
REPRODUCTION OF THE BLACK LAND CRAB, GECARCINUS RURICOLA, IN THE SAN ANDRES ARCHIPELAGO, WESTERN...
13
BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, research libraries, and research funders in the common goal of maximizing access to critical research. REPRODUCTION OF THE BLACK LAND CRAB, GECARCINUS RURICOLA, IN THE SAN ANDRES ARCHIPELAGO, WESTERN CARIBBEAN Author(s): Richard G. Hartnoll, Mark S. P. Baine, Arne Britton, Yolima Grandas, Jennifer James, Alejandro Velasco, and Michael G. Richmond Source: Journal of Crustacean Biology, 27(3):425-436. 2007. Published By: The Crustacean Society DOI: http://dx.doi.org/10.1651/S-2772.1 URL: http://www.bioone.org/doi/full/10.1651/S-2772.1 BioOne (www.bioone.org ) is a nonprofit, online aggregation of core research in the biological, ecological, and environmental sciences. BioOne provides a sustainable online platform for over 170 journals and books published by nonprofit societies, associations, museums, institutions, and presses. Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use . Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiries or rights and permissions requests should be directed to the individual publisher as copyright holder.
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of REPRODUCTION OF THE BLACK LAND CRAB, GECARCINUS RURICOLA, IN THE SAN ANDRES ARCHIPELAGO, WESTERN...

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, researchlibraries, and research funders in the common goal of maximizing access to critical research.
REPRODUCTION OF THE BLACK LAND CRAB, GECARCINUS RURICOLA,IN THE SAN ANDRES ARCHIPELAGO, WESTERN CARIBBEANAuthor(s): Richard G. Hartnoll, Mark S. P. Baine, Arne Britton, Yolima Grandas, Jennifer James,Alejandro Velasco, and Michael G. RichmondSource: Journal of Crustacean Biology, 27(3):425-436. 2007.Published By: The Crustacean SocietyDOI: http://dx.doi.org/10.1651/S-2772.1URL: http://www.bioone.org/doi/full/10.1651/S-2772.1
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in the biological, ecological, andenvironmental sciences. BioOne provides a sustainable online platform for over 170 journals and books publishedby nonprofit societies, associations, museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance ofBioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiriesor rights and permissions requests should be directed to the individual publisher as copyright holder.

JOURNAL OF CRUSTACEAN BIOLOGY, 27(3): 425–436, 2007
REPRODUCTION OF THE BLACK LAND CRAB, GECARCINUS RURICOLA, IN THE
SAN ANDRES ARCHIPELAGO, WESTERN CARIBBEAN
Richard G. Hartnoll, Mark S. P. Baine, Arne Britton, Yolima Grandas, Jennifer James,Alejandro Velasco, and Michael G. Richmond
(RGH, correspondence) Port Erin Marine Laboratory, University of Liverpool, Port Erin,
Isle of Man IM9 5ED, British Isles ([email protected]);
(MSPB) Motupore Island Research Centre, University of Papua New Guinea, PO Box 320,
University 134, National Capital District, Papua New Guinea;
(AB, YG, JJ, AV) CORALINA, San Luis Road, KM 26, PO Box 725, San Andres Island, Colombia;
(MGR) School of Life Sciences, Heriot-Watt University, Edinburgh, U.K.
A B S T R A C T
Reproductive biology of the black land crab, Gecarcinus ruricola (Linnaeus, 1758), has been studied in the San Andres Archipelago in the
Western Caribbean, on the islands of San Andres and Old Providence. Both sexes become mature at about 50 mm maximum carapace
width, and very few specimens below this size take part in the annual breeding migration. In females the ovaries mature progressively
from January to April, and laying occurs from May to July. There is no evidence that individual females lay more that once in a year. The
breeding migrations were monitored in 2003 and 2004 by recording specimens crossing the coast road (running 50-100 m from the upper
shore). On both San Andres and Old Providence migration was limited to specific stretches of the western shores. On both islands
migration occurred from late April to July, intensity varying with time. Migration peaks were not consistent between islands or years, and
could not be clearly linked to environmental factors. Migrating crabs were mostly female (. 80%), and a proportion were bearing ripe
eggs: on San Andres ovigerous female predominated. Thus some females mated and laid eggs on the landward side of the coast road,
others on the seaward side. Migrating females were on average larger than migrating males, in contrast to the situation in the non-
migrating populations. Log egg number varied isometrically with log carapace width (slope ;3.0): for 70 mm CW the calculated
fecundity was 85,000 eggs. Percentage reproductive investment, based on dry weight, ranged from 1.7 to 9.8%, with a mean of 4.95%.
Recruitment was only observed once, with a mass return of megalops to Old Providence in June 2004. All evidence points to very irregular
recruitment.
INTRODUCTION
The family Gecarcinidae are popularly referred to as ‘landcrabs’, though they are not the only terrestrial crabs, norindeed the crabs most completely adapted for a life on land(see Hartnoll, 1988). Gecarcinids spend most of their adultlife on land, but females must return to the sea to releasetheir larvae (even the cavernicolous Discoplax longipesA. Milne-Edwards, 1867 (Ng and Guinot, 2001)), which allundergo planktotrophic development. The terrestrial adap-tation of adult gecarcinids varies widely. The cavernicolousDiscoplax longipes is largely aquatic (Ng and Guinot,2001). Some, such as several Cardisoma spp., areessentially supratidal, frequently entering the sea on highspring tides, and always retaining access to free water.Others, such as various Gecarcinus and Gecarcoideaspecies, can exist many kilometres from the sea, away fromfree water, and enter the sea only after the often lengthyannual spawning migration.
There are three gecarcinids in the Caribbean. Cardisomaguanhumi (Latreille, 1852) and Gecarcinus lateralis(Freminville, 1835) are widespread both on Caribbeanislands and along the adjacent mainland coast. Both havebeen relatively well studied (for example: for C. guanhumisee Costlow and Bookhout, 1968; O’Mahoney and Full,1984; Wolcott, 1984: for G. lateralis see Klaassen, 1975;Bliss et al., 1978; Willems, 1982; Wolcott and Wolcott,1988; Capistran-Barrados et al., 2003), probably due to their
accessibility to mainland-based researchers. The thirdspecies, Gecarcinus ruricola (Linnaeus, 1758), is essentiallyrestricted to Caribbean islands—there are few records formainland America. Thus, the records for ‘Florida’ (seeKeith, 1985; Chace and Hobbs, 1969) are in fact confirmedonly by specimens for the island of Loggerhead Key, in theTortugas (Carson, 1967): Bliss et al. (1978) report it as‘‘quite rare’’ at Sabal Point in south east Florida, the solemainland location. The references to ‘Nicaragua’ by Chaceand Hobbs (1969) and Keith (1985) are not confirmed inany other sources, and as no specimens or locations arecited, this reference as a mainland site is dubious. Incontrast, its island distribution is widespread and welldocumented through the Greater and Lesser Antilles and theBahamas, from the western end of Cuba eastwards toBarbados (see Carson, 1967, Table 1, for a comprehensivesummary). It is also recorded from Curacao (off the coastof Venezuela). In the western Caribbean there are threeoutliers: the Swan Islands 150 km north of Honduras (Keith,1985), and Old Providence and San Andres Islands (jointlyknown as the San Andres Archipelago) 250 km east ofNicaragua (Carson, 1967 and Hartnoll et al., 2006b).
In the Caribbean, Gecarcinus ruricola is the mostterrestrial of the land crabs. It inhabits damp and shadedforest areas, where it burrows in the soil, or shelters underrocks or amongst tree roots. It is found many kilometresfrom the sea, and to considerable altitudes, .300 m in
425

Dominica (Chace and Hobbs, 1969), and .1000 m inJamaica (Britton et al., 1982). It is mainly nocturnal, exceptduring the breeding migration when large numbers may beactive by day. Nocturnal activity is stimulated by rainfall,but inhibited by bright moonlight (Hartnoll et al., 2006b). Inthe Caribbean, it is the most widely exploited gecarcinid forhuman consumption, though Cardisoma guanhumi isconsumed on a smaller scale (Maitland, 2002).
