
Reproductive status and body condition of Atlantic bluefin tuna in the Gulf of Maine, 2000–2002
13
RESEARCH ARTICLE Reproductive status and body condition of Atlantic bluefin tuna in the Gulf of Maine, 2000–2002 Jennifer Goldstein Scott Heppell Andrew Cooper Solange Brault Molly Lutcavage Received: 29 September 2006 / Accepted: 1 February 2007 Ó Springer-Verlag 2007 Abstract The reproductive status and body condition of 195 (‡185 cm curved fork length, CFL; assigned age 7 and above) Atlantic bluefin tuna were assessed in the Gulf of Maine during the commercial fishing season of June– October, 2000–2002. Given the distance between known spawning and feeding grounds, the prevailing paradigm for Atlantic bluefin tuna (Thunnus thynnus thynnus, L.) suggests that the most likely histological state for females arriving in the Gulf of Maine after spawning would be a resting or quiescent state with little or no perigonadal fat. Alternatively, the presence of mature or mature-inactive histological states in some females supports a more varied or individualistic model for bluefin reproduction. No relationship was found between body condition and reproductive status. Males were found in all reproductive stages, but were more likely to be in spawning condition (stages 4 and 5) or a mature-inactive state (stage 6) in June and July. Female bluefin tuna were found in stage 1 (immature or non-spawning) and stage 6 (mature-inac- tive). Stage 6 females were only present in June and July and smaller females (<235 cm CFL) were more likely to be in stage 6 than large females (>235 cm CFL) sampled during those same months. The presence of smaller fe- males in stage 6 arriving at the same time as larger fe- males in stage 1 indicates that Western Atlantic bluefin tuna may have an asynchronous reproductive schedule and may mature at a smaller size than the currently ac- cepted paradigm suggests. Introduction The reproductive biology of the western component of the Atlantic bluefin tuna (Thunnus thynnus) stock remains incomplete and impaired by large uncertainties, despite the depleted status and economic importance of this fishery. When, where, how often and at what size/age long-lived, highly migratory fish species reproduce are key life history characteristics needed by fisheries man- agers to determine the regenerative ability of the stock (Mather et al. 1995; Quinn and Deriso 1999; Jorgensen et al. 2006; Fromentin and Powers 2005). Bluefin tuna feed in energy-rich coastal areas at higher latitudes to increase somatic fat reserves that fuel reproduction and migration (Crane 1936; Bigelow and Schroeder 1953; Dragovich 1970; Chase 2002). Poor body condition brought about by low quality prey or reduced prey availability can affect reproductive success through re- duced fecundity (Hislop et al. 1978) and changes in reproductive schedule (Burton and Idler 1984, 1987; Hunter and Macewicz 1985; Larsson et al 1990; Rijnsdorp Communicated by J.P. Grassle. J. Goldstein (&) S. Brault Biology Department, University of Massachusetts, Boston, MA 02125, USA e-mail: [email protected] S. Heppell Department of Fisheries and Wildlife, Oregon State University, Corvallis, OR 97331, USA A. Cooper Department of Natural Resources, Institute for the Study of Earth, Oceans and Space, University of New Hampshire, Durham, NH 03824, USA M. Lutcavage Large Pelagics Research Center, University of New Hampshire, Durham, NH 03824, USA 123 Mar Biol DOI 10.1007/s00227-007-0638-8
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Reproductive status and body condition of Atlantic bluefin tuna in the Gulf of Maine, 2000–2002

RESEARCH ARTICLE
Reproductive status and body condition of Atlanticbluefin tuna in the Gulf of Maine, 2000–2002
Jennifer Goldstein Æ Scott Heppell Æ Andrew Cooper ÆSolange Brault Æ Molly Lutcavage
Received: 29 September 2006 / Accepted: 1 February 2007
� Springer-Verlag 2007
Abstract The reproductive status and body condition of
195 (‡185 cm curved fork length, CFL; assigned age 7
and above) Atlantic bluefin tuna were assessed in the Gulf
of Maine during the commercial fishing season of June–
October, 2000–2002. Given the distance between known
spawning and feeding grounds, the prevailing paradigm
for Atlantic bluefin tuna (Thunnus thynnus thynnus, L.)
suggests that the most likely histological state for females
arriving in the Gulf of Maine after spawning would be a
resting or quiescent state with little or no perigonadal fat.
Alternatively, the presence of mature or mature-inactive
histological states in some females supports a more varied
or individualistic model for bluefin reproduction. No
relationship was found between body condition and
reproductive status. Males were found in all reproductive
stages, but were more likely to be in spawning condition
(stages 4 and 5) or a mature-inactive state (stage 6) in
June and July. Female bluefin tuna were found in stage 1
(immature or non-spawning) and stage 6 (mature-inac-
tive). Stage 6 females were only present in June and July
and smaller females (<235 cm CFL) were more likely to
be in stage 6 than large females (>235 cm CFL) sampled
during those same months. The presence of smaller fe-
males in stage 6 arriving at the same time as larger fe-
males in stage 1 indicates that Western Atlantic bluefin
tuna may have an asynchronous reproductive schedule
and may mature at a smaller size than the currently ac-
cepted paradigm suggests.
Introduction
The reproductive biology of the western component of the
Atlantic bluefin tuna (Thunnus thynnus) stock remains
incomplete and impaired by large uncertainties, despite
the depleted status and economic importance of this
fishery. When, where, how often and at what size/age
long-lived, highly migratory fish species reproduce are
key life history characteristics needed by fisheries man-
agers to determine the regenerative ability of the stock
(Mather et al. 1995; Quinn and Deriso 1999; Jorgensen
et al. 2006; Fromentin and Powers 2005). Bluefin tuna
feed in energy-rich coastal areas at higher latitudes to
increase somatic fat reserves that fuel reproduction and
migration (Crane 1936; Bigelow and Schroeder 1953;
Dragovich 1970; Chase 2002). Poor body condition
brought about by low quality prey or reduced prey
availability can affect reproductive success through re-
duced fecundity (Hislop et al. 1978) and changes in
reproductive schedule (Burton and Idler 1984, 1987;
Hunter and Macewicz 1985; Larsson et al 1990; Rijnsdorp
Communicated by J.P. Grassle.
J. Goldstein (&) � S. Brault
Biology Department, University of Massachusetts,
Boston, MA 02125, USA
e-mail: [email protected]
S. Heppell
Department of Fisheries and Wildlife,
Oregon State University, Corvallis, OR 97331, USA
A. Cooper
Department of Natural Resources,
Institute for the Study of Earth, Oceans and Space,
University of New Hampshire, Durham, NH 03824, USA
M. Lutcavage
Large Pelagics Research Center,
University of New Hampshire,
Durham, NH 03824, USA
123
Mar Biol
DOI 10.1007/s00227-007-0638-8

1990; Rajisilta 1992; Rideout et al. 2000; Jorgensen et al.
2006). Seafood brokers’ records of fat and oil grades
indicate that bluefin tuna arriving in the Gulf of Maine are
usually in poor condition (Estrada et al. 2005), and that
overall body condition and quality has declined over the
past 14 years (Golet et al. 2006). Shifts in fat and energy
stores in relation to reproduction have been documented
for the eastern Atlantic bluefin tuna stock (Frade 1937;
Rodriguez-Roda 1963; Medina et al. 2002; Abascal et al.
2004), though there have been no previous studies of
reproduction or energy stores for bluefin tuna that forage
in the Gulf of Maine in summer and fall.
Mature bluefin tuna have been presumed to follow an
annual cycle of foraging from June through March off the
eastern US and Canadian coasts, followed by migration to
the Gulf of Mexico to spawn in April and May, although
Rivas (1978) suggested that some western fish might spawn
in the Mediterranean. The prevailing assumption is that the
Gulf of Maine assemblage is comprised of individuals from
the western Atlantic stock that matures at age 8–10 (�200
to 225 cm CFL, Turner et al. 1991) and shows spawning
site fidelity to the Gulf of Mexico or Florida Straits (Rivas
1954; NRC 1994; Mather et al. 1995; Nemerson et al.
2000; Block et al. 2005). An eastern Atlantic stock, which
primarily spawns in the Mediterranean, achieves sexual
maturity at age 3–5, or 103–110 cm FL (�108–115 CFL)
(Tiews 1963; Rodriguez-Roda 1964; Susca et al. 2000,
2001b; Medina et al. 2002; Corriero et al. 2005).
Recent electronic tagging data from bluefin tuna ini-
tiated in the western Atlantic have been used to support
this interpretation (Stokesbury et al. 2004; Block et al.
2001, 2005), or to call it into question (Lutcavage et al.
1999; Wilson et al. 2005; Fromentin and Powers 2005).
Tag reporting rates from the Gulf of Mexico for mature
sized bluefin tuna from all studies (N = 381) were only
3–11%, including individuals that were present well be-
fore the spawning season of April–June (Lutcavage et al.
