
a precision launch technique for insensitive munitions ... - DTIC
Upload
independentCategory
view
1download
0
Plant Molecular Biology 24: 561-569, 1994. © 1994 Kluwer Academic Publishers. Printed in Belgium. 561
Regulation of the rabl7 gene promoter in transgenic Arabidopsis wild-type, ABA-deficient and ABA-insensitive mutants
Josep Vilardell 1, Jos6 M.Martinez-Zapater 2, Adela Goday 1 Cesar Arenas i and Montserrat Pages 1,, 1Departamento de GenOtica Molecular, Centro de Investigacidn y Desarrollo, C.S.I.C., Jorge Girona Salgado 18-26, 08034 Barcelona, Spain (* author for correspondence)," 2 Departamento de Proteccidn Vegetal, I.N.I.A., Carretera de la Coru~a, Km 7.8111, Madrid, Spain
Received 15 September 1993; accepted in revised form 29 November 1993
Key words." Arabidopsis, ABA mutants, ABA-responsive genes, promoter, transgenic plants
Abstract
The abscisic acid-responsive gene rabl7 is induced during maize embryo maturation and in vegetative tissues under water stress conditions. To investigate how ABA is involved in the induction of the rabl7 gene, we present here a genetic approach to analyse the transcriptional regulation of the 1.3 kb rabl7 promoter fragment in transgenic wild-type Arabidopsis and mutants which are deficient (aba) and in- sensitive (abil, abi2 and abi3) to ABA. During seed development the rabl7 promoter fragment confers similar temporal and spatial regulation on the reporter gene GUS, both in transgenic wild-type and ABA-deficient and ABA-insensitive mutants. The rabl7 promoter was only active in embryo and en- dosperm during late seed development, although the ABA-deficient embryo mutant showed a reduction in the level of GUS activity. During germination rabl7 promoter activity decreases, and GUS activity is not enhanced by water stress in transgenic wild-type and mutant plants. In contrast, transcription of the Arabidopsis endogenous rab gene is stimulated by water stress, both in wild-type and ABA-insensitive mutants. Our data suggest that different molecular mechanisms mediate seed-specific expression and ABA water stress induction of the rabl7 gene and indicate strong conservation of the seed-specific regulatory mechanism for rab genes in monocot and dicot plants.
Introduction
Abscisic acid (ABA) is supposed to mediate a large number of physiological and developmental processes in plants such as embryo maturation, seed dormancy and the response of plants to os- motic stress [28]. The response of the plant cells to ABA include modulation of specific mRNAs and proteins in different tis sues [ 23, 21 ]. Although the molecular basis of ABA's effect on gene ex- pression has been the subject of numerous stud- ies, to date little progress has been made in the elucidation of the intermediates in the ABA sig- nal transduction pathway.
Many genes responsive to ABA (rab) are con- trolled at the transcriptional level. Functional dis- section of rab promoters by using transient ex- pression assays in protoplasts has revealed the existence of a specific sequence element (ABA- responsive element, ABRE) which confers ABA responsiveness upon the reporter gene [13, 14, 27]. Moreover, leucine zipper-type proteins bind- ing to the ABRE sequence have been identified [7, 17]. However, recent studies have shown that the expression of rab genes is also controlled by a variety of other processes and that changes in ABA levels are not always related with changes in gene expression [3, 16, 18].

562
The maize rab17 gene is expressed in different plant parts in response to exogenous ABA and osmotic stress [25]. Functional analysis of the rab 17 promoter showed developmental and ABA responsiveness of the reporter gene in transgenic tobacco and in transfected protoplasts [26]. However, no clear relationship between the en- dogenous ABA levels and the abundance of tab mRNAs was found during embryogenesis in maize embryo mutants deficient or insensitive to ABA [18, 19].
To further investigate the role of ABA in the control of rab gene expression we have initiated a genetic approach to characterize the transcrip- tional regulation of the maize ABA-responsive rabl7 promoter using transgenic Arabidopsis plants. The genetic tools existing in this species can allow the genetic dissection of the ABA- mediated signal transduction pathway. Moreover, a set of mutants which are deficient (aba) and insensitive (abiI, abi2 and abi3) to ABA have been isolated in Arabidopsis. These mutants are characterized genetically by their ability to re- spond to ABA and by a variety of ABA-inducible responses throughout the life cycle of the plant [4, 9, 10, 11]. Recently, the locus abi-3 has been cloned [5] and new abi-3 alleles have been iden- tified [15]. Thus, these mutations provide an in- valuable tool for the dissection of the ABA effects versus developmental and physiological cues in- volved in tab gene expression.
