
La cita "atanasiana" del "Apologeticum de tribus capitulis" de Julián de Toledo
Upload
independentCategory
view
1download
0
Hydrobiologia 439: 161–178, 2000.© 2000Kluwer Academic Publishers. Printed in the Netherlands.
161
Redescription and assignment of the chydoridIndialona ganapatiPetkovski, 1966 (Branchiopoda: Anomopoda: Aloninae) toIndialonini, new tribus
Alexey A. KotovA. N. Severtsov Institute of Ecology and Evolution, Leninsky Prospect 33, Moscow 117071, RussiaE-mail: [email protected]
Received 01 August 2000; in revised form 30 August 2000; accepted 01 September 2000
Key words: Cladocera, Anomopoda, Chydoridae,Indialona, morphology, systematics, Indialonini n. trib.,evolutionary relationships
Abstract
The chydoridIndialona ganapatiPetkovski, 1966 is redescribed based on material from the sublittoral planktonof Ajwa Reservoir, Ahmedabad, India, and a neotype is selected. Lateral head pores are recorded, and all thoraciclimbs are studied in detail. Gamogenetic specimens are described for the first time. Evolutionarily relationshipsof Indialona and other Aloninae are discussed. A new tribe Indialonini n. trib. is proposed for the sole genusIndialona, while all other genera of Aloninae, as far as is known, compose the tribe Alonini s. str. A diagnosis ofboth tribes and a key for their discrimination is proposed. Sharp differences between these tribes are found in somecharacters of the thoracic limbs. The limbs of the Indialonini are more oligomerized than those of the Alonini, anda strong oligomerization is characteristic of limbs I–II.
Abbreviations:IDL – inner distal lobe of limb I; ODL – outer distal lobe of limb I
Introduction
Based on the material from India, Petkovski (1966)described a remarkable, minute chydorid, assigned byhim to a new genus,Indialona. The generic diagnosiswas short and, though clearly defining this animal,lacked information on many important characters:
“Kleine Chydoride der Sub-fam. Aloninae Freymit Schalenmerkmalen der Gattung Alonella,deren Kopfschild aber Alona-artig erscheint.Postabdomen sehr eigentümlich, spezifisch fürdie Gattung gestaltet: am Distalende einfachverjüngt, Hinterrand distal ohne Bildung einerEcke oder Rundung. Bewehrung des Postabdo-mens ebenso sehr charakteristisch, obwohl zartund spärlich; aus 4 bis 5 Gruppen von jewenigen Börstchen bestehend. Mediane Kop-fporengarnitur aus einer einzigen Pore gebil-det. Mandibeln sind im Verbindungswinkel
des Kopfschildes und der Schale befestigt”(Petkovski, 1966: 158).
Smirnov (1971) expandedIndialonaby transferring toit three species described earlier by Daday as membersof other genera:Alona globulosaDaday, 1898,AlonamacronyxDaday, 1898 andEuryalona annandaleiDaday, 1908. Subsequently, the artificial status ofIndialona sensuSmirnov (1971) was demonstrated:Rajapaksa & Fernando (1985) returnedI. macronyxto the genusAlona’ and I. globulosabecame the typespecies of a new genus,Notoalona (Rajapaksa &Fernando, 1987b). As withEuryalona annandalei, itis currently not possible to elucidate its taxonomicalstatus, including its generic position. In the first de-scription of this species by Daday (1908), informationon many important characters is minimal. Particularly,the head pores and thoracic limbs ofE. annandaleiwere not described, which are currently the main cri-teria for including an animal into a given genus of the

162
Anomopoda. Unfortunately, material on this speciesis completely absent in the collection of Daday (Forró& Frey, 1982). Thus, the genusIndialona is to beregarded as monotypical for the time being.
After Petkovski (1966), only one partial redescrip-tion of the morphology ofI. ganapatiwas published(Rajapaksa & Fernando, 1987b). However, many im-portant details of this tiny animal remained unknown,and the place ofIndialonawithin Aloninae continuedto be unclear. No gamogenetic specimens were seen.It is necessary to say that besides the fact that themandibles articulate with the integument between thehead shield and the valves, there was no other proof ofits belonging to the subfamily Aloninae. In addition,some taxonomical problems are connected with thisanimal: the author did not establish types, and did notspecify a type locality (he had material from severalreservoirs in the vicinity of Ahmedabad).
The presence of ample material ofI. ganapatiinthe collection of Prof. Smirnov gave me a chance tore-study this interesting animal. This series of sampleswas collected in Ajwa reservoir and sent to Prof.Smirnov by Dr Ganapati, in honour of whom this spe-cies was named. Petkovski (1966) did not mark a typelocality, but Ajwa reservoir is one of the water bodiesfrom his list of studied material.
My investigation of the morphology ofIndialonaand a comparison with other genera of Aloninae hasresulted in the necessity to create a special tribe, Indi-alinini n. trib. for the genus as opposed to tribe Aloninis.str., including all other known Aloninae.
Materials and methods
Animals were selected from samples under a binocularstereoscopic microscope, placed on slides (in a dropof a glycerol-formaldehyde mixture) and studied un-der an optical microscopein toto. Twenty five adultfemales, one juvenile male I, one juvenile male IIand three adult males were dissected for analysis ofappendages, 15 specimens were lyophilised, mountedon an aluminium stub, coated with gold and examinedunder a scanning electron microscope (JEOL-840A).
Different systems of enumeration of setae on thelimbs of Chydoridae exist (Smirnov, 1971; Chiam-beng & Dumont, 1999). In the present article, I usea simple system of numbers consistent with my pre-vious article (Kotov, 2000): all marginal setae of (1)exopodites, (2) inner limb portions (endopodites), and(3) distal armature of gnathobases numbered from
distalmost to basalmost elements without attempts todetermine homologies between different limbs. Setaeof the inner portion of limb I are marked by letters,a–h.
Results
Family Chydoridae Stebbing, 1902 emend. Du-mont & Silva-Briano, 1998Subfamily Aloninae Dybowski & Grochowski,1894 emend. Frey, 1967
1. Tribe Alonini s. str.
DiagnosisBody of varying shape. Setae at ventral margin ofvalves forming a continuous row. Spine on basalsegment of antennal exopod present at least in rudi-mentary state. Limb I and II with numerous setae: 7–9anterior setae on limb I, 8 setae on corm of limb II,5–8 setae in filter plate II. Many marginal setae oncorm of limb III. There are from 6 to 4 well developed,plus one rudimentary, seta on exopodite IV. Normally4 (very rarely 3) setae on exopodite V. Limb VI presentor reduced. In males, 3–4 setae on IDL, gonoporesopen at distal end of postabdomen.
Littoral animals, only a few species facultativelypenetrate the plankton.
Type genusAlona Baird, 1843, with type speciesA. quadrangularis(O. F. Müller, 1785).
2. Tribe Indialonini n. trib.
DiagnosisBody of females are rounded and very high. Row ofsetae at ventral margin with gap in medium portionof valve. Spine on basal segment of antennal exopodabsent. Limbs I and II with greatly reduced numberof setae: only three setae on corm of limb I, only sixsetae on corm of limb II, four setae in filter plate II, oneof them rudimentary. A few (three) marginal setae oncorm of limb III. Only four setae on exopodite IV, onlythree setae on exopodite V. Limb VI absent. In males,only two setae on IDL, gonopores open far from distalend of postabdomen.
Obligate sublittoral (and, perhaps, pelagic) plank-ters.
Type (sole) genusIndialonaPetkovski, 1966 withone species,I. ganapatiPetkovski, 1966.

