplamente utilizada como genitor de outras variedades de cana-de-açúcar
(Tabela 1; Figura 4) para obtenção de cultivares que se desenvolvam nos
diversos tipos de solos presente nos canaviais brasileiros. A alta heterozigosidade
observada para a variedade RB72454 pode justificar a sua utilização como

31
genitora de outras variedades propiciando combinações genotípicas que
aumentam a capacidade de adaptação das variedades a possíveis mudanças
ambientais (Sebbenn et al., 2000). A variedade RB72454 foi usada como
genitora das variedades RB845210, RB867515, e RB835054. Com o aumento da
área de plantio na região norte e noroeste do Paraná em regiões de solo com
textura superficial franco-arenoso, elevada acidez, com baixa fertilidade natural
(Sambatti, 2003; Fidalski, 1997), os programas de melhoramento têm o desafio
de desenvolver variedades mais resistentes e com ampla adaptabilidade; a alta
heterozigosidade observada na variedade RB72454 favorece sua utilização no
desenvolvimento dessas novas variedades.
A variedade RB92579 liberada comercialmente em 2003, também
apresentou alta heterozigosidade (0,8080), e segundo o censo varietal de 2011, a
variedade passou do 19° lugar em plantio geral em 2010 (Censo
Varietal/PMGCA, 2010) para o terceiro lugar no censo varietal de 2011, sendo
mais explorada devido sua baixa floração que favorece maiores teores de
sacarose no colmo. A variedade RB867515 foi a variedade mais plantada em
2010 e 2011, ocupando 1.311.017 ha em 2011, obtendo excelente produtividade;
sua heterozigosidade foi alta (0,7920), porém menor do que a da sua genitora
RB72454 (0,8330). A alta heterozigosidade observada (Ho > 0,50) é uma
característica comum a todas as nove variedades de cana-de-açúcar que,
igualmente, apresentam alta produtividade (Tabela 1).
A diversidade genética estimada com base nos coeficientes de Identidade
Genética de Nei (1978) indicou uma base genética estreita para as nove
variedades de cana-de-açúcar. A diversidade genética calculada mostrou maior
identidade genética entre as variedades RB92579 e RB72454 com 86,31% de
similaridade e a menor similaridade encontrada foi entre as variedades
RB845210 e RB835054 (61,99%). A base genética estreita entre essas variedades
é demonstrada pelo baixo valor de variação entre o mais similar e o menos
similar (24,32%) para os primers utilizados neste trabalho. Muitas das variedades
utilizadas no presente trabalho têm genitores comuns em alguma etapa do
melhoramento como pode ser observado na Tabela 1 e na Figura 5, contribuindo

32
para o estreitamento da base genética entre as nove variedades, considerando os
locos SSR analisados.
Figura 5 - Genealogia de 4 variedades utilizadas nesse trabalho. A – RB72453; B – RB935744; C – RB92579; D – RB867515. (?): Policruzamento. RIDESA (2010).
Na Tabela 5 é possível observar as similaridades e as distâncias entre as
variedades segundo Nei (1978). A maior distância genética encontrada entre as
variedades RB935744 e RB92579 (0,4874), RB845210 e RB835054 (0,4782),
RB92579 e RB835054 (0,4749), RB855453 e RB92579 (0,4098), torna
promissora o cruzamento entre essas variedades gerando populações segregantes
mais divergentes entre si.