We have analysed the transcriptional regula- tion of the 1.3 kb rabl7 promoter fragment in wild-type transgenic Arabidopsis plants and in isogenic lines homozygous for mutations in the aba or the abi loci. Six transgenic plants carrying independent insertions of the rabl7/GUS were obtained. A similar pattern of GUS activity for all test transgenic lines was found. One transgenic wild-type line, La 1011, was crossed with ABA- deficient and ABA-insensitive mutants as de- scribed in Materials and methods. Here we present the detailed study of La 1011 and its de- rived isogenic mutant lines aba, abil, abi2 and abi3. We show that the chimeric rabl7/GUS gene is developmentally regulated during seed forma- tion in the wild-type and in the four mutants
tested; however, under water stress, no activity of the tab promoter can be detected in wild-type and mutant plants containing normal levels of ABA.
Materials and methods
Plant material
Arabidopsis thaliana (L.) Heynh. ecotype Lands- berg erecta (Ler) and mutants generated in this background affecting ABA synthesis (aba- 1) [ 9], or ABA responsiveness (abi-l-1, abi-2-1 and abi- 3-1) [10], were kindly provided by Maarten Koornneef (Agricultural University, Wageningen, Netherlands).
For germination seeds were surface-sterilized in 5~o hypochlorite, 0.02~o Triton X-100 then rinsed five times with sterile water before plating on minimal medium composed of mineral nutri- ents (8) and 0.7~o agar. Plated seeds were incu- bated 24 h at 4 °C and transferred to chambers where seedlings were grown at 22 °C under con- tinuous illumination. Transformed plants were grown to maturity in Aracons (Beta Tech, Bel- gium) in environmentally controlled growth rooms maintained for 16 h day/8 h night at 22 °C. For water stress plants were unwatered for the last 5 days before collection.
Transgenic plants
For stable transformation we used a construction in which the -1330/+ 29 region of the rabl7 pro- moter was ligated into the polylinker of the plas- mid pB1101.1 [27 ]. The construction was verified by restriction site analysis and sequencing. The plasmid containing the rabl7/GUS gene fusion was transformed into Agrobacterium tumefaciens (strain LBA4404) and transferred to Arabidopsis via root transformation [24]. Kanamycin-resis- tant plants regenerating from tissue culture (T1) generation were rooted and transferred to soil. Seeds from the T1 plants were harvested and the presence of T-DNA inserts was confirmed by ger- mination on GM plates containing 50 mg/1 of kanamycin. Lines homozygous for the T-DNA insertion were initiated from T2 plants that gave

100 ~o of kanamycin-resistant seedlings after self- ing. Six independent insertions were fixed in this way. After analysis of the expression of the chi- meric gene constructs in these transgenic lines the three lines Lal011, Lal012 and Lal013 showing higher levels of seed expression were used for further studies. Segregation of GUS activity was 3:1 in mature T2 seeds, indicating that the trans- formants contained one functional locus of the transgene per haploid genome. To study the same T-DNA insertions in different mutant back- grounds, homozygous ABA-mutant lines carry- ing the T-DNA insertions were generated by crossing homozygous mutants (aba-1, abi 1-1, abi 2-1 and abi 3-1) with the homozygous T-DNA wild-type lines Lal011, Lal012, Lal013, and se- lecting in the F2 generation for plants that were kanamycin resistant and showed the mutant phe- notype. The genotype of these plants was con- firmed by analyzing the phenotypes of the F3 derived plants. The aba and abil, abi2 and abi3 mutants derived by crossing with the line Lal011 were further analysed.
Molecular analysis
Genomic DNA was isolated as described by Del- laporta etal. [2]. Enzymatic amplification by PCR was performed using Taq DNA polymerase (Stratagene, La Jolla, CA) as described by Saiki et al. [22]. The primers used for amplifications were nucleotides from -758 to -738, and from -19 to -1 [25]. The plasmid grabl7 was used as positive template DNA. The conditions for the PCR and northern hybridization were as previ- ously described [26]. The Arabidopsis rabl8 cDNA [12] was kindly provided by Dr Tapio Palva (Uppsala Genetic Center, Uppsala, Swe- den).
