Role of increased sphingomyelinase activity in apoptosis and organ failure of patients with severe...
21
©2005 FASEB The FASEB Journal express article 10.1096/fj.04-2842fje. Published online July 28, 2005. Role of increased sphingomyelinase activity in apoptosis and organ failure of patients with severe sepsis Ralf A. Claus,* Alexander C. Bunck,* Clemens L. Bockmeyer,* Frank M. Brunkhorst,* Wolfgang Lösche,* Ralf Kinscherf, † and Hans-Peter Deigner ‡ *Department of Anesthesiology and Intensive Care Medicine, Friedrich Schiller University Jena, Erlanger Allee 101, Jena, Germany; † Department of Anatomy and Cell Biology III, University of Heidelberg, Im Neuenheimer Feld 307, 69120 Heidelberg, Germany; ‡ School of Chemical Sciences and Pharmacy University of East Anglia, Norwich, UK Corresponding authors: Ralf A. Claus, Department of Anesthesiology and Intensive Care Medicine, Division for Experimental Anesthesiology, Erlanger Allee 101, D-07747 Jena, Germany. E-mail: [email protected]; H.-Peter Deigner, School of Chemical Sciences and Pharmacy, University of East Anglia UEA, Norwich NR4 7TJ, United Kingdom. E-mail: [email protected] ABSTRACT Numerous studies support the notion that an activation of sphingomyelinases and a subsequent increase of the concentration of the bioactive lipid mediator ceramide are critical in the concert of inflammatory stimuli and to the induction of apoptosis during inflammation. Here we show that patients with severe sepsis exhibit an enhanced sphingolytic activity in comparison with controls [262 pmol/(mlxh) vs. 123.6 pmol/(mlxh), P<0.005]. During the clinical course, a further increase was paralleled by the severity of illness and by fatal outcome. Moreover, we show that oxidative stress may partially account for the increased activity through posttranslational modification of the enzyme. In a murine endotoxic shock model, administration of a low molecular weight inhibitor diminished the rise in enzymatic activity and improved the survival rate. In liver specimen, inhibition of activity correlated with a reduced rate of hepato-cellular apoptosis. Our data support the concept that activation of the plasmatic isoform of sphingomyelinase may play a critical role in the development of apoptosis and organ failure in sepsis. An inhibition of the secreted isoform of sphingomyelinase should be explored further as a potential target in the complicated puzzle of sepsis. Key words: inflammation • ceramide • endotoxin • oxidative stress • desipramine epsis frequently leads to multiple organ failure (MOF) and is the most common cause of mortality in intensive care units (ICUs). In numerous studies the diagnostic and therapeutical value of a variety of markers and mediators has been investigated (1, 2). Although various pathophysiological changes during the early and late stages of the disease have been analyzed in detail, a large number of biological mechanisms activated during sepsis are not yet fully understood. Since anti-cytokine strategies have failed so far, the modulation of signal S Page 1 of 21 (page number not for citation purposes)
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Role of increased sphingomyelinase activity in apoptosis and organ failure of patients with severe...

©2005 FASEB The FASEB Journal express article 10.1096/fj.04-2842fje. Published online July 28, 2005. Role of increased sphingomyelinase activity in apoptosis and organ failure of patients with severe sepsis Ralf A. Claus,* Alexander C. Bunck,* Clemens L. Bockmeyer,* Frank M. Brunkhorst,* Wolfgang Lösche,* Ralf Kinscherf,† and Hans-Peter Deigner‡
*Department of Anesthesiology and Intensive Care Medicine, Friedrich Schiller University Jena, Erlanger Allee 101, Jena, Germany; †Department of Anatomy and Cell Biology III, University of Heidelberg, Im Neuenheimer Feld 307, 69120 Heidelberg, Germany; ‡School of Chemical Sciences and Pharmacy University of East Anglia, Norwich, UK
Corresponding authors: Ralf A. Claus, Department of Anesthesiology and Intensive Care Medicine, Division for Experimental Anesthesiology, Erlanger Allee 101, D-07747 Jena, Germany. E-mail: [email protected]; H.-Peter Deigner, School of Chemical Sciences and Pharmacy, University of East Anglia UEA, Norwich NR4 7TJ, United Kingdom. E-mail: [email protected] ABSTRACT
Numerous studies support the notion that an activation of sphingomyelinases and a subsequent increase of the concentration of the bioactive lipid mediator ceramide are critical in the concert of inflammatory stimuli and to the induction of apoptosis during inflammation. Here we show that patients with severe sepsis exhibit an enhanced sphingolytic activity in comparison with controls [262 pmol/(mlxh) vs. 123.6 pmol/(mlxh), P<0.005]. During the clinical course, a further increase was paralleled by the severity of illness and by fatal outcome. Moreover, we show that oxidative stress may partially account for the increased activity through posttranslational modification of the enzyme. In a murine endotoxic shock model, administration of a low molecular weight inhibitor diminished the rise in enzymatic activity and improved the survival rate. In liver specimen, inhibition of activity correlated with a reduced rate of hepato-cellular apoptosis. Our data support the concept that activation of the plasmatic isoform of sphingomyelinase may play a critical role in the development of apoptosis and organ failure in sepsis. An inhibition of the secreted isoform of sphingomyelinase should be explored further as a potential target in the complicated puzzle of sepsis.
Key words: inflammation • ceramide • endotoxin • oxidative stress • desipramine
epsis frequently leads to multiple organ failure (MOF) and is the most common cause of mortality in intensive care units (ICUs). In numerous studies the diagnostic and therapeutical value of a variety of markers and mediators has been investigated (1, 2).
Although various pathophysiological changes during the early and late stages of the disease have been analyzed in detail, a large number of biological mechanisms activated during sepsis are not yet fully understood. Since anti-cytokine strategies have failed so far, the modulation of signal
S
Page 1 of 21(page number not for citation purposes)

transduction pathways mediating the converging cytokine actions has been proposed as a more promising therapeutic approach (3, 4).
Various bioactive lipids—sphingolipids in particular—have been implicated in the sophisticated signaling network that regulates immunological processes during inflammation (5). Key enzymes in the initiation of the sphingomyelin/ceramide signaling pathway are sphingomyelinases that catalyze the breakdown of sphingomyelin to the potent second messenger ceramide (6).
In this study we focused on SMPD1, which is a product of the acid sphingomyelinase gene (7) and an enzyme present in both plasma (pSMPD1) and intracellular lysosomes (8). Among the known types of sphingomyelinases, the secretory variant pSMPD1 is the only sphingomyelinase held responsible for an extracellular hydrolysis of sphingomyelin (7). pSMPD1 has been found to be secreted by macrophages, human skin fibroblasts, and human vascular endothelial cells (8, 9). In endothelial cells, apical as well as basolateral secretion is stimulated by a variety of proinflammatory mediators such as IL-1β, IFN-γ, IFN-β, TNF-α, and endotoxin (9). Moreover, a two- to threefold increase in plasma sphingolytic activity was observed in animal models after application of endotoxin or proinflammatory cytokines (TNF-α, IL-1β) (10, 11).
Indirect evidence for an altered pSMPD1 activity during systemic inflammation has previously been found. Increased ceramide/sphingomyelin ratio in septic patients as well as its association with a poor clinical outcome were reported by Drobnik et al. (12). Delogu et al. (13) observed an increased ceramide concentration in mononuclear cells of septic patients that was positively correlated with plasma TNF-α levels and was higher among patients with MOF than in those without MOF. Furthermore, raised pSMPD1 levels in children with hypercytokinemia due to hemophagocytic lymphohistiocytosis have also been reported (14).
To achieve a deeper insight into the role of pSMPD1 activity during sepsis, we analyzed the presence and activity in plasma of patients with sepsis and healthy controls and examined a potential association with clinical parameters of the acute phase response and multiple organ failure. Moreover, we addressed the question whether mechanisms other than a cytokine-stimulated secretion might be involved in the observed increase of pSMPD1 among septic patients. For this purpose we examined the immediate effects of oxidative stress on enzyme activity in a human ex vivo model. Moreover, we used an experimental animal model of endotoxic shock to evaluate the effect of an inhibition of SMPD1 by a novel, specifically designed carbazol-derivative (NB6) (15) on hepato-cellular apoptosis and mortality.
MATERIALS AND METHODS
Study population and control group
The study was conducted in the intensive care unit (ICU) of the Department of Anaesthesiology and Intensive Care Medicine, Friedrich-Schiller University, Jena, Germany. Procedures were reviewed and approved by the local ethics committee, and written consent was obtained whenever possible from the patient, otherwise from a legal representative. Twelve patients hospitalized in the ICU and meeting the clinical criteria of severe sepsis or septic shock, as defined according to the ACCP/SCCM Consensus conference (16), were enrolled prospectively.
Page 2 of 21(page number not for citation purposes)

The first clinical evaluation and blood sampling were performed within 6 h after diagnosis. All patients were treated with antibiotics empirically according to our ward protocol. Adjustments of antimicrobial treatment were performed as needed based on the results of resistance analysis. Levels of the observed plasma proteins were also studied in a nonseptic, age-matched control group, consisting of 13 healthy volunteers [median age: 61.0 years, interquartile range (IQR) 25.8].