163
GenusIndialonaPetkovski, 1966
New diagnosisAdult female: Body high, broadly rounded.
Dorsal margin regularly curved, postero-dorsal anglesmoothed, posterior margin straight, falling downsharply, postero-dorsal angle broadly rounded, ventralmargin weakly concave. No dorsal keel on head or restof body. Head shield ovoid, somewhat elongated, withsingle major head pore, and minute lateral head pores.
Labral keel broadly expanded, with well expressedapex, without denticles or setulation. A row of setaealong ventral rim of valves with a gap in the medianzone. Postabdomen of characteristic form stronglytapering distally, an acute-angled triangle in lateralview. No marginal postanal denticles, only five seriesof relatively long setules at postanal margin, analogousseries of setules laterally on surface of postabdomen.Postabdominal claw relatively long, with long, robustbasal spine and a group of 3–4 long setules on clawbasally to this spine.
Antennule short, not reaching tip of rostrum,without setulation. Slender antennular sensory setaat distance of 1/4 of antennular length from apex,markedly projecting beyond antennular tip. Secondantenna with short body, but with long swimmingsetae. Setae 0-0-3/1-1-3, spines 0-0-1/0-0-1. Apicalspines rudimentary; spine from basal segment of ex-opod absent. Five pairs of trunk limbs. Trunk limb Iwithout external accessory seta, its ODL with one longseta, IDL with two bisegmented setae of somewhat un-equal size. Endites on limb corm not expressed. Onlythree bisegmented setae of similar size on inner faceof limb corm, maxillar process absent, ejector hooksvery large. Trunk limb II with only six setae on limbcorm, deep incision delimits inner limb portion fromgnathobase. Only four setae in filter comb II, distal-most seta very short. Five setae in exopodite III. Onlythree marginal setae and three soft setae on inner por-tion of limb III, five setae in filter comn III. ExopoditeIV large, subovoid, with four setae, four marginal andthree soft setae on inner limb portion, filter comb IVwith four setae. Only three setae in exopodite V, filterplate V absent.
Ephippial female:This is larger than parthenogen-etic one, dorsal part of its valves prominent laterally.Entire carapace used as an ephippium.
Adult male: This has an elongated body, largehead, straight or slightly concave ventral margin ofvalves, and angle in antero-medial portion of vent-ral margin. Postabdomen generally similar to that in
female, gonopores open far from its distal end. Anten-nules large, with nine esthetascs, and additional maleseta subdistally. IDL of male with two greatly unequalnaked setae, ODL with one seta.
Very small with a size up to 305µm in partheno-genetic females, up to 315µm in ephippial females,up to 280µm in males.
One species,I. ganapatiPetkovski, 1966.
Indialona ganapati Petkovski, 1966
Petkovski, 1966: 157–165, Abb. 1–9; Smirnov, 1971:502, Fig. 640 (after Petkovski, 1966); Rajapaksa &Fernando, 1987b: 151, Figs 126–136.
Neotype: An adult parthenogenetic female withsingle egg in 4% formaldehyde with addition of 5%of glycerol, deposited to the collection of ZoologicalMuseum of Moscow State University, MGU-Ml-18,specimen selected from sample NNS-1998-175.
Petkovski (1966) did not specify a type and did notmark a type locality.
Neotype locality: Ajwa Reservoir, vicinity ofAhmedabad, India.
Material examined:Twelve ample samples, more than 100 specimens ofIndialona in each, all collected by Dr S. V. Ganapatiin Ajwa Reservoir on: (1) 23.04.1963 (NNS-1998-168); (2–3) 21.06.1963 (NNS-1998-169 & NNS-1998-170); (4) 19.07.1963 (NNS-1998-171); (5)29.08.1963 (NNS-1998-172); (6) 30.08.1963 (NNS-1998-173); (7) 20.12.1963 (NNS-1998-174); (8)28.04.1964 (NNS-2000-036); (9) 11.06.1964 (NNS-1998-175); (10) 28.08.1964 (NNS-1998-167); (11)25.09.1964 (NNS-1998-176); (12) Unknown year,sample marked by S. V. Ganapati as ‘a16 21/12’(NNS-1998-177).
All samples are from the collection of Prof. N. N.Smirnov, now at the Zoological Museum of MoscowState University, but not officially deposited to it.
DiagnosisAs for the genus.
Redescription
Parthenogenetic female
General: Minute animal. In lateral view bodyelongated in juveniles and very high, broadly roundedin large adults (Figs 1–2, 8 and 12–14) (height/ length

164
Figures 1–11. Indialona ganapati, females from Ajwa Reservoir, vicinity of Ahmedabad, India, collected 11.06.1964 by Dr S. V. Ganapati.Figures 1–2. A large adult female in lateral and anterior view;Figures 3–5. Its head, head shield and labrum;Figure 6. Ventral margin of valve;Figure 7. Postabdomen in lateral view;Figures 8–9. Juvenile female and its postabdomen in lateral view;Figures 10–11. Ephippial female inlateral and anterior view. Scale bar denotes 100µm.