1999; Stokesbury et al. 2004; Block et al. 2005; Wilson
et al. 2005; Sibert et al. 2006; F. Royer et al., unpub-
lished data). Most (N � 317) were found in a vast area
spanning from the Gulf Stream margins to the Azores,
where there is substantial mixing of the two purported
stocks (Lutcavage et al. 1999; Block et al. 2001, 2005;
Sibert et al. 2006). These results present the possibility
that (1) individual bluefin tuna do not spawn on an an-
nual cycle as previously assumed (Lutcavage et al. 1999;
Lioka et al. 2000; Corriero et al. 2003; Block et al. 2005;
Fromentin and Powers 2005), (2) a component of the
western stock is spawning somewhere other than the Gulf
of Mexico, e.g., in the central north Atlantic or Gulf
Stream edge (Mather et al. 1995; Lutcavage et al. 1999)
or (3) some of the electronically tagged fish have eastern
Atlantic origins but migrate west as juveniles and share
feeding grounds in US coastal waters (Rooker and Secor
2004; Block et al. 2005). Any of these scenarios impact
the quality of population assessments used to pro-
mote conservation and stock recovery for this depleted
species.
Ideally, reproductive parameters should be measured in
fish collected directly on the spawning grounds (NRC
1994; Farley and Davis 1998; Schaefer 1998, 2001;
Medina et al. 2002) because direct evidence of spawning
activity (migratory nucleus oocytes, post-ovulatory folli-
cles (POFs), and fully hydrated oocytes) are detectable for
£24 h after spawning in tunas (Hunter et al. 1986;
Schaefer 1996, 1998, 2001; Medina et al 2002; Corriero
et al. 2005). However, sampling in the Gulf of Mexico is
difficult due to a moratorium on bluefin tuna fishing there.
Also, if a portion of the stock is spawning elsewhere or if
the population structure more closely resembles a meta-
population (Fromentin and Powers 2005), the sizes of fish
in the Gulf of Mexico in April and May may not accu-
rately represent the spawning size range of the population
as a whole.
Despite their distance from known spawning grounds,
Gulf of Maine bluefin tuna offer alternative information on
size at maturity and timing of spawning. Though direct
evidence of spawning is unlikely to be detected, atretic
(degrading) vitellogenic oocytes, which indicate that
maturity has occurred during the last maturity cycle
(Heppell and Sullivan 1999; Corriero et al. 2003), are
detectable for a longer time period. Atretic follicles in
northern anchovy (Eugraulis mordax) were detectable for
as long as fourteen days after cessation of spawning
(Hunter and Macewicz 1985), but are likely to be resorbed
more quickly in tuna due to their higher metabolism and
relatively warm internal body temperature (Hunter et al.
1986). The presence of vitellogenic and atretic vitellogenic
oocytes in histological samples from Gulf of Maine bluefin
tuna would indicate that maturity had occurred recently,
even if spawning per se cannot be determined (Heppell and
Sullivan 2000).
Given the distance between known spawning and feed-
ing grounds, the prevailing paradigm suggests that the most
likely histological state for females arriving in the Gulf of
Maine after spawning would be a resting or quiescent state
with little or no perigonadal fat. Alternatively, the presence
of mature or mature-inactive histological states in some
females supports a more varied or individualistic model for
bluefin reproduction. The purpose of this study is to (1)
characterize the reproductive status of Gulf of Maine
bluefin tuna using histological techniques and (2) use
general linear models (GLMs) to determine whether the
reproductive characteristics of histological stage and gonad
weight could be separated on the basis of size/age and body
condition.
Mar Biol
123

Materials and methods
Sample collection
We asked 12 commercial fishermen participating in the
seasonal Gulf of Maine fishery for large and large-medium
bluefin (‡185 cm CFL) to collect stomachs, gonads, and
perigonadal fat. The minimum legal size for bluefin tuna
landed in the commercial sector of the fishery is 185 cm
CFL, so smaller sizes could not be sampled for this study.
Perigonadal fat is an adipose fat reserve attached directly to
the gonad, used primarily for the production of gametes but
also for general metabolism (Mourente et al. 2002; Abascal
et al. 2004; Corriero et al. 2003). It can be used as an
indicator of condition and foraging success because it ac-
crues rapidly over the summer and fall if feeding condi-
tions are appropriate. A 1-cm3 sample of muscle tissue was
frozen for hormonal analyses of 11-ketotestosterone and
17ß-oestradiol levels to determine whether a relationship
between hormone levels and maturational status was
detectable, similar to the study by Susca et al. (2000).
Sampling coincided with the commercial fishing season
June 1st through mid October 2000–2002. Fishermen were
asked to record capture location and gear. Either CFL or
dressed length (DL), which is the curved length of the fish
minus the head, was recorded. Dressed weights (DW) were
recorded rather than round weights, as most fishermen re-
move heads from fish at sea prior to obtaining a weight
dockside. DWs and DLs were converted to round weights
and CFLs when necessary by using the conversion factors
of DW · 1.25 = round weight (Anon. 2003) and
CFL = DL · 1.35 (S. Turner, unpublished data). Samples
were obtained from 195 fish (80 females and 115 males),
ranging in size from 185 to 291 cm CFL, and round
weights of 75–314 kg. The study area includes the Gulf of
Maine and some sampling also occurred east of Cape Cod
and south to the Great South Channel, all known foraging
areas for bluefin tuna (Crane 1936; Bigelow and Schroeder
1953; Caddy and Butler 1975; Chase 2002) (Fig. 1).
Histological analysis
Perigonadal fat was trimmed from the gonads and each
weighed separately to the nearest 0.1 g. A gonadosomatic
index was calculated by the following formula:
GSI ¼ gonad weight (kg)
RW (kg)� 100. ð1Þ
A 1.0 cm3 sub-sample from the midsection of the gonad
was preserved for histological analysis in 10% buffered
formalin. Gonads were staged according to Heppell and
Sullivan (1999) and stages were defined as follows:
Females
Stage 0: Largest germ cells are primary oocytes with a
conspicuous nucleus containing chromatin threads and a
single large nucleolus, stage 1: primary (perinucleolar)
stage oocytes; minor atresia may be present, stage 2:
vitellogenin independent growth, stage 3: early vitello-
genesis, stage 4: late vitellogenesis, stage 5: final oocyte
maturation and spawning, stage 6: degredation and pres-
ence of atretic follicles. If yolked, atretic oocytes are not
present, it may be impossible to determine whether the fish
is truly immature, or is mature but has had sufficient time
Fig. 1 Sampling locations for
bluefin tuna, Thunnus thynnus,
in the Gulf of Maine and
adjacent waters. Inset shows
study site in relation to the
western North Atlantic. Filledcircle 2000, filled triangle 2001,
plus symbol 2002
Mar Biol
123
since spawning to resorb evidence of maturation (Schaefer
1998; Corriero et al. 2003). These individuals are classified
as stage 1 fish, and termed ‘‘immature or non-maturing’’.
Males
Stage 1: no signs of spermatogenesis, stage 2: spermato-
cytes present, stage 3: spermatocytes and spermatids
present, stage 4: mature sperm in small quantities, stage 5:
fully mature testis, stage 6: collapsing crypts and tubules.
Stage 0 was not defined for males. Stages 1–3 are con-
sidered to be immature or non-maturing, stages 4–5 are
considered to be mature-active, and stage 6 is considered to
be mature-inactive for both males and females.
Calculation of Wr
To quantify changes in body condition over the season, we
calculated a condition factor, Wr, after Anderson and
Neumann (1996).
Wr ¼Wo
Ws
: ð2Þ
Wr is an index of the relative weight of a fish, where Wo is
the observed weight (in kg) of an individual and Ws (in kg)
is a length-specific standard weight predicted by a weight–
length regression constructed to represent the population.
The weight–length regression was calculated from the
National Marine Fisheries Service (NMFS) landings data
(Bluefin Tuna Landings Database, NMFS Northeast Re-
gional Office, Gloucester, MA, 01930, USA) collected
during 2000–2002. Only landings data from the New
England region were used to ensure that Ws was calculated
from the same assemblage of fish.
