
Role of Saponins in Plant Defense Against Specialist Herbivores
21
molecules Review Role of Saponins in Plant Defense Against Specialist Herbivores Mubasher Hussain 1,2,3,4,5,6 , Biswojit Debnath 1 , Muhammad Qasim 2,7 , Bamisope Steve Bamisile 2,3,5,6 , Waqar Islam 3,8 , Muhammad Salman Hameed 3,6,9 , Liande Wang 2,3,4,5,6, * and Dongliang Qiu 1, * 1 College of Horticulture, Fujian Agriculture and Forestry University, Fuzhou 35002, China; [email protected] (M.H.); [email protected] (B.D.) 2 State Key Laboratory of Ecological Pest Control for Fujian and Taiwan Crops, Fujian Agriculture and Forestry University, Fuzhou 350002, China; [email protected] (M.Q.); [email protected] (B.S.B.) 3 College of Plant Protection, Fujian Agriculture and Forestry University, Fuzhou 350002, China; [email protected] (W.I.); [email protected] (M.S.H.) 4 Key Laboratory of Integrated Pest Management for Fujian-Taiwan Crops, Ministry of Agriculture, Fuzhou 350002, China 5 Key Laboratory of Biopesticide and Chemical Biology, Ministry of Education, Fuzhou 350002, China 6 Institute of Applied Ecology and Research Centre for Biodiversity and Eco-Safety, Fujian Agriculture and Forestry University, Fuzhou 350002, China 7 Ministry of Agriculture Key Lab of Molecular Biology of Crop Pathogens and Insects, Institute of Insect Science, Zhejiang University, Hangzhou 3100058, China 8 College of Geography, Fujian Normal University, Fuzhou 350007, China 9 Faculty of Agricultural Sciences, Department of Plant Protection, Ghazi University, Dera Ghazi Khan 32200, Pakistan * Correspondence: [email protected] (L.W.); [email protected] (D.Q.) Academic Editor: David Popovich Received: 10 May 2019; Accepted: 27 May 2019; Published: 30 May 2019 Abstract: The diamondback moth (DBM), Plutella xylostella (Lepidoptera: Plutellidae) is a very destructive crucifer-specialized pest that has resulted in significant crop losses worldwide. DBM is well attracted to glucosinolates (which act as fingerprints and essential for herbivores in host plant recognition) containing crucifers such as wintercress, Barbarea vulgaris (Brassicaceae) despite poor larval survival on it due to high-to-low concentration of saponins and generally to other plants in the genus Barbarea. B. vulgaris build up resistance against DBM and other herbivorous insects using glucosinulates which are used in plant defense. Aside glucosinolates, Barbarea genus also contains triterpenoid saponins, which are toxic to insects and act as feeding deterrents for plant specialist herbivores (such as DBM). Previous studies have found interesting relationship between the host plant and secondary metabolite contents, which indicate that attraction or resistance to specialist herbivore DBM, is due to higher concentrations of glucosinolates and saponins in younger leaves in contrast to the older leaves of Barbarea genus. As a response to this phenomenon, herbivores as DBM has developed a strategy of defense against these plant biochemicals. Because there is a lack of full knowledge in understanding bioactive molecules (such as saponins) role in plant defense against plant herbivores. Thus, in this review, we discuss the role of secondary plant metabolites in plant defense mechanisms against the specialist herbivores. In the future, trials by plant breeders could aim at transferring these bioactive molecules against herbivore to cash crops. Keywords: bioactive molecule; biological management; host plant resistance; plant immunity; plant secondary metabolites; triterpenoids Molecules 2019, 24, 2067; doi:10.3390/molecules24112067 www.mdpi.com/journal/molecules
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Role of Saponins in Plant Defense Against Specialist Herbivores

molecules
Review
Role of Saponins in Plant Defense AgainstSpecialist Herbivores
Mubasher Hussain 1,2,3,4,5,6 , Biswojit Debnath 1 , Muhammad Qasim 2,7 ,Bamisope Steve Bamisile 2,3,5,6 , Waqar Islam 3,8 , Muhammad Salman Hameed 3,6,9,Liande Wang 2,3,4,5,6,* and Dongliang Qiu 1,*
1 College of Horticulture, Fujian Agriculture and Forestry University, Fuzhou 35002, China;[email protected] (M.H.); [email protected] (B.D.)
2 State Key Laboratory of Ecological Pest Control for Fujian and Taiwan Crops, Fujian Agriculture and ForestryUniversity, Fuzhou 350002, China; [email protected] (M.Q.); [email protected] (B.S.B.)
3 College of Plant Protection, Fujian Agriculture and Forestry University, Fuzhou 350002, China;[email protected] (W.I.); [email protected] (M.S.H.)
4 Key Laboratory of Integrated Pest Management for Fujian-Taiwan Crops, Ministry of Agriculture,Fuzhou 350002, China
5 Key Laboratory of Biopesticide and Chemical Biology, Ministry of Education, Fuzhou 350002, China6 Institute of Applied Ecology and Research Centre for Biodiversity and Eco-Safety, Fujian Agriculture and
Forestry University, Fuzhou 350002, China7 Ministry of Agriculture Key Lab of Molecular Biology of Crop Pathogens and Insects, Institute of Insect
Science, Zhejiang University, Hangzhou 3100058, China8 College of Geography, Fujian Normal University, Fuzhou 350007, China9 Faculty of Agricultural Sciences, Department of Plant Protection, Ghazi University, Dera Ghazi Khan 32200,
Pakistan* Correspondence: [email protected] (L.W.); [email protected] (D.Q.)
Academic Editor: David PopovichReceived: 10 May 2019; Accepted: 27 May 2019; Published: 30 May 2019
�����������������
Abstract: The diamondback moth (DBM), Plutella xylostella (Lepidoptera: Plutellidae) is a verydestructive crucifer-specialized pest that has resulted in significant crop losses worldwide. DBM iswell attracted to glucosinolates (which act as fingerprints and essential for herbivores in host plantrecognition) containing crucifers such as wintercress, Barbarea vulgaris (Brassicaceae) despite poorlarval survival on it due to high-to-low concentration of saponins and generally to other plants inthe genus Barbarea. B. vulgaris build up resistance against DBM and other herbivorous insects usingglucosinulates which are used in plant defense. Aside glucosinolates, Barbarea genus also containstriterpenoid saponins, which are toxic to insects and act as feeding deterrents for plant specialistherbivores (such as DBM). Previous studies have found interesting relationship between the hostplant and secondary metabolite contents, which indicate that attraction or resistance to specialistherbivore DBM, is due to higher concentrations of glucosinolates and saponins in younger leaves incontrast to the older leaves of Barbarea genus. As a response to this phenomenon, herbivores as DBMhas developed a strategy of defense against these plant biochemicals. Because there is a lack of fullknowledge in understanding bioactive molecules (such as saponins) role in plant defense againstplant herbivores. Thus, in this review, we discuss the role of secondary plant metabolites in plantdefense mechanisms against the specialist herbivores. In the future, trials by plant breeders couldaim at transferring these bioactive molecules against herbivore to cash crops.
Keywords: bioactive molecule; biological management; host plant resistance; plant immunity; plantsecondary metabolites; triterpenoids
Molecules 2019, 24, 2067; doi:10.3390/molecules24112067 www.mdpi.com/journal/molecules

Molecules 2019, 24, 2067 2 of 21
1. Introduction
The capacity of individual plant species to develop novel metabolites has been affirmed in chargeof their imperviousness to plant herbivores. Plants have developed surprising diversity of substanceprotections against plant herbivores in light of bioactive mixtures of low atomic weight. A case of thebioactive mixtures utilized by plants in this regard are the triterpenoid saponins (Figure 1); whichencourages plant immunity against a wide range of insect pests, pathogens, as well as other herbivores.Molecules 2019, 24, x FOR PEER REVIEW 3 of 22
3
Figure 1. Triterpenoids saponins identified in Barbarea vulgaris.
2. Plant Defense and Evolution
A variety of plants is susceptible to environmental disputes, but could not escape. In spite of this evident exposure, the Earth’s flora has developed to be highly abundant and diverse. It’s a reality that not all plants are entirely consumed, this could be as a result of top-down control [22], also to bottom-up mechanisms such as the direct defense of plants in response to herbivores [23]. Plants might play a major role in top-down control of herbivores by enrolling natural enemies of their enemies as an indirect defense [24]. Regarding a wide range of herbivores, direct plant defense mechanisms can demand structural adjustments, for example, trichomes, thistles, and silica bodies or assistance some other natural products. Furthermore, the auxiliary metabolites that potentially built up are lethal to herbivores, or attract the natural foes of the herbivores [25]. Disregarding their name, derivative metabolites have a vital impact on the chemical communication between plants and their surroundings. They are of basic significance for the appeal of pollinators (terpenes), protecting the plant as opposed to UV light (flavonoids), pathogens and herbivores (alkaloids, glucosinolates, saponins). The majority of plants comprises a significant range of plant derivatives [26]. From a developmental point, this range is mystified however even ineffectively understood.
The reciprocal process of adaptation within plants and their insect herbivores was observed by Stahl [27], and he proposed that the synthetic mixtures may be included. These above thoughts were advanced by Ehrlich and Raven [23] to deliver a hypothetical background for the compound nature of insect and plant communications. They proposed a well-ordered biochemical co-advancement amongst plants and bugs. Unexpectedly, some herbivore species build up a resistance against biochemical compounds that are dangerous and distasteful to different insect pests. Gradually these biocompounds may possibly act as feeding stimulant or attractant for a particular insect, which has changed according to certain conditions and even utilizes some biochemicals as a guard, from the
Figure 1. Triterpenoids saponins identified in Barbarea vulgaris.
Triterpenoid saponins are mostly found in dicotyledonous species whereas monocots mainlysynthesis steroidal saponins. Some leguminous crops such as: Pea, sugar beet, soybeans, cowpea,asparagus, and capsicum peppers have been reported to contain saponins [1]. Saponins are consideredone of the immeasurable and distinct groups of bio-plant items, and categorize secondary plantmetabolites with particular natural properties [2,3]. Saponins content in plants is dynamic, and itinfluences various biotic stimuli that are related to pest attack, pathogenic infection, plant mutualisticsymbioses with rhizobial bacteria and mycorrhizal fungi [2]. About over 200 different structures ofsaponins had so far been described [4]. Likewise, Khakimov, et al. [5] reported that blends of differentchemical structures are accumulated by saponin producing plants. The biological activity of saponinscan be attributed to the amphipathic properties of the constituting molecules, which consist of ahydrophobic triterpene or sterol backbone and a hydrophilic carbohydrate chain. Some saponins havepotent biological activities that are influenced by other aspects of their structure.
Saponins have been reported from different and unrelated plant families [5]. Whereas cereals areinsufficient in saponins, aside from a few species of grass, such as Panicum virgatum, Panicum coloratum,and Avena spp. [6–8]. Additional gene families which have been reported to be involved in saponinbiosynthesis and diversification are methyl and acyltransferases [9,10]. Aside crucifers, saponins occurconstitutively in many other plant species as part of their defense system. For Brassicaceae, just a couple