Sample collection
Blood specimens were obtained through central venous catheters. Plasma samples were collected in nonheparinized tubes for measurement of pSMPD1 (see below). Sodium citrate was added as an inhibitor of blood clotting, and all samples were strictly kept at 4°C, centrifuged (1000×g, 15 min, 4°C) within 30 min of collection, aliquoted, and stored at –80°C until assayed. A group of 13 healthy male individuals from the ICU and laboratory personnel formed the basis for determination of reference values for pSMPD1.
Additional clinical data
As soon as diagnosis of severe sepsis or septic shock was established, risk of death was evaluated by assessment of the Acute Physiology and Chronic Health Evaluation (APACHE) II scoring system at baseline (17). Furthermore, the Sepsis-related Organ Failure Assessment (SOFA) score as reported by Vincent et al. (18) was determined for each study day. Serum levels of creatinine, procalcitonin, lactate, C-reactive protein, thromboplastin time as well as the number of platelets were determined using commercially available test kits in the university’s Department for Clinical Chemistry and Laboratory Medicine.
Determination of pSMPD1 activity
pSMPD1 activity was determined by the hydrolysis of fluorescently labeled sphingomyelin (NBD-SM, Molecular Probes, Eugene, OR) as a substrate, chromatographic product separation, and image analysis as described previously (19) with the following modifications: the final composition of the reaction mixture was 10 µl plasma, 10 µl zinc (Zn2+) sulfate solution 2 mM, 1.0 nmol substrate in a final volume of 200 µl of 0.1% NP-40/62 mmol sodium acetate, pH 5.0. Incubation was performed at 37°C for 2 h; the extraction was carried out according to Bligh and Dyer (20). Product separation was completed using silica gel TLC plates 60 F254 (Merck, Darmstadt, Germany) and a mixture of chloroform/methanol/ammonium hydroxide (2 molar) 5/25/4 (v/v/v). The resulting NBD-labeled ceramide was assessed with a FLA-5 000 imaging system using the LBP filter set at 300 V/100 µm resolution (Fuji-Raytest, Berlin, Germany). Due to higher activity in murine plasma, plasma samples were diluted 1:5 with incubation buffer (sodium acetate, pH 5.0) before analysis. Specific activity of pSMPD1 was calculated by comparison with a calibration curve, which was linear for values corresponding to an activity of up to 2000 pmol/(mlxh).
Analysis of oxidative stress on enzyme activity
We examined the effects of 2,2'-azobis (2-amidinopropane) dihydrochloride (AAPH, Sigma, Deisenhofen, Germany) on pSMPD1 activity in the plasma of 12 healthy volunteers. AAPH is
Page 3 of 21(page number not for citation purposes)

frequently used as a model substance to simulate the effects of oxidative stress in human plasma (21). For this purpose, 10 μl of a 200 mM AAPH buffer solution was added to the reaction mixture, to give a final concentration of 10 mM. To illustrate a direct dependency between the observed effect and the induction of oxidative stress, 10 μl ascorbic acid (4 mM) in buffer solution was added to an otherwise identical set. A direct effect of the antioxidative ascorbic acid on enzyme activity was excluded; incubation time was extended to 3 h.
Characterization of NB 6
For in vivo inhibition of SMPD1, NB6, (3-carbazol-9-yl-propyl)-(2-(3,4-dimethoxy-phenyl)-ethyl)-methylamine was used (15). NB6 was solved in DMSO (Sigma) at a concentration of 125 mg ml–1, diluted to a final concentration of 0.8 mg ml–1 in NaCl (0.9%) and stored in aliquots at –20°C. DMSO (0.6% in isotonic NaCl solution) was used as vehicle control.
Experimental animals and animal model
Animal experiments were performed in accordance with local authorities (Landesverwaltungsamt Thüringen, 740.73-2684/02-08/98, 22.2684.04-02-30/4). Female NMRI mice (60–80 days old, weight 30–35 g) were used in this study. For the purpose of biochemical measurements and lethality experiments, endotoxic shock was triggered by intraperitoneal application of 0.9–1.2 mg endotoxin (E. coli, serotype 0111:B4, 2 mg ml–1, dissolved in NaCl 0.9%, dispersed by ultrasound) in 31 animals of the verum groups (groups 1–4) and 29 controls (groups 1c–4c). The amount of endotoxin applied in matched pairs of mice treated with vehicle or verum was varied slightly in order to obtain similar physiological responses and survival times in each of the experimental series. NB6 was applied by repetitive injections (i.e., every 24 h during the first 3 days after endotoxin challenge) in order to maintain adequate plasma levels of NB6 as well as to observe possible dose-dependent effects and toxicity of the substance. During the study term (maximum 7 days), the animals were kept under standardized conditions with access to food and water ad libitum.
For mortality studies, animals were randomly divided into four groups: group 1 (n=7), 1.2 mg endotoxin, 440 µg NB6 each in four repetitive injections (30 min before endotoxin; 1, 3, and 9 h after endotoxin injections); group 2 (n=7), 1.0 mg endotoxin, 440 µg NB6, four repetitive injections as in group 1; and group 3 (n=8) and group 4 (n=9) with identical treatment: 0.9 mg endotoxin, 800 µg NB6 given repetitively 30 min before endotoxin injection and at time 1, 3, 7, and 11 h after endotoxin administration along with controls (n=7 for groups 1c, 2c, and 3c and n=8 for group 4c) treated with equivalent amounts of endotoxin and vehicle (isotonic saline) injected at the same time schedule as given for the treatment groups.
For analysis of pSMPD1, animals were randomly divided into four groups (n=9 each), and citrated plasma samples were drawn 24 h before experimentation for determination of basal activity by retro-orbital blood collection (150 µl) and centrifugation (4 min, 4°C, 1000g). Groups received sublethal endotoxin dosages i.p. (group 1 and 2) or vehicle (group 3 and 4). NB6 was administered s.c. (group 2 and 4) in four repetitive injections (440 µg each, 30 min before experimentation; 1, 4, and 8 h after endotoxin/saline treatment); group 1 and 3 received saline vehicle at the same time schedule. After 24 h, plasma samples were drawn as described.
Page 4 of 21(page number not for citation purposes)

For preparation of histological samples, two additional groups of animals were pretreated with either NB6 (n=31, 880 µg per injection) or vehicle (n=31) 30 min before endotoxin application (1 mg). Repetitive dosages were administered 60 and 240 min after endotoxin injection. Seven hours after induction of endotoxic shock, mice were killed by cervical dislocation (performed under short-term anesthesia during spontaneous ventilation using isoflurane 5 vol%). For preparation of histological specimens, liver samples were harvested, immediately fixed with 4% paraformaldehyde, embedded in paraffin or embedded in Tissue-Tec, cryofixed in liquid nitrogen-cooled isopentane, and stored at –80°C until analyzed. In situ DNA fragmentation during apoptosis was identified by detection of TUNEL-positive cells with additional morphological characteristics of apoptosis such as cell shrinkage, chromatin condensation, and margination as described previously (22–24).
Data presentation and statistical analysis
Statistical analysis was carried out using the Winstat for Excel and the SPSS (13.0) software packages. The exact, nonparametric Mann-Whitney U test was applied for comparison of patients vs. healthy controls, survivor vs. nonsurvivor at baseline, and absolute changes of enzyme activity over the whole study period. Data are given as median (Q1, Q3).
Maximum and minimum values of SMPD1 activity were tested for an association with alterations in other clinical variables such as SOFA or procalcitonin plasma level. The set of two values for a variable of interest corresponding to these extremes was analyzed by the exact Wilcoxon test for paired samples. To estimate the magnitude of the difference between the two values corresponding to maximum and minimum pSMDP1 activities, the median difference was determined. Differences were defined to be positive if the clinical variable of interest had a higher value on the day with maximal pSMPD1 activity than on the day with minimal pSMPD1 activity. We considered values of P < 0.05 to be statistically significant. After log transformation, effects of AAPH on pSMPD1 activity were compared using the paired sample t test. Activity is shown as the means ± SD; P < 0.05 was considered significant.
Regarding the animal experiments, differences between the verum vs. placebo group were calculated using the logrank test and the Gehan-Wilcoxon test. Survival rate data are presented in Kaplan-Meier plots. To analyze hepatic apoptosis rates, results of NB-6 vs. placebo group were compared using the Mann-Whitney U test. For analysis of change in pSMPD1 activity, basal activity levels were compared with activity levels 24 h after endotoxin application using the Wilcoxon test. Activity levels at 24 h after endotoxin challenge were compared using the Mann-Whitney U test; P < 0.05 was considered significant.
RESULTS
Plasmatic activity of SMPD1
A method for determination of zinc (Zn2+)-dependent acid sphingomyelinase in human and murine plasma has been evaluated in the present study. Calculations of the amount of resulting ceramide due to the sphingolytic activity of pSMPD1 were performed on the basis of a calibration plot for human and murine samples in appropriate concentrations (Fig. 1A), and the presence of pSMPD1 was confirmed in plasma of two septic patients (Fig. 1B).
