
Reproductive Parameters and Productivity of the Yellow-Shouldered Parrot on Margarita Island,...
15
REPRODUCTIVE PARAMETERS AND PRODUCTIVITY OF THE YELLOW-SHOULDERED PARROT ON MARGARITA ISLAND, VENEZUELA: A LONG-TERM STUDY VIRGINIA SANZ 1 AND ADRIANA RODRI ´ GUEZ-FERRARO 2 PROVITA, Apartado 47552, Caracas 1041 – A, Venezuela Abstract. We studied the breeding biology of the Yellow-shouldered Parrot (Amazona barbadensis) on Margarita Island from March-August, 1990 to 1999. The timing of the different phases (egg-laying, hatching, fledging) was consistent over the years, except during 1998 when all phases were delayed. The average clutch size was 3.38 6 0.78 eggs per nest, with a range of one to five eggs per clutch, and most eggs survived until hatching (3.36 6 0.80 eggs per nest). Total clutch size and hatching success of this species on Margarita Island are among the highest in the genus Amazona, suggesting the Yellow- shouldered Parrot has a higher reproductive potential than other species of the genus. We detected interannual differences for some of the reproductive parameters, all in 1998, a year with an extreme drought. Egg losses totaled 20% and were caused by hatching failure, predation, and human disturbance. Forty-nine percent of nestlings were lost, mainly due to poaching. The number of fledglings per nesting pair averaged 1.27 6 1.61, but varied greatly among years. Thus, in relation to the average total clutch laid, each pair lost an average of 62% of its initial reproductive investment. Key words: Amazona barbadensis, breeding biology, Margarita Island, mortality factors, productivity, Venezuela, Yellow-shouldered Parrot. Para ´ metros Reproductivos y Productividad de Amazona barbadensis en la Isla de Margarita, Venezuela: un Estudio a Largo Plazo Resumen. Estudiamos la biologı ´a reproductiva de la cotorra Amazona barbadensis en la isla de Margarita entre marzo y agosto desde 1990 hasta 1999. Las fechas de las diferentes fases (puesta de huevos, eclosio ´ n, vuelo de pichones) fueron consistentes a trave ´s de los an ˜ os, excepto durante 1998, cuando todas las fases se atrasaron. El promedio del taman ˜o de nidada para la de ´cada fue de 3.38 6 0.78 huevos por nido, con un intervalo de uno a cinco huevos por nidada. El taman ˜ o promedio de nidada al momento de la eclosio ´n (3.36 6 0.80 huevos por nido) indica que la mayorı ´a de los huevos sobrevivio ´ hasta la eclosio ´n. El taman ˜o total de la nidada y el e ´xito de eclosio ´n de la especie en la isla de Margarita son de los ma ´ s altos entre las especies del ge ´nero Amazona, sugiriendo que A. barbadensis tiene un potencial reproductivo ma ´ s elevado que el de otras especies del ge ´nero. Detectamos diferencias interanuales en algunos de los para ´ metros reproductivos, todas correspondientes a 1998, un an ˜ o con una extrema sequı ´a. Las pe ´rdidas de huevos representaron un 20% durante el perı ´odo 199021999 y fueron causadas por fallos en la eclosio ´ n, depredacio ´ n y perturbaciones humanas. El 49% de los pichones se perdieron, principalmente por la extraccio ´n ilegal. El nu ´ mero promedio de volantones por pareja reproductiva fue 1.27 6 1.61, y este valor tambie ´n vario ´ considerablemente a trave ´s de los an ˜ os. Esto indica que, en relacio ´n con el taman ˜ o promedio de la nidada, cada pareja perdio ´ en promedio el 62% de su inversio ´ n reproductiva inicial. INTRODUCTION Population dynamics models are important conservation tools because they can predict the behavior of populations over time (Suther- land 2000). In fact, demographic models are commonly used to make management decisions regarding endangered species (Beissinger and Westphal 1998). Frequently, these models are based on the estimation of parameters related to mortality rate and fecundity. As the former Manuscript received 30 November 2004; accepted 4 August 2005. 1 Present address: Centro de Ecologı ´a, Instituto Venezolano de Investigaciones Cientı ´ficas, Apdo. 21827 Caracas, 1020-A, Venezuela. E-mail: [email protected] 2 Present address: Department of Biology, R223, University of Missouri-St. Louis, 8001 Natural Bridge Road, St. Louis, MO 63121-4499, USA. E-mail: [email protected] The Condor 108:178–192 # The Cooper Ornithological Society 2006 [178]
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Reproductive Parameters and Productivity of the Yellow-Shouldered Parrot on Margarita Island,...

REPRODUCTIVE PARAMETERS AND PRODUCTIVITY OF THEYELLOW-SHOULDERED PARROT ON MARGARITA ISLAND,
VENEZUELA: A LONG-TERM STUDY
VIRGINIA SANZ1
AND ADRIANA RODRIGUEZ-FERRARO2
PROVITA, Apartado 47552, Caracas 1041 – A, Venezuela
Abstract. We studied the breeding biology of the Yellow-shouldered Parrot (Amazonabarbadensis) on Margarita Island from March-August, 1990 to 1999. The timing of thedifferent phases (egg-laying, hatching, fledging) was consistent over the years, exceptduring 1998 when all phases were delayed. The average clutch size was 3.38 6 0.78 eggsper nest, with a range of one to five eggs per clutch, and most eggs survived until hatching(3.36 6 0.80 eggs per nest). Total clutch size and hatching success of this species onMargarita Island are among the highest in the genus Amazona, suggesting the Yellow-shouldered Parrot has a higher reproductive potential than other species of the genus. Wedetected interannual differences for some of the reproductive parameters, all in 1998, a yearwith an extreme drought. Egg losses totaled 20% and were caused by hatching failure,predation, and human disturbance. Forty-nine percent of nestlings were lost, mainly dueto poaching. The number of fledglings per nesting pair averaged 1.27 6 1.61, but variedgreatly among years. Thus, in relation to the average total clutch laid, each pair lost anaverage of 62% of its initial reproductive investment.
Key words: Amazona barbadensis, breeding biology, Margarita Island, mortalityfactors, productivity, Venezuela, Yellow-shouldered Parrot.
Parametros Reproductivos y Productividad de Amazona barbadensis
en la Isla de Margarita, Venezuela: un Estudio a Largo Plazo
Resumen. Estudiamos la biologıa reproductiva de la cotorra Amazona barbadensis enla isla de Margarita entre marzo y agosto desde 1990 hasta 1999. Las fechas de lasdiferentes fases (puesta de huevos, eclosion, vuelo de pichones) fueron consistentes a travesde los anos, excepto durante 1998, cuando todas las fases se atrasaron. El promedio deltamano de nidada para la decada fue de 3.38 6 0.78 huevos por nido, con un intervalo deuno a cinco huevos por nidada. El tamano promedio de nidada al momento de la eclosion(3.36 6 0.80 huevos por nido) indica que la mayorıa de los huevos sobrevivio hasta laeclosion. El tamano total de la nidada y el exito de eclosion de la especie en la isla deMargarita son de los mas altos entre las especies del genero Amazona, sugiriendo que A.barbadensis tiene un potencial reproductivo mas elevado que el de otras especies delgenero. Detectamos diferencias interanuales en algunos de los parametros reproductivos,todas correspondientes a 1998, un ano con una extrema sequıa. Las perdidas de huevosrepresentaron un 20% durante el perıodo 199021999 y fueron causadas por fallos en laeclosion, depredacion y perturbaciones humanas. El 49% de los pichones se perdieron,principalmente por la extraccion ilegal. El numero promedio de volantones por parejareproductiva fue 1.27 6 1.61, y este valor tambien vario considerablemente a traves de losanos. Esto indica que, en relacion con el tamano promedio de la nidada, cada parejaperdio en promedio el 62% de su inversion reproductiva inicial.
INTRODUCTION
Population dynamics models are importantconservation tools because they can predictthe behavior of populations over time (Suther-land 2000). In fact, demographic models are
commonly used to make management decisionsregarding endangered species (Beissinger andWestphal 1998). Frequently, these models arebased on the estimation of parameters relatedto mortality rate and fecundity. As the former
Manuscript received 30 November 2004; accepted 4 August 2005.1 Present address: Centro de Ecologıa, Instituto Venezolano de Investigaciones Cientıficas, Apdo. 21827
Caracas, 1020-A, Venezuela. E-mail: [email protected] Present address: Department of Biology, R223, University of Missouri-St. Louis, 8001 Natural Bridge
Road, St. Louis, MO 63121-4499, USA. E-mail: [email protected]
The Condor 108:178–192# The Cooper Ornithological Society 2006
[178]