Molecules 2019, 24, 2067 3 of 21
of species are known to yield saponins [11,12]. For example, the species from genus–rea {wintercress,B. vulgaris R. Br. (Brassicaceae)} are identified to create saponin; which are directly related to the plantdefense against specialist herbivore, such as the diamondback moth (Plutella xylostella) (Lepidoptera:Plutellidae) [5]. P. xylostella is a typical crucifer specialist that is known worldwide as a severe pest ofcruciferous crops, such as cabbage (Brassica oleracea) and oilseed rape (Brassica napus) [13,14].
Most of the glucosinolates–containing crucifers are all suitable hosts for the plant pest.The attractiveness of P. xylostella to these plant species is as a result of the glucosinolates content and itssecondary products; such as isothiocyanates [15]. These compounds have been found to stimulateoviposition by P. xylostella adults, as well as, feeding by the larvae [16]. A similar observation has alsobeen reported in cabbage butterfly (Pieris rapae); another crucifer specialist by Huang, Renwick andSachdev-Gupta [15]. In another related report, a highly feeding deterrent activity to P. xylostella larvaewas recorded in a chloroform extract of B. vulgaris leaves [17,18].
The management of P. xylostella has recorded minimum success as a result of its notoriousability to develop resistance to synthetic insecticides [13]. The ability of the pest to adapt plantsecondary metabolites for host plant recognition, feeding, and oviposition stimulants has also beenreported [19–21]. Moreover, inadequate knowledge of the biosynthetic paths and conducting systemsof saponins has additionally complicated its application for pest control. However, the prospect ofsaponins modification as direct plant defense strategies against pests has offered alternative controlmeasure for inclusion in an integrated pest management program for P. xylostella.
2. Plant Defense and Evolution
A variety of plants is susceptible to environmental disputes, but could not escape. In spite of thisevident exposure, the Earth’s flora has developed to be highly abundant and diverse. It’s a realitythat not all plants are entirely consumed, this could be as a result of top-down control [22], also tobottom-up mechanisms such as the direct defense of plants in response to herbivores [23]. Plants mightplay a major role in top-down control of herbivores by enrolling natural enemies of their enemiesas an indirect defense [24]. Regarding a wide range of herbivores, direct plant defense mechanismscan demand structural adjustments, for example, trichomes, thistles, and silica bodies or assistancesome other natural products. Furthermore, the auxiliary metabolites that potentially built up are lethalto herbivores, or attract the natural foes of the herbivores [25]. Disregarding their name, derivativemetabolites have a vital impact on the chemical communication between plants and their surroundings.They are of basic significance for the appeal of pollinators (terpenes), protecting the plant as opposedto UV light (flavonoids), pathogens and herbivores (alkaloids, glucosinolates, saponins). The majorityof plants comprises a significant range of plant derivatives [26]. From a developmental point, thisrange is mystified however even ineffectively understood.
The reciprocal process of adaptation within plants and their insect herbivores was observed byStahl [27], and he proposed that the synthetic mixtures may be included. These above thoughts wereadvanced by Ehrlich and Raven [23] to deliver a hypothetical background for the compound nature ofinsect and plant communications. They proposed a well-ordered biochemical co-advancement amongstplants and bugs. Unexpectedly, some herbivore species build up a resistance against biochemicalcompounds that are dangerous and distasteful to different insect pests. Gradually these biocompoundsmay possibly act as feeding stimulant or attractant for a particular insect, which has changed accordingto certain conditions and even utilizes some biochemicals as a guard, from the respective plant. It isuseful for the insect pest as the plant constitutes a habitation which is limited for other generalistinsect pests that are dissuaded by the biochemical compounds [28,29]. As a result, plants require newchemical admixtures to be ensured against these particular groups of insects. This procedure maybring about a proportional, well ordered “arms race” inside insect pests and its host plant, driving awide range of biochemical-barrier mixtures [30].
Molecules 2019, 24, 2067 4 of 21
3. Chemical Variety of Secondary Metabolites
Recently different ecological and evolutionary theories explain the chemical variety of secondaryplant compounds. Generally, plants are required to be able to compete for the vast range of abovegroundand belowground specialist herbivore. Consequently, they may incidentally be in a similar place;and compete with phytophagous arthropods, and other microorganisms, like virus, bacteria, as well asfungi [31–33]. As a result of insect pest’s diversity and the co-occurrence scope of bolstering plans,the plant requires mixtures of biochemicals for its defense. Thus, a wide range of biochemicals maygive such protection [34]. Additionally, a variety of biochemical compounds is required by plants forproducing strong physiological and biochemical effects to fight against different kinds of herbivores(see Table 1). Moreover, very lethal admixtures may have an adverse effect on many beneficial insectswhich are factually valuable for plants. Examples are pollinators and parasitoids [35,36]. Thus,the plant should have the capacity, to recognize phytophagous insect pests with comparable characters,as well as focus on its defense towards an exact body to maintain a strategic distance from such contraryimpact on beneficial insects. In this way, a substantial diversity of chemicals would be required witha high specificity [37]. As a result, chemical variety is strongly motivated by the development ofphytophagous insect pests. Since development is well on the way to request just a single or a coupleof phytophagous insect pests, alternate phytophagous arthropod will remain prevented by the oldbiochemical. As an outcome, a compound has a capacity to protect, and it might be helpful to extendthe biochemical range of a particular plant, in spite of substituting the old synthetic compounds with arecent chemical. Thus, the ethical force of the plant and phytophagous insect co-operations involves adistinct and dynamic arrangement of biochemical compounds.
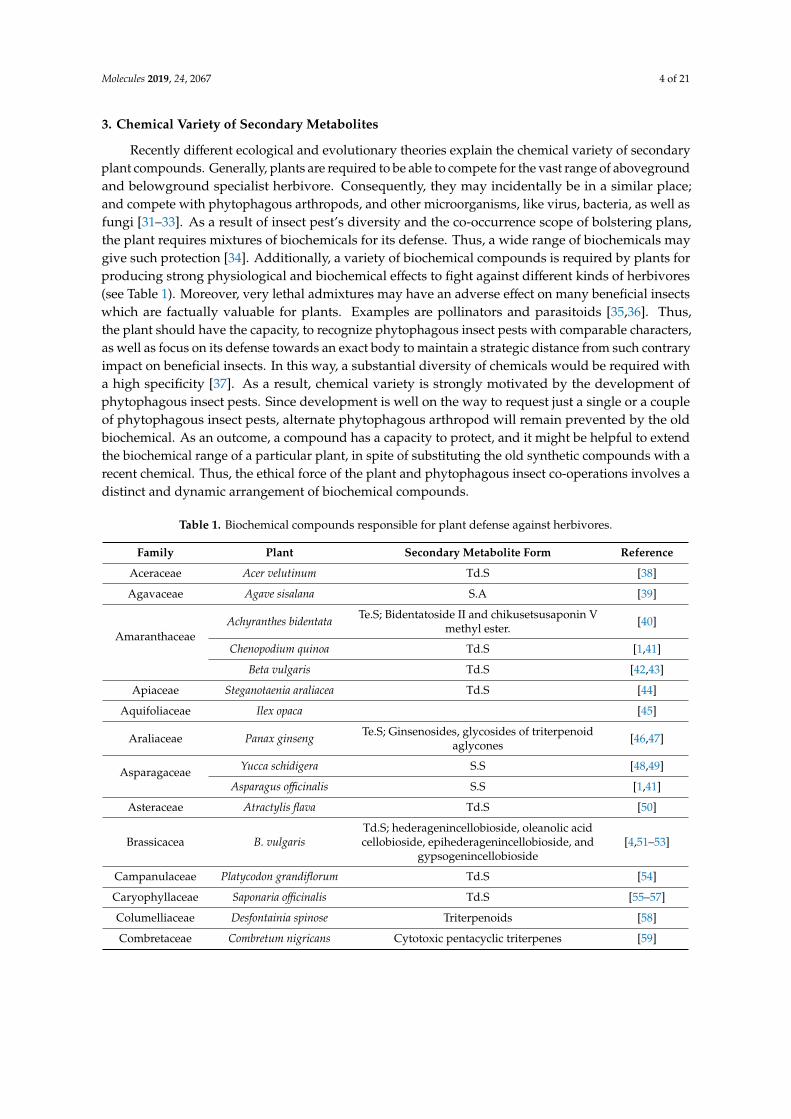
Table 1. Biochemical compounds responsible for plant defense against herbivores.
Family Plant Secondary Metabolite Form Reference
Aceraceae Acer velutinum Td.S [38]
Agavaceae Agave sisalana S.A [39]
AmaranthaceaeAchyranthes bidentata Te.S; Bidentatoside II and chikusetsusaponin V
methyl ester. [40]
Chenopodium quinoa Td.S [1,41]
Beta vulgaris Td.S [42,43]
Apiaceae Steganotaenia araliacea Td.S [44]
Aquifoliaceae Ilex opaca [45]
Araliaceae Panax ginseng Te.S; Ginsenosides, glycosides of triterpenoidaglycones [46,47]
Asparagaceae Yucca schidigera S.S [48,49]
Asparagus officinalis S.S [1,41]
Asteraceae Atractylis flava Td.S [50]
Brassicacea B. vulgarisTd.S; hederagenincellobioside, oleanolic acidcellobioside, epihederagenincellobioside, and
gypsogenincellobioside[4,51–53]
Campanulaceae Platycodon grandiflorum Td.S [54]
Caryophyllaceae Saponaria officinalis Td.S [55–57]
Columelliaceae Desfontainia spinose Triterpenoids [58]
Combretaceae Combretum nigricans Cytotoxic pentacyclic triterpenes [59]

Molecules 2019, 24, 2067 5 of 21
Table 1. Cont.
Family Plant Secondary Metabolite Form Reference
Compositae
Aster auriculatus Td.S [60]
As. tataricus Triterpene glycoside [61]
As. ageratoides Td.S [62]
As. batagensis Td.S [63–65]
As. bellidiastrum Td.S [66,67]
As. lingulatus Td.S [68,69]
As. scaber Td.S [70]
As. sedifolius Oleane-type saponins; AstersedifoliosideA, B and C [71]
As. yunnamensis Td.S [64,72,73]
CucurbitaceaeGynostemma pentaphyllum Gypenosides [74,75]
Momordica charantia Td.S [76]
Dioscoreae Dioscorea spp. Te.S, Dioscin [77,78]
FabaceaeGlycyrrhiza spp. Glycyrrhizin; Td.S [79,80]
Medicago sativa Td.S [81–83]
Desmodium adscendens Td.S [84,85]
Flacourtiaceae Aphloia madagascariensis Te.S [86]
Flacourtiaceae Aphloia theiformis Te.S [87]
Hippocastanaceae Aesculus spp. Td.S; Escins Polyhydroxyoleanene pentacyclictriterpenoid saponins; Aesculiosides [88–92]
Lamiaceae Salvia staminea Td.S, salvistamineol [93]
Lecythidaceae Petersianthus macrocarpus Td.S [94,95]
Barringtonia acutangula Monodesmosidic glucuronide saponins;Barringtosides A, B and C [96]
Liliaceae
Allium aflatunense S.S [97,98]
A. albanum S.S [99]
A. albiflorus S.S [100]
A. albopilosum S.G [101]
A. ampeloprasum S.S [102–104]
A. ascalonicum S.S [105]
A. cepa S.S; furostanol saponins, ceposide A, B, and C [106–109]
A. chinense S.S [110–112]
A. elburzense S.S [113]
A. erubescens S.S [114,115]
A. fistulosum S.S [116]
A. giganteum S.S [97,117–119]
A. jesdianum S.G [120]
A. karataviense S.S [121,122]
A. macleanii S.G [123]
A. macrostemon Furostanol glycosides [124]
A. narcissiflorum S.S [125–127]
A. nutans S.S [128,129]
A. ostrowskianum S.G [101]
A. porrum Spirostane-type saponin [130–132]
Molecules 2019, 24, 2067 6 of 21
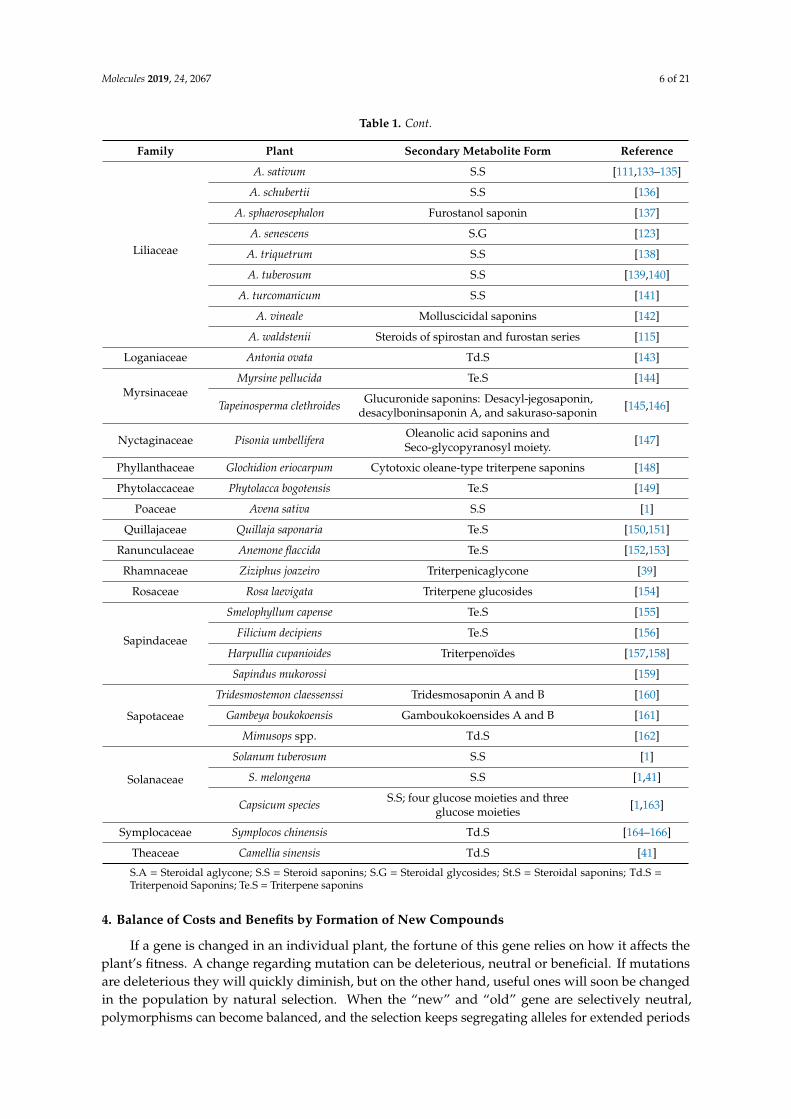
Table 1. Cont.
Family Plant Secondary Metabolite Form Reference
Liliaceae
A. sativum S.S [111,133–135]
A. schubertii S.S [136]
A. sphaerosephalon Furostanol saponin [137]
A. senescens S.G [123]
A. triquetrum S.S [138]
A. tuberosum S.S [139,140]
A. turcomanicum S.S [141]
A. vineale Molluscicidal saponins [142]
A. waldstenii Steroids of spirostan and furostan series [115]
Loganiaceae Antonia ovata Td.S [143]
MyrsinaceaeMyrsine pellucida Te.S [144]
Tapeinosperma clethroides Glucuronide saponins: Desacyl-jegosaponin,desacylboninsaponin A, and sakuraso-saponin [145,146]
Nyctaginaceae Pisonia umbellifera Oleanolic acid saponins andSeco-glycopyranosyl moiety. [147]
Phyllanthaceae Glochidion eriocarpum Cytotoxic oleane-type triterpene saponins [148]
Phytolaccaceae Phytolacca bogotensis Te.S [149]
Poaceae Avena sativa S.S [1]
Quillajaceae Quillaja saponaria Te.S [150,151]
Ranunculaceae Anemone flaccida Te.S [152,153]
Rhamnaceae Ziziphus joazeiro Triterpenicaglycone [39]
Rosaceae Rosa laevigata Triterpene glucosides [154]
Sapindaceae
Smelophyllum capense Te.S [155]
Filicium decipiens Te.S [156]
Harpullia cupanioides Triterpenoïdes [157,158]
Sapindus mukorossi [159]
Sapotaceae
Tridesmostemon claessenssi Tridesmosaponin A and B [160]
Gambeya boukokoensis Gamboukokoensides A and B [161]
Mimusops spp. Td.S [162]
Solanaceae
Solanum tuberosum S.S [1]
S. melongena S.S [1,41]
Capsicum species S.S; four glucose moieties and threeglucose moieties [1,163]
Symplocaceae Symplocos chinensis Td.S [164–166]
Theaceae Camellia sinensis Td.S [41]
S.A = Steroidal aglycone; S.S = Steroid saponins; S.G = Steroidal glycosides; St.S = Steroidal saponins; Td.S =Triterpenoid Saponins; Te.S = Triterpene saponins
4. Balance of Costs and Benefits by Formation of New Compounds
If a gene is changed in an individual plant, the fortune of this gene relies on how it affects theplant’s fitness. A change regarding mutation can be deleterious, neutral or beneficial. If mutationsare deleterious they will quickly diminish, but on the other hand, useful ones will soon be changedin the population by natural selection. When the “new” and “old” gene are selectively neutral,polymorphisms can become balanced, and the selection keeps segregating alleles for extended periods

