
Shortcomings of short hairpin RNA-based transgenic RNA interference in mouse oocytes
Upload
independentCategory
view
1download
0
Virology 397 (2010) 64–72
Contents lists available at ScienceDirect
Virology
j ourna l homepage: www.e lsev ie r.com/ locate /yv i ro
RNA induced polymerization of the Borna disease virus nucleoprotein
Miriam Hock a, Ina Kraus b, Guy Schoehn a,c, Marc Jamin a, Cornelia Andrei-Selmer d,Wolfgang Garten b, Winfried Weissenhorn a,⁎a Unit of Virus Host Cell Interactions (UVHCI) UMI 3265 Université Joseph Fourier-EMBL-CNRS, 6 rue Jules Horowitz 38042 Grenoble Cedex 9, Franceb Institut für Virologie, Philipps-Universität Marburg, Hans-Meerwein-Str. 2, 35043 Marburg, Germanyc Institut de Biologie Structurale Jean-Pierre Ebel, UMR 5075 CEA-CNRS-UJF, 41, rue Jules Horowitz, 38027 Grenoble Cedex 01, Franced Anatomisches Institut, Neuroanatomie, Universität Bonn, Nussallee 10, 53115 Bonn, Germany
⁎ Corresponding author. Fax: +33 476 209400.E-mail address: [email protected] (W. Weissenh
0042-6822/$ – see front matter © 2009 Elsevier Inc. Adoi:10.1016/j.virol.2009.11.016
a b s t r a c t
a r t i c l e i n f oArticle history:Received 3 August 2009Returned to author for revision29 October 2009Accepted 10 November 2009Available online 30 November 2009
Keywords:Borna disease virusNucleoproteinNucleocapsidPhosphoproteinGenomic ssRNA
The Borna disease virus (BDV) nucleoprotein (N) monomer resembles the nucleoprotein structures fromrabies virus (RABV) and vesicular stomatitis virus (VSV). We show that BDV N assembles into ring- andstring-like structures in the presence of 5′ genomic BDV RNA. RNA induced polymerization is partly RNA-specific since polymerization is inefficient in the presence of 3′ genomic BDV RNA or E. coli RNA. Mutagenesisof basic residues located in the cleft made up by the N- and C-terminal domains of N abrogate RNA-inducedpolymerization indicating that BDV N binds RNA similarly as observed in case of RABV and VSV N–RNAcomplexes. Bound RNA is not protected and sensitive to degradation. N–RNA polymers form complexes withthe phosphoprotein P as required for functional transcription or replication units. Our data indicate that BDVN utilizes similar structural principles for N–RNA and N–P–RNA complex formation as observed for relatednegative strand RNA viruses.
© 2009 Elsevier Inc. All rights reserved.
Introduction
In non-segmented negative-sense RNA viruses, the active tran-scription and replication complex, called ribonucleoprotein (RNP)complex (Murphy and Lazzarini, 1974; Szilágyi and Uryvayev, 1973)is composed of the viral RNA, the nucleoprotein N, the phosphopro-tein P and the viral polymerase L (Emerson and Wagner, 1972;Emerson and Yu, 1975; Pattnaik et al., 1992). The major RNPcomponent is N, which encapsidates the viral genome, thus formingN–RNA complexes to serve as a template for L (Emerson andWagner,1972). Because L cannot bind directly to N–RNA complexes, it isdependent on P, the polymerase cofactor (Horikami and Moyer,1995; Portner and Murti, 1986). P tethers the polymerase towardsthe N–RNA complexes, stabilizes the RNP complex and serves as ascaffold for L (Curran, 1998; Emerson and Schubert, 1987; Mellonand Emerson, 1978).
Borna disease virus (BDV) is the representative of the Bornaviridaein the orderMononegavirales. It is unique among the animal viruses ofthis order with respect to its transcription and replication in thenucleus, which provides access to the splicing machinery (Cubitt andde la Torre, 1994; Cubitt et al., 1994; Schneider et al., 1994; Tomonagaet al., 2000). BDV is noncytolytic, highly neurotropic and causesdiseases of the central nervous system (CNS) in a wide range ofvertebrates, like horses, sheep, rodents and parrots (Honkavuori et al.,
orn).
ll rights reserved.
2008; Richt and Rott, 2001; Rott and Becht, 1995). The BDVpolymerase complex consists of the nucleoprotein N, the phospho-protein P, the polymerase L and viral genomic RNA (Perez et al., 2003;Schneider et al., 2003; Yanai et al., 2006a), similar to other negativestrand RNA viruses (Albertini et al., 2008; Bourhis et al., 2006; Dolniket al., 2008).
The crystal structure of BDV N shows a homotetramer whereaseach protomer consists of two helical domains and N- and C-terminalextensions, involved in tetramer stabilization via domain exchange(Rudolph et al., 2003). An open question remained how BDV Ninteracts with RNA, although overall structural similarities withnucleoproteins from rhabdoviruses and vesiculoviruses suggestedsimilar modes of RNA interaction (Albertini et al., 2008; Luo et al.,2007). Rabies virus and VSV N are composed of two helical domainsthat bind RNA within the cleft made up by the two domains and useN- and C-terminal extensions for polymer stabilization (Albertiniet al., 2006; Green et al., 2006). BDV N does not spontaneouslyinteract with RNA like nucleoproteins from other negative strandRNA viruses, which often form ring-like or helical N–RNA structures(Albertini et al., 2007; Cox et al., 2009; Fooks et al., 1993; Green andLuo, 2006; Iseni et al., 1998; Mavrakis et al., 2002; Schoehn et al.,2001; Yang et al., 1998).
BDV genomes and antigenomes have uniformly trimmed 5′termini which might be involved in the control of virus replicationand immune evasion (Schneider et al., 2005, 2007; Habjan et al.,2008 #178). If there is a correlation between N–RNA interaction andthe trimming of the termini, however, remains to be elucidated.

Fig. 1. Molecular weight of BDV-P′ and BDV-N–P′ determined by multiangle laser lightscattering and refractometry combined with size-exclusion chromatography (MALLS).(A) BDV-P′ and (B) BDV-N–P′ SEC elution profiles revealing molecular weights of∼75 kDa for P′ and ∼235 kDa for N–P′. The dashed line shows the elution profilemonitored by excess refractive index (left ordinate axis). The black line shows themolecular mass distribution (right ordinate axis) determined by MALLS andrefractometry data. (C) Chemical cross-linking of N supports N tetramer formation insolution. Incubation of N with increasing amounts of glutaraldehyde produces newbands on SDS-PAGE and shows that the final cross-linked product of N (at 5 mMglutaraldehyde) migrates between the 150 and 250 kDa marker proteins, consistentwith tetramer formation.
65M. Hock et al. / Virology 397 (2010) 64–72
The N–RNA complex is the template for the viral polymerase. Thepolymerase L is bound to the N–RNA complex through P (P bindssimultaneously to L and to the N–RNA polymer) to position L on thetemplate. The assembly of this complex is stimulated by the non-structural BDV protein X(Poenisch et al., 2008; Schwardt et al., 2005),which is essential for viral replication (Poenisch et al., 2007). P plays arole in assembly and regulation of the polymerase complex viainteractions with X, N, L and itself (Schneider et al., 2004a;Schwemmle et al., 1998; Walker et al., 2000); its own activity isregulated by phosphorylation (Schmid et al., 2007; Schwemmle et al.,1997).
P oligomerization is required for the formation of an active poly-merase complex (Schneider et al., 2004a) similar to other negativestrand RNA polymerase complexes (Choudhary et al., 2002; Curran,1998; Möller et al., 2005). Oligomerization is likely achieved via apredicted coiled coil motif, overlapping with the interaction site of L(Schneider, 2005; Schneider et al., 2004a). Since the polymerasecomplex functions in the nucleus, P contains two nuclear localizationsignals and a nuclear export signal (Schwemmle et al., 1999; Shoyaet al., 1998; Yanai et al., 2006b) and was suggested to form dimers(Kliche et al., 1996). An N-terminally truncated isoform of P, P′ (aa 56–201) was found in BDV infected cells (Kobayashi et al., 2000);however, it does not seem to support reporter-gene expression in aviral minireplicon system (Schneider et al., 2004a). Similarly, BDV Nexists as two isoforms in BDV-infected cells, as full-length and as N-terminally truncated form (Kobayashi et al., 1998; Pyper and Gartner,1997; Schneider et al., 2004b). P requires an intact C-terminus for Ninteraction (Schwemmle et al., 1998) and contacts two different siteson N (Berg et al., 1998). P proteins from Rhabdoviruses and Sendaivirus contain two binding sites for N, one to keep N soluble and freefrom unspecific RNA and the other to bind to N–RNA forming thepolymerase complex (Curran et al., 1995; Masters and Banerjee, 1988;Mavrakis et al., 2003). However BDV N does not require P binding toprevent non-specific RNA interaction, since N oligomerizes intotetramers that do not complex RNA (Rudolph et al., 2003); thus theprecise role of N–P interaction in the absence of RNA is not known.