165
= 0.57–0.65 in juveniles, 0.68–0.75 in large adults),maximum height of body in its medium portion.Dorsal margin regularly curved, depression betweenhead and rest of body weakly expressed or absent.Postero-dorsal angle well expressed in juveniles, witha tendency to smoothing in adults. Posterior marginstraight, falling down sharply (frequently vertically),postero-dorsal angle broadly rounded. Ventral marginweakly concave.
In anterior view (Figs 2 and 15) the body ovoid,slightly compressed laterally, the maximum width ofthe body is in the mid-dorsal portion of valves. Nodorsal keel on head or rest of body. In ventral view,the body subovoid, posterior extremity acute-angled,maximum width of body posteriorly to fornices (Figs16 and 17). Parthenogenetic females carry 1–2 eggs.
Head: The head is very large in juveniles and re-latively small in adults, in both triangular-round inlateral view (Figs 1, 3, 8 and 18–20). Ventral margin ofhead is straight, dorsal margin is evenly arched. Eye islarger than ocellus. Rostrum well developed, pointingdownward. In juveniles the distance from the tip ofthe rostrum to the ocellus is markedly slightly smallerthan that between the ocellus and eye, but in adults thisdistance is approximately equal.
Head shield (Fig. 4) is ovoid, somewhat elong-ated, with the length being about 1.4–1.5 times itsmaximum width. Anterior (rostral) extremity is roun-ded and somewhat anteriorly to the middle, there areweakly developed, not projecting fornices: as a result,the maximum width of the head shield is at a level ofmandibular articulation. Lateral surface slightly con-cave anteriorly to fornix. Posterior extremity rounded.
Sole major head pore is an opening surrounded bya wide, thickened rim (Fig. 21). Minute lateral headpores are located at a level with the central majorhead pore or somewhat anteriorly, at about 0.25–0.35postpore distance.
Labrum: The labrum is large (Figs 3, 5 and 24).Distal labral plate is without setulation. Labral keel isbroadly expanded, with expressed apex ventrally, two–three hillocks in region of apex. Anterior margin ofkeel convex, without denticles or setulation, posteriormargin short, generally straight. No special lateral pro-jections on labrum and no special folds surrounding itsbase.
Valves:The valves have thin walls, generally roun-ded. Their height depends on extension of an infla-tion of brood pouch. Specific reticulation expressedin antero-ventral portion (Fig. 22). Other surface ofvalves smoothed.
In contrast to other Chydoridae, the row of setaealong ventral rim of valves not continuous (Figs 1,6 and 25). In the antero-ventral portion of valves,there are about 12–15 setae, the middle setae of thisgroup the longest. Medium portion of ventral mar-gin free of setae, a row of conical hillocks continuesthe row of anterior setae, the distances between theformer increasing posteriad. A small (4–5) series ofsetae increasing in size posteriad in posterior zone ofventral margin (Fig. 6). Row of setae followed pos-teriorly by a dense row of setules projecting beyondmargin of valve (Fig. 26). Posterior margin of valveswith fine submarginal setules, not projecting beyondmargin (Fig. 27).
Thorax: The thorax is without external traces ofsegmentation, with five limb pairs.Abdomenis short,with transverse rows of setules.
Postabdomen:The postabcomen is of character-istic form, strongly tapering distally, an acute-angledtriangle in lateral view (Figs 7 and 9). Preanal part ofdorsal margin straight or slightly convex. Anal mar-gin concave, postanal part weakly convex. Its preanaland postanal angles are not prominent, although quitedistinguishable. Dorso-distal angle absent. Base ofpostabdominal claws not very massive, ventral marginpractically straight.
No marginal postanal denticles, only five series ofrelatively long setules at postanal margin (Figs 28–30).An analogous series of setules laterally on postabdo-men surface, their size smaller. Bunch of robust setulesnear claw base. Anal portion with three rows of muchshorter setules.
Abdominal setae are on a special base, their lengthexceeding that of preanal part of postabdomen.
Postabdominal claw:This is relatively long, butrobust; basally massive; distally weakly and evenlytapered and curved. Dorsal margin of claw with nu-merous long setules of similar size. A row of setulesalong ventral margin.
A single, long, robust, almost straight basal spinelocated on claw at an angle of about 30◦ to it at somedistance from its base. A group of 3–4 long setules onclaw basally to this spine.
Antennule: This is relatively short and robust,tapering distally, not reaching tip of rostrum, withoutsetulation (Figs 3 and 34). Slender antennular sens-ory seta arises at 1/4 of antennular tip and markedlyprojects beyond it. Nine esthetascs of unequal lengtharise from a cuplike indentation at tip of antennule; noscallops or teeth around indentation. All esthetascs are
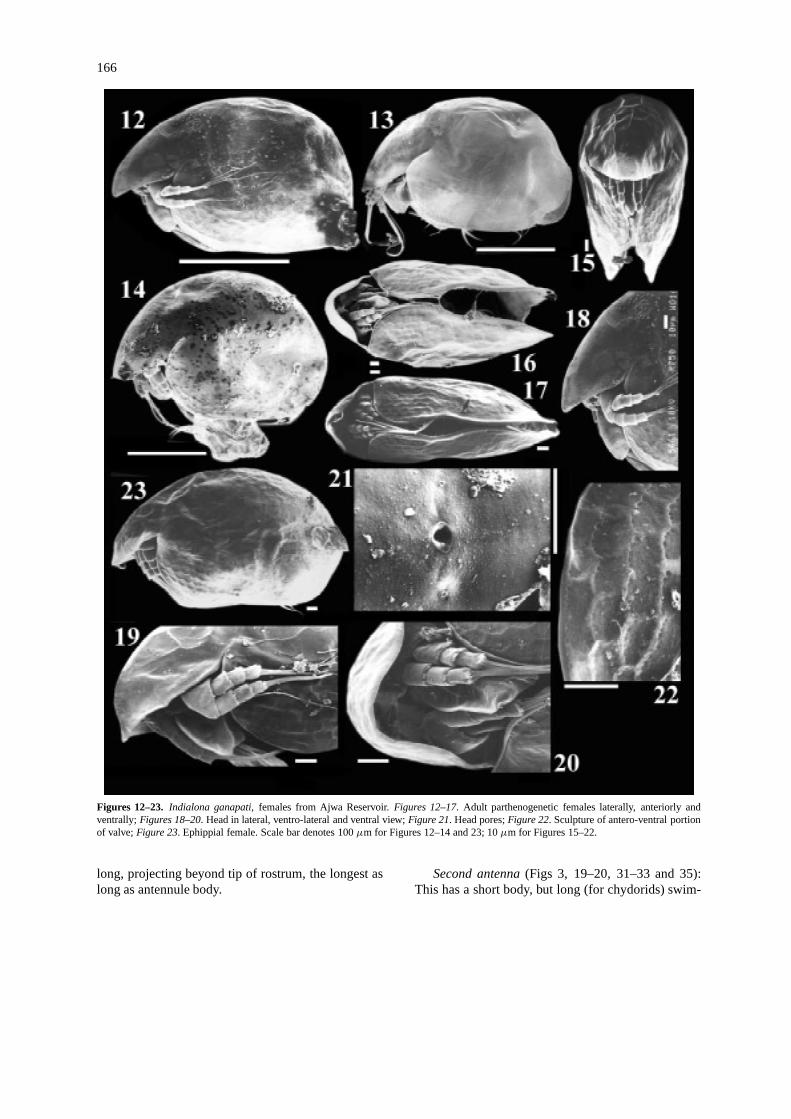
166
Figures 12–23. Indialona ganapati, females from Ajwa Reservoir.Figures 12–17. Adult parthenogenetic females laterally, anteriorly andventrally;Figures 18–20. Head in lateral, ventro-lateral and ventral view;Figure 21. Head pores;Figure 22. Sculpture of antero-ventral portionof valve;Figure 23. Ephippial female. Scale bar denotes 100µm for Figures 12–14 and 23; 10µm for Figures 15–22.
long, projecting beyond tip of rostrum, the longest aslong as antennule body.
Second antenna(Figs 3, 19–20, 31–33 and 35):This has a short body, but long (for chydorids) swim-

167
Figures 24–33. Indialona ganapati, females from Ajwa Reservoir.Figure 24. Labral keel in lateral view;Figures 25–27. Medium part ofventral margin, postero-ventral portion, and posterior margin of valve in inner view;Figures 28–30. Distal part of postabdomen in lateral,dorsal and latero-distal view;Figures 31–33. Second antenna laterally, postero-laterally and vetro-laterally. Scale bar denotes 10µm forFigures 24–26 and 28–33; 1µm for Figure 27.

168
ming setae, this feature specially characteristic injuveniles, where swimming setae almost reach pos-terior margin of valves. Coxal part with two relativelylong sensory setae. Basal segment distally with rudi-mentary setae on anterior surface (Figs 31–33), and arow of long, stout setules on ventral surface. Brancheswith robust segments, all segments cylindrical, withminute setules around distal margin.
Setae 0-0-3/1-1-3; apical members long, not setu-lated. Lateral seta from second endopod segment ofthe same size with them, seta from the first segmentshort, only slightly projecting bejond tip of endopod.Spines 0-0-1/0-0-1. Apical spines rudimentary; spinefrom basal segment of exopod absent, only a series ofparticularly large setules in this region (Figs 32–33).
Mandibles:These are short, thick, massive, articu-late at junction of head shield and carapace.
Trunk limbs:five pairs.Trunk limb I: large, with greatly reduced armature
for a chydorid (Fig. 36). Epipodite small, globular,without a projection. External accessory seta absent.ODL a small, cylindrical lobe with one long setaarmed with short setules distally. IDL large lobe withtwo series of setules, and only two bisegmented setaeof unequal size, both armed by relatively sparselydistributed setules.
Endites not individualized. Only three bisegmen-ted setae (Fig. 36 a–c), of similar size, on inner limbportion, all armed by long hairs, a small hillock nearmedium member. Two long ejector hooks of somewhatunequal size subdistally on outer portion of limb corm.A complicated system of series of long setules on pos-terior surface of limb. Maxillar process (=gnathobaseI) absent.
Limb II: Epipodite small, globular. Exopoditeprobably absent. Only six setae on ventral margin oflimb (Fig. 37: 1–6), status of distalmost members un-clear. Three distal elements feathered bilaterally withthin setules. Three basalmost elements armed unilat-erally with long, robust setules, thickening of whichincreasing basad. A deep incision delimits inner limbportion from gnathobase.
Distal armature of gnathobase with four elements(Fig. 37: 1–4). Only four setae in filter plate. Distal-most seta very short, second member is smaller thanthe other two.
Limb III: Pre-epipodite not found. Larg epipoditeovoid, but without a projection. Exopodite flat, relat-ively small. Distally, three long setae of unequal size(Fig. 38: 1–3), armed with long setules; laterally, twosetae of unequal size and similar armature (4–5).
Subdividing of inner limb portion into basal anddistal endites unclear. Only three marginal setae on in-ner limb portion (1–3), two distal members armed withlong setules, with long sensillum near second seta (thisis typical position for sensillum in all chydorids, seeDumont & Silva-Briano, 1997), definitely belongingto former distal endite. Status of third, rudimentary,seta unclear. Three relatively long soft seta of similarsize posteriorly on inner limb portion.
Distal armature of gnathobase III with four ele-ments (Fig. 39: 1–4), one of them a small, bottle-shaped sensillum located far from the others. Only fivesetae of similar size in filter comb.
Limb IV: Pre-epipodite a setulated hillock. Epipod-ite large, ovoid. Exopodite large, subovoid, with foursetae, size of which decreasing distad, and bunch oflong setules distally (Fig. 40: 1–4).
Marginally on the inner limb face, a row of foursetae unilaterally armed with long hairs (1–4). Distal-most member conical, each of others with inflatedbasal segment, and thin distal part. A small sensillumnear third seta. Three soft setae with size increasingbasad.
Distal armature of gnathobase with four elements:a globular sensillum on the border with inner limb por-tion, long setulated setae and a hook distally, and asmall receptor on anterior face on gnathobase. Filtercomb with four setae of similar size.
Limb V:Pre-epipodite and epipodite as in limb IV.Exopodite relatively large, size similar to that in limbIV, with three setulated setae, their size decreasing dis-tad (Fig. 41: 1–3). Distal part of inner limb portionprotruding as a very large (larger than exopodite) flap-like projection, fringed by long setules. Two robustsubmarginal setae on inner face of limb. A hillock inposition of distal armature of gnathobase.
Limb VI: absent.
Ephippial female and ephippiumEphippial females larger than parthenogenetic ones. Inlateral view, body somewhat deeper as compared withparthenogenetic female, postero-dorsal angle alwaysprominent. Posterior half of dorsal margin heavilychitinized (Figs 10–11 and 23). Dorsal part of car-apace slightly more pigmented, and with polygonalcells. In ventral view, dorsal part of valves, region ofresting egg locule, prominent laterally. Entire carapaceof female used as an ephippium. A single resting egg.Foamy material around the egg occupies part of theephippial chamber.