Molecules 2019, 24, 2067 7 of 21
of time [167]. Subsequent of the aggregate adjustment of expenses and advantages in the naturalecosystem of the plant portrays the variety among various groups of population and species in amountand kind of protection. Such modification can affect the competitiveness amongst genotypes and asa result the choice for a specific genotype [168]. Beside the undeniable advantages clarified in theprevious, large amounts of protection, without enemy violence, are thought to be expensive [169,170].The defense expenses are mostly visualized regarding the distribution of minimal assets from differentvigor upgrading capacities inside a plant, for example, photosynthesis, development, as well as newgeneration [171,172]. Though, those expenses are not evident, as were assumed for plant biochemicals,especially volatiles along with a particular amount of terpenoids by Dicke and Sabelis [24], andGershenzon [173]. Some defenses may demand ecological exchanges [168], so when supplies aredistributed to protect against a particular phytophagous insect, it can decrease the vigor of the plantwhen harm triggered by other non-target phytophagous insect increments. Eventually, it is expensivewhen protection admixtures discourage advantageous living bodies, for example, crop pollinators andexpected enemies of the phytophagous insect pests [174].
A diversity of plant defense chemical compounds can act as shields in contrast to insects, involvingalkaloids, flavonoids, glucosinolates, and phenolic acids [175]. Mostly chemical compounds productionis prompted by certain biotic or abiotic factors. Such a schematic arrangement is considered as fightagainst pathogens and frame insurance economically. Various chemical admixtures involved againstinsects are the saponins, which have distinctive chemical configurations commonly containing atriterpenoid and steroidal core with a differing quality of glycosylation structures. Saponins arepresented in References [176,177] particular 100 various plant categories, even though they mostlyare general in species from distinct families or genre, for example, Leguminosae and Liliaceae [178].Saponins are acquired independent from outside signals and lead to the innate immunity, so named ashypo anticipations because they introduced in individual plants.
The positive aspect of cumulating saponins is primary protection, which is not just for hugemeasures of vitality, as well as for pathogens to develop mod additionally that it makes feasible forpathogens to develop moderation. It voided when saponin antecedents cumulate and saponinstuff raises resultantly chemical changes of precursor molecules, which incited by pathogeniccontamination [179]. Perhaps, the saponin substance may build ideally to the limited quantitydue to the chemical response of deposited precursors for biochemical compound safety system orbecause of pathogen given debasement [180,181].
In the beginning, several studies data on the specific activity of saponins against insects werelimited to leguminous origins and extracts [182]. Hostettmann and Marston [183] indicated thatseveral high saponin plant parts from various families, including Aquifoliaceae, Theaceae, as wellas Leguminosae, are resistant to insects. Recently, a lot of studies showing the structural activity ofconcentrated or pure saponin fractions against insects have widely elaborated, and have influencedinsects such as aphids, beetles, caterpillars, and flies [1,184]. Nevertheless, the relevant studiesof the consequences of various saponins from different origins against insects of different feedingdifferentiation are still limited.
The behavior of insects changes with individual components of host food, some nutrients attractthe insects, while others repel. Hence, plants can synthesize some substances that are importantfor their significant exercises, while the auxiliary metabolites are included during the time spentco-development amongst plants and other living organisms, for example, insects [185,186]. P. xylostellais a serious pest of cruciferous crops with a cosmopolitan distribution [187]. P. xylostella has developedresistance to existing chemical insecticides including the Bt toxin [188], making it increasingly difficultto control [189]. The capacity of P. xylostella to quickly create imperviousness to insecticides, joined withtypically ecological and suitability risks, have fortified enthusiasm for optional controlling systems,for example, trap crops [190]. A trap crop proposed for P. xylostella is wintercress, B. vulgaris [191–194].It is a biennial or short-lived perennial plant native to temperate regions worldwide [195]. According tothe findings of Shinoda, Nagao, Nakayama, Serizawa, Koshioka, Okabe and Kawai [16], the response

Molecules 2019, 24, 2067 8 of 21
of P. xylostella larvae is to be suspected that there is a feeding-deterrent in a crucifer-B. vulgaris. Theyrecorded an adverse effect of the plant volatile compounds on the specialist pest larvae, as the feedingrate of larvae of P. xylostella was reduced on the plant. The feeding deterrent was isolated from B. vulgarisleaves and was identified through the structure to be a monodesmosidic triterpenoid saponin.
5. Larval Feeding Preference and Adult Oviposition Behavior
Larval feeding choice and adult oviposition for younger leaves when contrasted with moreseasoned leaves of a specific accommodating plant is a general pattern common with numerousphytophagous insects, particularly in connoisseurs, encompassing P. xylostella [196]. WheneverP. xylostella adults have an option of B. vulgaris and different cruciferous crops, despite the fact P.xylostella larvae cannot continue their lives on a limited range of B. vulgaris, as such as plants beingmuch supportive for oviposition of P. xylostella adults [16,197]. This non survivorship is thought to beas a result of saponins [196].
5.1. P. xylostella Larval Survival on Cotyledons and True Leaves within the Same Plant
Cotyledons represent the capacity of food storage for the improvement of plant, which is theprimary photosynthetic network for the plant after germination [198], cotyledons of brassicaceousplants contain varying contents of glucosinolates [199,200]. In Barbarea plants, glucosinolates thatmight protect the plants against generalist herbivores, were present in the cotyledons, while saponins,which could defend the plant against specialist herbivores like P. xylostella. Similarly, some saponinswere not present in cotyledons, indicating that there might be some other biochemical compoundswhich are responsible for plant defense against herbivores.
5.2. Saponins Presentation in B. Vulgaris Var Arcuata (Isolation and Identification)
The isolation and identification of a triterpenoid saponin, from the leaves of B. vulgaris, whichstrongly deters feeding of P. xylostella larvae and also the oleane type saponin was studied byShinoda, et al. [201]. Nielsen, et al. [202]) and Augustin, et al [4] found five triterpenoid saponins inB. vulgaris namely; 3-O-cellobiosyl-hederagenin (hederagenin cellobioside), 3-O-cellobiosyl-oleanoicacid (oleanolic acid), 3-O-cellobiosyl-gypsogenin (gypsogenincellobioside), 3-O-cellobiosylcochalicacid (cochalic acid cellobioside) and 3-O-cellobiosyl-4-epihederagenin (4-epihegragenin cellobioside)Hederagenin cellobioside and oleanolic acid (Figure 1), which make B. vulgaris resistant to P. xylostellaand are correlated with deterrence of adult P. xylostella females [4,52,203]. Shinoda, et al [16] discoveredthat this is not only the first feeding deterrent to P. xylostella found in the family Brassicaceae, but also thefirst oleanane-type saponin found in this family. So, advance clarification of the chemical configurationof saponins could enhance the development of hydrophobic analogs which may be characterized asfascinating insecticides and herbicides, which potentially required for ecologically more suitable thanpresent synthetic pesticide and herbicides.
6. Biological Significance of Saponins
Saponins are biochemical compounds or otherwise depicted as natural products, which havean extensive spectrum of natural performances. Numerous biological roles have been indicated forvarious saponins, including anti-inflammatory allelopathic action, anti-carcinogenic, mitigating cellreinforcement, heamolytic, hypocholesterolemic resistance stimulators, cell layer permeabilizingcharacteristics, as well as can influence feeding behavior, development, and cause mortality,development hindrance, limit the insects’ productiveness and protection against insects and othermicro-organisms.
6.1. Saponins Interference with the Feeding Behavior
Some previous reports are available indicating the inability of insect pests larvae to attackBrassicaceae species (B. vulgaris) due to triterpene (saponin), along with two sugars at the position

Molecules 2019, 24, 2067 9 of 21
of C3, which restrain the prosecution of the food uptake [201]. Saponins also showed strong effectsagainst other pathogens like fungi mollusks certain bacteria and viruses. In general, it is believed thatsuch biochemicals operate crucially in the plant protection against biotic, as well as abiotic factors, asreported from soybean saponins, which had shown detrimental effects against Tribolium castaneum,Bufo viridis and Lebistes reticulatus. Similarly, saponins were also observed to check the cholinesterases,as well as the proteolytic drive of other enzymes, like trypsin, chymotrypsin and papain, which leadstowards non-specific communication with other protein. Moreover, some studies reported that QuillajaSaponaria saponins induce fatality in living insects, and a potent cytotoxic activity on other insects likeDrosophila melanogaster cells [204].
6.2. Saponins Effects on Protein Digestion
The toxicity of saponins to various organisms linked to their interaction with biological membranes.Some saponins form complexes with proteins [205] and by this action, they apparently inhibitproteinases and affect digestion in insect gut [204,206,207]. The capability of saponins to penetrate thecell membrane and to induce apoptosis makes saponins cytotoxic to lepidopteran cells [204].
6.3. Enterotoxicity
Saponins are a group of steroidal or triterpenoid secondary plant metabolites, with divergentbiological activities [208–210], they are responsible for plant defense against antagonists; such asmollusks, pathogens and insects [211,212]. The combination of hydrophilic sugars and hydrophobicsapogenin enable saponins to incorporate into biological membranes. Toxicity of saponins to differentorganisms seems to be related to their interaction with biological membranes and might be related totheir soap-like properties. As a result, detoxification of saponins is probably regarded as enzymatichydrolysis of the glycosidic bonds, as already produced for fungi [213,214].
Many crucifer specialist insects, such as Pieris brassicae and Pieris rapae and Pieris nemorum withR-genes, are insusceptible to the defenses of B. vulgaris. By finding out the structures of saponins inB. vulgaris [16,215] has allowed for investigations into the mechanism by which these in susceptibleinsects can deal with the potentially toxic saponins. Badenes-Perez, Reichelt, Gershenzon andHeckel [196] reported that the struggle of B. vulgaris to the diamondback moth (DBM) is prompted bytwo different saponins; I) 3-0-b-cellobiosylhederagenin and II) 3-0-cellobiosyloleanolic acid, whichprevents the feeding of P. xylostella. Likewise, it had been reported that the combination of feedingdeterrents showed feeding deterrent habituation in other insects and the combination of saponins Iand II may also slow down feeding deterrent habituation in P. xylostella. Nevertheless, saponins I andII contain similar chemical structures; cross habituation might be easier as compared to compoundswith different chemical structures, which also indicate the synthesis of saponin-II could be after that ofsaponin-I [216–219].
Idris and Grafius [220] and Badenes-Pérez, et al. [221] showed that a small percentage of larvae ofa P. xylostella population collected from the field were able to survive on B. vulgaris, even though theydid not report the concentration of saponins in these plants. Further researches are required to verifyin any case being feeding deterrents, saponins I and II, might have a toxic effect on P. xylostella larvae.Badenes-Perez, Reichelt, Gershenzon and Heckel [196] observed that continuous feeding of neonatesof P. xylostella usually on resistant B. vulgaris, results in feeding signs [192].
Dissimilarly to glucosinolates, saponins I and II do not have all the earmarks of being expressedon the leaf covering of Barbarea [222]. Therefore, it is probably that neonates of P. xylostella encounterglucosinolates on the leaf surface and start feeding, while feeding is reduced when insects come intocontact with the saponins in the leaf tissue. Likewise saponins I and II, other saponins have beensegregated from P-type B. vulgaris var. arcuata, which are responsible for the resistance of this plantto P. nemorum [53,223]. Given the similarity in the resistance mechanisms of G-type B. vulgaris var.arcuata to both P. nemorum and P. xylostella, these saponins might be required in the resistance ofBarbarea to P. xylostella. Saponins display higher toxicity, even though the precise mode of action of

