![Belize Rural Development Strategy (BRADS) [in Spanish]](https://static.fdokumen.com/doc/165x107/6314a0a7c32ab5e46f0cf89b/belize-rural-development-strategy-brads-in-spanish.jpg)
RECONSTRUCTING INFANT DIET AND WEANING BEHAVIOR OF ANCIENT MAYA FROM LAMANAI, BELIZE USING LASER...
421
RECONSTRUCTING INFANT DIET AND WEANING BEHAVIOR OF ANCIENT MAYA FROM LAMANAI, BELIZE USING LASER ABLATION-INDUCTIVELY COUPLED PLASMA-MASS SPECTROMETRY (LA-ICP-MS) A Dissertation Presented by RHAN-JU SONG Submitted to the Graduate School of the University of Massachusetts Amherst in partial fulfillment of the requirements for the degree of DOCTOR OF PHILOSOPHY September 2004 Anthropology
Transcript of RECONSTRUCTING INFANT DIET AND WEANING BEHAVIOR OF ANCIENT MAYA FROM LAMANAI, BELIZE USING LASER...

RECONSTRUCTING INFANT DIET AND WEANING BEHAVIOR
OF ANCIENT MAYA FROM LAMANAI, BELIZE
USING LASER ABLATION-INDUCTIVELY COUPLED
PLASMA-MASS SPECTROMETRY
(LA-ICP-MS)
A Dissertation Presented
by
RHAN-JU SONG
Submitted to the Graduate School of the
University of Massachusetts Amherst in partial fulfillment
of the requirements for the degree of
DOCTOR OF PHILOSOPHY
September 2004
Anthropology

© Copyright by Rhan-Ju Song 2004
All Rights Reserved

RECONSTRUCTING INFANT DIET AND WEANING BEHAVIOR
OF ANCIENT MAYA FROM LAMANAI, BELIZE
USING LASER ABLATION-INDUCTIVELY COUPLED
PLASMA-MASS SPECTROMETRY
(LA-ICP-MS)
A Dissertation Presented
by
RHAN-JU SONG
Approved as to style and content by:
Alan H. Goodman, Chair
Dulasari Amarasiriwardena, Member
R. Brooke Thomas, Member
Ralph Faulkingham, Department Chair
Anthropology

DEDICATION
To my family.

ACKNOWLEDGMENTS
The end of a journey is a contemplative occasion. When one finally arrives at the
intended destination, reflection always competes with relief and happiness to crowd one’s
emotions. Like most lengthy trips, this dissertation has been shaped by the assistance,
experience, inspiration and insight of many individuals.
Foremost, this research has been nurtured by Dr. Alan H. Goodman (Hampshire
College), who has been the most encouraging, flexible and patient advisor since I first
emailed him in 1996. I thank him immensely for his enthusiastic support, which never
wavered, even as I moved to farther destinations. The other members of my committee,
Drs. Dula Amarasiriwardena, Brooke Thomas, as well as Drs. Debra Martin and Alan
Swedlund, are equally acknowledged for their wisdom, understanding, and friendship.
They continue to broaden my perspective and enlighten my approach.
I also thank Deb Martin for her enormous generosity, particularly her support and
willingness to accommodate my personal circumstances. Pete and I only hope that we
have provided plenty of amusing memories in return!
At the Department of Anthropology, I have benefited considerably from the academic
contributions of the faculty and I am proud to be associated with them. As well, I am
grateful to Shelley Bellor Richotte for help with many of the “Fifty Three Easy Steps”.
Fellow grad students have also furthered my education and research objectives. These
wonderful friends, who selflessly offered logistical support in many instances, include
Joseph Jones, Sacha Page, Ventura Perez, Flavia Stanley and Pam Stone. Ventura Perez
and Kathleen Brown Perez are especially thanked for their generous assistance,
indubitable character, and committed friendship.
v

At Hampshire College, my analysis was facilitated by Sue Keydel, Laurie Smith and
Kristin Shrout, who provided invaluable logistical assistance with the LA-ICP-MS
machinery, as well as access to computers and other equipment. Other individuals who
provided important research advice include Drs. Christine White, Lori Wright, Peter
Outridge, Don Reid, Louise Humphrey, Phil Kelleher and John Reid.
For the privilege of analyzing ancient Maya human remains, I owe immense thanks to
several individuals: Drs. Jaime Awe and Allan Moore, of the Belize Department of
Archaeology; Drs. Elizabeth Graham and David M. Pendergast, principal investigators of
the Lamanai Archaeological Project; and Drs. Hermann Helmuth and Paul F. Healy from
the Dept. of Anthropology at Trent University. Funding for this research was generously
provided by the Graduate School at the University of Massachusetts and the Wenner-
Gren Foundation for Anthropological Research and they are duly recognized.
Above all, I am grateful to my family for their infinite love, steadfast support and
patience. This remarkable bunch includes Kim, Nak, Ba, Christine, Dennis and Pete,
who have been the steady keel of my sometimes-flagging ship. This manuscript could
not have been completed without their involvement and I am forever indebted to them.
My husband, Peter A. Zubrzycki, was there when I formulated this research, when we
prepared the first tooth for analysis, and he continues to provide endless support. I thank
him wholeheartedly. During this undertaking, we have traveled from Amherst to
Toronto, driven through North and Central America to live in the jungles of Belize, criss-
crossed the Atlantic innumerable times and confronted many challenges together. This
document is as much a product of his participation and encouragement, and he remains
my most worthy partner in this journey.
vi

vii
ABSTRACT
RECONSTRUCTING INFANT DIET AND WEANING BEHAVIOR OF
ANCIENT MAYA FROM LAMANAI, BELIZE USING LASER ABLATION-
INDUCTIVELY COUPLED PLASMA-MASS SPECTROMETRY (LA-ICP-MS)
SEPTEMBER 2004
RHAN-JU SONG, B.A., UNIVERSITY OF TORONTO
M.A., TRENT UNIVERSITY
Ph.D., UNIVERSITY OF MASSACHUSETTS AMHERST
Directed by: Professor Alan H. Goodman
This investigation represents the first extensive application of Laser Ablation-
Inductively Coupled Plasma-Mass Spectrometry to ancient dental analysis and
paleodietary reconstruction. Enamel strontium composition is examined because it is a
reliable hard tissue indicator of diet during enamel formation in childhood.
Here, infant diet and weaning behavior of pre-contact and colonial period Maya from
Lamanai, Belize are reconstructed. Weaning is a critical dietary transition that has
adaptive significance for later life. Since the strontium-calcium (Sr/Ca) ratio of solid
food is high compared to that of breast milk, strontium composition of hard tissues
developing before, and after, food supplementation can infer the timing of food
introduction and weaning. Known timing of permanent enamel development allows
correlation of canine enamel Sr/Ca values with age in childhood, which is facilitated by
continuous laser microsampling.

viii
The results indicate that enamel Sr/Ca faithfully records a biogenic signal associated
with childhood dietary intake. The total Sr/Ca pattern generally follows the projected
model of strontium change, with food supplementation starting at around nine months of
age, which increases gradually until there is a substantial surge in food intake at
approximately two years of age. Lamanai children continue to nurse afterward, possibly
up to five years of age, but it comprises a minor nutritional component.
Significantly, the disadvantaged colonial Maya cohort has a reduced Sr/Ca pattern
compared to elite Postclassic Maya, suggesting that colonial children may have
exclusively breastfed for longer, delaying the age of food supplementation and weaning.
Colonial Maya also exhibit greater Sr/Ca variation, reflecting dietary shifts that can be
attributed to poorer nutrition and health. Female economic responsibilities partially
account for the differences in colonial childcare practices. Additionally, enamel variation
may reflect the different childhood origins (and weaning patterns) of possible migrants at
Lamanai. Ultimately, the nature of infant diet and enamel Sr/Ca can be linked to
interrelated ecological, political and economic factors.
LA-ICP-MS analysis reveals enamel Sr/Ca to be a sensitive gauge of the prolonged
and complex process of weaning and it proves to be an ideal method of capturing the
richness of early life history documented in sequentially-formed enamel.

ix
TABLE OF CONTENTS
Page
ACKNOWLEDGMENTS ……………………………………………………………….. v
ABSTRACT ……………………………………………………………………………. vii
LIST OF TABLES ……………………………………………………………………... xii
LIST OF FIGURES …………………………………………………………………….. xv
CHAPTER
1. INTRODUCTION TO RESEARCH.……………………………………………….... 1
1.1 Introduction…………………………………………………………….... 1
1.2 Cultural Context Of The Investigation……………………………….…. 6
1.3 Theoretical Framework…………………………………………….……. 7
2. THE MAYA OF LAMANAI, BELIZE.……………………………………………... 9
2.1 Lamanai Research History.…………………………………………….... 9
2.2 Environmental Setting.………………………………………………… 12
2.3 The Nature of Human Settlement at Lamanai.………………………… 14
2.4 Food Resources.………………………………………………………... 18
2.5 The Postclassic and Historical Periods at Lamanai....…………………. 19
2.6 The Biological Consequences of Maya-Spanish Contact at Lamanai..... 33
2.7 Bone Chemical Evidence of Diet at Lamanai………………………….. 40
2.8 The Repercussions of Maya-Spanish Contact on Infant Nutrition…….. 42
3. DENTAL HARD TISSUES AND THEIR APPLICATION
IN PALEONUTRITIONAL RESEARCH…………………………………………. 45
3.1 Dental Anthropology in Archaeological and
Paleonutritional Reconstruction.……………………………………….. 45
3.2 Dental Hard Tissues and Enamel Development.………………………. 49
3.3 Dental Enamel Composition.…………………………………………... 57
3.4 Diagenesis and Hard Tissue Preservation.……………………………... 61
3.5 The Nature of Hard Tissue Preservation at Lamanai.………………….. 67

x
4. THE ROLE OF STRONTIUM ANALYSIS IN PALEONUTRITION…………….. 70
4.1 Strontium Distribution in the Food Web.……………………………… 70
4.2 The Role of Strontium in Human Nutrition.…………………………… 83
4.3 Strontium Absorption and Hard Tissue Incorporation.…………………84
4.4 Variations in Strontium Absorption.…………………………………… 88
4.5 Applicability of Strontium in Hard Tissue Paleodietary Analysis.……. 98
4.6 Strontium Preservation in Fossilized Hard Tissues.…………………...100
4.7 The Effects of Maize Processing on Hard Tissue Sr/Ca Composition.. 102
5. BREASTFEEDING, WEANING AND INFANT HEALTH……………………… 108
5.1 The Nutritional Qualities of Human Breast Milk.…………………….. 108
5.2 Determining Factors in Breastfeeding and Weaning Behavior.………. 113
5.3 The Biological Bases for Breastfeeding Duration and Weaning Age.... 119
5.4 Methods of Identifying Weaning Patterns in the Past.………………...125
5.5 Hard Tissue Strontium and the Inferences for Infant Nutrition.……… 132
5.6 Enamel Sr/Ca and Infant Nutrition at Lamanai.………………………. 136
6. MATERIALS AND METHODS…………………………………………………... 140
6.1 Analytical Technique.………………………………………………… 140
6.2 Technological and Methodological Challenges.……………………… 144
6.3 Materials and Methods.……………………………………………….. 152
6.4 Sample Preparation.…………………………………………………... 155
6.5 A Protocol for LA-ICP-MS Analyses of Human Dental Enamel.……. 158
6.6 LA-ICP-MS Observations.……………………………………………. 164
6.7 Laser Ablation Details for Lamanai Canines.………………………… 167
6.8 Data Analysis.………………………………………………………… 169
7. RESULTS OF ANALYSIS………………………………………………………... 177
7.1 Major Findings.……………………………………………………….. 177
7.2 The Lamanai Sr/Ca Pattern.…………………………………………... 180
7.3 The Sr/Ca Patterns of Lamanai Cohorts.……………………………… 190
7.4 Sr/Ca Patterns among Lamanai Individuals.………………………….. 212
7.5 The Nature of Enamel Sr/Ca Change in Lamanai Individuals.………. 224
7.6 Determination of Dietary Sr/Ca from Enamel Sr/Ca.………………… 244

xi
8. DISCUSSION……………………………………………………………………… 252
8.1 The Implications of Maize Processing for Dietary Reconstruction…... 253
8.2 Sr/Ca Behavior in Lamanai Permanent Canine Enamel.……………... 264
8.3 The Nature of Ancient Maya Health and Nutrition at Contact.………. 268
8.4 Implications of Infant Weaning Behavior for Ancient Maya Society... 281
8.5 The Comparative Hard Tissue Strontium Data.………………………. 286
8.6 Recommendations for Future Research.……………………………… 294
9. CONCLUDING REMARKS.…………………………………………………….... 298
APPENDICES
A. DENTAL HEALTH OF LAMANAI CANINES…………………………………. 305
B. ABLATION DETAILS FOR LAMANAI CANINES.……………………………. 327
C. STATISTICAL RESULTS………………………………………………………... 336
BIBLIOGRAPHY.………………………………………………………………...…… 345

xii
LIST OF TABLES
Page
2.1 Lamanai chronology.……………………………………………………... 11
3.1 Concentrations of major components of human dental enamel
excluding water and organic material.………………………………….… 59
3.2 Range of element concentrations in human dental enamel.…………….... 59
4.1 Average Sr content of various foods in order of relative abundance,
with some traditional Maya foods italicized.……………………………... 73
4.2 Mean Sr/Ca values for common Maya animal and plant foods
from the Pasion region of Guatemala.………...………………………….. 76
4.3 Ranges of Sr and Zn content in mammalian bones..……………………... 78
5.1 Constituents of human milk.………………………………………….…. 109
5.2 Bone Sr/Ca changes from the Dor sample, Israel.………………….…… 134
5.3 Bone Sr/Ca changes from the Schleswig sample, Germany.………….… 135
6.1 Sample Breakdown.……………………………………………………... 154
6.2 LA-ICP-MS Operating Parameters.………………………..………….… 160
7.1 Summary of mean 86
Sr/43
Ca for 0.25 year intervals.……………………. 182
7.2 Results of ANOVA comparing Postclassic and Historical individuals..... 184
7.3 Mean 86
Sr/43
Ca ratios for intervals between 0.5 and 5.5 years of age…... 185
7.4 Mean total Sr/Ca data (x10-3
) for Postclassic and Historical cohorts.…... 187
7.5 Results of Single Factor ANOVA (two-tailed) comparing Postclassic
Children and Adults.…………………………………………………….. 191
7.6 Results of Single Factor ANOVA (two-tailed) comparing Postclassic
Females and Males.…………………………………………………….... 192
7.7 Results of Single Factor ANOVA (two-tailed) comparing Historical
Children and Adults.…………………………………………………….. 193

xiii
7.8 Results of Single Factor ANOVA (two-tailed) comparing Historical
Females and Males.…………………………………………………….... 194
7.9 Average Sr/Ca values for 0.25 year intervals of Postclassic Children.….. 213
7.10 Average Sr/Ca values for 0.25 year intervals of Postclassic Females…... 214
7.11 Average Sr/Ca values for 0.25 year intervals of Postclassic Males.…….. 215
7.12 Average Sr/Ca values for 0.25 year intervals of Historical Children.…... 217
7.13 Average Sr/Ca values for 0.25 year intervals of Historical Females……. 218
7.14 Average Sr/Ca values for 0.25 year intervals of Historical Males.…….... 219
7.15 Average greatest increase in Sr/Ca from infancy to early childhood
(0.5-5.5 yrs) among Postclassic and Historical period individuals…….... 221
7.16 Age-related bone-diet observed ratios (ORbone-diet).……………………... 244
7.17 Enamel vs. Dietary Sr/Ca values for Lamanai cohorts at various ages…. 245
8.1 Relative Sr and Ca data for Lamanai bone and soil samples.………….... 257
8.2 Mean bone and soil Sr/Ca of Pasion Maya sites in Guatemala…………. 258
8.3 Comparative age-related mean Sr/Ca (x 10-3
) values from various
clinical and paleodietary investigations.……………………………….... 287
A.1 Regression formulae for calculation of timing of enamel defect
formation in Lamanai canines.……………………………………......…. 314
A.2 Average ages of enamel defect formation in the Lamanai sample....….... 318
A.3 Dental health details for Lamanai canines.………………………...……. 320
B.1 Ablation details for Postclassic Lamanai maxillary canines.…...…..….... 328
B.2 Ablation details for Postclassic Lamanai mandibular canines.…….…..... 330
B.3 Ablation details for Historical Lamanai maxillary canines……………... 332
B.4 Ablation details for Historical Lamanai mandibular canines.………….... 334

xiv
C.1 Results of Single Factor ANOVAs comparing total Sr/Ca increase
between Postclassic children and adults………………………………… 336
C.2 Results of Single Factor ANOVAs comparing total Sr/Ca increase
between Postclassic females and males…………………………….…… 336
C.3 Results of Single Factor ANOVAs comparing total Sr/Ca increase
between Historical children and adults………………………………...... 336
C.4 Results of Single Factor ANOVAs comparing total Sr/Ca increase
between Historical females and males………………………………....... 337
C.5 Results of Single Factor ANOVAs comparing total Sr/Ca increase
between Postclassic and Historical children………………………....….. 337
C.6 Results of Single Factor ANOVAs comparing total Sr/Ca increase
between Postclassic and Historical females…………………………....... 337
C.7 Results of Single Factor ANOVAs comparing total Sr/Ca increase
between Postclassic and Historical males…………………………......… 337
C.8 Results of Single Factor ANOVAs comparing Postclassic children
and adults.……………….……………………………………………..... 338
C.9 Results of Single Factor ANOVAs comparing Postclassic females
and males.……………………………………...……………..………….. 339
C.10 Results of Single Factor ANOVAs comparing Historical children
and adults.…………………………………………………………..….... 340
C.11 Results of Single Factor ANOVAs comparing Historical females
and males.………………………………………………...…………….... 341
C.12 Results of Single Factor ANOVAs comparing Postclassic
and Historical children.………………………………………………….. 342
C.13 Results of Single Factor ANOVAs comparing Postclassic
and Historical females.…………………………………………………... 343
C.14 Results of Single Factor ANOVAs comparing Postclassic
and Historical males.……………………………………………...……... 344

xv
LIST OF FIGURES
Page
2.1 Map of Belize showing the location of Lamanai.………………………… 10
2.2 Drawing of Structure N10-2.……………………………………………... 20
2.3 Map illustrating the southeastern Maya frontier,
with provinces and sites indicated.………………….……………………. 22
2.4 Remains of the second Spanish church at Lamanai.……………………… 24
3.1 Cross-section of human tooth, with major anatomical features indicated... 50
3.2 Three-dimensional chemical configuration of hydroxyapatite.………..…. 51
5.1 Proposed model of Sr/Ca behavior during infancy and early childhood... 137
6.1 Tooth cross-section indicating correlation of laser ablation
position, development age and enamel Sr/Ca intensity ratio…..………... 142
6.2 Position of laser ablation trenches along secondary canine crown.……... 158
6.3 Schematic of instrumentation used in LA-ICP-MS.…………………….. 161
6.4 Example of LA-ICP-MS recording sheet used in this analysis.………… 162
6.5 Graph illustrating distinct patterns in Sr/Ca during LA-ICP-MS.………. 165
6.6 Photograph of ablated tooth section and micrometer scale.……………... 167
6.7 Graphical comparison of different Sr/Ca isotope ratios in enamel.……... 173
6.8 Adjusted comparison of different Sr/Ca isotope ratios.…………………. 174
7.1 Standard Error graph of mean 86
Sr/43
Ca for all Lamanai individuals.…... 181
7.2 Standard Error graph of mean Sr/Ca for Postclassic
and Historical individuals.………………………………………………. 183
7.3 Scatter plot distributions of total Sr/Ca for Lamanai cohorts.…………... 188
7.4 Sr/Ca patterns of Lamanai cohorts.……………………………………… 190
7.5 Average Sr/Ca for Postclassic and Historical individuals.……………… 196

xvi
7.6 Comparison of total Historical sample with Postclassic adults.………… 197
7.7 Comparison of Postclassic and Historical cohorts.……………………… 200
7.8 Comparison of Postclassic Children and Adults.…..……………………. 202
7.9 Comparison of Postclassic Females and Males.………………………… 204
7.10 Comparison of Historical Children and Adults.………………………… 206
7.11 Comparison of Historical Females and Males.………………………….. 207
7.12 Comparison of Postclassic and Historical Children.…………………….. 208
7.13 Comparison of Postclassic and Historical Females.…………………….. 209
7.14 Comparison of Postclassic and Historical Males.……………………….. 211
7.15 Graph of Sr/Ca change among Postclassic children.……………………. 213
7.16 Graph of Sr/Ca change among Postclassic females.…………………….. 216
7.17 Graph of Sr/Ca change among Postclassic males.………………………. 216
7.18 Graph of Sr/Ca change among Historical children.……………………... 217
7.19 Graph of Sr/Ca change among Historical females.……………………… 220
7.20 Graph of Sr/Ca change among Historical males.………………………... 220
7.21 Distribution of peak Sr/Ca values per age interval.……………………... 222
7.22 Graph illustrating Postclassic individuals
with a general increase in Sr/Ca over time.……………………………... 225
7.23 Graph illustrating Historical individuals
with a general increase in Sr/Ca over time.……………………………... 226
7.24 Graph illustrating Postclassic individuals
with fluctuations in Sr/Ca over time.……………………………………. 230
7.25 Graph illustrating Historical individuals
with fluctuations in Sr/Ca over time.……………………………………. 231

xvii
7.26 Graph illustrating Postclassic individuals
with a delayed increase in Sr/Ca over time.……………………………... 237
7.27 Graph illustrating Historical individuals
with a delayed increase in Sr/Ca over time.……………………………... 238
7.28 Histogram of age of significant Sr/Ca change in “delayed” individuals... 240
7.29 Graph illustrating Lamanai individuals with stable Sr/Ca over time.…… 242
7.30 Comparison of enamel Sr/Ca and dietary Sr/Ca in Lamanai cohorts.…... 246
A.1 Standards of enamel hypoplasia severity....……………………………... 310
A.2 Distribution of defect timings in Lamanai maxillary canines....………… 317
A.3 Distribution of defect timings in Lamanai mandibular canines....………. 317

1
CHAPTER 1
INTRODUCTION TO RESEARCH
1.1 Introduction
The dietary and nutritional status of people in the past can be inferred by numerous
bone and dental indicators. These indicators can be grouped into three levels of analysis,
namely surface manifestations of disease or undernutrition, microscopic or histological
features, and studies of the chemical composition of tissues.
With regard to chemical make-up, numerous attempts have been made to assess the
relationship between what is consumed during life with consequent hard tissue
composition. Significantly, the overwhelming majority of these investigations have
focused on the isotopic and elemental chemistry of bone for dietary inferences.
Hard tissue chemical analysis is based on the fundamental principle that different
food categories have distinct elemental and isotopic compositions and that these
differences are reflected in the tissues of their consumers. Thus, hard tissue chemical
analysis provides a direct reflection of ancient subsistence (the meal), unlike the indirect
inferences from faunal and floral data and material evidence associated with food
processing (the menu).
Quantitative and qualitative methods of dietary reconstruction, such as those based on
faunal and floral data, are subject to preservation and often misrepresent consumption
proportions. Such data, in addition to dental microwear, stature and skeletal pathologies
indicative of health and nutrition, discloses only relatively broad inferences about diet.
(Note, while dietary status refers to the sum of all foods ingested, regardless of nutrient

2
value, nutritional status, refers to the state of balance between diet, nutrient intake and
functional capacity.)
For the purposes of paleodietary reconstruction, hard tissue analysis encompasses
assessments of elemental composition and stable isotopes, which reflect those of ingested
food types. This approach is based on studies that linked animal feeding habits with
variations in bone chemistry and isotopic composition (DeNiro and Epstein 1978, 1981).
Specifically, the application of chemical methods to reconstruct ancient diet from hard
tissues is built on the pioneering work of Brown (1973) and Gilbert (1975), followed by
DeNiro and Epstein (1978), van der Merwe and Vogel (1978), Lambert and colleagues
(1979), and Schoeninger (1979).
Importantly, it should be stressed that diet is only one of many variables that affect
bone composition. Variables such as individual metabolism and digestion, which vary
with food types and meal conditions; meal preparation; unequal absorption of nutrients
by the body; and the role of other substances in the diet on total elemental absorption
(e.g., phytates in maize) all contribute to the expressed hard tissue chemistry of an
individual.
Hard tissue chemical composition is not static during life and can be distinct between
bones and teeth. In this regard, dental enamel only reflects chemistry during its
calcification, since the mineral phase, or hydroxyapatite, of enamel is essentially inert
once formed, while living bone continuously remodels at a rate of approximately 2-4%
calcium turnover per annum (Avioli 1988), or ten years for total replacement (Bumsted
1985: 544). Thus, bone potentially provides a chemical record of approximately the last
ten years of life, while teeth (enamel) record varying periods between in utero