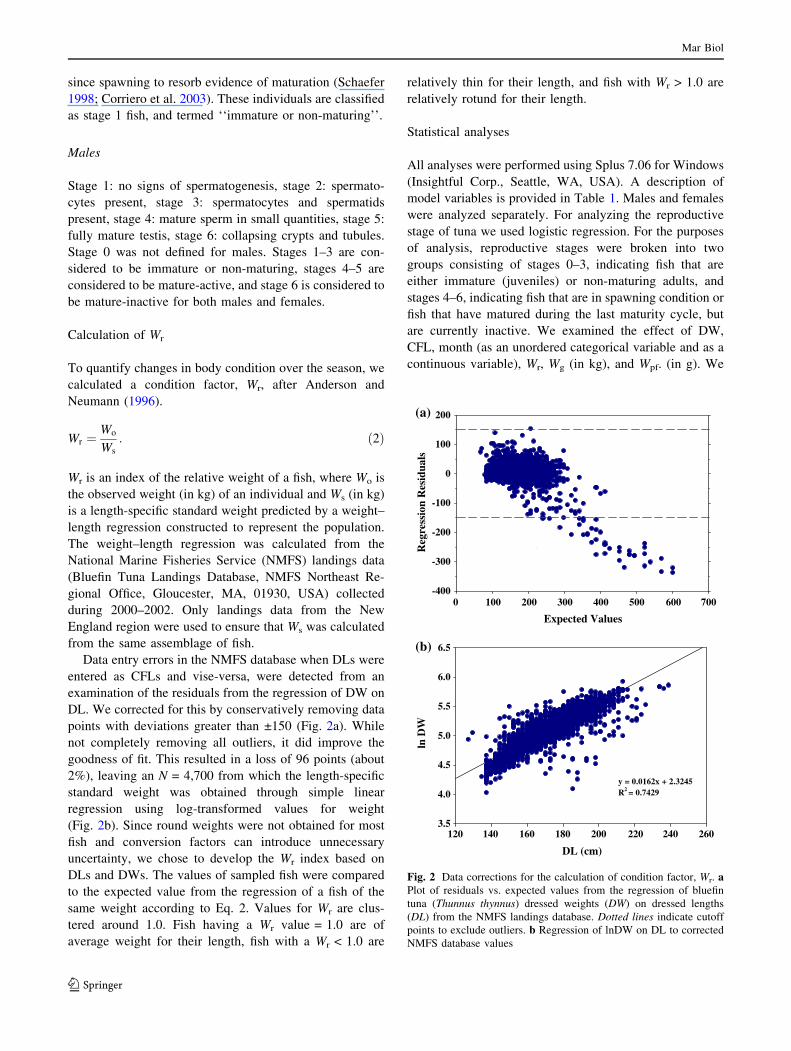
Data entry errors in the NMFS database when DLs were
entered as CFLs and vise-versa, were detected from an
examination of the residuals from the regression of DW on
DL. We corrected for this by conservatively removing data
points with deviations greater than ±150 (Fig. 2a). While
not completely removing all outliers, it did improve the
goodness of fit. This resulted in a loss of 96 points (about
2%), leaving an N = 4,700 from which the length-specific
standard weight was obtained through simple linear
regression using log-transformed values for weight
(Fig. 2b). Since round weights were not obtained for most
fish and conversion factors can introduce unnecessary
uncertainty, we chose to develop the Wr index based on
DLs and DWs. The values of sampled fish were compared
to the expected value from the regression of a fish of the
same weight according to Eq. 2. Values for Wr are clus-
tered around 1.0. Fish having a Wr value = 1.0 are of
average weight for their length, fish with a Wr < 1.0 are
relatively thin for their length, and fish with Wr > 1.0 are
relatively rotund for their length.
Statistical analyses
All analyses were performed using Splus 7.06 for Windows
(Insightful Corp., Seattle, WA, USA). A description of
model variables is provided in Table 1. Males and females
were analyzed separately. For analyzing the reproductive
stage of tuna we used logistic regression. For the purposes
of analysis, reproductive stages were broken into two
groups consisting of stages 0–3, indicating fish that are
either immature (juveniles) or non-maturing adults, and
stages 4–6, indicating fish that are in spawning condition or
fish that have matured during the last maturity cycle, but
are currently inactive. We examined the effect of DW,
CFL, month (as an unordered categorical variable and as a
continuous variable), Wr, Wg (in kg), and Wpf. (in g). We
Expected Values
0 100 200 300 400 500 600 700
Reg
ress
ion
Res
idua
ls
-400
-300
-200
-100
0
100
200
DL (cm)
120 140 160 180 200 220 240 260
ln D
W
3.5
4.0
4.5
5.0
5.5
6.0
6.5
y = 0.0162x + 2.3245R2 = 0.7429
(a)
(b)
Fig. 2 Data corrections for the calculation of condition factor, Wr. aPlot of residuals vs. expected values from the regression of bluefin
tuna (Thunnus thynnus) dressed weights (DW) on dressed lengths
(DL) from the NMFS landings database. Dotted lines indicate cutoff
points to exclude outliers. b Regression of lnDW on DL to corrected
NMFS database values
Mar Biol
123
analyzed gonad weight using generalized linear models
(GLMs) (e.g., McCullagh and Nelder 1989), and examined
the effect of DW, CFL, month (as an unordered categorical
variable and as a continuous variable), Wr, and Wpf (in g).
Main effects for the regression and GLMs were tested
using an alpha-level of 0.05, and interaction effects were
tested using an alpha-level of 0.10.
Results
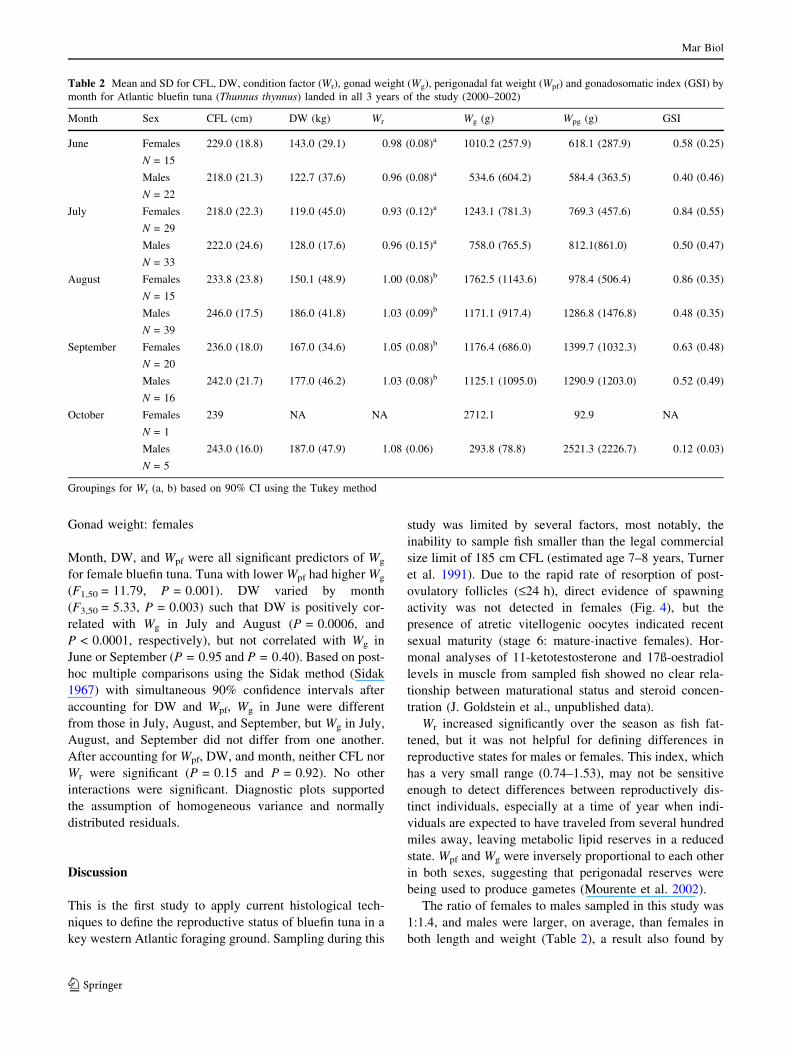
A summary of variables by month (Table 2) revealed that
sex ratios were skewed, with 59% of fish sampled being
male, and 41% female. Mean male CFL and DW were
larger than their corresponding female values in all months
except June, where females tended to be larger. The effect
of month on Wr was significant (P = 0.0001), with June
and July not different from each other, but less than August
and September based on 90% confidence intervals using
the Tukey method. Females had lower Wr overall
(P = 0.027), but the effect of month did not differ by sex
(P = 0.50). Wg was extremely variable, but females tended
to have a higher mean Wg in all months. Wpf did not differ
by sex (P = 0.47), nor did the effect of month differ by sex
(P = 0.26). Mean GSI values were highest in the months of
July and August (0.84 and 0.86) for females, and highest
for males in September (0.52). Females in the mature/
mature-inactive group had GSI values that were higher
than females in the immature /non-maturing group (0.63
vs. 0.47), but the two male functional groups had almost
identical GSI values (0.40 vs. 0.43).
Reproductive stage: males
Males were found in all reproductive stages except stage 0.
Males were more likely to be in a mature or a mature-
inactive state in June and July, with active stages
decreasing as the summer progressed (Fig. 3a). The most
parsimonious model for predicting the reproductive stage
of male bluefin tuna only contained the variable Month, as
an unordered categorical variable, (F4,111 = 7.37,
P < 0.0001). After accounting for month, DW, CFL, Wr,
Wg, and Wpf were each non-significant (P = 0.64, P = 0.52,
P = 0.41, P = 0.28, and P = 0.86, respectively). Month,
however, was always significant with these other variables
in the model. All interactions were similarly not significant.
Diagnostic plots supported the assumption of homogeneous
variance and normally distributed residuals.
Gonad weight: males
Wg was log transformed to correct for heterogeneous sto-
chasticity. The most parsimonious model for predicting Wg
of male bluefin tuna contained only Wpf and DW. Wpf was
negatively correlated with Wg (F1,91 = 24.85, P < 0.0001),
but DW was positively correlated with Wg (F1,91 = 28.8,
P < 0.0001). With Wpf and DW in the model, neither
month, CFL, nor Wr were significant (P = 0.51, P = 0.22,
P = 0.71, respectively). No interactions were significant.