Molecules 2019, 24, 2067 10 of 21
saponins remains unresolved, it was reported by Badenes-Perez, et al. [196] that saponins specificallytarget pest insects: Both the continuous insect cells and the primary midgut cells of Spodoptera littoralisshowed high sensitivity to Q. saponaria saponin. The phenomenon behind such synergistic mechanismsare unknown, but may include the ability of one biochemical to inhibit the detoxification of othercomponents or to up regulate the absorption of others from the gut.
More significantly, the saponins can cause great and quick in vivo enterotoxin results on the larvaeof S. littoralis, and with contents likewise those that can be presented in nature. Therefore, saponinsshowed substantial evidence for the potency in the control of pest insects, especially insect midgutepithelium as the primary target tissue. So, the insect midgut is an attractive target, as any damagingeffect on the midgut epithelial cells will result in starvation, leading towards slow insect mortality.As this component is not the same in midgut cells as the approach of Bacillus thuringiensis (Bt), it canlikewise be of assistance in the management of imperviousness to Bt. Furthermore, as aphids are notperceptive to the poisons of Bt, all observations propose that saponins may represent a noteworthyoutcome in developing new, substitute, environmentally favorable aphid control agents amongstintegrated pest management.
7. Limits of the Use of Saponins in Pest Management Control
Some saponins have heamolytic and cytotoxic effects which have the potential of inhibiting theprotease activity. Due to this constraint, it is difficult to apply in the field, as they might also be toxicto humans. The saponins function to protect host plant and to discourage phytophagous insectsusually is explained according to their performance in the body of the exposed organism, such asless food consumption, obstructions as well other poisons [224,225]. Mostly, saponins are known asdisincentives against insect pests, but their mode of action is yet relatively obscure, however it isidentified to interrupt cell sheets [213,226].
Moreover, it was assumed with respect to insects that insect resistance, on the base of ecdysteroidreceptor complex (EcR), may be due to particular steroidal saponins, which have resemblance with20-hydroxyecdysone (molting hormone) [227,228]. Even though, the saponins performance was notsupported by real resistance reaction to EcR communication, yet rather than loss of cellular unityconsiderably, due to the pervasion of the insect cell layer, as described by De Geyter, Swevers, Caccia,Geelen and Smagghe [204]. Plant-derived triterpenoid and steroidal saponins are very promising forthe development of botanical insecticides. Aside from cellular poisoning quality, saponins additionallyexhibited hindrance or anti-feedant drive against herbivores, especially insects. In a previous study,it was reported that saponins (aginosid) extracted from leek (Allium porrum) caused a noteworthyobstruction in response to two Lepidopteran insect pests; Peridroma saucia and Mamestra configurata [229],as well as in sucking insect pests [230]. In another related study, extracts from the roots of Saponariaofficinalis induced a reduction in the rate of oviposition by females of Tetranychus urticae [231]. The plantextracts were found to contain a mix of various saponins [232–234] which are suggested to be responsiblefor the acaricidal efficacy. However, the mechanism of action on mites needs to be further explored.
8. Conclusions and Recommendations
Glucosinolates and saponins play an important role in the plant defense against specialistherbivores. Preliminary data on the saponins’ performance was constrained to reports of leguminousreserves as well other by-products [182]. Thus, comparative studies on the role of saponins are stilllimited. The chemical basis of previously reported flea beetle resistance in the G-type of B. vulgarisvar. arcuata is unknown, but resistance is not correlated to glucosinolates or glucosinolate levels [235].Resistance may be due to the occurrence of a triterpenoid saponin, which made resistant to B. vulgarisagainst DBM [16]. Development in the interpretation of saponins biosynthetic system has beenobstructed due to a distinctive molecular configuration along with the complication of enzymes, relatedto two major superfamilies, such as I) cytochrome (P450) and II) glycosyltransferase (GT). The greaterpart of the Allium and Calamus species consist of saponins, which have a crucial role in health; as such

Molecules 2019, 24, 2067 11 of 21
saponins are responsible to decrease the level of garlic cholesterol, as well as enhance the anti-fungalfunction of garlic [236,237]. They display higher toxicity, even though the precise mode of actionof saponins remains unresolved. It has been reported by Badenes-Perez, et al. [196] that saponinsfrom Q. saponaria approach S. littoralis directly by affecting consistent insect midgut cells. It might beexemplified significant results in developing new, substitute, environmentally favorable control agentsamongst integrated pest management.
As the discovery of plant defense chemicals continues at its present rapid pace, the present studiesdiscussed above represent the role of plant secondary metabolites in plant defense against herbivores.Given the complex chemical structures of plants, which are not easy to fully understand, play the actualrole in defense mechanism (adaptations or counteradaptations) in plant–herbivore interactions. Thus,the structure activity studies of saponins as deterrents for specialist herbivore (such as P. xylostella),therefore, are useful for the deeper understanding of the components and the systems concerned withinsect resistance. However, a targeted isolation of these insect repellants will elucidate their structures.Therefore, the improvement of hydrophobic analogs might be regulated by a particular chemicalstructure of saponins, which may be characterized as interesting chemical sprays, for a particularrange of plants, and are (potentially) more natural, compared to the present synthetic herbicide usedagainst herbivores.
Author Contributions: M.H. conceived, designed and written the manuscript. M.Q. revised and helped in figurepreparation. B.D., W.I., M.S.H., and B.S.B. helped in the collection of information and writing the manuscript. L.W.and D.Q. conceived, revised and finalized the manuscripts. All authors approved the submission version.
Funding: This study was supported by National Natural Science Grant of China (Award no. 30400061), NaturalScience Foundation of Fujian Province, China (2011J01082) and Special Fund for Science and Technology Innovationof FAFU (CXZX2016107). And National Key Project of R&D of China (2018YFD0201500), Key Projects of Scienceand Technology of Fujian Province (2016N0005), and Research Fund for the International Collaborative Program(KXGH17004) with (CXZX2017211) from Fujian Agriculture and Forestry University (FAFU).
Acknowledgments: We are grateful to the researchers who have contributed to this field.
Conflicts of Interest: The authors declare no conflict of interest.
References
1. De Geyter, E.; Lambert, E.; Geelen, D.; Smagghe, G. Novel advances with plant saponins as natural insecticidesto control pest insects. Pest Technol. 2007, 1, 96–105.
2. Mugford, S.T.; Osbourn, A. Saponin synthesis and function. In Isoprenoid Synthesis in Plants and Microorganisms;Springer: New York, NY, USA, 2012; pp. 405–424.
3. Moses, T.; Papadopoulou, K.K.; Osbourn, A. Metabolic and functional diversity of saponins, biosyntheticintermediates and semi-synthetic derivatives. Crit. Rev. Biochem. Mol. Biol. 2014, 49, 439–462. [PubMed]
4. Augustin, J.M.; Drok, S.; Shinoda, T.; Sanmiya, K.; Nielsen, J.K.; Khakimov, B.; Olsen, C.E.; Hansen, E.H.;Kuzina, V.; Ekstrøm, C.T. UDP-glycosyltransferases from the UGT73C subfamily in Barbarea vulgaris catalyzesapogenin 3-O-glucosylation in saponin-mediated insect resistance. Plant Physiol. 2012, 160, 1881–1895.
5. Khakimov, B.; Kuzina, V.; Erthmann, P.Ø.; Fukushima, E.O.; Augustin, J.M.; Olsen, C.E.; Scholtalbers, J.;Volpin, H.; Andersen, S.B.; Hauser, T.P. Identification and genome organization of saponin pathway genesfrom a wild crucifer, and their use for transient production of saponins in Nicotiana benthamiana. Plant J. 2015,84, 478–490. [PubMed]
6. Patamalai, B.; Hejtmancik, E.; Bridges, C.; Hill, D.; Camp, B. The isolation and identification of steroidalsapogenins in Kleingrass. Vet. Hum. Toxicol. 1990, 32, 314–318. [PubMed]
7. Lee, S.T.; Stegelmeier, B.L.; Gardner, D.R.; Vogel, K.P. The isolation and identification of steroidal sapogeninsin switchgrass. J. Nat. Toxins 2001, 10, 273–281.
8. Osbourn, A.E. Saponins in cereals. Phytochemistry 2003, 62, 1–4. [CrossRef]9. Thimmappa, R.; Geisler, K.; Louveau, T.; O’Maille, P.; Osbourn, A. Triterpene biosynthesis in plants.
Annu. Rev. Plant Biol. 2014, 65, 225–257. [CrossRef] [PubMed]10. Ghosh, S. Biosynthesis of Structurally Diverse Triterpenes in Plants: The Role of Oxidosqualene Cyclases.
Proc. Indian Natl. Sci. Acad. 2016, 82, 1189–1210. [CrossRef]

Molecules 2019, 24, 2067 12 of 21
11. Nielsen, J.K.; Nagao, T.; Okabe, H.; Shinoda, T. Resistance in the plant, Barbarea vulgaris, and counter-adaptations in flea beetles mediated by saponins. J. Chem. Ecol. 2010, 36, 277–285. [CrossRef]
12. Badenes-Perez, F.R.; Gershenzon, J.; Heckel, D.G. Insect attraction versus plant defense: Young leaves highin glucosinolates stimulate oviposition by a specialist herbivore despite poor larval survival due to highsaponin content. Plos One 2014, 9, e95766.
13. Talekar, N.; Shelton, A. Biology, ecology, and management of the diamondback moth. Annu. Rev. Entomol.1993, 38, 275–301. [CrossRef]
14. Li, Z.; Feng, X.; Liu, S.-S.; You, M.; Furlong, M.J. Biology, Ecology, and Management of the DiamondbackMoth in China. Annu. Rev. Entomol. 2016, 61, 277–296.
15. Huang, X.; Renwick, J.; Sachdev-Gupta, K. Oviposition stimulants in Barbarea vulgaris for Pieris rapae andP. napi oleracea: Isolation, identification and differential activity. J. Chem. Ecol. 1994, 20, 423–438.
16. Shinoda, T.; Nagao, T.; Nakayama, M.; Serizawa, H.; Koshioka, M.; Okabe, H.; Kawai, A. Identification ofa triterpenoid saponin from a crucifer, Barbarea vulgaris, as a feeding deterrent to the diamondback moth,Plutella xylostella. J. Chem. Ecol. 2002, 28, 587–599.
17. Serizawa, H.; Shinoda, T.; Kawai, A. Occurrence of a feeding deterrent in Barbarea vulgaris (Brassicales:Brassicaceae), a crucifer unacceptable to the diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae).Appl. Entomol. Zool. 2001, 36, 465–470.
18. Newman, K. Feeding and oviposition preferences of the diamondback moth Plutella xylostella (Lepidoptera:Plutellidae) on six Brassicaceae host plant species. M.Sc Thesis 2014.
19. Hopkins, R.J.; van Dam, N.M.; van Loon, J.J. Role of glucosinolates in insect-plant relationships andmultitrophic interactions. Annu. Rev. Entomol. 2009, 54, 57–83. [CrossRef]
20. Ratzka, A.; Vogel, H.; Kliebenstein, D.J.; Mitchell-Olds, T.; Kroymann, J. Disarming the mustard oil bomb.Proc. Natl. Acad. Sci. USA 2002, 99, 11223–11228.
21. Renwick, J.A.A.; Haribal, M.; Gouinguené, S.; Städler, E. Isothiocyanates stimulating oviposition by thediamondback moth, Plutella xylostella. J. Chem. Ecol. 2006, 32, 755–766.
22. Hairston, N.G.; Smith, F.E.; Slobodkin, L.B. Community structure, population control, and competition.Am. Nat. 1960, 94, 421–425. [CrossRef]
23. Ehrlich, P.R.; Raven, P.H. Butterflies and plants: A study in coevolution. Evolution 1964, 18, 586–608.[CrossRef]
24. Dicke, M.; Sabelis, M.W. Does it pay plants to advertise for bodyguards? Towards a cost-benefit analysisof induced synomone production. In Causes and Consequences of Variation in Growth Rate and Productivity ofHigher Plants; SPB Academic Publishing: The Hague, The Netherlands, 1989; pp. 341–358.
25. Renwick, J.A.A. The chemical world of crucivores: Lures, treats and traps. Entomol. Exp. Et Appl. 2002, 104,35–42. [CrossRef]
26. Jones, C.G.; Firn, R.D.; Malcolm, S.B. On the evolution of plant secondary chemical diversity [and discussion].Philos. Trans. R. Soc. Lond. B Biol. Sci. 1991, 333, 273–280.
27. Stahl, E. Pflanzen und Schnecken: Eine Biologische Studie über die Schutzmittel der Pflanzen gegen Schneckenfrass;G. Fischer: Germany, 1888; Volume 1.
28. Cornell, H.V.; Hawkins, B.A. Herbivore responses to plant secondary compounds: A test of phytochemicalcoevolution theory. Am. Nat. 2003, 161, 507–522. [CrossRef] [PubMed]
29. Van der Putten, W.H. Plant defense belowground and spatiotemporal processes in natural vegetation. Ecology2003, 84, 2269–2280. [CrossRef]
30. Iwao, K.; Rausher, M.D. Evolution of plant resistance to multiple herbivores: Quantifying diffuse coevolution.Am. Nat. 1997, 149, 316–335. [CrossRef]
31. Van der Putten, W.H.; Vet, L.E.M.; Harvey, J.A.; Wäckers, F.L. Linking above-and belowground multitrophicinteractions of plants, herbivores, pathogens, and their antagonists. Trends Ecol. Evol. 2001, 16, 547–554.[CrossRef]
32. Van Dam, N.M.; Harvey, J.A.; Wäckers, F.L.; Bezemer, T.M.; van der Putten, W.H.; Vet, L.E. Interactionsbetween aboveground and belowground induced responses against phytophages. Basic Appl. Ecol. 2003, 4,63–77. [CrossRef]
33. Bezemer, T.M.; van Dam, N.M. Linking aboveground and belowground interactions via induced plantdefenses. Trends Ecol. Evol. 2005, 20, 617–624. [CrossRef]