169
Figures 34–41.Head and thoracic appendages of females ofIndialona ganapatifrom Ajwa Reservoir.Figures 34–35. First and secondantennae;Figure 36. Limb I in anterior view;Figure 37. Limb II in posterior view; Figures 38–39. Limb III in anterior view, and distalarmature of its gnathobase in posterior view;Figures 40–41. Limbs IV and V. Scale bar denotes 10µm.
MalesTwo juvenile instars, quite typical of all chydorids,were detected inIndialonamales. Juvenile males I ha-bitually similar with females of the same instar (Fig.42). Males II retain their juvenile form, in contrastto more higher and rounder females II, dorsal mar-
gin of the latter distinctly arched (Fig. 46). Maturemales (Fig. 50) markedly differ from females in (1)more elongated, lower body, (2) higher head withsharply pointed tip of rostrum, (3) straight or evenslightly concave dorsal margin of valves, the part thatis inflated in female as brood pouch, (4) expression

170
of special angle in antero-medial portion of ventralvalve margin, a smaller number of setae on ventralmargin, (5) antennules slightly protruding behind tipof rostrum, and (6) more powerful second antennaewith longer swimming setae. The majority of thesetraits are connected with a preservation of juvenilecharacters.
Shape and armature of postabdomen is generallysimilar to that of the female, but postabdomen ofthe adult female is somewhat shorter (Figs 43, 47and 51. Only in adult males, gonopores are visible,they open far from the distal end of the postabdomen,sub-ventrally. Postabdominal setae longer than that infemale, reaching postanal edge of anus. Postabdom-inal claw slightly shorter than in female, denticles onits dorsal side seem to be somewhat more robust, inthe bunch of setules near the basal spine the memberclosest to this spine particularly thick.
Antennules of male I as in female I (Fig. 44), thosein male II about the same relative shape and size asin female, but with short, thin additional seta sub-distally (Fig. 48). In adult males, this seta is longer,as long as sensory seta, and thicker. Antennule withnine esthetascs and no setules (Fig. 52).
In males, trunk limb I is not strongly modifiedin comparison with the female, except for the pres-ence of a clasper hook, located between the ODL andIDL (Figs 45, 49 and 53–54). In males I and II, itis slightly bent and relatively short, in male III thishook is large, slightly and evenly tapering distally,U-shaped with a wide gap, and supplied with twotransverse rows of setules subdistally. There is a bunchof setules and a hillock on the limb corm near the IDLin males II. In adult males, a copulatory brush of ro-bust denticles with special short setae having inflatedbase are present in this region of the limb corm, whichis strongly inflated. No additional male seta on IDL ofjuvenile males I and II, and adult male. IDL of malesI and II thick, relatively short, with two unequal setae,as in females. In adult males, the longest seta of IDLare much more elongated than in females, markedlylonger than ODL seta, its setules reduced as well ason the second, short seta. Ejector hooks in adult malesshorter than that in females, not reaching distal end oflimb corm.
SizeNeotype 280µm, females 155–305µm (70), juvenilefemales I 155–180µm (10), minimal size of repro-duction 205µm. Ephippial females 275–315µm (10).
Instar I males 178–195µm (5), males II 210–240µm(3), adult males 245–280µm (10).
Note on biology
Petkovski (1966) stated that he foundIndialona inplanktonic samples. Indeed, at least the majority, orall, of Ganapati’s samples studied by myself containplankton, without fragments of vegetation or mud. Un-fortunately, labels do not contain information on theplaces of collection. There were two dominant speciesof algae in these samples: the typical plankterPedi-astrumsp. (Chlorococcales) andSurirellasp. (Bacilla-ryophyta), the latter more characteristic for the littoralof lakes. These samples were, therefore, probably col-lected in the sublittoral.Indialona ganapatiappears tobe a sublittoral (or pelagic?) plankter that occurs allyear round. This mode of life is not characteristic ofmany chydorids.
Discussion
Comparative analysis of Indialonini and Alonini
Due to the fact that the new tribe includes only asingle species, it is not easy to subdivide its diagnosticfeatures into species, generic and tribe rank. Thepostabdomen of adultIndialona(long, conically taper-ing, without expressed dorso-distal angle) is moresimilar to that of aMoina (Moinidae) orPseudomoina(Macrothricidae), than to the Aloninae. A superficiallysimilar postabdomen, however, with expressed dorso-distal angle, is also found in some Chydorinae, i.e.Picripleuroxus(Smirnov, 1996). I think that this isa character of generic rank, since the postabdomenis different in different genera of Aloninae. AlthoughPetkovski (1966) stated that the presence of groups ofsetules instead of marginal denticles is an importantdiagnostic feature ofIndialona, the same armature ofthe postanal margin of postabdomen is characteristicof some other genera:Celsinotum(Frey, 1991),Le-beris (Smirnov, 1989; Frey, 1998) and someAlona(Smirnov, 1971; Frey, 1991; Sinev & Kotov, 2000),while other species ofAlona have normal denticles(Smirnov, 1971; Alonso, 1996). In juveniles ofIndia-lona, the postabdomen is shorter, robust and similarin external appearance with that inMonospilus(seeSmirnov, 1971: Fig. 647; Alonso, 1996: Fig. 172H).To my mind, a postabdomen ofMonospilus-type may

171
Figures 42–54. Indialona ganapati, males from Ajwa Reservoir, vicinity of Ahmedabad, India, collected 11.06.1964 by Dr S. V. Ganapati.Figures 42–45. Juvenile male I, its postabdomen, antennule and limb I;Figures 46–49. Juvenile male II, its postabdomen, antennule and limbI; Figures 50–54. Adult male, its postabdomen, antennule and limb I in inner and lateral view. Scale bar denotes 100µm for Figures 42, 46 and50; 10µm for others.