3
development to approximately 18-21 years of age, the entire period of deciduous and
permanent tooth crown formation.
Hard tissue elemental composition can be used to distinguish diet because they vary
in abundance in the environment. Prominent elements generally associated with animals
(and animal protein) include zinc, copper, molybdenum and selenium, while common
elements in plant material include barium, strontium, magnesium, manganese, cobalt,
nickel and vanadium (see Gilbert and Mielke 1985; Price 1989; Sandford 1993).
Additionally, because of the multi-elemental nature of hard tissue chemical studies, the
relationships between elements can also be used to indicate diet. For example, relatively
high levels of zinc, but low levels of strontium and magnesium can distinguish a diet rich
in animal protein, while high values for manganese, magnesium and strontium suggest
heavy reliance on plant foods.
Subsequently, the potentials of bone chemical analysis for dietary reconstruction have
been widely adopted. Utilizing bone elemental and isotopic composition, various studies
have examined the contribution of plants versus animals to human diet (Schoeninger
1979; Sillen and Kavanagh 1982); marine versus terrestrial components of diet (Ambrose
1991; Burton and Price 1990; Chisholm et al. 1982; Connor and Slaughter 1984; Nelson
et al. 1983; Schoeninger et al. 1983; Schoeninger and DeNiro 1984; Sealy and Sillen
1988); the transformation from hunting and gathering to agriculture (Schoeninger 1982;
Sillen 1981); infant nutrition and weaning (Sillen and Smith 1984) (see Chapter Five);
and the exploitation of certain species of plants such as maize (Bender et al. 1981;
Katzenberg et al. 1993; Lynott et al. 1986; Norr 1981; Reed 1994; Schoeninger 1979;

4
Schoeninger et al. 1990; Vogel and van der Merwe 1977; White 1986; White et al. 1993,
1994; Wright 1994, 1997, 1999; among others).
To date, the analysis of hard tissue (bone) composition to infer childhood nutrition
has been clouded by challenges in pinpointing precise periods of early development. In
particular, it is not fully understood how bone chemistry reflects diet and nutrition at
different stages of development. Related to this is the problem of not knowing the time
delay between ingestion of foodstuffs and their chemical incorporation in hard tissues.
Utilization of bone material to infer nutrition is also hampered by the organic nature
of bone, which renders it highly susceptible to post-depositional contamination and
deterioration (see Ambrose 1991; Armelagos et al. 1989; Chisholm 1989; DeNiro 1985;
Pate and Hutton 1988; Lambert et al. 1985; Price 1989b; Price et al. 1992; Sandford
1993; Sillen and Sealy 1995; Sillen et al. 1989, among others). Diagenesis is the sum of
all physical, chemical and biological processes that occur in the postmortem depositional
environment to alter hard tissue composition.
Furthermore, conventional techniques adopted until very recently are characterized by
complete dissolution of hard tissues for chemical analysis. Chemical signatures either
reflect the combined composition of hard tissues in the sample, which may include post-
mortem contaminants, or they reflect very specific locations of bone that cannot be
“aged” accurately due to bone turnover. Elemental and isotopic analyses of dissolved
samples mean that archaeological human remains are lost forever.
Fortunately, these shortcomings can be overcome by analyzing the chemical
composition of teeth by laser ablation-inductively coupled plasma-mass spectrometry
(LA-ICP-MS). LA-ICP-MS is a method of multielement analysis that can sample precise

5
regions of hard tissues to infer dietary intake (see Chapter Six). In this study, such an
application is focused on enamel, which is significantly more inorganic (97-98%) and
resistant to diagenesis than dentine, cementum and bone (see Chapter Three).
The appearance and composition of teeth reflect the health and dietary status of
individuals during childhood, the period when teeth are formed. Teeth are an ideal
analytical material because they are incremental biological structures that are not
significantly remodeled after formation, unlike bone, which undergoes constant turnover.
Since they sequentially calcify at a known rate, they are invaluable to dietary studies that
seek to associate dietary intake, in the form of tissue chemistry (and intra-tooth
variation), with developmental age. Enamel, in particular, has an inherent permanent
“time axis” (Lee et al. 1999), which can be recognized microscopically as sequential
Striae of Retzius. Teeth are thus an indelible record of early development, providing
meaningful clues to a critical period of human adaptation.
Recognizing the attributes of dental material that make them ideal for chemical
methods of paleonutritional reconstruction, this study focuses on enamel elemental
composition to infer dietary behavior of young children in the past. Secondary canines
are used since they trace diet and nutrition of individuals from approximately birth to five
or six years of age, the period when such teeth develop. Specifically, this research
focuses on changes in diet during weaning, a key maturational event in human life history
that has consequences for future survival (see Chapter Five).
Chemical analysis of canine teeth will focus on the element strontium (Sr) and its
ratio to calcium (Ca) in enamel (see Chapter Four). The application of strontium to
paleodietary reconstruction is based on the element’s relative concentration in breast milk

6
and solid foods. As strontium abundance differs dramatically between breast milk (low)
and food (high), it is postulated that the process of exclusive breast milk intake to dietary
supplementation and weaning should be visible as a change in strontium, relative to
calcium (Sr/Ca) (see Chapter Five). The duration of breast milk intake, and the age at
which children are weaned, are significant predictors of future growth and development.
1.2 Cultural Context of the Investigation
All told, the teeth will be analyzed as biological testimony of the indigenous
experience during the early Spanish contact period in the Americas. Changes in
indigenous health from the 16th
century A.D. onward will be traced through the health
status of children. The health of children is a sensitive indicator of overall population
health (Goodman and Armelagos 1989) and, importantly, it is a reflection of a “group’s
ability to manage its social and physical environment” (Swedlund and Ball 1998: 195).
Dental remains from the ancient Maya site of Lamanai, northern Belize, will be
employed in this study (see Chapter Two). Lamanai is noteworthy in Maya prehistory
for its lengthy archaeological record. Culturally, health will be examined within the
historical context of Maya resistance and cultural syncretism at the site. Regarding
Lamanai, I will address the question: How is the unique nature of native resistance and
syncretism at this “frontier” post reflected in the health and nutritional status of its
inhabitants? In particular, did the political, economic and social conditions of
colonialism, in the midst of some indigenous autonomy, effect dietary and/or health
changes among Maya women and children, and can these changes be recognized in
childhood nutritional status?

7
Drawing from archaeological, osteological and ethnographic evidence, specific
concerns include: 1) Are there temporal distinctions in breastfeeding and dietary
supplementation between pre-colonial and historical periods? 2) Did Spanish control of
economic activities, i.e., demands for agricultural products and human labor, affect
household patterns and childhood nutrition? 3) Did this demand for increased labor
result in any changes in weanling diet or duration? 4) Is the heterogeneous nature of the
colonial mission population, including Maya “refugees” from other areas, reflected in
childhood diet and weaning patterns? 5) Are there distinctions in weaning age, duration
or diet that could be related to socioeconomic status or gender?
1.3 Theoretical Framework
In terms of a research paradigm, interpretations of the enamel chemistry are situated
in a biocultural anthropology framework, which is rooted in the materialism of political
economy. According to Goodman and Leatherman (1998: 19-20), five main tenets of
political economic theory form the foundations of biocultural anthropology:
1) Examining biological variation in terms of social relations (of power) through
which individuals gain access to basic resources and labor (production and
distribution); importance of social processes
2) Recognition of links between the local and the global (macro-micro
interconnections)
3) History and historical contingency are critical to understanding the direction of
social change and the biological consequences of change
4) Humans are active agents in constructing their environments
5) Ideology and knowledge, of both subjects and researchers, are essential to
understanding human action

8
These concepts frame the objectives of this investigation. Understanding the status of
indigenous health and nutrition at the nexus of Maya-Spanish contact demands an
integrated and critical biocultural approach. It recognizes that children, as the most
vulnerable members of society, are the most impacted by the interactions between
biology, culture and ecology. Thus, beyond infant diet, this investigation considers the
roles of women in Maya society; childcare practices and individual decisions regarding
infant nutrition; access to food resources; disease ecology and the cultural, political and
economic factors that constrain or determine the weaning process.
Like recent dental research by Wright and Schwarcz (1998), this study adds to the
immense potentials of teeth for understanding the health and nutrition of past
populations. Methodologically, the application of LA-ICP-MS technology to this study
offers a minimally invasive means of (micro)sampling and multielement profiling that
takes advantage of the developmental databanks that are teeth. Thus, the structural
integrity of teeth is preserved at the same time a chronological profile of hard tissue
chemistry is detailed (see Chapter Six). It is an ideal method to capture the complex
history of a critical developmental period in the human life cycle.
As such, the present study represents an evolution of paleonutritional research in two
respects: it employs new, non-destructive, micro sampling technology in the form of
Laser Ablation- Inductively Coupled Plasma-Mass Spectrometry (LA-ICP-MS) to
analyze the nature of intra-tooth variation in the chemical composition of dental hard
tissues to trace dietary transitions and life history during early childhood.

9
CHAPTER 2
THE MAYA OF LAMANAI, BELIZE
2.1 Lamanai Research History
The present application of LA-ICP-MS analysis to paleonutrition utilizes dental
remains from the ancient Maya site of Lamanai, northern Belize (Figure 2.1). Lamanai is
the archaeological focus of research because of its unique continuity from prehistoric to
historical times and the interdisciplinary nature of long-term investigations.
Lamanai, or Indian Church as it was known in Belize since the 19th
century, and
which roughly means “submerged crocodile” in Maya, was excavated under the direction
of Drs. David M. Pendergast and Elizabeth Graham of the Royal Ontario Museum
primarily between 1974 and 1986.
Extensive research includes more than 25 years of ongoing archaeological excavation
and analysis (Pendergast and Graham, various; Loten 1985; Powis 2001); osteological
analyses of disease, dental morphology and bone chemistry (Coyston et al. 1999;
Helmuth and Pendergast 1989; Lang 1990; White 1986, 1988, 1997; White and Schwarcz
1989; White et al., 1994; Wright 1990); zooarchaeological reconstruction of the faunal
assemblage (Emery 1990, 1999; Lovisek, R.O.M.); botanical studies (Lambert and
Arnason 1978, 1979); and investigations of ethnohistorical records (Graham et al. 1989;
Jones 1989).
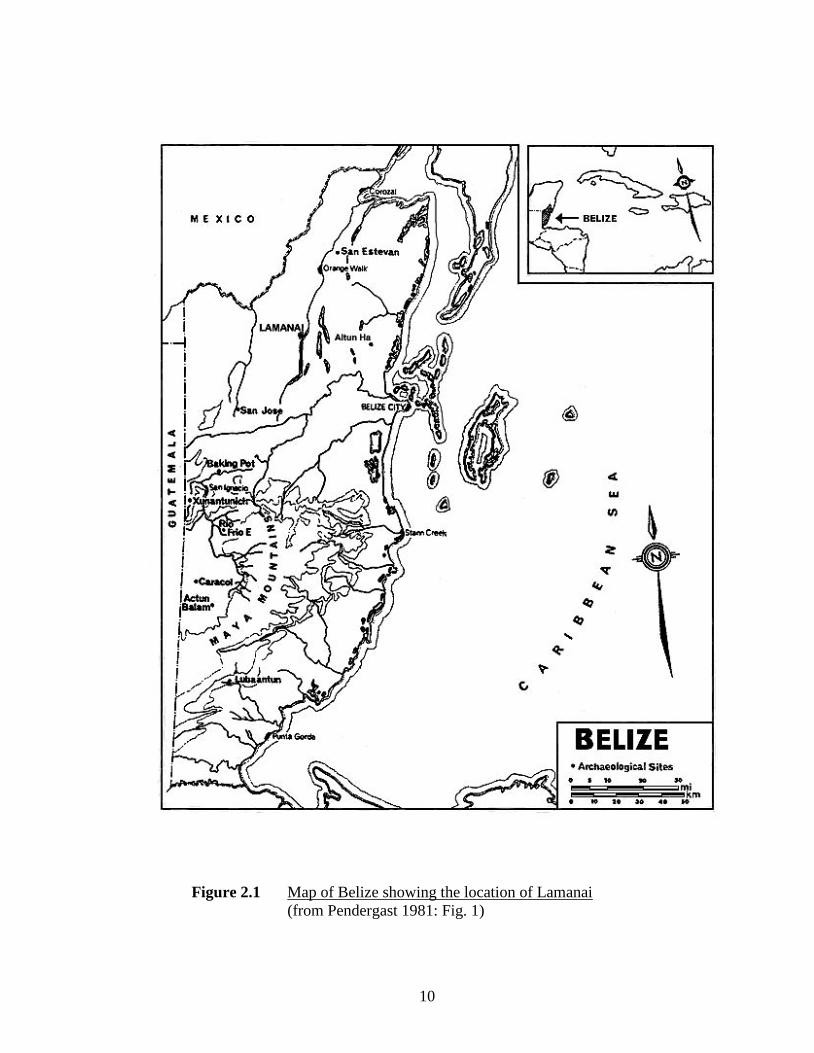
10
Figure 2.1 Map of Belize showing the location of Lamanai
(from Pendergast 1981: Fig. 1)

11
Archaeological work has resumed since 1997 by the principal directors heading the
Lamanai Archaeological Project, as well as site development carried out by the Tourism
Development Project since 2001 (Ministry of Tourism and Youth, Government of
Belize). Throughout, particular attention has been directed toward site restoration,
consolidation, and its development as a national park for tourism.
From these investigations, Lamanai emerges as an important ceremonial, trade and
administrative center in the Maya Lowlands during much of the Classic (A.D. 300-1000)
and Postclassic (A.D. 1000-1520) periods. It is noteworthy for having one of the longest
continuous settlements in the Lowlands, spanning from 1500 B.C. (or earlier) to A.D.
1675, and a dynamic archaeological record (Graham et al. 1989; Pendergast 1981, 1986,
1991a,b, 1992, 1993). The following chronology outlines the various cultural phases for
Lamanai.
Table 2.1 Lamanai chronology
Time Period Dates
Preclassic
Early 2500-1250 B.C.
Middle 1250-400 B.C.
Late 400 B.C.-A.D. 250
Classic
Early A.D. 250-400
Middle A.D. 400-650
Late A.D. 650-900
Terminal A.D. 900-1000
Postclassic
Early A.D. 1000-1100
Middle A.D. 1100-1400
Late A.D. 1400-1544
Historical A.D. 1544-1670

12
2.2 Environmental Setting
The site of Lamanai lies within the north-central coastal plain region of Belize,
Central America (Figure 2.1). It is situated at approximately 170 46’N latitude and 880
39’W longitude, while the country of Belize as a whole is located between the latitudes of
150 N and 180 N, which puts it on the fringes of the tropical (0-150 N) and subtropical
(20-300 N) latitudinal belts (Smith and Panton 1986: 48). Presently, the site is located in
the southern half of the Orange Walk District of Belize, approximately 70 km from the
Caribbean coast, within the 385 hectare Lamanai Archaeological Reserve.
The northern lowlands of Belize resemble the geology and landscape of much of the
Yucatan peninsula, with gently sloping limestone hills, calcium carbonate rich soils, and
level plains mainly under 100 meters above sea level. In the Lamanai Archaeological
Reserve, elevation averages around 30 meters above sea level. Rivers in the region are
generally slow-flowing because of the lack of gradient and often pass through badly
drained swampland (Furley and Crosbie 1976).
Lamanai falls within Thornthwaite’s (1948) Subtropical zone, or the Dry Tropical
climate zone according to the classification system of Wright and colleagues (1959: Figs.
II, VIII). The Dry Tropical zone, which encompasses all of northern Belize and most of
central Belize, has a mean annual temperature of more than 240C (750F). It generally
receives from 1500-1750 mm of rainfall per year, but not more than 2000 mm (Wright et
al. 1959).
The period between January and May has relatively dry weather, which is
characterized by less than 100 mm (4 inches) of rainfall (Furley and Crosbie 1976: 12).
Generally, the wet season occurs between June and December, but the region is prone to

13
radical fluctuations in rainfall from year to year. Thus, some years experience no real dry
season, while extended dry seasons often follow. Presently, the usual planting season for
maize, a primary staple of ancient and contemporary Maya and Latin American diets, is
April to May (Furley and Crosbie 1976).
Cyclones, in the form of tropical depressions, storms, or hurricanes, frequently
develop from the Caribbean Sea, and can drastically affect typical weather patterns
throughout Belize. Tropical storms and hurricanes are most active over Belize between
August and October, with peak occurrences during the month of September (Gibson
1983), but their number and regularity vary from year to year. Severe hurricane
conditions have caused widespread coastal flooding and alteration of portions of the coast
and offshore cays (e.g., Hurricane Hattie, 1961). While such destructive events cannot
currently be extrapolated from prehistoric contexts, weather records indicate that Belize
has experienced about three hurricane episodes, on average, every 10 years (Furley and
Crosbie 1976).
Regarding vegetation, the Lamanai area consists of several distinct types: broadleaf
forest, high bush, cohune ridge, swamp broadleaf forest (swamp [bajo] and riparian
forest), pine savanna, grasses, shrubs, tropical forest and alluvial shoreline (Lambert and
Arnason 1978; White 1986). Past and present evidence indicate that the region is rich in
tree species, some of which include mahogany, ceiba, sapodilla, breadnut, allspice,
chicle, cherry, cohune palm, cedar, gumbolimbo, copal, strangler fig, bay leaf palm, and
poisonwood, among many others (Hartshorn et al. 1984; Lambert and Arnason 1978;
Pohl 1990; Turner and Harrison 1983).

14
From an economic standpoint, approximately one third of the available tree species in
northern Belize provide fruit or leafy components that are suitable for human
consumption. Some include sapote, breadnut/ramon nut, allspice, cohune, strangler fig,
wild papaya, bay cedar, hogplum, guanacaste, Piper, and the pork-and-doughboy palm (J.
Eckenwalder, personal communication, 1996; Standley 1961).
The majority of plants around Lamanai do not contain edible components, but many
of these species are, and were, important sources of lumber, as well as ritual (copal) and
medicinal products (e.g., Acacia, bay leaf palm, bookut, cedar, copal, dogwood, Ficus,
gumbolimbo, mayflower, Mimosa, papaya, Piper, and the trumpet tree [see Roys 1931]).
2.3 The Nature of Human Settlement at Lamanai
Surveys at Lamanai between 1974-1976 located 718 structures (which now exceeds
720) within a 4.5 km2 area, or a settlement density of approximately 160 structures per
km2 (Pendergast 1981: 32). Compared to the rate of 367 structures per km
2 of habitable
terrain at Altun Ha (Pendergast 1979), and the 810 structures per km2 at wall-enclosed
Mayapan (Jones 1952), which represents the high end of Maya settlement density,
Lamanai inhabitants enjoyed relatively pleasant living conditions.
Numerous features can be recognized as having attracted settlers to Lamanai: fresh
water from the New River lagoon and New River, proximity to the Caribbean Sea, and
diverse food and trade resources. As stated above, the area is rich in vegetation diversity,
but numerous aquatic ecosystems are also close at hand: lagoon, river, estuary, coastal
reef and sea. As White (1988: 3) states, the ecological and resource diversity at Lamanai
makes it “an environmental microcosm for the entire Maya Lowlands”.

15
Archaeological evidence suggests that the site was inhabited by at least 1500 B.C.,
which is when pollen cores indicate the cultivation of maize (Pendergast, personal
communication to White, 1988). Such agriculture was achieved by several methods,
namely slash-and-burn milpa, raised fields and other irrigation techniques. Raised fields
have been identified in the northern limit of the site, but their date of origin is
questionable (Lambert et al. 1984). According to Lambert and Arnason (1979), the
production potential of the Lamanai area for maize far exceeded even the highest
population estimates, with the adoption of variable agricultural practices preventing the
likelihood of soil exhaustion (Lambert 1985).
Unlike most sites in the southern Lowlands, including important centers nearby (e.g.,
Altun Ha), Lamanai survived the Maya “Collapse” of the ninth century A.D., as indicated
by archaeological evidence of political, economic and demographic stability (Loten 1985;
Pendergast 1981, 1986, 1991a, 1992). In particular, architectural and material evidence
suggest continuity of Classic period ideology, religious belief and practices. This
stability is atypical for the Maya Lowlands at this time. While many centers were
abandoned during the Terminal Classic (AD 900-1000), after experiencing socio-
political, economic and ideological upheaval, Lamanai witnessed sustained construction
of monumental public and ceremonial architecture, as well as residential structures.
The reasons for this stability at Lamanai are numerous and interrelated and include:
1) the lakeside setting of the site, which provided numerous resources; 2) its access to the
New River, which was a busy route for trade and contact with many regions of
Mesoamerica; 3) their long-standing participation in trading networks; and 4) the moral
fortitude and personality of community leaders (Pendergast 1981, 1986, 1991a, 1992).

16
A principle factor for Lamanai’s endurance was its setting. Located on the western
shore of the New (or Dzuluinicob) River Lagoon, just south of its northern juncture with
the New River, Lamanai is situated at the head of a major riverine trade route. Presently,
the New River Lagoon is the largest body of open water in Belize.
Owing to its lakeside setting, settlement at Lamanai took on a “decidedly non-
standard settlement pattern, in which the usual arrangement of one or more ceremonial
precinct plaza groups, surrounded by zones of residential and other small structures, gives
way to a sort of massive strip development with not a single ceremonial grouping
resembling those generally encountered elsewhere” (Pendergast 1981: 32). This unusual
pattern is distinct from many lowland Maya centers, including nearby Altun Ha and San
Jose, but it served to control access of the upper reaches of the New River’s headwaters
(Pendergast 1981).
Beyond the diverse food resources of the lagoon and river (see below), Lamanai’s
location was probably more important as a means of communication with other parts of
Mesoamerica. This route connected the Northern Maya Lowlands, the Caribbean coast,
the Southern Lowland interior and southern regions of Mesoamerica to the inhabitants of
Lamanai. For instance, artifact and architectural styles and materials indicate that
Lamanai had a long history of extensive contact with foreigners from western and central
Mexico (Oaxaca), the Guatemalan highlands, the Peten, Yucatan, Honduras and Lower
Central America (Pendergast 1981, 1986, 1991a, 1992). Overall, the quantity and variety
of Late Classic and Postclassic trade goods reflect the site’s prosperity and status among
traders and merchants throughout Mesoamerica.

17
Importantly, during the Postclassic and Historical periods, the New River was the
conduit through which human migration, as well as the trade of materials and ideas,
flowed into territory controlled by Tipu (see Figure 2.3). This site in western Belize,
located on the Macal River near its junction with the Mopan River, was the political
center of the Dzuluinicob province and was a major site of native resistance during
colonial times.
Furthermore, Lamanai’s situation on the New River provided direct access to the
Caribbean Sea. Other than marine dietary resources, which were restricted to elites, the
sea also provided the inhabitants of Lamanai with a rich and varied material culture.
Based on the faunal evidence, numerous vertebrate and molluscan resources were
exploited by the Lamanai Maya throughout its occupation (Emery 1990, 1999). As at
nearby Altun Ha, marine resources such as bony fish, shark, stingray, manatee and shell
species (Spondylus, Strombus and Oliva, among many others) were available to higher
status inhabitants, and they were also important commodities involved in long-distance
trade to inland sites (Pendergast 1979).
Participation in far-reaching trade networks via the New River ensured that Lamanai
elites prospered, and this is clearly evident in the abundance of elaborate public
architecture at the site, including a 33 meter high temple pyramid as early as the Late
Preclassic (100 B.C.), and numerous equally monumental structures thereafter (see
Pendergast 1981, 1986, 1991a, 1992). Artifactual and iconographic evidence further
establish Lamanai as an important political, economic and ceremonial Lowland Maya
center for most of its history.

18
2.4 Food Resources
The diverse ecological setting of Lamanai also meant that its inhabitants had access to
significantly more varied dietary resources than were available at sites further inland.
Local resources were exploited from jungle, pine ridge, lacustrine, riverine and marine
ecosystems. The lagoon and river provided abundant resources such as thirty-one species
of freshwater fish, turtle and other water-based animals such as birds, manatee, eel,
shellfish, and snails (see White 1986: Table 1). Other local fauna include important food
resources such as deer, peccary, dog and turkey, in addition to rabbit, armadillo, large
rodents such as paca and agouti, opossum, tapir, pheasant, monkey and squirrel, among
others (Emery 1990, 1999; Hellmuth 1977; Shaw 1991; White 1986; Wing and Scudder
1991).
These resources supplemented a diet based on locally grown staples such as maize
(Zea mays), beans (Phaseolus sp.) and squash (Cucurbita sp.), which were probably
grown in nearby raised fields (Lambert and Arnason 1978, 1979; Pendergast 1991a).
Archaeobotanical evidence indicates that maize was probably produced at the level of
self-sufficiency during the Postclassic (Pendergast 1986).
A wide variety of other domesticated and wild plant foods were also available,
including chile peppers, sweet peppers, tomato, various root crops (manioc, jicama, sweet
potato) and tree crops such as avocado, cacao, cashew, coconut, fig, mango, mamey
apple, papaya, ramon, palm and guava (see Hellmuth 1977; Lentz 1999; Miksicek et al.
1991; Pohl 1990; Turner and Miksicek 1984; White 1986, 1999, among others). Cotton
and other fibers were also locally grown.