Molecules 2019, 24, 2067 13 of 21
34. Rask, L.; Andréasson, E.; Ekbom, B.; Eriksson, S.; Pontoppidan, B.; Meijer, J. Myrosinase: Gene familyevolution and herbivore defense in Brassicaceae. In Plant Molecular Evolution; Springer: The Netherlands,2000; pp. 93–113.
35. Poveda, K.; Steffan-Dewenter, I.; Scheu, S.; Tscharntke, T. Effects of below-and above-ground herbivores onplant growth, flower visitation and seed set. Oecologia 2003, 135, 601–605. [CrossRef]
36. Soler, R.; Harvey, J.A.; Kamp, A.F.D.; Vet, L.E.M.; Van der Putten, W.H.; Van Dam, N.M.; Stuefer, J.F.; Gols, R.;Hordijk, C.A.; Martijn Bezemer, T. Root herbivores influence the behaviour of an aboveground parasitoidthrough changes in plant-volatile signals. Oikos 2007, 116, 367–376. [CrossRef]
37. Fritz, R.S.; Simms, E.L. Plant Resistance to Herbivores and Pathogens: Ecology, Evolution, and Genetics; Universityof Chicago Press: Chicago, IL, USA, 1992.
38. Glénsk, M.; Włodarczyk, M.; Bassarello, C.; Pizza, C.; Stefanowicz, P.; Switalska, M. A Major Saponin fromLeaves Extract of Acer velutinum. Z. Für Nat. B 2009, 64, 1081–1086. [CrossRef]
39. Ribeiro, B.D.; Alviano, D.S.; Barreto, D.W.; Coelho, M.A.Z. Functional properties of saponins from sisal(Agave sisalana) and juá (Ziziphus joazeiro): Critical micellar concentration, antioxidant and antimicrobialactivities. Colloids Surf. A Physicochem. Eng. Asp. 2013, 436, 736–743. [CrossRef]
40. Mitaine-Offer, A.-C.; Marouf, A.; Hanquet, B.; Birlirakis, N.; Lacaille-Dubois, M.-A. Two Triterpene Saponinsfrom Achyranthes bidentata. Chem. Pharm. Bull. 2001, 49, 1492–1494. [CrossRef]
41. Francis, G.; Kerem, Z.; Makkar, H.P.; Becker, K. The biological action of saponins in animal systems: A review.Br. J. Nutr. 2002, 88, 587–605. [CrossRef] [PubMed]
42. Ridout, C.L.; Price, K.R.; Parkin, G.; Dijoux, M.G.; Lavaud, C. Saponins from sugar beet and the floc problem.J. Agric. Food Chem. 1994, 42, 279–282. [CrossRef]
43. Massiot, G.; Dijoux, M.-G.; Lavaud, C.; Le Men-Olivier, L.; Connolly, J.D.; Sheeley, D.M. Seco-glycosides ofoleanolic acid fromBeta vulgaris. Phytochemistry 1994, 37, 1667–1670. [CrossRef]
44. Lavaud, C.; Massiot, G.; Le Men-Olivier, L.; Viari, A.; Vigny, P.; Delaude, C. Saponins from Steganotaeniaaraliacea. Phytochemistry 1992, 31, 3177–3181. [CrossRef]
45. Potter, D.A.; Kimmerer, T.W. Inhibition of herbivory on young holly leaves: Evidence for the defensive roleof saponins. Oecologia 1989, 78, 322–329. [CrossRef]
46. Shin, B.-K.; Kwon, S.W.; Park, J.H. Chemical diversity of ginseng saponins from Panax ginseng. J. GinsengRes. 2015, 39, 287–298. [CrossRef] [PubMed]
47. Luo, H.; Sun, C.; Sun, Y.; Wu, Q.; Li, Y.; Song, J.; Niu, Y.; Cheng, X.; Xu, H.; Li, C.; et al. Analysis of thetranscriptome of Panax notoginseng root uncovers putative triterpene saponin-biosynthetic genes andgenetic markers. Bmc Genom. 2011, 12, S5. [CrossRef] [PubMed]
48. Miyakoshi, M.; Tamura, Y.; Masuda, H.; Mizutani, K.; Tanaka, O.; Ikeda, T.; Ohtani, K.; Kasai, R.; Yamasaki, K.Antiyeast steroidal saponins from Yucca schidigera (Mohave Yucca), a new anti-food-deteriorating agent.J. Nat. Prod. 2000, 63, 332–338. [CrossRef] [PubMed]
49. Piacente, S.; Pizza, C.; Oleszek, W. Saponins and phenolics of Yucca schidigera Roezl: Chemistry andbioactivity. Phytochem. Rev. 2005, 4, 177–190. [CrossRef]
50. Chabani, S.; Lavaud, C.; Benkhaled, M.; Harakat, D.; Long, C.; Haba, H. Three new oleanane-type triterpenesaponins from Atractylis flava. Phytochem. Lett. 2016, 15, 88–93. [CrossRef]
51. Khakimov, B.; Amigo, J.M.; Bak, S.; Engelsen, S.B. Plant metabolomics: Resolution and quantificationof elusive peaks in liquid chromatography–mass spectrometry profiles of complex plant extracts usingmulti-way decomposition methods. J. Chromatogr. A 2012, 1266, 84–94. [CrossRef] [PubMed]
52. Kuzina, V.; Ekstrøm, C.T.; Andersen, S.B.; Nielsen, J.K.; Olsen, C.E.; Bak, S. Identification of defensecompounds in Barbarea vulgaris against the herbivore Phyllotreta nemorum by an ecometabolomic approach.Plant Physiol. 2009, 151, 1977–1990. [CrossRef] [PubMed]
53. Kuzina, V.; Nielsen, J.K.; Augustin, J.M.; Torp, A.M.; Bak, S.; Andersen, S.B. Barbarea vulgaris linkagemap and quantitative trait loci for saponins, glucosinolates, hairiness and resistance to the herbivorePhyllotreta nemorum. Phytochemistry 2011, 72, 188–198. [CrossRef]
54. Ma, C.-H.; Gao, Z.-J.; Zhang, J.-J.; Zhang, W.; Shao, J.-H.; Hai, M.-R.; Chen, J.-W.; Yang, S.-C.; Zhang, G.-H.Candidate Genes Involved in the Biosynthesis of Triterpenoid Saponins in Platycodon grandiflorum Identifiedby Transcriptome Analysis. Front. Plant Sci. 2016, 7. [CrossRef] [PubMed]
55. Hostettmann, K.; Marston, A. Saponins; Cambridge University Press: New York, NY, USA, 2005.

Molecules 2019, 24, 2067 14 of 21
56. Koike, K.; Jia, Z.; Nikaido, T. New triterpenoid saponins and sapogenins from Saponaria officinalis.J. Nat. Prod. 1999, 62, 1655–1659. [CrossRef]
57. Jia, Z.; Koike, K.; Sahu, N.P.; Nikaido, T. Triterpenoid saponins from Caryophyllaceae family. In Studies inNatural Products Chemistry; Atta ur, R., Ed.; Elsevier: Athens, GA, USA, 2002; Volume 26, Part G; pp. 3–61.
58. Houghton, P.J.; Lian, L.M. Triterpenoids from Desfontainia spinosa. Phytochemistry 1986, 25, 1939–1944.[CrossRef]
59. Simon, G.; Dewelle, J.; Nacoulma, O.; Guissou, P.; Kiss, R.; Daloze, D.; Braekman, J.C. Cytotoxic pentacyclictriterpenes from Combretum nigricans. Fitoterapia 2003, 74, 339–344. [CrossRef]
60. Wang, C.Z.; Yu, D.Q. Triterpenoid Saponins from Aster auriculatus. J. Asian Nat. Prod. Res. 1998, 1, 1–14.[CrossRef]
61. Dongliang, C.; Yu, S. Terpenoid glycosides from the roots of Aster tataricus. Phytochemistry 1993, 35, 173–176.[CrossRef]
62. Sakai, K.; Nagao, T.; Okabe, H. Triterpenoid saponins from the ground part of Aster ageratoides var. ovatus.Phytochemistry 1999, 51, 309–318. [CrossRef]
63. Shao, Y.; Zhou, B.-N.; Lin, L.-Z.; Cordell, G.A. Two new triterpenoid saponins, asterbatanoside F and G, fromAster batangensis. Nat. Prod. Lett. 1994, 5, 233–240. [CrossRef]
64. Shao, Y.; Zhou, B.-N.; Lin, L.-Z.; Cordell, G.A. Triterpene saponins from Aster yunnanensis. Phytochemistry1995, 38, 1487–1492. [CrossRef]
65. Shao, Y.; Zhou, B.; Ma, K.; Wu, H.; Lin, L.; Cordell, G.A. Medicagenic acid saponins from Aster batangensis.Phytochemistry 1995, 39, 875–881. [CrossRef]
66. Schöpke, T.; Al-Tawaha, C.; Wray, V.; Nimtz, M.; Meyer, A.; Hiller, K. Triterpenoid saponins from the aerialparts of Aster bellidiastrum. Phytochemistry 1995, 40, 1489–1492. [CrossRef]
67. Schöpke, T.; Al-Tawaha, C.; Wray, V.; Nimtz, M.; Hiller, K. Triterpenoid saponins from Aster bellidiastrum.Phytochemistry 1997, 45, 125–132. [CrossRef]
68. Shao, Y.; Ho, C.-T.; Chin, C.-K.; Poobrasert, O.; Yang, S.-W.; Cordell, G.A. Asterlingulatosides C and D,cytotoxic triterpenoid saponins from Aster lingulatus. J. Nat. Prod. 1997, 60, 743–746. [CrossRef] [PubMed]
69. Shao, Y.; Ho, C.-T.; Chin, C.-K.; Rosen, R.T.; Hu, B.; Qin, G.-W. Triterpenoid saponins from Aster lingulatus.Phytochemistry 1997, 44, 337–340. [CrossRef]
70. Nagao, T.; Tanaka, R.; Okabe, H. Saponins from the compositae plants: Structures of the saponins fromAster scaber Thunb. In Saponins Used in Traditional and Modern Medicine; Springer: Boston, MA, USA, 1996;pp. 297–307.
71. Corea, G.; Iorizzi, M.; Lanzotti, V.; Cammareri, M.; Conicella, C.; Laezza, C.; Bifulco, M. AstersedifoliosideA–C, three new oleane-type saponins with antiproliferative activity. Bioorganic Med. Chem. 2004, 12,4909–4915. [CrossRef] [PubMed]
72. Shao, Y.; Zhou, B.-N.; Lin, L.-Z.; Cordell, G.A. Asteryunnanosides F and G: Two new triterpenoid saponinsfrom Aster yunnanensis. Planta Med. 1995, 61, 446–449. [CrossRef]
73. Shao, Y.; Zhou, B.-N.; Ma, K.; Wu, H.-M. Echinocystic acid saponins from Aster yunnanensis. J. Nat. Prod.1995, 58, 837–842. [CrossRef]
74. Shi, L.; Cao, J.-Q.; Shi, S.-M.; Zhao, Y.-Q. Triterpenoid saponins from Gynostemma pentaphyllum. J. AsianNat. Prod. Res. 2011, 13, 168–177. [CrossRef]
75. Yang, F.; Shi, H.; Zhang, X.; Yang, H.; Zhou, Q.; Yu, L.L. Two new saponins from tetraploid jiaogulan(Gynostemma pentaphyllum), and their anti-inflammatory and α-glucosidase inhibitory activities. Food Chem.2013, 141, 3606–3613. [CrossRef]
76. Keller, A.C.; Ma, J.; Kavalier, A.; He, K.; Brillantes, A.-M.B.; Kennelly, E.J. Saponins from the traditionalmedicinal plant Momordica charantia stimulate insulin secretion in vitro. Phytomedicine 2011, 19, 32–37.[CrossRef]
77. Waller, G.R.; Yamasaki, K. Saponins Used in Traditional and Modern Medicine; Springer Science & BusinessMedia: Chicago, IL, USA, 2013; Volume 404.
78. Qin, Y.; Wu, X.; Huang, W.; Gong, G.; Li, D.; He, Y.; Zhao, Y. Acute toxicity and sub-chronic toxicity ofsteroidal saponins from Dioscorea zingiberensis CH Wright in rodents. J. Ethnopharmacol. 2009, 126, 543–550.[CrossRef]