GUS assays
Transgenic plants were assayed histochemically for GUS enzyme activity as follows. Freshly cut tissues, young seedlings, embryos manually dis- sected from the seeds at different days after flow- ering (d.a.f.), and the seed coat with the attached
563
endosperm were immersed in 50 mM NaH2PO 4 pH 7.0, containing 0.1~o Triton X-100 (Sigma) and 1 mM 5-bromo-4-chloro-3-indolylglucu- ronide (X-Gluc) and incubated at 37 °C over- night. After incubation, samples were rinsed in 50 mM NaH2PO 4 pH 7.0 and cleared in three changes of 70~o ethanol. Levels of GUS activi- ties were quantitated by fluorometry. Typically, 2 to 2.5 mg of seeds (150 dry seeds) or 20 seedlings representing each independent transgenic line were pooled and three sets of assays were per- formed. In seedlings special care was taken to remove any rest of seed coat attached to seedlings to avoid GUS activity from the endosperm tissue. Frozen tissues were ground in liquid nitrogen and proteins extracted with 1 ml of buffer 50 mM NaH2PO4 pH 7.0, 10 mM 2-mercaptoethanol, 10mM EDTA, 0.1~o N-lauroylsarcosine, 0.1~o Triton X-100). After centrifugation for 3 min at 4 °C aliquots of the supernatant were used to assay after addition of 22 mg/50 ml of 4-methy- lumbelliferone (4-MU). Protein concentration in the extracts were measured using the Bradford [ 1 ] assay. Non transformed plants and transgenic wild-type Arabidopsis plants with the 35S/GUS gene were used as controls.
Results
Molecular analysis of wild-type and mutant trans- genic lines
To analyse the effect of mutations causing ABA- deficiency or insensitivity on the regulation of the rab promoter the transgenic wild-type line LA1011 carrying specific single insertion of the rab/GUS gene was crossed with ABA-deficient (aba) and ABA-insensitive (abil, abi2 and abi3) mutants. The four mutant plants carrying the same T-DNA insertion as Lal011 were isolated.
Molecular hybridization experiments were car- ried out to confirm that the transformants con- tained the complete insert. Total genomic DNA from Lal011 was digested and hybridized with the tab17 probe. Hybridization of the probe to a 1.3 kb fragment in Lal011 DNA indicated that it contained intact copies of the rab 17 promoter unit
564
(not shown). This hybridizing fragment was ab- sent in DNA from untransformed plants.
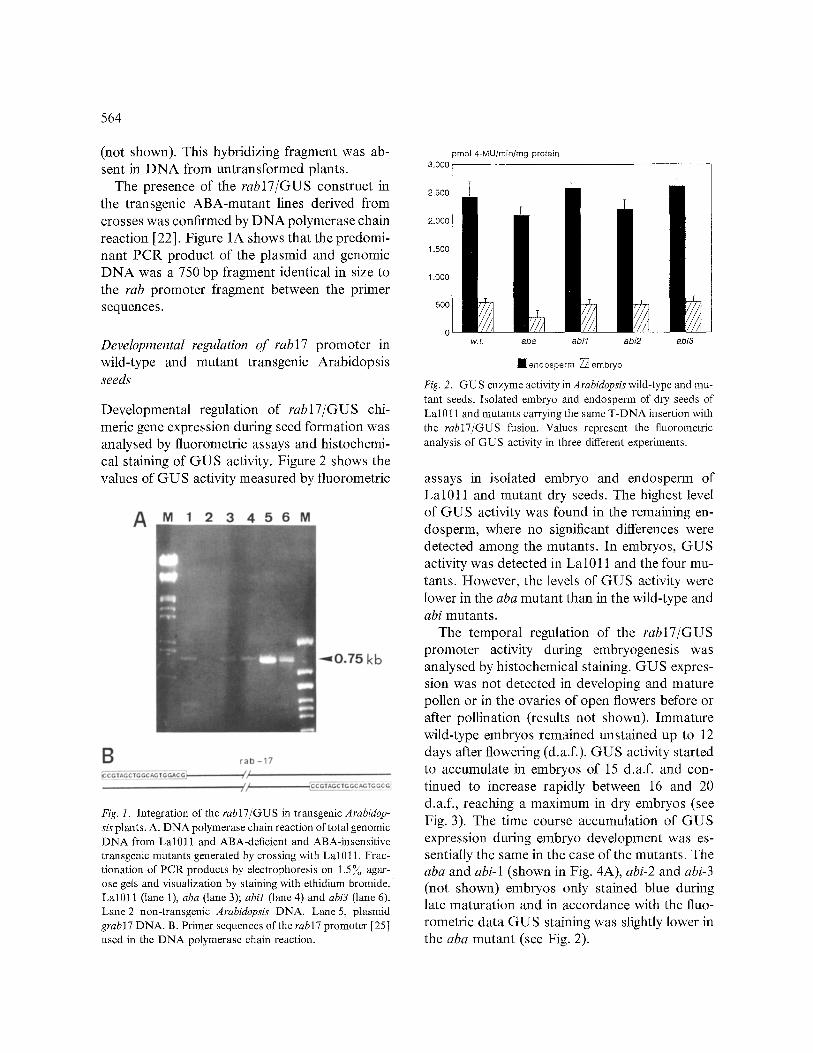
The presence of the rabl7/GUS construct in the transgenic ABA-mutant lines derived from crosses was confirmed by DNA polymerase chain reaction [22]. Figure 1A shows that the predomi- nant PCR product of the plasmid and genomic DNA was a 750 bp fragment identical in size to the tab promoter fragment between the primer sequences.