Despite this ecological and economic interest, G. ruricolahas been very little studied. Carson (1967, 1974) describedits symbiotic relation with flies of the genus Drosophila.Britton et al. (1982) compared its biometry, distribution andactivity with those of G. lateralis. Prahl and Manjarres(1983) briefly outlined its coastal distribution around OldProvidence. Mulder and Stam (1987) and Hoeven andWalters (1998) described distribution and populationstructure in Saba. Distribution and abundance have beenstudied in Dominica (Magin, 2003). Nevertheless, knowl-edge regarding the species is still very fragmentary,especially regarding reproduction. Stebbing (1893) andCalman (1911) describe the annual breeding migration, theformer in more detail. Hoeven and Walters (1988) brieflydiscuss reproduction in Saba. Sjogreen (1999) describes thebreeding migration in Old Providence Island.
In 2002, a comprehensive study programme was initiatedto examine the basic ecology and biology of the species in theSan Andres Archipelago, and the characteristics ofits exploitation. This was triggered by concern that it wasbeing overexploited. Previous papers from this study coverpopulation structure, stock size, and relative growth (Hartnollet al., 2006b), the landward return and description of themegalop stage (Hartnoll and Clark, 2006), and implicationsfor management of the catchery (Baine et al., in press). Thispaper reports on the reproductive biology of the species.Objectives to clarify are the size at maturity, the seasonalmaturation of the gonads, the reproductive migration,fecundity and reproductive investment, and recruitment.
MATERIALS AND METHODS
Study Area
The San Andres Archipelago is an extensive area of islands, banks and caysin the western Caribbean. It is an offshore province of Colombia, locatedsome 700 km northwest of mainland Colombia, and 250 km east of
Nicaragua (Fig. 1). Within the Archipelago Gecarcinus ruricola occursonly on the islands of San Andres and Old Providence/Santa Catalina. SanAndres (hereafter referred to as ‘SA’) has an area of 27 km2, a maximumaltitude of ;100 m, and consists of raised coral limestone (Parsons, 1956).The population is thought to exceed 80,000, and many areas have beendeveloped for tourism or converted to cultivation—in consequence theoriginal forest habitat has been much depleted. Old Providence (not to beconfused with New Providence Island in the Bahamas) has an area of21 km2, a maximum altitude of 360 m, and is connected to the small islandof Santa Catalina (area 1.3 km2) by a floating bridge. The two islands(hereafter jointly referred to as ‘OP’) are of volcanic origin, havea combined population of little over 4000, and are relatively undevelopedwith much of the forest ecosystem intact. References to ‘the Archipelago’relate to SA and OP combined—see Baine et al. (2007) for a review of thecurrent environmental and demographic status of the Archipelago.
Size at Maturity, and Seasonal Maturation of Ovaries
All references to crab size are to maximum carapace width (CW).Preliminary studies were made to determine the approximate size atmaturity, and the reproductive season. These studies, together withbackground data from other unpublished sources, indicated a size atmaturity of ;50 mm CW in each sex, and a breeding season from Aprilto July. As a result the following protocols were adopted. Specimenswere collected by the project staff, or when necessary purchased fromcrab catchers. Crabs were killed by freezing for ;30 min. at �208C, andwhen possible examined fresh. Similar studies were carried out on SA andOP—the indicated sample sizes apply to the work on each island.
To examine size at maturity, samples were collected only during theperiod immediately preceding the breeding season (namely from Februaryto May). This was to prevent the analysis being confounded by specimensof ‘mature’ size, but with undeveloped or spent gonads outside the breedingseason. For each sex ideally at least 10 specimens were collected withineach 10 mm CW size class: this was not possible for some sizes.
To examine the seasonal maturation of the ovaries, only females .60mm CW were sampled. This eliminated immature specimens from theanalysis. A sample of 20 females was examined in each month fromOctober to April.
For both analyses the following data were recorded from each specimen.For males: CW; appearance of vas deferens, categorized as immature,small, medium or large. ‘Immature’ are very small, translucent, and usuallyinvisible to the naked eye. The other categories are opaque white, and ofincreasing size. For females: CW; size of ovary, categorized as immature(¼ usually invisible), small, medium or large; colour of ovary, categorizedas translucent (immature), yellow-orange (maturing), or brown-green(mature); condition of spermathecae, categorized as unmated (translucentand effectively invisible), mated (opaque white, but flattened), or swollen(opaque white, rounded).
The Reproductive Migration
The observations on each island were of crabs observed crossing the coastalroad, which in the areas involved was usually between 50-100 m from theupper shore. Data are not available on the migration patterns further inland,
Table 1. Gecarcinus ruricola. Seasonal ripening of the ovaries from October 2003 to July 2004, determined by changes in ovary size and colour (Y/O¼yellow/orange; B/G¼brown/green). Only specimens .60 mm CW (above the size of maturity) were included in the analysis. Numbers of crabs in each classare listed.
Month
San Andres Old Providence
Ovary size Ovary colour Ovary size Ovary colour
Small Medium Large Y/O B/G Small Medium Large Y/O B/G
Oct 03 15 7 2 20 4 19 4 1 31 2Nov 03 12 7 1 20 0 — — — — —Dec 03 — — — — — 15 4 0 18 1Jan 04 16 2 0 14 4 12 8 0 9 11Feb 04 15 9 4 19 9 11 8 1 5 15Mar 04 5 8 4 13 4 5 4 11 7 15Apr 04 9 1 12 7 15 — — — — —May 04 7 3 10 8 12 7 1 3 5 6Jun 04 — — — — — 23 4 0 20 7Jul 04 — — — — — 19 4 1 11 13
426 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 27, NO. 3, 2007

where the sex composition may differ, nor on the activity of crabs at theshore: the precise timing of spawning behaviour by females may differ fromtheir migration patterns across the road. Logistic constraints precludedcollection of these additional data, which would have been valuable.
Surveys were carried out throughout the breeding season on the knownmigration areas (Fig. 2). When migration was sparse surveys wereconducted every third night or so, but the frequency was increased duringperiods of higher activity. The procedure was to drive and/or walk, asappropriate, along the coast road once during the evening, normally between2000 and 2200 (well after the time of dusk). Preliminary studies had shownthat activity at this time was representative of migration intensity. The roadwas divided into sections. On SA it was split into 1 km sections,conveniently indicated by roadside distance markers, from Km 1 to Km 15.On OP, since distances were smaller, and migrating numbers greater, theroad was divided into 13 sections over a distance of ;4 km. Within eachsection crabs encountered were sexed, CW measured, and presence of eggs(and in selected specimens, incubation stage) on females recorded. Unlessneeded for other studies, the crabs were released unharmed. It is important tonote the limitations of this procedure. Firstly, it was not normally possible toadequately discriminate seaward and landward migration: by the time thatcrabs were spotted by using lights, their activity had been disrupted.Secondly, no estimate of total numbers migrating per night could be made,only of the number on the road at the time of the survey. The methodprovides only comparable data over time, and between locations, on therelative numbers migrating. In these respects the data are reliable. On bothislands the coast road is of constant width, so comparisons between locationare valid. All personnel involved had training in the survey procedures, sosurveys on different dates are comparable.
Roadkills were also recorded during the surveys. They were counted,and where possible sex and presence of eggs recorded. These comple-mented the data on live crabs.
Fecundity and Reproductive Investment
Determination of fecundity and reproductive investment was made only onripe eggs. Females with eggs were collected during the reproductivemigrations. The eggs were removed, and the dry weight of both the female
(without eggs) and the egg mass were determined after drying to constantweight in an oven at 1008C. Reproductive investment (RI) was determinedby expressing egg mass dry weight as a percentage of female body dryweight (after removal of the egg mass). This is the most common means ofcalculating RI (see Hines, 1982).
To determine fecundity a subsample of eggs (;300) was removed fromthe egg masses of selected females before drying. These eggs were countedusing a Bogarov tray, dried as above, and weighed. The results were used todetermine the mean dry weight per egg. This value was used to convert thedry weight of egg mass to egg number. Log egg number was regressed onlog female CW.
Recruitment
A lookout was kept for returning juveniles during the breeding season, anddetailed observations were made of any occurrence. In addition, in theabsence of formal records, anecdotal information was sought regardingrecruitment events in earlier years.
RESULTS
Size at Maturity
For males from both islands the results (Fig. 3) are similar.Specimens less than 50 mm CW were either immature, or inthe early stage of maturity, with ‘small’ vas deferens.Specimens above 50 mm were in almost all cases maturing,and above 70 mm CW all had ‘medium’ or ‘large’ vasdeferens. The slight differences between the two islandsmust be treated with caution, since the data were collectedby different teams. A size at first maturity of 50 mm CW isa fair conclusion.
The results for females are very similar from the twoislands, so only those for OP are shown in Fig. 4. On allthree criteria—ovary size, ovary colour, and condition of
Fig. 1. The Caribbean, to show the location of the San Andres Archipelago.