33
Ao analisar a Tabela 5 é possível destacar a variedade RB92579 como a
que mais se distancia das outras variedades, exceto a variedade RB72454. Essa
variedade foi considerada pelos produtores do Nordeste brasileiro como a
variedade “Fantástica” por apresentar cerca de 30 a 40% de produtividade maior
que outras variedades cultivadas na região. A variedade RB92579 além de alta
produtividade apresenta ótimo perfilhamento, boa resistência a adversidades de
solo e clima (RIDESA, 2010), porém seu caldo é escuro prejudicando a sua
utilização pela indústria de açúcar no Brasil. Este é um exemplo de que para
algumas variedades de cana-de-açúcar, as excelentes características agronômicas
não estão associadas com as características desejáveis para o processamento
destas pela indústria de açúcar e álcool. Por isso é importante a caracterização
genética molecular destas variedades e as estimativas de divergência genética
entre elas, para orientar o melhoramento das características desejáveis pela
indústria. O agrupamento representado na Figura 6 organizou as nove variedades
em quatro grupos que representam a formação de grupos heteróticos em nível
molecular que podem ser usados pelos melhoristas, para orientar os cruzamentos
entre os grupos considerando as características morfoagronômicas e as de
interesse da indústria produtora de álcool e açúcar.
Tabela 5. Identidade (acima da diagonal) e Distância (abaixo da diagonal) genética de Nei (1978). SP813250(1); RB845210(2); RB935744(3); RB867515(4); RB036152(5); RB835054(6); RB855453(7); RB92579(8); RB72454(9)
Variedade 1 2 3 4 5 6 7 8 9 1 **** 0.8069 0.7030 0.7311 0.7717 0.6979 0.7649 0.6869 0.6654 2 0.2146 **** 0.7586 0.8160 0.6915 0.6169 0.8288 0.7283 0.7939 3 0.3524 0.2762 **** 0.7539 0.7163 0.8049 0.7961 0.6142 0.7330 4 0.3132 0.2034 0.2825 **** 0.7213 0.6522 0.8063 0.6703 0.7488 5 0.2592 0.3689 0.3336 0.3268 **** 0.6844 0.7182 0.6500 0.7183 6 0.3596 0.4782 0.2171 0.4273 0.3791 **** 0.7039 0.6219 0.6797 7 0.2680 0.1878 0.2281 0.2153 0.3310 0.3511 **** 0.6638 0.7514 8 0.3756 0.3171 0.4874 0.4000 0.4307 0.4749 0.4098 **** 0.8631 9 0.4074 0.2308 0.3106 0.2892 0.3309 0.3861 0.2859 0.1472 ****

34
Com base nas distâncias genéticas de Nei (1978) o dendrograma gerado
por agrupamento das variedades, pode ser observado na Figura 6.
A estimativa da divergência genética entre as nove variedades de cana-
de-açúcar quantificadas usando o coeficiente GST de Nei (1975) também mostrou
que apesar destas variedades apresentarem características morfoagronômicas
similares (Tabela 1) elas estão geneticamente estruturadas (GST = 0,7034).
Figura 6. Agrupamento obtido para as nove variedades de cana-de-açúcar, realizado através do método UPGMA e obtida a partir da análise de 70 alelos amplificados com primers para SSR.·.
Valores de GST ≥ 0,25 indicam que as populações analisadas apresentam
uma divergência genética muito alta decorrente de frequências diferentes dos
alelos nestas populações. Não foi verificada a presença de alelos exclusivos em
uma ou outra variedade, mas as frequências dos alelos comuns nas nove
variedades foram diferentes o suficiente para diferenciar geneticamente estas
variedades.
O dendrograma construído a partir da análise dos alelos como
marcadores dominantes, usando o programa NTSYSpc versão 2.1 (Rohlf, 2000)
para comparar as 74 plantas das nove variedades ilustra claramente a

35
variabilidade genética contida nos 17 locos microssatélites de cada uma das
variedades comerciais de cana-de-açúcar (Figura 7). A organização das plantas
de cada genótipo num determinado grupo mostra como as variedades estão
altamente estruturadas, e a organização das plantas das variedades RB92579 e
RB72454 num grupo, ilustra a maior similaridade (0,8631) observada entre estas
duas variedades. Este dendrograma ilustra também a presença de plantas com
100% de similaridade (clones) para os 17 locos microssatélites somente nas
variedades SP813250 (plantas 4 e 5; 7 e 8) que foi considerada como
geneticamente mais uniforme, RB935744 (plantas 21 e 22) e RB835054 (plantas
43,44 e 45).