Page 5 of 21(page number not for citation purposes)

To obtain further information on variations of sphingolytic activities in the course of endotoxic shock, we analyzed pSMPD1 activities in plasma samples from mice (n=9) 24 h after endotoxin challenge. Average activity in the saline group raised from 2458 to 5036 pmol/(mlxh). Next, an inhibitor of SMPD1 biosynthesis (NB6) (15) was administered to reduce ceramide formation. The schedules for dosage and application intervals were derived from cell culture experiments, and our previous experiences with testing of related compounds (25, 26). Preadministration of NB6 prevented the increase of pSMPD1 activity as compared with endotoxin [median value, 2797 pmol/(mlxh), P<0.05] but had no effect on the enzyme activity in animals without endotoxin challenge [2875 pmol/(mlxh)] (Fig. 2A). Absolute changes in SMPD1 activity in mice treated with endotoxin without NB6 pretreatment in comparison to basal levels are illustrated in Fig. 2B (P<0.05).
Beneficial effects of SMPD1 inhibition in an endotoxic shock model
In an NMRI mice model, endotoxic shock was triggered by i.p. endotoxin administration. As illustrated in the Kaplan-Meier diagram, analysis including all subgroups (details are given in Materials and Methods) showed an improved survival rate of NB6-treated mice compared with saline (Fig. 3). Twenty-six of 29 animals died in the saline-treated group during the period of administration of NB6 or vehicle of 3 days (89.7%), whereas the mortality rate was significantly lower in the NB6-treated group (20 of 31, 64.5%). Subgroup analysis revealed neither significant toxic effects nor increasing efficacy with increasing doses of NB6 injected (data not shown).
Previous studies indicated that the liver is a major organ responsible for perpetuation of sepsis-associated MOF, and induction of apoptosis during sepsis has been shown in human (27) and animal (28) tissue. To further characterize a potential antiapoptotic effect of NB6 in a short-term experimental design, we analyzed hepatic DNA fragmentation using the TUNEL method and determination of morphological changes during apoptosis. After triggering endotoxic shock, we were able to demonstrate a significantly diminished number of stained cells (Fig. 4A) in the NB6-treated group (25%) (Fig. 4C) vs. the saline-treated group (43%) (Fig. 4B) (P<0.0001) 8 h after endotoxin administration.
pSMPD1 activity in critically ill patients
Encouraged by the results obtained in these animal experiments, we prospectively analyzed pSMPD1 activity in a group of critically ill patients. Main demographic, baseline clinical, and microbiological characteristics of the patients are summarized in Table 1. There were no statistically significant differences in ICU stay [nonsurvivors (NS): 12.0 vs. survivors (S): 19.0 days], accumulated ICU days of severe sepsis or septic shock (NS: 10.0 vs. S: 12.0 days) and APACHE II score (NS: 25.0 vs. S: 22.0 points). However, nonsurvivors were older (NS: 68 vs. S: 47 years). There were no differences in the SOFA score and lactate levels on the first day of diagnosis of sepsis/septic shock (Table 2), and procalcitonin levels were even higher in survivors (P<0.05).
Significantly elevated levels of pSMPD1 activity were found in patients in comparison with healthy controls: the median for the pSMPD1 activity in the control group was 123.6 pmol/(mlxh) [first quartile (Q1), 76.7; third quartile (Q3), 181.9] as opposed to a median of 262.3 pmol/(mlxh) (Q1, 234.4, Q3, 370.4) for the pSMPD1 activity in patients with sepsis at
Page 6 of 21(page number not for citation purposes)

baseline (P<0.005). The comparison between nonsurvivors and survivors revealed no significant difference for median pSMPD1 activities at baseline: 247.6 pmol/(mlxh) (Q1, 185.0; Q3, 320.1) vs. 264.7 pmol/(mlxh) (Q1, 259.9; Q3, 376.8) (Table 2).
In nonsurvivors (n=7) sphingolytic activity increased during the study period [+77.4 pmol/(mlxh), (Q1, –2.7; Q3, 365.8)] compared with a decrease in survivors [–252.1 pmol/(mlxh), (Q1, –254.8; Q3, –108.2)]. Considering intraindividual changes over the study period, there were significant differences between survivors and nonsurvivors. As shown in Fig. 5, we found a further increase among nonsurvivors by ~24% (Q1, 2.4; Q3, 143.9), but a decrease among survivors by ~65% (Q1, –66.9; Q3, –41.6) was observed (P<0.02).
Comparison of pSMPD1 activity with other clinical parameters
To obtain further insight into factors influencing pSMPD1 activity, we compared pSMPD1 activities with parameters relevant for clinical evaluation. We found that SOFA scores were significantly higher on days with maximal pSMPD1 activity than on days with minimal pSMPD1 activity [median difference: 4 points (1.5; 5.5) P<0.005 (one-sided)]. The same relationship was observed for procalcitonin [median difference: 1.84 ng/ml (0.3, 13.6), P<0.05 (one-sided), data not shown].
Effects of oxidative stress on enzyme activity
Qiu and colleagues observed an increase in enzymatic activity after oxidation of a C-terminally located cysteine group in recombinant SMPD1 (29). Thus, we addressed the question whether prooxidative conditions affect activity of pSMPD1 in human plasma. More specifically, we assessed the immediate effect of oxidative stress known to be increased under inflammatory conditions, such as observed during sepsis.
Basal pSMPD1 activity was determined as 122.7 ± 21.5 pmol/(mlxh). As expected, addition of ascorbic acid did not result in a significant change in sphingolytic activity [117.8±19.9 pmol/(mlxh)]. However, addition of the oxidant AAPH to the plasma was followed by a substantial rise in pSMPD1 activity by 87% to a value of 207.2 ± 26.1 pmol/(mlxh). This increase was significantly attenuated by parallel incubation with ascorbic acid. (Fig. 6).
DISCUSSION
This study shows for the first time that circulating pSMPD1 enzyme activity is markedly elevated in patients with sepsis. In fact, the observed plasma activities in septic patients clearly exceed the enzymatic activities reported to induce biological effects in cell culture systems (25, 26). The patients selected for our study were included prospectively and are deliberately heterogeneous to evaluate the general role of elevated plasma SMPD1 activity. Patients featured differences and diverse degrees of concomitant diseases, variable sources of infection, a broad range of sepsis severity as reflected by differing levels of inflammatory markers. Despite this heterogeneity, a uniform and broad increase in the activity of the sphingomyelin-cleaving pSMPD1 was observed; in patients with fatal outcome, an increase in sphingolytic activity over time could be demonstrated. The absolute changes of pSMPD1 activities during the observation period on ICU allowed discrimination between surviving and nonsurviving patients displaying an inverse trend of pSMPD1 activity. Our data further suggest that this increase is paralleled by
Page 7 of 21(page number not for citation purposes)

changes of established severity markers such as SOFA and procalcitonin known to predict outcome.
In keeping with these findings, the results of our animal studies provide further evidence for a crucial role of SMPD1 in the pathogenesis of systemic inflammation and subsequent organ failure, with impact on survival rate and apoptosis of end organs such as the liver.
Various mechanisms may be involved in the observed increase of SMPD1 activity in plasma of septic patients. Up to now, strong evidence has been provided for a cytokine- and endotoxin-stimulated secretion of pSMPD1 from endothelial cells (10, 30). However, in critically ill patients not only cytokines but also the release of reactive oxygen species contributes to the pathophysiology of systemic inflammation (31–33), oxidative stress is known to modulate the catalytic activity of a variety of enzymes (34). In our ex vivo setting, incubation of plasma in the presence of AAPH, as oxidative stimulus, resulted in a pronounced rise in sphingolytic activity. So far, an increase of enzymatic hydrolysis by oxidative modification has only been shown for the recombinant protein (29). Qiu et al. made the modification by a “cysteine switch” or a copper-induced dimerization via disulfide bond formation responsible for an increase in enzymatic hydrolysis (29). Our data suggest that this mechanism of posttranslational modification is also of relevance to the secreted sphingomyelinase in human plasma. Hence, oxidative stress induced by systemic inflammation might well have contributed to the increase in pSMPD1 activity that we observed in septic patients as well as in the endotoxic animal model.
Ceramide, the product of sphingomyelin hydrolysis, is known to induce the production of reactive oxygen species in endothelial cells (35). Under oxidative conditions, it may thus be part of a self-perpetuating positive feedback mechanism based on the posttranslational modification of pSMPD1. A similar positive feedback mechanism that implicates increased ceramide levels in the transcriptional up-regulation of SMPD1 activity, was previously suggested by our group (15). Macrophages show a marked increase of SMPD1 synthesis when exposed to hexanoyl-ceramide in cell culture experiments. Drobnik et al. (12) recently described an increased ceramide-to-sphingomyelin ratio in septic patients. While the altered ratio may in itself be a consequence of the elevated pSMPD1 levels, ceramide could possibly function as a stimulus for a transcriptional up-regulation of the pSMPD1 synthesis.