Reproductive stage: females
All females, with the exception of one fish in stage 2, were
found in either stage 1 or stage 6 (Fig. 3b). No length or
weight data was available for the female in stage 2, so it
was excluded from the model. All females landed in Au-
gust, September and October were in stage 1. Month, as an
unordered categorical variable, was a significant factor in
predicting the reproductive stage of female bluefin tuna
(F3,55 = 18.51, P < 0.0001). Females were most likely to
be in a mature-inactive state (stage 6) in June or July. Only
one female was observed in October; it was not in stage 6,
and was not included in this analysis due to the small
sample size. Even after accounting for month and Wg, DW
was also significant (F1,55 = 36.33, P < 0.0001), with lar-
ger females less likely to be in stage 6. When accounting
for month and DW, females with higher Wg were more
likely to be in stage 6 (F1,55 = 40.10, P < 0.0001). With
month, Wg, and DW in the model, Wr was not significant
(P = 0.92); nor was Wpf (P = 0.90). Interactions between
month and Wg, month and DW, and Wg and DW were all
nonsignificant (P = 0.33, P = 0.90, and P = 0.32, respec-
tively). Diagnostic plots supported the assumption of
homogeneous variance and normally distributed residuals.
Table 1 Conditional and
morphometric descriptor
variables for Atlantic bluefin
tuna, Thunnus thynnus, used in
logistic regression and GLM
analyses
Variable Abbreviation Units Description
Dressed wt. DW kg Fish weight—gilled, gutted, head removed
Curved fork length CFL cm Length measured along the contour of the body,
from tip of upper lip to fork of tail, touching
top of pectoral fin
Condition factor Wr None Relative weight of a fish (see text, Eq. 1)
Perigonadal fat wt. Wpf g Weight of perigonadal fat reserve
Gonad wt. Wg g Weight of gonads
Mar Biol
123
Gonad weight: females
Month, DW, and Wpf were all significant predictors of Wg
for female bluefin tuna. Tuna with lower Wpf had higher Wg
(F1,50 = 11.79, P = 0.001). DW varied by month
(F3,50 = 5.33, P = 0.003) such that DW is positively cor-
related with Wg in July and August (P = 0.0006, and
P < 0.0001, respectively), but not correlated with Wg in
June or September (P = 0.95 and P = 0.40). Based on post-
hoc multiple comparisons using the Sidak method (Sidak
1967) with simultaneous 90% confidence intervals after
accounting for DW and Wpf, Wg in June were different
from those in July, August, and September, but Wg in July,
August, and September did not differ from one another.
After accounting for Wpf, DW, and month, neither CFL nor
Wr were significant (P = 0.15 and P = 0.92). No other
interactions were significant. Diagnostic plots supported
the assumption of homogeneous variance and normally
distributed residuals.
Discussion
This is the first study to apply current histological tech-
niques to define the reproductive status of bluefin tuna in a
key western Atlantic foraging ground. Sampling during this
study was limited by several factors, most notably, the
inability to sample fish smaller than the legal commercial
size limit of 185 cm CFL (estimated age 7–8 years, Turner
et al. 1991). Due to the rapid rate of resorption of post-
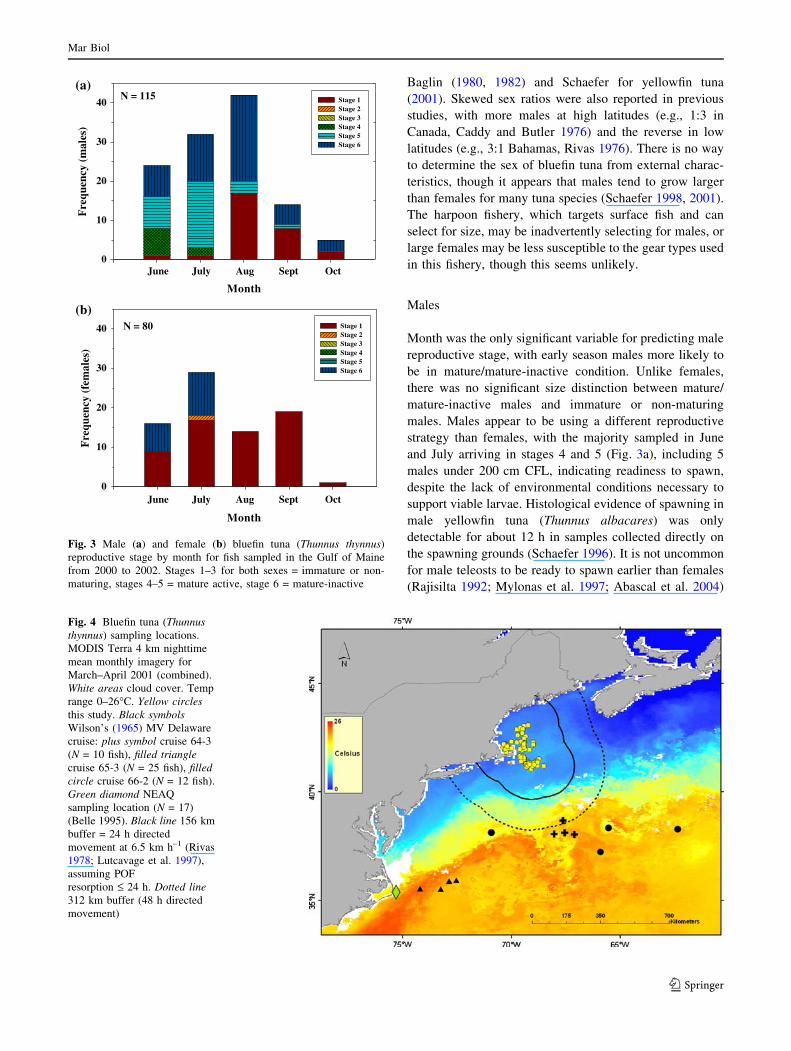
ovulatory follicles (£24 h), direct evidence of spawning
activity was not detected in females (Fig. 4), but the
presence of atretic vitellogenic oocytes indicated recent
sexual maturity (stage 6: mature-inactive females). Hor-
monal analyses of 11-ketotestosterone and 17ß-oestradiol
levels in muscle from sampled fish showed no clear rela-
tionship between maturational status and steroid concen-
tration (J. Goldstein et al., unpublished data).
Wr increased significantly over the season as fish fat-
tened, but it was not helpful for defining differences in
reproductive states for males or females. This index, which
has a very small range (0.74–1.53), may not be sensitive
enough to detect differences between reproductively dis-
tinct individuals, especially at a time of year when indi-
viduals are expected to have traveled from several hundred
miles away, leaving metabolic lipid reserves in a reduced
state. Wpf and Wg were inversely proportional to each other
in both sexes, suggesting that perigonadal reserves were
being used to produce gametes (Mourente et al. 2002).
The ratio of females to males sampled in this study was
1:1.4, and males were larger, on average, than females in
both length and weight (Table 2), a result also found by
Table 2 Mean and SD for CFL, DW, condition factor (Wr), gonad weight (Wg), perigonadal fat weight (Wpf) and gonadosomatic index (GSI) by
month for Atlantic bluefin tuna (Thunnus thynnus) landed in all 3 years of the study (2000–2002)
Month Sex CFL (cm) DW (kg) Wr Wg (g) Wpg (g) GSI
June Females
N = 15
229.0 (18.8) 143.0 (29.1) 0.98 (0.08)a 1010.2 (257.9) 618.1 (287.9) 0.58 (0.25)
Males
N = 22
218.0 (21.3) 122.7 (37.6) 0.96 (0.08)a 534.6 (604.2) 584.4 (363.5) 0.40 (0.46)
July Females
N = 29
218.0 (22.3) 119.0 (45.0) 0.93 (0.12)a 1243.1 (781.3) 769.3 (457.6) 0.84 (0.55)
Males
N = 33
222.0 (24.6) 128.0 (17.6) 0.96 (0.15)a 758.0 (765.5) 812.1(861.0) 0.50 (0.47)
August Females
N = 15
233.8 (23.8) 150.1 (48.9) 1.00 (0.08)b 1762.5 (1143.6) 978.4 (506.4) 0.86 (0.35)
Males
N = 39
246.0 (17.5) 186.0 (41.8) 1.03 (0.09)b 1171.1 (917.4) 1286.8 (1476.8) 0.48 (0.35)
September Females
N = 20
236.0 (18.0) 167.0 (34.6) 1.05 (0.08)b 1176.4 (686.0) 1399.7 (1032.3) 0.63 (0.48)
Males
N = 16
242.0 (21.7) 177.0 (46.2) 1.03 (0.08)b 1125.1 (1095.0) 1290.9 (1203.0) 0.52 (0.49)
October Females
N = 1
239 NA NA 2712.1 92.9 NA
Males
N = 5
243.0 (16.0) 187.0 (47.9) 1.08 (0.06) 293.8 (78.8) 2521.3 (2226.7) 0.12 (0.03)
Groupings for Wr (a, b) based on 90% CI using the Tukey method
Mar Biol
123
Baglin (1980, 1982) and Schaefer for yellowfin tuna
(2001). Skewed sex ratios were also reported in previous
studies, with more males at high latitudes (e.g., 1:3 in
Canada, Caddy and Butler 1976) and the reverse in low
latitudes (e.g., 3:1 Bahamas, Rivas 1976). There is no way
to determine the sex of bluefin tuna from external charac-
teristics, though it appears that males tend to grow larger
than females for many tuna species (Schaefer 1998, 2001).