36
Figura 7. Agrupamento obtido para as 74 plantas das nove variedades de cana-de-açúcar, realizado através do método UPGMA e obtido a partir da análise de 70 alelos amplificados com primers para SSR. I- SP13250; II-RB036152; III-RB935744; IV-RB835054; V- RB845210; VI- RB867515; VII- RB855453; VIII- RB72454; IX – RB92579.
I
II
III
IV
V
VI
VII
VIII
IX
VIII

37
A amplitude de variação alélica entre as nove variedades foi baixa (0,6142
entre as variedades RB92579 e RB935744, e 0,8631 entre as variedades
RB92579 e RB72454) concordando com a base genética estreita que tem sido
descrita para os genótipos de cana-de-açúcar (Barbosa, 2000; Almeida, 2005;
Duarte-Filho et al., 2010; Silva et al., 2012). Por outro lado, no dendrograma da
Figura 7 está representada uma base genética ampla (0,45 – 1,00) quando oito ou
dez exemplares de plantas de cada variedade foram comparados analisando os 17
locos microssatélites. Os estudos com outras variedades de cana-de-açúcar
comparam somente as variedades, tomando como amostras bulk de cada
variedade (Singh et al., 2008; 2011), numa análise que não identifica os
genótipos heterozigotos para expressar a variação contida dentro de cada
variedade.
No presente estudo, considerando o número de alelos (4,12 alelos/locos),
os valores de heterozigosidade média observados (de 0,575 a 0,833), e os índices
de similaridade genética entre as plantas (0,44 a 1,00), como parâmetros para a
análise de diversidade genética nas nove variedades de cana-de-açúcar usando os
17 primers de microssatélites, foi possível verificar uma diversidade genética alta
e uma base genética ampla para as amostras da espécie Saccharum spp..

38
5 CONCLUSÃO
5.1 A análise dos locos Ugmsm38, Segms240, Segms1069, Smc226CG,
EstB130, Smc1237FL, Smc165A, Ugsm117, EstB41, EstB145, Smc1047HA,
Smc2017FL, EstC66, mSscir56, e EstB50, mostrou que existe uma diversidade
genética alta dentro das variedades SP813250, RB845210, RB935744,
RB867515, RB036152, RB835054, RB855453, RB92579, e RB72454 de cana-
de-açúcar, e uma divergência genética alta entre estas variedades.
5.2 Os locos Ugsm148 e Ugsm59 (100% de plantas homozigotas) podem ser
indicados para monitorar a estabilidade dos nove genótipos de cana-de-açúcar,
quando for necessário.
5.3 Os primers para os locos Segms1069, EstB60, Smc226CG, Est130,
Smc1237FL, Smc165A, EstB41, Smc1047HA, Smc2017FL, EstC66, e mSscir56
(Ho > 0,50), são promissores para avaliar a diversidade genética dentro e entre
variedades de cana-de-açúcar.
5.4 Os locos SSR em variedades de cana-de-açúcar são marcadores promissores
para avaliar a real diversidade genética em nível molecular de variedades que
apresentam características morfoagronômicas uniformes, contrariando o
postulado de que existe uniformidade genética em variedades comerciais de
cana-de-açúcar.

39
6 REFERÊNCIAS
ALMEIDA, C.M.C.V.; DIAS, L.A.S.; OKABE, E.T.; MEDEIROS, J.R.P. Variability in genetic resources of cação in Rondonia, Brazil. Crop Breending and Applied Biotechnology, v.5, p.318-324, 2005.
BARBOSA, G.V.S.; SOUZA, A.J.R.; ROCHA, A.M.C.; RIBEIRO, C.A.G.; FERREIRA, J. L. C.; SOARES, L.; CRUZ, M. M.; SILVA, W. C. M. Novas variedades RB de cana-de-açúcar para Alagoas. Programa de Melhoramento Genético de Cana-de-Açúcar, 2000. 16p. (Boletim Técnico Programa de Melhoramento Genético de Cana-de-Açúcar).
BARBOSA, M.H.P. Study of genetic divergence in sugarcane varieties grown in Brazil using the parentage coefficient. International Sugar Jornal, v.103, p. 294-295, 2001.
BARRETO, A.; SIMONS, J.P. Identification de progenies Y progenitors por el analisis Del numero cromosomico en Saccharum. Turrialba, v.32, p.169-180, 1982.
BORÉM, A.; VIEIRA, M.L.C.; COLLI, W. Glossário de Biotecnologia. 2ed. UFV, 2009, 186p.
BORÉM, A.; CAIXETA, E.T. Marcadores Moleculares. 2ed. UFV, p.33-54, 2009.
BORBA. T.C.O.; MENDES, C.A.; GUIMARAES, E.P.; BRUNES, T.O.; FONSECA, J.R.; BRONDANI, R.V.; BRONDANI, C. Genétic variability of brasilian rice landraces determined by SSR markers. Pesquisa Agropecuaria Brasileira, v.44, p.706-712, 2009.
BOTSTEIN, D.; WHITE, R.L.; SKOLNICK, M; DAVIS, R.W. Construction of a genetic linkage map in man using restriction fragment length polymorphism. American Journal of Human Genetics, v.32, p 315 - 330, 1980.
CANA WEB/DADOS E ESTATÍSTICA. Disponível em: http://www.jornalcana.com.br. Acesso em: 19 jan. 2012.
CORDEIRO, G.M.; PAN, Y.B.; HENRY, R.J. Sugarcane microsatellites for the assessment of genetic diversity in sugarcane germoplasm. Plant Science, v.165, p.181-189, 2003.
CORDEIRO G.M.; TAYLOR G.O.; HENRY; R.J. Characterisation of microsatellite markers from sugarcane (Saccharum sp.), a highly polyploid species. Plant Science, v.155, p.161-168, 2000.