The actual sites of action and the effects mediated by pSMPD1 in detail still await further exploration. In view of our findings and based on various other studies, two main targets for pSMPD1 seem most plausible. First, there is considerable evidence for an extracellular sphingolipid metabolism that appears to be important in a variety of diseases. pSMPD1 represents a key enzyme in starting off the gradual breakdown of inert sphingomyelin into downstream bioactive mediators, and previous findings suggest that SMPD1 is moderately elevated in chronic disease states such as diabetes (36) and atherosclerosis (9, 30). Consistent with our findings and the concept of an extracellular sphingolipid metabolism, Drobnik (12) described elevated ratios of plasma ceramide and sphingomyelin in septic patients. This could reflect an elevated activity of pSMPD1. Furthermore, the authors describe a correlation between the increased ratio with poor outcome in their patients’ cohort. Lightle (10) provided evidence that probably both an increased pSMPD1 and an elevated hepatic de novo synthesis of ceramide may account for increased plasma ceramide levels observed in the acute phase response and endotoxic shock.
Page 8 of 21(page number not for citation purposes)

Second, elevated pSMPD1 levels in the plasma may influence the intracellular sphingolipid metabolism directly and lead to an in situ generation of intracellular ceramide. The incubation of cells with exogenous sphingomyelinases is a widely used in vitro model to illustrate the biological effects of intracellular ceramide generation. As sphingomyelin is preferentially distributed in the outer leaflet of cellular membranes (8), after inflammation triggered secretion, pSMPD1 gains unlimited access to its substrate and may thus produce huge amounts of ceramide. In this context, Garzia-Ruiz et al. affirm that there is a selective sensitivity of distinct cell types to different types of exogenously added sphingomyelinases (37). Compatible with our results indicating a decreased hepatic apoptosis rate after SMPD1 inhibition, they demonstrated that hepatocytes are selectively induced to enter apoptosis by an exogenous acid sphingomyelinase. In contrast, this effect could not be elicited by exposure to a neutral, bacterial sphingomyelinase.
A variety of mediators of the acute phase response such as endotoxin, interleukin 1β, and TNF-α induce hepatic apoptosis via early hepatic generation of ceramide (38). The progression of sepsis in BALB/c mice after intravenous application of Escherichia coli preparations resulted in a very similar extent of apoptotic cell death in liver biopsy specimens (39). Using an identical method (TUNEL staining), we found that nearly 40% of hepatic cells underwent apoptosis 8 and 24 h postinjection. Similarly, the disseminated endothelial apoptosis seen in endotoxemia and known to be implicated in the pathogenesis of organ dysfunction (40) has been attributed to the sequential action of TNF-α and ceramide (41).
Many of the deleterious effects of an activation of sphingomyelin hydrolysis are abrogated in SMPD1−/− mice, which display an improved survival rate after injection of endotoxin in lethal dosages (10). Again, this is consistent with our findings of improved long-term survival after inhibition of SMPD1 synthesis by NB6. NB6 is known to affect activities of stress-inducible kinases in human smooth muscle cells (25, 26). Furthermore, NB6 was shown to inhibit transcription of sphingomyelinase. This, however, does not rule out additional inhibitory mechanisms. Due to structural similarities to desipramine, NB6 may similarly interfere directly with enzyme-membrane interactions (42).
Inhibition of the hydrolytic activity of SMPD1 may have a therapeutic potential and can be achieved by the novel blocker NB6. In line with that, Goggel and colleagues (43) provided the first promising evidence that SMPD1 inhibition constitutes a potential therapeutic target in the treatment of PAF-induced pulmonary edema and acute lung injury. Equally, the inhibition of the sphingomyelinase pathway may be a therapeutic approach in alleviating the sphingolipid-mediated myocardial depression induced by endotoxin (44). In fact, antiapoptotic therapeutic strategies for critically ill patients targeting effectors that modulate sepsis-induced cell death have shown encouraging results (45, 46), although their impact on outcome in sepsis-related MOF in the critical care unit at present is unknown.
ACKNOWLEDGMENTS
This manuscript is dedicated to the memory of our friend and colleague Erik Beutmann (1972–2004), whose contributions in the initial phase of this project are gratefully acknowledged.
We confirm that we have no conflicting financial interests.
Page 9 of 21(page number not for citation purposes)

REFERENCES
1. Bochud, P. Y., and Calandra, T. (2003) Pathogenesis of sepsis: new concepts and implications for future treatment. BMJ 326, 262–266
2. Riedemann, N. C., Guo, R. F., and Ward, P. A. (2003) Novel strategies for the treatment of sepsis. Nat. Med. 9, 517–524
3. Papathanassoglou, E. D., Moynihan, J. A., and Ackerman, M. H. (2000) Does programmed cell death (apoptosis) play a role in the development of multiple organ dysfunction in critically ill patients? a review and a theoretical framework. Crit. Care Med. 28, 537–549
4. Hotchkiss, R. S., and Karl, I. E. (2003) The pathophysiology and treatment of sepsis. N. Engl. J. Med. 348, 138–150
5. Baumruker, T., and Prieschl, E. E. (2002) Sphingolipids and the regulation of the immune response. Semin. Immunol. 14, 57–63
6. Pettus, B. J., Chalfant, C. E., and Hannun, Y. A. (2002) Ceramide in apoptosis: an overview and current perspectives. Biochim. Biophys. Acta 1585, 114–125
7. Schissel, S. L., Keesler, G. A., Schuchman, E. H., Williams, K. J., and Tabas, I. (1998) The cellular trafficking and zinc dependence of secretory and lysosomal sphingomyelinase, two products of the acid sphingomyelinase gene. J. Biol. Chem. 273, 18250–18259
8. Goni, F. M., and Alonso, A. (2002) Sphingomyelinases: enzymology and membrane activity. FEBS Lett. 531, 38–46
9. Marathe, S., Schissel, S. L., Yellin, M. J., Beatini, N., Mintzer, R., Williams, K. J., and Tabas, I. (1998) Human vascular endothelial cells are a rich and regulatable source of secretory sphingomyelinase. Implications for early atherogenesis and ceramide-mediated cell signaling. J. Biol. Chem. 273, 4081–4088
10. Lightle, S., Tosheva, R., Lee, A., Queen-Baker, J., Boyanovsky, B., Shedlofsky, S., and Nikolova-Karakashian, M. (2003) Elevation of ceramide in serum lipoproteins during acute phase response in humans and mice: role of serine-palmitoyl transferase. Arch. Biochem. Biophys. 419, 120–128
11. Langmann, T., Buechler, C., Ries, S., Schaeffler, A., Aslanidis, C., Schuierer, M., Weiler, M., Sandhoff, K., de Jong, P. J., and Schmitz, G. (1999) Transcription factors Sp1 and AP-2 mediate induction of acid sphingomyelinase during monocytic differentiation. J. Lipid Res. 40, 870–880
12. Drobnik, W., Liebisch, G., Audebert, F. X., Frohlich, D., Gluck, T., Vogel, P., Rothe, G., and Schmitz, G. (2003) Plasma ceramide and lysophosphatidylcholine inversely correlate with mortality in sepsis patients. J. Lipid Res. 44, 754–761
Page 10 of 21(page number not for citation purposes)

13. Delogu, G., Famularo, G., Amati, F., Signore, L., Antonucci, A., Trinchieri, V., Di Marzio, L., and Cifone, M. G. (1999) Ceramide concentrations in septic patients: a possible marker of multiple organ dysfunction syndrome. Crit. Care Med. 27, 2413–2417