427HARTNOLL ET AL.: REPRODUCTION OF GECARCINUS RURICOLA
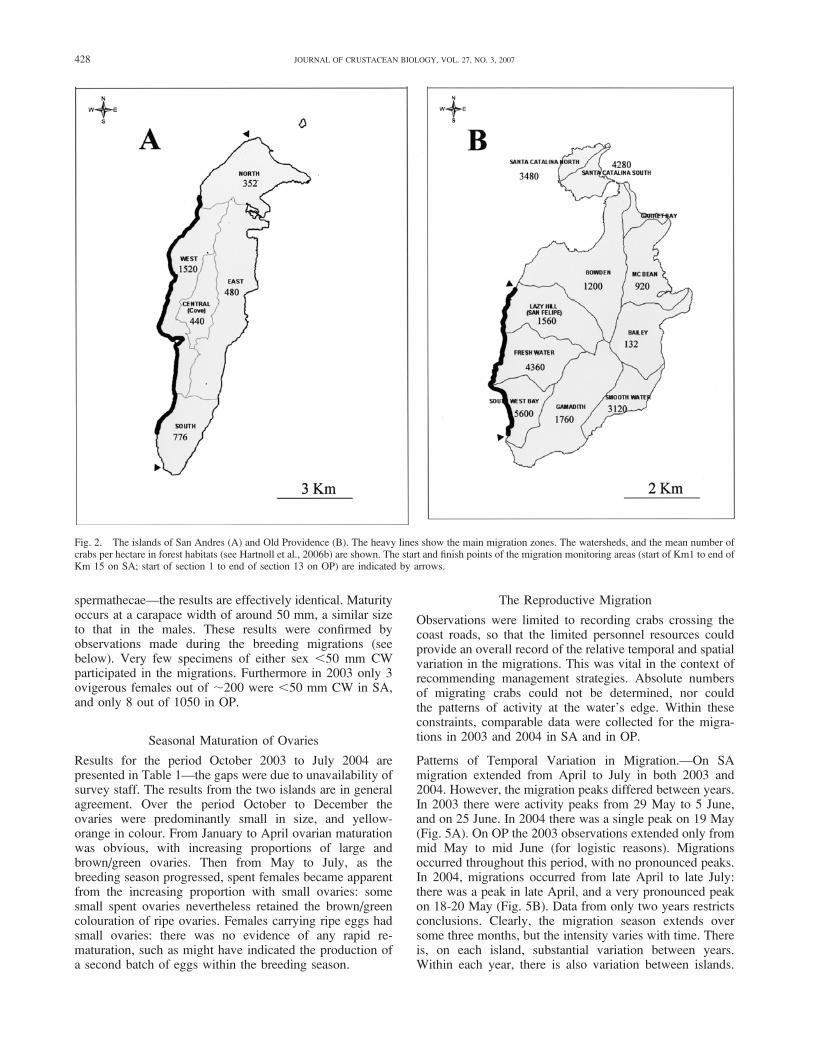
spermathecae—the results are effectively identical. Maturityoccurs at a carapace width of around 50 mm, a similar sizeto that in the males. These results were confirmed byobservations made during the breeding migrations (seebelow). Very few specimens of either sex ,50 mm CWparticipated in the migrations. Furthermore in 2003 only 3ovigerous females out of ;200 were ,50 mm CW in SA,and only 8 out of 1050 in OP.
Seasonal Maturation of Ovaries
Results for the period October 2003 to July 2004 arepresented in Table 1—the gaps were due to unavailability ofsurvey staff. The results from the two islands are in generalagreement. Over the period October to December theovaries were predominantly small in size, and yellow-orange in colour. From January to April ovarian maturationwas obvious, with increasing proportions of large andbrown/green ovaries. Then from May to July, as thebreeding season progressed, spent females became apparentfrom the increasing proportion with small ovaries: somesmall spent ovaries nevertheless retained the brown/greencolouration of ripe ovaries. Females carrying ripe eggs hadsmall ovaries: there was no evidence of any rapid re-maturation, such as might have indicated the production ofa second batch of eggs within the breeding season.
The Reproductive Migration
Observations were limited to recording crabs crossing thecoast roads, so that the limited personnel resources couldprovide an overall record of the relative temporal and spatialvariation in the migrations. This was vital in the context ofrecommending management strategies. Absolute numbersof migrating crabs could not be determined, nor couldthe patterns of activity at the water’s edge. Within theseconstraints, comparable data were collected for the migra-tions in 2003 and 2004 in SA and in OP.
Patterns of Temporal Variation in Migration.—On SAmigration extended from April to July in both 2003 and2004. However, the migration peaks differed between years.In 2003 there were activity peaks from 29 May to 5 June,and on 25 June. In 2004 there was a single peak on 19 May(Fig. 5A). On OP the 2003 observations extended only frommid May to mid June (for logistic reasons). Migrationsoccurred throughout this period, with no pronounced peaks.In 2004, migrations occurred from late April to late July:there was a peak in late April, and a very pronounced peakon 18-20 May (Fig. 5B). Data from only two years restrictsconclusions. Clearly, the migration season extends oversome three months, but the intensity varies with time. Thereis, on each island, substantial variation between years.Within each year, there is also variation between islands.
Fig. 2. The islands of San Andres (A) and Old Providence (B). The heavy lines show the main migration zones. The watersheds, and the mean number ofcrabs per hectare in forest habitats (see Hartnoll et al., 2006b) are shown. The start and finish points of the migration monitoring areas (start of Km1 to end ofKm 15 on SA; start of section 1 to end of section 13 on OP) are indicated by arrows.
428 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 27, NO. 3, 2007
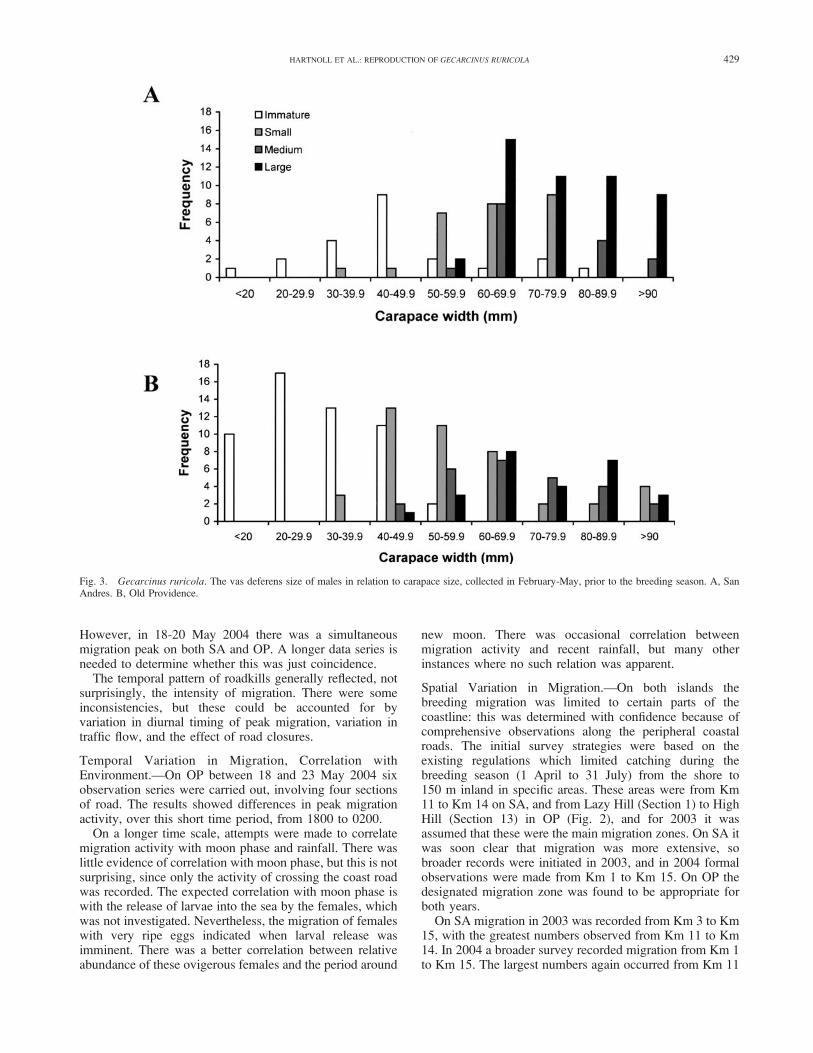
However, in 18-20 May 2004 there was a simultaneousmigration peak on both SA and OP. A longer data series isneeded to determine whether this was just coincidence.