The aim of our study was to understand BDV N–RNA and N–Pinteractions. We show that P′ oligomerizes into tetramers. It interactswith BDV-N and the complex assembles into a hetero-octamer, withfour P′ bound to the N-tetramer. P′–N interaction requires the last fiveC-terminal P residues to form a stable complex with a KD of 1.66 μM.Tetrameric N is destabilized in the presence of 5′ genomic BDV RNAwhich leads to the formation of N–RNA polymers. Similar N–RNApolymers are formed in the presence of P′ leading to P′–N–RNApolymers. Electron microscopy analyses of N–RNA complexesrevealed irregular “open” ring-like and string-like assemblies withthe RNA exposed and accessible for degradation. The N or N–Ppolymers stayed intact after RNA degradation suggesting thatpolymerization is mainly stabilized by N–N interactions. N–RNAinteraction is mediated via recognition of basic residues within thecleft of the N- and C-terminal domains similar to the observednucleoprotein–RNA recognition of other negative strand-RNA viruses.
Results
Properties of N–P′ oligomers employed to form N–P–RNA complexes
We performed all studies presented here with an N-terminallydeleted fragment of P, P′ (Kobayashi et al., 2000) since full-lengthP has a high tendency to degrade and precipitate (data notshown). P′ and N–P′ complexes were expressed as recombinantproteins in E. coli. They eluted from a size exclusion chromato-graphy column as single peaks. The molecular weight determinedby multi-angle laser light scattering (MALLS) revealed 75.57±0.35 kDa for P′ corresponding to a tetramer (the theoretical MW ofa tetramer is 75.397 kDa) (Fig. 1A) and of 235.3±9 kDa for the N–
P′ complex consistent with a P′ tetramer interacting with an Ntetramer (calculated MW of the hetero-octamer is 233.99 kDa)(Fig. 1B). N tetramer formation was further confirmed by chemicalcross linking, which revealed intermediate states of N (migratingat ∼ 75 and ∼140 kDa) and the fully cross linked Nmigrating betweenthe 150 and 250 kDa marker proteins (Fig. 1C).
We then determined the affinity of the N–P′ interaction bysurface plasmon resonance (SPR) measurements. Binding of BDV-P′to immobilized BDV-N was performed with different analyte (P′)concentrations. The experiments showed rapid protein–proteinassociation and fast dissociation (Fig. 2A). The binding associationcurves enabled the calculation of the KD by plotting SPR values foreach injection as a function of the concentration of BDV-P′ (0.8–16 μM). A bimolecular binding model was determined based on theinteraction data and used to compute the binding constant (KD) of ∼

Fig. 2. Characterization of the N–P′ interaction. (A) Specificity of BDV-P′ binding toBDV-N plot of sensorgrams associated to injection of BDV-P (c=0–16 μM) toimmobilized BDV-N. The arrows indicate the association and dissociation phaserespectively. The dissociation constant was evaluated according to a 1:1 bindingmodel with drifting base line as 1.66 μM. (B) Decrease of interaction between BDV-P′and BDV-N due to blocking binding sites on BDV-P′ by a DEWDIIP-peptide. BDV-P′(6.4 μM) was injected over the sensor chip surface coupled with BDV-N after priorincubation at 4 °C of 30 min with the peptide (c=0–280 μM). (C) Pull down assay ofN with C-terminal P′ mutants. All pull downs were performed with Ni2+-agarosebeads and Streptactin resin simultaneously. Proteins were separated by 16% SDS-PAGE and visualized by Coomassie brilliant blue stain. The asterisk indicates thebands of the different P proteins; lanes 1 and 4 show P′; lanes 2 and 5 show P′-CΔ1and lanes 3 and 6 show P′-CΔ5. Lanes 1–3 show Ni2+-, lanes 4–6 indicate Streptactin-purified bands corresponding to His-N and strep-P′ proteins.
66 M. Hock et al. / Virology 397 (2010) 64–72
1.66 μM, fitting the experimental data with a χ2 of 1.6. Individualrate constants were determined for kon, ka to be 1.8 × 103 (1/ms)and for the koff, kd to be 3 × 10−3 (1/s). Since deletion of the C-terminal four residues of P′ abrogates N interaction (Schwemmleet al., 1998) a peptide composed of the C-terminal 7 aa was used asanalyte. Although no response signal was detected with this peptidealone (data not shown), it showed an effect in a competition assay.Incubation of the peptide with full-length BDV-P′ as analyte prior toinjection over immobilized BDV-N decreased the SPR-signals pro-portional to the increase of peptide concentration (Fig. 2B). Thisconfirmed that the C-terminal seven P′ residues are important for Ninteraction, which was further corroborated by pull-down assays.Deletion mutants of P′ lacking either 1 (P′-CΔ1) or 5 C-terminalamino acids (P′-CΔ5) were used to pull down N upon co-expression.P′ was labeled with a Strep-TagII (Strep-P′) and N was His-tagged(His-N). Full-length Strep-P′ co-eluted with His-N (Fig. 2C, lanes 1and 4) as well as Strep-P′-CΔ1 (Fig. 2C, lanes 2 and 5) but not withStrep-P′-CΔ5 using pull down via the Strep or the His tags (Fig. 2C,lanes 3 and 6). Together these results suggest that a short C-terminallinear peptide region of P is required for a low affinity interactionwith N.
N–RNA and N–P′–RNA interaction
Although RABV and VSV nucleoprotein expression in E. coli orinsect cells induces the spontaneous uptake of cellular RNA, whichresults in the formation of N–RNA polymers (Iseni et al., 1998;Mavrakis et al., 2002; Schoehn et al., 2001, 2004; Spehner et al., 1991),such a phenomenon is not observed upon recombinant BDV Nexpression (Rudolph et al., 2003). We thus prepared BDV genomicRNA fragments derived from the 5′ and the 3′ ends and incubated Nand N–P′ complexes with genomic RNA alone or in combination withincreasing concentrations of urea in order to induce N tetramerdestabilization. Both N and N–P′ bands shift to a new position in anative gel when incubated with BDV genomic 5′-trailer RNA (125nucleotides) in the absence of urea (Fig. 3A and B). In contrast no gelshifts were observed when N and N–P′ were incubated with BDVgenomic 3′-leader RNA (125nucleotides) (Fig. 3C andD; lanes 1 and2)and E. coli RNA (Figs. 3E and F). The efficiency of N–RNA andN–P′–RNAcomplex formation with 5′ genomic RNA improved with increasingurea concentrations leading to the appearance of prominent newbands in both cases (Figs. 3A and B; lanes 4, 5 and 6). Notably N andN–P′ incubation with 4 M urea did not change their mobility on a nativegel indicating that the urea treatment did not affect the oligomeriza-tion or conformation of N and N–P complexes (Figs. 3A and B; lane 7).Some complex formation was observed when N and N–P′ wereincubatedwith 3′ genomic RNA or even E. coli RNA (of variable length)under 4Murea conditions leading to the appearance of a ladder of newbands (Figs. 3E and F; lane 6). To verify the presence of P′ in the N–P′–RNA complex, the bands corresponding to N–P′ and N–P′–RNA wereexcised from a native gel, the proteins eluted from the gel and sepa-rated on SDS-PAGE. This indicated that the N–P′ complex containedP′ as expected, as well as the N–P′–RNA complex (Fig. 3G). We thencompared complex formation of 5′ trailer RNA and 5′ antigenomicRNA with N using DIG labeled RNA. This indicated N interactionwith the 5′ antigenomic RNA in the presence and absence of urea(Fig. 3H, lanes 2 and 3); however the band shift produced by 5′antigenomic RNA N-interaction are less compact than those producedby N interaction with 5′ trailer RNA (Figs. 3H, lanes 5 and 6). Theseresults indicate that encapsidation of RNA by N is more efficient in thepresence of 5′ genomic RNA. Since viral nucleoproteins need torecognize RNA in a non-sequence-specific manner, the preference for5′ genomic RNA might be explained by the formation of a hairpinstructure adopted by the 5′ RNA segment that might specificallyinteract with N. Secondly, we show that N–RNA complexes are stillable to bind to P as required for a functional N–RNApolymer present ina nucleocapsid.