14. Takahashi, T., Abe, T., Sato, T., Miura, K., Takahashi, I., Yano, M., Watanabe, A., Imashuku, S., and Takada, G. (2002) Elevated sphingomyelinase and hypercytokinemia in hemophagocytic lymphohistiocytosis. J. Pediatr. Hematol. Oncol. 24, 401–404
15. Deigner, H. P., Claus, R., Bonaterra, G. A., Gehrke, C., Bibak, N., Blaess, M., Cantz, M., Metz, J., and Kinscherf, R. (2001) Ceramide induces aSMase expression: implications for oxLDL-induced apoptosis. FASEB J. 15, 807–814
16. Members of the American College of Chest Physicians/Society of Critical Care Medicine Consensus Conference Committee (1992) Definitions for sepsis and organ failure and guidelines for the use of innovative therapies in sepsis. Chest 11, 1644–1655
17. Knaus, W. A., Draper, E. A., Wagner, D. P., and Zimmerman, J. E. (1985) APACHE II: a severity of disease classification system. Crit. Care Med. 13, 818–829
18. Vincent, J. L., Moreno, R., Takala, J., Willatts, S., De Mendonca, A., Bruining, H., Reinhart, C. K., Suter, P. M., and Thijs, L. G. (1996) The SOFA (Sepsis-related Organ Failure Assessment) score to describe organ dysfunction/failure. On behalf of the Working Group on Sepsis-Related Problems of the European Society of Intensive Care Medicine. Intensive Care Med. 22, 707–710
19. Loidl, A., Claus, R., Deigner, H. P., and Hermetter, A. (2002) High-precision fluorescence assay for sphingomyelinase activity of isolated enzymes and cell lysates. J. Lipid Res. 43, 815–823
20. Bligh, E. G., and Dyer, W. J. (1959) A rapid method for total lipid extraction and purification. Can. J. Biochem. Physiol. 37, 911–917
21. Kontush, A., and Beisiegel, U. (1999) Measurement of oxidizability of blood plasma. Methods Enzymol. 299, 35–49
22. Kinscherf, R., Claus R., Wagner, M., Gehrke, C., Kamencic, H., Hou, D., Nauen, O., Schmiedt, W., Kovacs, G., Pill, J., Metz, J., and Deigner, H. P. (1998) Apoptosis caused by oxidized LDL is manganese superoxide dismutase and p53 dependent. FASEB J. 12, 461–467
23. Haberkorn, U., Altmann, A., Kamencic, H., Morr, I., Traut, U., Henze, M., Jiang, S., Metz, J., and Kinscherf, R. (2001) Glucose transport and apoptosis after gene therapy with HSV thymidine kinase. Eur. J. Nucleic Med. 28, 1690–1696
24. Haberkorn, U., Bellemann, M. E., Brix, G., Kamencic, H., Morr, I., Traut, U., Altmann, A., Doll, J., Blatter, J., and Kinscherf, R. (2001) Apoptosis and changes in glucose transport early after treatment of Morris hepatoma with gemcitabine. Eur. J. Nucleic Med. 28, 418–425
Page 11 of 21(page number not for citation purposes)

25. Loidl, A., Sevcsik, E., Riesenhuber, G., Deigner, H. P., and Hermetter, A. (2003) Oxidized phospholipids in minimally modified low density lipoprotein induce apoptotic signaling via activation of acid sphingomyelinase in arterial smooth muscle cells. J. Biol. Chem. 278, 32921–32928
26. Loidl, A., Claus, R., Ingolic, E., Deigner, H. P., and Hermetter, A. (2004) Role of ceramide in activation of stress-associated MAP kinases by minimally modified LDL in vascular smooth muscle cells. Biochim. Biophys. Acta 1690, 150–158
27. Mahidhara, R., and Billiar, T. R. (2000) Apoptosis in sepsis. Crit. Care Med. 28, 4 Suppl, N105–N113
28. Cobb, J. P., Laramie, J. M., Stormo, G. D., Morrissey, J. J., Shannon, W. D., Qiu, Y., Karl, I. E., Buchman, T. G., and Hotchkiss, R. S. (2002) Sepsis gene expression profiling: murine splenic compared with hepatic responses determined by using complementary DNA microarrays. Crit. Care Med. 30, 2711–2721
29. Qiu, H., Edmunds, T., Baker-Malcolm, J., Karey, K. P., Estes, S., Schwarz, C., Hughes, H., and Van Patten, S. M. (2003) Activation of human acid sphingomyelinase through modification or deletion of C-terminal cysteine. J. Biol. Chem. 278, 32744–32752
30. Wong, M. L., Xie, B., Beatini, N., Phu, P., Marathe, S., Johns, A., Gold, P. W., Hirsch, E., Williams, K. J., Licinio, J., and Tabas, I. (2000) Acute systemic inflammation up-regulates secretory sphingomyelinase in vivo: a possible link between inflammatory cytokines and atherogenesis. Proc. Natl. Acad. Sci. USA 97, 8681–8686
31. Hotchkiss, R. S., Tinsley, K. W., and Karl, I. E. (2003) Role of apoptotic cell death in sepsis. Scand. J. Infect. Dis. 35, 585–592
32. Cohen, J. (2002) The immunopathogenesis of sepsis. Nature 420, 885–891
33. Ayala, A., Lomas, J. L., Grutkoski, P. S., and Chung, S. (2003) Fas-ligand mediated apoptosis in severe sepsis and shock. Scand. J. Infect. Dis. 35, 593–600
34. Nakashima, I., Kato, M., Akhand, A. A., Suzuki, H., Takeda, K., Hossain, K., and Kawamoto, Y. (2002) Redox-linked signal transduction pathways for protein tyrosine kinase activation. Antioxid. Redox Signal. 4, 517–531
35. Zhang, D. X., Yi, F. X., Zou, A. P., and Li, A. L. (2002) Role of ceramide in TNF-alpha-induced impairment of endothelium-dependent vasorelaxation in coronary arteries. Am. J. Physiol. Heart Circ. Physiol. 283, H1785–H1794
36. Gorska, M., Baranczuk, E., and Dobrzyn, A. (2003) Secretory Zn2+-dependent sphingomyelinase activity in the serum of patients with type 2 diabetes is elevated. Horm. Metab. Res. 35, 506–507
Page 12 of 21(page number not for citation purposes)

37. Garcia-Ruiz, C., Colell, A., Mari, M., Morales, A., Calvo, M., Enrich, C., and Fernandez-Checa, J. C. (2003) Defective TNF-alpha-mediated hepato-cellular apoptosis and liver damage in acidic sphingomyelinase knockout mice. J. Clin. Invest. 111, 197–208
38. Jones, B. E., Lo, C. R., Srinivasan, A., Valentino, K. L., and Czaja, M. J. (1999) Ceramide induces caspase-independent apoptosis in rat hepatocytes sensitized by inhibition of RNA synthesis. Hepatology 30, 215–222
39. Yu, S. L., Chen, H. W., Yang, P. C., Peck, K., Tsai, M. H., Chen, J. J., and Lin F. Y. 2004. Differential gene expression in gram-negative and gram-positive sepsis. Am. J. Respir. Crit. Care Med. 169, 1135–1143
40. Aird, W. C. (2003) The role of the endothelium in severe sepsis and multiple organ dysfunction syndrome. Blood 101, 3765–3777
41. Haimovitz-Friedman, A., Cordon-Cardo, C., Bayoumy, S., Garzotto, M., Mc, M., Loughlin, R., Gallily, C. K., Edwards, E. H., III, Schuchman, Z., Fuks, R., et al. (1997) Lipopolysaccharide induces disseminated endothelial apoptosis requiring ceramide generation. J. Exp. Med. 186, 1831–1841
42. Kolzer, M., Werth, N., and Sandhoff, K. (2004) Interactions of acid sphingomyelinase and lipid bilayers in the presence of the tricyclic antidepressant desipramine. FEBS Lett. 559, 96–98
43. Goggel, R., S. Winoto-Morbach, G., Vielhaber, Y., Imai, K., Lindner, L., Brade, H., Brade, S., Ehlers, A. S., Slutsky, S., Schutze, E., Gulbins, S., and Uhlig, S. (2004) PAF-mediated pulmonary edema: a new role for acid sphingomyelinase and ceramide. Nat. Med. 10, 155–160
44. Favory, R., Lancel, S., Marchetti, P., Mordon, S., Chopin, C., Formstecher, P., and Neviere, R. (2004) Endotoxin-induced myocardial dysfunction: evidence for a role of sphingosine production. Crit. Care Med. 32, 495–501
45. Hotchkiss, R. S., Tinsley, K. W., Swanson, P. E., Chang, K. C., Cobb, J. P., Buchman, T. G., Korsmeyer, S. J., and Karl, I. E. (1999) Prevention of lymphocyte cell death in sepsis improves survival in mice. Proc. Natl. Acad. Sci. USA 96, 14541–14546
46. Oberholzer, C., Oberholzer, A., Clare-Salzler, M., and Moldawer, L. L. (2001) Apoptosis in sepsis: a new target for therapeutic exploration. FASEB J. 15, 879–892
Received August 2, 2004; accepted June 13, 2005.
Page 13 of 21(page number not for citation purposes)
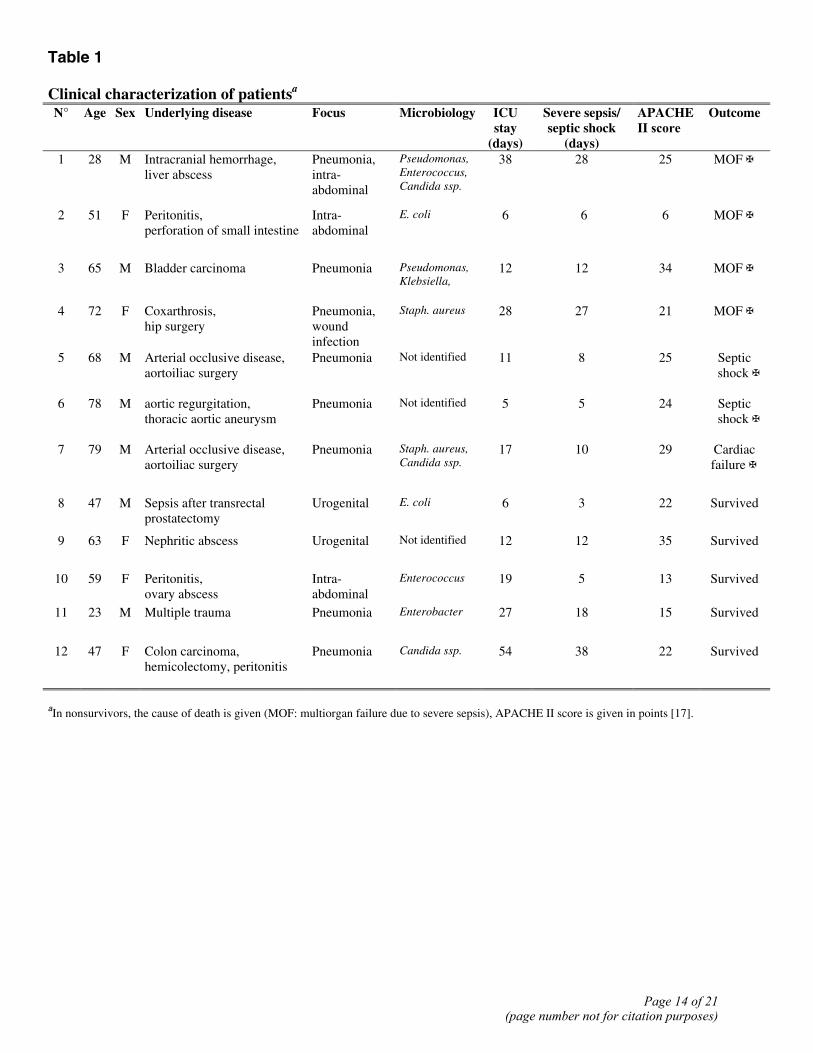
Table 1 Clinical characterization of patientsa
N° Age Sex Underlying disease Focus Microbiology ICU stay
(days)
Severe sepsis/ septic shock
(days)
APACHE II score
Outcome
1 28 M Intracranial hemorrhage, liver abscess
Pneumonia, intra-abdominal
Pseudomonas, Enterococcus, Candida ssp.