19
Ironically, despite ecological diversity, zooarchaeological data suggest that consumed
food diversity diminished significantly during the Late Postclassic and continued in this
way into the Historical period. At these times, Lamanai Maya relied increasingly on
cultivated foods and a small number of animal species, particularly riverine ones, to the
exclusion of other resources. Exploited animal resources included turtle, fish, birds,
turkey, and curassow (Emery 1990, 1999). This was supplemented with a small amount
of imported marine foodstuffs (primarily among elites), as well as imported salt
(Pendergast 1986).
2.5 The Postclassic and Historical Periods at Lamanai
In the Postclassic period (A.D. 1000-1520), Lamanai Maya shifted settlement toward
the southern limit of the Classic period site core, which was eventually abandoned
(Pendergast 1986, 1991a). The Postclassic sample of 115 burials consists of individuals
interred in Structure N10-2, a predominantly Middle Postclassic ceremonial structure in
the Postclassic central precinct, and Structure N10-4, a Middle to Late Postclassic
structure adjacent to N10-2. Male and female adults are equally represented and
subadults are likewise interred in the same context.
Based on burial context, construction and grave offerings, Structure N10-2
individuals are considered high in socioeconomic status, and because of its adjacent
proximity, Structure N10-4 burials are also likely privileged (Pendergast 1981). Hard
tissue chemical composition, reflecting dietary intake, supports the lack of status (or sex)
differentiation in the Postclassic sample (Coyston et al. 1999; White 1986). Skeletal
evidence suggests that Postclassic individuals were generally healthy (White 1986).

20
The largest number of Postclassic burials derives from Structure N10-2, which,
alongside N10-1, was the ceremonial focal point for the Terminal Classic and Postclassic
community until the 15th
or early 16th
century (Pendergast 1986). At least fifty
Postclassic burials were excavated from Str. N10-2 (see Figure 2.2).
Figure 2.2 Drawing of Structure N10-2 (Pendergast 1981: Fig. 17)
Structure N10-4 was a Classic period structure that continued in use into the
Postclassic as a burial mound, with two or more small structures atop. Forty-seven
Postclassic burials, all derived from a single construction period, were interred overlying
the remains of an Early Classic structure (Pendergast 1981). The majority of these
remains are contemporaneous with the later remains of N10-2, dating to the late 15th
to
early 16th
century A.D.
While the entire Postclassic sample is generally “privileged” in socioeconomic status
(i.e., greater access to resources and social power), as inferred by interment context and
burial assemblage, several individuals are particularly notable, namely N10-2/9, N10-
2/20 and N10-4/46 (Pendergast 1981, 1992). The wealth of burial artifacts, including
copper and gold sheet objects, distinguishes these male individuals as nobles or

21
community leaders. (Teeth from Individuals N10-2/20 and N10-4/46 are included in the
present analysis.) According to Pendergast (1991b), the primary individual of burial
N10-4/46 is probably the most important in the Postclassic group based on two accounts:
1) its very late date, of the early 16th
century (post A.D. 1525), is from a period which is
little evidenced elsewhere at the site; and 2) it is possibly the last pre-Hispanic ruler of
Lamanai.
The Historical period (AD 1544-1670) at Lamanai is notable for a Spanish presence
that interfered with aspects of Maya lifeways amidst preservation of some cultural
autonomy. Rich archaeological and ethnohistorical documentation indicate that the site
was an important frontier center of Maya resistance during the Historical period, with
components of indigenous culture existing side-by-side with Christian churches and
ideology (Jones 1989; Graham et al. 1989; Pendergast 1991b, 1993).
Unfortunately, notwithstanding historical accounts by Landa (1566), Lizana (1633),
Lopez de Cogolludo (1654, 1688), and Oviedo (1535-1547), there is difficulty detailing
all aspects of Maya life at the time of contact, due to both Spanish ignorance of
indigenous material culture, and the intentional concealment of Maya activity from
Spanish writers (Jones 1989).
As a result, reconstructing indigenous culture at that time has relied on integrating
ethnographic and historical accounts with the archaeological record. This approach is
problematic at times, with discrepancies occurring between the two “truths” (see Jones
1989; Rogers and Wilson 1993). Nevertheless, combining these complementary modes
of cultural reconstruction has been fruitful for the southeastern Maya frontier, due to
invaluable ethnohistorical documents and abundant archaeological data for the sites of

22
Figure 2.3 Map illustrating the southeastern Maya frontier, with provinces and sites
indicated (adapted from Jones 1989)

23
Lamanai and Tipu (see Figure 2.3) (Jones 1989, 1998; Jones et al. 1986; Graham 1991;
Graham et al. 1989; Pendergast 1981, 1986, 1991a, b, 1993).
Notably, however, while Tipu is well documented in historical sources, there are
scant written records of Lamanai (Jones 1989). As stated by Jones (1989), this fact is
perplexing as Lamanai has the remains of two ramada churches (structure with a pole
and palm-thatch covered nave), one more substantial than at Tipu, numerous Christian
Maya burials and domestic architecture.
The few historical references that do exist indicate that the Spanish first visited
Lamanai, which was usually spelled Lamanay, shortly after 1544 (Jones 1989;
Pendergast 1992). At this time, it was established as an encomienda (royal grant to
Spaniards for the right of tribute from native populations, and usually composed of one or
more towns) following the 1543-1544 conquest of the region (Jones 1989). Up until
then, at least the southern third of the Postclassic settlement was still functioning as a
community, alongside a satellite community near the northern boundary, but the ancient
ceremonial core remained abandoned (Pendergast 1981, 1986).
The first Christian church at Lamanai (Structure N12-11) was erected around A.D.
1545-1550, 0.75 km south of the Postclassic ceremonial center, for the relocated scattered
settlement. It was utilized until almost the end of the 16th
century (Pendergast 1991b).
Following the practice common throughout the colonized Americas, it was built over an
earlier indigenous ceremonial structure. At Lamanai, it entailed the destruction of most
of a small, fresco-decorated temple dating to the 15th
century for an earthen platform that
supported a masonry and perishable material structure. As succinctly stated by
Pendergast (1991b: 341), this structural superposition had “the eminently practical aim of

24
perpetuating precontact patterns of activity while supplanting one form of religious
practice with another”.
This original church was modest, with a nave measuring only 6 x 9 meters
(Pendergast 1991b: 342). It served a small local community that operated on the visita
(circuit-riding) system, which saw Spanish priests circulating among many different
parishes, aided by local Maya religious figures (Pendergast 1991b).
The second church was built around A.D. 1600, just north of the first church, and was
much larger than the original, with substantial masonry architecture (Graham et al. 1989)
(see Figure 2.4). Unlike the first church, architectural traits of this one were fully
European and resembled other churches in the Yucatan (Pendergast 1991b).
Figure 2.4 Remains of the second Spanish church at Lamanai
In 1637, Lamanai was listed as a reduccion town, or a compact community composed
of the forced resettlement of the native population from nearby villages, and its Maya

25
citizens paid tribute to Salamanca de Bacalar (Jones 1989). Based on tribute payment
records, a population of 72 people is suggested at this time (Archivo General de Indios,
Seville, in Jones 1989: 117, 310). By now considered a vacant encomienda made up of
“runaway Indians” from the Bacalar region, many of who were later reduced back to San
Juan Extramuros at Bacalar, it was known as a trouble-making community (Jones 1989).
The period between 1638 and 1677 was a notable period of resistance and rebellion in
the Lamanai and Tipu areas (Jones 1989). At this time, the Yucatan and southeast
periphery were especially turbulent, with increasing Maya migration to the frontiers of
Belize. Accompanying this were epidemic disease, successive famines, frontier
rebellions inspired by millennial prophecy, increasing piracy and foreign control of
logwood cutting and export, and repartimiento (forced advances of cash or goods in
return for native-produced foodstuffs, crafts or natural products) exploitation (Graham et
al. 1989; Jones 1989).
Based on ethnohistorical and archaeological evidence, it appears that unlike most pre-
Columbian Maya sites, Lamanai witnessed a dynamic Maya presence during colonial
times. As stated earlier, Lamanai’s continuity was primarily due to its geographic
location. Even during colonial times this fact ensured the Maya community’s survival.
At this time, Spanish administrative control in the Maya Lowlands was based at
Merida in the Yucatan, with regions south of Chetumal Bay practically unknown
frontiers. Even to natives of the north, the southern portion of the Maya Lowlands was
considered frontier territory and the land of foreigners. The New River in northern
Belize was often called Dzuluinicob in ancient times, meaning “foreign people” in Maya
(Lopez de Cogolludo, 1688 [5th
ed., 1971]), and the same term also applied to the region

26
south of the New River Lagoon in central and west-central Belize (Jones 1989). As a
frontier, numerous Maya communities existed along the New and Belize Rivers outside
the control of Spanish forces.
The southeastern provinces of the Maya “frontier” were a dynamic breeding ground
for both active resistance and cultural syncretism, and it was a transitional zone that
“connected the native world of the colonized Yucatan to the north with the genuinely
independent Maya world of the Itzas of the Peten” (Jones 1989: 123). Both people and
ideas were safely hidden here and, consequently, Spanish attempts to pacify these
provinces (e.g., establishment of the villa at Salamanca de Bacalar) were geared toward
monitoring indigenous resistance movements, potential rebellions and Maya runaways
from the north, rather than for the purposes of collecting tribute (Jones 1989).
Besides being in the Maya “frontier”, Lamanai was strategically placed midway along
a major riverine route between Bacalar, the important Spanish administrative center
located near Chetumal Bay that controlled the ancient province of Chetumal, and Tipu,
the political heart of the Dzuluinicob province (Graham et al. 1989; Jones 1989) (see
Figure 2.3). Tipu is located in a small valley in the foothills of the Maya Mountains and
was the last of a series of visita missions extending south-southwest from the villa of
Salamanca de Bacalar (located more than 200 km away by river and land) (Figure 2.3).
The Dzuluinicob province was composed of indigenous Yucatec Maya speakers and it
became the primary center of Maya resistance to colonial rule (Jones 1989).
Maintaining its pre-Columbian past, Lamanai acted as an important hub for trade and
communications during the 16th
and 17th
centuries (Graham et al. 1989; Pendergast
1991b, 1993). However, in contrast to the relative stability of Tipu, Lamanai, as a

27
reduction center for runaways from the Bacalar region, experienced more population flux
and instability (Graham et al. 1989; Jones 1989). Despite its proximity to the
administrative center of Bacalar though, which made it vulnerable to Spanish control,
native Maya material culture at Lamanai was well represented.
Architecturally, there was a noticeable change from a pre-Columbian tradition to
Spanish influenced masonry at Lamanai, albeit on a limited scale, but there was no
significant alteration to the pre-Hispanic settlement pattern, which was already restricted
by the lakeside location (Graham et al. 1989; Pendergast 1981, 1986, 1991a, b).
Compared to many other Spanish-controlled colonial centers, which generally exhibited
European town plans, the situation at Lamanai was unique and likely reflects Spanish
efforts to attract Maya refugees (Graham et al. 1989; Pendergast 1991b).
In terms of small-scale material culture it appears that there was also general
continuity of artifacts. To a limited extent, some types of European artifacts were
accepted, but the material culture of colonial period Maya remained surprisingly
traditional and significantly more numerous than Spanish imports. This is particularly
visible in ceramics and lithic tool production (Graham 1991; Graham et al. 1989;
Pendergast 1991b). In total, Spanish material culture was “never more than an overlay on
that of the Maya” (Graham et al. 1989: 1258).
The European imports that appear in the archaeological record at Lamanai most often
occur in burial contexts and they were most certainly material symbols of socio-economic
status (Graham et al. 1989). Historical documents suggest that gifts such as glass beads,
rosaries, axes, machetes, knives, needles, silver and glass earrings and necklaces were
given by priests to encourage religious conversion (Villagutierre Soto-Mayor 1983).

28
In response to religious proselytizing, which, in addition to the seizure of land and
resources, was the main goal of Spanish colonists, Maya groups throughout the Lowlands
and Highlands resisted. Other than material cultural stability, important insight into
various forms of community resistance can be drawn from ethnohistorical texts and
demographic records. Such evidence suggests that in response to heavy tribute and
taxation demands, cultural and religious persecution, economic constraints, brutality,
massacres, famine, and disease, many Maya protested Spanish hegemony by violent
protest and out-migration (Clendinnen 1987; Farriss 1984, 1993; Jones 1977, 1989, 1998;
Landa in Tozzer 1941; Restall 1997; Varner and Varner 1983).
Native resistance also came in the form of upholding cultural traditions, as well as
cultural syncretism with European customs. Language, a fundamental symbol of culture
and group identity, was an important mechanism through which colonial period Maya
resisted Spanish influence. Spanish terms were adopted, but the general rule was not to
borrow Spanish words if there was an equally appropriate Maya term (Farriss 1984). In
contexts involving discourse between Maya and Spanish, it was usually Yucatec Maya
that was employed, through the use of interpreters (Farriss 1984).
The minimal extent of Spanish language conversion during colonial times is at odds
with the enormous efforts undertaken to instill Christianity. This may be attributed to the
moral fortitude of Maya communities to maintain language above all else, as a clear
symbol of identity, but it was also due to the laxity of Spanish attempts to impart their
language (Archivo General de la Nacion, 1790-1805, in Farriss 1984).
On the other hand, Maya culture during the 16th
and 17th
centuries also reflects a great
deal of cultural syncretism with Spanish traditions, namely in the form of religious

29
acceptance. The main features of Maya/Christian religious syncretism include:
integration of the vast Maya pantheon with Christian saints (represented physically as
figurines or censers); adoption of Christian holy days such as Good Friday and Easter,
which were secondary in importance compared to the syncretic All Souls’ Day (“a
merger between the Catholic concept of restless souls in purgatory and Maya notions of
the afterlife” [Farriss 1984]); expression of sacred Christian imagery within native
contexts (e.g., crucifixes upon altars surrounded by offerings); and the incorporation of
copal incense, ritual balche drinks, offerings of maize, turkey and deer, pre-Columbian
songs, dances and traditional instruments with Christian rituals (Clendinnen 1987; Farriss
1984, 1993; Jones 1977, 1989, 1998; Miller and Farriss 1979; among others).
It is obvious in reading the historic accounts of Spanish priests, leaders and scribes
that despite efforts to Christianize native communities, and despite the appearance of
conversion to Christianity, there was an active preservation of traditional Maya religious
practices. Entradas and other passages through Maya communities frequently
encountered native “idols” in houses and temples, even practically “under the noses of
clerics”, and as a typical example, Juan Garzon (1568, cited in Jones 1989: 49) stated that
they found “so many idols that they couldn’t easily be counted”.
These “idols” were anthropomorphic and zoomorphic censers, whistles and figurines
that likely had multiple functions and various religious and non-religious significance.
They were commonly found in churches, caves, and shrines and recesses of Maya homes
(Jones 1989; Farriss 1984). At both Tipu and Lamanai, such pre-Columbian-style
effigies were found in refuse deposits and offerings in colonial period buildings,
testament to the fact that native religion was not entirely displaced by Christianity

30
(Graham 1991; Graham et al. 1989; Pendergast 1991). In one case at Lamanai, a small
bat effigy vessel was deposited at the base of a ruined Maya temple prior to construction
of the overlying church platform (Pendergast 1991b: Fig. 16-3b). Besides being an
expression of resistance, it may represent an attempt to “appease(ing) the old gods for the
destruction of their temple” (Pendergast 1991b: 343).
The Lamanai burials themselves attest to the degree to which Maya selectively
adopted features of Christianity. This sample represents the largest skeletal assemblage
from the site, with approximately 179 individuals identified. While sex distribution is
generally equal, there are more Historical period subadults compared to the Postclassic
sample (White et al. 1994).
The first cemetery was located just south of the first church and contained the remains
of more than 25 individuals, who were uniformly adults, suggesting that a separate
children’s cemetery is located elsewhere (Pendergast 1991b).
The second graveyard and church (YDL-85), from which dental remains have been
collected for this analysis, contained individuals of mixed age and sex that are dated to
A.D. 1570-1640. It is believed that both graveyards contain exclusively Maya
individuals (Pendergast 1981, 1986). Owing to their shared circumstances as “reduced”
Maya under Spanish authority, and based on the burial artifacts and bone chemical
evidence (Coyston et al. 1999; White 1986), they are considered similar in
socioeconomic status.
However, for some Maya, this seeming “equality” in death only reflects relative
social circumstances prior to their demise. The fluid nature of population movement and
interactions in the southeastern provinces after Spanish contact suggests that colonial

31
Lamanai was a diverse settlement. Until further analysis is undertaken, the nature of the
archaeological and ethnohistorical records precludes identification of individual Maya in
the colonial cemetery, but it is likely that the Historical assemblage includes: 1)
“commoner” Maya native to Lamanai; 2) members of formerly elite Postclassic
(Lamanai) families; and 3) Maya immigrants from the Yucatan and other adjacent areas,
of varied socioeconomic background. This heterogeneous colonial population likely
experienced dissimilar life experiences prior to arrival at Lamanai, especially childhood
diet patterns and health histories. Such variations may reflect differences in access to
food resources and social relations (power), epidemic disease, famine, re-settlement and
violence associated with the “conquest”.
Despite the Spanish failure to completely stifle Maya beliefs and rituals, there was,
nevertheless, a strict adherence to an entirely Christian mode of burial in the Lamanai
cemeteries at the end of the 16th
century (Graham 1991; Jones et al. 1986; Pendergast
1991b). At Tipu, even after Spanish abandonment of the site (post 1701), the Christian
interment pattern of supine positioning, with the head to the west facing east and with the
hands drawn over the stomach or chest, continued to be practiced in the native Maya
cemetery (Graham 1991; Graham et al. 1989).
To explain this acceptance of Christianity, traditional interpretations, including those
of priests at the time, usually branded the Maya as opportunistic: people who simply
adopted enough Christianity to deceive their oppressors (out of necessity for survival),
without believing what they were taught (see Jones 1989; Graham 1991). At Lamanai,
this is reflected in several caches of Maya religious figurines, which were deposited into
the church platform structure unbeknownst to Spanish clerics (Pendergast 1991b).

32
However, it has become increasingly apparent that Christianity was not “simply a
veneer on a pre-Columbian base” (Graham 1991: 331). Burial evidence, the extent of
religious syncretism of sacred icons, images and rituals, as well as archaeological
indications of regular church maintenance (at Tipu), suggest that despite the preservation
of fundamental Maya beliefs, many colonial period Maya were not superficial, but
practicing, Christians (Graham 1991). Colonial period Maya may have rejected Spanish
autonomy and imperialism, particularly as it was manifest in the clergy, but it appears
that they were active followers of a religion that shared fundamental precepts such as the
heavens, an underworld, an afterlife and multiple religious figures (“saints”) (Farriss
1984).
In total, the syncretism of Christianity with the polytheistic, animatistic Maya religion
demonstrates great resilience in the face of dominating forces. This adaptive ability can
be attributed to the experience of Maya communities with foreigners throughout their
long history, particularly Mexican traders. Time and time again, Northern Lowland
Maya in particular, including Lamanai, encountered visitors who imparted linguistic,
architectural, material cultural and religious influences. In this way, it was a
Mesoamerican tradition that conquering groups introduced new religious cults that
inevitably coexisted with indigenous beliefs (Farriss 1984, 1993). Unfortunately, the root
of much tension between Spanish and Maya lay in this very fact, which was at odds with
the exclusivity of Christianity, a religion that demanded complete dedication, i.e., the
total abandonment of Maya beliefs and deities (Farriss 1993).
Inevitably, the Maya would prevail at Lamanai. Despite being practitioners of
Christianity, according to visiting Franciscan friars Fuensalida and Orbita, the local

33
community destroyed and burnt the church between 1640 and 1641 (Pendergast 1981;
Jones 1989). This rebellion effectively ended Spanish control of Lamanai, as well as
most of Belize, until 1695 (Jones 1989).
Free from the rule of Spanish officials, however, Lamanai Maya continued to revere
the church as hallowed ground, even erecting a stela and traditional Maya altar in the
entrance and nave areas and maintaining ritual activity (Graham et al. 1989; Pendergast
1991b). As Pendergast (1991b: 352) recognizes, “the events that followed the burning of
the church show that although the Spaniards had clearly failed to Hispanicize the Maya,
they had succeeded, probably more fully than they realized, in Christianizing them”.
Slightly later, and until the end of the 17th
century (around 1675), there was limited
and intermittent settlement by one or more families in the masonry chancel of the second
church (Pendergast 1986), as well as south of the main historical period community
(Graham et al. 1989). Thereafter, the site remained abandoned until British colonists
established a short-lived sugar mill between the 1850’s to the early 1880’s.
2.6 The Biological Consequences of Maya-Spanish Contact at Lamanai
In retrospect, the acceptance of Christianity by indigenous Maya, and their
“conquest” by the Spanish in general, can be tied to the psychological and physiological
climate of the time. Prior to contact, widely known Maya prophecies foretold of
impending foreign invasion and catastrophe (Landa in Tozzer 1941; Roys 1933, 1954).
Upon subjugation by Spanish conquistadors, foreign microbes aided in the eventual
decimation of many indigenous groups, primarily affecting vulnerable young and elderly
individuals.

34
As elsewhere in the New World, Maya populations were substantially diminished
after Spanish contact. Unfortunately, this era was also preceded by a series of natural
disasters in the Yucatan that would weaken indigenous survival thereafter. In this way,
the Maya Katun Prophecies of the Books of Chilam Balam refute the notion of pristine
health among indigenous groups before Spanish contact (see Roys 1933, 1954). In
particular, a series of famines and epidemics, most likely pneumonia, greatly affected
Maya demographics around A.D. 1400 (Landa in Tozzer 1941). These events followed
record droughts and locust swarms that devastated maize crops (Roys 1933, 1954).
Epidemic infectious diseases are thought to have been introduced into the Maya area
in 1517, when Spanish soldiers and their African slaves, both probably stricken with
smallpox, arrived in the Yucatan from Cuba (Clendinnen 1987). Between then and the
founding of Merida in 1542 and Bacalar and Valladolid in 1544 (that is, prior to the
Spanish arrival at Lamanai), the Maya population in the Yucatan declined from a
conservative figure of at least 800,000 to about 250,000 survivors (Clendinnen 1987;
Farriss 1984).
Notable epidemics in the Yucatan occurred in 1566, 1569-1570, 1575-1576, 1609
(typhus), 1648-1650 (yellow fever, smallpox), 1659 (measles, smallpox), 1692 –1693 and
1699 (Farriss 1984: 61). Famines in the same area are documented for 1535-1541, 1564,
1571-1572, 1575-1576, 1604, 1618, 1627-1631, 1650-1653, 1692-1693, and 1700
(Farriss 1984).
Worse yet, in the rest of Central America beyond the Yucatan and Belize, a
population loss of almost 90% has been estimated for the period between 1520 and 1580
(Gerhard 1979; Lovell 1985). In Guatemala, The Annals of the Cakchiquels document

35
three major epidemics that depleted native populations after the ‘Great Revolt’ of 1498.
In 1523, 1559-1562 and 1576, entire towns were wiped out by infectious disease,
documentation of which suggests death by influenza (McBryde 1940), measles or
smallpox (Cook 1998).
All told, Old World epidemics of smallpox, measles, influenza, bubonic and
pneumonic plague, yellow fever, typhus and cholera were primarily responsible for
native deaths by infectious disease in the post-Columbian New World (see Cook 1998;
Dobyns 1983; Denevan 1992; Verano and Ubelaker 1992, among others). Three
introduced parasitic infections that could have been the greatest potential contributors to
debilitating anemia include dysentery (Manter 1967; Schmidt and Roberts 1981), malaria
and hookworm (Bruce-Chwatt 1965; Dunn 1965; Friedlander 1977).
Unlike the intensively colonized Yucatan, however, in the southeastern Maya
frontier, early records indicate that political territories remained viable during early
colonial times despite the economic and demographic collapse (Jones 1989).
Nonetheless, the frontier provinces also suffered from epidemics of smallpox, yellow
fever and malaria (see Cook 1998; Farriss 1984; Gerhard 1979; Jones 1989).
In the case of Lamanai, the site’s location put it on a route of potential rapid
transmission of disease. As a frontier for runaways and an outpost in the visita system, it
was likely the refuge of fleeing Maya infected with disease (Pendergast 1991b). For
instance, at Tipu, during the “great general famine” of 1647 to 1650 in the Yucatan,
which followed one of the worst smallpox epidemics in the peninsula’s history, many
Maya migrated to the site in search of food and shelter (Jones 1989).