Electron microscopy of N–RNA polymers
N was incubated with BDV genomic 5′-RNA and protein–RNAcomplexes were separated from RNA-free protein by glycerolgradient centrifugation producing a single band on native gelelectrophoresis (Fig. 6B; lane 2). Negatively stained electronmicroscopy images revealed open circular and rod-like structuresfor N–RNA complexes (Fig. 4A). The circular polymers have an outerdiameter of ∼10–11 nm and the rods show a length of ∼12–14 nm.The absence of the typical shape of N tetramers (Fig. 4B) in the N–RNA complex images and the width of the N–RNA rods (35±4 Å)indicated that the tetramer (∼59×59×87 Å; Rudolph et al., 2003)disassembled; this led most likely to N protomers lining up alongthe RNA chain, as demonstrated for VSV and RABV N–RNA (Albertiniet al., 2006; Green et al., 2006). The circular complexes seemed to beless compact compared to EM images of negatively stained RABVand VSV N–RNA rings (Chen et al., 2004; Schoehn et al., 2001). Theapparent lack of ring closure observed for the complexes is likelydue to the assembly of the polymer employing 5′ genomic BDV RNA.Part of the 5′ trailer RNA might adopt a hairpin conformation that is

Fig. 3. N–RNA and N–P′–RNA gel shift experiments. (A, C, E) BDV-N and (B, D, F) BDV-N–P′ incubated with BDV-5′-Trailer-(A, B), BDV-3′-Leader (C, D) and total E. coli RNA (E, F) inpresence of 0–4 M Urea, subjected to 6.5% native PAGE. (G) Presence of P′ in RNA-shift bands. Bands (marked by ⁎) corresponding to N–P′ (exN–P′) and N–P′–RNA (exN–P′–RNA)were excised from a native gel and extracted from gel slices prior to separation on a 16% SDS-PAGE together with a control of purified N–P′ complex. Bands were visualized withCoomassie Brilliant Blue. (H) Gel shift analysis of DIG-labeled 5′ trailer RNA and 5′ antigenomic RNA. The positions of free RNA and N–RNA complexes are indicated.
67M. Hock et al. / Virology 397 (2010) 64–72
recognized by N as a polymerization initiation signal and preventingring closure of the N-polymer.
BDV N sequesters RNA in a cleft between the N- and C-terminal domains
The global structural similarity of BDV N and rhabdoviridae N(Albertini et al., 2006; Green et al., 2006) suggested that the cleftbetween the N- and C-terminal domains of N harbors the RNAinteraction sites. In order to test this hypothesis, N mutants weregenerated and analyzed for their ability to form polymers in thepresence of 5′ genomic RNA. All purified N mutants elute in mono-disperse peaks from a gel filtration chromatography column andmigrate as single bands on a native gel indicating that themutations did not unfold N. The mutation of Lys164Asp, Arg165Asp,Lys242Ala and Arg297Asp were identified as critical contact pointsfor the RNA, since single mutations and double mutationsabrogated N–RNA complex formation as evidenced by native gelanalysis (Fig. 5A; lanes 1–6). A Glu161Ser mutation and thedeletion of 24 C-terminal residues had no significant effect andproduced band shifts on a native gel, indicating polymer formation(Fig. 5A; lanes 7–10). Wild type N produces a band shift under thesame conditions (Fig. 4A; lanes 11, 12). Thus, residues K164, R165,K242 and R297 from both the N-terminal and the C-terminal
domains are implicated in RNA coordination, indicating that astretch of RNA could bind within the cleft made up by the N- andC-terminal domains (Fig. 5B).
To test whether the RNA in complex with N or N–P′ issequestered by N or partly exposed, the degradation of the RNAwas analyzed. Glycerol gradient centrifugation purified N–RNA andN–P′–RNA complexes were incubated with increasing concentrationsof RNase A, divided into equal volumes and subjected to denaturingand native PAGE. This revealed that the RNA was not protected inboth BDV N– and N–P′–RNA complexes, indicated by the rapiddegradation of RNA at low RNase A concentrations (Fig. 6A).However, the positions of the protein bands did not change on thenative gel (Fig. 6B and C), suggesting that N can form stable polymersin the absence of RNA once the polymers have been formed.
RNA degradation was further confirmed by analyzing N–RNA andN–P′–RNA complexes formed with DIG-labeled BDV-5′-RNA incu-bated with 0.05 μg/ml RNase A. The higher sensitivity of this assayrevealed the appearance of two small RNA bands upon RNase Atreatment of RNA alone, N–RNA and N–P′–RNA complexes (Fig. 6D).Although we could not determine the size of the small RNAs, thebands might represent the 5′ RNA region that could adopt a hairpinstructure; it is thus most likely that this region is not directlyincorporated into the nucleoprotein RNA interaction cleft. The
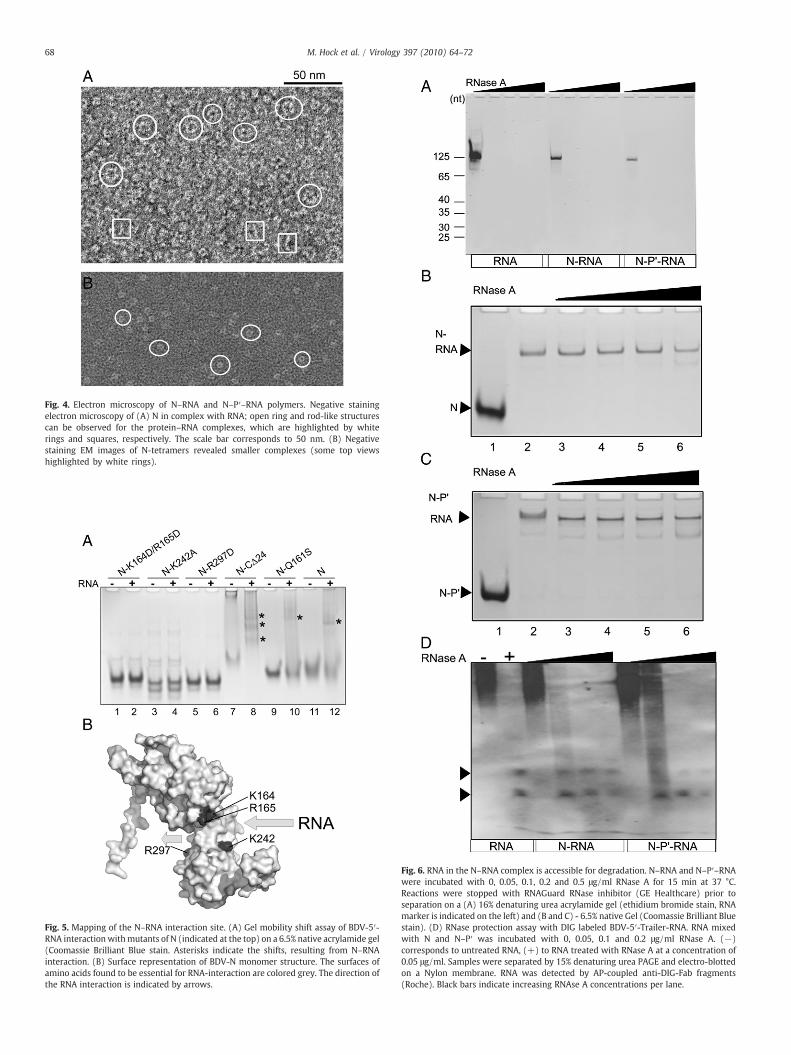
Fig. 4. Electron microscopy of N–RNA and N–P′–RNA polymers. Negative stainingelectron microscopy of (A) N in complex with RNA; open ring and rod-like structurescan be observed for the protein–RNA complexes, which are highlighted by whiterings and squares, respectively. The scale bar corresponds to 50 nm. (B) Negativestaining EM images of N-tetramers revealed smaller complexes (some top viewshighlighted by white rings).
Fig. 5. Mapping of the N–RNA interaction site. (A) Gel mobility shift assay of BDV-5′-RNA interaction withmutants of N (indicated at the top) on a 6.5% native acrylamide gel(Coomassie Brilliant Blue stain. Asterisks indicate the shifts, resulting from N–RNAinteraction. (B) Surface representation of BDV-N monomer structure. The surfaces ofamino acids found to be essential for RNA-interaction are colored grey. The direction ofthe RNA interaction is indicated by arrows.
Fig. 6. RNA in the N–RNA complex is accessible for degradation. N–RNA and N–P′–RNAwere incubated with 0, 0.05, 0.1, 0.2 and 0.5 μg/ml RNase A for 15 min at 37 °C.Reactions were stopped with RNAGuard RNase inhibitor (GE Healthcare) prior toseparation on a (A) 16% denaturing urea acrylamide gel (ethidium bromide stain, RNAmarker is indicated on the left) and (B and C) - 6.5% native Gel (Coomassie Brilliant Bluestain). (D) RNase protection assay with DIG labeled BDV-5′-Trailer-RNA. RNA mixedwith N and N–P′ was incubated with 0, 0.05, 0.1 and 0.2 μg/ml RNase A. (−)corresponds to untreated RNA, (+) to RNA treated with RNase A at a concentration of0.05 μg/ml. Samples were separated by 15% denaturing urea PAGE and electro-blottedon a Nylon membrane. RNA was detected by AP-coupled anti-DIG-Fab fragments(Roche). Black bars indicate increasing RNAse A concentrations per lane.