172
have been characteristic of the hypothetic ancestor ofthe Indialonini, its elongation having occurred in thecourse of the evolution ofIndialonaafter its separationfrom the main branch of alonine-like anomopods.
The interrupted row of setae along the ventral valvemargin is, probably, a good diagnostic character of theIndialonini. This transformation probably took placeafter the separation of tribe Indialonini. Note that inmany Aloninae, the valve setae are shortest in the me-dium part of the ventral margin. So a complete loss ofthese setae is the final step of their reduction.
The absence of a spine on the basal segment of an-tennal exopod in Indialonini is a good trait for the sep-arating Indialonini from Alonini, where this spine var-ies from rudimentary to quite developed. InGraptole-beris (Alonso, 1996), and in someAlona, i.e. A. lab-rosa (Sinev & Kotov, 2000), this spine is very small.Its size is an important systematic feature, not onlyfor Chydoridae, but also for Ilyocryptidae (Štifter,1991; Kotov, 1999) and Macrothricidae (Silva-Briano,1998).
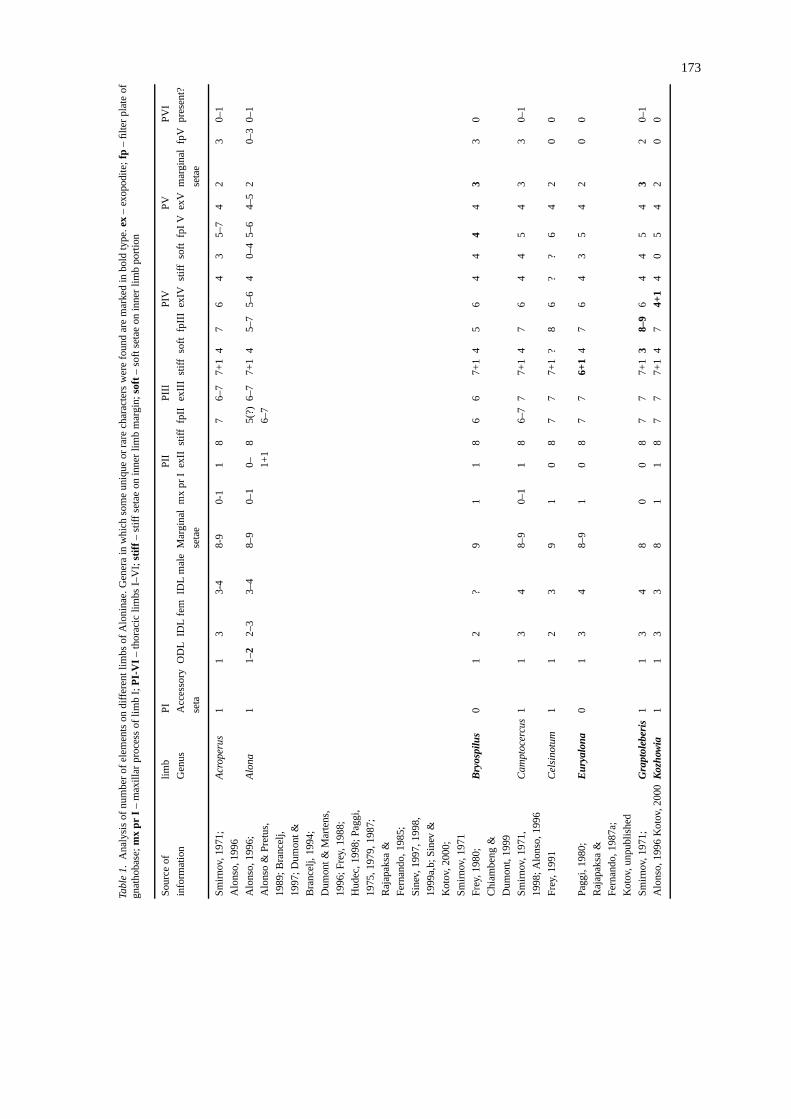
The sharpest differences between Indialonini andAlonini lay in the thoracic limbs. All limbs of theformer are more oligomerized, and especially strongoligomerization is characteristic for limbs I–II of theIndialonini.
Indialonais unique in having only two setae on theIDL of the male limb I. In males of all Alonini, the IDLI bears 3–4 setae (see Table 1).
In Alonini (see pictures of limbs ofTretocephalaambiguaas an example of a representative of thistribe, Figs 55–63) there are three endites on the cormof limb I, each endite bears 2–3 large posterior setae(Fig. 57 a–i). In addition, on each endite there is 0–1 anterior seta (1–3), usually small, and frequentlya single sensillum is located near each of them (thelatter was not discussed by Dumont & Silva-Briano,1997, because they studiedAlonaandCamptrocercuswhich have reduced sensillae on limb I). Thus, thereare maximally nine posterior and three anterior setaeon the corm of limb I (all of them are developed inTretocephalain at least a rudimentary state). In manyAlonini, this number is more or less reduced. In mostAlona the majority or all anterior setae are absent,and present only in few species, such asA. affinis, A.phreaticaor A. labrosa(Alonso, 1996; Sinev & Kotov,2000). In some other Alonini, the number of posteriorsetae is reduced, by an absence of the basalmost setaon endite I (i in Figs 55 and 57, 10th seta in terms ofChiambeng & Dumont, 1999). There are only seven ofthese setae on the corm of limb I inNotoalona globu-
losadue to the absence of a seta on the second endite,while all nine setae are present inN. freyi (Rajapaksa& Fernando, 1987b). In Indialonini, it is not possibleto distinguish endites on limb I, and they have onlythree setae (I think these are members of the posteriorrow, the anterior elements are absent). Thus, two tribesare definitely separated in this character. Among theAnomopoda, the first limb ofIndialona is maximallysimilar to that inBosminopsis(Kotov, 1997), in whichthere are also three setae on the limb corm, and it isnot possible to distinguish endites. However, the firstlimb of the Bosminidae sharply differs from that inChydoridae (includingIndialona): in the former, theIDL is greatly reduced in females, and developed onlyin males, while an additional seta is present in femalesof Bosminidae, homologous to the seta of the copulat-ory brush of chydorid males, and absent in females.In addition, the ODL bears two long setae in Bos-minidae. Therefore, the similarity between limb I inIndialonaand Bosminopsis is a result of independentevolution. Moreover, in the second genus of the Bos-minidae,Bosmina, there are seven setae on corm oflimb I (Kotov, 1997).
The second limb of the Indialonini is also stronglysimplified. Exopodite II is absent, and there are onlysix setae on limb corm in contrast to the eight scrapersof the Alonini (Fig. 58: 1–8). Limb II inIndialona issimilar toBosminopsistoo (see Kotov, 1997, Fig. 1C)!In Alonini, there are hillocks and rudimentary setae –a remainder of the second, posterior, line of elements.In Indialona, they are absent. Finally, in the Alonini,the filter comb consists of 5–8 setae, in contrast to four(with small distalmost elements) inIndialona.
There are only five setae on exopodite III in In-dialona. Exopodite III bears 6–7 setae in the majorityof Alonini (Fig. 59: 1–7), but there are only 3–5 inLeydigia, and four inSpinalona, even less than inIn-dialona. A more important diagnostic feature of thethird limb of Indialonini is the presence of only threemarginal posterior setae on the limb corm. In Alonini,this number is markedly higher, with a maximum ofeight elements. The eighth element is a sensillum inthe majority of genera (Figs 59 and 60: 1–7), butin Nicsmirnovius, one is a true seta (Chiambeng &Dumont, 1999). The minimal number of posteriorelements in Alonini is not clear due to an absenceof information onNotoalona globulosa(Rajapaksa &Fernando, 1987b), in which this number is small, butnot precisely known.
Thoracic limbs IV–V in Indialona are oligomer-ized too, but the degree of oligomerization is less than
173
Table
1.A
naly
sis
ofnu
mbe
rof
elem
ents
ondi
ffere
ntlim
bsof
Alo
nina
e.G
ener
ain
whi
chso
me
uniq
ueor
rare
char
acte
rsw
ere
foun
dar
em
arke
din
bold
type
.ex
–ex
opod
ite;fp
–fil
ter
plat
eof
gnat
hoba
se;m
xpr
I–
max
illar
proc
ess
oflim
bI;P
I-V
I–
thor
acic
limbs
I–V
I;stif
f–
stiff
seta
eon
inne
rlim
bm
argi
n;sof
t–so
ftse
tae
onin
ner
limb
port
ion
Sou
rce
oflim
bP
IP
IIP
IIIP
IVP
VP
VI
info
rmat
ion
Gen
usA
cces
sory
OD
LID
Lfe
mID
Lm
ale
Mar
gina
lm
xpr
Iex
IIst
ifffp
IIex
IIIst
iffso
ftfp
IIIex
IVst
iffso
ftfp
IVex
Vm
argi
nal
fpV
pres
ent?
seta
seta
ese
tae
Sm
irnov
,197
1;A
croperu
s1
13
3-4
8-9
0-1
18
76–
77+
14
76
43
5–7
42
30–
1
Alo
nso,
1996
Alo
nso,
1996
;A
lona
11–
22–
33–
48–
90–
10–
85(
?)6–
77+
14
5–7
5–6
40–
45–
64–
52
0–3
0–1
Alo
nso
&P
retu
s,1+
16–
7
1989
;Bra
ncel
j,
1997
;Dum
ont&
Bra
ncel
j,19
94;
Dum
ont&
Mar
tens
,
1996
;Fre
y,19
88;
Hud
ec,1
998;
Pag
gi,
1975
,197
9,19
87;
Raj
apak
sa&
Fer
nand
o,19
85;
Sin
ev,1
997,
1998
,
1999
a,b;
Sin
ev&
Kot
ov,2
000;
Sm
irnov
,197
1
Fre
y,19
80;
Bry
ospi
lus
01
2?
91
18
66
7+1
45
64
44
43
30
Chi
ambe
ng&
Dum
ont,
1999
Sm
irnov
,197
1,C
am
pto
cerc
us1
13
48–
90–
11
86–
77
7+1
47
64
45
43
30–
1
1998
;Alo
nso,
1996
Fre
y,19
91C
els
inotu
m1
12
39
10
87
77+
1?
86
??
64
20
0
Pag
gi,1
980;
Eur
yalo
na0
13
48–
91
08
77
6+1
47
64
35
42
00
Raj
apak
sa&
Fer
nand
o,19
87a;
Kot
ov,u
npub
lishe
d
Sm
irnov
,197
1;G
rapt
oleb
eris
11
34
80
08
77
7+13
8–9
64
45
43
20–
1
Alo
nso,
1996
Kot
ov,2
000K
ozho
wia
11
33
81
18
77
7+1
47
4+1
40
54
20
0