Developmental regulation of rabl7 promoter in wild-type and mutant transgenic Arabidopsis seeds
Developmental regulation of rab17/GUS chi- meric gene expression during seed formation was analysed by fluorometric assays and histochemi- cal staining of GUS activity. Figure 2 shows the values of GUS activity measured by fluorometric
Fig. 1. Integration of the rab17/GUS in transgenic Arabidop- sis plants. A. DNA polymerase chain reaction of total genomic DNA from Lal011 and ABA-deficient and ABA-insensitive transgenic mutants generated by crossing with Lal0 l 1. Frac- tionation of PCR products by electrophoresis on 1.5 % agar- ose gels and visualization by staining with ethidium bromide. Lal011 (lane 1), aba (lane 3); abil (lane 4) and abi3 (lane 6). Lane 2 non-transgenic Arabidopsis DNA. Lane 5, plasmid grabl7 DNA. B. Primer sequences of the rabl7 promoter [25] used in the DNA polymerase chain reaction.
pmol 4-MU/min/mg protein 3,000
2.500
2.000
1.500
1.000
500
0 w. t, aba ab i l abi2 abi3
• endosperm []embryo
Fig. 2. GUS enzyme activity in Arabidopsis wild-type and mu- tant seeds. Isolated embryo and endosperm of dry seeds of Lal011 and mutants carrying the same T-DNA insertion with the rabl7/GUS fusion. Values represent the fluorometric analysis of GUS activity in three different experiments.
assays in isolated embryo and endosperm of Lal011 and mutant dry seeds. The highest level of GUS activity was found in the remaining en- dosperm, where no significant differences were detected among the mutants. In embryos, GUS activity was detected in Lal011 and the four mu- tants. However, the levels of GUS activity were lower in the aba mutant than in the wild-type and abi mutants.
The temporal regulation of the rabl7/GUS promoter activity during embryogenesis was analysed by histochemical staining. GUS expres- sion was not detected in developing and mature pollen or in the ovaries of open flowers before or after pollination (results not shown). Immature wild-type embryos remained unstained up to 12 days after flowering (d.a.f.). GUS activity started to accumulate in embryos of 15 d.a.f, and con- tinued to increase rapidly between 16 and 20 d.a.f., reaching a maximum in dry embryos (see Fig. 3). The time course accumulation of GUS expression during embryo development was es- sentially the same in the case of the mutants. The aba and abi-1 (shown in Fig. 4A), abi-2 and abi-3 (not shown) embryos only stained blue during late maturation and in accordance with the fluo- rometric data GUS staining was slightly lower in the aba mutant (see Fig. 2).