68 M. Hock et al. / Virology 397 (2010) 64–72

69M. Hock et al. / Virology 397 (2010) 64–72
presence of a specific tertiary RNA structure within the 5′ genomicRNA is further supported by the fact that the same small RNAfragments are produced upon RNase A treatment of the RNA alone(Fig. 6C).
Discussion
Nucleoproteins from negative strand RNA viruses such as VSV andRABV encapsidate their RNA genome and protect it from the hostileenvironment in the host cell. The interaction of N with RNA inducespolymerization of N along the RNA genome and each protomerinteracts with a short stretch of RNA in a non-sequence-specificmanner (Albertini et al., 2006; Green et al., 2006). The overall struc-tural similarity of BDV N and VSV or RABV N suggested that BDV Nmight coordinate RNA in a similar way (Albertini et al., 2008; Luo etal., 2007). We employed an assay that permitted destabilization of theN tetramers leading to the formation of N–RNA polymers in vitro. Onedimension of the polymers, their width, resembles that of nucleo-capsids present in BDV (Kohno et al., 1999). Mutagenesis of basicresidues located within the cleft made by the N- and C-terminaldomains confirmed this region as major RNA binding site. The boundRNA was sensitive to enzymatic RNA degradation suggesting that it isexposed in the N–RNA complex. This is in contrast to previous reportsthat showed that isolated BDV RNPs and virus particles are resistantagainst RNA degradation (Cubitt and de la Torre, 1994; Richt et al.,1993). This might be due to the fact that the strings of N–RNAcomplexes might be packaged tighter in the nucleocapsid due to Mthat interacts with RNA and potentially with N (Chase et al., 2007;Mayer et al., 2005; Neumann et al., 2009) or due to the presence of fulllength P in the wild type RNPs. In contrast, in vitro assembled com-plexes might constitute loose beads on a string. Another explanationfor the observed degradationmight be the partial exposure of the RNAwithin a single N molecule, leading to accessibility of the phosphatebackbone or even looping out of RNA segments within one N orbetween subsequent N molecules. Finally we cannot exclude thepossibility that aberrant RNA encapsidation in vitro, leaving multipleRNA segments available for nuclease attack, might have contributedto the RNA degradation sensitivity observed here. Furthermore, theusage of different RNases might have affected the outcome. BDV RNPswere protected from degradation by Micrococcus nuclease, but hadnot been tested for RNase A degradation before (Cubitt and de laTorre, 1994). This is consistent with RNA protection observed in caseof N–RNA complexes from RABV, whereas RNA present in nucleo-capsids was entirely protected from micrococcal nuclease degrada-tion, albeit RNase A treatment produced small RNA fragments of fourto nine nucleotides (Iseni et al., 1998). Sensitivity to RNasedegradation was further reported for RNA within influenza virusRNPs and Mumps virus N–RNA (Baudin et al., 1994; Cox et al., 2009;Duesberg, 1969) similar to our results. A requirement for RNAprotection might depend on the replication site within the cell. Bothinfluenza and Borna disease viruses replicate in the nucleus, protectedfrom the RNA degradation machinery. Additional factors such as viralmatrix proteins interacting with RNA themselves (Gomis-Rüth et al.,2003; Hoenen et al., 2005; Money et al., 2009; Neumann et al., 2009)or the formation of a compact structure of the RNPs might protectthem during the transport in the cytosol to the sites of virus assembly.In contrast viruses such as RABV and VSV might have to protect theirgenomes better since they replicate in the cytosol.
BDV N alone or in complex with P′ formed preferentially com-plexes with 5′ genomic RNA indicating that N has the capacity todiscriminate between RNA sequences, which is important for thespecific encapsidation of genomic RNA during the replication cycle.This is consistent with previous studies on RABV suggesting that the 5′leader RNA plays an important role in conferring specific RNAencapsidation (Blumberg et al., 1983; Kouznetzoff et al., 1998). Thisprocess can be further modulated by P (Yang et al., 1998) and by
phosphorylation of N (Toriumi and Kawai, 2004). The in vitrodegradation of a 5′ genomic RNA fragment revealed that two smallregions are protected from complete degradation by RNase A,independent of the presence of N or N–P′. This suggests that part ofthe 5′ genomic RNA used in the study adopts a secondary or tertiarystructure that is insensitive to RNase A degradation. We hypothesizethat this is most likely the extreme 5′ end. A defined RNA structurecould be recognized by N and thus serve as a starting signal toencapsidate RNA in a sequence unspecific manner as observed forRABV and VSV N–RNA interactions. Furthermore, RNA recognition bythe N tetramer might help to destabilize the tetramer thus exposingthe RNA binding sites. This hypothesis is supported by the fact that Ntetramers do not interact with 3′ genomic RNA or E. coli RNA in theabsence of urea as a destabilizing agent. Even in the presence of urea,neither N nor N–P tetramers are efficiently destabilized by 3′ genomicRNA or by E. coli RNA when compared to the effect of 5′ genomic RNAand to some extend of the 5′ antigenomic RNA. Since the 3′ end of thegenome is complementary to the 5′ region, it would be expected thatthe 5′ antigenome might adopt a similar conformation that permits Ninteraction, as we observe in the current study. The EM images of N–RNA complexes do not reveal closed ring-like structures as observedfor other negative strand RNA viruses (Iseni et al., 1998; Maclellanet al., 2007; Schoehn et al., 2001). This is most likely due to thepresence of the 5′ end of the RNA genome that might adopt a definedstructure that itself is not incorporated into the RNA binding cleft of N.Our data also suggest that the N tetramers have to disassemble uponRNA interaction. Firstly, the appearances of the EM images suggest theabsence of N tetramers as building blocks of the polymers. Secondly,important basic residues required for RNA binding were mapped tothe cleft that itself is sequestered in the interior of the N tetramer(Rudolph et al., 2003).
Depletion of the RNA from the N–RNA and N–P′–RNA polymersindicate that polymerization is mainly stabilized via N–N proteininteractions as shown for empty VSV ring-like nucleocapsids (Zhanget al., 2008). This suggests that polymerization is also most likelysupported by domain exchange between protomers within thepolymer similar to RABV and VSV N–RNA polymerization (Albertiniet al., 2006; Green et al., 2006).
The N–RNA complex constitutes the target for the polymerasecomplex composed of L and the phosphoprotein P. P regulates trans-cription and replication by linking the N–RNA complex to L(Schneider et al., 2004a). Although it has been speculated that Pforms dimers, trimers or tetramers based on gel filtration results(Schneider, 2005; Schneider et al., 2004a) our analysis clearly showsthat P forms tetramers in solution similar to P from Sendai virus(Tarbouriech et al., 2000b). Although Cys 125 of BDV-P was impli-cated previously in disulfide cross-linked dimerization (Kliche et al.,1996) MALLS analysis of N–P complexes produced the same results ina reducing buffer (data not shown), indicating that Cys cross linking isnot required for oligomerization. Our results suggest further thattetramer formation of P does not depend on phosphorylation asshown for Ps from Rhabdoviruses (Gerard et al., 2007; Gigant et al.,2000).
We show further that P interaction with N requires a shortC-terminal region of P including the last seven amino acids as pre-viously reported (Schneider, 2005; Schwemmle et al., 1998). Wedemonstrate that this region binds N with a micromolar affinitysimilar to other N–P interactions; the C-terminal nucleocapsid-binding domain of measles virus P binds the nucleoprotein NTAIL
peptide with an affinity constant of 13 μM (Kingston et al., 2004)and the Sendai virus NTAIL–PX interaction has an affinity of ∼57 μM(Houben et al., 2007). A low binding affinity is consistent with amodel proposed for Sendai virus, whereas P is “cart wheeling” alongthe N–RNA during RNA-synthesis, by employing on-and-off interac-tions (Kolakofsky et al., 2004). Therefore a low binding affinitybetween P and N proteins is energetically beneficial for movement of

70 M. Hock et al. / Virology 397 (2010) 64–72
the L protein along the nucleocapsid. Since P proteins from negativestrand RNA viruses adopt a modular structure (Gerard et al., 2009;Karlin et al., 2003; Tarbouriech et al., 2000a) whereas the singledomains are flexibly linked, BDV P might be similarly organized byutilizing a C-terminal domain for interaction with N tetramers aswell as N–RNA polymers. The short N interaction site might furtherensure that multiple Ps can simultaneously interact with at least fourNs present in N–RNA polymers. In summary our study provides afirst model of the RNA interaction mode of BDV N and suggests thatsuch N–RNA polymers could be targets for low affinity interactionwith tetrameric P and thus provide a link to the polymerase L duringreplication and transcription.