The temporal pattern of roadkills generally reflected, notsurprisingly, the intensity of migration. There were someinconsistencies, but these could be accounted for byvariation in diurnal timing of peak migration, variation intraffic flow, and the effect of road closures.
Temporal Variation in Migration, Correlation withEnvironment.—On OP between 18 and 23 May 2004 sixobservation series were carried out, involving four sectionsof road. The results showed differences in peak migrationactivity, over this short time period, from 1800 to 0200.
On a longer time scale, attempts were made to correlatemigration activity with moon phase and rainfall. There waslittle evidence of correlation with moon phase, but this is notsurprising, since only the activity of crossing the coast roadwas recorded. The expected correlation with moon phase iswith the release of larvae into the sea by the females, whichwas not investigated. Nevertheless, the migration of femaleswith very ripe eggs indicated when larval release wasimminent. There was a better correlation between relativeabundance of these ovigerous females and the period around
new moon. There was occasional correlation betweenmigration activity and recent rainfall, but many otherinstances where no such relation was apparent.
Spatial Variation in Migration.—On both islands thebreeding migration was limited to certain parts of thecoastline: this was determined with confidence because ofcomprehensive observations along the peripheral coastalroads. The initial survey strategies were based on theexisting regulations which limited catching during thebreeding season (1 April to 31 July) from the shore to150 m inland in specific areas. These areas were from Km11 to Km 14 on SA, and from Lazy Hill (Section 1) to HighHill (Section 13) in OP (Fig. 2), and for 2003 it wasassumed that these were the main migration zones. On SA itwas soon clear that migration was more extensive, sobroader records were initiated in 2003, and in 2004 formalobservations were made from Km 1 to Km 15. On OP thedesignated migration zone was found to be appropriate forboth years.
On SA migration in 2003 was recorded from Km 3 to Km15, with the greatest numbers observed from Km 11 to Km14. In 2004 a broader survey recorded migration from Km 1to Km 15. The largest numbers again occurred from Km 11
Fig. 3. Gecarcinus ruricola. The vas deferens size of males in relation to carapace size, collected in February-May, prior to the breeding season. A, SanAndres. B, Old Providence.
429HARTNOLL ET AL.: REPRODUCTION OF GECARCINUS RURICOLA

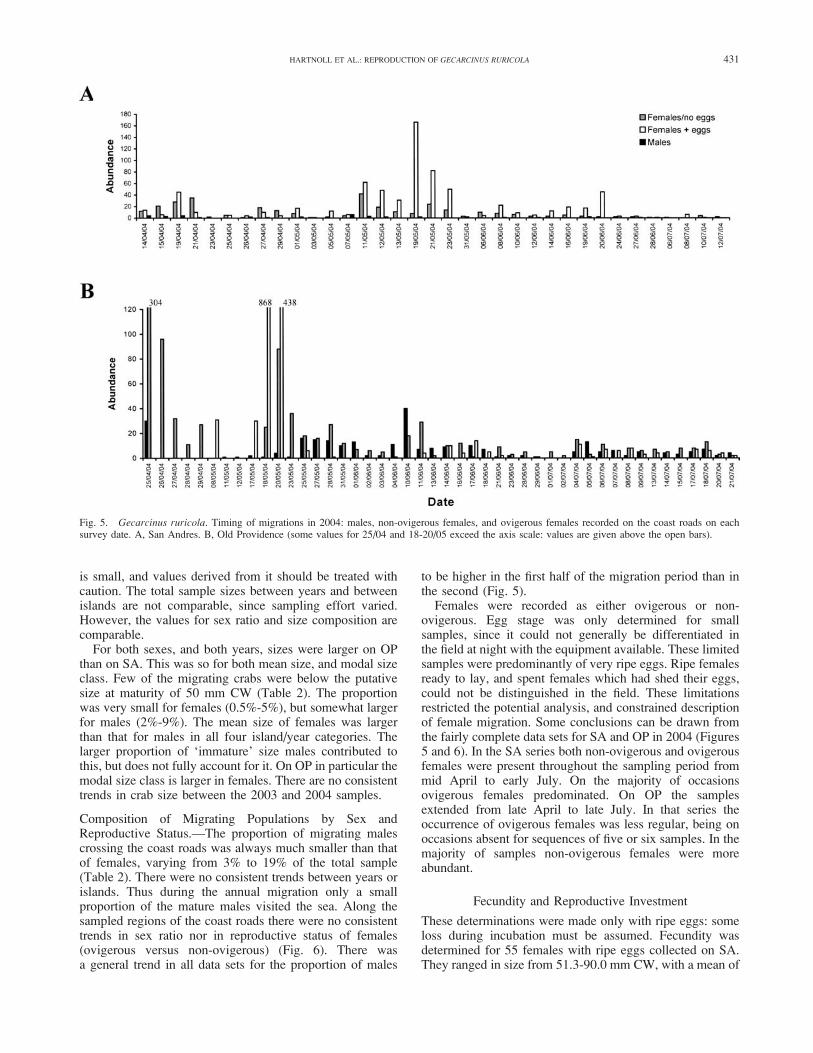
to Km 14, but there were also substantial numbers from Km4 to Km 8 (Fig. 6A). The 2004 data permit a comparison ofthe distribution of live crabs and roadkills. The patterns arebroadly similar, except that roadkills were proportionatelyhigher from Km 3 to Km 7: this reflects the greater trafficdensity in that more populated area.
On OP migration was recorded in all sections of the studyarea. In 2003 the highest numbers were in sections 10 to 12,with substantial numbers also in sections 7 to 9, and 13. In2004 there was a very clear peak in section 11, and lesserabundance in sections 10 and 12 (Fig. 6B). In 2004
distribution of live crabs and roadkills were compared. Themaximum of road kills was found in section 11, butproportionately very high numbers in sections 2 to 7. Thelatter abundance was due to higher traffic density, and thefact that these sections lay outside the usual road closurelimits (normally sections 10 to 13).
Size Composition of Migrating Populations.—Details of thesize composition of migrating crabs, based on samplessexed and measured whilst crossing the coast roads, arepresented in Table 2. The sample of males from SA in 2004
Fig. 4. Gecarcinus ruricola. Reproductive condition in relation to carapace size for females from Old Providence collected in February-May, prior to thebreeding season. A, ovary size. B, ovary colour. C, spermatheca condition.
430 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 27, NO. 3, 2007
is small, and values derived from it should be treated withcaution. The total sample sizes between years and betweenislands are not comparable, since sampling effort varied.However, the values for sex ratio and size composition arecomparable.
For both sexes, and both years, sizes were larger on OPthan on SA. This was so for both mean size, and modal sizeclass. Few of the migrating crabs were below the putativesize at maturity of 50 mm CW (Table 2). The proportionwas very small for females (0.5%-5%), but somewhat largerfor males (2%-9%). The mean size of females was largerthan that for males in all four island/year categories. Thelarger proportion of ‘immature’ size males contributed tothis, but does not fully account for it. On OP in particular themodal size class is larger in females. There are no consistenttrends in crab size between the 2003 and 2004 samples.
Composition of Migrating Populations by Sex andReproductive Status.—The proportion of migrating malescrossing the coast roads was always much smaller than thatof females, varying from 3% to 19% of the total sample(Table 2). There were no consistent trends between years orislands. Thus during the annual migration only a smallproportion of the mature males visited the sea. Along thesampled regions of the coast roads there were no consistenttrends in sex ratio nor in reproductive status of females(ovigerous versus non-ovigerous) (Fig. 6). There wasa general trend in all data sets for the proportion of males
to be higher in the first half of the migration period than inthe second (Fig. 5).
Females were recorded as either ovigerous or non-ovigerous. Egg stage was only determined for smallsamples, since it could not generally be differentiated inthe field at night with the equipment available. These limitedsamples were predominantly of very ripe eggs. Ripe femalesready to lay, and spent females which had shed their eggs,could not be distinguished in the field. These limitationsrestricted the potential analysis, and constrained descriptionof female migration. Some conclusions can be drawn fromthe fairly complete data sets for SA and OP in 2004 (Figures5 and 6). In the SA series both non-ovigerous and ovigerousfemales were present throughout the sampling period frommid April to early July. On the majority of occasionsovigerous females predominated. On OP the samplesextended from late April to late July. In that series theoccurrence of ovigerous females was less regular, being onoccasions absent for sequences of five or six samples. In themajority of samples non-ovigerous females were moreabundant.