174
Table
1.
Contin
ued.
Sou
rce
oflim
bP
IP
IIP
IIIP
IVP
VP
VI
info
rmat
ion
Gen
usA
cces
sory
OD
LID
Lfe
mID
Lm
ale
Mar
gina
lm
xpr
Iex
IIst
ifffp
IIex
IIIst
iffso
ftfp
IIIex
IVst
iffso
ftfp
IVex
Vm
argi
nal
fpV
pres
ent?
seta
seta
ese
tae
Raj
apak
sa&
Kurz
ia1
13
48
00
87
77+
14
76
44
54
23
0
Fer
nand
o,19
86;
Hud
ec,2
000
Sm
irnov
,198
9;
Fre
y,19
98L
eber
is1
12–
?9
10
87
6?
?9–
6?
?7
42
00
102+
1
Sar
s,18
85;Y
oon
&L
eydig
ia0
13
39
00–
18
6–83
–57+
14
6–7
64
45
42
20
Kim
,199
3;A
lons
o,
1996
;Kot
ov,
unpu
blis
hed
Val
divi
aV
illar
,L
eydig
iopsi
s?
13
?8
?0
87
77+
14
76
44
54
20
0
1984
;Rey
&
Vas
quez
,198
6
Alo
nso,
1996
Monosp
ilus
11
2+1
?8
01
88
77+
14
76
43
54
20
0
Chi
ambe
ng&
Nic
smirnov
ius
01
3?
81
08
76
84
86
40
54
22
0
Dum
ont,
1999
Raj
apak
sa&
Noto
alo
na
01
3?
7–9
00–
18
77
?–8
4?7
64
?–3
53–
42
00
Fer
nand
o,19
87b
Sm
irnov
,197
1;O
xyur
ella
01
23
80
08
77
7+1
47
64
35
42
00
Mic
hael
&F
rey,
1983
Kot
ov,2
000
Para
kozh
ow
ia1
13
38
11
87
77+
14
76
43
54
20
1
Sm
irnov
,197
1;R
hyn
chota
lona1
12
38
11
87
77+
14
76
43
54
23
0–
Alo
nso,
1996
1?
Ciro
s-Pe
rez
&S
pina
lona
0?1
3?
80
18
54
7+1
55
65
32
32
00
Elia
s-G
utier
rez,
1997
Fre
y,19
65;A
lons
o,Tre
toce
phala
01
34
91r
08
87
7+1
47
64
35
42
00
1996
;Kot
ov,
unpu
blis
hed
ALO
NIN
I,in
0–1
1–2
2–3
3–4
7–9
0–1
0–1
85–
83–
7?–
83–
55–
4–6
4–5
0–4
2–7
3–5
2–3
0–3
0–1
tota
l10
Pre
sent
edda
taIn
dial
ona
01
22
30
06
45
33
54
43
43
20
0

175
Figures 55–63.Thoracic limbs ofTretocephala ambiguafrom Lake Kravtsevo, Stavropolsky Kray, Russia.Figures 55–56. Limb I and its distalportion;Figure 57. Its inner face, somewhat schematically;Figure 58. Limb II; Figures 59–60. Limb III and distal armature of its gnathobase;Figures 61–62. Limb IV and distal armature of its gnathobase;Figure 63. Limb V. Scale bar deotes 100µm.
in limbs I–III. As a result, they are more similar tolimbs of Alonini (Figs 61–63). InIndialona, there areonly four setae on exopodite IV. Until recently, it wasconsidered, that there are seven setae on the exopod-ite IV in Chydorinae, and six in Aloninae (Smirnov,1971). However, it is shown nowadays that in somemembers of the Aloninae, this number is less, i.e. five
in someAlona (Dumont & Brancelj, 1994), and evenfour plus one rudimentary hillock inKozhowia. As aresult, a diagnosis of the subfamily must be amended(Kotov, 2000). Indialona has four setae on exopod-ite IV, an extremely rare for chydorids number, butthis character is obviously of Alonina-nature, becausethe same number was previously found in another