38 28 25 MOF ✠
2 51 F Peritonitis, perforation of small intestine
Intra-abdominal
E. coli
6 6 6 MOF ✠
3 65 M Bladder carcinoma Pneumonia Pseudomonas, Klebsiella,
12 12 34 MOF ✠
4 72 F Coxarthrosis, hip surgery
Pneumonia, wound infection
Staph. aureus
28 27 21 MOF ✠
5 68 M Arterial occlusive disease, aortoiliac surgery
Pneumonia Not identified 11 8 25 Septic shock ✠
6 78 M aortic regurgitation, thoracic aortic aneurysm
Pneumonia Not identified 5 5 24 Septic shock ✠
7 79 M Arterial occlusive disease, aortoiliac surgery
Pneumonia Staph. aureus, Candida ssp.
17 10 29 Cardiac failure ✠
8 47 M Sepsis after transrectal prostatectomy
Urogenital E. coli
6 3 22 Survived
9 63 F Nephritic abscess Urogenital Not identified 12 12 35 Survived
10 59 F Peritonitis, ovary abscess
Intra-abdominal
Enterococcus 19 5 13 Survived
11 23 M Multiple trauma Pneumonia Enterobacter 27 18 15 Survived
12 47 F Colon carcinoma, hemicolectomy, peritonitis
Pneumonia Candida ssp. 54 38 22 Survived
aIn nonsurvivors, the cause of death is given (MOF: multiorgan failure due to severe sepsis), APACHE II score is given in points [17].
Page 14 of 21(page number not for citation purposes)
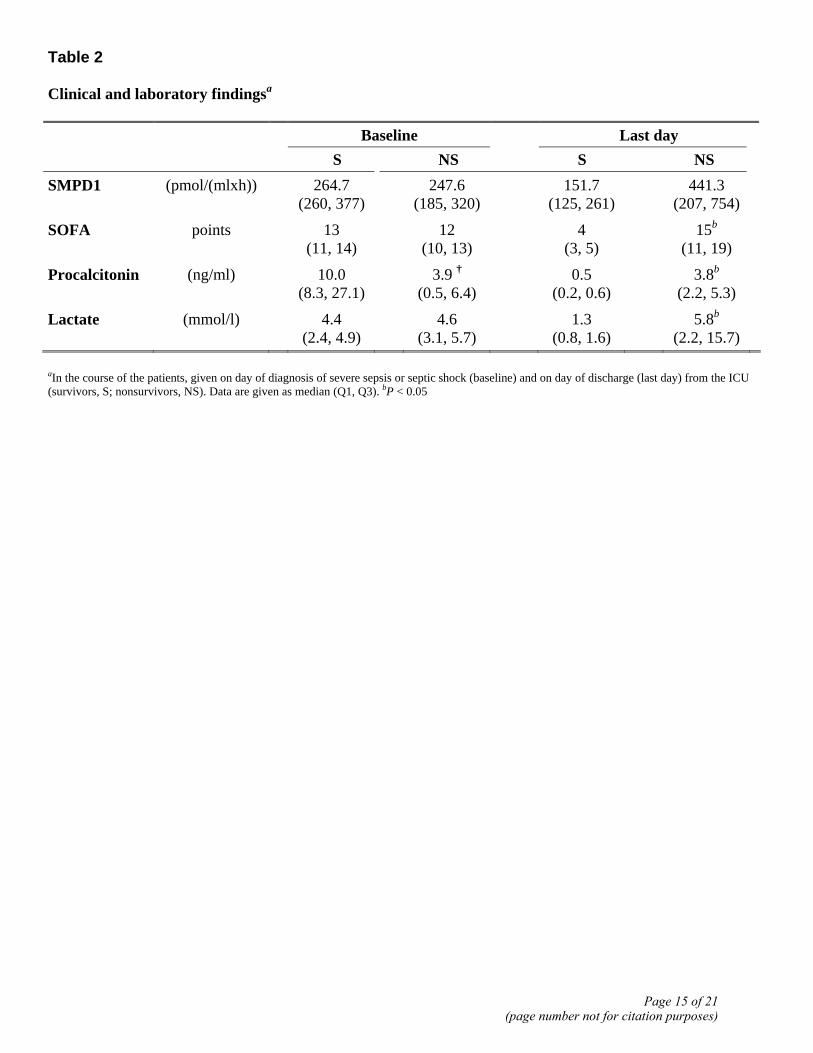
Table 2 Clinical and laboratory findingsa Baseline Last day
S NS S NS SMPD1 (pmol/(mlxh)) 264.7
(260, 377) 247.6
(185, 320) 151.7
(125, 261) 441.3
(207, 754)
SOFA points 13 (11, 14)
12 (10, 13)
4 (3, 5)
15b
(11, 19)
Procalcitonin (ng/ml) 10.0 (8.3, 27.1)
3.9 † (0.5, 6.4)
0.5 (0.2, 0.6)
3.8b
(2.2, 5.3)
Lactate (mmol/l) 4.4 (2.4, 4.9)
4.6 (3.1, 5.7)
1.3 (0.8, 1.6)
5.8b (2.2, 15.7)
aIn the course of the patients, given on day of diagnosis of severe sepsis or septic shock (baseline) and on day of discharge (last day) from the ICU (survivors, S; nonsurvivors, NS). Data are given as median (Q1, Q3). bP < 0.05
Page 15 of 21(page number not for citation purposes)
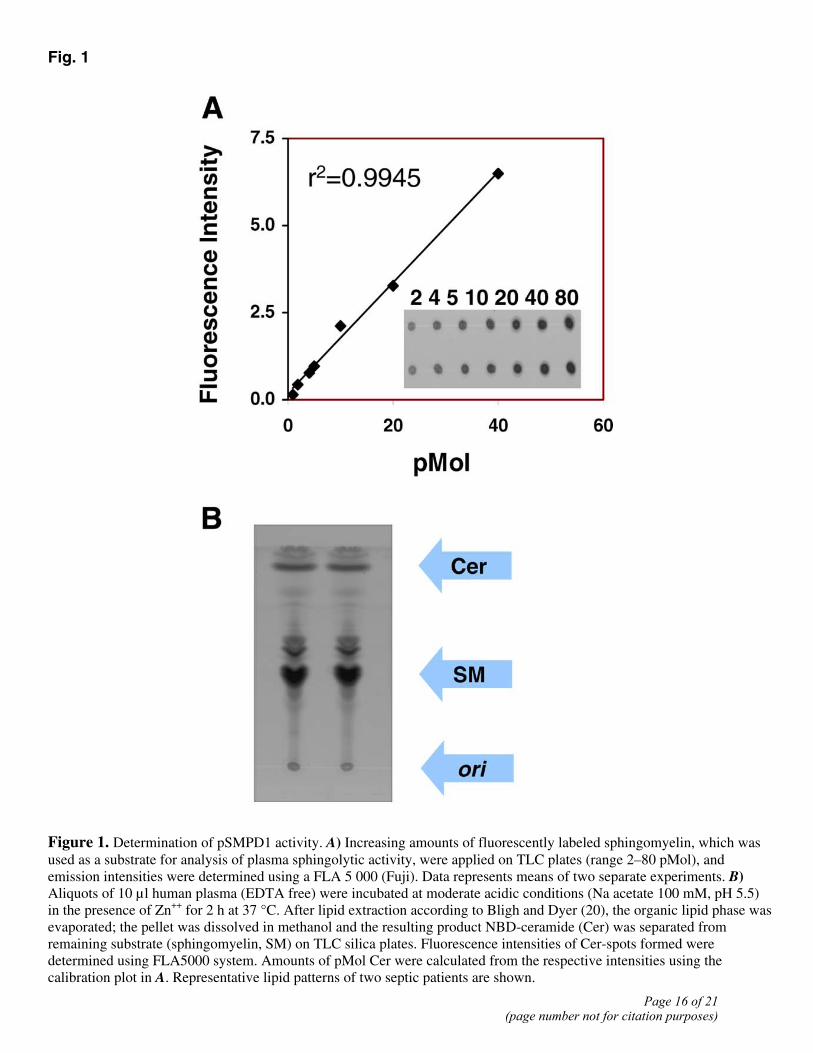
Fig. 1
Figure 1. Determination of pSMPD1 activity. A) Increasing amounts of fluorescently labeled sphingomyelin, which was used as a substrate for analysis of plasma sphingolytic activity, were applied on TLC plates (range 2–80 pMol), and emission intensities were determined using a FLA 5 000 (Fuji). Data represents means of two separate experiments. B) Aliquots of 10 µl human plasma (EDTA free) were incubated at moderate acidic conditions (Na acetate 100 mM, pH 5.5) in the presence of Zn++ for 2 h at 37 °C. After lipid extraction according to Bligh and Dyer (20), the organic lipid phase was evaporated; the pellet was dissolved in methanol and the resulting product NBD-ceramide (Cer) was separated from remaining substrate (sphingomyelin, SM) on TLC silica plates. Fluorescence intensities of Cer-spots formed were determined using FLA5000 system. Amounts of pMol Cer were calculated from the respective intensities using the calibration plot in A. Representative lipid patterns of two septic patients are shown.