Materials and methods
Protein expression and purification
BDV-N (370 aa) and BDV-N mutants 1–346 and 43–370 werecloned into the pCDFDuet-1 vector (Novagen), containing an N-terminal His-Tag. 1–346 comprises aa 1–346 and 43–370 comprisesaa 43–370 of BDV N. Single and double aa substitution mutants N-K164D/R165D, N-K242A and N-R297D were cloned via site directedmutagenesis according to (Imai et al., 1991) with pCDFDuet-1containing full-length N as a template. K164D/R165D correspondsto Lys 164 and Arg 165 exchanged against an Asp each. K242Acorresponds to Lys 242 changed to Ala, R297D contains an Asp insteadof Arg at position 297. Gln 161 is substituted by Ser in Q161S. Proteinswere expressed in E. coli BL21 (DE3)-Gold cells. Cells were harvestedby centrifugation, resuspended in buffer 1 (20 mM Hepes pH 7.8;100 mM NaCl) and lysed by sonication in presence of proteaseinhibitors (Complete EDTA-free protease inhibitors, Roche). Thecleared supernatant was loaded onto a Ni2+-charged ChelatingSepharose (GE Healthcare) column. The column was washed in buffer1 plus 1 MNaCl and 1M KCl followed by extensive washes with buffer1 plus 50 mM Imidazole, pH 7.8. Proteins were eluted with Buffer 1plus 500 mM imidazole, pH 7.8. The His-tag was cleaved off overnightat 4 °C with TEV protease (1:100 w/w) in presence of 1 mM DTT and1 mM EDTA, pH 8.0. The tag was removed by a second Ni2+-ChelatingSepharose column after dialysis into buffer 1. Further purification wascarried out via a MonoQ anion exchange column (GE Healthcare).Proteins were eluted by performing a salt gradient with buffer 1 plus1 M NaCl.
A Superdex 200 size-exclusion column (GE Healthcare) was usedas a final purification step in buffer 1. Protein concentration wasdetermined by absorbance at 280 nm under denaturing conditions(6 M guanidinium hydrochloride).
BDV-N–P′ was co-expressed from pCDFDuet-1, but P′ remaineduntagged in order to co-elute with His-tagged N. Expression andpurification was performed as described for BDV-N. BDV-P′ (146 aa,from aa 56–201) and C-terminal deletion P′ mutants P′-CΔ1 and P′-CΔ5 were cloned into pASK45-plus vector (IBA), containing an N-terminal StrepTagII. P′-CΔ1 comprises aa 56–200 and P′-CΔ5corresponds to aa 56–196. Proteins were expressed in E. coli BL21(DE3)-Gold cells. Production of cleared lysates was identical withthat of BDV-N. Lysates were passed over a Streptactin sepharosecolumn (IBA) and washed with buffer 1, followed by buffer 1 plus1 M NaCl and 1 M KCl and buffer 1 plus 10 mM ATP, 10 mM MgCl2.Elution was performed with buffer 1 plus 10 mM Desthiobiotin.Further purification steps were performed as described for BDV-N.BDV-N, N–P′, P′ and P′ mutants were cloned by standard PCRmethods, N mutants were cloned according to Imai et al. (1991). Allconstructs were sequenced.
Size exclusion chromatography and multi-angle laser scattering.Size exclusion chromatography was performed with a Shodex ProteinKW-804 HPLC column (300×8.0 mm) (Lata et al., 2008). Briefly, thecolumn was equilibrated in buffer 1 and proteins were separated at
25 °C with a flow rate of 0.8 ml/min. On-line detection was performedby multi-angle laser light scattering (MALLS) using a DAWN-EOSdetector (Wyatt Technology Corp., Santa Barbara, CA) equipped witha laser emitting at 690 nm and by refractive indexmeasurement usinga RI2000 detector (Schambeck SFD). Light scattering intensities weremeasured at different angles relative to the incident beam, andanalysis of the data was performed with the ASTRA software (WyattTechnology Corp.). Weight-averaged (Mw) molecular weights wereobtained from the molecular weight distribution across the elutionpeak. For both P′ and N–P′ complexes the molecular mass wasconstant across the elution peak. The polydispersity factor (MW/MN)equaled 1.00±0.02 for P′ and 1.00±0.026 for N–P′ indicating mono-disperse species in both cases. For regular size exclusion chromato-graphy (SEC), stokes radii were determined by calibrating the columnwith globular proteins of known stokes radii.
Surface plasmon resonance (SPR) measurements of the N–P′ interaction
To study the direct interaction between BDV-N and BDV-P′ weused surface plasmon resonance (SPR) technique on a Biacore 2000instrument and reagents from Biacore AB (Germany). Immobilizationof the ligand (BDV-N) was performed via amino coupling on a CM5sensor chip. BDV-N (treated with 10 mM acetate buffer, pH 4.5) wasinjected over a flow cell (activated with EDC/NHS) and immobilizedto 8800 response units (RU). The flow cell was then deactivated withethanol amine (EA). Another flow cell, activated with EDC/NHS anddeactivated with EA was used as a reference cell. Immobilization andinteraction analysis were performed at 25 °C, using HBS as runningbuffer. Aliquots of 60 μl of BDV-P′ (0.8–16 μM) were injected at a flowrate of 20 μl/min over sensor chip surfaces. The signal in the referencecell was subtracted online during all measurements. Alternative,aliquots of 25 μl BDV-P′ (6.4 μM) incubated with a DEWDIIP-peptide(0–280 μM) were injected over sensor chip. Furthermore 6.4 μM ofeach BDV-P mutant was injected over sensor chip. Evaluation ofbinding specificity and kinetics of interaction were performed usingthe BIAevaluation 3.2RC1 software (Biacore).
RNA transcription and purification
BDV-3′-Leader, 5′-Trailer RNA and 5′ antigenomic RNA (BDV He/80/FR strain genome, GenBank AJ311522; nucleotides 1–125, 8784–8909 and 1–125 (antigenome) respectively) were cloned into pGem-9zf (Novagen) and in-vitro transcribed after cleavage of the plasmidwith NsiI (3′-RNA) and XbaI (5′-RNA). The genome sequence entry inthe NCBI nucleotide database for BDV He/80/FR corresponds to thegenome as described (Pleschka et al., 2001), lacking two nucleotidesat the 5′ trailer for entire complementarity with the 3′ leader andends with nucleotide 8909. Transcription reaction was carried out for3–4 h at 37 °C, modified from Price et al. (1995) with 40 mM Tris, pH8.0, 1 mM Spermidine, 5 mM DTT, 0.01% Triton-X, 0.1 mg T7-Polymerase (EMBL, Heidelberg), 4 mM NTPs each, 25 mM MgCl2 and1–2 mg template DNA. RNA purification by denaturing PAGE wasperformed according to Price et al. (1995). Alternatively, BDV-5′-Trailer-RNA and antigenomic RNA were transcribed and DIG body-labeled from pGem-9zf, “DIG Northern starter kit” (Roche) accordingto the manufacturer's protocol.
N–RNA and N–P′–RNA interaction
SEC purified N and N–P′ were incubated overnight at 4 °C withBDV-3′-Leader-, 5′-Trailer- and E. coli RNA (Sigma) at a molar ratio of1:2 respectively, in buffer 3 (20 mM Hepes, 20 mM NaCl, pH 7.8) andin absence and presence of urea (1, 2, 3 and 4 M). The effect of urea onN and N–P′ alone was tested by incubation with 4 M urea. Sampleswere analyzed by 6.5% native PAGE and stained with CoomassieBrilliant Blue after verifying RNA presence by Methylene Blue

71M. Hock et al. / Virology 397 (2010) 64–72
staining. Furthermore, N, N–P′ and P′were incubated overnight at 4 °Cwith BDV-5′-Trailer RNA (125 nucleotides) at a molar ratio of 1:2respectively, to verify that P′ does not interact with RNA (data notshown). Samples were analyzed by 6.5% native PAGE and stainedwithCoomassie Brilliant Blue after verifying RNA presence by MethyleneBlue stain. The same experiments were performed with N carryingspecific point mutations.