parameter is difficult to estimate (Beissinger2002), such models are usually based onfecundity data and, generally, indirect methodsare used to quantify them (Caughley 1978). Thevalidity of such models depends on preciseparameter estimation and the accuracy withwhich those parameters represent the behaviorof the species over time (Beissinger andWestphal 1998). Reproductive parameters andpopulation productivity constitute especiallyrelevant information for endangered or com-mercially exploited species. Only by knowingreproductive parameters will it be possible toestimate and accurately predict the maximumpotential of a species’ population to grow andrecover (Sutherland 2000). Determining thecritical factors that affect the breeding cycle iscrucial to deciding which aspects should bemanaged, and at what level, to achieve a certainpopulation size.
The Psittacidae family has one of the highestproportions of threatened species, with onethird of all known parrot species considered atrisk (Bennett and Owens 1997, BirdLife In-ternational 2000, Snyder et al. 2000). Repro-ductive information for threatened species isfundamental for developing recovery programs.In Australia, population models of severalparrot species have been produced based onlife tables generated with annual productivityand survival data (Saunders 1980, 1986, Rowleyand Chapman 1991). Because of a lack of data,this is not possible for most of the parrotspecies in the rest of the world. Although therehas been a recent increase in studies of breedingbiology of Neotropical members of the genusAmazona (Snyder et al.1987, Gnam and Rock-well 1991, Enkerlin-Hoeflich 1995, Martuscelli1995, Koenig 2001, Seixas and Mourao 2002,Renton and Salinas-Melgoza 2004), most areshort term. Data derived from short-termstudies may be inadequate to formulate reliablepredictions from population models. Only twomore extensive studies have compiled sufficientdata to conduct population viability analysesfor two Amazona species (A. vittata and A.barbadensis, Lacy et al. 1989, Rodrıguez andRojas-Suarez 1994, Rodrıguez et al. 2004).
The Yellow-shouldered Parrot (Amazonabarbadensis) is one of the most endangeredparrot species in Venezuela because of illegalpet trade and habitat loss (Desenne and Strahl1991, Collar et al. 1994, Rodrıguez and Rojas-
Suarez 1995). Its total population size isestimated at 10 000 individuals and it isconsidered Vulnerable at a global level (Bird-Life International 2000). This species is patchilydistributed throughout the arid zones of thenorthern coast of Venezuela, and on theCaribbean islands of Margarita, La Blanquilla,and Bonaire (Netherlands Antilles; Forshaw1989, Collar et al. 1994, Juniper and Parr 1998).The Yellow-shouldered Parrot went extinct onAruba during the 1950s (Forshaw 1989), andprobably also from Curacao (Juniper and Parr1998). Although there are no records of itspresence on the last island, it probably livedthere considering the species’ pattern of geo-graphical distribution.
In this paper, we present data on the breedingbiology of the Yellow-shouldered Parrot onMargarita Island from 1990 to 1999, andanalyze the relationship between reproductiveparameters and environmental fluctuations. Ananalysis of the role of climate in reproduction isparticularly important on Margarita Islandbecause the precipitation is so unpredictable(Goldbrunner 1962). The main objectives of ourstudy were to determine the average values andyearly fluctuations of reproductive parametersand productivity, and to identify the factorsresponsible for total and partial nest losses.
METHODS
STUDY AREA
Our study was conducted in the MacanaoPeninsula (300 km2) of Margarita Island(10u519–11u119N and 63u469–64u249W), off thenortheastern coast of Venezuela (Fig. 1). Ele-vation of the Macanao Peninsula ranges fromsea level to 760 m. Average annual temperatureis 27uC, ranging from 24uC to 31uC; meanannual rainfall is 522 6 224 mm (range: 214–1164 mm), with a distinct dry season fromJanuary to May (Ministerio del Ambiente y losRecursos Naturales, unpubl. data). This semi-arid tropical climate favors a mixture of opencactus communities and thorn forest, withcolumnar cacti (Stenocereus griseus and Sub-pilocereus repandus) and legumes (Prosopisjuliflora, Caesalpinia coriaria, and Cercidiumpraecox) as the dominant vegetation, althoughseasonal riverbeds can support deciduous for-ests (Hoyos 1985). Currently these galleryforests are being drastically reduced because
PRODUCTIVITY OF YELLOW-SHOULDERED PARROTS 179

of open-pit sand mining activities, resulting inloss of nesting and roosting habitat for theYellow-shouldered Parrot.
The main study area included La Chica Creek(Fig. 1) and surrounding areas in the northeast ofthe peninsula between 0 and 100 m abovesea level. An open-pit sand mine is situated alongthe creek between 40 and 80 m above sea level.
NEST LOCATIONS AND MONITORING
Breeding biology data were collected as part ofa conservation program developed for thisspecies on Margarita Island from 1990 to1999. The main activities in this programinvolved nest monitoring and guarding nestsfrom poachers in La Chica Creek and nearbyareas. Nest guarding was initiated when ap-proximately 50% of the nestlings born ina season were 15 days old and lasted until thelast fledgling left the nest. Guarding of nestswas conducted from 06:00 to 19:00 seven daysa week, and additional night guarding was alsocarried out during some years (1997–1999) from20:00 to 02:00. During these surveillanceactivities some members of the research teamwalked along La Chica Creek looking forpoachers or their tracks, while others watchedthe nesting area from fixed points on hilltops.These strategies, however, did not prevent allpoaching because not all active nests could bewatched at the same time and every day.
During the first breeding season, active nestswere located based on information from localpeople. In following years, we carried outsystematic searches by checking trees withcavities at the beginning of the breeding season,including former nest sites. Each cavity wasinspected with a small flashlight and a mirrorand was considered active if we found a nestingfemale, eggs, or chicks. We recorded the dateand the status of the nest when an active nestwas found, and we checked nests every 2–3 days in order to determine nest fate and theduration of the incubation and nestling periods.These data were estimated annually and for thewhole decade.
From 1995 to 1999 we marked nestlings withaluminum color bands, using a different colorfor each year, which allowed us to recognize theage of fledglings at a distance. Fledglings can beeasily differentiated from adult parrots by iriscolor (brown or orange vs. bright scarlet red inadults) and the length of tail feathers, which areshorter in fledglings.
REPRODUCTIVE PARAMETERS
We estimated the following reproductive pa-rameters: total clutch laid, clutch size athatching, brood size hatched, and brood sizeat fledging, following Gnam and Rockwell(1991). All these parameters were calculatedannually and for the whole decade.
FIGURE 1. Location of the study site (La Chica Creek) in Macanao Peninsula, Margarita Island,northern Venezuela.
180 VIRGINIA SANZ AND ADRIANA RODRIGUEZ-FERRARO