36
Hard tissue evidence reflects the deterioration of population health in the early
Historical period. For instance, enamel histology, in the form of defective Wilson bands,
indicates that Lamanai Maya suffered more severe, acute, episodic stresses during
colonial times (Wright 1990). Wilson bands appear as darkened bands of enamel in SEM
sections and represent brief periods of ameloblastic growth disruption (Rose 1977, 1979).
The Lamanai historical sample is distinguished by three times more Wilson bands
than Postclassic teeth (Wright 1990). Overwhelmingly, they occur in Lamanai teeth at
the time of weaning. (Microdefect evidence does not reflect a universal colonial
experience however. Historical Maya at Tipu actually have less Wilson bands on average
than pre-contact Maya [Danforth 1989]. This likely reflects differing socio-political
climates: Tipu was not a “reduced” community like Lamanai and was only marginally
controlled by Spanish authorities [Cohen et al. 1994; Graham et al. 1989].)
The mean number of shallow enamel hypoplasias per Lamanai individual is also
significantly higher in Historical individuals compared to earlier periods, and such dental
defects also peak at the time of infant weaning (Wright 1990). The higher rate of such
shallow defects reflect chronic, low-grade stress, and contrasts White’s (1986) findings
for more severe enamel hypoplasias, which are not significantly different from earlier
periods (nor differentiated by age or sex).
Together, the prevalence of Wilson bands and enamel hypoplasia indicate the co-
existence of different kinds of stress in early childhood: brief episodes of severe stress
within periods of chronic low-level health disturbance (Wright 1990). The nature of
Wilson bands is thought not to reflect the epidemics that were recorded ethnohistorically
elsewhere, but rather, lesser childhood developmental disturbances that were survived.

37
The sources of such short-term, acute, ill health are thought to be malaria and Old World
parasitic infections (Wright 1990).
Skeletal evidence of anemia or infectious disease, in the form of porotic hyperostosis,
also indicates that colonial period Maya of both sexes experienced significantly greater
physiological stress (17.0% of Historical Lamanai Maya adults, compared to 9.0% of
Postclassic adults) (White 1986; White et al. 1994). At Lamanai, anemic conditions,
particularly among women and children, might have arisen from several interrelated
factors, namely helminthic or other parasitic infections, heavy reliance on an iron-poor
maize diet, and excessive fluid loss in a tropical climate (White et al. 1994). (Additional
risk factors for women, such as blood loss from menstruation and increased iron needs
with parturition, were postulated for sex differences in porotic hyperostosis prevalence
among adults, but no distinctions were observed [White 1986].)
European diseases such as dysentery, malaria and hookworm could have been the
major causes of anemia and porotic hyperostosis (White et al. 1994). Such skeletal
evidence supports an epidemiological climate of chronic stress in Historical times. The
chronic nature of porotic hyperostosis indicates that either children were increasingly
subjected to infectious disease or that there was a greater likelihood of survival from
early illness (White et al. 1994). Importantly, White and colleagues (1994) believe that
the combined skeletal and dental data reflect a common weaning age, between two and
six years, and the period of highest risk for infectious and parasitic diseases among
children.
Due to the pathogenic attack, it is likely that Maya at the time of contact were also
victims of psychological trauma (despair, suicide) that further eroded physiological well-

38
being and survival (following Blakey 1994; Locke and Hornig-Rohan 1983; Neel 1977).
One way populations respond to witnessing the mass deaths of young and old include
reduced fecundity (see Stannard 1991). Furthermore, loss of older individuals is
particularly detrimental to cultural continuity, since elders are the primary bearers of oral
tradition, i.e., mythology and religion.
Despite escaping death by infectious disease, some survivors still faced other
invisible adversaries. In particular, Maya women, like others elsewhere, were
incapacitated by sexually transmitted diseases. These infections, in addition to smallpox
and tuberculosis, often result in female infertility among survivors (Stannard 1990, 1991;
among others). As elsewhere in the New World, it was both increased mortality rates and
reduced birth rates that led to substantial declines in indigenous population after Spanish
contact.
Numerous lines of evidence offer insight into the nature of Maya diet at Lamanai
after Spanish contact. In general, reduced communities such as Lamanai, which lack the
stability of large established enduring communities, are vulnerable to inconsistent and
unreliable agricultural production (Graham et al. 1989; Pendergast 1991b). This is due to
the fact that reducciones impose heavy burdens on the subsistence activities of the often-
smaller permanent population (Pendergast 1991b: 345).
As a result of disruptions in subsistence practice, as indicated by faunal and floral
data, it has been thought that diet quality may have declined after Spanish contact
(Pendergast 1991b). In particular, the instability of reduced settlements may have
negatively affected agricultural production and diminished per capita intake of protein

39
(i.e., hunting success) (Pendergast 1991b). Such dietary disruptions might account for
some decline in health during the historical period.
Ethnohistorical records shed light on the diet of Maya from the Cholti-Lacandon area
south of Lamanai (Hellmuth 1977; Thompson 1938) and those from the Yucatan (Landa
1566, in Tozzer 1941; Martinez Hernandez, in Marcus 1982). In addition, the Maya
codices and the sacred Quiche Maya text, the Popul Vuh, are invaluable documents. All
such sources point to the sacred role of maize in human creation and subsistence. As a
staple, its cultivation dictated daily activities and resonated throughout all aspects of
Maya culture.
By the time of Spanish contact, however, it appears that most forms of intensive
maize agriculture, i.e., raised fields, terracing, hydraulic agriculture, drained fields and
other irrigation systems, were abandoned, with only traditional slash and burn (milpa) or
swidden cultivation of mixed crops utilized (Marcus 1982; Tozzer 1941). At the same
time, maize shortages were recorded throughout the Yucatan, as well as regions of the
Lowlands toward the Gulf of Honduras and the Lacandon area of Chiapas, Mexico
(White 1986).
Exacerbating the natural disasters that diminished maize production (drought, locusts,
crop disease), Spanish forces also held campaigns to intentionally destroy fields and
homes (Archivo General de Centro America 1937, in Hellmuth 1977). Once reduced to
Spanish-controlled territory, Maya were forced to increase maize production. In the early
18th
century, such intensified cultivation was necessitated by imposed taxation, export
demands and personal consumption (Hellmuth 1977). As a result of crop failure and
purposeful destruction, sometimes by the Maya themselves, many Maya also relied on

40
root crop (jicama) (Roys 1933) and breadnut (ramon) consumption (Puleston 1982),
which were considered either poisonous or unwholesome by the Spanish (Roys 1933).
Nevertheless, bone chemical evidence suggests that dietary quality did not
significantly change from the Postclassic to Historical period at Lamanai, with continued
high maize dependency and supplementation with land herbivores and aquatic resources
(White 1986; White and Schwarcz 1989). Compared to the Classic period, however,
Postclassic and colonial faunal diversity was reduced. Faunal evidence indicates
continued heavy consumption of large turtles (Dermatemys), fish and birds (turkey,
curassow) for all social strata (Pendergast 1991b: 345), but this is accompanied by a
reduction in large mammal (peccary, deer) exploitation (Emery 1999).
The Lamanai dietary evidence, indicating dietary stability from pre-contact to post-
contact times, suggests that while cultural changes were indisputably significant to Maya
lifeways, it was not sufficient enough to alter dietary and agricultural practices in the
early Historical period (White et al. 1994). [Still, as reflected in enamel Wilson bands,
the arrival of Old World infectious diseases was enough to change epidemiological
conditions among indigenous Maya (White et al. 1994; Wright 1990).]
2.7 Bone Chemical Evidence of Diet at Lamanai
Advantageously, bone chemical analyses by White (1986) provide a hard tissue
comparison for LA-ICP-MS results. Lamanai bone chemistry indicates that from the
Preclassic to Early Classic period, strontium levels were decreasing, suggesting increased
maize intake, which comprised 50% of the total diet. This trend eventually reverses
during the Late to Terminal Classic when strontium levels rise, pointing to increased

41
consumption of other, non-maize, foods as important dietary sources. At this time, maize
consumption declined to 37% of the diet (White 1986: 297).
In the Postclassic, this trend reverses, with bone isotopic signatures indicating that
maize consumption at Lamanai reached its highest level, comprising two-thirds or more
(up to 70%) of the diet (White 1986, 1988; White and Schwarcz 1989). In terms of
elemental composition, fresh maize (and especially alkali-processed maize) is notable for
having very low strontium levels (see Chapter Four). In Mesoamerica, maize was, and
continues to be, widely consumed after alkali processing with calcium hydroxide (lime),
which improves nutritional quality and facilitates absorption (Katz et al. 1974). The
effect of maize consumption on enamel element composition during infancy is discussed
in Chapters Four, Five and Seven.
Postclassic evidence of high maize consumption is accompanied by bone chemical
and faunal evidence that suggest minimal exploitation of available lagoon resources and
reliance on fewer animal species (fish, turkey, curassow) (Emery 1990, 1999).
After Spanish contact, 15
N and 13
C analyses of bone suggest that Maya dietary
habits, and presumably agricultural practices, were minimally affected (White 1986).
Isotopic values of historical period remains suggest that maize consumption continued to
dominate 60-70% of the diet (White 1986), which mirrors ethnographic observations of
contemporary Maya diet (Benedict and Steggerda 1937).
At this time, as previously in the Postclassic, children were consuming the same high
level of maize as adults and were likely surviving on diets marginal for protein and iron
(White 1986; White and Schwarcz 1989). As White (1986) notes, such high intake of
protein-deficient maize would have had significant repercussions for children, whose

42
protein requirements are higher than those of adults. In fact, children cannot consume
enough maize to meet their protein demands without adequate supplementation (Behar
1968). Combined with the fact that children are rarely weaned onto a diet containing
meat, Lamanai children likely suffered from illnesses related to protein calorie
malnutrition and iron deficiency anemia (White 1986, 1988). Indeed, this is supported by
skeletal and dental pathologies such as porotic hyperostosis and enamel Wilson bands.
Overall, Postclassic and Historical samples indicate the same level of heavy maize
reliance, with protein supplementation from deer, peccary, fish, turtle and seasonal
exploitation of freshwater mollusks (Emery 1986, 1999; White and Schwarcz 1989).
Dental evidence, in the form of high caries incidence and substantial calculus deposits,
also support the conclusion of dietary stability (White 1986; White et al. 1994). In
White’s (1986) opinion, such evidence refutes the notion that Spanish demands for
increased labor, including that of women, impacted Maya subsistence activities.
2.8 The Repercussions of Maya-Spanish Contact on Infant Nutrition
The myriad dynamic processes that characterize Maya-Spanish contact, i.e.,
resistance, syncretism and transculturation, ecological, biological and cultural
devastation, undoubtedly had numerous consequences on children’s health. Even though
subsistence strategies at Lamanai were not significantly affected, Spanish demands for
increased labor may have affected maternal care and infant health, and this is an
important subject of inquiry in this study.
In ancient times (e.g., Postclassic), it was likely that Maya mothers breastfed their
infants for periods of up to four years (see Chapter Five). While supplementation with

43
non-breast milk foods likely began after six months of age, due to diminished nutritional
quality of breast milk, prolonged nursing is indicated by ethnohistorical sources (Tozzer
1941).
With Spanish colonization, it is postulated that socio-politico-economic factors
altered infant feeding patterns, with dire consequences for children. Historical
documents reveal that Spanish extortions of tribute from Maya men and women greatly
exceeded the legal limits of the colonial system (Graham et al. 1989: 1255). Besides
textiles, agricultural products were in significant demand (Clendinnen 1982; Graham et
al. 1989; Restall 1995). Spaniards took up residence in Maya towns to oversee
collections, but local Maya officials primarily organized the political, economic and
religious administrations alongside Spanish authorities (Graham et al. 1989; Pendergast
1991b). Lamanai and Tipu Maya themselves participated in the exchange of European
products throughout the Yucatan peninsula, which involved Spanish priests, secular
authorities, Maya merchants and Maya migrating from the Yucatan to the southeastern
frontier (Graham et al. 1989).
In this new, colonial, socio-economic climate, the survival of Lamanai children was
compromised. A significant factor may have been reduced lactation and earlier weaning
age resulting from Spanish demands on female labor for cotton trade goods (see
Clendinnen 1982; Restall 1995; White 1986). Such demands on women’s time and
bodies may have undermined childcare practices and further impacted on the health and
survival of infants who were increasingly stressed in historical times, according to bone
and dental evidence (White et al. 1994; Wright 1990). Changes in weaning behavior can
be recognized in the strontium composition of secondary canine enamel, which reflects

44
relative breast milk intake and food supplementation during infancy and early childhood
(see Chapter Five).
On the other hand, native resistance to colonial conditions may have sustained
traditional childrearing practices. It is possible that Maya women resisted Spanish
demands on their time by maintaining intensive childcare practices such as prolonged
breastfeeding. This continuity would be an extension of the preservation of traditional
Maya subsistence overall, which is indicated by bone chemical evidence (White 1986).
In this case, enamel strontium would continue to indicate a prolonged, gradual, reduction
in breast milk intake accompanied by increasing food supplementation.
As the most vulnerable members of a society, children’s health and nutrition are
impacted by changes in almost every aspect of their social and physical environment. At
Lamanai, the cultural, physical and epidemiological upheaval of Spanish colonization
provides a dynamic backdrop to observe the physiological consequences on indigenous
populations. Importantly, teeth provide reliable hard tissue records of an individual’s diet
and nutrition during childhood and can document invaluable life history during such
turbulent historic times. The applicability of dental tissue analysis to paleonutritional and
archaeological reconstruction is discussed in Chapter Three.
Finally, as will be discussed in Chapter Five, it is clear that the nature of infant food
supplementation, breast milk consumption and weaning has significant ramifications for
adult survival and population health status. The effect of colonization on indigenous
infant feeding is thus a vital question in reconstructions of the Maya past.

45
CHAPTER 3
DENTAL HARD TISSUES
AND THEIR APPLICATION IN PALEONUTRITIONAL RESEARCH
3.1 Dental Anthropology in Archaeological and Paleonutritional Reconstruction
For as long as researchers have analyzed human remains to understand the past, teeth
have proven to be invaluable pieces of evidence. More so than bone, which is prone to
physical and chemical degradation, teeth can provide a wealth of biological information
that far exceeds their relative proportion in the human skeleton. As evidenced in the
literature, dental anthropology maintains a significant role within physical anthropology
and archaeology (e.g., Alt et al. 1998; Brothwell 1963; Cruwys and Foley 1986;
Goodman and Capasso 1992; Hillson 1986, 1996; Kelley and Larsen 1991; Moggi-
Cecchi 1995; Scott and Turner 1997).
Teeth can shed light on three major areas of archaeological inquiry. These research
fields are equally assisted by both the macroscopic nature of teeth, as well as the inherent
chemical composition of dental tissues. The first concerns the strong genetic basis of
dental growth and development, which renders teeth an important marker of genetic
affinity (‘biodistance’) and population dynamics (e.g., Hanihara 1979; Hillson 1996;
Jacobi 2003; Lang 1990; Lukacs 1989; Lukacs and Hemphill 1991, 1993; Turner et al.
1991; Scott and Turner 1997). Studies have examined metric and non-metric dental traits
to reconstruct evolutionary history; the nature of group boundaries and movement of

46
individuals; as well as the role of population structure on ancient health and nutrition (see
Larsen 1997).
Chemically, distinctions in dental isotopic signatures (e.g., strontium and lead) related
to geological environment can also be used to trace population movements and origins
(Budd et al. 1996; Sealy et al. 1995; Grupe 1995; Grupe et al. 1997; Gulson et al. 1997;
Montgomery et al. 1998; Price et al. 1994, 2001; Prohaska et al. 2002; Reid et al. 2001).
This is an area of significant advancement in archaeology, as well as modern biological
and forensic science (e.g., Tremlett 2003).
Teeth also reflect general health status during dental development. Beyond indicating
the age at death, an individual’s health, hygiene and nutritional history can be indicated
by pathological conditions such as enamel hypoplasia, hypocalcification, caries,
periodontitis, abscesses and Wilson bands, in addition to non-pathological calculus
deposits (Cohen and Armelagos 1984; Danforth 1989; Goodman and Capasso 1992;
Goodman and Rose 1990; Lukacs 1992; Marks and Rose 1985; Rose 1977; Wright 1990;
among many others).
Additionally, retardation of growth and development due to environmental pollutant
exposure (e.g., lead) during childhood can be discerned from dental chemistry (Budd et
al. 1996, 1998; Evans et al. 1994; Fergusson and Purchase 1987; Goodman et al. 2003;
Outridge et al. 1995; Sharon 1988; Tunstall and Amarasiriwardena 2002).
The third, and most significant, contribution teeth provide for archaeological
reconstruction is paleodietary inference. As the fundamental mechanism directly
involved with mastication, teeth provide dietary biographies at two levels: macroscopic
evidence associated with food intake and inherent chemical composition. Macroscopic

47
indicators such as caries and calculus give a generalized account of relative carbohydrate
(e.g., maize) consumption. Likewise, dental morphology, attrition and wear patterns
provide general indications of dietary components and food preparation techniques
(Brace 1995; Brace and Mahler 1971; Cohen and Armelagos 1984; Grine and Kay 1988;
Harmon and Rose 1988; Lalueza Fox et al. 1996; Lukacs and Pal 1993; Molnar 1971;
Molnar and Molnar 1990; Powell 1985; Teaford 1991).
In contrast, the chemical composition of teeth provides direct evidence of food intake.
Chemical analyses offer both qualitative and quantitative inferences of paleodiet and
represent an evolution of dental anthropology in paleonutritional research. As mentioned
in Chapter One, the assumption that hard tissue chemistry accurately reflects the
chemical composition of consumed plants and animals is central to its application in
paleodietary research.
The analytical chemistry methods used to evaluate hard tissue chemistry and
paleodiet have traditionally involved bone collagen and bone mineral using mass
spectrometric (atomic absorption spectroscopy [AAS]), X-ray fluorescence (XRF), and
neutron activation techniques (see Curzon and Featherstone 1983; Gilbert and Mielke
1985; Price 1989a; Sandford 1993a). Other methods of hard tissue analysis include
isotope dilution-thermal ionization mass spectrometry (ID-TIMS) and proton induced X-
ray emission (PIXE) (Ahlberg and Akelsson 1976; Chaudhri and Ainsworth 1981;
Johansson et al. 1995; Lane and Peach 1997).
For trace element analysis of hard tissues, X-ray fluorescence and atomic absorption
spectroscopy have been the most widely used, with AA more popular due to its
availability and operation ease. However, AA analysis is hindered by its inability to

48
quantify multiple elements at the same time. As a result, the development of inductively
coupled plasma-atomic emission spectroscopy (ICP-AES) and inductively coupled
plasma-mass spectrometry (ICP-MS), which evolved from ICP-AES, have become
popular analytical methods due to their multielement capabilities, speed, and high
sensitivity.
Unfortunately, the traditional means of sample introduction to AAS, ICP-AES and
ICP-MS entail destruction of important time dependent information, which often involves
crude mechanical sampling and dissolution of bulk dental tissues (see Chapter Six). This
fails to capture the subtle gradations in composition, and thus, dietary intake, that is
inherent in sequentially calcified dental tissues.
Due to the compactness of teeth and the depth of stored biological data, the analysis
of dental tissues ideally requires microsampling techniques. The most suitable methods
currently available include laser sampling coupled with mass spectrographic analyzers
and electron or proton microprobes with X-ray emission detectors (e.g., PIXE) (Outridge
et al. 1995). According to Outridge and colleagues (1995: 164), laser ablation (LA) ICP-
MS may be the “most promising microprobe technique for quantitative elemental
analyses of biological microstructures”. This approach is utilized for this analysis and is
elaborated in Chapter Six (Methodology).

49
3.2 Dental Hard Tissues and Enamel Development
Human dental development is characterized by seven main physiological stages:
1) bud; 2) cap; 3) bell; 4) dentinogenesis; 5) amelogenesis; 6) appositional dentine and
enamel maturation (resorption); and 7) eruption.
The initial “bud” phase of (deciduous) tooth germ development takes place at about
six weeks after fertilization, when mesenchyme cells assume arch-shaped zones along the
developing jaws, while epithelial cells form the ten “swellings” in the jaw that will
become the enamel organs for deciduous teeth (Hillson 1996). Epithelium is the layer of
tissue that lines an embryo’s developing mouth. Underlying this layer are mesenchyme
cells, which will ultimately develop into the connective tissues of the body: bone,
cartilage, muscle, tendons, blood vessels, dentine and cementum.
The cap stage is characterized by the proliferation of dental tissues. At this time,
mesenchyme cells form the dental papilla and dental follicle, which will develop into
odontoblasts (dentine forming cells) and cementoblasts (cementum forming cells),
respectively. Later in this stage, the enamel organ differentiates into the internal enamel
epithelium, which will eventually differentiate into ameloblasts and form enamel matrix .
During the bell stage, dental cells differentiate further. Now, odontoblasts develop,
which secrete predentine matrix, and inner enamel epithelium cells differentiate into
ameloblasts, which secrete enamel matrix on top of the predentine. The enamel organs
are fully differentiated by the fourth month in utero, with dentinogenesis and
amelogenesis taking place soon afterwards (see below).
Human teeth are composed of three different hard tissues, namely cementum, dentine
and enamel (see Figure 3.1). Cementum is the thin calcified layer that covers tooth roots,

50
while dentine makes up the interior of tooth crowns and roots, and enamel forms the
crown exterior. Comparatively, mature enamel is the most inorganic: 97-98%
hydroxyapatite, with no organic collagen component. Dentine and cementum have a
lower inorganic component: 75% and 65%, respectively, which resembles bone (65%).
Rose et al. 1985
Figure 3.1 Cross-section of human tooth, with major anatomical features indicated
Cementum is the thin layer of bone-like connective tissue that surrounds the dentine
of tooth roots and forms the connecting tooth plates of dentine and enamel.
Ultrastructurally and biomechanically, it resembles bone, which is why it is often called a
bone of attachment (Scott and Symons 1974). Owing to their shared mesenchymal
origin, the apatite crystals of cementum are similar in size and composition to those of
bone and dentine. Likewise, cementum is mesodermal (mesenchymal) in origin, with a
collagenous organic matrix. Inorganic apatite comprises approximately 70% by weight
of cementum, with 24-26 wt % being organic content, and water amounting to 4-6 wt %
(Carlson 1990: 534).

51
Dentine is formed by odontoblast cells and makes up the interior of tooth crowns and
roots. It is the first dental tissue to mineralize, followed by enamel and cementum. Like
bone and cementum, dentine is a mesodermal, mineralized, connective tissue. Compared
to enamel, it is less inorganic (70-75 wt %), with an 18-21% organic component made up
of collagenous proteins, and the remaining 4-12 wt % composed of water (Carlson 1990:
533). Moreover, dentine in contact with the pulp cavity, or secondary (circumpulpal)
dentine has an even higher organic content. Unlike enamel, but comparable to secondary
cementum, secondary dentine continues to develop during the lifetime of a tooth.
Overall, dentine is less dense and more permeable than enamel.
The inorganic component of bone, cementum, dentine and enamel consists of calcium
phosphate crystallites, which mineralize after organic matrix secretion. Human hard
tissue crystallites are a carbonate hydroxyapatite, which are apatite minerals. Apatite
refers to a diverse group of phosphate minerals sharing the same basic structural type:
A4B6(MO4)6X2. Typical biological apatite contains calcium ions in both the A and B sites
of the lattice; a phosphate group such as PO4 in the MO4 position, and hydroxyl (OH) or
other ions in the X position (Carlson 1990: 531). The resulting chemical formula for the
inorganic crystal lattice of human bone and dental tissues is Ca10(PO4)6(OH)2 (see Figure
3.2).
Figure 3.2 Three-Dimensional
Chemical Configuration
of Hydroxyapatite
[Ca10(PO4)6OH2]

52
Apatite crystallites in dentine resemble those of bone in size and dimension,
averaging 2-4 m in length, and 0.3 m in width (Carlson 1990). This is noticeably
smaller than enamel crystallites, which range from 16-100 m in length, and 4 m in
width (Carlson 1990).
Finally, enamel is the hardest human tissue and it differs fundamentally from both
dentine and cementum. Notably, enamel derives from ectodermal (epithelial) fetal
tissues and has a non-collagenous organic matrix. It is formed by protein-secreting
ameloblasts from the inner enamel epithelium. Ninety per cent of the organic component
is amelogenin, which are proteins unique to enamel matrix.
While highly inorganic and hard when mature, enamel starts off as a porous
crystalline matrix filled with water and organic material inhabiting intercrystalline
spaces. Water, proteins and lipids form the aqueous matrix that surrounds enamel
crystals, which totals approximately 3-4% of mature enamel by weight (almost 15% by
volume) (Curzon and Featherstone 1983). Specifically, water amounts to approximately
2% of enamel by weight (12% by volume) (Carlson 1990), while organic material makes
up the remaining 1% by weight (2% by volume) (Jenkins 1978).
Enamel growth corresponds with dentine, which develops from adjacent odontoblast
cells. In general, the rate of enamel deposition is stable unless physiological disruptions
affect ameloblast secretion. In humans, the rate of enamel matrix secretion is estimated
at 4-4.5 m per day (Schour and Hoffman 1939; Massler and Schour 1946).
Amelogenesis is characterized by an initial secretion of enamel matrix accompanied
by some (primary) mineralization. Deciduous enamel formation begins at around four
months in utero, with the first tooth eruptions at about six months after birth, and all

53
deciduous teeth fully erupted by about two and a half years. Among permanent teeth,
central incisors, first molars and upper canines commence enamel formation earliest, at
around the time of birth (ending 4.5, 3.5 and 5 years later, respectively), while third
molars are the last developing teeth (generally between 9 and 13 years of age [Anderson
et al. 1976; Moorrees et al. 1963]).
In this study, permanent canine crowns are considered to begin enamel development
at between birth and six postnatal months and be complete by 5-5.5 years of age (see
Appendix A). Some variations in the rate of dental development exist between
populations, as well as between males and females (who are slightly advanced), but the
disparities are minimal, e.g., 2-3%, and are not considered significant (see Appendix A).
During the initial migration of ameloblasts from the dentine substrate, cone-shaped
Tomes’ processes develop at the end of each cell. With the completion of amelogenesis,
these processes eventually become reduced and disappear, resulting in a thin layer of
non-prismatic enamel at the enamel-dentine junction of the enamel surface (Scott and
Symons 1974).
Upon reaching full enamel thickness, ameloblasts switch from enamel matrix
secretion to absorption of protein and water. This subsequent phase of enamel
maturation, or secondary mineralization, results in chemical and structural changes to
enamel. It is a protracted and complex process that is thought to involve three phases of
calcification, starting from the surface to the dentinoenamel junction and then back
toward the surface, finalized by tertiary mineralization of the narrow subsurface layer,
which is the most mineralized of all enamel layers (Suga 1982).