Page 16 of 21(page number not for citation purposes)
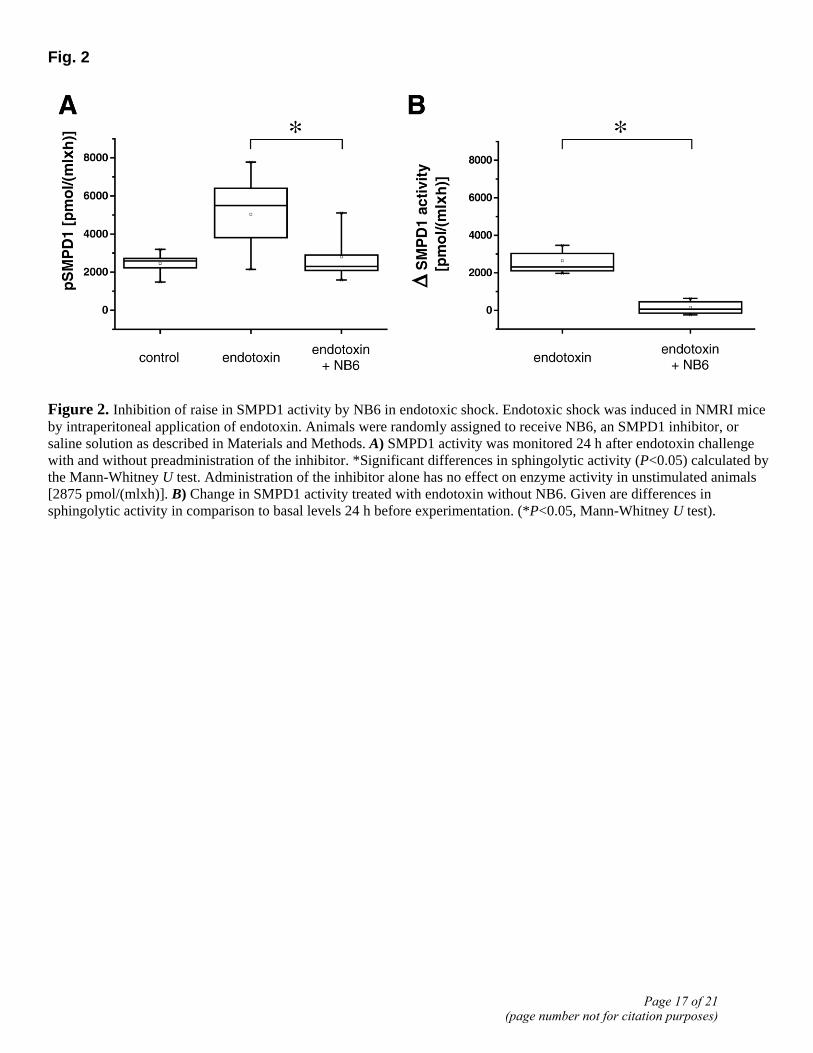
Fig. 2
Figure 2. Inhibition of raise in SMPD1 activity by NB6 in endotoxic shock. Endotoxic shock was induced in NMRI mice by intraperitoneal application of endotoxin. Animals were randomly assigned to receive NB6, an SMPD1 inhibitor, or saline solution as described in Materials and Methods. A) SMPD1 activity was monitored 24 h after endotoxin challenge with and without preadministration of the inhibitor. *Significant differences in sphingolytic activity (P<0.05) calculated by the Mann-Whitney U test. Administration of the inhibitor alone has no effect on enzyme activity in unstimulated animals [2875 pmol/(mlxh)]. B) Change in SMPD1 activity treated with endotoxin without NB6. Given are differences in sphingolytic activity in comparison to basal levels 24 h before experimentation. (*P<0.05, Mann-Whitney U test).
Page 17 of 21(page number not for citation purposes)
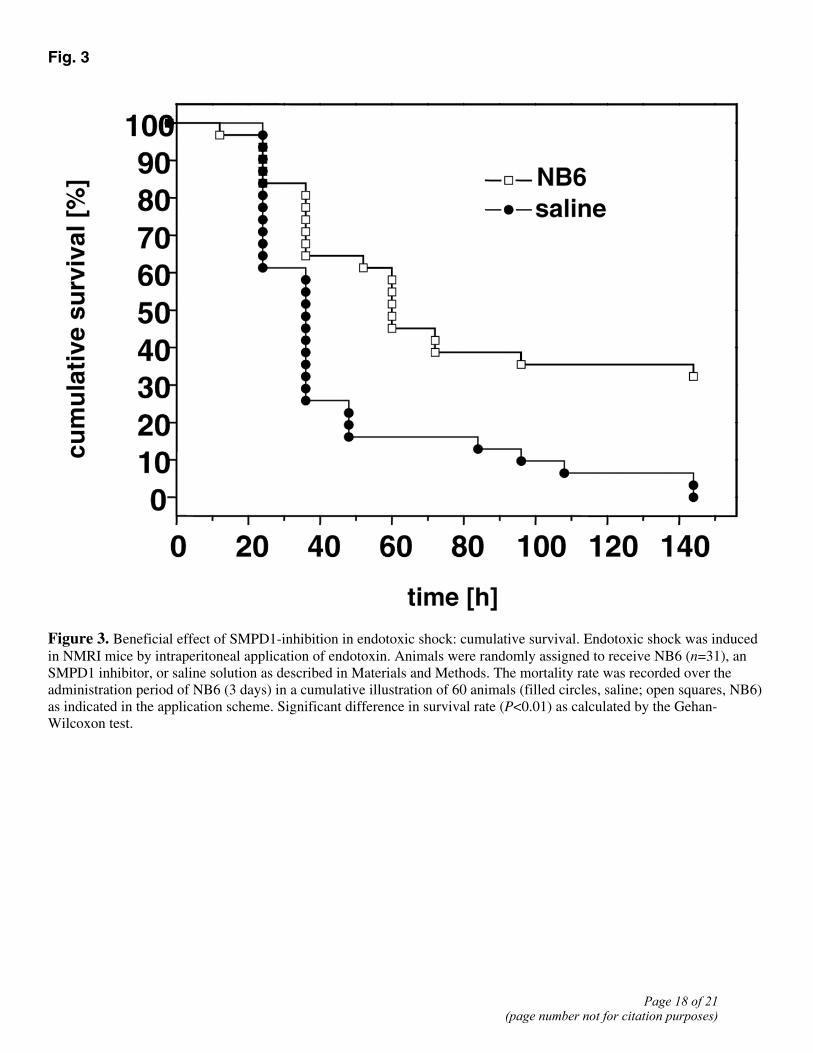
Fig. 3
Figure 3. Beneficial effect of SMPD1-inhibition in endotoxic shock: cumulative survival. Endotoxic shock was induced in NMRI mice by intraperitoneal application of endotoxin. Animals were randomly assigned to receive NB6 (n=31), an SMPD1 inhibitor, or saline solution as described in Materials and Methods. The mortality rate was recorded over the administration period of NB6 (3 days) in a cumulative illustration of 60 animals (filled circles, saline; open squares, NB6) as indicated in the application scheme. Significant difference in survival rate (P<0.01) as calculated by the Gehan-Wilcoxon test.