Fecundity and Reproductive Investment
These determinations were made only with ripe eggs: someloss during incubation must be assumed. Fecundity wasdetermined for 55 females with ripe eggs collected on SA.They ranged in size from 51.3-90.0 mm CW, with a mean of
Fig. 5. Gecarcinus ruricola. Timing of migrations in 2004: males, non-ovigerous females, and ovigerous females recorded on the coast roads on eachsurvey date. A, San Andres. B, Old Providence (some values for 25/04 and 18-20/05 exceed the axis scale: values are given above the open bars).
431HARTNOLL ET AL.: REPRODUCTION OF GECARCINUS RURICOLA
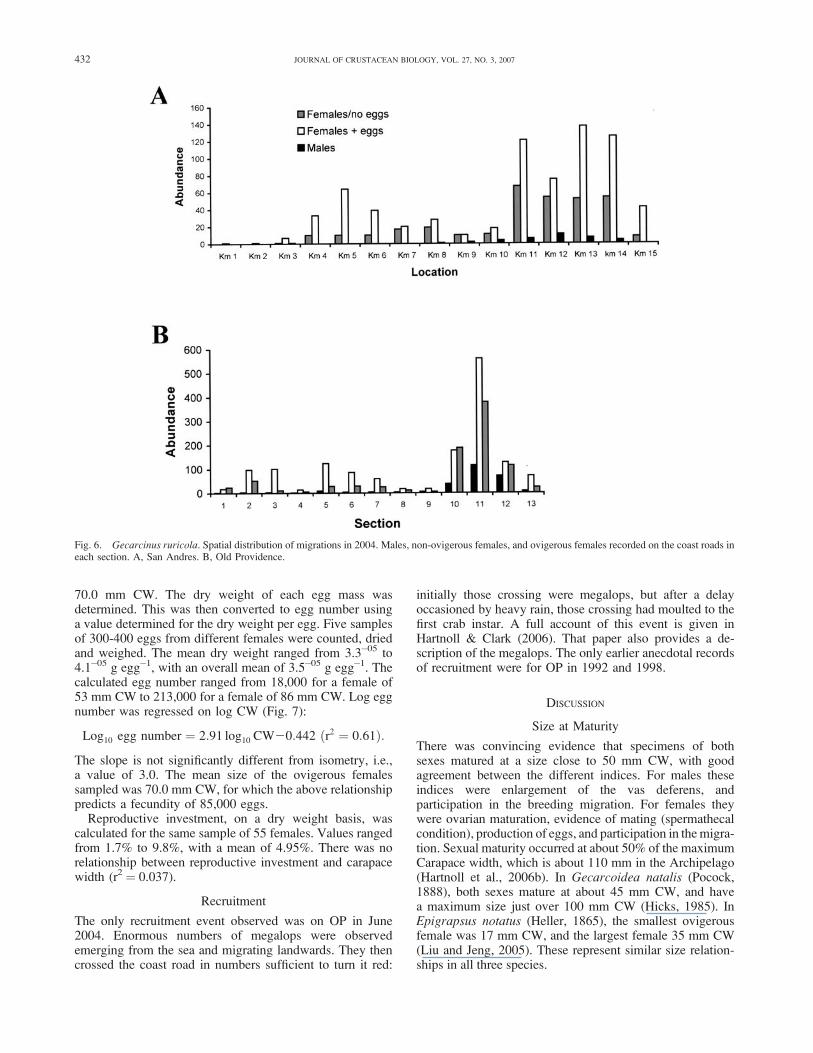
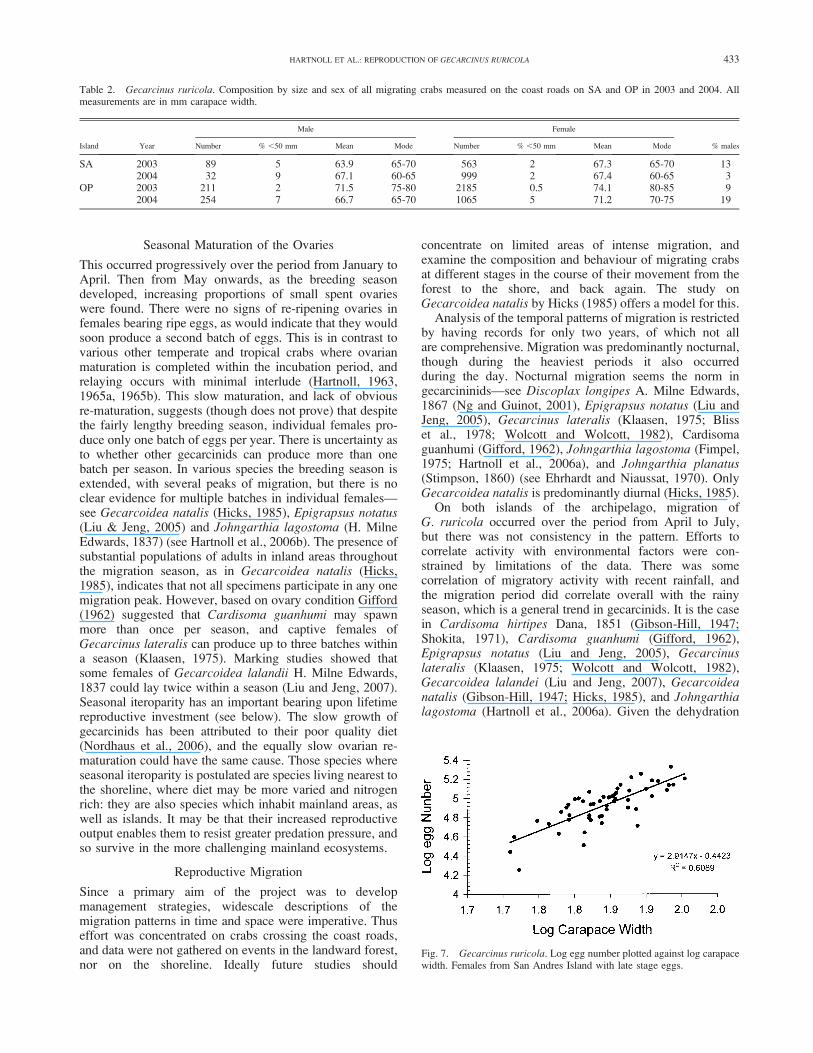
70.0 mm CW. The dry weight of each egg mass wasdetermined. This was then converted to egg number usinga value determined for the dry weight per egg. Five samplesof 300-400 eggs from different females were counted, driedand weighed. The mean dry weight ranged from 3.3�05 to4.1�05 g egg�1, with an overall mean of 3.5�05 g egg�1. Thecalculated egg number ranged from 18,000 for a female of53 mm CW to 213,000 for a female of 86 mm CW. Log eggnumber was regressed on log CW (Fig. 7):
Log10 egg number ¼ 2:91 log10 CW20:442 ðr2 ¼ 0:61Þ:
The slope is not significantly different from isometry, i.e.,a value of 3.0. The mean size of the ovigerous femalessampled was 70.0 mm CW, for which the above relationshippredicts a fecundity of 85,000 eggs.
Reproductive investment, on a dry weight basis, wascalculated for the same sample of 55 females. Values rangedfrom 1.7% to 9.8%, with a mean of 4.95%. There was norelationship between reproductive investment and carapacewidth (r2 ¼ 0.037).
Recruitment
The only recruitment event observed was on OP in June2004. Enormous numbers of megalops were observedemerging from the sea and migrating landwards. They thencrossed the coast road in numbers sufficient to turn it red:
initially those crossing were megalops, but after a delayoccasioned by heavy rain, those crossing had moulted to thefirst crab instar. A full account of this event is given inHartnoll & Clark (2006). That paper also provides a de-scription of the megalops. The only earlier anecdotal recordsof recruitment were for OP in 1992 and 1998.
DISCUSSION
Size at Maturity
There was convincing evidence that specimens of bothsexes matured at a size close to 50 mm CW, with goodagreement between the different indices. For males theseindices were enlargement of the vas deferens, andparticipation in the breeding migration. For females theywere ovarian maturation, evidence of mating (spermathecalcondition), production of eggs, and participation in the migra-tion. Sexual maturity occurred at about 50% of the maximumCarapace width, which is about 110 mm in the Archipelago(Hartnoll et al., 2006b). In Gecarcoidea natalis (Pocock,1888), both sexes mature at about 45 mm CW, and havea maximum size just over 100 mm CW (Hicks, 1985). InEpigrapsus notatus (Heller, 1865), the smallest ovigerousfemale was 17 mm CW, and the largest female 35 mm CW(Liu and Jeng, 2005). These represent similar size relation-ships in all three species.