Electron microscopy
BDV-N was purified by SEC and analyzed by electron microscopyusing negative staining with sodium silicotungstate (pH 7.0). Thegrids were observed using a JEOL 2010 FEG electron microscopeworking at 100 kV with a nominal magnification of 40.000(Schoehn et al., 2008). Gel filtration purified N was incubated withBDV-5′-Trailer-RNA (125 nucleotides) in buffer 3 and purified byglycerol gradient centrifugation. The gradient was continuous from45% glycerol at the bottom to 25% at the top. Glycerol was bufferedwith buffer 3. Samples were spun for 16 h at 4 °C and 40,000 rpm inan ultracentrifuge (Beckman). Samples present in the bottomfractions were collected and analyzed by electron microscopy.
RNAse protection assay
N–RNA and N–P′–RNA were purified as described above forelectron microscopy analyses. Complexes were incubated for 15 minat 37° with 0.05, 0.1, 0.2 and 0.5 μg/ml RNase A (Roche). Controlswere performed with buffer 3 instead of RNase A. After incubationsamples were treated with RNAGuard RNase inhibitor (GE Health-care) and stored on ice prior to further treatment. One half was boiledwith 8 M Urea before subjection to a 16% denaturing polyacrylamide-urea gel. The gel was stained with ethidium bromide. The other halfwas mixed with a 4× native PAGE sample buffer and samples wereseparated by 6.5% native PAGE. The bands were visualized withCoomassie Brilliant Blue staining.
RNase protection assay with DIG-labeled RNA
DIG-labeled BDV-5′-RNA and BDV-5′-antigenomic RNA (125nucleotides) were incubated with N, N–P′ or buffer 3 as a control.N–RNA and N–P′–RNA were incubated for 15 min at 37 °C with 0.05,0.1 and 0.2 μg/ml RNase A. After incubation samples were treatedwith RNAGuard RNase inhibitor (GE Healthcare) and subjected to 18%denaturing urea PAGE. Samples were electro-blotted to a nylonmembrane (Sigma) according to Günzl et al. (2002). Band detectionwas performed with alkaline phosphatase-conjugated anti-DIG-Fab-fragments (DIG northern starter kit, Roche), following the manufac-turer's protocol for direct detection.
Acknowledgments
This work was supported by SFB 593 (Marburg) and SFB 535(Giessen) of the Deutsche Forschungsgemeinschaft (W.G.).We thankDr. T. Crepin for help with RNA synthesis and appreciate the access tothe platforms provided by the Partnership for Structural Biology (PSB,Grenoble).
References
Albertini, A.A.V., Wernimont, A.K., Muziol, T., Ravelli, R.B.G., Clapier, C.R., Schoehn, G.,Weissenhorn, W., Ruigrok, R.W.H., 2006. Crystal structure of the rabies virusnucleoprotein–RNA complex. Science (New York, N.Y.) 313 (5785), 360–363.
Albertini, A.A.V., Clapier, C.R., Wernimont, A.K., Schoehn, G., Weissenhorn, W.,Ruigrok, R.W.H., 2007. Isolation and crystallization of a unique size category ofrecombinant Rabies virus Nucleoprotein–RNA rings. J. Struct. Biol. 158 (1),129–133.
Albertini, A.A.V., Schoehn, G., Weissenhorn, W., Ruigrok, R.W.H., 2008. Structuralaspects of rabies virus replication. Cell. Mol. Life Sci.: CMLS 65 (2), 282–294.
Baudin, F., Bach, C., Cusack, S., Ruigrok, R.W., 1994. Structure of influenza virus RNP: I.Influenza virus nucleoprotein melts secondary structure in panhandle RNA andexposes the bases to the solvent. EMBO J. 13 (13), 3158–3165.
Berg, M., Ehrenborg, C., Blomberg, J., Pipkorn, R., Berg, A.L., 1998. Two domains of theBorna disease virus p40 protein are required for interaction with the p23 protein.J. Gen. Virol. 79 (Pt. 12), 2957–2963.
Blumberg, B.M., Giorgi, C., Kolakofsky, D., 1983. N protein of vesicular stomatitis virusselectively encapsidates leader RNA in vitro. Cell 32 (2), 559–567.
Bourhis, J.-M., Canard, B., Longhi, S., 2006. Structural disorder within the replicativecomplex of measles virus: functional implications. Virology 344 (1), 94–110.
Chase, G., Mayer, D., Hildebrand, A., Frank, R., Hayashi, Y., Tomonaga, K., Schwemmle,M., 2007. Borna disease virus matrix protein is an integral component of the viralribonucleoprotein complex that does not interfere with polymerase activity.J. Virol. 81 (2), 743–749.
Chen, Z., Green, T.J., Luo, M., Li, H., 2004. Visualizing the RNAmolecule in the bacteriallyexpressed vesicular stomatitis virus nucleoprotein–RNA complex. Structure(London, England: 1993) 12 (2), 227–235.
Choudhary, S.K., Malur, A.G., Huo, Y., De, B.P., Banerjee, A.K., 2002. Characterization ofthe oligomerization domain of the phosphoprotein of human parainfluenza virustype 3. Virology 302 (2), 373–382.
Cox, R., Green, T.J., Qiu, S., Kang, J., Tsao, J., Prevelige, P.E., He, B., Luo, M., 2009.Characterization of a mumps virus nucleocapsid-like particle. J. Virol. 83 (21),11402–11406.
Cubitt, B., de la Torre, J.C., 1994. Borna disease virus (BDV), a nonsegmented RNA virus,replicates in the nuclei of infected cells where infectious BDV ribonucleoproteinsare present. J. Virol. 68 (3), 1371–1381.
Cubitt, B., Oldstone, C., Valcarcel, J., Carlos de la Torre, J, 1994. RNA splicing contributesto the generation of mature mRNAs of Borna disease virus, a non-segmentednegative strand RNA virus. Virus Res. 34 (1), 69–79.
Curran, J., 1998. A role for the Sendai virus P protein trimer in RNA synthesis. J. Virol. 72(5), 4274–4280.
Curran, J., Marq, J.B., Kolakofsky, D., 1995. An N-terminal domain of the Sendaiparamyxovirus P protein acts as a chaperone for the NP protein during the nascentchain assembly step of genome replication. J. Virol. 69 (2), 849–855.
Dolnik, O., Kolesnikova, L., Becker, S., 2008. Filoviruses: interactions with the host cell.Cell. Mol. Life Sci.: CMLS 65 (5), 756–776.
Duesberg, P.H., 1969. Distinct subunits of the ribonucleoprotein of influenza virus.J. Mol. Biol. 42 (3), 485–499.
Emerson, S.U., Schubert, M., 1987. Location of the binding domains for the RNApolymerase L and the ribonucleocapsid template within different halves of the NSphosphoprotein of vesicular stomatitis virus. Proc. Natl. Acad. Sci. U. S. A. 84 (16),5655–5659.
Emerson, S.U.,Wagner, R.R., 1972. Dissociation and reconstitution of the transcriptase andtemplate activities of vesicular stomatitis B and T virions. J. Virol. 10 (2), 297–309.
Emerson, S.U., Yu, Y., 1975. Both NS and L proteins are required for in vitro RNAsynthesis by vesicular stomatitis virus. J. Virol. 15 (6), 1348–1356.
Fooks, A.R., Stephenson, J.R., Warnes, A., Dowsett, A.B., Rima, B.K., Wilkinson, G.W.,1993. Measles virus nucleocapsid protein expressed in insect cells assembles intonucleocapsid-like structures. J. Gen. Virol. 74 (Pt. 7), 1439–1444.
Gerard, F.C.A., Ribeiro, E. d. A., Albertini, A.A.V., Gutsche, I., Zaccai, G., Ruigrok, R.W.H.,Jamin, M., 2007. Unphosphorylated rhabdoviridae phosphoproteins form elongat-ed dimers in solution. Biochemistry 46 (36), 10328–10338.
Gerard, F.C.A., Ribeiro, E. d. A., Leyrat, C., Ivanov, I., Blondel, D., Longhi, S., Ruigrok,R.W.H., Jamin, M., 2009. Modular organization of rabies virus phosphoprotein.J. Mol. Biol. 388 (5), 978–996.
Gigant, B., Iseni, F., Gaudin, Y., Knossow, M., Blondel, D., 2000. Neither phosphorylationnor the amino-terminal part of rabies virus phosphoprotein is required for itsoligomerization. J. Gen. Virol. 81 (Pt. 7), 1757–1761.
Gomis-Rüth, F.X., Dessen, A., Timmins, J., Bracher, A., Kolesnikowa, L., Becker, S., Klenk,H.D., Weissenhorn, W., 2003. The matrix protein VP40 from Ebola virusoctamerizes into pore-like structures with specific RNA binding properties.Structure (London, England: 1993) 11 (4), 423–433.