Molecules 2019, 24, 2067 15 of 21
79. Xu, R.; Zhao, W.; Xu, J.; Shao, B.; Qin, G. Studies on Bioactive Saponins from Chinese Medicinal Plants.In Saponins Used in Traditional and Modern Medicine; Waller, G.R., Yamasaki, K., Eds.; Springer US: Boston,MA, USA, 1996; pp. 371–382.
80. Augustin, J.M.; Kuzina, V.; Andersen, S.B.; Bak, S. Molecular activities, biosynthesis and evolution oftriterpenoid saponins. Phytochemistry 2011, 72, 435–457. [CrossRef]
81. Massiot, G.; Lavaud, C.; Guillaume, D.; Le Men-Olivier, L. Reinvestigation of the sapogenins andprosapogenins from alfalfa (Medicago sativa). J. Agric. Food Chem. 1988, 36, 902–909. [CrossRef]
82. Massiot, G.; Lavaud, C.; Besson, V.; Le Men-Olivier, L.; Van Binst, G. Saponins from aerial parts of alfalfa(Medicago sativa). J. Agric. Food Chem. 1991, 39, 78–82. [CrossRef]
83. Oleszek, W.; Price, K.R.; Colquhoun, I.J.; Jurzysta, M.; Ploszynski, M.; Fenwick, G.R. Isolation andidentification of alfalfa (Medicago sativa L.) root saponins: Their activity in relation to a fungal bioassay.J. Agric. Food Chem. 1990, 38, 1810–1817. [CrossRef]
84. Rastogi, S.; Pandey, M.M.; Rawat, A.K.S. An ethnomedicinal, phytochemical and pharmacological profileof Desmodium gangeticum (L.) DC. and Desmodium adscendens (Sw.) DC. J. Ethnopharmacol. 2011, 136,283–296. [CrossRef] [PubMed]
85. McManus, O.; Harris, G.; Giangiacomo, K.; Feigenbaum, P.; Reuben, J.; Addy, M.; Burka, J.F.; Kaczorowski, G.;Garcia, M. An activator of calcium-dependent potassium channels isolated from a medicinal herb. Biochemistry1993, 32, 6128–6133. [CrossRef] [PubMed]
86. Dijoux, M.-G.; Lavaud, C.; Massiot, G.; Le Men-Olivier, L.; Sheeley, D.M. A saponin from leaves of Aphloiamadagascariensis. Phytochemistry 1993, 34, 497–499. [CrossRef]
87. Gopalsamy, N.; Vargas, D.; Guého, J.; Ricaud, C.; Hostettmann, K. Saponins from leaves of Aphloia theiformis.Phytochemistry 1988, 27, 3593–3595. [CrossRef]
88. Kimura, H.; Ogawa, S.; Jisaka, M.; Kimura, Y.; Katsube, T.; Yokota, K. Identification of novel saponins fromedible seeds of Japanese horse chestnut (Aesculus turbinata Blume) after treatment with wooden ashes andtheir nutraceutical activity. J. Pharm. Biomed. Anal. 2006, 41, 1657–1665. [CrossRef]
89. Kimura, H.; Ogawa, S.; Katsube, T.; Jisaka, M.; Yokota, K. Antiobese effects of novel saponins from edibleseeds of Japanese horse chestnut (Aesculus turbinata Blume) after treatment with wood ashes. J. Agric.Food Chem. 2008, 56, 4783–4788. [CrossRef]
90. Zhang, Z.; Li, S. Cytotoxic triterpenoid saponins from the fruits of Aesculus pavia L. Phytochemistry 2007, 68,2075–2086. [CrossRef]
91. Zhang, Z.; Li, S.; Zhang, S.; Gorenstein, D. Triterpenoid saponins from the fruits of Aesculus pavia.Phytochemistry 2006, 67, 784–794. [CrossRef] [PubMed]
92. Yoshikawa, M.; Murakami, T.; Matsuda, H.; Yamahara, J.; Murakami, N.; Kitagawa, I. Bioactive saponins andglycosides. III. Horse chestnut.(1): The structures, inhibitory effects on ethanol absorption, and hypoglycemicactivity of escins Ia, Ib, IIa, IIb, and IIIa from the seeds of Aesculus hippocastanum L. Chem. Pharm. Bull. 1996,44, 1454–1464. [CrossRef]
93. Topcu, G.; Altiner, E.N.; Gozcu, S.; Halfon, B.; Aydogmus, Z.; Pezzuto, J.; Zhou, B.-N.; Kingston, D.G. Studieson di-and triterpenoids from Salvia staminea with cytotoxic activity. Planta Med. 2003, 69, 464–467.
94. Massiot, G.; Xiang-Fei, C.; Lavaud, C.; Le Men-Olivier, L.; Delaude, C.; Viari, A.; Vigny, P.; Duval, J. Saponinsfrom stem bark of Petersianthus macrocarpus. Phytochemistry 1992, 31, 3571–3576. [CrossRef]
95. El Izzi, A.; Benie, T.; Thieulant, M.-L.; Le Men-Olivier, L.; Duval, J. Stimulation of LH release from culturedpituitary cells by saponins of Petersianthus macrocarpus: A permeabilizing effect. Planta Med. 1992, 58, 229–233.[CrossRef]
96. Pal, B.C.; Chaudhuri, T.; Yoshikawa, K.; Arihara, S. Saponins from Barringtonia acutangula. Phytochemistry1994, 35, 1315–1318. [CrossRef]
97. Kawashima, K.; Mimaki, Y.; Sashida, Y. Steroidal saponins from Allium giganteum and A. aflatunense.Phytochemistry 1991, 30, 3063–3067. [CrossRef]
98. Mimaki, Y.; Kuroda, M.; Sashida, Y. Steroidal saponins from the bulbs of Allium aflatunense. Natural Med.1999, 53, 88–93.
99. Ismailov, A.; Tagiev, S.; Rasulov, E. Steroid saponins and sapogenins from Allium rubellum and Allium albanum.Khimiia Prir. Soedin. Chem. Natural Compound. 1976, 12, 495. [CrossRef]
100. Ismaılov, A.; Aliev, A. Determination of the steroid saponins in the onion (Allium albiflorus) that grows inAzerbaijan. Farmatsiia 1976, 25, 17.

Molecules 2019, 24, 2067 16 of 21
101. Mimaki, Y.; Kawashima, K.; Kanmoto, T.; Sashida, Y. Steroidal glycosides from Allium albopilosum and A.ostrowskianum. Phytochemistry 1993, 34, 799–805. [CrossRef]
102. MoRITA, T.; USHIROGUCHI, T.; HAYASHI, N.; MATSUURA, H.; ITAKURA, Y.; Fuwa, T. Steroidal saponinsfrom elephant garlic, bulbs of Allium ampeloprasum L. Chem. Pharm. Bull. 1988, 36, 3480–3486. [CrossRef]
103. Matsunaga, S.; Fusetani, N.; Nishikawa, H.; Takamura, S.; Saito, T. New antifungal and cytotoxic steroidalsaponins from the bulbs of an elephant garlic mutant. Biosci. Biotechnol. Biochem. 1998, 62, 1904–1911.
104. Mimaki, Y.; Kuroda, M.; Sashida, Y. Steroidal saponins from the bulbs of Allium ampeloprasum. Natural Med.1999, 53, 134–137.
105. Fattorusso, E.; Iorizzi, M.; Lanzotti, V.; Taglialatela-Scafati, O. Chemical composition of shallot(Allium ascalonicum Hort.). J. Agric. Food Chem. 2002, 50, 5686–5690. [CrossRef]
106. Corea, G.; Fattorusso, E.; Lanzotti, V.; Capasso, R.; Izzo, A.A. Antispasmodic saponins from bulbs of redonion, Allium cepa L. var. Tropea. J. Agric. Food Chem. 2005, 53, 935–940. [CrossRef]
107. Dini, I.; Tenore, G.C.; Trimarco, E.; Dini, A. Furostanol saponins in Allium caepa L. Var. tropeana seeds.Food Chem. 2005, 93, 205–214. [CrossRef]
108. Li, C.-J.; Yuan, L.; Ji, T.-F.; Yang, J.-B.; Wang, A.-G.; Su, Y.-L. Furostanol saponins from the seeds of Allium cepaL. Fitoterapia 2014, 99, 56–63. [CrossRef]
109. Lanzotti, V.; Romano, A.; Lanzuise, S.; Bonanomi, G.; Scala, F. Antifungal saponins from bulbs of whiteonion, Allium cepa L. Phytochemistry 2012, 74, 133–139. [CrossRef]
110. Kuroda, M.; Mimaki, Y.; Kameyama, A.; Sashida, Y.; Nikaido, T. Steroidal saponins from Allium chinense andtheir inhibitory activities on cyclic AMP phosphodiesterase and Na+ K+ ATPase. Phytochemistry 1995, 40,1071–1076. [CrossRef]
111. Peng, J.-P.; Yao, X.-S. 19 new steroidal saponins from Allium plants: Isolation, structural elucidation andeffect on blood coagulability. In Saponins Used in Traditional and Modern Medicine; Springer: Boston, MA,USA, 1996; pp. 511–526.
112. Jiang, Y.; Wang, N.-L.; Yao, X.-S.; Kitanaka, S. Steroidal saponins from the bulbs of Allium chinense. Stud.Plant Sci. 1999, 6, 212–219.
113. Barile, E.; Zolfaghari, B.; Sajjadi, S.E.; Lanzotti, V. Saponins of Allium e lburzense. J. Nat. Prod. 2004, 67,2037–2042. [CrossRef]
114. Chincharadze, D.; Kel’Ginbaev, A.; Gorovits, M.; Eristavi, L.; Gorovits, T.; Abubakirov, N. Steroid saponinsand sapogenins of Allium. XV. Eruboside B from Allium erubescens. Chem. Nat. Compd. 1979, 15, 442–446.[CrossRef]
115. Kravets, S.; Vollerner, Y.S.; Gorovits, M.; Abubakirov, N. Steriods of the spirostan and furostan series fromplants of the genus Allium. Chem. Nat. Compd. 1990, 26, 359–373. [CrossRef]
116. Do, J.C.; Jung, K.Y.; Son, K.H. Steroidal saponins from the subterranean part of Allium fistulosum. J. Nat. Prod.1992, 55, 168–173. [CrossRef]
117. KELGINBAEV, A.; Gorovits, M.; Gorovits, T.; Abubakirov, N. Steroidal saponins and sapogenins of Allium.9. Aginoside structure. Khimiya Prir. Soedin. 1976, 4, 480–486.
118. SAsHiDA, Y.; KAwASHIMA, K.; MIMAKI, Y. Novel polyhydroxylated steroidal saponins from Alliumgiganteum. Chem. Pharm. Bull. 1991, 39, 698–703. [CrossRef]
119. MIMAKI, Y.; NIKAIDO, T.; MATSUMOTO, K.; SASHIDA, Y.; OHMOTO, T. New steroidal saponins from thebulbs of Allium giganteum exhibiting potent inhibition of cAMP phosphodiesterase activity. Chem. Pharm.Bull. 1994, 42, 710–714. [CrossRef]
120. Mimaki, Y.; Kuroda, M.; Fukasawa, T.; Sashida, Y. Steroidal glycosides from the bulbs of Allium jesdianum.J. Nat. Prod. 1999, 62, 194–197. [CrossRef]
121. Mimaki, Y.; Kuroda, M.; Fukasawa, T.; SASHIDA, Y. Steroidal saponins from the bulbs of Allium karataviense.Chem. Pharm. Bull. 1999, 47, 738–743. [CrossRef]
122. Vollerner, Y.S.; Abdullaev, N.; Gorovits, M.; Abubakirov, N. Steroid saponins and sapogenins of Allium. XX.Structure of karatavisoides E and F. Chem. Nat. Compd. 1984, 20, 64–68. [CrossRef]
123. Inoue, T.; Mimaki, Y.; Sashida, Y.; Nishino, A.; Satomi, Y.; Nishino, H. Steroidal glycosides from Alliummacleanii and A. senescens, and their inhibitory activity on tumour promoter-induced phospholipid metabolismof HeLa cells. Phytochemistry 1995, 40, 521–525. [CrossRef]
124. Peng, J.; Yao, X.; Kobayashi, H.; Ma, C. Novel furostanol glycosides from Allium macrostemon. Planta Med.1995, 61, 58–61. [CrossRef]