FACTORS AFFECTINGREPRODUCTIVE SUCCESS
We distinguished between partial and total nestlosses. In the latter case, no egg hatched or nonestling fledged from a nest. When a nestlingdisappeared from the nest three or four daysbefore the estimated date of fledging with nosigns of poaching or predation, we consideredthat it had fledged successfully. This couldoverestimate reproductive success and produc-tivity, but the rate of recovery of the populationindicates that the error, if any, is small, and thisis a reasonable measure of fledging success.
When losses occurred (either partial or total),we recorded the most probable cause: infertileor unviable eggs (eggs that had not hatched oneweek to ten days after the expected hatchingdate), nest abandonment (10 days after theexpected hatching date no eggs had hatched),predation (missing or broken eggs without anyapparent cause, or nestlings gone from the nestwith feathers in the nest interior), humandisturbance (broken eggs or dead nestlingsbecause of stones or sticks thrown into thenest. As most of the nests are deep and dark,some poachers use this method to verify thepresence of a female or chicks to determinewhether a nest is active), poaching (nestlingsmissing from the nest with evidence of illegalextraction, e.g., human tracks, tree crotchesplaced in the nest tree to climb it, leaves of thespiny bromeliad [Bromelia humilis], used bypoachers to extract chicks from deep nests, onthe ground, or trees with machete or axe cutsnear the cavity), and deaths from unknowncauses (dead nestlings in the nest without anyexternal sign that allowed us to determine thecause of death).
For analysis, we separated the breedingperiod into two stages: prehatching and nest-ling. For each of these phases we estimated thepercentage of eggs or nestlings lost (number ofeggs or nestlings lost due to each cause dividedby the total clutch laid or number of nestlings athatch), and the percentage of total nest losses(number of nests lost in each stage divided bythe total number of active nests at the beginningof each stage). In both cases, percentages werecalculated annually and for the whole decade.We included data on nests from areas close toLa Chica Creek only during the prehatchingstage, when there was no effect of poachers.When analyzing causes of losses in the nestling
period we pooled data on natural and fosternests to increase sample size. During fosternests management, nestlings (confiscated fromnests at high risk of poaching or from nestswhere the brood was partially depredated) weretransferred to nests within the study area to beraised by ‘‘foster’’ parents, which had broodsizes lower than the population average (Al-bornoz et al. 1994, Sanz and Rojas-Suarez1997). As this technique could increase mortal-ity in the receptor nest, before pooling data wetested for the absence of significant differencesin mortality for each mortality factor betweenboth kinds of nests using Fisher’s exact tests.
PRODUCTIVITY
We calculated hatching success (proportion ofhatched eggs from the total number of eggslaid), fledging success (proportion of fledglingsfrom the total number of eggs laid), andnumber of fledglings per nesting pair, bothannually and for the whole decade. Thesemeasures were calculated for natural nests only,excluding the foster nests in the nestling period.
STATISTICAL ANALYSES
We used nonparametric (Kruskal-Wallis) anal-ysis of variance (Conover 1998) to examinewhether the average length of the incubationperiod and the average age of nestlings atfledging varied among years. When significantdifferences were detected, we also conductedchi-square contingency analyses. Differencesbetween causes of egg and nestling losses andtotal losses during the prehatching and nestlingstages were determined using chi-square tests(Siegel and Castellan 1995). Annual fluctua-tions of each of the causes of loss were alsotested with two-way contingency tables. Annualdifferences in reproductive parameters werecompared for different clutch sizes using two-way contingency tables, with the years and theclutch size categories (one–two, three, and four)as factors. The frequencies of one-, two-, andfive-egg clutches were low, thus we groupedone- and two-egg clutches together and did notconsider five-egg clutches for these analyses.When the chi-square values indicated signifi-cant differences, the identification of these wasdone using a standardized residual analysis(Siegel and Castellan 1995), with values ofstandardized residuals (st. resid.) greater than1.96 being significantly different.
PRODUCTIVITY OF YELLOW-SHOULDERED PARROTS 181

To analyze the influence of rainfall on clutchsize, we ran a stepwise multiple regressionbetween the average annual clutch size andthe following variables: 1) total rainfall of theprevious nonbreeding period, 2) total numberof rainy days of the previous nonbreedingperiod, 3) number of days with rainfall lessthan 6 mm (daily average of days when anyprecipitation occurred) during the nonbreedingperiod, and 4) total rainfall of the previous year(from January to December). We defined thenonbreeding period as the time period startingon August of the previous year until the monthin which egg-laying commenced in the analyzedyear. The total rainfall of the previous non-breeding period and the number of days withrainfall less than 6 mm were correlated (r 5
20.94), thus the latter variable was excludedfrom the analysis.
Descriptive statistics represent averages 6
SD. All statistical analyses were conductedusing StatView 5.0 (SAS Institute 1998), withsignificance accepted at P # 0.05.
RESULTS
We monitored 346 parrot nests from 1990 to1999, with a range of 20 in 1991 to 50 during1999. However, because we could not determineclutch size of all active nests, we reportreproductive parameters for 322 nests.
TIMING OF BREEDING
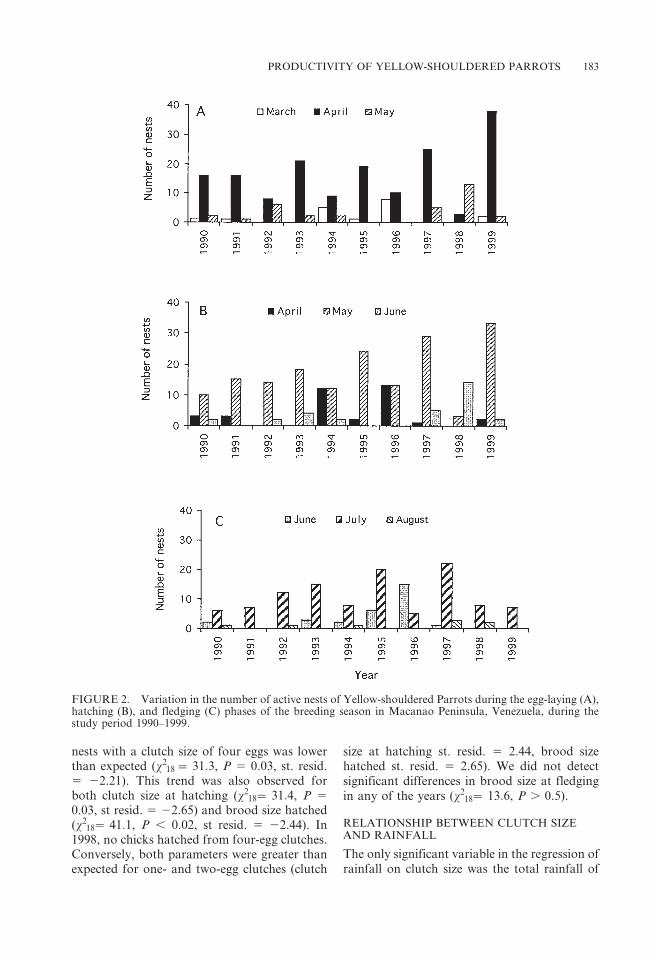
Egg-laying usually started the last week ofMarch or the first week of April, with slightannual variation (Fig. 2A), and lasted from fiveto eight weeks. Eggs hatched from April to June(Fig. 2B) and chicks fledged from June toAugust (Fig. 2C). The duration of the fledgingperiod varied among nests, extending from fiveto ten days for clutches of three and four eggs.The timing of the different nest stages wasconsistent across years except during 1998,when the phases of laying and hatching weredelayed relative to all other years.
The incubation period began when the firstegg was laid and lasted approximately onemonth (27.1 6 1.4 days; Table 1). Length of theincubation period varied among years (H 5
46.8, df 5 9, P , 0.001). The number of eggsthat were incubated for 26 days was greaterthan expected in 1990 (x 4
2 ~ 11:3, P 5 0.02, st.resid. 5 5.0). In 1991, the number of eggsincubated for both 26 and 27 days was higher
than expected (x 42 ~ 88:2, P , 0.001; 26 days
st. resid. 5 22.7 and 27 days st. resid. 5 10.7).
Only one of the pair (sex undetermined)incubated the eggs, and the other member of thepair brought food to the incubating bird. Afterhatching, both parents often entered the nest,presumably to feed nestlings, and remainedthere for 3–40 min (16.3 6 12.5 min, n 5 12).When nestlings were small, adults brought foodthree times a day: in the morning (08:30 to10:00), early afternoon (14:30 to 16:00), and lateafternoon (17:00 to 18:30). As nestlings grew,feeding frequency was reduced to twice a day(morning and afternoon). Nestlings werebrooded throughout the night for approximate-ly two weeks after hatching and the nestlingperiod lasted around two months (58.5 6
3.1 days; Table 1). We found no interannualdifferences in the average age at fledging (H 5
11.9, df 5 9, P 5 0.22).
Fledglings remain with their parents for atleast one month and up to one year becausethey are partially dependent on adults forfeeding. On three occasions, we observedjuveniles from the previous year (identified bycolor bands) soliciting food from their parentswho had a new clutch.
REPRODUCTIVE PARAMETERS
The average total clutch laid by the Yellow-shouldered Parrot during our study was 3.38 6
0.78 eggs per nest, with a range of one to fiveand a mode of four eggs per clutch (Table 2). In1993, 1994, and 1998, the mode was reduced tothree eggs per nest. In 1992, one nest had anextraordinary clutch of seven eggs. We regular-ly observed three parrots close to this nest,suggesting the trio was composed of twofemales and a male, and both females laid eggsin the nest. The average clutch size at hatching(3.36 6 0.80 eggs per nest) was almost equal tothe average total clutch laid, which indicatesthat very few partial losses occurred during theincubation period and most of the eggs surviveduntil hatching (Table 2). The brood size hatched(3.09 6 0.92 nestlings per nest) was slightlysmaller than the two previous parameters and,as expected, brood size at fledging decreasedfurther, to 2.95 6 1.05 fledglings per nest(Table 2).
In 1998, some reproductive parameters dif-fered from other years because of an extremedrought from January to June. The number of
182 VIRGINIA SANZ AND ADRIANA RODRIGUEZ-FERRARO
nests with a clutch size of four eggs was lowerthan expected (x 18
2 ~ 31:3, P 5 0.03, st. resid.5 22.21). This trend was also observed forboth clutch size at hatching (x 18
2 ~ 31:4, P 5
0.03, st resid. 5 22.65) and brood size hatched(x 18
2 ~ 41:1, P , 0.02, st resid. 5 22.44). In1998, no chicks hatched from four-egg clutches.Conversely, both parameters were greater thanexpected for one- and two-egg clutches (clutch
size at hatching st. resid. 5 2.44, brood sizehatched st. resid. 5 2.65). We did not detectsignificant differences in brood size at fledgingin any of the years (x 18
2 ~ 13:6, P . 0.5).
RELATIONSHIP BETWEEN CLUTCH SIZEAND RAINFALL
The only significant variable in the regression ofrainfall on clutch size was the total rainfall of
FIGURE 2. Variation in the number of active nests of Yellow-shouldered Parrots during the egg-laying (A),hatching (B), and fledging (C) phases of the breeding season in Macanao Peninsula, Venezuela, during thestudy period 1990–1999.
PRODUCTIVITY OF YELLOW-SHOULDERED PARROTS 183