The harpoon fishery, which targets surface fish and can
select for size, may be inadvertently selecting for males, or
large females may be less susceptible to the gear types used
in this fishery, though this seems unlikely.
Males
Month was the only significant variable for predicting male
reproductive stage, with early season males more likely to
be in mature/mature-inactive condition. Unlike females,
there was no significant size distinction between mature/
mature-inactive males and immature or non-maturing
males. Males appear to be using a different reproductive
strategy than females, with the majority sampled in June
and July arriving in stages 4 and 5 (Fig. 3a), including 5
males under 200 cm CFL, indicating readiness to spawn,
despite the lack of environmental conditions necessary to
support viable larvae. Histological evidence of spawning in
male yellowfin tuna (Thunnus albacares) was only
detectable for about 12 h in samples collected directly on
the spawning grounds (Schaefer 1996). It is not uncommon
for male teleosts to be ready to spawn earlier than females
(Rajisilta 1992; Mylonas et al. 1997; Abascal et al. 2004)
Fig. 4 Bluefin tuna (Thunnusthynnus) sampling locations.
MODIS Terra 4 km nighttime
mean monthly imagery for
March–April 2001 (combined).
White areas cloud cover. Temp
range 0–26�C. Yellow circlesthis study. Black symbolsWilson’s (1965) MV Delaware
cruise: plus symbol cruise 64-3
(N = 10 fish), filled trianglecruise 65-3 (N = 25 fish), filledcircle cruise 66-2 (N = 12 fish).
Green diamond NEAQ
sampling location (N = 17)
(Belle 1995). Black line 156 km
buffer = 24 h directed
movement at 6.5 km h–1 (Rivas
1978; Lutcavage et al. 1997),
assuming POF
resorption £ 24 h. Dotted line312 km buffer (48 h directed
movement)
Month
June July Aug Sept Oct
Month
June July Aug Sept Oct
Fre
quen
cy (
mal
es)
0
10
20
30
40 Stage 1 Stage 2 Stage 3 Stage 4 Stage 5 Stage 6
N = 115 F
requ
ency
(fe
mal
es)
0
10
20
30
40 Stage 1 Stage 2 Stage 3 Stage 4 Stage 5 Stage 6
N = 80
(a)
(b)
Fig. 3 Male (a) and female (b) bluefin tuna (Thunnus thynnus)
reproductive stage by month for fish sampled in the Gulf of Maine
from 2000 to 2002. Stages 1–3 for both sexes = immature or non-
maturing, stages 4–5 = mature active, stage 6 = mature-inactive
Mar Biol
123

and to continue spermiating after females have ceased
spawning (Burton and Idler 1984), but there is little
information regarding the post-spawning condition of
bluefin tuna males. Whether males producing active sperm
are capable of spawning in the absence of appropriate
environmental cues is unknown.
Females
Females entering the Gulf of Maine in June and July were
more likely to be in stage 6 (mature-inactive) than in stage 1
(immature or non-maturing) (Fig. 3b). After July, no fe-
males were found to be in a mature-inactive state. Females
exhibited two reproductive schedules, with all females
between 235 and 265 cm in the immature or non-maturing
state (stage 1), while smaller females (185–235 cm) were
significantly more likely to be in a mature-inactive state
(stage 6). Given the assigned age of females over 235 cm
(>>10 years), they are most likely mature fish traveling
from distant spawning grounds that have had sufficient time
to resorb oocytes and atretic follicles. The presence of rel-
atively small females (185–235 cm) in the mature-inactive
state is unexpected because (1) it indicates that some bluefin
tuna of assigned age 7–8 years are mature, despite the
apparent absence of this size/age class in the Gulf of Mexico
during the spawning season (Block et al. 2005), and (2)
maturity should not be detectable in animals that have
spawned at such great distances from the Gulf of Maine
(e.g., Gulf of Mexico or Mediterranean Sea). There are
several scenarios that could explain this finding.
Scenario I: natural variation
Large and small bluefin tuna could be arriving from the
same previous location with differences in reproductive
state reflecting normal variation in resorption time for fish
of different sizes/ages. Given that energetic reserves are
likely to be in short supply after spawning and migration,
rapid resorption of oocytes and follicles by all size classes
seems likely. Furthermore, we would expect to see a
greater proportion of larger fish rather than smaller fish
with unresorbed follicles, as the former should have greater
endogenous energy reserves.
Scenario II: arrival from different areas
Alternatively, different size/age classes may be arriving in
the Gulf of Maine from different spawning locations.
Similar results have been found for Pacific bluefin (Thun-
nus orientalis), which segregate to different spawning re-
gions based on size (Itoh 2006). Older, experienced
spawners may be coming from the Gulf of Mexico region
or more remote locations such as the Windward Passage
and other Caribbean areas, where adult bluefin tuna ‘‘in
near spawning condition’’ were documented in the RV
Crawford Cruise 62 of 19 April–8 June 1961 (Rathjen
1961). Smaller mature-inactive fish may have come from a
spawning location closer to the Gulf of Maine, lacking the
time to completely resorb vitellogenic oocytes. Although
the Gulf Stream margin has been dismissed as an important
spawning area for bluefin tuna because few larvae have
been found there (McGowan and Richards 1989), bluefin
spawning on its southern extension, the Loop Current, is
well established (Richards 1976). Mather hypothesized
(1973) that young bluefin tuna reaching maturity for the
first time may spawn on the northern edge of the Gulf
Stream, or in its warm core eddies. In tagging studies ini-
tiated in the Gulf of Maine, F. Royer et al. (unpublished
data) discerned seasonal patterns of behavior, and found
that some bluefin tuna moved from wintering areas off the
northeast continental slope to areas along the Gulf Stream
edge and Sargasso Sea during the spawning season of
April–June. Smaller mature-inactive fish may be coming
from this region rather than the Gulf of Mexico. It is also
possible that some of the sampled fish may have Medi-
terranean origins (Rooker and Secor 2004; Block et al.
2005), which mature at a younger age (3–5 years) than
members of the Western stock. This may explain why 7–8-
year-old fish in our sample were mature, but not why those
signs were detectable so far away from Mediterranean
spawning grounds
Scenario III: skipping a reproductive event
A third scenario may be that not all fish are spawning
annually, a hypothesis advanced to explain the presence of
spawning sized bluefin tuna located in the central north
Atlantic and elsewhere during the presumed spawning
period (Lutcavage et al. 1999; Wilson et al. 2005; Sibert
et al. 2006). Mature sized bluefin tuna sampled at feeding/
wintering grounds during the spawning season in the
Mediterranean had ovaries with no signs of previous or
imminent spawning, but displayed high levels of vitello-
genic atresia (Corriero et al. 2003), similar to mature fish
from our study, which has often been interpreted as a sign
of cessation of spawning or failure to attain final oocyte
maturation (Hunter et al. 1986; Mylonas et al. 1997; Farley
and Davis 1998; Schaefer 1998). Smaller mature-inactive
fish may have skipped spawning and so did not travel to
remote spawning locations.
Recent models on the Northeast Arctic cod stock, also a
long-lived, migratory, iteroparous species, showed that
skipped spawning was quite common, especially among
smaller size classes and younger fish, and was more com-
mon among females, and when feeding conditions were
poor and the energetic cost of migration was high
Mar Biol
123

(Jorgensen et al. 2006). Rideout et al. (2005) lists 21 spe-
cies in which skipped spawning has been documented, and
cites poor nutritional condition of non-spawners as the
most common reason. This phenomenon may be quite
common, but often goes undocumented due to the diffi-
culties in identifying non-reproductive individuals (Rideout
et al. 2005). Evidence from captive breeding experiments
(Lioka et al. 2000) and long-term historical data sets on the
Mediterranean trap fishery (Royer and Fromentin 2006)
suggests that some bluefin tuna may have a 2–3 year
spawning cycle, contrary to the common presumption of an
annual reproductive cycle.
Spatial-temporal patterns
Evidence for a more varied spawning cycle than the cur-
rently accepted paradigm suggests can be seen by exam-
ining historical records for the months of January–June.
Reproductive condition was recorded for 49 bluefin tuna
sampled during exploratory longline cruises conducted by
the MV Delaware in the 1960s (Wilson 1965). Although
qualitative in nature, gonad status estimated by the chief
scientist (P. Wilson, unpublished data, see Appendix 1)
indicates fish sampled along the Gulf Stream edge and
adjacent waters (Fig. 4) in March and April (mean CFL
198 ± 12.3 cm) were found in a mature or mature-inactive
state, including spent, ripening, and running (males, milt)
(Wilson 1965). In contrast, studies on bluefin tuna of
similar size sampled by coastal fisheries (with the excep-
tion of males in this study), have found little evidence of
active spawning. Seventeen fish (164–226 cm CFL,
x = 203 cm, males, 207 cm, females) sampled in January–
March 1995 off coastal North Carolina showed no signs of
spawning activity (Belle 1996), however, most were cap-
tured in winter sea surface temperatures <24�C (Fig. 4).