Molecules 2019, 24, 2067 17 of 21
125. Lazur’Evskii, G.; Krokhmalyuk, V.; Kintya, P. Structure of steroid glycosides of Allium narcissiflorum Wills.Dokl. Biochem. 1975, 221, 151.
126. Krokhmalyuk, V.; Kintya, P. Steroid saponins. XIII. The structure of alliumosides D and E fromAllium narcissiflorum. Chem. Nat. Compd. 1976, 12, 165–168. [CrossRef]
127. Mimaki, Y.; Satou, T.; Ohmura, M.; Sashida, Y. Steroidal saponins from the bulbs of Allium narcissiflorum.Nat. Med. 1996, 50, 308.
128. Akhov, L.; Musienko, M.; Piacente, S.; Pizza, C.; Oleszek, W. Structure of steroidal saponins from undergroundparts of Allium nutans L. J. Agric. Food Chem. 1999, 47, 3193–3196. [CrossRef]
129. Akhov, L.; Musienko, M.; Shishova, Y.; Polishuk, V.; Oleszek, W. Biological Activity of Deltoside from AlliumNutans L. In Saponins in Food, Feedstuffs and Medicinal Plants; Springer: Dordrecht, The Netherlands, 2000;pp. 227–231.
130. Harmatha, J.; Mauchamp, B.; Arnault, C.; Sláma, K. Identification of a spirostane-type saponin in theflowers of leek with inhibitory effects on growth of leek-moth larvae. Biochem. Syst. Ecol. 1987, 15, 113–116.[CrossRef]
131. Carotenuto, A.; Fattorusso, E.; Lanzotti, V.; Magno, S. Spirostanol saponins of Allium porrum L. Phytochemistry1999, 51, 1077–1082. [CrossRef]
132. Fattorusso, E.; Lanzotti, V.; Taglialatela-Scafati, O.; Di Rosa, M.; Ianaro, A. Cytotoxic saponins from bulbs ofAllium porrum L. J. Agric. Food Chem. 2000, 48, 3455–3462. [CrossRef]
133. MATSUURA, H.; USHIROGUCHI, T.; ITAKURA, Y. Further studies on steroidal glycosides from bulbs, rootsand leaves of Allium sativum L. Chem. Pharm. Bull. 1989, 37, 2741–2743. [CrossRef]
134. Matsuura, H. Saponins in garlic as modifiers of the risk of cardiovascular disease. J. Nutr. 2001, 131,1000S–1005S. [CrossRef]
135. Lanzotti, V. Bioactive Saponins from Allium and Aster Plants. Phytochem. Rev. 2005, 4, 95–110. [CrossRef]136. Kawashima, K.; Mimaki, Y.; Sashida, Y. Steroidal saponins from the bulbs of Allium schubertii. Phytochemistry
1993, 32, 1267–1272. [CrossRef]137. Mimaki, Y.; Satou, T.; Kuroda, M.; Kameyama, A.; Sashida, Y.; Li, H.-y.; Harada, N. A new furostanol saponin
with six sugars from the bulbs of Allium sphaerosephalon structural elucidation by modern NMR techniques.Chem. Lett. 1996, 25, 431–432. [CrossRef]
138. Corea, G.; Fattorusso, E.; Lanzotti, V. Saponins and Flavonoids of Allium triquetrum. J. Nat. Prod. 2003, 66,1405–1411. [CrossRef] [PubMed]
139. Zou, Z.-M.; Yu, D.-Q.; Cong, P.-Z. A steroidal saponin from the seeds of Allium tuberosum. Phytochemistry2001, 57, 1219–1222. [CrossRef]
140. Sang, S.; Mao, S.; Lao, A.; Chen, Z.; Ho, C.-T. New steroid saponins from the seeds of Allium tuberosum L.Food Chem. 2003, 83, 499–506. [CrossRef]
141. Pirtskhalava, G.; Gorovits, M.; Gorovits, T.; Abubakirov, N. Steroid saponins and sapogenins of Allium. XVI.Turoside C from Allium turcomanicum. Chem. Nat. Compd. 1979, 15, 446–452. [CrossRef]
142. Chen, S.; Snyder, J.K. Diosgenin-bearing, molluscicidal saponins from Allium vineale: An NMR approach forthe structural assignment of oligosaccharide units. J. Org. Chem. 1989, 54, 3679–3689. [CrossRef]
143. Magid, A.A.; Bobichon, H.; Borie, N.; Lalun, N.; Long, C.; Moretti, C.; Lavaud, C. Cytotoxic triterpenoidsaponins from the stem bark of Antonia ovata. Phytochemistry 2010, 71, 429–434. [CrossRef]
144. Lavaud, C.; Massiot, G.; Moretti, C.; Le Men-Olivier, L. Triterpene saponins Frommyrsine pellucida.Phytochemistry 1994, 37, 1671–1677. [CrossRef]
145. Kitagawa, I.; Yoshikawa, M.; Kobayashi, K.; Imakura, Y.; Im, K.; Ikenishi, Y. Saponin and sapogenol. XXVIII.Reinvestigation of the branching positions in the glucuronide moieties of three glucuronide saponins:Desacyl-jegosaponin, desacyl-boninsaponin A, and sakuraso-saponin. Chem. Pharm. Bull. 1980, 28, 296–300.[CrossRef]
146. Lavaud, C.; Pichelin, O.; Massiot, G.; Le Men-Olivier, L.; Sevenet, T.; Cosson, J. Sakuraso-saponin fromTapeinosperma clethroides. Fitoterapia 1999, 70, 116–118. [CrossRef]
147. Lavaud, C.; Beauvière, S.; Massiot, G.; Le Men-Olivier, L.; Bourdy, G. Saponins from Pisonia umbellifera.Phytochemistry 1996, 43, 189–194. [CrossRef]
148. Nhiem, N.X.; Thu, V.K.; Van Kiem, P.; Van Minh, C.; Tai, B.H.; Quang, T.H.; Cuong, N.X.; Yen, P.H.; Boo, H.-J.;Kang, J.-I.; et al. Cytotoxic oleane-type triterpene saponins from Glochidion eriocarpum. Arch. Pharmacal Res.2012, 35, 19–26. [CrossRef] [PubMed]

Molecules 2019, 24, 2067 18 of 21
149. Montoya, G.; Arango, G.J.; Unger, M.; Holzgrabe, U. Saponins from Phytolacca bogotensis usingHPLC-ESI/multi-stage tandem mass spectrometry. Phytochem. Anal. 2009, 20, 465–474. [CrossRef]
150. Bankefors, J.; Nord, L.I.; Kenne, L. Structural classification of acyl-substituted Quillaja saponins byelectrospray ionisation ion trap multiple-stage mass spectrometry in combination with multivariate analysis.Rapid Commun. Mass Spectrom. 2008, 22, 3851–3860. [CrossRef] [PubMed]
151. Kensil, C.R.; Patel, U.; Lennick, M.; Marciani, D. Separation and characterization of saponins with adjuvantactivity from Quillaja saponaria Molina cortex. J. Immunol. 1991, 146, 431–437.
152. Zhan, C.; Li, X.; Zhao, Z.; Yang, T.; Wang, X.; Luo, B.; Zhang, Q.; Hu, Y.; Hu, X. Comprehensive Analysisof the Triterpenoid Saponins Biosynthetic Pathway in Anemone flaccida by Transcriptome and ProteomeProfiling. Front. Plant Sci. 2016, 7. [CrossRef] [PubMed]
153. Zhao, L.; Chen, W.-M.; Fang, Q.-C. Triterpenoid saponins from Anemone flaccida. Planta Med. 1990, 56, 92–93.[CrossRef]
154. Yuan, J.-Q.; Yang, X.-Z.; Miao, J.-H.; Tang, C.-P.; Ke, C.-Q.; Zhang, J.-B.; Ma, X.-J.; Ye, Y. New triterpeneglucosides from the roots of Rosa laevigata Michx. Molecules 2008, 13, 2229–2237. [CrossRef]
155. Lavaud, C.; Voutquenne, L.; Massiot, G.; Le Men-Olivier, L.; Delaude, C. Saponines triperpéniques deSmelophyllum capense (Sapindaceae). Bull. De La Société R. Des Sci. De Liège 1994, 63, 455–463.
156. Lavaud, C.; Voutquenne, L.; Massiot, G.; Le Men-Olivier, L.; Das, B.C.; Laprévote, O.; Serani, L.; Delaude, C.;Becchi, M. Saponins from the stem bark of Filicium decipiens. Phytochemistry 1998, 47, 441–449. [CrossRef]
157. Dimbi, M.Z.; Warin, R.; Delaude, C.; Huls, R.; Mpuza, K. Triterpenoides de Harpullia cupanioides. Bull. DesSociétés Chim. Belg. 1983, 92, 473–484. [CrossRef]
158. Voutquenne, L.; Lavaud, C.; Massiot, G.; Delaude, C. Saponins from Harpullia cupanioides. Phytochemistry1998, 49, 2081–2085. [CrossRef]
159. Shiau, I.-L.; Shih, T.-L.; Wang, Y.-N.; Chen, H.-T.; Lan, H.-F.; Lin, H.C.; Yang, B.-Y.; Ko, C.-H.; Murase, Y.Quantification for saponin from a soapberry (Sapindus mukorossi Gaertn) in cleaning products by achromatographic and two colorimetric assays. J. Fac. Agr. Kyushu Univ. 2009, 54, 215–221.
160. Massiot, G.; Lavaud, C.; Delaude, C.M.; Van Binst, G.; Miller, S.P.F.; Fales, H.M. Saponins from Tridesmostemonclaessenssi. Phytochemistry 1990, 29, 3291–3298. [CrossRef]
161. Wandji, J.; Tillequin, F.; Mulholland, D.A.; Shirri, J.C.; Tsabang, N.; Seguin, E.; Verite, P.; Libot, F.; Fomum, Z.Pentacyclic triterpenoid and saponins from Gambeya boukokoensis. Phytochemistry 2003, 64, 845–849. [CrossRef]
162. Lavaud, C.; Massiot, G.; Becchi, M.; Misra, G.; Nigam, S. Saponins from three species of Mimusops.Phytochemistry 1996, 41, 887–893. [CrossRef]
163. De Lucca, A.; Boue, S.; Palmgren, M.; Maskos, K.; Cleveland, T. Fungicidal properties of two saponins fromCapsicum frutescens and the relationship of structure and fungicidal activity. Can. J. Microbiol. 2006, 52,336–342. [CrossRef] [PubMed]
164. Li, X.-H.; Shen, D.-D.; Li, N.; Yu, S.-S. Bioactive triterpenoids from Symplocos chinensis. J. Asian Nat. Prod. Res.2003, 5, 49–56. [CrossRef] [PubMed]
165. Li, B.; Abliz, Z.; Tang, M.; Fu, G.; Yu, S. Rapid structural characterization of triterpenoid saponins in crudeextract from Symplocos chinensis using liquid chromatography combined with electrospray ionizationtandem mass spectrometry. J. Chromatogr. A 2006, 1101, 53–62. [CrossRef]
166. Fu, G.-M.; Wang, Y.-H.; Gao, S.; Tang, M.-J.; Yu, S.-S. Five new cytotoxic triterpenoid saponins from the rootsof Symplocos chinensis. Planta Med. 2005, 71, 666–672. [CrossRef] [PubMed]
167. Mitchell-Olds, T.; Clauss, M.J. Plant evolutionary genomics. Curr. Opin. Plant Biol. 2002, 5, 74–79. [CrossRef]168. Simms, E.L.; Rausher, M.D. Costs and benefits of plant resistance to herbivory. Am. Nat. 1987, 130, 570–581.
[CrossRef]169. Vrieling, K.; Smit, W.; van der Meijden, E. Tritrophic interactions between aphids (Aphis jacobaeae Schrank),
ant species, Tyria jacobaeae L. and Senecio jacobaea L. lead to maintenance of genetic variation in pyrrolizidinealkaloid concentration. Oecologia 1991, 86, 177–182. [CrossRef]
170. Cipollini, D. Variation in the expression of chemical defenses in Alliaria petiolata (Brassicaceae) in the fieldand common garden. Am. J. Bot. 2002, 89, 1422–1430. [CrossRef] [PubMed]
171. Van der Meijden, E.; Wijn, M.; Verkaar, H.J. Defence and regrowth, alternative plant strategies in the struggleagainst herbivores. Oikos 1988, 51, 355–363. [CrossRef]
172. Herms, D.A.; Mattson, W.J. The dilemma of plants: To grow or defend. Q. Rev. Biol. 1992, 67, 283–335.[CrossRef]