54
Mineralization begins in the earliest secreted enamel at the dentine junction and then
extends outward as newly formed (immature) enamel develop. These “gradients of
mineralization” extend over many weeks in any one part of the developing crown
(Beynon et al. 1998: 353). It progress in an occluso-cervical direction and can vary in
timing depending on location, e.g., lateral tooth surfaces (buccal cervix) take longer to
mineralize than inner approximal enamel (Beynon et al. 1998; Liversidge 1995).
Unfortunately, enamel mineralization research currently cannot attribute a precise
time period for the complex process in permanent teeth (L. Humphrey, personal
communication, 2004), especially considering the extent of intratooth variation. The
main challenge has been the inability of radiographic techniques to identify the earliest
stage of enamel calcification, which is overestimated by X-rays (Hess et al. 1932;
Winkler 1995), while it underestimates the age of crown completion (see Beynon et al.
1998; Liversidge 1995).
Importantly, in this study, it must be remembered that relative age of individuals is
based on incremental enamel structures, which are the result of the first, mostly organic,
stage of enamel formation. The maturation phase witnesses the bulk of enamel
mineralization and it is the main period measured when elements such as strontium and
calcium are quantified by LA-ICP-MS. Since mineralization occurs over several weeks
after enamel deposition, this time delay is a source of error that must be recognized in
this investigation. However, until further research elucidates the timing of the maturation
process, the significance of this factor is uncertain.
During mineralization, the majority of crystal growth of the already initiated enamel
crystallites occurs (growing to 5-10 m in diameter), and the amino acid composition of

55
enamel protein changes (Carlson 1990; Hillson 1996). With a considerable decrease in
the volume of organic material and water, enamel matures into a harder and denser
calcified tissue that is almost 97 % inorganic by weight (86% by volume) (Curzon and
Featherstone 1983). Strontium is one element that is primarily incorporated in enamel
during the maturation phase.
For the purposes of this study, it is the inorganic composition of dental enamel that is
most relevant for elaboration. The inorganic component of human enamel is organized at
three structural levels: 1) the individual apatite crystal, 2) bundles of crystallites, or
enamel “prisms”, and 3) Hunter-Schreger bands, or prism layers.
Enamel apatite crystals are approximately 3-4 m in diameter and 10 m or more in
length (Orams et al. 1976). Similarly oriented apatite crystals occur in long clusters of
tightly packed hexagonal prisms or “rods”, which are roughly perpendicular to the tooth’s
surface. They arise from the dentinoenamel junction and extend to just below the surface
of the tooth. Enamel prisms are the morphologic unit of enamel and they can differ in
size, shape, orientation and relative density. Importantly, enamel apatite crystals are
imperfect structures and contain calcium deficient areas (Featherstone et al. 1981b).
Metal ions (e.g., strontium) or metal-fluoride complexes can incorporate in these
deficient areas and are an important component of enamel (see below).
Hunter-Schreger bands are layers of similarly oriented enamel prisms that are parallel
to one another but angled differently from prisms in adjacent bands. Such bands are also
differentiated by prism shape (Carlson 1990).
Morphologically, tooth crowns develop as fronts of corresponding enamel and
dentine bands (see Figure 3.1). Enamel growth bands appear histologically as successive

56
Striae of Retzius, which are regularly spaced, at 30-40m apart in the occlusal half of the
crown, and 15-20m apart in the cervical half (Hillson 1996). These striae record the
variable activity of ameloblasts during amelogenesis.
Physiological stressors during enamel development can result in pronounced,
darkened, striations. The neonatal line is a pronounced stria of Retzius that records the
time of birth in deciduous teeth and permanent first molars (Schour 1936) and can offer
insight into the relative timing and rate of dentine and enamel development before and
after this event.
Between adjacent striae of Retzius, there are approximately 7 to 8 cross-striations
(Newman and Poole 1974), or “pinch and swell” structures, spaced at regular intervals
about 4 to 8 microns apart along the length of enamel prisms (Boyde 1976). Current
research indicates a range of 6 to 12 cross-striations between brown striae of Retzius
(Beynon 1992; Bromage and Dean 1985; Bullion 1987; FitzGerald et al. 1996). Cross-
striations are caused by cyclical variation in the rate of enamel matrix secretion, with
swellings representing faster secretion, and constrictions, reduced secretion (Boyde 1976,
1989). The spacing of cross-striations in human teeth correspond well with the estimated
enamel matrix secretion rate of 4-4.5 m per day (Schour and Hoffman 1939; Massler
and Schour 1946), as well as with independent estimates for crown formation time (see
Hillson 1996). As a result, enamel cross-striations are thought to represent a 24 hourly,
or circadian, rhythm (Boyde 1976; Dean 1987; FitzGerald 1995, 1998), while striae of
Retzius represent weekly developmental features (Newman and Poole 1974).

57
3.3 Dental Enamel Composition
Since dental enamel is primarily inorganic, the analysis of enamel composition is
primarily concerned with elemental concentration. Elements fall under two main
categories: bulk elements (or macrominerals) and trace elements. They can be
cumulative in human hard tissues, e.g. lead, or they can fix in tissues up to a threshold
level, at which point the majority of the intake (or surplus beyond the needs of the
organism) is excreted in hair, urine, perspiration and/or feces. In this case, the element
maintains a relatively constant level within species.
In the human body, more than 99% by weight is composed of carbon (C), hydrogen
(H), oxygen (O), nitrogen (N), sulphur (S), phosphorus (P), calcium (Ca), magnesium
(Mg), sodium (Na), potassium (K) and chlorine (Cl), which are bulk or macro elements.
The first six bulk elements (C, H, O, N, S, P) form the major nutrients, namely proteins,
lipids, carbohydrates and nucleic acids. All bulk elements are essential for life.
Trace elements are minerals that occur in amounts of less than 0.01% of the body
mass. They can be separated into three categories: essential trace elements, metabolic
trace elements, and non-essential trace elements. In humans, fifteen elements are
considered “essential” trace elements, which are required for proper nutrition and
development of an individual. They are particularly vital for all biochemical processes
since they are part of important enzymes. All trace elements can be toxic in excessive
amounts, although some are noteworthy for physiological impairment even at low levels,
e.g., cadmium, mercury and lead (Larsen 1997).
Essential trace elements are those that are: 1) present in all healthy tissues of living
organisms; 2) occur in concentrations that are relatively constant between animals of the

58
same species; 3) can induce the same structural and physiological abnormalities and
dysfunctions (specific biochemical changes) in all species if removed from the body; 4)
can prevent or reverse such abnormalities if restored to the body; and 5) can prevent or
reverse such biochemical changes if the element is maintained (its deficiency or excess is
prevented or reversed) (Cotzias 1967). Indeed, both a deficiency and toxicity of essential
trace elements can disrupt body homeostasis and impart adverse physiological effects.
Essential trace elements in humans include iron (Fe), zinc (Zn), copper (Cu),
manganese (Mn), nickel (Ni), cobalt (Co), molybdenum (Mo), selenium (Se), chromium
(Cr), iodine (I), fluorine (Fl), tin (Ti), silicon (Si), vanadium (V), and arsenic (As)
(Underwood 1977). In life, these elements are provided by food and water, but ionic
exchange in the burial environment can also enrich or deplete these elements in fossilized
tissues. Other elements not considered “essential”, but which also play important roles in
human nutrition, include boron (B), barium (Ba), bromine (Br) and strontium (Sr) (see
below).
With fossilized hard tissues, trace elements are studied within the context of the
inorganic portion of bones and teeth, despite their presence in (significantly less) organic
portions. Excluding water and some organic material, the major components of human
dental enamel include Ca, P, CO3 (carbonate), Na, Mg, Cl and K. Enamel is
approximately 36% calcium in content (by weight), while phosphorus, which is present
almost entirely as phosphate, is between 16 and 18% (Curzon and Featherstone 1983).
The following tables outline the concentrations of bulk and trace elements in human
enamel (from Curzon and Featherstone 1983):

59
Table 3.1 Concentrations of major components of human dental enamel
excluding water and organic material
(Curzon and Featherstone 1983: Table 1)
Component Concentration range in percentage
(dry weight)
Ca 33.6-39.4
P 16.1-18.0
CO3 2.7-5.0
Na 0.25-0.90
Mg 0.25-0.90
Cl 0.19-0.30
K 0.05-0.30
Table 3.2 Range of element concentrations in human dental enamel
(adapted from Curzon and Featherstone 1983: Table 2)
Concentration
in ppm Elements
> 1000 Cl, Mg, Na
100-1000 K, S, Si, Sr, Zn
10-100 Al, B, Ba, Fe, Pb
1-10 Br, Cd, Cr, Cu, I, Mn, Mo, Rb, Sn, Ti
0.1-0.9 Ag, Be, Co, Hg, Li, Nb, Ni, Sb, Se, W, Zr
<0.1 As, Au, Bi, Ce, Cs, Dy, Er, Eu, Ga, Gd, Ge, Hf,
Ho, In, Ir, La, Lu, Nd, Os, Pb, Pr, Pt, Re, Rh, Ru,
Sc, Sm, Ta, Tb, Te, Tl, Tm, V, Y
(Together with calcium (Ca), highlighted elements are those quantified by
LA-ICP-MS in this study.)

60
Common trace elements in biological apatite include Al, Ba, Br, Fe, Pb, Sr and Zn.
Elements that exhibit wide variations in enamel include Pb, Mn, Se, Sr and Zn (see
Curzon and Featherstone 1983). The principal source of variation for many elements,
including strontium, in foodstuffs and eventually, hard tissues, is the underlying geology
(see below).
In teeth, there is general compositional homogeneity within enamel growth bands,
with variation more typical of intra-band differences, i.e., between enamel of different
developmental ages (Dolphin et al., in press; Goodman et al. 2003; Kang et al. 2004).
Any enamel chemical heterogeneity that occurs within bands is the result of individual
variation in mineralization during dental development or post-mortem external
contamination.
According to Curzon and Featherstone (1983), trace element composition in enamel
generally does not change during the course of enamel development, although certain
elements merit attention. Of twenty-four trace elements that have been studied for age
changes, fifteen have been observed to remain stable, five generally decrease (Cu, Fl,
Mn, Sr and Zn) and five can increase (Co, Pb, K, Na and Se) (see Curzon and
Featherstone 1983: Table 4). Age-related variations in element abundance can be
attributed to dietary changes (e.g., infant to adult foods), physiological changes (e.g.,
discrimination capacity, pregnancy, lactation), and accumulated pollution exposure.
However, extensive current research suggests that trace element behavior is probably
more complex and variable in humans. For example, strontium levels can increase and
decrease at different ages in the life of an individual depending on nutrition and digestive
system discrimination (see Chapter Four). Infants undergoing the transition from

61
exclusive breastfeeding to solid food reliance reflect differences in dietary (and thus hard
tissue) Sr/Ca that is due to the clear distinction between breast milk and solid food
strontium abundance. Breast milk has an extremely low Sr/Ca composition, while solid
foods and water have significantly higher concentrations. This fact forms the basis of
this investigation on ancient weaning. At the same time, hard tissue Sr/Ca is recognized
to be age-dependent on gastrointestinal maturation and strontium discrimination, which
develop during early childhood (see Chapter Four).
In light of the multielement analytical capabilities of ICP-MS, Se, Cu, Mn, Ba, Fe, Pb,
Sr, Zn and Mg were quantified from permanent canine enamel (see Table 3.2). Such
elements were chosen for their nutritional implications and potential for future research,
but for this investigation, the primary focus is strontium (Sr) (see Chapter Four).
3.4 Diagenesis and Hard Tissue Preservation
Concomitant with the extensive application of hard tissue chemistry to paleonutrition,
considerable research into the taphonomic processes that govern hard tissue preservation
was undertaken (DeNiro 1985; Ezzo 1994; Grupe and Herrmann 1988; Kohn et al. 1999;
Kyle 1986; Lambert et al. 1984, 1990; Pate and Brown 1985; Pate and Hutton 1988;
Price 1989; Price et al. 1992; Radosevich 1993; Sandford 1993; Sillen 1981, 1986; Sillen
and Sealy 1995; Sillen et al. 1989). Most researchers today recognize that prior to any
hard tissue chemical analysis, some assessment of postdepositional environment and
diagenesis must be determined to control for contaminated remains. Diagenesis is the
sum of all physical, chemical, and biological processes which occur in the post-mortem

62
depositional environment (Price 1989; Radosevich 1993; Sandford 1993; Sillen et al.
1989). All fossilized hard tissues undergo some degree of diagenetic alteration.
In short, the degree to which diagenesis can affect hard tissues is determined
generally by several factors: 1) time; 2) nature of the surrounding geological environment
(groundwater, soil pH, available elements); 3) age of individual at death (subadult bones
are less mineralized/more porous); and 4) type of hard tissue (compact cortical bone vs.
porous cancellous bone; bone vs. teeth) (Parker and Toots 1970, 1980).
Numerous diagenetic mechanisms can degrade the chemical integrity of human hard
tissues, most notably bone. Post-mortem factors include soil pH, organic matter content
(including bacteria, algae and fungi), mineralogy and texture, temperature, soil enzymes,
abundance and distribution of rainfall, and groundwater movement (Pate and Hutton
1988; Piepenbrink 1986; Hanson & Buikstra 1987). Such factors can enrich, deplete or
substitute original biogenic elements in hydroxyapatite by promoting soil element
precipitation and ionic exchange (Kyle 1986; Parker and Toots 1980; Pate and Hutton
1988; Schoeninger 1982).
The availability of insoluble or soluble and exchangeable ions in the soil solution of a
burial environment is the major factor determining whether elements will be chemically
mobile and available for contamination (Radosevich 1993). Ion availability is affected
by factors such as “weatherability” and solubility of a specific mineral, soil porosity, pH,
temperature and precipitation (Mitchell 1957).
The rate and intensity of soil weathering is highly dependent on water and
temperature. For example, exchangeable ions will make a greater contribution to soil
solution, and thus, hard tissue permeation, in more humid (hot and wet) climates, or in

63
areas where irrigation is practiced (Pate and Hutton 1988). In these environments, there
will be rapid weathering and downward soil leaching, compared to arid regions, where
there is slow weathering of bedrock and soils and upward evaporation of groundwater
(e.g., precipitation of calcite).
Soil pH plays a dominant role in determining the availability of various elements to
the soil solution. Bone hydroxyapatite is relatively insoluble at high pH levels (alkaline
conditions), but its solubility increases as pH decreases to 6.5 or 6, and increases rapidly
below a pH of 6.0 (Lindsay 1979; Pate and Brown 1985). Acidity in soil and/or
groundwater can result in the leaching of elements such as Zn, Cu, and Mn from organic
material (Gilbert 1985). Additionally, Mg, Sr and Zn can replace Ca heteroionically
(exchange with natural tissue constituents) at the surface of bone in acidic conditions
(Lambert et al. 1985b).
Lipids, humic acid (decayed plant matter), fulvic acid, and carbonates can also alter
hard tissue collagen isotopic values (Ambrose 1986, 1990, 1991, 1993; Chisholm 1989;
DeNiro 1985; DeNiro and Hastorf 1985; Heaton et al. 1986; Sealy et al. 1987; van der
Merwe 1989). Bone is susceptible to such alteration but tooth enamel is not significantly
affected and can preserve its dietary isotopic signature for up to several million years
(Ambrose 1993: 62).
Overall, the results of many studies indicate that barium, calcium, copper, manganese,
iron, aluminum, phosphorus, potassium, silicon and vanadium are elements that often
exhibit post-mortem alteration (enrichment or depletion) in human bones (Hancock et al.
1987; Keeley et al. 1977; Kyle 1986; Lambert et al. 1979, 1982, 1984, 1985; Parker &
Toots 1970, 1974, 1980; Pate and Brown 1985; Price & Kavanagh 1982; Sillen 1981b).

64
Elements that generally resist significant diagenetic change include magnesium,
strontium and zinc (Ezzo 1994; Kyle 1986; Lambert et al. 1979, 1982, 1984, 1990;
Sandford 1993; among others).
Of the hard tissues, dental remains maintain their chemical integrity most effectively
and are the best-preserved human tissues. Among dental tissues, enamel, particularly
core enamel, is the most resistant to chemical alteration in both the living mouth
environment and the post-mortem diagenetic environment, while dentine is the most
exposed and least resistant to alteration.
Unlike bone and dentine, enamel resists significant diagenetic alteration due to its
highly inorganic, acellular, nature, larger apatite crystal size, greater crystallinity,
hardness, and impermeability (Lee-Thorp 1989; Parker and Toots 1980). These
characteristics render enamel the hardest tissue in the human body, yet at the same time,
it is more brittle and has lower compressive strength than dentine.
The advantage of enamel’s low organic composition is also underscored by the highly
resistant nature of the organic material itself. In this case, the proteins and peptides of
mature enamel are tightly bound into the mineral structure and have been known to
survive in fossil enamel even millions of years old (Hillson 1996: 228).
Fully mature enamel is essentially fixed and removed from contact with cellular
elements, but it is not entirely inert, as ion exchange can take place between saliva and
the surface layer of enamel. Despite the fact that apatite crystals are generally
impermeable and more resistant to diagenetic alteration than bone, enamel prisms can
adsorb ions onto its surface that may eventually become incorporated into the surface
enamel. Major structural elements such as Ca and P, and to a lesser extent, Mg, Na and

65
chloride, remain relatively consistent, but there are substantial differences in carbonate
and trace element abundance (Carlson 1990; Puech et al. 1986).
In the living mouth, the outer few micrometers of enamel are regularly attacked by
acid from plaque bacterial metabolism, resulting in some enamel demineralization. The
partial demineralization that occurs is followed by remineralization as the partially
dissolved crystals “regrow with preferential exclusion of some ions and inclusion of
others” (Curzon and Featherstone 1983: 132). For instance, as a natural protective
mechanism against caries, fluoride, strontium and zinc can be incorporated during
remineralization or maturation of surface enamel (Curzon and Featherstone 1983) (see
Section 4.3 for in vivo incorporation of strontium). Such in vivo incorporation of
elements can be significant, with Reitznerová and colleagues (2000) identifying saliva
de- and re-mineralization of enamel as far as 150 μm from the enamel surface.
After death, enamel elemental composition can also be altered by diagenetic factors.
In particular, ions of barium (Ba), lead (Pb), magnesium (Mg), potassium (K), sodium
(Na) and strontium (Sr) from surrounding groundwater can replace calcium in enamel
crystal structures. Strontium can readily substitute for calcium in hydroxyapatite crystals
because of similar chemical and physical properties (see Section 4.3).
Other notable mineral exchanges in fossilized dental tissues include phosphate
substitution by carbonate ions, substitution of hydroxyl ions by fluorine and chlorine
(Carlson 1990); and enrichment of Fe, Mn, Si, Al, Ba and possibly Cu due to
contamination of enamel and dentine apatite with secondary minerals (Kohn et al. 1999).
Besides enamel chemistry, the post-mortem preservation of enamel can also be
affected on a gross level by the burial matrix. Despite its highly inorganic composition,

66
enamel is susceptible to adverse environmental conditions such as very acid soils.
Enamel can dissolve in such contexts and appear pitted and eroded but, interestingly,
some organic remnants of dentine can withstand the acidity (Stead et al. 1986).
Finally, outside of natural processes of diagenesis, cultural treatment of human
remains is an equally important factor in hard tissue preservation. In cremation, the
enamel of erupted teeth can flake away from dentine, but unerupted teeth are somewhat
protected by their bony crypts, and can retain their enamel (McKinley 1994). The
heating of bone affects its structural composition and isotopic signatures (DeNiro 1985),
but in teeth, limited findings indicate that there are only changes in tooth appearance and
microstructure (Shipman et al. 1984). Reflecting their hard, inorganic nature, enamel
prism structure is still visible even after temperatures of 8000C or more (Shipman et al.
1984).
In light of the evidence, enamel is the ideal hard tissue for chemical and sequential
analysis, and for the present paleodietary study, in particular, because of three main
qualities: 1) it is the best preserved and least diagenetically-affected human hard tissue;
2) unlike dentine and bone, which reflect an elemental signature accumulated just prior to
tooth loss or death, enamel preserves a faithful and indelible record of childhood dietary
(elemental) intake during the period of crown development, which is well understood;
and 3) non-developmental incorporation of elements in enamel can be recognized and
eliminated through several acid washing methods (see Chapter Six), and can be avoided
by sampling inner enamel.

67
3.5 The Nature of Hard Tissue Preservation at Lamanai
In light of the various factors potentially disguising the “true” chemical nature of hard
tissues, most notably bone, a sample-specific assessment of diagenesis is required for
archaeological remains. Fortunately, for Lamanai, White (1986) has accounted for
numerous controls (see Beck 1985; Gilbert 1977) to exclude all possibilities of
diagenesis. The skeletal remains of Lamanai individuals from all cultural periods at the
site are notably well preserved for sub-tropical conditions and relatively free of
significant diagenetic alteration (White 1986).
A major precondition for utilizing fossilized hard tissue samples in chemical analyses
is stability of the post-mortem environment. Generally, one can state that hard tissues
will be well preserved in soils of neutral or slightly alkaline pH, while acid soils inhibit
good preservation (Keeley et al. 1977). Low soil acidity (pH>6.5) will indicate low ionic
exchange and fewer available elements for contamination. Reduced exposure to
groundwater will also minimize the availability of exchangeable mineral ions. At
Lamanai, all human remains were buried in matrix above the water table and soil pH was
an acceptable mean of 6.8 (Lambert and Arnason 1984).
Comparison of elemental values between hard tissues and surrounding matrix will
give an indication of chemical stability, particularly trace elements that are known to be
diagenetically altered after deposition (see Keeley et al. 1977; Kyle 1986; Lambert et al.
1979, 1984; Nelson & Sauer 1984; Pate and Hutton 1988; Waldron 1981, 1983). For
instance, significant levels of iron, manganese and lead in fossilized tissues are reliable
indicators of diagenesis (Buikstra et al. 1989; Francalacci and Tarli 1988; Lambert et al.
1990; Runia 1987b; Waldron 1981, 1983). Rubidium and yttrium, which are common in

68
surrounding (calcareous) soil matrix, but normally absent in bone, can also be examined.
In the Lamanai remains, White (1986: 160) found no correspondence between soil and
bone values over time for elements relevant to paleonutritional analysis. Significantly,
for this investigation, Lamanai soil samples exhibit substantially lower Sr levels than
human bone remains (White and Schwarcz 1989: Table 2).
Due to the calcareous geology of the Maya Lowlands, precipitation of calcium into
hard tissues is a major concern (see Pate and Brown 1985). This limestone environment
is rich in calcium carbonate/calcite (CaCO3) and gypsum (CaSO4.2H2O), which are
common pore-filling minerals (White 1986).
Weathered hydroxyapatite in poorly preserved bone will have reduced Ca content,
which can be replaced by Ca ions from surrounding soil, and can actually result in Ca
enrichment (White and Hannus 1983). A high Ca/P ratio can identify calcium carbonate
addition because it results in the dilution of phosphorus (the theoretical ratio of Ca:P for
hydroxyapatite is 2.16) (Sillen 1989; White 1986: 121). Analyses of Ca/P ratios in
Lamanai bone suggest that there is no significant Ca enrichment over time, reflecting
good skeletal preservation.1 Thus, using Ca as an internal control is valid (White 1986).
Simple detection of diagenesis can also be accomplished by comparing different
fossil animal bones of known stratigraphic context at the site. The elemental and isotopic
integrity of hard tissues of animals with well-documented opposing diets can be
compared to test the post-mortem environment (e.g., Nelson et al. 1983; Pate and Brown
1 White (1986) notes, however, that a high mean ash content of 94.4% in Lamanai bones suggests diagenetic loss of organic
material. Leaching of calcium may have occurred, but it is not considered problematic because there is no significant difference in the
Ca/P ratio of Lamanai bones over time (i.e., Ca leaching was constant over time) (White 1986: 159). White (1986) also observed that
absolute and ratioed values of Mg and Sr with Ca were in accordance. While the Lamanai bones may exhibit some Ca leaching, dental enamel is more highly inorganic and resistant to such taphonomic processes and is probably minimally affected in this analysis.