the previous nonbreeding period (R2 5 0.57,R2 adj. 5 0.52, F1,8 5 10.6, P , 0.02). Thisvariable explained about half of the variation(52%), but if the relationship was establishedthrough a logarithmic regression, this valueincreased to more than 60% (R2 5 0.68, R2 adj.5 0.64).
Although total rainfall of the previous year(from January to December) was not significantin the regression model, this variable influencedwhether clutch size the following year washigher or lower than average. If the previousyear had higher-than-average rainfall, clutchsize was also higher than average, as occurred in
TABLE 1. Annual estimates for the duration of incubation and age of nestlings at fledging for the Yellow-shouldered Parrot in Macanao Peninsula, Venezuela, 1990–1999.
Year
Incubation (days) Age at fledging (days)
Mean 6 SD Range n Mean 6 SD Range n
1990 26.5 6 1.2 24–29 15 58.8 6 2.5 54–62 231991 26.0 6 0.8 24–29 62 58.9 6 4.4 53–66 191992 26.9 6 1.5 25–29 9 59.3 6 2.5 55–65 151993 27.0 6 1.3 25–30 18 58.5 6 2.5 56–63 101994 28.5 6 1.2 27–30 13 61.0 6 2.7 58–63 31995 27.1 6 1.8 24–30 20 58.5 6 1.8 56–62 271996 28.1 6 1.4 26–30 14 58.7 6 3.1 54–65 321997 27.9 6 1.5 25–30 11 58.7 6 2.9 54–65 271998 28.8 6 1.0 27–30 8 56.5 6 4.0 52–63 141999 27.4 6 2.0 25–30 5 55.7 6 4.2 50–61 91990–1999 27.1 6 1.4 24–30 175 58.5 6 3.1 50–66 179
TABLE 2. Annual and total values of reproductive parameters and productivity per nesting pair of Yellow-shouldered Parrots in Macanao Peninsula, Venezuela, 1990–1999.
Year
Total clutch laidClutch size at
hatchingBrood sizehatcheda
Brood size atfledging
Fledglings pernesting pair
Mean 6 SD(range), n
Mean 6 SD(range), n
Mean 6 SD(range), n
Mean 6 SD(range), n
Mean 6 SD(range), n
1990 3.36 6 0.85(2–5), 22
3.30 6 0.98(1–5), 20
2.86 6 1.06(1–4), 21
2.62 6 1.19(1–4), 13
1.79 6 1.58(0–4), 19
1991 3.60 6 0.68(2–5), 20
3.63 6 0.68(2–5), 19
3.32 6 0.75(2–5), 19
3.50 6 1.05(2–5), 6
1.31 6 1.85(0–5), 16
1992 3.71 6 0.56(3–5), 21
3.53 6 0.61(2–4), 19
3.21 6 0.83(1–4), 24
3.00 6 1.31(1–4), 8
1.71 6 1.82(0–4), 14
1993 3.32 6 0.75(1–5), 31
3.44 6 0.65(2–5), 25
3.03 6 0.84(1–5), 31
2.88 6 1.13(1–4), 8
0.87 6 1.49(0–4), 23
1994 3.13 6 0.73(2–5), 30
3.22 6 0.75(1–5), 27
2.71 6 1.04(1–5), 31
3.00 6 1.26(2–5), 6
0.72 6 1.41(0–5), 25
1995 3.29 6 0.92(1–4), 45
3.32 6 0.91(1–4), 44
3.20 6 0.90(1–4), 41
2.81 6 1.03(1–4), 21
1.74 6 1.60(0–4), 34
1996 3.31 6 0.90(1–5), 42
3.33 6 0.89(1–5), 40
3.21 6 0.93(1–5), 38
3.20 6 0.95(1–4), 20
1.78 6 1.76(0–4), 36
1997 3.45 6 0.65(2–4), 47
3.46 6 0.67(2–4), 41
3.28 6 0.75(1–4), 40
3.00 6 0.84(1–4), 18
1.54 6 1.63(0–4), 35
1998 2.88 6 0.60(2–4), 17
2.64 6 0.50(2–3), 14
2.31 6 0.70(1–3), 16
2.29 6 0.76(1–3), 7
0.87 6 1.19(0–3), 15
1999 3.57 6 0.77(1–5), 47
3.45 6 0.88(1–4), 40
3.30 6 0.94(1–4), 40
3.33 6 1.37(1–5), 6
0.37 6 1.03(0–4), 30
1990–1999 3.38 6 0.78(1–5), 322
3.36 6 0.80(1–5), 289
3.09 6 0.92(1–5), 301
2.95 6 1.05(1–5), 113
1.27 6 1.61(0–4), 247
a Sample sizes could increase between clutch size at hatching and brood size hatched because the lastparameter includes nests found with nestlings, but only nests for which number of hatched eggs was estimatedreliably were included.
184 VIRGINIA SANZ AND ADRIANA RODRIGUEZ-FERRARO