Fig. 6. Gecarcinus ruricola. Spatial distribution of migrations in 2004. Males, non-ovigerous females, and ovigerous females recorded on the coast roads ineach section. A, San Andres. B, Old Providence.
432 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 27, NO. 3, 2007
Seasonal Maturation of the Ovaries
This occurred progressively over the period from January toApril. Then from May onwards, as the breeding seasondeveloped, increasing proportions of small spent ovarieswere found. There were no signs of re-ripening ovaries infemales bearing ripe eggs, as would indicate that they wouldsoon produce a second batch of eggs. This is in contrast tovarious other temperate and tropical crabs where ovarianmaturation is completed within the incubation period, andrelaying occurs with minimal interlude (Hartnoll, 1963,1965a, 1965b). This slow maturation, and lack of obviousre-maturation, suggests (though does not prove) that despitethe fairly lengthy breeding season, individual females pro-duce only one batch of eggs per year. There is uncertainty asto whether other gecarcinids can produce more than onebatch per season. In various species the breeding season isextended, with several peaks of migration, but there is noclear evidence for multiple batches in individual females—see Gecarcoidea natalis (Hicks, 1985), Epigrapsus notatus(Liu & Jeng, 2005) and Johngarthia lagostoma (H. MilneEdwards, 1837) (see Hartnoll et al., 2006b). The presence ofsubstantial populations of adults in inland areas throughoutthe migration season, as in Gecarcoidea natalis (Hicks,1985), indicates that not all specimens participate in any onemigration peak. However, based on ovary condition Gifford(1962) suggested that Cardisoma guanhumi may spawnmore than once per season, and captive females ofGecarcinus lateralis can produce up to three batches withina season (Klaasen, 1975). Marking studies showed thatsome females of Gecarcoidea lalandii H. Milne Edwards,1837 could lay twice within a season (Liu and Jeng, 2007).Seasonal iteroparity has an important bearing upon lifetimereproductive investment (see below). The slow growth ofgecarcinids has been attributed to their poor quality diet(Nordhaus et al., 2006), and the equally slow ovarian re-maturation could have the same cause. Those species whereseasonal iteroparity is postulated are species living nearest tothe shoreline, where diet may be more varied and nitrogenrich: they are also species which inhabit mainland areas, aswell as islands. It may be that their increased reproductiveoutput enables them to resist greater predation pressure, andso survive in the more challenging mainland ecosystems.
Reproductive Migration
Since a primary aim of the project was to developmanagement strategies, widescale descriptions of themigration patterns in time and space were imperative. Thuseffort was concentrated on crabs crossing the coast roads,and data were not gathered on events in the landward forest,nor on the shoreline. Ideally future studies should
concentrate on limited areas of intense migration, andexamine the composition and behaviour of migrating crabsat different stages in the course of their movement from theforest to the shore, and back again. The study onGecarcoidea natalis by Hicks (1985) offers a model for this.
Analysis of the temporal patterns of migration is restrictedby having records for only two years, of which not allare comprehensive. Migration was predominantly nocturnal,though during the heaviest periods it also occurredduring the day. Nocturnal migration seems the norm ingecarcininids—see Discoplax longipes A. Milne Edwards,1867 (Ng and Guinot, 2001), Epigrapsus notatus (Liu andJeng, 2005), Gecarcinus lateralis (Klaasen, 1975; Blisset al., 1978; Wolcott and Wolcott, 1982), Cardisomaguanhumi (Gifford, 1962), Johngarthia lagostoma (Fimpel,1975; Hartnoll et al., 2006a), and Johngarthia planatus(Stimpson, 1860) (see Ehrhardt and Niaussat, 1970). OnlyGecarcoidea natalis is predominantly diurnal (Hicks, 1985).
On both islands of the archipelago, migration ofG. ruricola occurred over the period from April to July,but there was not consistency in the pattern. Efforts tocorrelate activity with environmental factors were con-strained by limitations of the data. There was somecorrelation of migratory activity with recent rainfall, andthe migration period did correlate overall with the rainyseason, which is a general trend in gecarcinids. It is the casein Cardisoma hirtipes Dana, 1851 (Gibson-Hill, 1947;Shokita, 1971), Cardisoma guanhumi (Gifford, 1962),Epigrapsus notatus (Liu and Jeng, 2005), Gecarcinuslateralis (Klaasen, 1975; Wolcott and Wolcott, 1982),Gecarcoidea lalandei (Liu and Jeng, 2007), Gecarcoideanatalis (Gibson-Hill, 1947; Hicks, 1985), and Johngarthialagostoma (Hartnoll et al., 2006a). Given the dehydration
Table 2. Gecarcinus ruricola. Composition by size and sex of all migrating crabs measured on the coast roads on SA and OP in 2003 and 2004. Allmeasurements are in mm carapace width.
Island Year
Male Female
% malesNumber % ,50 mm Mean Mode Number % ,50 mm Mean Mode
SA 2003 89 5 63.9 65-70 563 2 67.3 65-70 132004 32 9 67.1 60-65 999 2 67.4 60-65 3
OP 2003 211 2 71.5 75-80 2185 0.5 74.1 80-85 92004 254 7 66.7 65-70 1065 5 71.2 70-75 19
Fig. 7. Gecarcinus ruricola. Log egg number plotted against log carapacewidth. Females from San Andres Island with late stage eggs.
433HARTNOLL ET AL.: REPRODUCTION OF GECARCINUS RURICOLA

risk during the extensive overland migrations to the sea theadvantages of a humid environment are clear. In Gecarcinusruricola, there was evidence that the migration of femaleswith ripe eggs was greater at the period around new moon,suggesting that larval release may be concentrated at suchperiods. Lunar rhythms of egg hatching are recorded inother land crabs. In Gecarcoidea natalis, it is concentratedin the last quarter (Hicks, 1985), as it is in G. lalandii (Liuand Jeng, 2007). There was greater larval release at fullmoon in Columbia for Gecarcinus lateralis (Klaasen, 1975),and immediately after full moon in Bermuda (Wolcott andWolcott, 1982). In Epigrapsus notatus, larval release is inthe period following full moon (Liu and Jeng, 2005). InCardisoma guanhumi, spawning is concentrated near thefull moon (Gifford, 1962).
The spatial limits of migration were consistent on eachisland, both with regard to our observations, and in relationto anecdotal data from previous years. The data are reliable,since the continuous coast roads on each island facilitatecomprehensive observation. Road kills would clearlyindicate if migrations were occurring in other areas (otherthan on Santa Catalina, where there is no continuous road).The intensity of migration appears to differ on the basis ofobservations of live crabs, and of road kills. However, thiscan be explained by differences in traffic density, and by theeffect of selective road closures. The reasons for the limitedmigration zones are not obvious. The migration zones areadjacent to the areas of highest crab density (Hartnoll et al.,2006b), and on OP (no data are available for SA) they arethe same as the zones of recruitment (Hartnoll & Clark,2006). Thus the parsimonious explanation is that the adultcrabs predominantly remain in the areas close to where theyrecruited, and later retrace their steps to the sea. OnChristmas Island, however, both the migration of the adultsand the return of the megalops occurred on all sides of theisland (Hicks, 1985).
In Gecarcinus ruricola, the crabs crossing the coast roadwere predominantly females, males always comprising lessthat 20% of the total, often much less. It is clear that fewmales reached the sea, and it must be assumed that matingactivity is concentrated on the landward side of the coastroad. In Johngarthia lagostoma, females also predominate5:1 at the shoreline (Hartnoll et al., 2006a). This contrastswith Gecarcoidea natalis, where males and females migrateto the sea in similar numbers, and only after both indulge in‘dipping’ in the sea, does courtship and mating occur(Hicks, 1985). In contrast, in the cavernicolous Discoplaxlongipes, no males were observed in the migration (Ng andGuinot, 2001). There is a clear inter-specific variation fromno migrating males, to males and females migrating in equalnumbers. The structure of the female genital opening helpsto explain this variation. In the gecarcinids the femaleopenings are normally occluded by a rigid calcifiedoperculum: see Hartnoll (1968a) for Gecarcinus ruricola,and Hartnoll (unpublished) for Cardisoma, Gecarcoideaand Johngarthia. It is probably a general feature of thefamily. Local decalcification allows this operculum tobecome mobile for a period of days, and only during thatinterval is the female able to copulate, and to lay eggs (seeHartnoll, 1968b for example). This short term availability of
females entrains male behaviour. Thus where females lay intheir residential area (as in Discoplax), they are notsubsequently available to males, so no males migrate.Conversely, in Gecarcoidea natalis females lay only afterdipping in the sea, so there must be a mass male migration tobenefit from this limited shoreline female availability.Gecarcinus ruricola occupies an intermediate status. Somefemales lay (and therefore mate) prior to the final migrationto the sea, whilst others mate and lay only near to the shore.Thus a proportion of males migrate with the females, butfemales are in the majority.