40
CRUZ, C.D.; REGAZZI, A.J.; CARNEIRO, P.C.S. Modelos biométricos aplicados ao melhoramento genético. Universidade Federal de Viçosa, 3ed., v.1, 2004. 480p.
D’HONT, A.; GRIVET, L.; FELDMAN, P.; RAO, S.; BERDING, N.; GLASZMANN, J.C. Characterization of the double genome structure of modern sugarcane cultivars (Saccharum spp.) by molecular genetics. Molecular and General Genetics, v.250, p. 405-413, 1996.
DUARTE-FILHO, L.S.C.; SILVA, P.P.; SANTOS,; J.M. ;BARBOSA, G.V.S.; RAMALHO-NETO, C.E.; SOARES, L.; ANDRADE, J.C.F.; ALMEIDA, C. Genetic similarity among genotypes of sugarcane estimated by SSR and coefficient of parentage. Sugar Tech, v.12, p.145-149, 2010.
EPE - EMPRESA DE PESQUISA ENERGÉTICA. Disponível em: http://www.epe.gov.br. Acesso em: 21 dez. 2011.
FERREIRA, M.E.; GRATTAPAGLIA, D. Introdução do uso de marcadores moleculares em análise genética. EMBRAPA/CENARGEM, 1998. 220p.
FERREIRA, F. Caracterização cromossômica em cana-de-acúcar (Saccharum spp., Poaceae. Unicamp, 94p. Dissertação (Mestrado) - Programa de Pós-graduação do Instituto de Biologia, Universidade Estadual de Campinas, Campinas, 2010.
FIDALSKI, J. Fertilidade do solo sob pastagens, lavouras anuais e permanentes na região noroeste do Paraná. Revista Unimar, v.19, p.853-861, 1997.
FIGUEIREDO, P. Breve história da cana-de-açúcar e do papel do instituto agronômico no seu estabelecimento no Brasil. In: Dinardo-Miranda, L.L; Vasconcelos, A.C.M.; Landell, M.G.A. (Ed.). Cana-de-açúcar. IAC, 2008.
HAMADA, H.; PETRINO, M.G.; KAKUNAGA, T. A novel repeated element with Z-DNA-forming potential is widely found in evolutionarily diverse eukaryotic genomes. Proceedings of the National Academy of Sciences of the United State of America, v.79, p. 6465-6469, 1982.
HOISINGTON, D.; KHAIRALLAH, M.; GONZÁLEZ-LÉON, D. Laboratory Protocols: CIMMYT Applied Molecular Genetics Laboratory. 2ed. CIMMYT, 1994, 50p.
JACOB, H.J.; LINDPAINTER, K.; LINCOLN, S.E.; KASUMI,K.; BUNKE R.K.; MAO, Y. P.; GANTEN, D.; DZAU, V. J.; LANDER, E.S. Genetic mapping of a gene causing hypertensive rat . Cell, v.67, p.213-224, 1991.
KORBIE, D.J.; MATTICK, J.S. Touchdown PCR for increased specificity and sensitivity in PCR amplification. Nature Protocols, v.3, p.1452-1456, 2008.