1990, 1991, 1997, and 1999 (Fig. 3). Conversely,if rainfall in the previous year was lower thanaverage, clutch size was lower than the averageclutch size, as in the years 1993 to 1996 and1998 (Fig. 3). During the whole decade, therewas no instance where rainfall of the previousyear was higher than average and clutch size thefollowing year was lower than average. Theopposite occurred only in 1992 (Fig. 3).
FACTORS AFFECTINGREPRODUCTIVE SUCCESS
Prehatching period. Egg losses totaled 20% (209eggs lost from a total of 1071 eggs laid) duringour study. Hatching failure (8% of total eggslaid, n 5 85) was the greatest cause of egg losses(x 3
2 ~ 42:4, P , 0.001). Most of the unhatchedeggs were left inside the nest to avoid disturbingthe chicks. In some cases, however, such eggswere removed and their contents examined.Only one of 20 eggs removed containeda partially developed embryo. Other causes ofegg losses included predation (6%, n 5 62),human perturbation, mainly stones thrown intothe nest by local people (3%, n 5 35), andabandonment of nests where none of the eggshatched (3%, n 5 27). In the latter category weonly included clutches of two or more eggs,because for one-egg clutches it was not possibleto discriminate between nest abandonment andunviable eggs.
Causes of egg losses varied among years(Fig. 4A). Hatching failure was significantlyhigher than expected in 1994 and lower thanexpected in 1999 (x 9
2 ~ 20:0, P 5 0.02, st. resid.1994 5 2.81, st. resid. 1999 5 22.12). The lowhatching success in 1994 was due to multipleeggs within the same nest failing to hatch (4nests from a total of 30), which is unusual.Predation was significantly lower than expectedin 1995 and 1996, when no eggs were lost tothat cause, but it was higher than expected in1999 (14%; x 9
2 ~ 42:8, P , 0.001, st. resid. 19955 22.93, st. resid. 1996 5 22.80, st. resid. 19995 4.22). Human-related losses were significant-ly higher than expected in 1992 and 1997(x 9
2 ~ 31:2, P , 0.001, st. resid. 1992 5 3.91,st. resid. 1997 5 2.04). The number of eggslost due to clutch abandonment was signifi-cantly higher than expected in 1993 becauseof the flooding of two nests after an intensestorm (x 9
2 ~ 22:9, P , 0.01, st. resid. 1993 5
3.01).
Total losses of clutches (12% during theprehatching period; Table 3) resulted fromcauses similar to those for partial losses,including predation (46%), clutch abandonmentresulting from flooded nests, death of theincubating female, or one-egg clutches (29%),and human perturbations (24%). We did notfind significant differences among causes oftotal losses during the prehatching phaseamong years (x 9
2 ~ 5:0, P 5 0.83).Nestling period. Chick losses were higher than
egg losses (49%, 467 of 959 nestlings). Incontrast to the prehatching phase, humans werethe main cause of lost nestlings (x 9
2 ~ 139:6, P, 0.001). Poaching caused the loss of 29% (n 5
276) of nestlings, whereas natural factorscaused the loss of 20% (10%, n 5 98, deathsfrom undetermined causes and 10%, n 5 93,predation). In spite of the nest guarding systemimplemented in the study area, annual poachingranged from 6%–60%. Poaching was low during1990, 1993, and 1995 (x 9
2 ~ 97:4, P , 0.001, st.resid. 1990 5 23.14, st. resid. 1993 5 22.23, st.resid. 1995 5 22.20; Fig. 4B) but increasedsubstantially in 1999, when it caused the loss of60% of all nestlings (st. resid. 1999 5 6.67).Poaching may have increased in 1999 becausedeterioration of the socio-economic situation oflocal inhabitants (e.g., unemployment, drugconsumption and trafficking) caused morepeople to poach and sell parrot chicks. Poach-ing increased in 1994 because one of the guardspoached nests.
Nestling mortality from undetermined causesalso fluctuated among years. It was significantlyhigher in 1995 (x 9
2 ~ 27:4, P , 0.01, st. resid.
FIGURE 3. Positive relationship between meanclutch size and total precipitation of the previousyear. Dashed lines represent mean clutch size (3.38)and mean rainfall (362 mm) for 1990–1999. Numbersby each point indicate the year.
PRODUCTIVITY OF YELLOW-SHOULDERED PARROTS 185

FIGURE 4. Annual fluctuations in the relative frequencies of causes of nest losses of Yellow-shoulderedParrots in the prehatching (A) and nestling (B) periods in Macanao Peninsula, Venezuela, 1990–1999, given aspercentages of total number of eggs laid (A, n 5 1071) or nestlings hatched (B, n 5 959).
TABLE 3. Number of nests with total failure in the prehatching and nestling periods of Yellow-shoulderedParrots in Macanao Peninsula, Venezuela, 1990–1999.
Year
Total nesting failure Total brood failure
Total numberof clutches Failed clutches
Total numberof broods Failed broods
1990 23 2 20 61991 20 1 18 91992 26 2 21 81993 37 6 28 111994 35 4 29 181995 45 4 39 141996 42 4 38 141997 47 7 39 131998 21 3 17 81999 50 9 36 25
1990–1999 346 42 285 126
186 VIRGINIA SANZ AND ADRIANA RODRIGUEZ-FERRARO

1995 5 3.10; Fig 4B). In most years, only one ortwo nestlings, from different nests, died fromunknown causes. In 1995, however, wholebroods died in four of the 12 nests wherenestlings died from unknown causes. Althoughinexplicable nestling deaths occurred in differ-ent cavities, unexplained deaths were foundrepeatedly in six cavities over several years andthose cavities accounted for 41% of all thedeaths.
Number of depredated nestlings was higherthan expected in 1991 and lower in 1999(x 9
2 ~ 26:5, P , 0.01, st. resid. 1991 5 2.67,st. resid. 1999 5 23.36; Fig. 4B). The low levelof predation during 1999 was likely becausemany nestlings were poached at early ages,leaving fewer nestlings exposed to predators.The main predators during the nestling periodwere snakes, mainly the boa constrictor (Boaconstrictor), and to a lesser degree, the Amazontree boa (Corallus ruschenbergerii). These spe-cies often remained inside the cavity after anattack, making identification possible. In mostof the attacks, snakes only fed on one or twonestlings; however, the rest of the brood usuallydied as a result of wounds or crushing. Theseboas also depredated five nesting females andanother female was wounded on the wing.Including this last female, snakes depredated2% of the breeding females monitored duringour study.
Total brood losses were high during thenestling phase (44%; Table 3), mostly asa consequence of poaching (60%) and predation(24%). Mortality from undetermined causes
represented 12% of the total and a few broods(4%) were lost to some combination of causes.Interannual differences were a result of theincrease in poaching in 1999 (x 9
2 ~ 20:4, P ,
0.03, st. resid. 1999 5 2.17).
PRODUCTIVITY
The average hatching success for the period1990–1999 was 0.81 6 0.38 and ranged froma minimum of 0.75 in 1993 to a maximum of0.90 in 1996 (Fig. 5). An approximate estimateof fertility can be calculated after subtractingthe unhatched eggs from nests with total losses(Smith and Saunders 1986); this measure isapproximate because causes other than infertil-ity can reduce hatching. This calculation givesan average fertility value of 0.89 (range 0.81–0.94), which did not vary among years(x 9
2 ~ 12:5, P 5 0.19). Conversely, fledgingsuccess was only 0.38 6 0.44 for the sameperiod, with a minimum of 0.11 in 1999 anda maximum of 0.54 in 1995 (Fig. 5).
The average number of fledglings per nestingpair was 1.27 6 1.61 and ranged from 0.37fledglings per pair in 1999 to 1.79 fledglings perpair in 1990 (Table 2). Productivity as a functionof total clutch size was highest for five-eggclutches (3.00 6 2.37 fledglings per nest),intermediate for three- and four-egg clutches(1.24 6 1.34 and 1.52 6 1.78 fledglings per nest,respectively), and lowest for one- and two-eggclutches (0.22 6 0.44 and 0.26 6 0.66 fledglingsper nest, respectively). Finally, given that theaverage total clutch was 3.38 eggs per pair andeach pair produced an average of 1.27 fledg-
FIGURE 5. Mean annual and total hatching and fledging success (6 SD) of Yellow-shouldered Parrots inMacanao Peninsula, Venezuela, 1990–1999.
PRODUCTIVITY OF YELLOW-SHOULDERED PARROTS 187