Molecules 2019, 24, 2067 19 of 21
173. Gershenzon, J. Metabolic costs of terpenoid accumulation in higher plants. J. Chem. Ecol. 1994, 20, 1281–1328.[CrossRef] [PubMed]
174. Strauss, S.Y.; Siemens, D.H.; Decher, M.B.; Mitchell-Olds, T. Ecological costs of plant resistance to herbivoresin the currency of pollination. Evolution 1999, 53, 1105–1113. [CrossRef] [PubMed]
175. Rosenthal, G.; Janzen, D. Herbivores: Their interaction with plant secondary metabolites; Academic Press:New York, NY, USA, 1979; 718p.
176. Hostettmann, K.; Marston, A.; Wolfender, J.L. In Strategy in the search for new biologically active plantconstituents. In Proceedings-Phytochemical Society of Europe; Oxford University Press Inc.: Oxford, UK, 1995;p. 17.
177. Abreu, A.C.; McBain, A.J.; Simoes, M. Plants as sources of new antimicrobials and resistance-modifyingagents. Nat. Prod. Rep. 2012, 29, 1007–1021. [CrossRef]
178. Harborne, J.B.; Baxter, H. Dictionary of Plant Toxins; John Wiley & Sons: UK, 1996.179. Morrissey, J.P.; Osbourn, A.E. Fungal resistance to plant antibiotics as a mechanism of pathogenesis.
Microbiol. Mol. Biol. Rev. 1999, 63, 708–724. [PubMed]180. Szakiel, A.; Paczkowski, C.; Henry, M. Influence of environmental abiotic factors on the content of saponins
in plants. Phytochem. Rev. 2011, 10, 471–491. [CrossRef]181. Sampaio, B.L.; Edrada-Ebel, R.; Da Costa, F.B. Effect of the environment on the secondary metabolic profile of
Tithonia diversifolia: A model for environmental metabolomics of plants. Sci. Rep. 2016, 6, 29265. [CrossRef]182. Applebaum, S.; Birk, Y. Herbivores: Their Interactions with Secondary Plant Metabolites; Acad. Press: New York,
NY, USA, 1979; pp. 539–566.183. Hostettmann, K.; Marston, A. Chemistry and Pharmacology of Natural Products, Saponin; Cambridge University
Press: London, UK, 1995.184. Purkayastha, J.; Arora, R.; Singh, L. Sustainable and Novel Eco-friendly Approaches Towards Integrated
Disease and Vector Management. In Herbal Insecticides, Repellents and Biomedicines: Effectiveness andCommercialization; Springer: New Delhi, India, 2016; pp. 11–23.
185. Alias, E.E.M.; Zeinelabdin, M.H.; Bashir, N.H.H.; Assad, Y.O.H.; Abdelbagi, O.M. Hypoglycemic and toxiceffects of saponins from the fruit of bitter apple [Citrullus colocynthis (L.) Schrad] on the internal organs ofNorway rat [Rattus norvegicus (Berkenhout)]. Gezira J. Agric. Sci. 2015, 13, 1–17.
186. Fuenzalida, T. Plant natural defense against insects: Role of secondary metabolites. Santiago 2015, 1–24.187. Furlong, M.J.; Wright, D.J.; Dosdall, L.M. Diamondback moth ecology and management: Problems, progress,
and prospects. Annu Rev Entomol 2013, 58, 517–541. [CrossRef]188. Heckel, D.G.; Gahan, L.J.; Liu, Y.B.; Tabashnik, B.E. Genetic mapping of resistance to Bacillus thuringiensis
toxins in diamondback moth using biphasic linkage analysis. Proc. Natl. Acad. Sci. USA 1999, 96, 8373–8377.[CrossRef]
189. Tabashnik, B.E.; Huang, F.; Ghimire, M.N.; Leonard, B.R.; Siegfried, B.D.; Rangasamy, M.; Yang, Y.; Wu, Y.;Gahan, L.J.; Heckel, D.G.; et al. Efficacy of genetically modified Bt toxins against insects with differentgenetic mechanisms of resistance. Nat. Biotechnol. 2011, 29, 1128–1131. [CrossRef]
190. Shelton, A.M.; Badenes-Perez, F.R. Concepts and applications of trap cropping in pest management.Annu. Rev. Entomol. 2006, 51, 285–308. [CrossRef]
191. Badenes-Perez, F.R.; Shelton, A.M.; Nault, B.A. Evaluating trap crops for diamondback moth, Plutella xylostella(Lepidoptera: Plutellidae). J. Econ. Entomol. 2004, 97, 1365–1372. [CrossRef]
192. Lu, J.H.; Liu, S.S.; Shelton, A.M. Laboratory evaluations of a wild crucifer Barbarea vulgaris as a managementtool for the diamondback moth Plutella xylostella (Lepidoptera: Plutellidae). Bull. Entomol. Res. 2004, 94,509–516. [CrossRef] [PubMed]
193. Shelton, A.M.; Nault, B.A. Dead-end trap cropping: A technique to improve management of the diamondbackmoth, Plutella xylostella (Lepidoptera: Plutellidae). Crop Prot 2004, 23, 497–503. [CrossRef]
194. Badenes-Perez, F.R.; Shelton, A.M.; Nault, B.A. Using yellow rocket as a trap crop for diamondback moth(Lepidoptera: Plutellidae). J. Econ. Entomol. 2005, 98, 884–890. [CrossRef]
195. MacDonald, M.A.; Cavers, P.B. The biology of Canadian weeds: 97. Barbarea vulgaris R. Br. Can. J. Plant Sci.1991, 71, 149–166. [CrossRef]
196. Badenes-Perez, F.R.; Reichelt, M.; Gershenzon, J.; Heckel, D.G. Using plant chemistry and insect preferenceto study the potential of Barbarea (Brassicaceae) as a dead-end trap crop for diamondback moth (Lepidoptera:Plutellidae). Phytochemistry 2014, 98, 137–144. [CrossRef]

Molecules 2019, 24, 2067 20 of 21
197. Newman, K.; You, M.; Vasseur, L. Diamondback Moth (Lepidoptera: Plutellidae) Exhibits Oviposition andLarval Feeding Preferences Among Crops, Wild plants, and Ornamentals as Host Plants. J. Econ. Entomol.2016, tow002. [CrossRef]
198. Boege, K.; Marquis, R.J. Facing herbivory as you grow up: The ontogeny of resistance in plants. Trends Ecol.Evol. 2005, 20, 441–448. [CrossRef] [PubMed]
199. Petersen, B.; Chen, S.; Hansen, C.; Olsen, C.; Halkier, B. Composition and content of glucosinolates indeveloping Arabidopsis thaliana. Planta 2002, 214, 562–571. [CrossRef] [PubMed]
200. Wallace, S.K.; Eigenbrode, S.D. Changes in the glucosinolate–myrosinase defense system in Brassica junceacotyledons during seedling development. J. Chem. Ecol. 2002, 28, 243–256. [CrossRef]
201. Agerbirk, N.; Olsen, C.E.; Bibby, B.M.; Frandsen, H.O.; Brown, L.D.; Nielsen, J.K.; Renwick, J.A.A. ASaponin Correlated with Variable Resistance of Barbarea vulgaris to the Diamondback Moth Plutella xylostella.J. Chem. Ecol. 2003, 29, 1417–1433. [CrossRef] [PubMed]
202. Nielsen, N.J.; Nielsen, J.; Staerk, D. New resistance-correlated saponins from the insect-resistant cruciferBarbarea vulgaris. J. Agric. Food Chem. 2010, 58, 5509–5514. [CrossRef] [PubMed]
203. van Mölken, T.; Kuzina, V.; Munk, K.R.; Olsen, C.E.; Sundelin, T.; van Dam, N.M.; Hauser, T.P. Consequencesof combined herbivore feeding and pathogen infection for fitness of Barbarea vulgaris plants. Oecologia 2014,175, 589–600. [CrossRef]
204. De Geyter, E.; Swevers, L.; Caccia, S.; Geelen, D.; Smagghe, G. Saponins show high entomotoxicity by cellmembrane permeation in Lepidoptera. Pest Manag. Sci. 2012, 68, 1199–1205. [CrossRef]
205. Potter, S.M.; Jimenez-Flores, R.; Pollack, J.; Lone, T.A.; Berber-Jimenez, M.D. Protein-saponin interaction andits influence on blood lipids. J. Agric. Food Chem. 1993, 41, 1287–1291. [CrossRef]
206. Soetan, K.; Ajibade, T.; Akinrinde, A. Saponins; A Ubiquitous Phytochemical: A Review of its Biochemical,Physiological and Pharmacological Effects. Recent Prog. Med. Plants 2014, 43, 1–24.
207. Amtul, J.S.; Shakoori, A.R. Potential of Azadirachtin and Neem (Azadirachta indica) Based Saponins asBiopesticides for In vitro Insect Pests Cellulase (Beta-1, 4-Endoglucanase) Enzyme Inhibition and In vivoRepellency on Tribolium castaneum. Br. Biotechnol. J. 2014, 4, 904–917.
208. Goławska, S.; Łukasik, I.; Wójcicka, A.; Sytykiewicz, H. Relationship between saponin content in alfalfa andaphid development. Acta Biol. Crac. Ser. Bot. 2012, 54, 39–46. [CrossRef]
209. Tava, A.; Odoardi, M. Saponins from Medicago spp.: Chemical characterization and biological activity againstinsects. In Saponins Used in food and Agriculture; Springer: Boston, MA, USA, 1996; pp. 97–109.
210. Vincken, J.-P.; Heng, L.; de Groot, A.; Gruppen, H. Saponins, classification and occurrence in the plantkingdom. Phytochemistry 2007, 68, 275–297. [CrossRef] [PubMed]
211. Lee, H.-A.; Lee, H.-Y.; Seo, E.; Lee, J.; Kim, S.-B.; Oh, S.; Choi, E.; Choi, E.; Lee, S.E.; Choi, D. CurrentUnderstandings on Plant Nonhost Resistance. Mol. Plant-Microbe Interact. 2017, 30, 5–15. [CrossRef]
212. Dowd, P.F.; Berhow, M.A.; Johnson, E.T. Differential activity of multiple saponins against omnivorous insectswith varying feeding preferences. J. Chem. Ecol. 2011, 37, 443–449. [CrossRef]
213. Osbourn, A. Saponins and plant defence—A soap story. Trends Plant Sci. 1996, 1, 4–9. [CrossRef]214. Osbourn, A.E.; Wubben, J.P.; Melton, R.E.; Carter, J.P.; Daniels, M.J. Saponins and plant defense.
In Phytochemical Signals and Plant-Microbe Interactions; Springer: Boston, MA, USA, 1998; pp. 1–15.215. Chaieb, I. Saponins as insecticides: A review. Tunis. J. Plant Prot. 2010, 5, 39–50.216. Akhtar, Y.; Isman, M.B. Binary mixtures of feeding deterrents mitigate the decrease in feeding deterrent
response to antifeedants following prolonged exposure in the cabbage looper, Trichoplusia ni (Lepidoptera:Noctuidae). Chemoecology 2003, 13, 177–182. [CrossRef]
217. Akhtar, Y.; Isman, M.B. Larval exposure to oviposition deterrents alters subsequent oviposition behaviorin generalist, Trichoplusia ni and specialist, Plutella xylostella moths. J. Chem. Ecol. 2003, 29, 1853–1870.[CrossRef]
218. Kumar, R.; Bhardwaj, U.; Kumar, P.; Mazumdar-Leighton, S. Midgut serine proteases and alternative hostplant utilization in Pieris brassicae L. Front. Physiol. 2015, 6, 95. [CrossRef]
219. Frisch, T.; Agerbirk, N.; Davis, S.; Cipollini, D.; Olsen, C.E.; Motawia, M.S.; Bjarnholt, N.; Møller, B.L.Glucosinolate-related glucosides in Alliaria petiolata: Sources of variation in the plant and different metabolismin an adapted specialist herbivore, Pieris rapae. J. Chem. Ecol. 2014, 40, 1063–1079. [CrossRef] [PubMed]
220. Idris, A.B.; Grafius, E. The potential of using Barbarea vulgaris in insecticide-resistant diamondback mothmanagement. Resist. Pest Manag. (USA) 1994, 6, 7–8.

Molecules 2019, 24, 2067 21 of 21
221. Badenes-Pérez, F.R.; Márquez, B.P.; Petitpierre, E. Can flowering Barbarea spp. (Brassicaceae) be usedsimultaneously as a trap crop and in conservation biological control? J. Pest Sci. 2016, 4, 1–11. [CrossRef]
222. Badenes-Pérez, F.R.; Reichelt, M.; Gershenzon, J.; Heckel, D.G. Phylloplane location of glucosinolatesin Barbarea spp. (Brassicaceae) and misleading assessment of host suitability by a specialist herbivore.New Phytol. 2011, 189, 549–556. [CrossRef]
223. Vermeer, K.M.C.A. Genetics of Insect Resistance to Plant Defence; Wageningen University: Wageningen, TheNetherlands, 2014.
224. Wittstock, U.; Gershenzon, J. Constitutive plant toxins and their role in defense against herbivores andpathogens. Curr. Opin. Plant Biol. 2002, 5, 300–307. [CrossRef]
225. Mithöfer, A.; Maffei, M.E. General mechanisms of plant defense and plant toxins. In Plant Toxins; SpringerAG: Switzerland, 2016; pp. 1–22.
226. Sparg, S.G.; Light, M.E.; Van Staden, J. Biological activities and distribution of plant saponins. J. Ethnopharmacol.2004, 94, 219–243. [CrossRef]
227. Dinan, L. Phytoecdysteroids: Biological aspects. Phytochemistry 2001, 57, 325–339. [CrossRef]228. Harder, M.J.; Tello, V.E.; Giliomee, J.H. The acaricidal effect of ethanolic extracts of Chenopodium quinoa Willd.
on Tetranychus urticae Koch (Acari: Tetranychidae). Afr. Entomol. 2016, 24, 50–60. [CrossRef]229. Nawrot, J.; Koul, O.; Isman, M.; Harmatha, J. Naturally occurring antifeedants: Effects on two polyphagous
lepidopterans. J. Appl. Entomol. 1991, 112, 194–201. [CrossRef]230. Goławska, S.; Sprawka, I.; Łukasik, I. Effect of saponins and apigenin mixtures on feeding behavior of the
pea aphid, Acyrthosiphon pisum Harris. Biochem. Syst. Ecol. 2014, 55, 137–144. [CrossRef]231. Pavela, R. Extract from the roots of Saponaria officinalis as a potential acaricide against Tetranychus urticae.
J. Pest Sci. 2017, 90, 683–692. [CrossRef]232. Wina, E.; Muetzel, S.; Becker, K. The impact of saponins or saponin-containing plant materials on ruminant
production—A review. J. Agric. Food Chem. 2005, 53, 8093–8105. [CrossRef]233. Tamura, Y.; Miyakoshi, M.; Yamamoto, M. Application of saponin-containing plants in foods and cosmetics.
In Alternative Medicine; InTech: London, UK, 2012.234. Jia, Z.; Koike, K.; Nikaido, T. Major triterpenoid saponins from Saponaria officinalis. J. Nat. Prod. 1998, 61,
1368–1373. [CrossRef] [PubMed]235. Agerbirk, N.; Olsen, C.E.; Nielsen, J.K. Seasonal variation in leaf glucosinolates and insect resistance in two
types of Barbarea vulgaris ssp. arcuata. Phytochemistry 2001, 58, 91–100. [CrossRef]236. Borah, B.; Phukon, P.; Hazarika, M.P.; Ahmed, R.; Sarmah, D.K.; Wann, S.B.; Das, A.; Bhau, B.S. Calamus
leptospadix Griff. a high saponin yielding plant with antimicrobial property. Ind. Crop. Prod. 2016, 82, 127–132.[CrossRef]
237. Lanzotti, V.; Barile, E.; Antignani, V.; Bonanomi, G.; Scala, F. Antifungal saponins from bulbs of garlic,Allium sativum L. var. Voghiera. Phytochemistry 2012, 78, 126–134. [CrossRef]
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open accessarticle distributed under the terms and conditions of the Creative Commons Attribution(CC BY) license (http://creativecommons.org/licenses/by/4.0/).




