Green, T.J., Luo, M., 2006. Resolution improvement of X-ray diffraction data of crystals ofa vesicular stomatitis virus nucleocapsid protein oligomer complexed with RNA.Acta Crystallogr., D Biol. Crystallogr. 62 (Pt. 5), 498–504.
Green, T.J., Zhang, X., Wertz, G.W., Luo, M., 2006. Structure of the vesicular stomatitisvirus nucleoprotein–RNA complex. Science (New York, N.Y.) 313 (5785), 357–360.
Günzl, A., Palfi, Z., Bindereif, A., 2002. Analysis of RNA–protein complexes byoligonucleotide-targeted RNase H digestion. Methods (San Diego, Calif.) 26 (2),162–169.
Habjan, M., Andersson, I., Klingström, J., Schümann, M., Martin, A., Zimmermann, P.,Wagner, V., Pichlmair, A., Schneider, U., Mühlberger, E., Mirazimi, A., Weber, F.,2008. Processing of genome 5' termini as a strategy of negative-strand RNA virusesto avoid RIG-I-dependent interferon induction. PLoS One 3 (4), e2032.
Hoenen, T., Volchkov, V., Kolesnikova, L., Mittler, E., Timmins, J., Ottmann, M., Reynard,O., Becker, S., Weissenhorn, W., 2005. VP40 octamers are essential for Ebola virusreplication. J. Virol. 79 (3), 1898–1905.
Honkavuori, K.S., Shivaprasad, H.L., Williams, B.L., Quan, P.L., Hornig, M., Street, C.,Palacios, G., Hutchison, S.K., Franca, M., Egholm, M., Briese, T., Lipkin, W.I., 2008.Novel Borna virus in psittacine birds with proventricular dilatation disease. Emerg.Infect. Dis. 14 (12), 1883–1886.
Horikami, S.M., Moyer, S.A., 1995. Alternative amino acids at a single site in the Sendaivirus L protein produce multiple defects in RNA synthesis in vitro. Virology 211 (2),577–582.
Houben, K., Marion, D., Tarbouriech, N., Ruigrok, R.W.H., Blanchard, L., 2007. Interactionof the C-terminal domains of Sendai virus N and P proteins: comparison of

72 M. Hock et al. / Virology 397 (2010) 64–72
polymerase–nucleocapsid interactions within the paramyxovirus family. J. Virol. 81(13), 6807–6816.
Imai, Y., Matsushima, Y., Sugimura, T., Terada, M., 1991. A simple and rapid method forgenerating a deletion by PCR. Nucleic Acids Res. 19 (10), 2785.
Iseni, F., Barge, A., Baudin, F., Blondel, D., Ruigrok, R.W., 1998. Characterization of rabiesvirus nucleocapsids and recombinant nucleocapsid-like structures. J. Gen. Virol. 79(Pt. 12), 2909–2919.
Karlin, D., Ferron, F., Canard, B., Longhi, S., 2003. Structural disorder and modularorganization in Paramyxovirinae N and P. J. Gen. Virol. 84 (Pt. 12), 3239–3252.
Kingston, R.L., Baase, W.A., Gay, L.S., 2004. Characterization of nucleocapsid binding bythe measles virus and mumps virus phosphoproteins. J. Virol. 78 (16), 8630–8640.
Kliche, S., Stitz, L., Mangalam, H., Shi, L., Binz, T., Niemann, H., Briese, T., Lipkin, W.I.,1996. Characterization of the Borna disease virus phosphoprotein, p23. J. Virol. 70(11), 8133–8137.
Kobayashi, T., Shoya, Y., Koda, T., Takashima, I., Lai, P.K., Ikuta, K., Kakinuma, M., Kishi,M., 1998. Nuclear targeting activity associated with the amino terminal region ofthe Borna disease virus nucleoprotein. Virology 243 (1), 188–197.
Kobayashi, T., Watanabe, M., Kamitani, W., Tomonaga, K., Ikuta, K., 2000. Translationinitiation of a bicistronic mRNA of Borna disease virus: a 16-kDa phosphoprotein isinitiated at an internal start codon. Virology 277 (2), 296–305.
Kohno, T., Goto, T., Takasaki, T., Morita, C., Nakaya, T., Ikuta, K., Kurane, I., Sano, K., Nakai,M., 1999. Fine structure and morphogenesis of Borna disease virus. J. Virol. 73 (1),760–766.
Kolakofsky, D., Le Mercier, P., Iseni, F., Garcin, D., 2004. Viral DNA polymerase scanningand the gymnastics of Sendai virus RNA synthesis. Virology 318 (2), 463–473.
Kouznetzoff, A., Buckle, M., Tordo, N., 1998. Identification of a region of the rabies virus Nprotein involved in direct binding to the viral RNA. J. Gen. Virol. 79 (Pt. 5), 1005–1013.
Lata, S., Roessle, M., Solomons, J., Jamin, M., Gottlinger, H.G., Svergun, D.I., Weissenhorn,W., 2008. Structural basis for autoinhibition of ESCRT-III CHMP3. J. Mol. Biol. 378(4), 818–827.
Luo, M., Green, T.J., Zhang, X., Tsao, J., Qiu, S., 2007. Structural comparisons of thenucleoprotein from three negative strand RNA virus families. Virol. J. 4, 72.
Maclellan, K., Loney, C., Yeo, R.P., Bhella, D., 2007. The 24-angstrom structure ofrespiratory syncytial virus nucleocapsid protein–RNA decameric rings. J. Virol. 81(17), 9519–9524.
Masters, P.S., Banerjee, A.K., 1988. Complex formation with vesicular stomatitis virusphosphoprotein NS prevents binding of nucleocapsid protein N to nonspecific RNA.J. Virol. 62 (8), 2658–2664.
Mavrakis, M., Kolesnikova, L., Schoehn, G., Becker, S., Ruigrok, R.W.H., 2002.Morphology of Marburg virus NP–RNA. Virology 296 (2), 300–307.
Mavrakis, M., Iseni, F., Mazza, C., Schoehn, G., Ebel, C., Gentzel, M., Franz, T., Ruigrok,R.W.H., 2003. Isolation and characterisation of the rabies virus N degrees-Pcomplex produced in insect cells. Virology 305 (2), 406–414.
Mayer, D., Baginsky, S., Schwemmle, M., 2005. Isolation of viral ribonucleoproteincomplexes from infected cells by tandem affinity purification. Proteomics 5 (17),4483–4487.
Mellon, M.G., Emerson, S.U., 1978. Rebinding of transcriptase components (L and NSproteins) to the nucleocapsid template of vesicular stomatitis virus. J. Virol. 27 (3),560–567.
Möller, P., Pariente, N., Klenk, H.-D., Becker, S., 2005. Homo-oligomerization of Marburgvirus VP35 is essential for its function in replication and transcription. J. Virol. 79(23), 14876–14886.
Money, V.A., McPhee, H.K., Mosely, J.A., Sanderson, J.M., Yeo, R.P., 2009. Surface featuresof a Mononegavirales matrix protein indicate sites of membrane interaction. Proc.Natl. Acad. Sci. U. S. A. 106 (11), 4441–4446.
Murphy, M.F., Lazzarini, R.A., 1974. Synthesis of viral mRNA and polyadenylate by aribonucleoprotein complex from extracts of VSV-infected cells. Cell 3 (1), 77–84.
Neumann, P., Lieber, D., Meyer, S., Dautel, P., Kerth, A., Kraus, I., Garten, W., Stubbs, M.T.,2009. Crystal structure of the Borna disease virus matrix protein (BDV-M) revealsssRNA binding properties. Proc. Natl. Acad. Sci. U. S. A. 106 (10), 3710–3715.
Pattnaik, A.K., Ball, L.A., LeGrone, A.W., Wertz, G.W., 1992. Infectious defectiveinterfering particles of VSV from transcripts of a cDNA clone. Cell 69 (6), 1011–1020.
Perez, M., Sanchez, A., Cubitt, B., Rosario, D., de la Torre, J.C., 2003. A reverse geneticssystem for Borna disease virus. J. Gen. Virol. 84 (Pt. 11), 3099–3104.
Pleschka, S., Staeheli, P., Kolodziejek, J., Richt, J.A., Nowotny, N., Schwemmle, M., 2001.Conservation of coding potential and terminal sequences in four different isolatesof Borna disease virus. J. Gen. Virol. 82 (Pt. 11), 2681–2690.
Poenisch, M., Wille, S., Ackermann, A., Staeheli, P., Schneider, U., 2007. The X protein ofBorna disease virus serves essential functions in the viral multiplication cycle.J. Virol. 81 (13), 7297–7299.
Poenisch, M., Staeheli, P., Schneider, U., 2008. Viral accessory protein X stimulates theassembly of functional Borna disease virus polymerase complexes. J. Gen. Virol. 89(Pt. 6), 1442–1445.