176
genus of the subfamily. There are only three setaeon exopodite V inIndialona, and only in two otherrepresentatives of Alonini,Spinalona anophtalmaandNotoalona freyi, the same number occurs. In the ma-jority of Aloninae, there are four setae, and five inAlona hudeci(Hudec, 1998; Sinev, 1999a). No doubta small, solitary distalmost seta (1 in Fig. 63 forTre-tocephala) is reduced inIndialona. In general, limbsIV–V in Indialonini have a minimal set of elements,but that number may also occur in some Alonini.
It is interesting that both in Alonini and India-lonini, the distal armature of gnathobases II–IV con-sists of four setae, a structure not subject of oligo-merization within the subfamily. We may certainlyfind each of four setae in all Aloninae, and the ele-ments on my pictures, marked by the same numbers,are homologues. The same number of homologouselements is characteristic of gnathobases II–IV in Chy-dorinae (Smirnov, 1971),Eurycercus(Fryer, 1963;Frey, 1975),BosminaandBosminopsis(Kotov, 1997),Ilyocryptus(Kotov, 1999) and many genera of Mac-rothricidae s. lat. (Dumont & Silva-Briano, 1998).Generally, the number of elements in the distal arma-ture of the gnathobase is a conservative trait within the‘chydorid’ branch of the Anomopoda (Kotov, 1998).
Why not a new subfamily?
Petkovski (1966) gave only one proof of the alonine-nature ofIndialona, viz. the type of mandibular articu-lation with the head shield. I confirm that this is reallyof the Aloninae-type.
I also found lateral head pores inIndialona thatwere very small and were located rather far from themain pores. This too is a typical trait of the Aloninaein contrast with the Chydorinae with small pores onmidline of head shield. The external appearance of thesystem of both main and lateral pores inIndialona isunique for Aloninae, and a good diagnostic feature ofthe genus. There is some superficial similarity of thissystem betweenIndialonaandEurycercus. However,upon closer examination, essential differences may befound: in Eurycercus(1) the main ‘pore’ is actuallya window in the cuticle with five additional minutepores, (2) in all subgenera, lateral pores are locatednear the major one (Frey, 1975). Lateral pores locatedfar from the midline of the head shield are character-istic of Saycia(Frey, 1971; Smirnov, 1996), and alsoof Bosmina(Goulden & Frey, 1963; Kotov, 1998).
Finally, four setae on exopodite IV is an alonine,not a chydorine feature (see above).
There is a number of traits, especially of thoraciclimbs, in whichIndialonamarkedly differs from othergenera of both Aloninae and Chydorinae, however Ido not create a new subfamily for the genusIndialona,because I believe thatIndialona is an extremely spe-cialised animal, that originated from a typical alonina-like ancestor. Earlier, Wolterek (1920) proposed atheory of the origin of planktonic Bosminidae fromlittoral Chydorus-like animals. This statement is notcorrect, because bosminids are more primitive thanchydorines in many traits, especially the limbs (Kotov,1997, 1998). However,Indialona does not possessmore primitive features than the Alonini, and differsfrom them only in a series of advanced characters,which can be considered to have originated after theseparation of tribe Indialonini from the main branchof the Aloninae. This transformation was linked to areduction in body size and transition to a planktonicmode of life. Wolterek’s (1920) hypothesis can beapplied to the Aloninae for an explanation of the evol-ution of Indialona. No doubt the ancestor ofIndialonawas a true littoral animal, and penetration to the sub-littoral plankton was a secondary phenomenon. Onlyfew species of Chydorinae and Aloninae temporarilypenetrate the plankton of some reservoirs (Fryer, 1968;Smirnov, 1971).
Key for the discrimination of the tribes ofAloninae
1 (2) Many (seven or more) setae on corm of limb I,eight stiff setae on limb II . . . . . . . . . Alonini s. str.Additional characters: animals of various sizes, aspine on the basal segment of the second antennaepresent in at least rudimentary state.
2 (1) Only three setae on the corm of limb I, only sixsetae on the corm of limb II . . . Indialonini n. trib.Additional characters: minute animals, spine onbasal segment of exopod of second antenna absent.
Acknowledgements
I am grateful to Prof. N. N. Smirnov for this chanceto study his samples and for help at different phases ofmy work, and to Prof. H. J. Dumont for criticism andlinguistic corrections. Special thanks also to my wifeDr M. A. Gololobova for the determination of algae in

177
the samples, Dr I. B. Mertsalov for help in preparationof material and Mr V. N. Antropov for technical as-sistance in the work with SEM. The study was partlysupported by a grant from the Russian Foundation forBasic Research (99-04-48969), and a scholarship ofRussian Government to young scientists.
References
Alonso, M., 1996. Fauna Iberica. Crustacea Branchiopoda. Mus.Nac. Cien. Natur. Consejo Superior de Investigaciones Cienti-ficas, Madrid: 486 pp.
Alonso, M. & J. L. Pretus, 1989.Alona iberica, new species: firstevidence of noncosmopolitanism within theA. karuacomplex(Cladocera, Chydoridae). J. crust. biol. 9: 459–476.
Brancelj, A., 1997.Alona stochin. sp. – the third cave-dwellingcladoceran (Crustacea: Cladocera) from the Dinaric region.Hydrobiologia 360: 47–54.
Chiambeng, G. Y. & H. J. Dumont, 1999. New semi-terrestrialchydorids from the tropical rainforest of southwest Cameroon(Africa): Nicsmirnovius camerounensisn.gen. n.sp. andBryos-pilus africanusn.sp (Crustacea: Anomopoda). Hydrobiologia391: 259–270.
Ciros-Pérez, J. & M. Elías-Gutiérrez, 1997.Spinalona anophtalma,n. gen. n. sp. (Anomopoda, Chydoridae) a blind epigean clado-ceran from the Neovolcanic Province of Mexico. Hydrobiologia353: 19–28.
Daday, E. , 1908. Entomostraca et Hydrachnidae e Tibet. Rec.Indian Mus. 2: 323–341.
Dumont, H. J. & A. Brancelj, 1994.Alona alsafadiin.sp. from Ye-men, a primitive, groundwater-dwelling member of theA. kauragroup. Hydrobiologia 281: 57–64.
Dumont, H. J. & K. Martens, 1996. The freshwater microcrustaceaof Easter Island. Hydrobiologia 325: 83–99.
Dumont, H. J. & M. Silva-Briano, 1997. Sensory and glandularequipment of the trunk limbs of the Chydoridae and Macro-thricidae (Crustacea: Anomopoda). Hydrobiologia 360: 33–46.
Dumont, H. J. & M. Silva-Briano, 1998. A reclassification of theanomopod families Macrothricidae and Chydoridae, with thecreation of a new suborder, the Radopoda (Crustacea: Branchio-poda). Hydrobiologia 384: 119–149.
Forró, L. & D. G. Frey, 1982. Cladocera taxa described by E. Daday,together with a catalogue of pertinent material in the HungarianNatural History Museum. Misc. zool. Hungarica 1: 91–132.
Frey, D. G., 1965. A new genus of Chydoridae (Cladocera). Int. Rev.ges. Hydrobiol. 50: 153–168.
Frey, D. G., 1967. Phylogenetic relationships in the family Chydor-idae (Cladocera). Proc. Symp. on Crustacea, Marine Biol. Assoc.India, 12–15 Jan. 1965, Ernakulam, Part 1: 29–37.
Frey, D. G., 1971. Worldwide distribution and ecology ofEurycer-cusandSaycia(Cladocera). Limnol. Oceanogr. 16: 254–308.
Frey, D. G., 1975. Subgeneric differentiation withinEurycer-cus (Cladocera, Chydoridae) and a new species from NorthernSweden. Hydrobiologia 46: 263–300.
Frey, D. G., 1980. The non-swimming chydorid Cladocera of wetforests, with descriptions of a new genus and two new species.Int. Rev. ges. Hydrobiol. 65: 613–641.
Frey, D. G., 1988.Alona weineckiStuder on the subantarctic islands,not Alona rectangulaSars (Chydoridae, Cladocera). Limnol.Oceanogr. 33: 1386–1411.
Frey, D. G., 1991. A new genus of alonine chydorid cladoceransfrom athalassic saline waters of New South Wales, Australia.Hydrobiologia 224: 11–48.
Frey, D. G., 1998. Expanded description ofLeberis aenigmatosaSmirnov (Anomopoda: Chydoridae): a further indication ofthe biological isolation between western and eastern Australia.Hydrobiologia 367: 31–42.
Fryer, G., 1963. The functional morphology and the feeding mech-anism of the chydorid cladoceranEurycercus lamellatus(O. F.Müller). Trans. r. Soc. Edinb. 65: 335–381.
Fryer, G., 1968. Evolution and adaptive radiation in the Chydor-idae (Crustacea: Cladocera): a study of comparative functionalmorphology and ecology. Phil. Trans. r. Soc., Lond. B. 254:221–385.
Fryer, G., 1999. A comment on a recent phylogenetic analysis ofcertain orders of the branchiopod Crustacea. Crustaceana 72:1039–1050.
Goulden, C. E. & D. G. Frey, 1963. The ocurrence and significanceof lateral head pores in the genusBosmina(Cladocera). Int. Rev.ges. Hydrobiol. 48: 513–522.
Hudec, I., 1998. Anomopoda (Crustacea: Branchiopoda) from someVenezuelan tepuis. Hydrobiologia 377: 205–211.
Hudec, I., 2000. Subgeneric differentiation withinKurzia (Crusta-cea: Anomopoda: Chydoridae) and a new species from CentralAmerica. Hydrobiologia 421: 165–178.
Kotov, A. A., 1997. Structure of thoraic limbs ofBosminopsisdeitersiRichard, 1895. Hydrobiologia 360: 25–32.
Kotov, A. A., 1998. Sravnitelniy analiz morfologii Anomopoda(Crustacea, Branchiopoda). Ph. D. Thesis, Institute of Eco-logy and Evolution, Moscow: 309 pp. (Comparative analysis ofmorphology of the Anomopoda (Crustacea, Branchiopoda). InRussian).
Kotov, A. A., 1999. Morphology and variability ofIlyocryptusagilis Kurz, 1878 andIlyocryptuscf. cuneatusŠtifter, 1988 fromLake Glubokoe, Moscow Area, central Russia (Branchiopoda:Anomopoda). Arthropoda Selecta 8: 3–22.
Kotov, A. A., 2000. Analysis ofKozhowiaVasiljeva & Smirnov,1969 (Chydoridae, Anomopoda, Branchiopoda), and a descrip-tion of Parakozhowian. gen. Hydrobiologia 437: 17–56.
Michael, R. G. & D. G. Frey, 1983. Assumed amphi-atlantic distri-bution ofOxyurella tenuicaudis(Cladocera, Chydoridae) deniedby a new species from North America. Hydrobiologia 106: 3–35.
Olesen, J., 1998. A phylogenetic analysis of the Conchostraca andCladocera (Crustacea, Branchiopoda, Diplostraca) Zool. J. linn.Soc. 122: 491–536.
Paggi, J. C., 1975. Sobre dos Cladoceros Chydoridae nuevos para lafauna Argentina. Physis (Buenos Aires) B 34: 139–150.
Paggi, J. C., 1979. Aportes al Conocimiento de la fauna Argentinade Cladoceros. II. SobreAlona eximiaKiser, 1948. Neotropica25: 177–185.
Paggi, J. C., 1980. Aportes al Conocimiento de la fauna Argentinade Cladoceros. II.Euryalona fasciculataDaday, 1905 yEury-alona occidentalisSars, 1901. Rev. Asoc. Cienc. Nat. Literal.11: 145–160.
Paggi, J. C., 1987. Estudos limnologicos en la Isla Deception, Shet-land del sur, Antarctida: aspectos taxonomicos de los Crustaceosdel zooplancton. Dir. Nat. Antarct., Contr. No 338: 1–34.
Petkovski, T. K., 1966. Eine neue Cladoceren-Gattung aus demwesten IndiensIndialona ganapatin. gen. et n. sp. Fragm.Balcan. Maced. Sci. nat. 5: 157–165.
Rajapaksa, R. & C. H. Fernando, 1985. Reallocation ofIndialonamacronyx(Daday, 1898) (Cladocera, Chydoridae) to the genusAlona. Can. J. Zool. 63: 970–976.