The size composition of the migrating crabs wassurprising, in that overall males were smaller than females.This is in contrast to size distributions determined both inthe population studies, and from the market samplingsurveys (Hartnoll et al., 2006b), where males were larger ineach case. From the sex composition of migrating crabs it isclear that only a small proportion of males migrate close tothe sea, and it may be that these are ones which have hadless success in mating during earlier stages of the migration.Since larger males in a variety of crabs have been shown tobe competitively superior, it could be expected that theresidual unmated migrant males will be generally smaller.
The females crossing the coast roads comprised bothsome without eggs, and also a proportion bearing eggs. Thelatter were almost entirely composed of females with veryripe eggs, and it is postulated that they were proceeding tothe sea for imminent larval release. Females with early stageeggs were very rare, and this concurs with the generalpattern in gecarcinids—see Epigrapsus notatus (Liu andJeng, 2005), Gecarcinus lateralis (Bliss et al., 1978),Gecarcoidea natalis (Hicks, 1985), and Johngarthialagostoma (Hartnoll et al., 2006a). In gecarcinids generallyfemales lay near the end of the migration, and incubate theeggs within burrows or other refuges, presumably to protectthem from desiccation (Klaassen, 1975; Bliss et al., 1978),and emerge only to progress to the sea for hatching.Klaassen (1975) suggests that Gecarcinus lateralis mustrelease its larvae within 10 hours of emerging from itsburrow. Exceptionally, in the cavernicolous Discoplaxlongipes eggs are laid and incubated in the caves wherethey live, and the females with ripe eggs migrate directly tothe sea (Ng and Guinot, 2001). In Gecarcinus ruricola, theproportion of ovigerous females crossing the coast road washigher on SA than on OP. This could be because the coastroad is generally further from the sea on OP, providing moresuitable areas for shelter, laying and incubation on theseaward side.
Fecundity and Reproductive Investment
Fecundity and reproductive investment could only bedetermined from ripe egg masses, since early eggs wererarely encountered. Thus since there must be some egg lossduring incubation, values will be lower that those de-termined for early eggs, as is the standard protocol (seeHines, 1982). However, the difference is unlikely to exceed10% (Oh & Hartnoll, 1999). Fecundity was determined asbetween 18,000 and 213,000, with a calculated 85,000 eggsat 70 mm CW. In Gecarcinus lateralis, egg number rangedfrom 20,000 to 110,000, but relation with CW was notclarified (Klaasen, 1975). In Epigrapsus notatus, egg
434 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 27, NO. 3, 2007

number ranged from 12,000 to 58,000 eggs, with ;45,000at 30 mm CW. In Cardisoma guanhumi, numbers rangedfrom 20,000 to .600,000 (Henning, 1975), but againwithout relation to body size being defined. These data arehard to compare; nevertheless, the egg number forG. ruricola at 70 mm CW (;70 g DW) is well below theglobal estimate of 250,000 for crabs of that weight (Hines,1982).
Reproductive investment averaged ;5% per brood. Sinceonly a single brood is postulated each year, annual RI is also5%. The reproductive period probably extends over 10 yearsor more, giving a lifetime RI of .50%. The RI is a betterbasis than fecundity for comparing reproductive outputbetween species, since it is not confounded by variation inegg size. Values for other land crabs are not available, butfor a wide range of marine crabs the RI per brood averages10%, though a number of marine species have values of 5%or less (Hines, 1982; Hartnoll, 2006). However, if the valuefor G. ruricola is typical of gecarcinids, then it is relativelylow. There are two potential explanations for this low value.One is the problem of accumulating sufficient resources forovarian maturation on a low-energy and low-nitrogen diet ofpredominantly plant material (see discussion and referencesin Nordhaus et al., 2006). An alternative suggestion is thatthe size of the egg mass is constrained by the need toprovide an adequate oxygen supply to the developingembryos in the interior of the egg mass. In marine crabs,there can be reduction of oxygen levels within the egg mass,and the females employ special behaviour patterns to detectand remedy such a problem (see discussion and referencesin Baeza and Fernandez, 2002). No studies have been madeon terrestrial crabs, such as G. ruricola, which do not accessfree water during incubation. For them the problems ofoxygenation could be greater, limiting viable egg mass size.
The lifetime RI of .50% for G. ruricola is hard tocompare with other species, since the life span of most crabsis poorly documented. However, it is lower than the annualRI for the small number of free-living tropical marinespecies which have been studied (Hartnoll, 2006). Sodespite its longevity, G. ruricola nevertheless has a relativelylow lifetime fecundity. This will increase the problems ofpopulation maintenance.
Recruitment
Only a single recruitment event was observed, on OP inJune 2004. This is fully described in Hartnoll and Clark(2006), and little can be added to the discussion in thatpaper. The only other record of mass recruitment inG. ruricola was in Jamaica in 2006, where again there areno other recent records (Karl Aiken, personal communica-tion). A common feature of the two events above is that theyboth occurred close to full moon: further records are neededto see if this is a consistent aspect. Recruitment is spasmodicand unpredictable, seemingly a general feature of gecarci-nids. Consequently the long duration of reproductive life,exceeding 10 years or so, must be an important factor insustaining populations in the face of uncertain recruitment.Populations can be heavily biased to larger sizes, notably asin Johngarthia lagostoma on Ascension Island (Hartnollet al., 2006a), suggesting recruitment bottlenecks. However,
it may be that sampling bias plays a substantial role ingenerating such distributions.
ACKNOWLEDGEMENTS
This investigation was supported by the UK Darwin Initiative for theSurvival of Species, project 162/11/015, ‘Sustainable management of theblack land crab, Gecarcinus ruricola, Colombia’. Many thanks to the non-author staff of the Columbian Government Agency CORALINA in OldProvidence and San Andres for help with the collection and analysis ofmaterial, and for logistic support in the Archipelago. Thanks also to thoseisland crab catchers who assisted in the fieldwork—without their expertisethis study would not have been possible. We also thank the Natural HistoryMuseum, London, for the loan of material of Cardisoma and Gecarcoidea.Dr Karl Aiken of the University of West Indies, Jamaica, generouslyprovided unpublished details of the mass recruitment event in Jamaica.
REFERENCES
Baeza, J. A., and M. Fernandez. 2002. Active brood care in Cancer setosus(Crustacea: Decapoda): the relationship between female behaviour,embryo oxygen consumption and the cost of brooding. FunctionalEcology 16: 241-251.
Baine, M., M. Howard, S. Kerr, G. Edgar, and V. Toral. 2007. Coastal andmarine resource management in the Galapagos Islands and theArchipelago of San Andres: Issues, problems and opportunities. Oceanand Coastal Management 50: 148-173.
———, ———, E. Taylor, J. James, A. Velasco, Y. Grandas, and R. G.Hartnoll. in press. The development of management options for the blackland crab (Gecarcinus ruricola) catchery in the San Andres Archipelago,Colombia. Ocean and Coastal Management.
Bliss, D. E., J. van Montfrans, M. van Montfrans, and J. R. Boyer. 1978.Behaviour and growth of the land crab Gecarcinus lateralis (Freminville)in Southern Florida. Bulletin of the American Museum of NaturalHistory 160: 111-152.
Britton, J. C., G. C. Kroh, and C. Golightly. 1982. Biometric and ecologicalrelationships in two sympatric Caribbean Gecarcinidae (Crustacea:Decapoda). Journal of Crustacean Biology 2: 207-222.
Calman, W. T. 1911. The Life of Crustacea. Methuen & Co., London.Capistran-Barrados, A., O. Defoeo, and P. Moreno-Casasola. 2003. Density
and population structure of the red land crab Gecarcinus lateralis ina tropical semi-deciduous forest in Veracruz, Mexico. Interciencia 28:323-345.