41
LAGERCRANTZ, U.; ELLEGREN, H.; ANDERSON, L. The abundance of various polymorphic microsatellite motifs differs between plants vertebrates. Nucleic Acids Research, v.21, p.1111-1115, 1993.
LIMA, M.L.A.; GARCIA, A.A.F.; OLIVEIRA, K.M.; MATSUOKA, S.; ARIZONO, H.; SOUZA JR., C.L.; SOUZA, A.P. Analysis of genetic similarity detected by AFLP and coefficient of parentage among genotypes of sugar cane (Saccharum spp.). Theorical and Applied Genetic, v.104, p.30-38, 2002.
LITT, M.; LUTY, J.A. A hypervariable microsatellite revealed by in-vitro amplification of a dinucleotide repeat within the cardiac muscle actin gene. The American Journal of Human Genectics, v.44, p.397- 401, 1989.
MATSUOKA, S.; GARCIA, A.A.F.; ARIZONO, H. Melhoramento de cana-de-açúcar. In: BOREM, A. (Ed.). Melhoramento de espécies cultivadas, Imprensa Universitária, p.205-251, 1999.
MATSUOKA, S.; GARCIA, A.A.F.; ARIZONO, H. Melhoramento de cana-de-açúcar. In: BOREM, A. (Ed.). Melhoramento de espécies cultivadas, 2ed. UFV, p.225–274, 2005.
MCINTYRE, L.; AITKEN, K.; BERDING, N.; CASU, R.; DRENTH, J.; JACKSON, P.; JORDAN, D.; PIPERIDIS, G.; REFFAY, N.; SMITH, G.; TAO, Y.Z.; WHAN, V. Identification of DNA markers linked to agronomic traits in sugarcane in Australia, in: XXIV Proceeding of International Society of Sugar Cane Technology, v.24, p.560-562, 2001.
MEIRELLES, P.G. Divergência genética entre linhagens de milho estimadas por microssatélites e correlação com desempenho de híbridos simples. 92p. Tese (Doutorado) - Programa de Pós-graduação em Agronomia, Universidade Estadual Paulista Julio Mesquita Filho, 2009.
MING, R.; MOORE, P.H.; WU, K.; D´HONT, A.; GLAZMANN, J.C.; TEW, T.L.; MIRKOV, T.E.; DA SILVA, J.; JIFON, J.; RAI, M.; SCHNELL, R.J.; BRUMBLEY, S.M.; LAKSHMANAN, P.; COMSTOCK, J.C.; PATERSON, A.H. Sugarcane improvement through breeding and biotechnology. Plant Breeding Reviews, v.27, p.15-118, 2006.
MINISTÉRIO DO DESENVOLVIMENTO, INDÚSTRIA E COMÉRCIO EXTERIOR. Disponível em http://www.mdic.gov.br. Acesso em: 19 jan. 2012.
NEI, M. Estimation of average heterozygosity and genetic distance from a small number of individuals, Genetics v.89, p.583-590, 1978.
OLIVEIRA, K.M.; PINTO, L.R.; MARCONI, T.G.; MOLLINARI, M.; ULIAN, E.C.; CHABREGAS, S.M.; FALCO, M.C.; BURNQUIST, W.; GARCIA, A.A.F.; SOUZA, A.P. Characterization of new polymorphic functional markers for sugarcane. Genome, v.52, p.191-209, 2008.