565
Fig. 3. Pattern of GUS activity in developing embryos of rabl7/GUS transformed Arabidopsis wild-type plants. Isolated embryos from Lal011 were incubated in X-Gluc at 37 °C for 6 h. GUS staining of embryos. From top left to bottom right: 3 d.a.f.; 6 d.a.f.; 8 d.a.f.; 10 d.a.f.; 14 d.a.f.; 16 d.a.f.; 18 d.a.f.; 20 d.a.f.; dry embryo. Magnification x 40.
Microscopic analysis of GUS expression in seed sections of both transgenic wild-type and mutants (Fig. 4B), revealed that in mature seeds there is a gradual change in the levels of GUS expression primarily detected in endosperm and in the epidermal and subepidermal cells of the embryo.
Regulation of the rabl 7promoter in transgenic seed- lings
In order to investigate the activity of the rabl7 promoter in transgenic plants, GUS activity was analyzed by fluorometry in Lal011 and ABA mu- tants during germination. Relatively weak GUS

566
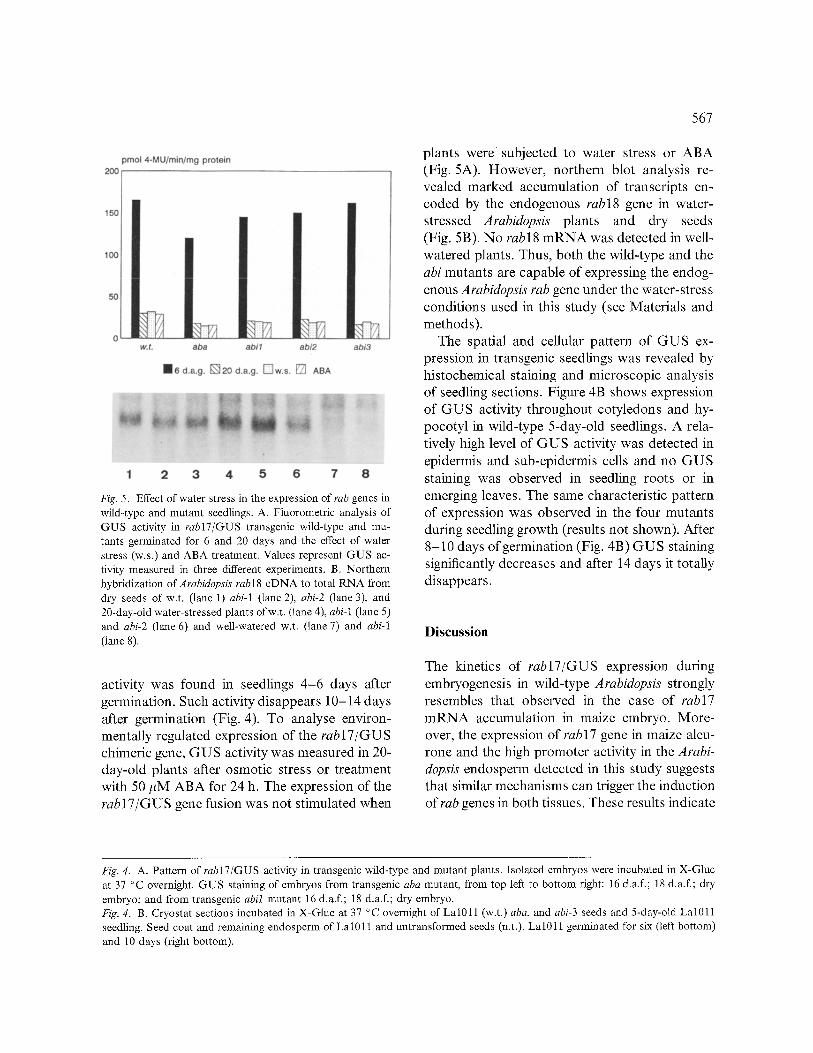
Fig. 5. Effect of water stress in the expression of rab genes in wild-type and mutant seedlings. A. Fluorometric analysis of GUS activity in rabl7/GUS transgenic wild-type and mu- tants germinated for 6 and 20 days and the effect of water stress (w.s.) and ABA treatment. Values represent GUS ac- tivity measured in three different experiments. B. Northern hybridiz ation of A rabidopsis rab 18 cDNA to total RNA from dry seeds of w.t. (lane 1) abi-1 (lane 2), abi-2 (lane 3), and 20-day-old water-stressed plants ofw.t. (lane 4), abi-1 (lane 5) and abi-2 (lane 6) and well-watered w.t. (lane 7) and abi-1 (lane 8).