69
1985, Price and Kavanagh 1982; Sillen 1981a, b, 1986, 1992; Wessen et al. 1978; White
1986, among others).
The range of animal species and plant resources available to the population at the
time should obviously be known, and in the case of Lamanai, faunal, paleobotanical,
ethnohistorical and archaeological evidence provide numerous sources of insight. Other
than variations due to semi-domestication (animal maize consumption), differences in
trophic position due to diet (herbivory vs. omnivory vs. carnivory) were clearly
discernible in Sr/Ca values of diverse Lamanai fauna, indicating good post-mortem
preservation (White 1986). Ancient Lamanai fauna also do not exhibit evidence of
significant post-mortem elemental deposition or leaching, and like the human remains,
faunal Ca/P ratios indicate chemical stability over time (White 1986).
In total, soil analysis, faunal controls and comparisons with human bone chemistry
indicate that Lamanai hard tissues are wholly suitable for chemical analysis and are
accurate indicators of in vivo dietary incorporation. Utilization of dental enamel, which
is more completely mineralized and impermeable to diagenesis than bone, means that
external and post-mortem effects on paleodietary research are especially limited. This
fact, combined with the timing of enamel development during a critical stage in life
history, highlights the uniqueness and applicability of dental tissues to health and
nutritional research.

70
CHAPTER 4
THE ROLE OF STRONTIUM ANALYSIS IN PALEONUTRITION
4.1 Strontium Distribution in the Food Web
Strontium (Sr) is a soft, silvery yellow, alkaline earth metal (atomic number 38) that
has a cubic face-centered crystal structure. It was first identified in Scotland in 1790 by
Adair Crawford and William Cruikshank as a component of the mineral strontianite. It
naturally occurs in the lithosphere as four stable isotopes (in order of % abundance): 88
Sr
(82.56%), 86
Sr (9.86%), 87
Sr (7.02%) and 85
Sr (0.56%), with an average atomic weight of
87.63. Fourteen radioactive isotopes of strontium also exist (e.g., 89
Sr and 90
Sr), which
have made strontium the focus of extensive research on nuclear fallout since the 1950’s.
Like other trace elements, its relative abundance in organisms is determined by species-
specific absorption ability and environmental availability, which have important
implications for paleonutritional reconstruction.
Importantly, the consideration of strontium (and other alkaline elements) in living
organisms and their fossilized hard tissues must be viewed as a ratio relative to the total
amount of available calcium (Sr/Ca). Strontium and calcium share a very close, direct,
relationship in living organisms, including shared metabolism and distribution (see
below). (In contrast, Sr has an inverse relationship with Mg, so that the two relationships
[Sr/Ca, Sr/Mg] should be reflected in correspondingly distinct ratios). Significantly, the
amount of Sr absorbed by organisms is wholly dependent on relative dietary calcium
abundance, with high Ca associated with reduced Sr absorption, and vice versa.

71
Consequently, for human dietary research, including understanding trophic level
fractionation, it is the Sr/Ca ratio that is relevant.
Following Beck’s (1985) and Lambert and coworkers’ (1982) suggestion to use
absolute Sr values instead of Sr/Ca ratios, White (1986) has examined both kinds of data
but found that Sr/Ca ratios closely mirrored patterns of absolute Sr values (reflecting
good Ca preservation). This is also the case for strontium data in this investigation.
It is, therefore, important to recognize the Ca abundance of consumed foods, in
addition to the Sr content and Sr/Ca ratio. As outlined by Sillen and Kavanagh (1982:
70), the efficacy of hard tissue Sr/Ca ratios for paleodietary research is determined by
“a) the Sr/Ca ratios entering food chains via water and plant foods, and b) the degree and
number of discrimination steps to which alkaline earths are subjected before reaching
human skeletons”.
In the natural environment, strontium is introduced at the plant level, through soil and
water, and is successively discriminated against as one moves up the food chain.
Specifically, Sr is found in the minerals celestite and strontianite. Variables in the
underlying geological substrate of the local environment will determine the amount of
strontium available to plants and animals (Toots and Voorhies 1965). Such factors,
which ultimately affect the Sr/Ca ratio of groundwater, include: 1) concentration of trace
elements in the parent rock; 2) rate and extent of weathering; 3) drainage of soils; and 4)
soil pH (Mitchell 1955, 1957).
Rocks such as shale and granite have the highest Sr/Ca ratios, while limestone
(calcium carbonate), despite having a high absolute concentration of Sr, has a low Sr/Ca
ratio due to a much higher abundance of Ca (Sillen and Kavanagh 1982). Dolomite and
non-volcanic rocks also contain relatively low Sr/Ca values (Odum 1957).

72
In their survey of American rivers and streams, Skougstad and Horr (1960, 1963)
found that areas with high rates of evaporation, low annual rainfall, high salinity and
highly soluble parent material had relatively high Sr concentrations. In general, Sr/Ca
ratios of watercourses remain constant within one major river or lake drainage system
(Sillen and Kavanagh 1982).
Schroeder and colleagues (1972) maintain that intake of foods make up the greatest
contribution of trace elements in humans. However, attesting to the importance of
geology and water sources, in some regions, water may contribute substantial proportions
of certain elements such as Sr, so that water quality is an important determinant of total
Sr intake and hard tissue chemistry (Wolf et al. 1973). Among contemporary Hopi
women, higher levels of bone strontium, compared to their nursing (and food
supplemented) children, has been attributed to greater intake of (relatively high Sr) water
in the form of tea and coffee (Kuhnlein and Calloway 1979).
Overall, the composition of food and water, which is related to local soil geology, is
very much interrelated. Plants growing in soil with relatively high available Sr/Ca ratios
will have generally high Sr/Ca ratios, while soils rich in Ca but deficient in Sr will
produce crops with significantly lower Sr/Ca ratios. Among foods, strontium
concentration (µg/g, or ppm) is most abundant in nuts and spices, followed by seafood,
root and leafy vegetables, legumes, grains, fruit, meat, dairy products, maize and lastly,
fats and oils (Rosenthal 1981; Schroeder et al. 1972) (see Table 4.1).

73
Table 4.1 Average Sr content of various foods (of diverse origin) in order of
relative abundance (with some traditional Maya foods italicized)
(adapted from Schroeder et al. 1972: Table 7)
Food Type Strontium (µg/g) Relative Abundance
Highest
Condiments Cinnamon 118.75
and Dry Spices Allspice 79.17
Cocoa 29.88
Nutmeg 13.90
Nuts Brazil 107.43
Pecan 13.63
Seafood Clam 25.93
Anchovy 16.46
Shrimp 6.18
Sardine (canned) 5.77
Vegetables Kale 117.28
Parsley 6.20
Carrot 2.65
Sweet Potato 1.09
Squash 0.94
Onion 0.65
Tomato 0.48
Green Pepper 0.29
Legumes Dry Lentils 5.07
Lima Bean 4.38
Green Beans 1.09
Red Beans 0.65
Grains Wheat 3.36
Millet 1.29
Barley 0.98
Fruit Strawberry 2.29
Grape 1.72
Banana 0.88
Meat Beef and fat 1.44
Chicken Leg 1.24
Pork Liver 0.76
Chicken Breast 0.52
Dairy Butter (unsalted) 0.80
Milk (whole dairy) 0.50
MAIZE Corn, sweet (fresh) 0.52
Fats and Oils Pork fat 0.14
Corn Oil < 0.10
Lowest

74
Staples such as cereal grains have low Ca contents, but also extremely low Sr values,
resulting in relatively low Sr/Ca ratios (Runia 1987b). According to Rosenthal (1981),
maize is noteworthy because it has “very low” Sr values relative to most foods, even
meat. Data in Table 4.1 reflect this, but both Schroeder and colleagues (1972), and
Rosenthal (1981), who cites their findings, do not specify strontium composition relative
to calcium abundance (Sr/Ca), which is more appropriate for studies of human
metabolism, nutrition and hard tissue incorporation. Additionally, food Sr values are
averaged and derive from different geographical areas, so that diverse geological
conditions, such as soil and water, are not accounted for in the differences.
In contrast, while it is not disputed that maize is low in total mineral content, others
characterize maize as having relatively high Sr/Ca values, like other plants (Burton and
Wright 1995). Compared to 0.52 ppm (Schroeder et al. 1972), Kunhlein and Calloway
(1979) have actually reported a maize strontium concentration of 60 ppm for modern blue
maize from the American Southwest, which is the traditional staple of the Hopi.
Kuhnlein and Calloway (1979) attribute a declining maize dependence to the difference
in tooth dentine between 17th
century Hopi and contemporary individuals, who exhibit
lower Sr values. The extent of the difference is not specified, but bone strontium levels
for the ancient Hopi averaged 478 μg/g (ppm) (Kuhnlein and Calloway 1979).
According to Kuhnlein and Calloway (1979), the high Sr abundance is derived from
dietary components such as blue maize, culinary plant ash (added during maize
processing), local green plants, native salts, as well as the clay of cooking and serving
vessels. Local water from the Colorado River is also notably high in Sr (higher than any
other river in the U.S.), and it is a significant factor determining food levels (Kuhnlein

75
and Calloway 1979). Yet, the strontium abundance cited by Kuhnlein and Calloway
actually refers to ash-processed maize, rather than raw maize. Burton and Wright (1995)
have observed that the Hopi practice of processing maize with saltbush plant ash
(bivilviki) produces a total maize Sr/Ca value that is, in fact, dominated by the Sr/Ca ratio
of the ash source, rather than the maize, which is higher in Sr/Ca. Ash-processed blue
maize consumed by the Hopi has an average Sr/Ca value of 8.48 x 10-3
(Kuhnlein 1981),
which is significantly higher than other known maize values (see Section 4.4).
The overall Sr/Ca ratio of raw maize appears to be relatively high due to its very low
calcium content. Wright (1994) reports on one untreated maize cob from Guatemala with
a Sr/Ca ratio significantly greater than most animal foods in the Maya area (Table 4.2).
Common animals consumed by the ancient Maya include deer, peccary, turkey, water
fowl, turtle, fish, large rodents and various freshwater snails (Emery 1999; Shaw 1991).
Sr/Ca values of some ancient and modern animal samples from Guatemala are
outlined in Table 4.2. Note, however, that Sr/Ca values refer to animal bone and shell
remains, rather than fleshy parts. Lamanai Maya would have consumed the meat of such
animals with a reduced Sr/Ca ratio compared to the hard tissues. In this case, the Sr/Ca
ratio of untreated maize would be enriched compared to animal foods (excluding the
turtle sample analyzed by Wright 1994).
However, when maize is treated with lime to facilitate cooking and consumption, its
Sr/Ca signature is indeed considerably reduced due to an enormous calcium contribution
(see Bressani et al. 2002; Burton and Wright 1995; Wright 1994). The effect of alkali
(lime) processing of maize on total Sr/Ca dietary contribution and hard tissue
composition is discussed in Section 4.7.

76
Table 4.2 Mean Sr/Ca values for common Maya animal and plant foods from the
Pasion region of Guatemala (ratios based on element data in Wright 1994:
Appendix D, E)
Food Item
Mean Sr/Ca (x10-3
)
S.D. (x10-3
)
N
ANCIENT FAUNAL BONE
White-tailed deer (Odocoileus virginianus) 0.551 0.261 3
Brocket deer (Mazama americana) 0.474 - 1
Peccary (Tayassuidae) 0.705 0.256 2
Turtle (Dermatemys) 1.260 0.260 4
Large rodent (Agouti paca) 0.241 - 1
MODERN FAUNAL BONE/SHELL
White-tailed deer (Odocoileus virginianus) 0.800 1.039 2
Brocket deer (Mazama americana) 0.278 - 1
Peccary (Tayassuidae) 0.237 0.200 4
Large rodent (Agouti paca) 0.594 0.695 3
Common turkey (Meleagris gallopavo) 0.154 - 1
Fish (colorada, guapote) 0.933 0.320 3
Freshwater snail (jute) (Pachychilus) 0.465 0.039 2
Freshwater snail (apple) (Pomacea) 0.672 0.301 4
MODERN PLANTS
Corn (Zea mays)
Squash (Cucurbita sp.)
Sweet pepper (Capsicum sp.)
1.129
1.446
0.239
-
-
-
1
1
1
Moving up the food chain, all plants will have lower Sr/Ca and Ba/Ca ratios than
their associated soils because of their tendency to preferentially assimilate calcium over
strontium and barium at the soil/plant interface, which is a process termed
“biopurification” (Elias et al. 1982; Burton et al. 1999).
Besides geological source, strontium values in plants differ significantly between
distinct species due to variations in calcium “biopurification”. In fact, the differences in
both Sr/Ca and Ba/Ca ratios of distinct plant species growing at the same location far

77
exceed (beyond an order of magnitude) the ratio differences among plants of the same
species from different locales (Burton et al. 1999). As stated by Burton and colleagues
(1999: 613), this inter-species variability, despite sharing the same soil conditions, can
exceed the differences due to trophic level biopurification, thus constituting a major
source of variation within a local food web. Still, according to Runia (1987b), it is the
difference in distribution of Sr and Ca that is most significant for plant material, rather
than the degree of discrimination against Sr, which is more relevant for animals including
humans (see below).
Within plants, there are important differences in Sr/Ca ratios between plant parts.
While leaves and shoots will most likely have Sr/Ca ratios resembling those of the soil
solution, plant roots and root crops will have a Sr/Ca ratio 1.5-10 times higher than leaves
or shoots (Runia 1987b: 602). It is thus important to note that the Sr/Ca ratios in specific
products such as cereal grains and roots will differ from the overall plant ratio, which in
this case is lower than the grain and root portions (Runia 1987b).
In mammals, the discrimination of strontium occurs in the digestive system (Kostial
et al. 1969; Lengemann 1963; Spencer et al. 1960; Taylor et al. 1962; Wasserman 1963).
Kidneys also contribute to reduced strontium levels by excreting the element more
rapidly than calcium (Walser and Robinson 1963; Spencer et al. 1960).
As stated above, “biopurification” is the tendency for organisms, both plant and
animal, to preferentially absorb calcium at the expense of other alkaline elements like
strontium and barium, resulting in the increased “purification” of calcium with each step
in a food chain (Elias et al. 1982). This preferential elimination of Sr and Ba relative to
Ca (fractionation) results in a reduction in Sr/Ca and Ba/Ca ratios relative to dietary

78
ratios, which is then reflected in hard tissues. [Comparatively, Ba is slightly more
fractionated than Sr (Elias et al. 1982).] The degree of Sr discrimination from plants to
humans is approximately 0.25% (Brown 1973).
Strontium, like most trace elements, enters the body through food intake and
digestion, distributing into the bloodstream as soluble ions. In older children and adults,
the alimentary tract is the primary site of Sr discrimination and absorption, depending on
relative Ca intake. In the case of developing fetuses and infants, the placenta and
mammary gland will be the main locations of discrimination and absorption (Comar
1963; Comar et al. 1955; Lough et al. 1963).
The overall reduction in Sr/Ca, like Ba/Ca, occurs with ascending trophic position in
the food web. Thus, strontium is more concentrated at the lower plant level, and
decreases, as does the Sr/Ca ratio, as it moves up the food chain through herbivores to
omnivores to carnivores (see Table 4.3).
Table 4.3 Ranges of Sr and Zn content in mammalian bones
(from Rheingold et al. 1983)
Mammal Type
Sr (ppm)
Zn (ppm)
Carnivore
100-300
175-250
(bobcat, wolf, mink, weasel)
Omnivore 150-400 120-200
(rat, beaver, dog, bear)
Herbivore 400-500 90-150
(cattle, deer, goat, rabbit)

79
The Sr/Ca ratios of animal tissues, including bone, are lower than their dietary ratios.
Among consumers, herbivores have lower Sr/Ca (and Ba/Ca) ratios than those of the
plants they consume, but higher ratios than other types of animals. Plant Sr/Ca variability
and species-specific consumption of different plants also means that some variation will
also exist within herbivores (Sillen and Lee-Thorp 1994). For instance, leaf-eaters will
exhibit lower Sr/Ca ratios than those herbivores eating high fiber diets (Sillen 1992).
Grazers, and possibly to a lesser extent, browsers, consume plant material with Sr/Ca
ratios that likely resemble Sr/Ca ratios in the reference material (soil and water) Runia
1987b). In general, plant material has approximately three times more strontium than
animal soft tissues (Schroeder et al. 1972). Carnivores will have the lowest Sr/Ca values
of the consumers because, in addition to their digestive system’s discrimination of
consumed Sr, they have ingested tissues that have already discriminated against Sr when
the animal was alive.
An Observed Ratio can be used to quantify the discrimination against strontium (or
differential distribution of Sr) in food chains (Comar et al. 1957). In this case, the
Observed Ratio (OR) = Sr/Ca in sample (e.g., bone/teeth)
Sr/Ca in precursor (e.g., diet)
The OR can be used to express the Sr/Ca relationships between plant/soil, milk/diet
and bone/diet or, essentially, the movement of Sr and Ca through the environment (see
Sillen and Kavanagh 1982). Conversely, the “biopurification” of calcium (discrimination
against strontium) or “discrimination factor” (DF) can be quantified as the inverse of the
OR (Elias et al. 1982; Rosenthal 1981). Notably, the OR values of plants express the

80
relationship between the Sr/Ca ratios in soils and various plant and plant parts (or
differences in the distribution of Sr and Ca in plants), rather than the discrimination
against Sr that is expressed in animal OR’s (Runia 1987b).
Numerous authors have quantified strontium discrimination in actual food chains (see
Sillen and Kavanagh 1982: Table 2). Sr/Ca OR values (bone/diet) for various adult
herbivores and omnivores (e.g., pigs, sheep, mice, cows and dogs, among others) range
from 0.16 to 0.35 (Alexander et al. 1956), but generally measure between 0.20 and 0.30
(Sillen and Kavanagh 1982: 73). Human adults have an average ORbone-diet value of 0.18
(Rivera and Harley 1965).
In their examination of South African food webs, Gilbert, Sealy and Sillen (1994)
clearly demonstrated trophic level fractionation of Sr/Ca. While plants have Sr/Ca values
(not OR) that range from 2.05-41.79 (with 80% in the range of 2.05-10.26), herbivore
bones exhibit ratios between 0.96-5.52, and carnivores from 0.39-2.31 (Gilbert et al.
1994: Table 1). Sr/Ca and Ba/Ca values for herbivores and carnivores overlap
considerably and both exhibit greater reduction in Ba/Ca than Sr/Ca because mammals
discriminate against Ba more than Sr (Elias et al. 1982). In general, their Sr/Ca ranges
are consistent with plant/consumer differences in other studies of the same region (Sealy
and Sillen 1988; Sillen 1992).
Similarly, Elias and colleagues (1982) found a four-fold reduction in Sr/Ca from
sedge leaves to voles, which concurs with findings in Sillen and Kavanagh’s (1982: Table
2) survey. Increased fractionation took the form of a further six-fold reduction in Sr/Ca
from voles to predating pine martens (Elias et al. 1982). Comparable data includes

81
Burton and colleagues’ (1999) 80% reduction in Sr/Ca for a bobcat/hare predator-prey
pair, as well as other findings by Price (1985) and Price, Smick and Chase (1986).
In light of the Sr/Ca distinctions between different foods and types of consumers, one
usefulness of Sr in paleodietary reconstruction concerns dietary regimens. Toots and
Voorhies (1965) were the first researchers to test this by differentiating bone Sr/Ca
between Pliocene herbivores and carnivores. Skeletons of individuals ingesting large
quantities of meat are expected to contain relatively low levels of Sr, while those relying
on a greater percentage of vegetable foods should exhibit higher Sr/Ca ratios (Brown
1973, 1974; Fuchs 1978; Gilbert 1975; Pate & Brown 1985; Price & Kavanagh 1982;
Schoeninger 1979a, 1981, 1982, 1985; Sealy & Sillen 1988; Sillen 1981a, 1986; Sillen
and Kavanagh 1982; Szpunar 1977). Brown (1973, 1974) has found that in herbivorous
humans, the average Sr/Ca ratio is around 1:2000, while meat-eating humans display a
mean ratio of 1:4800.
Seafood has been thought to concentrate much more strontium than terrestrial
resources due to high seawater strontium levels. Strontium is the most abundant trace
element in seawater, averaging 8 ppm (Schroeder et al. 1972). High Sr levels are
correlated with water salinity (Odum 1951), depth and temperature (Angino et al. 1966;
see White 1986). The abundance of Sr in seafood is due to compounds in shell such as
aragonite and strontianite, which accumulate Sr rather than maintain a maximum
threshold reflecting surrounding seawater (Schoeninger and Peebles 1981).
Given that there is high strontium concentration in seawater, and assuming that the
intake of similarly Sr-enriched marine foods would elevate tissue levels in consumers,
numerous researchers have examined human hard tissue Sr to assess the proportion of

82
seafood intake (Benfer 1984; Burton and Price 1990; Connor and Slaughter 1984; Gilbert
et al. 1994; Schoeninger and Peebles 1981; Sealy and Sillen 1988).
Nonetheless, while seawater is high in Sr, like terrestrial food webs, it decreases
(“biopurifies”) with increasing trophic position in the marine environment, so that marine
diets are not equally high in Sr (Burton and Price 1999). As Burton and Price (1999:
235) note, even humans consuming significant quantities of marine mammals and fish are
still eating large amounts of meat, which can be expected to have lower, rather than
elevated, Sr/Ca values, that are comparable to terrestrial omnivores and carnivores.
Consumption of seafood (including mollusks and crustaceans) has an insignificant
effect on human tissues because, like mammals, very little Sr and Ca concentrates in
edible portions of seafood, with most alkaline earth metals contained in hard tissues.
Strontium content in the soft tissues of fish and marine gastropods represent only
approximately 0.1% of the total Sr in marine tissues (Sillen and Kavanagh 1982: 79). (In
contrast, barium is very low in seawater, averaging 0.006 ppm compared to strontium’s 8
ppm [Schroeder et al. 1972]. Because it is absorbed at one-fifth the rate of Sr [Marcus
and Wasserman 1965], the Ba/Ca ratio of marine foods [flesh and hard tissues] is even
further reduced compared to Sr/Ca.)
As Burton and Price (1999) insist, hard tissue Sr/Ca is not a sensitive indicator of
marine resource consumption and cannot be used to differentiate between marine and
terrestrial diets. However, as it faithfully represents marine foods as a meat source, the
assertion that “bone strontium meaningfully tracks dietary Sr/Ca ratios” (Burton and
Price 1999: 235) is further strengthened.

83
4.2 The Role of Strontium in Human Nutrition
As stated above, strontium is considered a non-essential trace element in human and
animal nutrition. The definition of “essentiality” is based on an element’s influence on
“important physiological functions”, but in cases where the deficiency of an element is
not widely associated with unequivocal functional impairment (detrimental effects), its
nutritional necessity is less clear. Despite being non-essential, strontium’s metabolic role
in animals and humans can be demonstrated by the various abnormalities that develop in
its absence. For instance, in experimental animals, withholding of strontium from a
purified diet results in reduced hard tissue calcification and slowed growth rates (Rygh
1949).
Additionally, because it can substitute for calcium in calcium-deficient areas of
apatite crystals, strontium supplementation has been found to reduce the rate of bone loss
and increase bone density (re-mineralization) (McCaslin and Janes 1959, 1981; Shorr and
Carter 1952; among others). It may also act as a cariostatic agent for teeth, resulting in
lower caries incidence in areas of high strontium levels in drinking water, soil and foods
(see Curzon 1983).
The replacement or substitution of Ca by Sr entails changes in physical and chemical
properties of enamel apatites (Curzon 1983; Featherstone and Nelson 1980). For
instance, LeGeros and colleagues (1976) found that the presence of Sr limits the
incorporation of carbonate. Carbonate affects apatite structure, increases apatite
reactivity to acid, and has been associated with enamel crystal defects, dislocations and
increased solubility and dissolution (Curzon and Featherstone 1983; Cutress 1972;
Dedhiya et al. 1974; Featherstone et al. 1981; Nelson 1981).