Page 18 of 21(page number not for citation purposes)
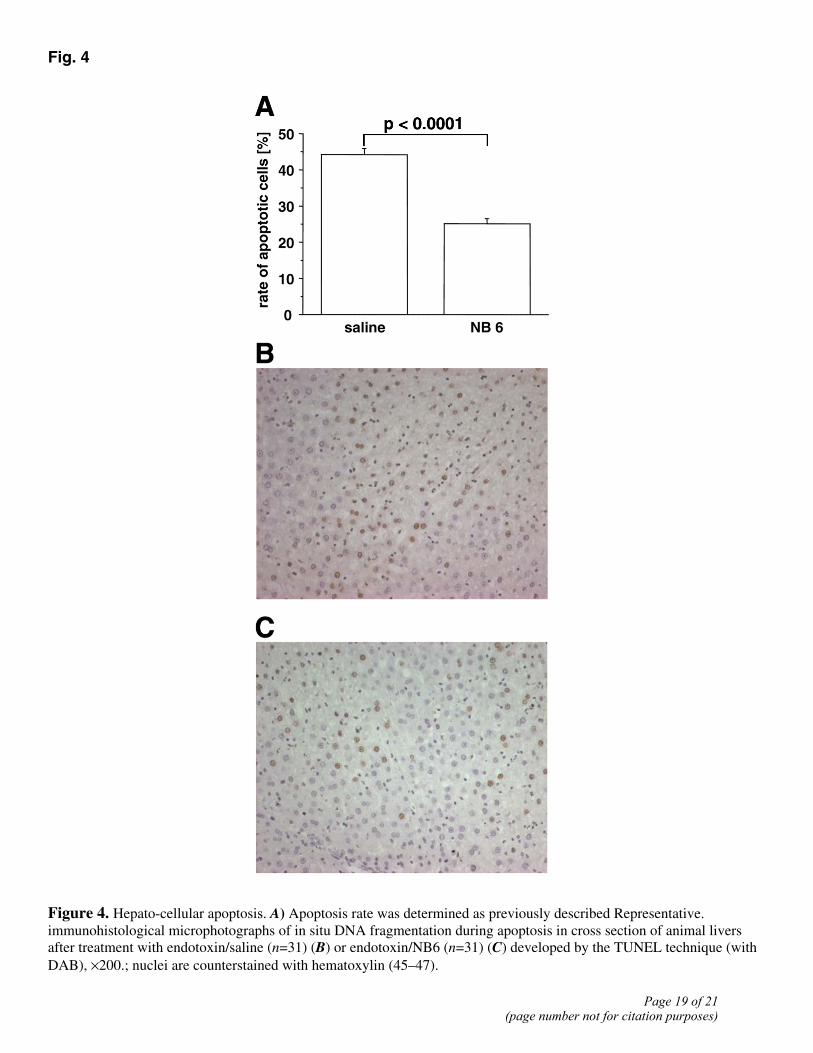
Fig. 4
Figure 4. Hepato-cellular apoptosis. A) Apoptosis rate was determined as previously described Representative. immunohistological microphotographs of in situ DNA fragmentation during apoptosis in cross section of animal livers after treatment with endotoxin/saline (n=31) (B) or endotoxin/NB6 (n=31) (C) developed by the TUNEL technique (with DAB), ×200.; nuclei are counterstained with hematoxylin (45–47).
Page 19 of 21(page number not for citation purposes)
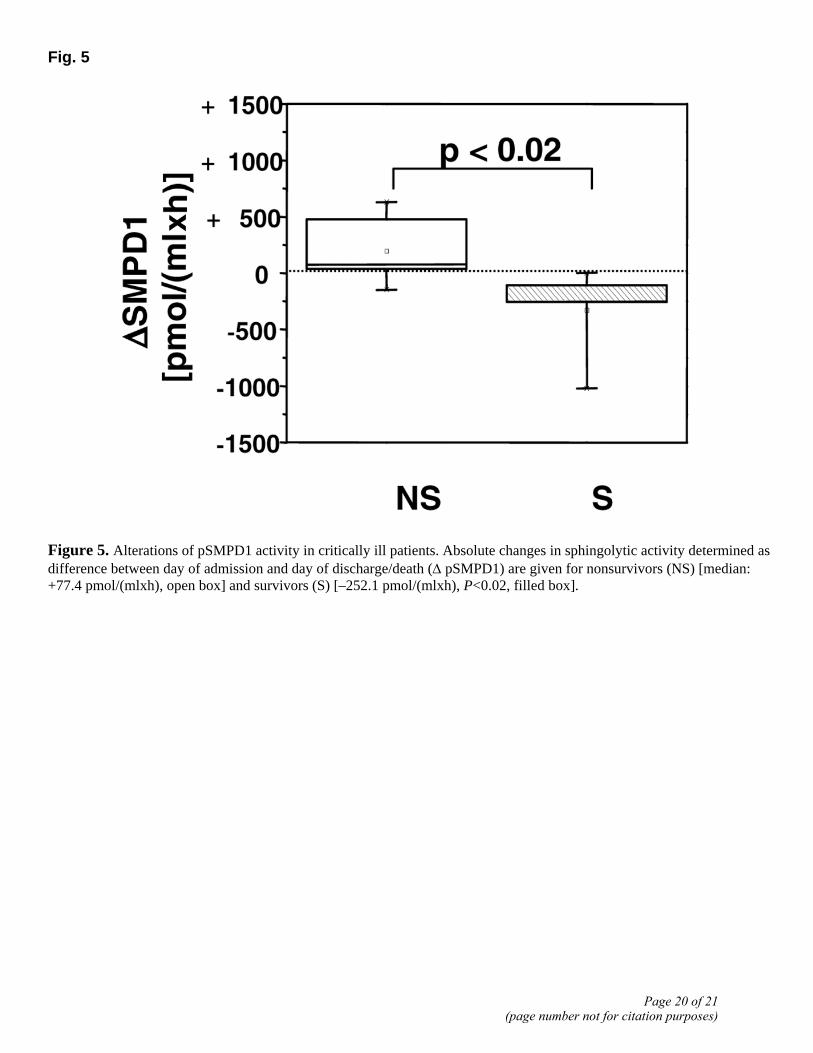
Fig. 5
Figure 5. Alterations of pSMPD1 activity in critically ill patients. Absolute changes in sphingolytic activity determined as difference between day of admission and day of discharge/death (Δ pSMPD1) are given for nonsurvivors (NS) [median: +77.4 pmol/(mlxh), open box] and survivors (S) [–252.1 pmol/(mlxh), P<0.02, filled box].
Page 20 of 21(page number not for citation purposes)
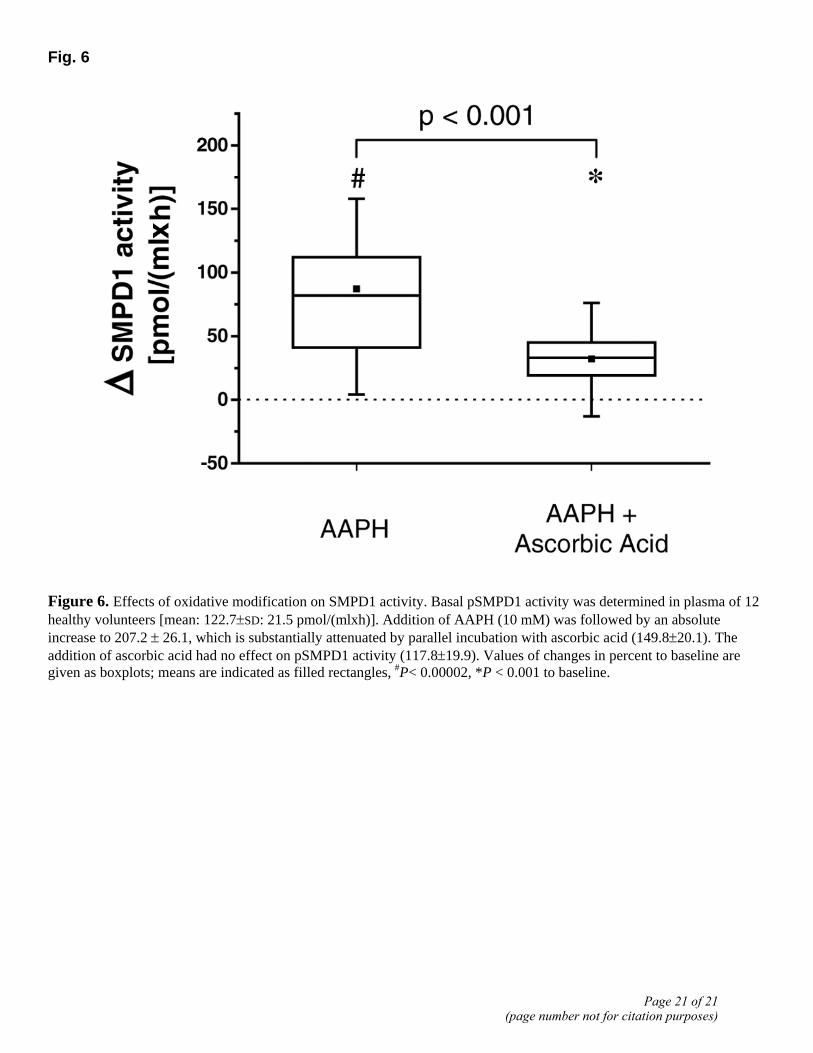
Fig. 6
Figure 6. Effects of oxidative modification on SMPD1 activity. Basal pSMPD1 activity was determined in plasma of 12 healthy volunteers [mean: 122.7±SD: 21.5 pmol/(mlxh)]. Addition of AAPH (10 mM) was followed by an absolute increase to 207.2 ± 26.1, which is substantially attenuated by parallel incubation with ascorbic acid (149.8±20.1). The addition of ascorbic acid had no effect on pSMPD1 activity (117.8±19.9). Values of changes in percent to baseline are given as boxplots; means are indicated as filled rectangles, #P< 0.00002, *P < 0.001 to baseline.
Page 21 of 21(page number not for citation purposes)