activity was found in seedlings 4-6 days after germination. Such activity disappears 10-14 days after germination (Fig. 4). To analyse environ- mentally regulated expression of the rabl7/GUS chimeric gene, GUS activity was measured in 20- day-old plants after osmotic stress or treatment with 50 #M ABA for 24 h. The expression of the rabl7/GUS gene fusion was not stimulated when
567
plants were subjected to water stress or ABA (Fig. 5A). However, northern blot analysis re- vealed marked accumulation of transcripts en- coded by the endogenous rabl8 gene in water- stressed Arabidopsis plants and dry seeds (Fig. 5B). No rabl8 m R N A was detected in well- watered plants. Thus, both the wild-type and the abi mutants are capable of expressing the endog- enous Arabidopsis tab gene under the water-stress conditions used in this study (see Materials and methods).
The spatial and cellular pattern of GUS ex- pression in transgenic seedlings was revealed by histochemical staining and microscopic analysis of seedling sections. Figure 4B shows expression of GUS activity throughout cotyledons and hy- pocotyl in wild-type 5-day-old seedlings. A rela- tively high level of GUS activity was detected in epidermis and sub-epidermis cells and no GUS staining was observed in seedling roots or in emerging leaves. The same characteristic pattern of expression was observed in the four mutants during seedling growth (results not shown). After 8-10 days of germination (Fig. 4B) GUS staining significantly decreases and after 14 days it totally disappears.
Discussion
The kinetics of rabl7/GUS expression during embryogenesis in wild-type Arabidopsis strongly resembles that observed in the case of rabl7 m R N A accumulation in maize embryo. More- over, the expression of rabl7 gene in maize aleu- rone and the high promoter activity in the Arabi- dopsis endosperm detected in this study suggests that similar mechanisms can trigger the induction of tab genes in both tissues. These results indicate
Fig. 4. A. Pattern of rab17/GUS activity in transgenic wild-type and mutant plants. Isolated embryos were incubated in X-Gluc at 37 °C overnight. GUS staining of embryos from transgenic aba mutant, from top left to bottom right: 16 d.a.f.; 18 d.a.f.; dry embryo; and from transgenic abil mutant 16 d.a.f.; 18 d.a.f.; dry embryo. Fig. 4. B. Cryostat sections incubated in X-Gluc at 37 °C overnight of Lal011 (w.t.) aba, and abi-3 seeds and 5-day-old Lal011 seedling. Seed coat and remaining endosperm of Lal011 and untransformed seeds (n.t.). Lal011 germinated for six (left bottom) and 10 days (right bottom).

568
that seed-specific transcription factors in Arabi- dopsis are capable of correctly regulating the maize tab 17 promoter. Furthermore, microscopic analy- sis of GUS activity in embryo sections indicated that no strict border between different cell types exists. This suggests that ubiquitous more than cell-specific factors control expression of the chi- meric gene.
It has been reported that various ABA-induced responses are decreased or eliminated in Arabi- dopsis mutants carrying ABA deficiency and in- sensitivity mutations [ 11 ]. Seed dormancy is re- duced in all the mutants. Moreover, a stage specificity to the effects of these mutations has been shown. The abi3 mutation primarily affects seed development, while abil and abi2 exert major effects during vegetative growth [4]. However, the 1.3 kb rabl7 promoter fragment shows the same temporal and spatial regulation on the re- porter gene GUS during embryo maturation in both wild-type and ABA-deficient and ABA- insensitive mutants. Our previous studies have shown that tab mRNAs are expressed during late embryo development of the maize viviparous mu- tants [18, 19]. These mutants are impaired in ABA responses and the viviparous embryos are devoid of desiccation. Taken together, these data seem to indicate that the rabl7 gene is an integral part of the embryo developmental programme and that specific expression of rabl7 gene in late em- bryogenesis does not depend on the presence of high levels of endogenous ABA in the tissues whether in maize or Arabidopsis. Moreover, seed- specific expression of the rabl7 gene seems to be relatively independent of alterations in embryo dormancy or desiccation.