Portner, A., Murti, K.G., 1986. Localization of P, NP, and M proteins on Sendai virusnucleocapsid using immunogold labeling. Virology 150 (2), 469–478.
Price, S.R., Ito, N., Oubridge, C., Avis, J.M., Nagai, K., 1995. Crystallization of RNA-proteincomplexes: I. Methods for the large-scale preparation of RNA suitable forcrystallographic studies. J. Mol. Biol. 249 (2), 398–408.
Pyper, J.M., Gartner, A.E., 1997. Molecular basis for the differential subcellularlocalization of the 38- and 39-kilodalton structural proteins of Borna diseasevirus. J. Virol. 71 (7), 5133–5139.
Richt, J.A., Rott, R., 2001. Borna disease virus: a mystery as an emerging zoonoticpathogen. Vet. J. (Lond. Eng., 1997) 161 (1), 24–40.
Richt, J.A., Clements, J.E., Herzog, S., Pyper, J., Wahn, K., Becht, H., 1993. Analysis of virus-specific RNA species and proteins in Freon-113 preparations of the Borna diseasevirus. Med. Microbiol. Immunol. 182 (5), 271–280.
Rott, R., Becht, H., 1995. Natural and experimental Borna disease in animals. Curr. Top.Microbiol. Immunol. 190, 17–30.
Rudolph, M.G., Kraus, I., Dickmanns, A., Eickmann, M., Garten, W., Ficner, R., 2003.Crystal structure of the Borna disease virus nucleoprotein. Structure (Lond. Eng.,1993) 11 (10), 1219–1226.
Schmid, S., Mayer, D., Schneider, U., Schwemmle, M., 2007. Functional characterizationof the major and minor phosphorylation sites of the P protein of Borna diseasevirus. J. Virol. 81 (11), 5497–5507.
Schneider, U., 2005. Novel insights into the regulation of the viral polymerase complexof neurotropic Borna disease virus. Virus Res. 111 (2), 148–160.
Schneider, P.A., Schneemann, A., Lipkin, W.I., 1994. RNA splicing in Borna disease virus,a nonsegmented, negative-strand RNA virus. J. Virol. 68 (8), 5007–5012.
Schneider, U., Naegele, M., Staeheli, P., Schwemmle, M., 2003. Active Borna disease viruspolymerase complex requires a distinct nucleoprotein-to-phosphoprotein ratio butno viral X protein. J. Virol. 77 (21), 11781–11789.
Schneider, U., Blechschmidt, K., Schwemmle, M., Staeheli, P., 2004a. Overlap ofinteraction domains indicates a central role of the P protein in assembly andregulation of the Borna disease virus polymerase complex. J. Biol. Chem. 279 (53),55290–55296.
Schneider, U., Naegele, M., Staeheli, P., 2004b. Regulation of the Borna disease viruspolymerase complex by the viral nucleoprotein p38 isoform. Brief Report. Arch.Virol. 149 (7), 1409–1414.
Schneider, U., Schwemmle, M., Staeheli, P., 2005. Genome trimming: a unique strategyfor replication control employed by Borna disease virus. Proc. Natl. Acad. Sci. U. S. A.102 (9), 3441–3446.
Schneider, U., Martin, A., Schwemmle, M., Staeheli, P., 2007. Genome trimming by Bornadisease viruses: viral replication control or escape from cellular surveillance? Cell.Mol. Life Sci.: CMLS 64 (9), 1038–1042.
Schoehn, G., Iseni, F., Mavrakis, M., Blondel, D., Ruigrok, R.W., 2001. Structure ofrecombinant rabies virus nucleoprotein–RNA complex and identification of thephosphoprotein binding site. J. Virol. 75 (1), 490–498.
Schoehn, G., Mavrakis, M., Albertini, A., Wade, R., Hoenger, A., Ruigrok, R.W.H., 2004.The 12 A structure of trypsin-treated measles virus N–RNA. J. Mol. Biol. 339 (2),301–312.
Schoehn, G., El Bakkouri, M., Fabry, C.M.S., Billet, O., Estrozi, L.F., Le, L., Curiel, D.T.,Kajava, A.V., Ruigrok, R.W.H., Kremer, E.J., 2008. Three-dimensional structure ofcanine adenovirus serotype 2 capsid. J. Virol. 82 (7), 3192–3203.
Schwardt, M., Mayer, D., Frank, R., Schneider, U., Eickmann, M., Planz, O., Wolff, T.,Schwemmle, M., 2005. The negative regulator of Borna disease virus polymerase isa non-structural protein. J. Gen. Virol. 86 (Pt. 11), 3163–3169.
Schwemmle, M., De, B., Shi, L., Banerjee, A., Lipkin, W.I., 1997. Borna disease virus P-protein is phosphorylated by protein kinase C epsilon and casein kinase II. J. Biol.Chem. 272 (35), 21818–21823.
Schwemmle, M., Salvatore, M., Shi, L., Richt, J., Lee, C.H., Lipkin, W.I., 1998. Interactionsof the Borna disease virus P, N, and X proteins and their functional implications.J. Biol. Chem. 273 (15), 9007–9012.
Schwemmle, M., Jehle, C., Shoemaker, T., Lipkin, W.I., 1999. Characterization of themajor nuclear localization signal of the Borna disease virus phosphoprotein. J. Gen.Virol. 80 (Pt. 1), 97–100.
Shoya, Y., Kobayashi, T., Koda, T., Ikuta, K., Kakinuma, M., Kishi, M., 1998. Two proline-rich nuclear localization signals in the amino- and carboxyl-terminal regions of theBorna disease virus phosphoprotein. J. Virol. 72 (12), 9755–9762.
Spehner, D., Kirn, A., Drillien, R., 1991. Assembly of nucleocapsidlike structures inanimal cells infected with a vaccinia virus recombinant encoding the measles virusnucleoprotein. J. Virol. 65 (11), 6296–6300.
Szilágyi, J.F., Uryvayev, L., 1973. Isolation of an infectious ribonucleoprotein fromvesicular stomatitis virus containing an active RNA transcriptase. J. Virol. 11 (2),279–286.
Tarbouriech, N., Curran, J., Ebel, C., Ruigrok, R.W., Burmeister, W.P., 2000a. On thedomain structure and the polymerization state of the Sendai virus P protein.Virology 266 (1), 99–109.
Tarbouriech, N., Curran, J., Ruigrok, R.W., Burmeister, W.P., 2000b. Tetrameric coiled coildomain of Sendai virus phosphoprotein. Nat. Struct. Biol. 7 (9), 777–781.
Tomonaga, K., Kobayashi, T., Lee, B.-J., Watanabe, M., Kamitani, W., Ikuta, K., 2000.Identification of alternative splicing and negative splicing activity of a nonseg-mented negative-strand RNA virus, Borna disease virus. Proc. Natl. Acad. Sci. U. S. A.97 (23), 12788–12793.
Toriumi, H., Kawai, A., 2004. Association of rabies virus nominal phosphoprotein (P)with viral nucleocapsid (NC) is enhanced by phosphorylation of the viralnucleoprotein (N). Microbiol. Immunol. 48 (5), 399–409.
Walker, M.P., Jordan, I., Briese, T., Fischer, N., Lipkin, W.I., 2000. Expression andcharacterization of the Borna disease virus polymerase. J. Virol. 74 (9), 4425–4428.
Yanai, H., Hayashi, Y., Watanabe, Y., Ohtaki, N., Kobayashi, T., Nozaki, Y., Ikuta, K.,Tomonaga, K., 2006a. Development of a novel Borna disease virus reverse geneticssystem using RNA polymerase II promoter and SV40 nuclear import signal.Microbes Infect./Institut Pasteur 8 (6), 1522–1529.
Yanai, H., Kobayashi, T., Hayashi, Y., Watanabe, Y., Ohtaki, N., Zhang, G., de la Torre, J.C.,Ikuta, K., Tomonaga, K., 2006b. A methionine-rich domain mediates CRM1-dependent nuclear export activity of Borna disease virus phosphoprotein. J. Virol.80 (3), 1121–1129.
Yang, J., Hooper, D.C., Wunner, W.H., Koprowski, H., Dietzschold, B., Fu, Z.F., 1998. Thespecificity of rabies virus RNA encapsidation by nucleoprotein. Virology 242 (1),107–117.
Zhang, X., Green, T.J., Tsao, J., Qiu, S., Luo, M., 2008. Role of intermolecular interactions ofvesicular stomatitis virus nucleoprotein in RNA encapsidation. J. Virol. 82 (2),674–682.
Copyright © 2022 FDOKUMEN




