lings, each pair lost on average 62% of its initialreproductive investment.
DISCUSSION
Analysis of the different reproductive param-eters and productivity of the Yellow-shoulderedParrot indicated that major losses occurredduring the nestling period, primarily as a conse-quence of human activities. Illegal poachingsignificantly reduced productivity, and is clearlythe main reason for the species’ initial popula-tion decline and current endangerment (Rodrı-guez et al. 2004).
Clutch size of the Yellow-shouldered Parroton Margarita Island is one of the highestrecorded among members of the genus Ama-zona, including the population of the samespecies on La Blanquilla Island, Venezuela(Table 4). This attribute, together with the highhatching success of the species, suggests theYellow-shouldered Parrot has a higher repro-ductive potential than many species in thisgenus. However, the number of fledglingsproduced per breeding pair was low comparedwith other species of the genus, a consequenceof the high percentage of nestlings that werepoached. Reducing poaching levels is key forincreasing the parrot population, as is evidentfrom the years when the guarding and surveil-lance program was executed successfully at LaChica Creek. During the decade of manage-
ment the population increased from a probablenumber of 1300 parrots in 1990 to 2350 in 1999.In other reproductive areas the poaching levelcan be as high as 95% of total nests (VS andARF, pers. obs.). Illegal poaching varies greatlyamong years because it depends on multiplefactors, such as the number and attitude ofpeople involved in this activity, the effectivenessof control strategies, the socio-economic situa-tion of the country, and the availability ofa market (either local or international) for thechicks (Wright et al. 2001). In the absence ofpoaching, productivity could increase up to2.16 fledglings per nest, a very high valuecompared with other Amazona species (Table 4).
The second most important reason for lowproductivity was predation, which is an impor-tant limiting factor during the reproductiveperiod for most avian species (Ricklefs 1969,Skutch 1985). When we added all the lossescaused by predation during the two periods(prehatching and nestling), predation was re-sponsible for 15% of the losses of all eggs laidby Yellow-shouldered Parrots. Comparison ofthis value with other Amazona species isdifficult because of the different ways in whicheach author expresses such values. Our value of15%, however, is similar to those reported byEnkerlin-Hoeflich (1995) for three Amazonaspecies on mainland Mexico, which fluctuatedbetween 10% and 19% (but could be as high as
TABLE 4. Measurements of reproductive success of several Amazona species.
SpeciesMean clutchsize (range)
Hatchingsuccess (%)
Fledglings perbreeding pair Source
A. aestiva (1–6) 70 1.03 Seixas and Mourao 2002A. agilis 3.1 85 0.95 Koenig 2001A. autumnalis 2.7 (1–4) – 0.94 Enkerlin-Hoeflich 1995A. barbadensis 3.4 (1–5) 80 1.27 This studyA. barbadensisa 2.3 (1–4) – – Rodrıguez-Ferraro and Sanz,
unpubl. dataA. brasiliensis 3.6 (3–4) 63 2.19b; 0.03c Martuscelli 1995A. collaria 3.0 66 .0.28 Koenig 2001A. finschi 2.6 (2–4) 90 0.99 Renton and Salinas-Melgoza 2004A. leucocephala bahamensis 3.6 (2–5) 51 0.80 Gnam and Rockwell 1991A. l. caymanensis 3.2 (2–5) – 1.80 Bradley 1986 in Gnam and
Rockwell 1991A. oratrix 2.6 (2–3) – 0.82 Enkerlin-Hoeflich 1995A. ventralis 2.8 (2–4) 93 2.55 Snyder et al. 1987A. viridigenalis 3.4 (2–5) – 1.43 Enkerlin-Hoeflich 1995A. vittata 3.0 (2–4) 84 1.80 Snyder et al. 1987
a La Blanquilla Island population.b Protected population.c Population under poaching pressure.
188 VIRGINIA SANZ AND ADRIANA RODRIGUEZ-FERRARO

30%), but considerably lower than that reportedfor the Black-billed Parrot (A. agilis) inJamaica, where losses can reach 37% (Koenig2001). For Puerto Rican (A. vittata), Blue-fronted (A. aestiva), and Lilac-crowned (A.finschi) Parrots, predation was identified asthe main factor reducing productivity (Snyderet al. 1987, Seixas and Mourao 2002, Rentonand Salinas-Melgoza 2004).
Based on evidence found in depredated nestscombined with fauna resident in the MacanaoPeninsula, potential egg predators includetropical rat snakes (Spillotes pullatus) andstriped hog-nosed skunks (Conepatus semistria-tus), whereas Robinson’s mouse opossums(Marmosa robinsoni) may be potential nestlingpredators. Two uncommon nestling predatorsincluded an ocelot (Leopardus pardalis) anda centipede (Scolopendra sp.). Raptors areknown to prey on psittacines (Snyder et al.1987), and may feed on juveniles and, occa-sionally, on nesting females on MargaritaIsland. We did not observe any hawks attackingwild parrots, but we did find a parrot band ina Harris’s Hawk (Parabuteo unicinctus) pellet,indirect evidence that the hawk fed on a juvenileYellow-shouldered Parrot. On several occasionswe found ants inside a cavity where the wholebrood was dead, but we do not know if antscaused the deaths. Given that in some nests antsfed on dead nestlings without affecting the liveones, we suggest that ants entered the nest afterthe deaths occurred.
Unknown causes were responsible for a sub-stantial number of nestling deaths. In nine casesin which the whole brood died of unknowncauses, the most likely reason was nest aban-donment after the death of one of the parents,resulting in nestling starvation or death fromexposure to low temperatures or rain. In otherpsittacene species, however, it has occasionallybeen reported that one member of a pair cansuccessfully raise the brood even after the othermember has died, e.g., female Puerto RicanParrots (Snyder et al. 1987) and male PinkCockatoos (Cacatua leadbeateri, Rowley andChapman 1991). An alternative explanation isthat these deaths could have been caused bydiseases, especially viral diseases, which aredifficult to detect. However, veterinary studiesconducted from 1993 to 1996 demonstratedthat the parrot population at La Chica Creek isfree of diseases that are common in parrots.
Salmonella was the only bacteria present infecal cultures, and was found in only five of the66 nestlings sampled from 33 nests (a sample of23% of average total annual nests). All thesenestlings looked outwardly healthy and fledgedsuccessfully. None of the sampled nestlings hadantibodies against New Castle disease orpsittacosis, indicating they were not exposedto these diseases (Hoogesteijn and Dıaz 1993,Hoogesteijn et al. 1994). Three of the nestlingsthat died during 1999 were in a wet cavity andhad furuncles around their cloacae, thus theirdeaths may have been caused by infection.Brood reduction may also have caused somedeaths; six of 36 recovered nestlings were belowaverage weight. At least six nestlings might havedied as a consequence of mistakes in themanagement of foster nests. These nestlingswere introduced into cavities where deaths hadoccurred prior to the introduction; thus, it ispossible they died as a consequence of pre-existing problems in that cavity. After theseevents, the technique was improved to reducelosses (Sanz and Rojas-Suarez 1997).
Ectoparasites may also reduce breeding pro-ductivity, especially for birds that nest incavities and reuse them every year (Ricklefs1969). During the veterinary studies, the num-ber of ectoparasites found was minimal. Onlytwo nestlings had one tick each and anotherhad a louse (Hoogesteijn and Dıaz 1993,Hoogesteijn et al. 1994). We found no cases ofinfection by Warble flies (Philornis pici), re-ported as an important nestling parasite forPuerto Rican (Snyder et al. 1987) and Lilac-crowned Parrots (Renton 2002).
The fertility value of 90% estimated for theYellow-shouldered Parrot is high, but is withinthe reported range for Australian parrot species(from 77% to 91%; Smith and Saunders 1986,Rowley and Chapman 1991, Smith 1991). Thus,losses due to infertility constitute a reasonableproportion as expected in a healthy population.Hatching success (80%) is also within theexpected range of values ($60%) for altricialand cavity-nesting species (Nice 1957, Koenig1982). One factor that can increase the annualproductivity of a pair is the ability to laya second clutch after the loss of the first. Asparrots were not individually banded, it was notpossible to know whether the same pair oranother one laid a new clutch in a cavity when itwas reused during the same breeding season
PRODUCTIVITY OF YELLOW-SHOULDERED PARROTS 189