Baglin’s (1982) sampling in the Mid-Atlantic Bight found
no evidence of spawning activity in June, July and August
but all fish sampled were assigned age 7 or younger.
Conclusions
The range of post-reproductive states present well outside
of known spawning areas from this study and from his-
torical cruise data combined with recent electronic tagging
results lends evidence to a more asynchronous model for
bluefin tuna reproduction than previously assumed. Males
of commercial sizes (185–263 cm CFL) and small females
(185–235 cm CFL) arriving in June and July in mature or
mature-inactive condition may indicate that spawning has
occurred outside of known spawning areas, or spawning
has been omitted during the last maturity cycle. This result
is particularly striking given that the mean age of elec-
tronically tagged bluefin tuna present on the spawning
grounds are ages 11 and above (‡241 cm CFL) (Block
et al. 2005) and recent analyses on longline data in the Gulf
of Mexico estimate the age of 50% maturity to be 12 years
(Diaz and Turner 2006), although this finding may be
highly biased by size selectivity of longline sampling
(Davis and Farley 2001; Corriero et al. 2003). The exis-
tence of oceanic spawning areas remains a possibility, and
must be addressed by more extensive sampling, and a
complete understanding of the bluefin tuna reproductive
schedule. Sampling that includes fish smaller than 185 cm
CFL should be included to determine the lower size/age
limit of this maturity curve.
Modeling approaches on tuna (Fromentin and Fonteneau
2001) and other species (Stearns and Crandall 1984; Gar-
rod and Horwood 1984; Gunderson 1997; Jorgensen et al.
2006) and applied studies (Hearn and Polacheck 2003)
have shown that life history traits like size and age at
maturity shift in response to environmental stress such as
low food availability and fishing pressure. Age 8 (200 cm
CFL) has been assigned as the age of 100% knife-edge
maturity for western Atlantic bluefin tuna based on sizes
observed on known spawning grounds (Turner et al. 1991)
and not on histological evidence, an approach that has been
criticized for its non-quantitative nature (Schaefer 2001).
Determination of size/age at maturity depends first on
accurately assigning age, which has proven difficult for this
species, particularly in the larger size classes (Turner and
Terceiro 1994). A recent study on bluefin tuna sampled in
Canadian waters indicates that the growth curve most
commonly used to assign ages for the western Atlantic
stock may have shifted (Neilson and Campana 2006),
suggesting that growth curves need to be revised for this
stock (Restrepo et al. 2006).
Our data did not allow us to find a significant link be-
tween body condition and reproductive state but condition
should not be ruled out as a possible factor for determining
differences in migration routes or reproductive status. We
are currently developing a more precise index of condition
based on % lipid in muscle tissue to provide a more
accurate measure of energetic state. Future work will in-
clude muscle tissue biopsies and/or blood samples taken
from electronically tagged individuals which could prove
useful in determining whether sex, (Susca et al. 2000;
Heppell and Sullivan 2000) and/or lipid content of muscle
correlates with migratory and reproductive behavior or
spatial range as has been found in herring (Rajasilta 1992)
and European eels (Larsson et al. 1990)
Acknowledgments We thank Mike Blanchard, Matt Bunnell, John
Caldwell, Bill Chaprales, Rocky Chase, Scott Drabinowicz, Mark
Godfried, Eric and John Hesse, Jeff Tutein, Dave and Greg Wa-
Mar Biol
123
linski, Cape Quality Bluefin and Fresh Water Fish for collecting
samples. We also thank Jennifer Bowdoin and Jennifer Albright for
help with sampling, Kurt Schaefer and Antonio Medina for advice
on histology, Chris Bridges for hormonal analysis, Ben Galuardi for
spatial analysis, and Frank Cyganowski and the late Peter C. Wilson
for use of unpublished materials. This work was supported by
NOAA Grant NA04NMF4550391 to M. Lutcavage. All work was
done under compliance with UNH IACUC and NOAA Exempted
Fishing Permits.
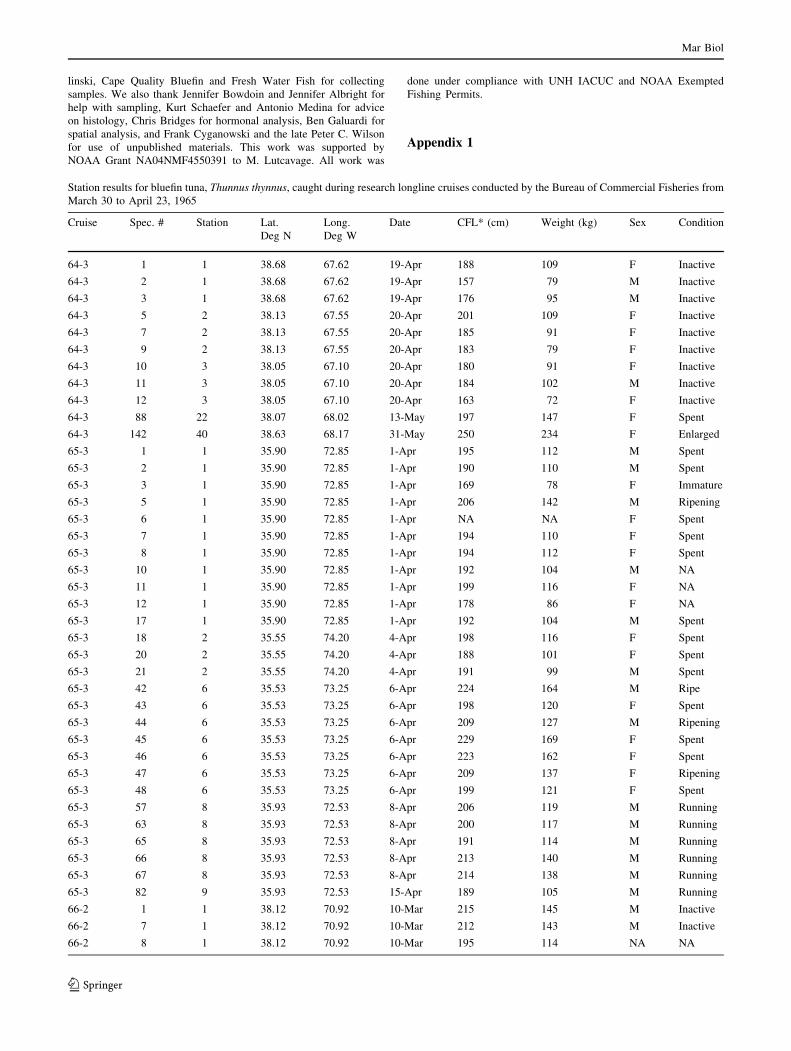
Appendix 1
Station results for bluefin tuna, Thunnus thynnus, caught during research longline cruises conducted by the Bureau of Commercial Fisheries from
March 30 to April 23, 1965
Cruise Spec. # Station Lat.
Deg N
Long.