84
Strontium substitution is thought to reduce enamel dissolution and caries incidence,
either by increasing the size and surface area of apatite crystals, or by reducing carbonate
incorporation (Curzon 1983: 299). In total, strontium can function very similarly to
calcium. Accordingly, strontium can be considered important (though not essential) for
human nutrition since its deficiency does result in some degree of sub-optimal biological
function, which can be prevented and reversed by its restoration to the body.
4.3 Strontium Absorption and Hard Tissue Incorporation
Strontium has a vital relationship to calcium in that its absorption and incorporation
in hard tissues are dependent on the concentration and availability of calcium in the body.
Increased intake of dietary calcium will reduce the level of strontium absorption and
retention (and Sr/Ca ratio), while reduced available calcium will result in greater uptake
of strontium (Thompson 1963). This discrimination against strontium exists because of
the relatively lower absorption rate, but higher urinary excretion, of strontium compared
with calcium (Curzon 1983). Since calcium is an essential nutrient, it is tightly regulated
in the body, with its absorption dependent on the balance between blood and extracellular
fluid calcium levels. Comparatively, calcium, strontium and barium are absorbed at a
ratio of 10:5:1 (Marcus and Wasserman 1965). [As a result, while Sr/Ca is reduced at
higher trophic levels in a food web, Ba/Ca is even further reduced (Elias et al. 1982).]
As outlined by Sillen and Kavanagh (1982), the mechanisms of absorption of Sr and
Ca are both qualitatively and quantitatively similar. For instance, strontium is bound by
calcium-binding proteins necessary for the transfer of calcium across intestinal

85
membranes, but this protein has a higher affinity for calcium than for strontium (Ingersoll
and Wasserman 1971; Menczel and Mor 1972; Wasserman 1963).
Besides calcium content, strontium absorption is also affected by all the factors that
influence calcium absorption, namely vitamin D, age and metabolic status (Sillen and
Kavanagh 1982). Comparatively, because calcium is an essential nutrient, but strontium
has no known independent metabolic function, normal adults absorb 40-80% of ingested
Ca, but only 20-40% of ingested Sr (Spencer et al. 1960, 1973). Estimates for Sr
absorption among animals are even as low as less than 10% of ingested Sr (Elias et al.
1982; Wasserman 1963).
The average adult diet generally provides 1 to 3 mg Sr/day, but the total intake is also
dependent on water quality and strontium content (Schroeder et al. 1972). As well,
differences in Sr intake can be related to animal milk consumption. In populations that
consume animal milk beyond childhood, the average Sr intake is approximately 1.4 mg
Sr/g Ca (Sr/Ca ratio of 1.4 x 10-3
) per day, while those who do not consume animal milk,
and who rely on grains as a staple, can average 2.7 – 4.9 mg Sr/g Ca (Sr/Ca ratio of 2.7 -
4.9 x 10-3
) (Rosenthal 1981). For instance, higher Sr/Ca values in the hard tissues of
individuals from East Asia have been attributed to lower calcium intake from milk
avoidance (after childhood) (Schroeder et al. 1972). While American children and adults
average 96 ppm and 110 ppm in bone strontium, respectively, Asian children and adults
have bone Sr values averaging 320 ppm and 190 ppm (Schroeder et al. 1972).
Among adults, providing that calcium levels remain within normal physiological
limits, the relative absorption of Sr and Ca does not vary a great deal (Wasserman and
Comar 1960; Spencer et al. 1961; Wasserman 1963) (but see below for variations in

86
absorption due to age, sex and other factors). As a result, Sillen and Kavanagh (1982)
maintain that medium or high levels of calcium in the diet are unlikely to have a
significant effect on the applicability of Sr/Ca analysis to paleodietary reconstruction,
since the discrimination against Sr is generally unaffected. However, they warn that it
should not be used in cases where extreme calcium deprivation is suspected.
While the majority of dietary strontium is excreted renally, or during lactation and
placental transfer among women, a small amount of strontium is circulated in the
bloodstream for eventual hard tissue incorporation. In erupted teeth, in vivo exchange of
elements primarily occurs in dentine via the pulp cavity blood supply, but saliva also acts
as a medium for element exchange in enamel.
Alkaline earth elements such as strontium and calcium are notable calcified tissue
seekers, with most strontium incorporating into the hydroxyapatite fraction of bones and
teeth after absorption (Boyde et al. 1958). In fact, strontium and calcium are deposited in
calcified tissues in greater quantities than most other trace elements (Turekian and Kulp
1956). In humans, approximately 99% of body Sr and Ca is stored in hard and
connective tissues (Avioli 1988; Elias et al. 1982; Schroeder et al. 1972). In enamel,
strontium is primarily incorporated during the maturation phase.
Strontium incorporates into bone and dental apatite by replacing calcium ions. This
is the case for all divalent cations, i.e., Ca, Sr, Mg, Ba, Pb and Zn. In life, Sr2+
is
incorporated in bone as an isovalent replacement for Ca2+
. Up to four of the Ca ions in
human enamel apatite can be replaced by Sr. This can be attributed to strontium’s close
similarity to calcium in chemical and physical properties, most notably electron

87
configuration, ionization energy and ionic size (Sr = 0.112 nm; Ca = 0.099 nm) (Curzon
1983: 296; Schoeninger 1979a).
As stated above, the incorporation of Sr in apatite crystals results in some structural
modification. In synthetic hydroxyapatite, the addition of Sr can increase the size of
each crystallite (LeGeros et al. 1977) and limit carbonate incorporation (Curzon 1983;
LeGeros et al. 1976). Fortunately, a small amount of strontium exchange for calcium
does not adversely affect apatite formation or calcification, but large amounts may inhibit
calcification and have been shown to produce “strontium rickets” in experimental
animals (Storey 1961); dentine hypomineralization and enamel hypoplasia in rats
(Curzon 1983); and enamel mottling in humans (Curzon and Spector 1977).
Importantly, strontium levels in hard tissues are not directly proportional to the
amount of strontium in the diet, but rather, to the mean Sr/Ca ratio of the diet (Burton and
Wright 1995). With preferential calcium absorption, the amount of strontium absorbed
and retained in hard tissues is only approximately 20% that of calcium (Elias et al. 1982).
At each trophic level, hard tissue Sr/Ca will drop to approximately one-fifth of the dietary
Sr/Ca value (Elias et al. 1982). As summarized by Burton and Wright (1995: 274):
(Sr/Ca)hard tissues (0.2)(Sr/Ca)diet
As previously stated, knowledge of the total Ca contribution to a diet is essential.
The dependence of Sr on Ca intake means that high calcium foods will have a
disproportionately large effect on hard tissue Sr/Ca, while low Ca intake will have a
negligible, or even reverse, effect on hard tissue Sr/Ca ratios (Burton and Wright 1995:
275). Consequently, high intake of maize enriched with Ca from alkali treatment, which

88
lowers the Sr/Ca ratio, has a potentially significant effect on hard tissue composition (see
Chapter Eight).
4.4 Variations in Strontium Absorption
Studies of enamel strontium content indicate wide disparities in concentration, with
values as low as 13 ppm, to as high as 1400 ppm, in permanent teeth from dissimilar
geographic regions and time periods (see Curzon 1983: Table 15-6; Curzon and
Featherstone 1983: Table 3; Curzon and Losee 1977; Lane and Peach 1997: Table 2;
Losee et al. 1971; Wolf et al. 1973). In a survey of 24 countries around the world,
Thurber and colleagues (1958) found means that ranged from 101-344 ppm, while
Turekian and Kulp’s (1956) investigation of contemporary skeletons from 14 countries
gave values between 40 and 740 ppm. Comparisons of bone Sr between various regions
of the United Kingdom gave an overall coefficient of variation of 32.8%, ranging from
21.6 - 47.8% variation within counties (see Sillen and Kavanagh 1982: Table 5).
Such variation can be attributed to various factors, namely: distinct geology (water,
food sources); diverse analytical methods (spectrography, neutron activation, X-ray
emission, mass spectrometry, optical emission, atomic absorption); sample size variation;
different developmental and temporal ages of hard tissue samples; and variable caries
prevalence (which can reflect different Sr levels of the environment and tooth).
Within teeth, Steadman and authors (1958) have found a consistent distribution of Sr
across the total depth of enamel, but most evidence indicates that there are recognizable
variations in Sr/Ca between different enamel layers, most notably in the surface layer of
permanent teeth (Curzon 1983). (Similarly, slight Sr variation has been found between

89
different bone types such as cancellous ribs and cortical bone within the same individual
[Tanaka et al. 1981]. This variation, which corresponds with age-related differences, has
been associated with metabolic rates, but in archaeological samples, it is also possible
that diagenesis is accounting for some variation, as cortical bone is more stable and
resistant to post-mortem degradation than cancellous bone [see White 1986].)
In enamel, Little and Barrett (1976) have found a decreasing gradient of strontium
concentration from the surface inward, which suggested surface enamel incorporation of
strontium from the oral environment after tooth eruption. When they examined caries
rate, they found that teeth from areas of high caries prevalence had less surface enamel
strontium than teeth from low caries regions, which had at least twice as much strontium
(and fluoride) in surface enamel. Curzon (1983) has noted, however, that the variation in
enamel strontium and caries indices within groups is too large to ascertain a clear
relationship, as Spector and Curzon (1979) actually found no significant relationship
between surface enamel strontium and individual caries prevalence.
Conversely, Sr demineralization has also been observed in surface enamel, resulting
in increased Sr concentration toward the enamel-dentine junction (Reitznerová et al.
2000). According to Reitznerová and colleagues (2000), it is likely that pH changes in
the oral environment are responsible.
Between the two dentitions, it has been postulated that there might be higher trace
element levels in permanent enamel compared to deciduous enamel (Vrbič et al. 1987).
This was based on the fact that while some trace elements are bound to the mineral
structure or organic matrix of enamel during enamel formation, other elements are

90
adsorbed on the enamel surface later in life, i.e., post-development, which would give
permanent enamel more time to acquire (surface) elements.
Vrbič and co-workers (1987) found that of eight elements measured, three (Cd, Cu,
V) were actually higher in deciduous enamel, but five (Al, Ba, Li, Mo and Sr) occurred in
higher concentrations in permanent enamel, although only Ba (p<0.01), Li (p<0.01) and
Sr (p<0.02) were highly significant. In this study, average Sr concentration in primary
teeth was 74.2 μg/g (ppm), while the mean value for permanent teeth was 106.7 μg/g
(Vrbič et al. 1987: Table II). However, these results are absolute strontium values, rather
than relative Sr/Ca ratios, which are more relevant for comparison. In addition, a search
of the literature failed to uncover other findings or supporting evidence.
Variations aside, strontium concentration commonly ranges from 100-200 ppm in
enamel (Brudevold & Steadman 1956), or 111 ppm on average (Carlson 1990: Table 1),
150-250 ppm in bone ash (Aufderheide 1989), and 100-600 ppm in dentine (Rowles
1967), or 94 ppm on average (Carlson 1990: Table 1).
Within such ranges, strontium absorption differs by age, sex and reproductive status.
The most important metabolic factor affecting absorption of alkaline earth metals in
animals is developmental age, since discrimination and relative absorption of such
minerals is dependent on gastrointestinal capacity (maturation). For instance, Sowden
and Stitch (1957) found that average strontium abundance in nursing infants was 79.1 +
20 μg/g (ppm), which increased regularly with age, reaching 114 + 28 μg/g in adults.
Bedford et al. (1960) and Schroeder and colleagues (1972) have also observed the same
positive trend with age.

91
However, the evidence has not been consistent, with some studies finding higher
Sr/Ca in children compared to adults (Brown 1973; Lengemann 1963; Lough et al. 1963);
and others reporting no significant age difference except in fetal bone (Szpunar 1977;
Turekian and Kulp 1956). It is believed that factors mentioned previously are responsible
for the inconsistency of findings, i.e., differences in geology, preservation, analytical
method, sample size, age categories and caries prevalence.
In the total human lifespan, average bone strontium levels generally decrease with
age (Curzon and Featherstone 1983). This general pattern is characterized by several
notable fluctuations however. Among children, there are age-related variations in Sr/Ca
enamel composition due to dietary changes that are significant. For this study, in
particular, the variation in strontium concentration prior to age-related decline is the
premise from which infant dietary behavior will be based.
Early life is characterized by relatively low Sr/Ca levels, with newborn infants
generally having the lowest Sr/Ca ratios in a population due to placental discrimination of
Sr during fetal development (Comar 1963; Comar et al. 1955, 1957; Spencer et al. 1956).
Compared to mothers’ tissues, fetal hard tissues are approximately half the Sr/Ca
concentration (Comar 1963). In their examination of maternal and stillborn skeletons,
Knizhnikov and Marei (1967) found that the observed ratio (OR) of infant bone Sr/Ca to
maternal diet Sr/Ca was only 0.08, while the OR of maternal bone Sr/Ca to maternal diet
was 0.20, which was in the normal adult range (Sillen and Kavanagh 1982). Similarly,
Beninson and colleagues (1964) found an adult OR (maternal bone/maternal diet) of 0.23,
while the stillborn OR (stillborn bone/maternal diet) was only 0.013 for Sr/Ca. Results
by Sowden and Stitch (1957) also reveal low Sr content in infant hard tissues.

92
However, young animals actually consistently exhibit less discrimination against
strontium than adult animals (see Sillen and Kavanagh 1982: Table 3). This is due to the
higher absorptive efficiency of alkaline earths in young animals, or rather, reduced
discrimination (less urinary excretion) of Sr by immature digestive systems. Newborns
and infants up to 9 months of age are observed to have bone Sr/Ca comparable to their
diet (OR ~ 1.00) (Lough et al. 1963; Rivera and Harley 1965; Straub et al. 1961).
Alkaline elements present in milk are absorbed at a higher rate than those of other
foods since such elements are more completely absorbed from liquids than solid foods
(Marcus and Lengemann 1962). The absorption of Sr from milk increases proportionally
to Ca due to an enhancement produced by lactose (Lengemann 1963). Vitamin D and the
amino acids lysine and arginine also act to increase Sr absorption in milk (Comar et al.
1957).
While relative Sr absorption may be higher in infants, the absolute amounts of Sr
available to infants and young animals are significantly lower than the average adult diet,
since milk is exceptionally low in Sr due to mammary gland discrimination (Lough et al.
1963; Twardock 1963). Indeed, compared to calcium, the transfer of strontium from
blood to milk is 10-14 times less (Comar et al. 1961; Wasserman et al. 1958). Unlike the
relatively high Ca content of human milk, Sr values range between 0.14-0.35 mg Sr/g Ca
(Rehnberg et al. 1969; Rosenthal 1981), or a breast milk Sr/Ca ratio of 0.14 - 0.35 x 10-3
.
Exclusively breastfed infants display the lowest Sr/Ca values until they are
supplemented with non-breast milk foods and liquids (e.g., cereal gruels), which have
significantly higher Sr/Ca abundance (see Chapter Five). For instance, Central American
corn is reported to have Sr/Ca values of 1.13 x 10-3
(Wright 1994) and 2.51 x 10-3

93
(LaVigne 2002), while some North American corn even reaches 7.14 x 10-3
(Shacklette
1980; Watt and Merrill 1963) and 8.48 x 10-3
(Kuhnlein 1981), which is likely due to
processing techniques (see Section 4.1). Increased food supplementation and decreased
breast milk intake is postulated to increase the level of Sr/Ca in infant hard tissues
developing at the time, in effect tracking the process of weaning (see Mays 2003; Sillen
and Kavanagh 1982).
In their study, Bryan and Loutit (1964) found that British infants from birth to six
months of age had a mean bone value of 0.2 mg Sr/g Ca, or a Sr/Ca ratio of 0.2 x 10-3
.
After six months of age, when infants began food supplementation, bone Sr/Ca rose
steeply and continued to increase slowly thereafter until adulthood, when bone values
average around 0.33 mg Sr/g Ca (Sr/Ca of 0.33 x 10-3
).
Despite maturation of the gastrointestinal tract after the first year of life (Sillen and
Kavanagh 1982), and a concomitant increase in the discrimination of Sr, higher food
strontium intake still results in rising levels of Sr. Thus, while infant hard tissue strontium
is primarily influenced by diet, i.e., breast milk and eventual solid food intake, after the
first birthday, gastrointestinal maturation (discrimination) plays an important role in
variations in hard tissue Sr/Ca. By about 8-10 years of age, with fully mature
gastrointestinal systems and improved Sr discrimination, reduced subadult Sr/Ca values
generally resemble those of adults (Rivera and Harley 1965). However, strontium levels
are the most varied in children, particularly between the ages of 4 and 13 (Lambert et al.
1979; Sillen and Kavanagh 1982). High values also at adolescence reflect peak periods
of metabolic activity (Lambert et al. 1979; Loutit 1967).

94
Thereafter, strontium levels among adults will stabilize between the ages of twenty
and fifty years (Lambert et al. 1979). In later adulthood, reduced gastrointestinal
discrimination (rather than dietary changes) results in slightly increasing strontium
values, although not to the extent of subadults. For instance, Hodges and colleagues
(1950) found that adult cadavers 42-75 years of age had bone strontium values averaging
around 0.59 mg Sr/g Ca (Sr/Ca ratio of 0.59 x 10-3
).
Since strontium variability in bones of children and adolescents is greater than among
adults, Sillen and Kavanagh (1982) have insisted that bone strontium should only be
compared between adults. However, this variability in children, when applied to dental
chemical analysis, provides a vital comparative tool for nutritional research. As a
permanent record of elemental incorporation during crown development in childhood,
such variation is highly useful to assess differences in infant nutrition between
individuals of a population through time and space.
In terms of variation due to sex, the results do not entirely agree. Lambert and co-
workers (1979) have found significantly higher Sr levels among males, but Price and
colleagues (1986) have observed the opposite, with females having higher strontium
levels, which were partly attributed to reproductive status (see below).
The hard tissue Sr/Ca values of modern western populations do not reflect significant
differences between adult males and females (Turekian and Kulp 1956b), but Sillen and
Kavanagh (1982) note that this equality might not characterize prehistoric populations. In
contrast to contemporary western women, in ancient times, it is expected that relatively
more women were either pregnant or lactating for most of their adult lives (Sillen and
Kavanagh 1982). Many studies, of both modern and ancient skeletons, have reported

95
relatively higher Sr/Ca levels in the bones of adult females compared to males (Blakely
1989; Brown 1973; Gilbert 1975; Lambert et al. 1979; Price and Kavanagh 1982;
Schroeder et al. 1972; Sillen 1981a; Vuorinen et al. 1996).
Higher levels of hard tissue strontium in women are related to metabolic changes
during pregnancy and lactation. Numerous studies indicate that pregnancy increases the
absorption of all alkaline earths, most notably calcium (Allen 1998; Allen et al. 1994;
Hayslip et al. 1989; Heaney and Skillman 1971; Kostial et al. 1969; Ritchie et al. 1998;
among others). Maternal calcium levels are particularly lowered during pregnancy
because of fetal development demands. Despite continued discrimination against
strontium by the placenta and mammary gland, a slightly reduced discrimination with
pregnancy entails a significant net increase in the Sr/Ca ratio of maternal plasma, and
ultimately, hard tissues (Kostial et al. 1969; Blanusa et al. 1970).
Physiological differences in hard tissue Sr/Ca due to pregnancy and lactation
obviously have ramifications for status distinctions in archaeological populations.
Several studies associate relatively high bone strontium (inferring high plant intake) with
“low status” individuals, while more privileged individuals have lower strontium values
that are attributed to greater access to meat resources (Brown 1973; Gilbert 1975;
Lambert et al. 1979; Price and Kavanagh 1982; Schoeninger 1979a,b; Sillen 1981; White
1986; White and Schwarcz 1989; Wolfsperger 1993).
For instance, at Chalcatzingo, Mexico, high status individuals (as determined by
grave associations such as ceramics, ground stone, figurines, jade) were found to have the
lowest mean bone strontium levels, which were associated with less reliance on maize,
and greater access to meat protein (Schoeninger 1979). In contrast, lower status

96
individuals, in poorly furnished graves, provided very high mean bone strontium values,
which was associated with greater reliance on plant foods such as maize.
However, in light of better understanding of strontium and calcium metabolism and
hard tissue incorporation, as well as the effects of alkali processing on maize Sr/Ca
values (see Section 4.7), this status association needs re-examination. According to
Burton and Wright (1995), the calcium-rich lime used to process maize results in
extremely low Sr/Ca maize products (gruel, masa, tortillas) that are indistinguishable
from other low Sr/Ca foods such as meat.
This explains the supposed “discrepancy” in Tipu Maya bones found by Bennett
(1986), who assumed that high maize intake should entail high bone Sr/Ca. In that case,
while carbon isotopic data inferred high maize intake (between 65-75% of total diet), this
seemingly did not correspond with the low Sr/Ca content of bones. Presently, this can be
explained by the high Ca content of lime-treated maize ingested by Tipu Maya.
Similarly, White (1986) found that Lamanai females with relatively low bone Sr
levels had supporting (high) Mg values that pointed to greater maize intake, contrasting
the traditional high Sr-high maize association. Relatively low levels of bone zinc also
suggest reduced meat (and seafood) consumption among Lamanai women (White 1986).
At Lamanai, differences in strontium values were also interpreted by White and
Schwarcz (1989) to reflect status distinctions in meat intake. In this case, elite Early
Classic tomb individuals (male and female) exhibited significantly higher strontium
concentrations than non-tomb individuals (White 1986; White and Schwarcz 1989).
Together with isotopic data (15
N and 13
C values), the authors attributed these patterns
in hard tissue chemistry to greater elite consumption of imported reef-marine foods, but

97
reduced consumption of more “ordinary”, locally-produced, maize, compared to non-
elites. Furthermore, differences in isotopic 15
N values between male and female elites
suggest that elite males exhibit even greater distinction, with the greatest access to
imported foodstuffs (White and Schwarcz 1989).
Yet, despite the high strontium content of seawater and marine animal hard tissues, it
is the soft tissues of such seafood that are consumed, which are, like meat, notably low in
Sr/Ca (see Burton and Price 1999). In this case, marine foods should be indistinguishable
from terrestrial meat and cannot be inferred by hard tissue Sr/Ca.
In light of sex differences, it is possible that, in other cases, individuals of lower
socioeconomic status may have actually been females who had children, in which case,
metabolic factors, rather than status-related dietary access, are responsible for higher
Sr/Ca values (Sillen and Kavanagh 1982). Still, females are often accorded lower
socioeconomic status (and less access to high-protein status foods such as meat) than men
in archaeological populations, so that the association of high Sr/Ca values (high plant
intake) with low social status is not entirely incorrect.
Clearly, any assessment of status associations with Sr/Ca values in hard tissues must
be evaluated in light of the sex of each individual. Higher female strontium levels may
be due to differential food (meat vs. plant) intake, but pregnancy and lactation also
influence relative Sr absorption and hard tissue incorporation. Further, if maize is the
major staple of the population, as it is for the ancient Maya, then the relatively reduced
Sr/Ca ratio of lime-processed maize must be considered (see Section 4.7). Finally,
attributing sex or status factors to distinctions in dietary intake and hard tissue Sr/Ca must

98
also rely on comparative isotopic (e.g., carbon) evidence, as well as archaeological
context (e.g., burial assemblage).
4.5 The Applicability of Strontium in Hard Tissue Paleodietary Analysis
In recognition of the myriad factors affecting hard tissue chemistry, a number of
criteria have been amassed to determine the appropriateness of an element for
paleodietary research (see Beck 1985; Ezzo 1994; Gilbert 1977; Gilbert and Mielke 1985;
Parker and Toots 1980; Price 1989; Sandford 1993; among others). These limiting
characteristics include:
1) The element must be hard tissue-seeking, i.e., it concentrates in hard tissues and is
incorporated into the hydroxyapatite crystal, rather than remain surface-bound on the
crystal
2) There must be documentation of a direct relationship between the levels of an element in
the diet and the levels present in human skeletal tissue. Moreover, the element values in
different food types should be distinguishable by trophic level (biopurification), or some
other distinction (e.g., tissue-specific occurrence)
3) The element levels in the skeletal tissue must be adequate to allow for accurate and
consistent measurement and such levels should be most diagnostic of an expected range of
consumption
4) The element preferably should not be an essential nutrient, so that it is not under tight
homeostatic regulation (since this feature of essential nutrients means they are largely
non-fluctuating)
5) The element should mimic the activity of a hard tissue-seeking essential element (e.g., Ca)
in biological systems
6) Concentrations of an element in human hard tissues either should be unaffected by
disease, or the diseases which alter the concentration should be identifiable in skeletal
remains
7) Concentrations of an element in human tissues should not be subject to significant
diagenetic change, either by leaching or enrichment, during extended periods of interment
8) Such elements have the potential to test nutritional problems using other skeletal
indicators