Carson, H. L. 1967. The association between Drosophila carcinophilaWheeler and its host, the land crab Gecarcinus ruricola (L.). AmericanMidland Nauralist 78: 324-343.
———. 1974. Three flies and three islands: Parallel evolution inDrosophila. Proceedings of the National Academy of Science 71:3517-3521.
Chace, F. A., and H. H. Hobbs. 1969. The freshwater and terrestrialdecapod crustaceans of the West Indies with special reference toDominica. Bulletin of the U.S. National Museum 292: 1-258.
Costlow, J. D., and C. G. Bookhout. 1968. The complete larval develop-ment of the land-crab, Cardisoma guanhumi Latreille in the laboratory(Brachyura, Gecarcinidae). Crustaceana Supplement 2: 259-270.
Ehrhardt, J.-P., and P. Niaussat. 1970. Ecologie et physiologie dubrachyoure terrestre Gecarcinus planatus Stimpson (d’apres les individsde l’atoll de Clipperton). Bulletin de la Societe Zoologique de France 95:41-54.
Fimpel, E. 1975. Phanomene der Landadaptation bei terrestrischen undsemiterrestrischen Brachyura der brasilianischen Kuste (Malacostraca,Decapoda). Zoologische Jahrbucher Systematik 102: 173-214.
Gibson-Hill, C. A. 1947. Field notes on the terrestrial crabs. Bulletin of theRaffles Museum 18: 43-52.
Gifford, C. A. 1962. Some observations on the general biology of the landcrab, Cardisoma guanhumi (Latreille), in south Florida. BiologicalBulletin 123: 207-223.
Hartnoll, R. G. 1963. The biology of Manx spider crabs. Proceedings of theZoological Society of London 141: 423-496.
———. 1965a. The biology of spider crabs: a comparison of British andJamaican species. Crustaceana 9: 1-16.
———. 1965b. Notes on the marine grapsid crabs of Jamaica. Proceedingsof the Linnean Society of London 176: 113-147.
———. 1968a. Morphology of the genital ducts in female crabs. Journal ofthe Linnean Society (Zoology) 47: 279-300.
435HARTNOLL ET AL.: REPRODUCTION OF GECARCINUS RURICOLA

———. 1968b. Reproduction in the burrowing crab Corystes cassivelaunus(Pennant, 1777) (Decapoda, Brachyura). Crustaceana 15: 165-170.
———. 1988. Evolution, systematics and geographical distributionpp. 6-54. In, W. W. Burggren and B. R. McMahon (eds.), Biology ofLand Crabs. Cambridge University Press.
———. 2006. Reproductive investment in Brachyura. Hydrobiologia 557:31-40.
———, and P. Clark. 2006. A mass recruitment event in the land crabGecarcinus ruricola (Linnaeus, 1758) [Brachyura: Grapsoidea: Gecarci-nidae], and a description of the megalop. Zoological Journal of theLinnean Society 146: 149-164.
———, T. Mackintosh, and T. J. Pelembe. 2006a. Johngarthia lagostoma(H. Milne Edwards, 1837) on Ascension Island: a very isolated land crabpopulation. Crustaceana 79: 197-215.
———, M. S. P. Baine, Y. Grandas, J. James, and H. Atkin. 2006b.Population biology of the black land crab, Gecarcinus ruricola, in theSan Andres Archipelago, Western Caribbean. Journal of CrustaceanBiology 26: 316-325.
Henning, H. G. 1975. Okologische, ethologische und sinnesphysiologischean der Landkrabbe Cardisoma guanhumi Latreille (Decapoda, Bra-chyura) in Nordcolumbien. Forma et Functio 8: 253-304.
Hicks, J. W. 1985. The breeding behaviour and migrations of the terrestrialcrab Gecarcoidea natalis. Australian Journal of Zoology 33: 127-142.
Hines, A. S. 1982. Allometric constraints and variables of reproductiveeffort in brachyuran crabs. Marine Biology 69: 309-320.
Hoeven, E. C. van der, and L. M. H. Walters. 1988. Meer over de Ecologieen de Populatiedynamiek van de Landkrab Gecarcinus ruricola(Latreille) op Saba. De Mogelijkheden tot ‘‘Crabfarming’’ op Saba.Proceedings of the Saba Crab Research Centre 2: 1-36.
Keith, D. E. 1985. Shallow-water and terrestrial brachyuran crabs of Roatanand the Swan Islands. Sarsia 70: 251-278.
Klaassen, F. 1975. Okologische und ethologische Untersechungen zurFortpflanzungsbiologie von Gecarcinus lateralis (Decapoda, Brachyura).Forma et Functio 8: 101-174.
Liu, H.-C., and M.-S. Jeng. 2005. Reproduction of Epigrapsus notatus(Brachyura: Gecarcinidae) in Taiwan. Journal of Crustacean Biology 25:135-140.
———, and ———. 2007. Some aspects of the reproduction ofGecarcoidea lalandii (Brachyura: Gecarcinidae) in Taiwan. ZoologicalStudies 46: 347-354.
Magin, C. 2003. Development of a National Strategy for SustainableWildlife Use for the Commonwealth of Dominica. Wildlife SurveyReport. Unpublished Report, Fauna and Flora International, Cambridge.
Maitland, D. P. 2002. Convergent design of Caribbean and Philippinebamboo land-crab traps. Journal of Crustacean Biology 22: 497-501.
Mulder, A., and E. Stam. 1987. Ecologische en PopulatiedynamischeWaarnemingen aan de Landkrab Gecarcinus ruricola (Latreille) op Sabaand Ein Simulatiensmodel voor de geexploiteerde Populatie. Proceedingsof the Saba Crab Research Centre 1: 1-40.
Ng, P. K. L., and D. Guinot. 2001. On the land crabs of the genusDiscoplax A. Milne. Edwards, 1867 (Crustacea: Decapoda: Brachyura:Gecarcinidae), with description of a new cavernicolous species from thePhilippines. The Raffles Bulletin of Zoology 49: 311-338.
Nordhaus, I., M. Wolff, and K. Diele. 2006. Litter processing andpopulation food intake of the mangrove crab Ucides cordatus in a highintertidal forest in northern Brazil. Estuarine Coastal and Shelf Science67: 239-250.
Oh, C.-W., and R. G. Hartnoll. 1999. Brood loss during incubation inPhilocheras trispinosus (Decapoda) in Port Erin Bay, Isle of Man.Journal of Crustacean Biology 19: 467-476.
O’Mahoney, P. M., and R. J. Full. 1984. Respiration of crabs in air andwater. Comparative Biochemistry and Physiology 79A: 275-282.
Parsons, J. J. 1956. San Andres and Providencia. English speaking islandsin the western Caribbean. Public Geography 12: 1-84.
Prahl, H. Von, and G. Manjarres. 1983. Cangrejos gecarcınidos (Crustacea:Gecarcinidae) de la Isla de Providencia, Columbia. Caribbean Journal ofScience 19: 31-34.
Shokita, S. 1971. On the spawning habits of the land crab Cardisomahirtipes Dana from Ishigaki Island, in the Ryuku Islands. BiologicalMagazine of Okinawa 7: 27-32.
Sjogreen, M. 1999. Estudio bioecologico de la poblacion de cangrejosGecarcinus ruricola en las isles de Providencia y Santa Catalina.Dissertation, Pontificia Universidad Javeriana, Bogota.
Stebbing, T. R. R. 1893. A History of Crustacea. Kegan Paul, Tench,Trubner & Co., London.
Willems, K. A. 1982. Larval development of the land crab Gecarcinuslateralis lateralis (Freminville, 1835) (Brachyura: Gecarcinidae) rearedin the laboratory. Journal of Crustacean Biology 2: 180-201.
Wolcott, T. G. 1984. Uptake of interstitial water from soil: mechanisms andecological significance in the ghost crab Ocypode quadrata and twogecarcinid land crabs. Physiological Zoology 57: 161-184.
———, and D. L. Wolcott. 1982. Larval loss and spawning behaviour inthe land crab Gecarcinus lateralis (Freminville). Journal of CrustaceanBiology 4: 477-485.
———, and ———. 1988. Availability of salts is not a limiting factor forthe land crab Gecarcinus lateralis (Freminville). Journal of ExperimentalMarine Biology and Ecology 120: 199-219.
RECEIVED: 6 June 2006.ACCEPTED: 4 December 2006.
436 JOURNAL OF CRUSTACEAN BIOLOGY, VOL. 27, NO. 3, 2007




