42
OTT, J. Strategies for characterizing highly polymorphic markers in human gene mapping. American Journal of Human Genetics, v. 51, p. 283–290, 1992.
PALHARES, A.C. Mapeamento genético de marcadores AFLP e de retrotransposons em cana-de-açucar (Saccharum spp.). 97p. Dissertação (Mestrado) - Programa de Pós-graduação em Ciências, Universidade de São Paulo, 2010.
PALOMINO, E.C.; MORI, E.S.; ZIMBACK, L.; TAMBARUSSI, E.V.; MORAES, C.B. Genetic diversity of common bean genotypes of Carioca commercial group using RAPD markers. Crop Breeding and Applied Biotechnology, v.5, p.80-85, 2005.
PAN, Y-B.; CORDEIRO, G.M.; HENRY, R.J.; SCHNELL, R.J. Microsatellite fingerprints of Louisiana sugarcane varieties and breeding lines, in: Xth International Conference on the Status of Plant and Animal Genome Research, p.45. 2002.
PAN, Y-B. Highly polymorphic microsatellite DNA markers for sugarcane germplasm evaluation and variety identity testing. Sugar Tech, v.8, p.246-256. 2006.
PANDEY, A.; SOCCOL, C.; SOCCOL, V. Biotechnological potential of agro-industrial residues. I: Sugarcane bagasse. Bioresource Technology, v.74, p.69, 2000.
PARIDA, S.K.; KALIA, S.K.; KAUL, S.; DALAL, V.; HEMAPRABHA, G.; SELVI, A.; PANDIT, A.; SINGH, A.; GAIKWAD, K.; SHARMA, T.R.; SRIVASTAVA, P.S.; SINGH, N.K.; MOHAPATRA, T. Informative genomic microsatellite markers for efficient genotyping applications in sugarcane. Theorical and Applied Genetic, v.118, p.327-338, 2009.
POWELL, W.; MACHRAY, G.C.; PROVAN, J. Polymorphism revealed by simple sequence repeats. Trends Plant Science, v.1, p.215–222, 1996.
RALLO, P.; DORADO, G.; MARTÍN, A. Development os simple sequence repeats (SRRs) in olive tree (Olea europaea L.). Theoretical and Applied Genétics, v.101, p.984-989, 2000.
RIDESA. Catálogo nacional de variedades “RB” de cana-de-açúcar. 1ed. UFPR, 136p., 2010.
ROHLF, F.J. NTSYS-pc: Numerical Taxonomy and Multivariate Analysis System, version 2.1. Exeter Software: Setauket, 2000.
RUZENE, D.S.; SILVA, D.P.; VICENTE, A.A.; TEIXEIRA, J.A.; AMORIM, M.T.P.; GONÇALVES, A.R. Cellulosic films obtained from the treatment of sugarcane bagasse fibers with N-methylmorpholine-N-oxide (NNMO). Applied Biochemistry and Biotechnology, v.154, p.217-226, 2009.

43
SALEM, K.F.M.; EL-ZANATY, A.M.; ESMAIL, R.M. Acessing Wheat (Triticum aestivum L.) Genetic Diversity Using Morphological Characters and Microsatellites Markers. World Journal of Agriculture Sciences, v.4, p.538 – 544, 2008.
SEBBENN, A.M.; KAGEYAMA, P.Y.; SIQUEIRA, A.C.M.F.; ZANATTO, A.C.S. Sistema de cruzamento em populações de Cariniana legalis Mart. O. Ktze.: implicações para a conservação e o melhoramento genético. Scientia Forestalis, v.58, p.25-40, 2000.
SCHUSTER, I.; VIEIRA, E.S.N.; PADILHA, L. Marcadores moleculares no pós-melhoramento. In: BORÉM, A.; CAIXETA, E.T. Marcadores moleculares. Universidade Federal de Viçosa, 2006. p.205-230.
SAMBATTI, J.A.; SOUZA JUNIOR, I.G.; COSTA, A.C.S.; TORMENA, C.A. Estimativa da acidez potencial pelo método do pH SMP em solos da formação Caiuá: noroeste do estado do Paraná. Revista Brasileira de Ciência do Solo, v. 27, p.257-264, 2003.
SELVI, A.; NAIR, N.V.; NOYER, J.L.; SINGH, N.K.; BALASUNDARAM, N.; BANSAL, K.C.; KOUNDAL, K.R.; MOHAPATRA, T. Genomic constitution and genetic relationship among the tropical and subtropical Indian sugarcane cultivars revealed by AFLP. Crop Science, v.45, p.1750-1757, 2005.
SILVA, C.M. Diversidade genética em variedades de cana-de-açúcar (Saccharum spp.) micropropagadas e propagadas vegetativamente. 45p. Tese (Doutorado) – Programa de Pós-graduação em Agronomia, Universidade Estadual de Maringá, Maringá, 2006a.
SILVA, P.P. Divergência genética em genótipos de cana-de-açúcar (Saccharum spp.) através de caracteres morfoagronômicos e por marcadores moleculares. 96p. Dissertação (Mestrado) – Universidade Federal de Alagoas, Rio Largo, 2006b.
SILVA, G.C.; OLIVEIRA, F.J.; ANUNCIAÇÃO-FILHO, C.J.; SIMOES-NETO, D.E.; MELO, L.J.O.T. Divergência Genética entre Genótipos de Cana-de-açúcar. Revista Brasileira de Ciências Agrarias, v.6, p.52-58, 2011.
SILVA, A.L. Caracterização molecular e variabilidade genética de cultivares brasileiras de trigo, obtidas por marcadores microsstélites. 2010. 61p. Dissertação (mestrado) – Universidade Paranaense, 2010.
SILVA, C.M.; MANGOLIN, C.A.; MOTT, A.S.; MACHADO, M.F.P.S. Genetic diversity associated with in vitro and conventional buck propagation of Saccharum variety using RAPD analysis. Plant Breeding, v.127, p. 160-165, 2008.