after the total loss of a first clutch. However, re-laying appears limited in the Yellow-shoulderedParrot, as in the majority of species of the genusAmazona.
EFFECT OF CLIMATE ON PRODUCTIVITY
In the case of the Yellow-shouldered Parrot,minimal losses due to climate may be related tothe fact that the species inhabits a warm andextremely dry environment. Losses caused bynest flooding are few and limited to years withoverpowering rains. Even though MargaritaIsland is in the Caribbean, it is not reached byhurricanes, which severely affect other parrotspecies in the Bahamas and Cayman Islands(Wiley 1991), as well as on Greater (Snyder etal. 1987, Wiley 1991) and Lesser Antilles (Evans1991, Christian et al. 1996). As the weather iswarm year-round, adequate nestling thermo-regulation is not as critical as it might be forother Amazona species living in colder climates.
However, climatic factors such as precipita-tion do affect productivity. This was clear in1993 and 1994, when most breeding pairs hadthree-egg clutches instead of the more frequentfour-egg clutch. Clutch size, however, wassignificantly reduced only in 1998, one of thedriest periods in the last 50 years in theMacanao Peninsula. The low precipitation inthat year also affected the number of activenests and delayed the timing of breeding. Weused rainfall as an indirect measure of theamount of food available during the studyperiod, because several studies have determinedthat a direct relationship between rainfall andflowering-fruiting periods exists for most plantspecies inhabiting arid zones (Gibbs and Grant1987, Guevara et al. 1992, Bancroft et al. 2000,Renton 2002). Thus, the production of primaryfood sources (fruits and seeds) of the Yellow-shouldered Parrot (Stolk 1997, Sanz 2004) ishighly influenced by the amount of rainfall ina given year. A greater abundance of availablefood would result in heavier female parrots. Inseveral bird species, heavier females tend to laylarger clutches or larger eggs and thus havegreater reproductive success (Ankney andMacInnes 1978, Newton et al. 1983, Jarvinenand Vaisanen 1984). Interestingly, there alsoseems to be a relationship between low averagerainfall and an increase in the percentage ofunhatched eggs. The physical condition ofnesting females (affected by food availability)
may affect their incubation constancy (Martin1987). Therefore, the high percentage of un-hatched eggs in 1994 could have resulted fromthis year being the second driest of the decadeand coming after a below-average rainfall year.
In conclusion, this species is facing highlyvariable ecological pressures over time, withboth natural factors and human pressures(poaching) showing significant interannualfluctuations. This variation, which affects re-productive parameters and productivity, indi-cates that short-term studies can be misleadingwhen used as guidelines for conservation andmanagement programs, and emphasizes theneed for long-term monitoring programs.
The information gathered during this decadeof study indicates that the population ofYellow-shouldered Parrots in the MacanaoPeninsula has a great recovery potential dueto the high values of their reproductive param-eters (clutch size and hatching success). In theabsence of poaching, steady growth of thepopulation would be possible. An effectiveprogram to reduce poaching is crucial fora viable population to persist in the MacanaoPeninsula in the long term.
ACKNOWLEDGMENTS
We are especially grateful to Juan Francisco Penoth,Pablo Antonio Millan and Jesus Marcano for fieldassistance and their huge contributions to the successof this study, and to the other project coordinatorsFranklin Rojas-Suarez, Mariana Albornoz, AnaTrujillo, and Carolina Bertsch for their contributionto data collection. We thank Stuart Strahl andAlejandro Grajal for their support during differentphases of the project. We would also like to thank thefamily Salazar Meneses for allowing us to conductthis long-term project at the Fundo San Francisco,and all the volunteers that helped with fieldwork.Funds to support the Yellow-shouldered Parrotconservation program were provided by the WildlifeConservation Society, the British Embassy in Car-acas, BirdLife International, U.S. Fish and WildlifeService, Papageien Fund, American Bird Conservan-cy, and the Golden Ark Foundation. Parrot moni-toring and manipulation were conducted with per-missions from Direccion de Fauna - Ministerio delAmbiente y de los Recursos Naturales de Venezuela.Donald Brightsmith, John G. Blake, James W. Wiley,and an anonymous reviewer made useful commentson a previous version of the manuscript.
LITERATURE CITED
ALBORNOZ, M., F. ROJAS-SUAREZ, AND V. SANZ.1994. Conservacion y manejo de la cotorracabeciamarilla (Amazona barbadensis) en la islade Margarita, Estado Nueva Esparta, p. 197–
190 VIRGINIA SANZ AND ADRIANA RODRIGUEZ-FERRARO

207. In G. Morales, I. Novo, D. Bigio, A. Luy,and F. Rojas-Suarez [EDS.], Biologıa y conserva-cion de los psitacidos de Venezuela. GraficasGiavimar, Caracas, Venezuela.
ANKNEY, C. D., AND C. D. MACINNES. 1978.Nutrient reserves and reproductive performanceof female Lesser Snow Geese. Auk 95:459–471.
BANCROFT, G. T., R. BOWMAN, AND R. J. SAWICKI.2000. Rainfall, fruiting phenology, and thenesting season of White-crowned Pigeons in theupper Florida Keys. Auk 117:416–426.
BEISSINGER, S. R. 2002. Population viability analysis:past, present, future, p. 5–17. In S. R. Beissingerand D. R. McCullough [EDS.], Populationviability analysis. University of Chicago Press,Chicago.
BEISSINGER, S. R., AND M. I. WESTPHAL. 1998. Onthe use of demographic models of populationviability in endangered species management.Journal of Wildlife Management 62:821–841.
BENNETT, P. M., AND I. P. F. OWENS. 1997.Variation in extinction risk among birds: chanceor evolutionary predisposition? Proceedings ofthe Royal Society of London Series B 264:401–408.
BIRDLIFE INTERNATIONAL. 2000. Threatened birdsof the world. Lynx Edicions and BirdLifeInternational, Barcelona and Cambridge, UK.
CAUGHLEY, G. 1978. Analysis of vertebrate popula-tions. John Wiley & Sons, Bath, UK.
CHRISTIAN, C. S., T. E. LACHER JR., M. P. ZAMORE,T. D. POTTS, AND G. W. BURNETT. 1996. Parrotconservation in the Lesser Antilles with somecomparison to the Puerto Rican efforts. Bi-ological Conservation 77:159–167.
COLLAR, N. J., M. J. CROSBY, AND A. J. STATTERS-
FIELD. 1994. Birds to watch 2: the world list ofthreatened birds. BirdLife International, Cam-bridge, UK.
CONOVER, W. J. 1998. Practical nonparametricstatistics. 3rd ed. Wiley & Sons, New York.
DESSENE, P., AND S. D. STRAHL. 1991. Trade and theconservation status of the family Psittacidae inVenezuela. Bird Conservation International1:153–169.
ENKERLIN-HOEFLICH, E. C. 1995. Comparativeecology and reproductive biology of three speciesof Amazona parrots in northeastern Mexico.Ph.D. dissertation, Texas A & M University,Austin, TX.
EVANS, P. G. H. 1991. Status and conservation ofImperial and Red-necked Parrots Amazonaimperialis and A. arausiaca on Dominica. BirdConservation International 1:11–32.
FORSHAW, J. M. 1989. Parrots of the world.Lansdowne Editions, Melbourne.
GIBBS, H. L., AND P. R. GRANT. 1987. Ecologicalconsequences of an exceptionally strong El Ninoevent on Darwin’s Finches. Ecology 68:1735–1746.
GNAM, R. S., AND R. F. ROCKWELL. 1991. Re-productive potential and output of the BahamaParrot Amazona leucocephala bahamensis. Ibis133:400–405.
GOLDBRUNNER, A. W. 1962. El clima de la isla deMargarita. Memoria Sociedad Ciencias Natur-ales La Salle 22:146–155.
GUEVARA, M., Y. BERGERON, R. MCNEIL, AND A.LEDUC. 1992. Seasonal flowering and fruitingpatterns in tropical semi-arid vegetation inNortheastern Venezuela. Biotropica 24:64–76.
HOOGESTEIJN, A., AND M. DIAZ. 1993. Informe delestado de salud de Amazona barbadensis en laPenınsula de Macanao, isla de Margarita.Technical report. Provita, Caracas, Venezuela.
HOOGESTEIJN, A., M. GONZALEZ, AND M. DIAZ.1994. Informe del estado de salud de Amazonabarbadensis en la Penınsula de Macanao. Tech-nical report. Provita, Caracas, Venezuela.
HOYOS, J. 1985. Flora de la isla de Margarita.Sociedad Fundacion La Salle de Ciencias Nat-urales, Caracas, Venezuela.
JARVINEN, A., AND R. A. VAISANEN. 1984. Re-production of Pied Flycatchers (Ficedula hipo-leuca) in good and bad breeding seasons ina northern marginal area. Auk 101:439–450.
JUNIPER, T., AND M. PARR. 1998. Parrots. A guide toparrots of the world. Yale University Press, NewHaven, CT.
KOENIG, S. E. 2001. The breeding biology of Black-billed Parrot Amazona agilis and Yellow-billedParrot Amazona collaria in Cockpit Country,Jamaica. Bird Conservation International11:205–225.
KOENIG, W. D. 1982. Ecological and social factorsaffecting hatchability of eggs. Auk 99:526–536.
LACY, R. C., N. R. FLESNESS, AND U. S. SEAL. 1989.Puerto Rican Parrot Amazona vittata populationviability analysis and recommendations. CaptiveBreeding Specialist Group, Apple Valley, MN.
MARTIN, T. E. 1987. Food as a limit on breedingbirds: a life-history perspective. Annual Reviewof Ecology and Systematics 18:453–487.
MARTUSCELLI, P. 1995. Ecology and conservation ofthe Red-tailed Amazon Amazona brasiliensis insouth-eastern Brazil. Bird Conservation Interna-tional 5:405–420.
NEWTON, I., M. MARQUISS, AND A. VILLAGE. 1983.Weights, breeding, and survival in EuropeanSparrowhawks. Auk 100:344–354.
NICE, M. M. 1957. Nesting success of altricial birds.Auk 74:305–321.
RENTON, K. 2002. Influence of environmentalvariability on the growth of Lilac-crownedParrot nestlings. Ibis 144:331–339.
RENTON, K., AND A. SALINAS-MELGOZA. 2004.Climatic variability, nest predation, and repro-ductive output of Lilac-crowned Parrots (Ama-zona finschi) in tropical dry forest of westernMexico. Auk 121:1214–1225.
RICKLEFS, R. E. 1969. An analysis of nestingmortality in birds. Smithsonian Contributionsto Zoology 9:1–48.
RODRIGUEZ, J. P., L. FAJARDO, I. HERRERA, A.SANCHEZ, AND A. REYES. 2004. Yellow-shoul-dered Parrot (Amazona barbadensis) on theislands of Margarita and La Blanquilla, Vene-zuela, p. 361–370. In H. R. Akcakaya, M.Burgman, O. Kindvall, C. Wood, P. Sjoren-
PRODUCTIVITY OF YELLOW-SHOULDERED PARROTS 191