Deg W
Date CFL* (cm) Weight (kg) Sex Condition
64-3 1 1 38.68 67.62 19-Apr 188 109 F Inactive
64-3 2 1 38.68 67.62 19-Apr 157 79 M Inactive
64-3 3 1 38.68 67.62 19-Apr 176 95 M Inactive
64-3 5 2 38.13 67.55 20-Apr 201 109 F Inactive
64-3 7 2 38.13 67.55 20-Apr 185 91 F Inactive
64-3 9 2 38.13 67.55 20-Apr 183 79 F Inactive
64-3 10 3 38.05 67.10 20-Apr 180 91 F Inactive
64-3 11 3 38.05 67.10 20-Apr 184 102 M Inactive
64-3 12 3 38.05 67.10 20-Apr 163 72 F Inactive
64-3 88 22 38.07 68.02 13-May 197 147 F Spent
64-3 142 40 38.63 68.17 31-May 250 234 F Enlarged
65-3 1 1 35.90 72.85 1-Apr 195 112 M Spent
65-3 2 1 35.90 72.85 1-Apr 190 110 M Spent
65-3 3 1 35.90 72.85 1-Apr 169 78 F Immature
65-3 5 1 35.90 72.85 1-Apr 206 142 M Ripening
65-3 6 1 35.90 72.85 1-Apr NA NA F Spent
65-3 7 1 35.90 72.85 1-Apr 194 110 F Spent
65-3 8 1 35.90 72.85 1-Apr 194 112 F Spent
65-3 10 1 35.90 72.85 1-Apr 192 104 M NA
65-3 11 1 35.90 72.85 1-Apr 199 116 F NA
65-3 12 1 35.90 72.85 1-Apr 178 86 F NA
65-3 17 1 35.90 72.85 1-Apr 192 104 M Spent
65-3 18 2 35.55 74.20 4-Apr 198 116 F Spent
65-3 20 2 35.55 74.20 4-Apr 188 101 F Spent
65-3 21 2 35.55 74.20 4-Apr 191 99 M Spent
65-3 42 6 35.53 73.25 6-Apr 224 164 M Ripe
65-3 43 6 35.53 73.25 6-Apr 198 120 F Spent
65-3 44 6 35.53 73.25 6-Apr 209 127 M Ripening
65-3 45 6 35.53 73.25 6-Apr 229 169 F Spent
65-3 46 6 35.53 73.25 6-Apr 223 162 F Spent
65-3 47 6 35.53 73.25 6-Apr 209 137 F Ripening
65-3 48 6 35.53 73.25 6-Apr 199 121 F Spent
65-3 57 8 35.93 72.53 8-Apr 206 119 M Running
65-3 63 8 35.93 72.53 8-Apr 200 117 M Running
65-3 65 8 35.93 72.53 8-Apr 191 114 M Running
65-3 66 8 35.93 72.53 8-Apr 213 140 M Running
65-3 67 8 35.93 72.53 8-Apr 214 138 M Running
65-3 82 9 35.93 72.53 15-Apr 189 105 M Running
66-2 1 1 38.12 70.92 10-Mar 215 145 M Inactive
66-2 7 1 38.12 70.92 10-Mar 212 143 M Inactive
66-2 8 1 38.12 70.92 10-Mar 195 114 NA NA
Mar Biol
123
References
Abascal FJ, Megina C, Medina A (2004) Testicular development in
migrant and spawning bluefin tuna (Thunnus thynnus (L.) from
the eastern Atlantic and Mediterranean. Fish Bull 102:407–417
Anderson R, Neumann R (1996) Length, weight and associated
structural indices. In: Murphy B, Willis D (eds) Fisheries
techniques, 2nd ed. American Fisheries Society, Bethesda, pp
447–481
Anonymous (2003) Report of the sixth GFCM-ICCAT meeting on
stocks of large pelagic fishes in the Mediterranean. Col Vol Sci
Pap ICCAT 55:1–84
Baglin RE Jr (1980) Length–weight relationships of western Atlantic
bluefin tuna, Thunnus thynnus. Fish Bull 77:995–1000
Baglin RE Jr (1982) Reproductive biology of western Atlantic bluefin
tuna. Fish Bull 80:121–134
Belle S (1996) Biological sampling of bluefin tuna off Cape Hatteras,
North Carolina. Final report to the New England Aquarium
Corporation, NOAA req no. 43AANF503279, Boston, 12 p
Bigelow HB, Schroeder WC (1953) Fishes of the Gulf of Maine. Fish
Bull 53:1–577
Block BA, Dewar H, Blackwell SB, Williams TD, Prince ED, Farwell
CJ, Boustany A, Teo SL, Seitz A, Walli A, Fudge D (2001)
Migratory movements, depth preferences, and thermal biology of
Atlantic bluefin tuna. Science 293:1310–1314
Block BA, Teo SLH, Walli A, Boustany A, Stokesbury MJW, Farwell
C, Weng K, Dewar H, Williams TD (2005) Electronic tagging
and population structure of Atlantic bluefin tuna. Nature
434:1121–1126
Burton MP, Idler DR (1984) The reproductive cycle in winter
flounder, Pseudopleuronectes americanus (Walbaum). Can J
Zool 62:2563–2567
Burton MP, Idler DR (1987) An experimental investigation of the
non-reproductive post-mature state in winter flounder. J Fish
Biol 30:643–650
Caddy JF, Butler MJA (1976) Recent catch trends and age compo-
sitions in Canadian coastal fisheries for giant bluefin tuna
(Thunnus thynnus), and their relevance to assessment of the
Northwest Atlantic large fish stock. ICCAT Col Vol Sci Pap
5:244–257
Chase B (2002) Differences in the diet of Atlantic bluefin tuna
(Thunnus thynnus) at five seasonal feeding grounds on the New
England continental shelf. Fish Bull 100:168–180
Corriero A, DeSantis S, DeFlorio M, Acone F, Bridges CR, de la
Serna JM, Megelofonou P, DeMetrio G (2003) Histological
investigation on the ovarian cycle of the bluefin tuna in the
western and central Mediterranean. J Fish Biol 62:1–12
Corriero A, Karakulak S, Santamaria N, Deflorio M, Spedicato D,
Addis P, Desantis S, Cirillo F, French-Farrugia A, Vassallo-
Agius R, de la Serna J, Oray Y, Cau A, Megalofonou P,
DeMetrio G (2005) Size and age at sexual maturity of female
bluefin tuna (Thunnus thynnus L. 1758) from the Mediterranean
Sea. J Appl Ichthyol 21:483–486
Crane J (1936) Notes on the biology and ecology of giant tuna
Thunnus thynnus, L. observed at Portland, Maine. Zoologica
212:207–212
Davis TLO, Farley JH (2001) Size distirbution of southern bluefin
tuna (Thunnus maccoyii) by depth on their spawning ground.
Fish Bull 99:381–386
Diaz GA, Turner SC (2006) Size frequency distribution and age
estimation of BFT in the Gulf of Mexico during the spawning
season. ICCAT Col Vol Sci Pap SCRS/2006/090
Dragovich A (1970) The food of bluefin tuna, (Thunnus thynnus) in
the western North Atlantic ocean. Trans Am Fish Soc 99:726–
731
Estrada JA, Lutcavage M, Thorrold SR (2005) Diet and trophic
position of Atlantic bluefin tuna (Thunnus thynnus) inferred from
stable carbon and nitrogen isotope analysis. Mar Biol 147:37–45
Farley JH, Davis TLO (1998) Reproductive dynamics of southern
bluefin tuna, Thunnus maccoyii. Fish Bull 96:223–236
Frade F (1937) Recherches sur la maturiete sexuelle du thon rouge.
Bull Soc Portugaise Sci Naturelles Lisbonne 12:243–250
Fromentin JM, Fonteneau A (2001) Fishing effects and life history
traits: a case-study comparing tropical versus temperate tunas.
Fish Res 53:133–150
Fromentin JM, Powers JE (2005) Atlantic bluefin tuna: population
dynamics, ecology, fisheries and management. Fish Fish 6:281–
306
Garrod DJ, Horwood JW (1984) Reproductive strategies and the
response to exploitation. In: Potts GW, Wooton RJ (eds) Fish
reproduction. Academic, London, pp 367–384
Golet W, Cooper A, Campbell R, Lutcavage M (2006) Decline in
condition factor of Atlantic bluefin tuna in the Gulf of Maine.
Fish Bull (in press)
Gunderson DR (1997) Trade-off between reproductive effort and
adult survival in oviparous and viviparous fishes. Can J Fish
Aquat Sci 54:990–998
Hearn WS, Polacheck T (2003) Estimating long-term growth-rate
changes of southern bluefin tuna (Thunnus maccoyii) from two
periods of tag-return data. Fish Bull 101:58–74
Appendix 1 continued
Cruise Spec. # Station Lat.
Deg N
Long.
Deg W
Date CFL* (cm) Weight (kg) Sex Condition
66-2 9 1 38.12 70.92 10-Mar 211 129 M Some milt
66-2 10 1 38.12 70.92 10-Mar 205 128 M Inactive
66-2 11 1 38.12 70.92 10-Mar 218 144 M Inactive
66-2 12 1 38.12 70.92 10-Mar 210 133 M Inactive
66-2 42 4 38.33 65.53 13-Mar 221 145 M Some milt
66-2 49 4 38.33 65.53 13-Mar 222 154 M Inactive
66-2 54 6 38.28 62.35 15-Mar 192 104 F Ripening
66-2 63 6 38.28 62.35 15-Mar 201 120 F Ripening
66-2 108 9 37.23 65.90 27-Mar 196 116 M NA
* Fork length (FL) converted to curved FL after Parrack and Phares (1979) for comparison with values in this study
Mar Biol
123

Heppell SA, Sullivan CV (1999) Gag (Mycteroperca microlepsis)
vitellogenin: purification, characterization, and use for enzyme-
linked immunosorbent assay (ELIZA) of female maturity in
three species of grouper. Fish Physiol Biochem 20:361–374
Heppell SA, Sullivan CV (2000) Identification of gender and
reproductive maturity in the absence of gonads: muscle tissue
levels of sex steroids and vitellogenin in gag (Mycteropercamicrolepsis). Can J Fish Aquat Sci 57:148–159
Hislop JRG, Robb AP, Gauld JA (1978) Observations on effects of
feeding level on growth and reproduction in haddock, Melan-ogrammus aeglefinus (L.) in captivity. J Fish Biol 13:85–98
Hunter JR, Macewicz BJ (1985) Rates of atresia in the ovary of
captive and wild northern anchovy, Engraulis mordax. Fish Bull
83:119–136
Hunter JR, Macewicz BJ, Sibert JR (1986) The spawning frequency
of skipjack tuna, Katsuwonus pelamis, from the south Pacific.