44
SIMÕES NETO, D.E.; MOREIRA, C.N.; LIMA, R.O.R.; MELO, L.J.O.T. Desempenho de genótipos de cana-de-açúcar (Saccharum spp) em diferentes ambientes do Estado de Pernambuco. In: CONGRESSO NACIONAL DA STAB, p.29-33, 1999.
SINGH, R.K.; SRIVASTAVA, S.; SINGH, S.P.; SHARMA, M.L.; MOHOPATRA, T.; SINGH, N.K.; SINGH, S.B. Indentification of new microsatellites DNA markers for sugar and related traits in sugarcane. Sugar Tech, v.10, p.327-333, 2008.
SOUSA, L.R.L. Viabilidade do uso de cinzas de bagaço de cana-deaçúcar (CBC) residuais do Estado de Goiás como adição mineral ao Cimento Portland, 120 p. Dissertação de Mestrado. Universidade Federal de Goiás, 2009.
WANG, Z.; WEBER, J.L.; ZHONG, G.; TANKSLEY, S.D. Survey of plant short tandem DNA repeats. Theoretical and Applied Genétics, v.88, p.1-6, 1994.
WEBER, Z.; MAY, P.E. Abundant class of human DNA polymorphism which can be typed using the polymerase chain reaction. The American Journal of Human Genectics, v.44, p.388-396, 1989.
WILLIAMS, J.G.K.; KUBELIK, A.R.; LIVAK, K.J.; RAFALSKI, J.A.; TINGEY, S.V. DNA polymorphism amplified by arbitrary are useful as genetic markers. Nucleic Acids Research, v.18, p.6531-6535, 1990.
VARSHNEY, R.K.; GRANER, A.; SORRELLS, M.E. Genic microsatellite markers in plants: features and applications. Trends in Plant Science, v.23, p.48–55, 2005.
VIEIRA, E.S.N.; SCHUSTER, I.; SILVA, R.B.; OLIVEIRA, M.A.R. Variabilidade genética em cultivares de soja determinada com marcadores microssatelites em gel de agarose. Pesquisa Agropecuaria Brasileira, v.44, p. 1460-1466, 2009.
VOS, P.; HOGERS, R.; BLEEKER, M.; REIJANS, M.; VAN DE LEE, T.; HORNES, M.; FRIJTERS, A.; POT, J.; PELEMAN, J.; KULPER, M.; ZABEAU, M. AFLP: a new technique for DNA fingerprinting. Nucleic Acid Research, v.23, p.4404 - 4414, 1995.
YEH, F.C.; BOYLE, T.Y.Z.; XIYAN, J.M. POPGENE version 1.31: Microsoft Window – based free ware for population genetic analysis. University of Alberta and Center for International Forestry Research, 1999.
ZANIN, G.M.; SANTANA, C.C.; BON, E.P.S.; GIORDANO, R.C.L.; MORAES, F.F.; ANDRIETTA, S.R.; CARVALHO NETO, C.C.; MACEDO, I.C., FO, D.L.; RAMOS, L.P., FONTANA, J.D. Brazilian Bioethanol Program. Applied Biochemistry and Biotechnology, v.84–86, p.1147-1161, 2000.




