Gulve, J. Hattfield, and M. McCarthy [EDS.],Species conservation and management: casestudies. Oxford University Press, Oxford.
RODRIGUEZ, J. P., AND F. ROJAS-SUAREZ. 1994.Analisis de viabilidad poblacional de tres pobla-ciones de psitacidos insulares de Venezuela,p. 97–113. In G. Morales, I. Novo, D. Bigio,A. Luy, and F. Rojas-Suarez [EDS.], Biologıa yconservacion de los psitacidos de Venezuela.Graficas Giavimar, Caracas, Venezuela.
RODRIGUEZ, J. P., AND F. ROJAS-SUAREZ. 1995.Libro Rojo de la Fauna Venezolana. Provita andFundacion Polar, Caracas, Venezuela.
ROWLEY, I., AND G. CHAPMAN. 1991. The breedingbiology, food, social organization, demographyand conservation of the Major Mitchell or PinkCockatoo, Cacatua leadbeateri, on the margin ofthe western Australian wheatbelt. AustralianJournal of Zoology 39:211–261.
SANZ, V. 2004. Ecologıa de Amazona barbadensis(Aves: Psittacidae): caracterizacion y uso delhabitat en la Penınsula de Macanao (Isla deMargarita) a diferentes escalas espaciales ytemporales. Ph.D. dissertation, UniversidadCentral de Venezuela, Caracas, Venezuela.
SANZ, V., AND F. ROJAS-SUAREZ. 1997. Los nidosnodriza como tecnica para incrementar elreclutamiento de la cotorra cabeciamarilla (Ama-zona barbadensis, Aves: Psittacidae). Vida Sil-vestre Neotropical 6:8–14.
SAS INSTITUTE. 1998. StatView. Version 5.0. SASInstitute, Inc., Cary, NC.
SAUNDERS, D. A. 1980. Food and movements of theshort-billed form of the White-tailed BlackCockatoo. Australian Wildlife Research 7:257–269.
SAUNDERS, D. A. 1986. Breeding season, nestingsuccess and nestling growth in Carnaby’s Cock-atoo, Calyptorhynchus funereus lattirostris, over16 years at Coomallo creek, and a method forassessing the viability of populations in otherareas. Australian Wildlife Research 13:261–273.
SEIXAS, G. H. F., AND G. MOURAO. 2002. Nestingsuccess and hatching survival of the Blue-frontedAmazon (Amazona aestiva) in the Pantanal ofMato Grosso do Sul, Brazil. Journal of FieldOrnithology 73:399–409.
SIEGEL, S., AND N. J. CASTELLAN. 1995. Estadısticano parametrica: aplicada a las ciencias de laconducta. Editorial Trillas, Mexico D.F., Mex-ico.
SKUTCH, A. 1985. Clutch size, nesting success, andpredation on nests of Neotropical birds, re-viewed. Ornithological Monographs 36:575–594.
SMITH, G. T. 1991. Breeding ecology of the westernLong-billed Corella, Cacatua pastinator pastina-tor. Wildlife Research 18:91–110.
SMITH, G. T., AND D. A. SAUNDERS. 1986. Clutchsize and productivity in three sympatric speciesof cockatoo (Psittaciformes) in the south-west ofwestern Australia. Australian Wildlife Research13:275–285.
SNYDER, N., P. MCGOWAN, J. GILARDI, and A.GRAJAL [EDS.]. 2000. Parrots. Status survey andconservation action plan 2000–2004. IUCN,Gland, Switzerlandand Cambridge, UK.
SNYDER, N. F. R., J. W. WILEY, AND C. B. KEPLER.1987. The parrots of Luquillo: natural historyand conservation of the Puerto Rican Parrot.Western Foundation of Vertebrate Zoology, LosAngeles, CA.
STOLK, M. E. 1997. Dieta y disponibilidad de losrecursos alimenticios de la cotorra cabeciama-rilla (Amazona barbadensis) en la Penınsula deMacanao, isla de Margarita. Undergraduatethesis, Universidad Simon Bolıvar, Caracas,Venezuela.
SUTHERLAND, W. J. 2000. The conservation hand-book. Research, management, and policy. Black-well Science, Oxford.
WILEY, J. W. 1991. Status and conservation ofparrots and parakeets in the Greater Antilles,Bahama Islands, and Cayman Islands. BirdConservation International 1:187–214.
WRIGHT, T. F., C. A. TOFT, E. ENKERLIN-HOEFLICH,J. GONZALEZ-ELIZONDO, M. ALBORNOZ, A.RODRIGUEZ-FERRARO, F. ROJAS-SUAREZ, V.SANZ, A. TRUJILLO, S. R. BEISSINGER, V.BEROVIDES, X. GALVEZ, A. T. BRICE, K. JOYNER,J. EBERHARD, J. GILARDI, S. E. KOENIG, S.STOLESON, P. MARTUSCELLI, J. M. MEYERS, K.RENTON, A. M. RODRIGUEZ, A. C. SOSA-ASANZA, F. J. VILELLA, AND J. W. WILEY. 2001.Nest poaching in Neotropical parrots. Conserva-tion Biology 15:710–720.
192 VIRGINIA SANZ AND ADRIANA RODRIGUEZ-FERRARO




![The Parrot [1936]](https://static.fdokumen.com/doc/165x107/6333183ba6138719eb0a744a/the-parrot-1936.jpg)