Fish Bull 84:895–903
Itoh T (2006) Sizes of adult bluefin tuna Thunnus orientalis in
different areas of the western Pacific Ocean. Fish Sci 72:53–62
Jorgensen C, Ernande B, Fiksen O, Dieckmann U (2006) The logic of
skipped spawning in fish. Can J Fish Aquat Sci 63:200–211
Larsson P, Hamrin S, Okla L (1990) Fat content as a factor inducing
migratory behavior in the eel (Anguilla anguilla L.) to the
Sargasso Sea. Naturwissenshaften 77:488–490
Lioka C, Kani K, Nhhala H (2000) Present status and prospects of
technical development of tuna farming. Cahiers Options Medit-
erraneennes 47:275–285
Lutcavage M, Kraus S, Hoggard W (1997) Aerial assessment of
bluefin tuna in the Bahama Banks-Straits of Florida, 1995. Fish
Bull 95:300–310
Lutcavage ME, Brill RW, Skomal GB, Chase BC, Howey PW (1999)
Results of pop-up satellite tagging of spawning size class fish in
the Gulf of Maine: do North Atlantic bluefin tuna spawn in the
mid-Atlantic? Can J Fish Aquat Sci 56:173–177
Mather FJ III (1973) The bluefin tuna situation. In: Proceedings
of 16th annual international game fish research conference,
pp 93–120
Mather FJ III, Mason JM, Jones AC (1995) Historical document: life
history and fisheries of Atlantic bluefin tuna. NOAA Technical
Memorandum, NMFS SEFSC-370
McCullagh P, Nelder JA (1989) Generalized linear models, 2nd edn.
Chapman Hall/CRC, Florida
McGowan MF, Richards WJ (1989) Bluefin tuna, Thunnus thynnus,
larvae in the Gulf Stream off the southeastern United States:
shipboard observations of their movement. Fish Bull 87:615–631
Medina A, Abascal FJ, Megina C, Garcıa A (2002) Stereological
assessment of the reproductive status of female Atlantic northern
bluefin tuna during migration to the Mediterranean spawning
grounds through the Strait of Gibraltar. J Fish Biol 60(1):203–
217
Mourente G, Megina C, Dıaz-Salvago E (2002) Lipids in female
northern bluefin tuna (Thunnus thynnus thynnus L.) during
sexual maturation. Fish Physiol Biochem 24:351–363
Mylonas CC, Woods LC III, Zohar Y (1997) Cyto-histological
examination of post-vitellogenesis and final oocyte maturation in
cative-reared striped bass. J Fish Biol 50:34–49
Neilson JD, Campana SE (2006) Pilot study of bluefin tuna age
validation. ICCAT Col Vol Sci Pap SCRS/2006/077
Nemerson D, Berkeley S, Safina C (2000) Spawning site fidelity in
Atlantic bluefin tuna, Thunnus thynnus : the use of size-
frequency analysis to test for the presence of migrant east
Atlantic bluefin tuna on Gulf of Mexico spawning grounds. Fish
Bull 98:118–126
NRC (1994) An assessment of Atlantic bluefin tuna. National
Academy Press, Washington, DC
Parrack ML, Phares PL (1979) Aspects of the growth of Atlantic
bluefin tuna determined from mark-recapture data. ICCAT Col
Vol Sci Pap 8:356–366
Quinn T II, Deriso R (1999) Quantitative fish dynamics. Oxford
University Press, New York, pp 86–207
Rajasilta M (1992) Relationship between food, fat, sexual maturation,
and spawning time of Baltic herring (Clupea harengus membras)
in the Archipelago Sea. Can J Fish Aquat Sci 49:644–654
Rathjen WF (1961) Memorandum, US Dept. of the Interior, Bureau of
Commercial Fisheries, Exploratory Fishing and Gear Research
Base, Gloucester, May 9, 1961
Restrepo VR, Roderıguez-Marın E, Cort JL, Rodrıguez-Cabello C(2006) Are the growth curves currently used for Atlantic bluefin
tuna statistically different? ICCAT Col Vol Sci Pap SCRS 2006/
079
Richards WJ (1976) Spawning of bluefin tuna (Thunnus thynnus) in
the Atlantic ocean and adjacent seas. ICAAT Col Vol Sci Pap
2:267–278 (SCRS/75/97)
Rideout RM, Burton MPM, Rose GA (2000) Observations on mass
atresia and skipped spawning in northern Atlantic cod from
Smith Sound, Newfoundland. J Fish Biol 57:1429–1440
Rideout RM, Rose GA, Burton MPM (2005) Skipped spawning in
female iteroparous fishes. Fish Fish 6:50–72
Rijnsdorp AD (1990) The mechanism of energy allocation over
reproduction and somatic growth in female North Sea plaice,
Pleuronectes platessa L. Neth J Sea Res 25:279–290
Rivas LR (1954) A preliminary report on the spawning of the western
North Atlantic bluefin tuna (Thunnus thynnus) in the Straits of
Florida. Bull Mar Sci Gulf Caribbean Miami 4:302–322
Rivas LR (1976) Variation in sex ratio, size differences between
sexes, and change in size and age composition in Western North
Atlantic giant bluefin tuna (Thunnus thynnus) ICCAT Col Vol
Sci Pap 5:297–301
Rivas LR (1978) Preliminary models of annual life history cycles of
the north Atlantic bluefin tuna. In: Sharp GD, Dizon AD (eds)
The physiological ecology of tunas. Academic, New York, pp
369–393
Rodrıguez-Roda J (1963) Investigaciones sobre tunidos. Publicaci-
ones Tecnicas de la Junta de Estudios de Pesca. Direccion
General de Pesca Maritıma, Madrid 2:39–77
Rodrıguez-Roda J (1964) Biologıa del atun, Thunnus thynnus (L.), de
la costa sudatlantica de Espana. Invest Pesqu 25:33–146
Rooker JR, Secor DH (2004) Stock structure and mixing of Atlantic
bluefin tuna: evidence from stable d13C and d18O isotopes in
otoliths. Col Vol Sci Pap ICCAT 56:1115–1120
Royer F, Fromentin JM (2006) Environmental noise in spawning
areas: the case of Atlantic bluefin tuna (Thunnus thynnus). Fish
Oceanogr (in press)
Schaefer KM (1996) Spawning time, frequency, and batch fecundity
of yellowfin tuna, Thunnus albacares, near Clipperton Atoll in
the eastern Pacific Ocean. Fish Bull 94:98–112
Schaefer KM (1998) Reproductive biology of the yellowfin tuna
(Thunnus albacares) in the eastern Pacific ocean. Bull IATTC
21:489–528
Schaefer KM (2001) Reproductive biology of tunas. In: Block BA,
Stevens ED (eds) Tuna: physiology, ecology and evolution.
Academic, San Diego, pp 225–271
Sibert JR, Lutcavage ME, Nielsen A, Brill RW, Wilson SG (2006)
Inter-annual variation in large scale movement of Atlantic
bluefin tuna (Thunnus thynnus) determined from popup satellite
archival tags. Can J Fish Aquat Sci (in press)
Sidak Z (1967) Rectangular confidence regions for the means of
multivariate normal distributions. J Am Stat Assoc 62:626–633
Stearns SC, Crandall RE (1984) Plasticity for age and size at sexual
maturity: a life history response to unavoidable stress. In: Potts
Mar Biol
123

GW, Wooton RJ (eds) Fish reproduction. Academic, London,
pp 13–33
Stokesbury MJW, Teo SLH, Seitz A, O’Dor RK, Block BA (2004)
Movement of Atlantic bluefin tuna (Thunnus thynnus) as
determined by satellite tagging experiments initiated off New
England. Can J Aquat Sci 61:1976–1987
Susca V, Corriero A, Bridges CR, DeMetrio G (2000) Study of the
sexual maturity of female bluefin tuna: purification and partial
characterization of vitellogenin and its use in an enzyme-linked
immunosorbent assay. J Fish Biol 58:815–831
Susca V, Corriero A, Deflorio M, Bridges CR, DeMetrio G (2001b)
New results on the reproductive biology of the bluefin tuna
(Thunnus thynnus) in the Mediterranean. Col Vol Sci Pap
ICCAT 52:745–751
Tiews K (1963) Synopsis of biological data on bluefin tuna Thunnusthynnus (Linnaeus) 1758 (Atlantic and Mediterranean). FAO fish
rep synopsis no.56 2:422–481
Turner S, Terceiro M (1994) Estimation of west Atlantic bluefin tuna,
Thunnus thynnus, age composition with length composition
analysis. ICCAT Col Vol Sci Pap 42:173–180
Turner S, Restrepo V, Elkund A (1991) A review of the growth of
Atlantic bluefin tuna, Thunnus thynnus. ICCAT Col Vol Sci Pap
35:271–293
Wilson P (1965) Cruise Report, MV Delaware Cruise 65-3, March
30–April 23, 1965. Pelagic-oceanic explorations. May 20, 1965.
USFWS Bureau of Commercial Fisheries
Wilson S, Lutcavage M, Brill R, Genovese M, Cooper A, Everly A
(2005) Movements of bluefin tuna (Thunnus thynnus) in the
northwestern Atlantic ocean recorded by pop-up satellite archi-
val tags. Mar Biol 146:409–423
Mar Biol
123




