During germination the levels of GUS expres- sion gradually disappear, and neither wild-type or mutant plants express GUS after osmotic stress. It has been reported that several ABA-responsive gene promoters, the rice rabl6B and carrot DC8 promoter, are not active in vegetative organs of transgenic tobacco plants following desiccation or ABA treatment [6, 27]. Our previous results showed that a rabl7/CAT transgene was ex- pressed in transgenic 6-day-old tobacco seedlings following water stress or ABA treatment, al-
though the promoter activity was not studied in adult plants [26]. In maize, water stress and/or ABA induce high levels of rabl7 mRNA in all tissues tested. Moreover, the endogenous rab mRNA in Arabidopsis is induced by water stress both in wild-type and abi mutants. This apparent difference in rabl7 regulation may reflect either that the 1.3 kb promoter fragment lacks specific cis elements required for water stress regulation, or that the monocot cis elements are not recog- nized by the specific transcription factors existing in the vegetative tissues of Arabidopsis, a dicot plant.
The different forms of regulation of the rabl7/ GUS in embryo and vegetative tissues of trans- genic wild-type Arabidopsis and mutants raises some questions on the role of ABA on rab gene activation and strongly suggest that alternative pathways involving different molecular mecha- nisms may be responsible for seed-specific ex- pression and ABA-osmotic stress induction of this gene. Indeed, preliminary evidence of tissue- specific factors involved in the regulation of tab genes has been recently obtained. Distinct regu- latory protein factors with apparently similar af- finity for the ABRE sequence have been detected in embryo and vegetative tissues of maize [20]. In conclusion, the regulation of the rabl7 promoter and its characteristics presented in this study pro- vide a starting point for addressing questions con- cerning the cis elements and protein factors in- volved in the expression of tab genes in embryo and vegetative tissues in both monocot and dicot plants, and for investigating the role of ABA in transcriptional regulation.
Acknowledgements
The authors gratefully acknowledge Dr Maarten Koornneef for supplying the aba and abi mutants, and Dr Tapio Palva for the Arabidopsis rabl8 cDNA clone. Rosa Raz is thanked for excellent technical assistance and Belen Nadal for help in the cryostat sections. This research was sup- ported by Grant BIOT CT 90-0207 from the EC BRIDGE Program to M.P. and J.M.M.Z.

References
1. Bradford MM: A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein dye-binding. Anal Biochem 72: 248-254 (1976).
2. Dellaporta SL, Wood J, Hicks JB: Molecular Biology of Plants: A Laboratory Course Manual, pp. 36-37, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY (1984).
3. Espelund M, Saeboe-Larssed S, Hughes DW, Galau GA, Larsen F, Jakobsen KS: Late embryogenesis-abundant genes encoding proteins with different numbers of hydro- philic repeats are regulated differentially by abscisic acid and osmotic stress. Plant J 2:241-252 (1992).
4. Finkelstein RR, Somerville CR: Three classes of abscisic acid (ABA)-insensitive mutations of Arabidopsis define genes that control overlapping subsets of ABA responses. Plant Physiol 94:1172-1179 (1990).
5. Giraudat J, Hauge BM, Valon C, Smalle J, Parcy F, Goodman HM: Isolation of the Arabidopsis ABI3 gene by positional cloning. Plant Cell 4:1251-1261 (1992).
6. Goupil P, Hatzopoulos P, Franz G, Hempel FD, You R, Sung ZR: Transcriptional regulation of a seed-specific carrot gene DC8. Plant Mol Biol 18:1049-1063 (1992).
7. Guiltinan MJ, Marcotte WR, Quatrano RS: A plant leu- cine zipper protein recognizes an abscisic acid response element. Science 250:267-270 (1990).
8. Haughn G, Somerville CR: Sulfonylurea-resistant mu- tants of Arabidopsis thaliana. Mol Gen Genet 204: 430- 434 (1986).
9. KoornneefM, Jorna ML, Brinkhorst-van der Swan DEC, Karssen CM: The isolation of abscisic acid (ABA)- deficient mutants by selection of induced revertants in non-germinating gibberellin sensitive lines of Arabidopsis [haliana (L.) Heynh. Theor Appl Genet 61:385-393 (1982).