178
Rajapaksa, R. & C. H. Fernando, 1986. Tropical species ofKurzia(Crustacea, Cladocera), with a description ofKurzia brevilabrissp. nov. Can. J. Zool. 64: 2590–2602.
Rajapaksa, R. & C. H. Fernando, 1987a. A redescription ofEury-alona orientalis (Daday, 1898), with a consideration of theother species in the genusEuryalona(Cladocera: Chydoridae).Hydrobiologia 150: 75–90.
Rajapaksa, R. & C. H. Fernando, 1987b. Redescription and assign-ment ofAlona globulosaDaday, 1898 to a new genusNotoalonaand a description ofNotoalona freyisp. nov. Hydrobiologia 144:131–153.
Rey, J. & E. Vasquez, 1986. Contribution à la connaissance desCladocères néotropicaux: redescription deLeydigiopsis ornataDaday, 1905 (Crustacea, Cladocera). Ann. Limnol. 22: 169–176
Sars, G. O., 1885. On some Australian Cladocera, raised from driedmud. Forh. VidenskSelsk. Krist. 8: 1–46.
Sharma, B. K. & S. Sharma, 1990. On the taxonomic status of somecladoceran taxa (Crustacea: Cladocera) from Central India. Rev.Hydrobiol. trop. 23: 105–113.
Silva-Briano, M., 1998. A revision of Macrothricid-like anomo-pods. Ph.D. Thesis, Ghent University: 388 pp.
Sinev, A. Y., 1997. Review of theaffinis-group of Alona Baird,1843, with the description of a new species from Australia(Anomopoda Chydoridae). Arthropoda Selecta 6: 47–58.
Sinev, A. Y., 1998.Alona ossianisp. n., a new species of theAlonaaffiniscomplex from Brazil, deriving from the collection of G. O.Sars (Anomopoda Chydoridae). Arthropoda Selecta 7: 103–110.
Sinev, A. Y., 1999a.Alona costataSars, 1862versusrelated pa-laeotropical species: the first example of close relations betweenspecies with a different number of main head pores amongChydoridae (Crustacea: Anomopoda). Anhropoda Selecta 8:131–148.
Sinev, A. Y., 1999b.Alona werestschaginisp. n., new speciesof genusAlona Baird, 1843, related toA. guttataSars, 1862(Anomopoda, Chydoridae). Arthropoda Selecta 8: 23–30.
Sinev, A. Y. & A. A. Kotov, 2000. Redescription ofAlona lab-rosa Vasiljeva & Smirnov, 1969 (Chydoridae, Anomopoda,Branchiopoda). Hydrobiologia 439: 125–139.
Smirnov, N. N., 1971. Chydoridae fauni mira. Fauna SSSR. Rakoo-braznie 1 (2): 1–531 (Chydoridae of the world fauna. In Russian).
Smirnov, N. N., 1989. Tropicheskie Cladocera. 2. Novie vidy se-meystv Chydoridae, Macrothricidae i Moinidae tropicheskoyAvstralii. Zoologicheskii zhurnal 68: 51–59 (Tropical Cladocera.2. New species of Chydoridae, Macrothricidae and Moinidaefrom tropical Australia. In Russian).
Smirnov, N. N., 1995a. Check-list of the Australian Cladocera(Crustacea). Arthropoda Selecta 4: 3–6.
Smirnov, N. N., 1995b. Semejstvo Chydoridae. In Alekseev, V.R. (ed.), Opredelitel Bespozvonochnih Rossii i SopredelnihTerritoriy. 2. Crustacea: 58–60 (Family Chydoridae. In Russian).
Smirnov, N. N., 1996. Cladocera: the Chydorinae and Sayciinae(Chydoridae) of the world. Guides to the Identification of the Mi-croivertebrates of the Continental Waters of the World 11. SPBAcad. Publ., Amsterdam: 197 pp.
Smirnov, N. N., 1998. A revision of the genusCamptocercus(Anomopoda, Chydoridae, Aloninae). Hydrobiologia 386: 63–83.
Štifter, P., 1991. A review of the genusIlyocryptus (Crustacea:Anomopoda) from Europe. Hydrobiologia 225: 1–8.
Valdivia Villar, R. S., 1984. K morfologii neotropicheskih rakoo-braznih roda Leydigiopsis (Cladocera, Chydoridae). Zool.Zhurnal 63: 1572–1575 (On the morphology of Neotropicalcrustaceans of genusLeydigiopsis(Cladocera, Chydoridae. InRussian).
Wolterek, R., 1920. Variation und Artbildung. Analytische undexperimentelle Untersuchungen an pelagischen Daphniden undanderen Cladoceren. Francke Verl., Berlin, 151 ss.
Yoon, S. M. & W. Kim, 1993. Redescription of two chydorid spe-cies of genusLeydigiaKurz, 1875 (Branchiopoda, Anomopoda,Chydoridae) from Korea. Korean J. Zool. 36: 380–390.
Copyright © 2022 FDOKUMEN




