10. Koornneef M, Reuling G, Karssen CM: The isolation and characterization of abscisic acid-insensitive mutants ofArabidopsis thaliana. Physiol Plant 61:377-383 (1984).
11. Koornneef M: Genetic aspects of abscisic acid. In: Blonstein AD, King PJ (eds) Genetic Approach to Plant Biochemistry, pp. 35-54. Springer-Verlag, Vienna (1986).
12. Lang V, Palva ET: The expression of rab-related gene, rab18, is induced by abscisic acid during the cold accli- mation process ofArabidopsis thaliana (L.) Heynh. Plant Mol Biol 20:951-962 (1992).
13. Marcotte WR, Bayley ChC, Quatrano RS: Regulation of a wheat promoter by abscisic acid in rice protoplasts. Nature 335:454-457 (1988).
14. Mundy J, Yamaguchi-Shinozaki K, Chua NH: Nuclear proteins bind conserved elements in the abscisic acid- responsive promoter of a rice RAB gene. Proc Natl Acad Sci USA 87:1406-1410 (1990).
569
15. Nambara E, Naito S, McCourt P: A mutant of Arabidop- sis which is defective in seed development and storage protein accumulation is a new abi3 allele. Plant J 2: 435- 442 (1992).
16. Nordin K, Heinno P, Palva ET: Separate signal pathways regulate the expres sion of a low-temperature-induced gene in Arabidopsis thaliana (L.) Heynh. Plant Mol Biol 16: 1061-1071 (1991).
17. Oeda K, Salinas J, Chua NH: A tobacco bZip transcrip- tion activator (TAF-1) binds to a G-box-like motif con- served in plant genes. EMBO J 10:1793-1802 (1991).
18. Pla M, Goday A, Vilardell J, Gomez J, Pages M: Differ- ential regulation of ABA-induced 23-25 kDa proteins in embryo and vegetative tissues of the viviparous mutants of maize. Plant Mol Biol 13:385-394 (1989).
19. Pla M, Gomez J, Goday A, Pages M: Regulation of the abscisic acid-responsive gene rab28 in maize viviparous mutants. Mol Gen Genet 230:394-400 (1991).
20. Pla M, Vilardell J, Guiltinan MJ, Marcotte WR, Niogret MF, Quatrano RS, Pages M: The cis-regulatory element CCACGTGG is involved in ABA and water-stress re- sponses of the maize gene rab28. Plant Mol Biol 21: 259-266 (1993).
21. Quatrano RS, Guiltinan MJ, Marcotte WR: Regulation ofgene expression by abscisic acid. In: Verma DPS (ed) Control of Plant Gene Expression. Telford Press, Cald- well, NJ, in press (1993).
22. Saiki RK, Scharf S, Faloona F, Mullis KB, Horn GT, Erlich HA, Arnheim N: Enzymatic amplification ofglobin genomic sequences and restriction site analysis for diag- nosis of sickle cell anaemia. Science 230:1350-1354 (1985).
23. Skriver K, Mundy J: Gene expression in response to abscisic acid and osmotic stress. Plant Cell 2:503-512 (1990).
24. Valvekens D, Van Montagu M, Van Lijsebettens M: Agrobacterium tumefaciens-mediated transformation of Arabidopsis thaliana root explants by using kanamycin selection. Proc Natl Acad Sci USA 85:5536-5540 (1988).
25. Vilardell J, Goday A, Freire MA, Torrent M, Martinez MC, Torne JM, Pages M: Gene sequence, developmen- tal expression, and protein phosphorylation of RAB-17 in maize. Plant Mol Biol 14:423-432 (1990).
26. Vilardell J, Mundy J, Stilling B, Leroux B, Pla M, Frey- ssinet G, Pages M: Regulation of the maize rabl7 gene promoter in transgenic heterologous systems. Plant Mol Biol 17:985-993 (1991).
27. Yamaguchi-Shinozaki K, Mino M, Mundy J, Chua NH: Analysis of an ABA-responsive rice gene promoter in transgenic tobacco. Plant Mol Biol 15:905-912 (1990).
28. Zeevaart JAD, Creelman RA: Metabolism and physiol- ogy of abscisic acid. Annu Rev Plant Physiol Plant Mol Biol 39:439-473 (1988).
Copyright © 2022 FDOKUMEN




