99
As stated by Ezzo (1994: 8), and also applicable to dental hard tissues,
…for an element to be a valid paleodietary indicator it should be incorporated into the
structure of bone hydroxyapatite at levels that are proportionate to levels in the diet, it
should not be an essential nutrient or one subject to metabolic regulation, and it should
be present in bone at levels that exceed contributions likely to occur as a result of
postdepositional processes.
On the whole, strontium, barium, zinc and magnesium have been the most widely
examined elements in paleodietary research due to their sensitivity to dietary intake and
post-mortem stability in human hard tissues. However, according to many researchers
(Beck 1985; Brown 1973; Burton and Price 1990; Klepinger 1993; Lambert et al. 1979,
1982, 1985, 1990; Price et al., 1985, 1986, 1992; Radosevich 1993; Sandford 1993;
Schoeninger 1979; Sealy and Sillen 1988; Sillen 1992; Sillen and Kavanagh 1982; Toots
and Voorhies 1965, among others), only strontium and zinc meet all the necessary
requirements and can be considered the most appropriate elements for paleodietary
reconstruction. In part, increased understanding of the effects of diagenesis on hard
tissue chemistry has significantly altered the perception that all dietary elements can be
used to infer diet.
Notably, however, Ezzo (1994) feels that only strontium and barium are valid
elements for hard tissue paleodietary research. Zinc should be excluded because 1)
changes in zinc intake of adults do not significantly correspond with relative zinc levels
in bone; and 2) there is no well-founded physiological or biochemical model for its
incorporation in hard tissues (Ezzo 1994).
Unlike many other elements, there is clear understanding of the complex nature of
strontium metabolism, particularly the realization that its eventual makeup in hard tissues

100
is a result of diet, age and health status, as well as post-depositional factors.
Distinctively, strontium has been extensively studied with respect to 1) its varied
distribution in food chains; 2) its physiological basis in human and animal nutrition,
particularly regarding discrimination; 3) the nature of in vivo hard tissue incorporation; 4)
its post-mortem stability/diagenetic alteration in hard tissues; and, importantly, 5) clear
demonstration that its variation in dietary components corresponds with variations in hard
tissue composition.
4.6 Strontium Preservation in Fossilized Hard Tissues
Since paleodietary reconstruction relies on fossilized human hard tissues, the
compositional integrity of such tissues in the post-mortem environment is a principal
consideration. Fortunately, the use of strontium in bone and dental chemical analysis is
fully warranted. Overwhelming evidence from studies of both hard tissue indicators of
diet and hard tissue diagenesis indicate that Sr/Ca in fossilized human hard tissues is a
faithful reflection of in vivo incorporation (Burton and Price 1999; Ezzo 1994; Lambert et
al. 1985; Pate and Brown 1985; Schoeninger 1979b; Sillen and Kavanagh 1982; White
1986; among others). Strontium’s post-mortem chemical stability is attributed to its very
stable 2+ cation position in apatite mineral, which is typical of all alkaline earth metals,
including calcium, barium and magnesium.
Still, there has been some conflicting evidence as to its post-mortem integrity in
human hard tissues, most notably bone. While generally stable, strontium values in fossil
bone can be depleted or enriched by mineral dissolution of the hydroxyapatite structure.
This can occur by microbial decomposition of collagen, and consequent weathering by

101
organic and carbonic acids formed by the decomposition (Pate & Brown 1985: 488). In
addition, acidic soil conditions can result in the replacement of surface bone Ca by soil
Sr, Mg and Zn, resulting in their enrichment in bone (Lambert et al. 1985b).
The strontium content of fossil bones and teeth is usually higher than the strontium
levels of surrounding sediment matrix (Wyckoff and Doberenz 1968), although this is
expected because of primary enrichment in living tissues from dietary and water intake,
and similarly high calcium content of fossils (Sillen and Kavanagh 1982).
During fossilization, almost all minor and trace elements are enriched, rather than
depleted (Carlson 1990: 545). This is in contrast to major elements (e.g., calcium and
phosphorus), which generally remain stable post-mortem, although carbonate has been
found to deplete slightly (Fosse et al. 1981; Puech et al. 1986).
Parker & Toots (1970, 1974, 1980) have maintained that human hard tissues are not
subject to diagenetic strontium addition. Their analyses of bone, dentine and enamel
from fossil Subhyracodon indicate consistent, equal amounts of strontium, while sodium
concentrations are lower (due to leaching/diagenetic loss), and fluorine concentrations are
higher (due to diagenetic enrichment) in bone and dentine relative to enamel.
However, Sillen and Kavanagh (1982) challenge this interpretation of the stability of
strontium in fossilized hard tissues. They suggest that the equal strontium levels between
bone, dentine and enamel actually reflect chemical equilibrium of the remains with the
post-mortem environment. Firstly, strontium from adult bone and tooth enamel can not
be compared because enamel is not remodeled in adulthood, as is bone, and therefore
reflects circulating element levels of the young animal, while bone reflects adult

102
circulating levels. Regardless of dietary stability, metabolic differences between young
individuals and adults should reflect different elemental levels.
Strontium contamination has also been found in human skeletons from freshwater
and coastal shell middens (the source of contaminating Sr), which were in concentrations
much higher than would be expected for normal reliance on seafood (Gilbert et al. 1994;
Kyle 1986; Schoeninger and Peebles 1981; Sealy and Sillen 1988). In addition, Sr
diagenesis is evident in fossils from Hayonim Cave, Israel, which failed to exhibit Sr
distinctions associated with fossil herbivore, carnivore and human omnivore remains
(Sillen 1981b).
Fortunately, the minimal evidence of diagenetic alteration of strontium has been
restricted to fossil bones, rather than teeth. Strontium is consistently observed to retain
its biogenic concentration and isotope ratio in enamel tissue, unlike dentine, which has
even been shown to undergo complete turnover of original biogenic Sr (Budd et al.
2000). Almost certainly, this fact renders the analysis of strontium in dental enamel the
most reliable method to reconstruct ancient diet and nutrition from human remains.
4.7 The Effects of Maize Processing on Hard Tissue Sr/Ca Composition
In examining maize’s Sr/Ca imprint in hard tissues, one must consider the
“paradoxical effect” of maize, or the nature of hard tissue Sr/Ca to change in a direction
contrary to that of the increasing maize component, due to low mineral content (Burton
and Wright 1995). Increased intake of lime-treated maize decreases the relative
contribution of other dietary components, but calcium abundance in the enriched maize
dampens the total Sr/Ca ratio so that it mistakenly appears like some other (low Sr/Ca)

103
foodstuff is increasing instead of maize (Katzenberg 1984; Burton and Wright 1995). In
total, high calcium foods will have a disproportionately large effect on hard tissue Sr/Ca
(Burton and Wright 1995).
To shed light on the nature of Sr/Ca patterns in Lamanai enamel, the dietary diversity
and food processing techniques of ancient Maya need to be assessed. Like most
populations in the New World, the Maya relied on maize as a dietary staple. However,
untreated, dried, ripe maize has marginal nutritional value, particularly in the essential
amino acids lysine and tryptophan, as well as niacin, an important member of the vitamin
B complex (Katz et al. 1974, 1975; among many others). Raw maize has relatively high
Sr/Ca compared to breast milk and meat products, but it is also notably low in total
mineral content, especially calcium.
Fortuitously, human societies across time and space have processed maize with
techniques that, while originally intended to facilitate digestibility, also improved its
nutritional value (see Katz et al. 1974, 1975; Bressani 1990; Bressani et al. 1990, 2002).
These treatments include the use of lime, wood/plant ash, lye and burnt shell ash to
produce alkaline solutions that soften and remove the external pericarp of maize (see
Katz et al. 1974), with lime and shell being the most common sources of alkali for the
ancient Maya.
In Mesoamerica, the process of alkali treatment, or nixtamalization, generally entails:
1) near boiling of dried maize in a 5% solution of lime in water for 30-50 minutes
(mixing maize with lime produces a dilute calcium hydroxide solution that is basic, or
alkaline, in pH); 2) discarding of the maize pericarp after cooling the mixture; 3)
thorough washing and draining of the remaining maize; and 4) fine grinding of the

104
remaining maize into a dough called masa (Katz et al. 1974: 174; 1975). This dough can
be made into tortillas by cooking on a hot clay or metal griddle, or used to make
sourdough, tamales, maize gruel or a stew ingredient.
During the process of lime soaking and heating, maize notably loses some
constituents. These include essential amino acids (excluding lysine), thiamin, riboflavin,
niacin, nitrogen, iron, zinc, fat and crude fiber (Bressani 1990; Bressani et al. 1958, 1990,
2002). However, the overall outcome is an enhancement of nutritional (biochemical)
quality, particularly a significant increase in bioavailable calcium, doubling (or more) of
essential amino acids, improved protein quality, and the release of otherwise almost
unavailable niacin (see Katz et al. 1974; Bressani et al. 1958, 1990, 2002).
As malnutrition can result from high consumption of untreated maize, it has been
concluded that societies that relied on maize as a major dietary staple (half to two thirds
of total diet) likely practiced a form of alkali processing (Katz et al. 1974; 1975). This is
supported by ethnographic analogue and cultural continuity of such cooking practices in
Latin America today, but it is also inferred by archaeological evidence. This includes
lime-encrusted vessels at Teotihuacan, Mexico dating to 100 B.C. (C.C. Kolb, pers.
comm. to Katz et al. 1974: 183), Terminal Classic (or earlier) jars with lime deposits at
Lamanai, as well as lime-coated colanders from Postclassic Lamanai (White 1986: 85).
Observations by Landa (1566, in Tozzer 1941) also indicate that Maya in the Yucatan
treated maize with lime at the time of Spanish contact.
Significantly for this study, the act of alkali processing produces maize that is
enriched in calcium by orders of magnitude (Bressani et al. 2002; Burton and Wright
1995; Wright 1994). For instance, Wright (1994) found that tortillas treated with lime

105
exhibit Ca values six to twenty times higher than unprocessed maize. Bressani and
colleagues (2002) found that this process increased Ca by as much as 24 times in the
maize germ. Such calcium enhancement can be viewed as an evolutionary adaptation
that played a key role in preventing calcium deficiency, which is significant for
populations whose diet can verge on such deficiency due to low consumption of animal
products such as milk (Bressani et al. 1958, 2002).
High Ca renders maize Sr (as well as Ba) ineffective (or “invisible”) in influencing
hard tissue Sr/Ca to resemble its relative concentration in untreated maize (Burton and
Wright 1995). This is because increased calcium intake will reduce the level of strontium
absorption and retention by the body. Hard tissue strontium is not directly correlated
with the particular Sr/Ca ratio of plant or meat foods, but rather the average dietary Sr/Ca
ratio, which can be affected by certain cooking practices. When examining Maya diet,
one must consider that hard tissue strontium will be “disproportionately sensitive” to high
calcium foods such as alkali-treated maize (Burton and Wright 1995). According to
Burton and Wright (1995: 278), a specific food such as maize must make up more than
90% of the total diet before the dietary and hard tissue Sr/Ca will reflect that food’s Sr/Ca
ratio.
Katzenberg (1984) and Szpunar (1977) previously recognized this in studies of
ancient bone chemistry and maize diets, but, according to Burton and Wright (1995), they
had incorrectly attributed the “paradoxical” effect to low Sr in maize, rather than its low
total Ca (mineral) content. Similarly, Price and Kavanagh (1982) found that despite
archaeological and isotopic evidence for increasing maize intake in American Late
Woodland groups (Bender et al. 1981), bone Sr values were low instead of high,

106
contradicting the general association of elevated Sr values with high plant consumption
(see Chapter Four).
To illustrate the effect of food Ca on the Sr/Ca ratio, Burton and Wright (1995)
examined the combined effects of two and three component diets on the total Sr/Ca ratio.
In this case, meat, beans and corn represent some major items in a traditional Maya diet.
The authors found that in a diet composed of meat and beans, the high Ca and Sr levels of
beans, compared to meat, were over represented in the overall bone Sr/Ca ratio. As they
state, unless the diet was mostly composed of meat products, the total dietary Sr/Ca ratio
(and bone Sr/Ca) would almost entirely be derived from the beans (Burton and Wright
1995: 278).
In a diet composed of meat, beans and (alkali-treated) maize, even when meat levels
remain constant, increasing maize consumption results in a decreasing Sr/Ca ratio, as the
high Ca (low Sr/Ca) maize signature dominates the total ratio (Burton and Wright 1995).
Diets with the same plant/meat ratio can exhibit different Sr/Ca and Ba/Ca due to
variations in Sr, Ba and Ca in the plant foods ingested. Conversely, diverse diets can
result in similar Sr/Ca and Ba/Ca ratios in hard tissues, particularly if high calcium plants
provide the main source of dietary calcium (Burton and Wright 1995; Burton et al. 1999).
In total, for paleodietary reconstruction based on strontium, more research is needed to
assess the Sr/Ca relationships in various food items and their combined effect on hard
tissue composition.1
1 Cleary, for any paleodietary reconstruction based on hard tissue Sr/Ca data, the carbon and nitrogen isotopic composition of
comparable hard tissues should also be analyzed to determine if low Sr/Ca actually reflects high meat intake or high alkali- treated
maize intake. The repercussions of misinterpreting such scenarios for status- or gender-related differences in diet are considerable.
For Lamanai, White (1986) has examined 15N and 13C isotopic values in bone to reconstruct diet and the chemical distinctions
related to gender and socioeconomic status. Such evidence, in addition to strontium results, indicate that status distinctions among the
ancient Maya can be recognized in hard tissue composition reflecting relative maize and meat intake (also see Coyston et al. 1999; Gerry 1993; White 1999; White and Schwarcz 1989; White et al. 1993; Wright and White 1996) (see Section 4.4).

107
On a practical level, it is unlikely that Maya infants consumed a maize diet that was
not processed according to the treatment of adult maize foods, simply for digestion
purposes. Maize gruel is produced by boiling ground maize dough in water, often with
the addition of flavorings such as honey and seasonings. The dough is derived from
ground maize that was likely processed with lime or other alkali ingredients (e.g., shells
[Nations 1979]). In this case, one expects the Sr/Ca ratio to be reduced compared to the
Sr/Ca ratio of raw maize. Based on Wright’s (1994) observations of calcium enrichment
in tortillas, one can expect the Sr/Ca ratio of lime-treated maize to be reduced six to
twenty times compared to raw maize. (However, the extent of calcium enrichment in
maize atole is unknown and may differ from tortilla values due to its watery base [see
Chapter Eight].)
Nevertheless, Lamanai mothers would have consumed the same calcium-rich maize
during lactation, producing a reduced Sr/Ca breast milk that would be further diminished
due to mammary gland discrimination. Infants consuming low Sr/Ca breast milk will
also have low hard tissue values, and despite lime processing, maize weanling foods
(watery gruel) will remain enriched in Sr/Ca compared to breast milk (see Chapters Five
and Eight). The transition from exclusive nursing to maize supplementation should
therefore be reflected in the increasing Sr/Ca composition of an individual’s hard tissues
developing at that age. This is the fundamental premise of this investigation and it
capitalizes on the reliability of hard tissue strontium to chronicle dietary intake and the
weaning process.

108
CHAPTER 5
BREASTFEEDING, WEANING AND INFANT HEALTH
Breastfeeding is an intricate dance between mother and child,
during which the child’s physical, cognitive, and emotional well-being
are nurtured, and through which they flourish.
(Dettwyler 1995: 65)
5.1 The Nutritional Qualities of Human Breast Milk
As universally acknowledged, breast milk is the perfect food for young infants,
containing more than 200 recognized constituents with physiological significance. As a
food, it contains a vast array of bioactive compounds that are required for normal growth
and development, and which confer immunity to infants with immature immune systems.
These highly bioavailable constituents include enzymes, growth factors such as
epidermal growth factor (EGF), prostaglandins and prolactin, hormones, peptides,
glycoproteins, amino acids and glycoproteins responsible for intestinal maturation, and
immunological protection against microbial pathogens in the form of IgA antibodies,
leukocytes, lysozyme, lymphokines and lactoferrin (see Table 5.1).
Generally, lactation can be divided into three stages, based on milk composition: 1)
the immediate period (up to one week) after birth, when breast milk is referred to as
colostrum; 2) a period of “transitional milk” from one to three weeks after birth; and 3) a
period of “mature” breast milk from three weeks onward (Institute of Medicine 1991).

109
Table 5.1 Constituents of human milk
(from Institute of Medicine 1991: 114)
Classes of Constituents in Human Milk
Protein and Nonprotein Nitrogen Compounds Carbohydrates
Proteins Lactose
Caseins Oligosaccharides
-Lactalbumin Bifidus factors
Lactoferrin Glycopeptides
Secretory IgA and other immunoglobulins
-Lactoglobulin Lipids
Lysozyme Triglycerides
Enzymes Fatty acids
Hormones Phospholipids
Growth factors Sterols and hydrocarbons
Nonprotein Nitrogen Compounds Fat-soluble vitamins
Urea A and carotene
Creatine D
Creatinine E
Uric acid K
Glucosamine
-Amino nitrogen Minerals
Nucleic acids Macronutrient Elements
Nucleotides Calcium
Polyamines Phosphorus
Magnesium
Water-Soluble Vitamins Potassium
Thiamin Sodium
Riboflavin Chlorine
Niacin Sulfur
Pantothenic acid Trace Elements
Biotin Iodine
Folate Iron
Vitamin B6 Copper
Vitamin B12 Zinc
Vitamin C Manganese
Inositol Selenium
Choline Chromium
Cobalt
Cells
Leukocytes
Epithelial cells

110
However, the concentration of human milk constituents varies considerably both
within and among women, i.e., at different stages of feeding, diurnally, from day to day,
and at different stages of lactation. Maternal nutrition determines human milk
composition in three ways: 1) current dietary intake, 2) nutrient stores, and 3) alterations
in nutrient utilization influenced by hormonal characteristics of lactation (Institute of
Medicine 1991). Specifically, milk composition is affected by all the factors that
influence plasma composition, hormones, and non-nutritional influences such as nursing
frequency, environmental conditions (e.g., exposure to infections) and length of gestation.
Abundant evidence indicates that short-term decreases in dietary intake and/or
quality, even among undernourished women, will not significantly affect nutrient content
in breast milk (Dorea 1999, 2000; Institute of Medicine 1991; Laskey et al. 1998;
Lonnerdal 1986a,b; Prentice et al. 1994). In such cases, women are still able to produce
milk with adequate protein, fat, carbohydrates, folate and most minerals, including iron
and zinc (of which breast milk is a good source) (Dallman 1986; Institute of Medicine
1991; Lonnerdal 1986; Lonnerdal et al. 1981; Murray et al. 1978; Siimes et al. 1984). To
maintain nutrient levels in the face of malnutrition, nutrients for milk synthesis derive
from maternal stores or body tissues.
In general, major minerals such as Ca, P and Mg are tightly regulated in maternal
serum, so that maternal intake of such nutrients will not significantly alter breast milk
composition (Cross et al. 1995; Institute of Medicine 1991; Krebs et al. 1997; Laskey et
al. 1998; Prentice et al.1994). Other factors that do not appear to consistently and
significantly alter breast milk Ca (and P) concentration include maternal malnutrition
(unless there is severe calcium deficiency), maternal age, race, socioeconomic status,

111
infant gestation length, metabolic disorders, stage of lactation, milk volume, child
weaning, smoking habit, calcium and vitamin D supplementation, prolonged medication
use (e.g., oral contraceptives) and sampling technique (DeSantiago-Soledad et al. 2002;
Dorea 1999; King et al. 1992; Laskey et al. 1998; Prentice and Barclay 1991).
Thus, while previous dietary guidelines for pregnant and lactating women
recommended significant increases in Ca intake, this is no longer the case in North
America (see Allen 1998). During pregnancy and lactation, Ca mobilization to the
developing child is maintained by increased efficiency of maternal metabolism, which is
not affected by maternal Ca consumption (Allen 1998; Cross et al. 1995; King et al.
1992; Krebs et al. 1997; Laskey et al. 1998; Sowers et al. 1993). Even with lactation-
induced bone loss, particularly the whole-body mineral content of trabecular bone in the
spine and femoral neck (e.g., Laskey et al. 1998), epidemiological data indicates that
there is a subsequent complete replacement of such mineral loss (Cummings et al. 1995;
Koetting and Wardlaw 1988).
Breast milk calcium content is still variable, however, as indicated by data surveyed
over the last fifty years from around the globe (see Dorea 1999, 2000). Mean calcium
concentrations have ranged from 84 to 462 mg/L (with a median of 252 mg/L) (Dorea
1999), though most data reflect a range of 200-300 ppm (Krachler et al. 1999), or a mean
Ca concentration of approximately 280 mg/L in mature human milk (Institute of
Medicine 1991: Table 6-1). Comparative data for Maya women consuming a traditional
diet is not known, as is the effect of local geology on Maya breast milk, but data for rural
Mexican Otomi women subsisting on a similar high-maize, low-fat/protein diet indicates

112
mean breast milk Ca concentrations of 268 µg/g (ppm) at four months of lactation and
240 µg/g (ppm) at six months of lactation (Villalpondo et al. 1998: Table 3).
Within individuals, noticeable differences in the concentration of calcium over time,
that are unrelated to nutritional status, include significant increase in Ca content during
early lactation (1-3 months), followed by a decline after 6 months of nursing, to a level
lower than in earlier stages (Karra et al. 1988). Similarly, cross-culturally, in both healthy
and malnourished mothers, zinc steadily decreases over the course of lactation (Karra et
al. 1988).
To an extent, maternal nutrition during lactation has little impact on infant growth.
This has been shown by studies in Taiwan (Adair and Pollitt 1985), Columbia (Herrera et
al. 1980) and Guatemala (Lechtig and Klein 1980), among others. Whitehead and Paul
(1984) found that the growth of breastfed infants in the first four to six months of life did
not differ significantly between industrialized and developing countries.
However, chronic malnutrition and ill health in pregnant and lactating mothers will
result in some physiological consequences for developing offspring. The nutritional
status of exclusively breastfed infants is dependent on three major factors: 1) infant
nutrient stores, which are determined by placental nutrient transfer in utero, gestation
length and birth weight; 2) amount and bioavailability of nutrients provided by breast
milk; and 3) environmental and genetic factors that affect the infant’s rate of nutrient
utilization and nutritional status, such as growth, infections and disease (see Institute of
Medicine 1991).
Allen (1994) has found that lactating women are more likely to suffer from
micronutrient deficiencies that can ultimately affect breast milk composition and infant

113
development. Other studies have also associated increased infant growth with maternal
supplementation and breast milk quality (Delgado et al. 1982; Herrera et al. 1980). In
particular, chronic malnutrition will affect vitamin levels in breast milk, particularly
vitamins A, B6, B12 and D (Institute of Medicine 1991). Some studies have also found
correlations between maternal Zn intake and Zn content in breast milk, especially in cases
of Zn supplementation (Karra et al. 1988).
5.2 Determining Factors in Breastfeeding and Weaning Behavior
Despite being the principal means of infant nutrition, breastfeeding is a highly
variable behavior that is subject to numerous biological and cultural demands. Current
Western beliefs promote prolonged breastfeeding, noting the nutritional, immunological
and emotional benefits. Conversely, analyses of the nutrient quality of breast milk and
the growth requirements of young children suggest that infants require additional
nutrients by six months of age. For instance, if breast milk is the only consumed food,
there is a risk of iron deficiency by the age of six to nine months (Institute of Medicine
1991). Some exclusively breast-fed rural Maya infants even begin to falter as early as
three months of age (Rivera and Ruel 1997). As a result, weaning is an event of great
importance to the well-being of children and is significant in nutritional research.
In discussing infant health and nutrition, weaning is more appropriately defined as the
complete cessation of breast milk intake, rather than the process of dietary
supplementation, which is also known as “supplementary” or “complementary” feeding.
Such definitions are important, as many weaning studies are not clear about whether they

114
refer to the onset of weaning, the rate that new foods replace breast milk, or the complete
cessation of nursing (see Schurr 1997).
In this study, examination of Sr/Ca changes in permanent dental enamel represents
the introduction and rate of non-breast milk supplementation among infants. Rather than
being a single, abrupt event, weaning is the outcome of an extended, gradual, process that
involves two dietary transitions. Supplementation, or the gradual introduction and
increased consumption of non-milk weanling foods (beikost) after a period of exclusive
breastfeeding, represents the first transition, or the onset of weaning. This process is
accompanied by the concomitant decline of breast milk intake at a variable rate, which is
replaced with alternate sources of liquid. Over many months, supplementation is stepped
up, while breastfeeding is reduced, until the child is weaned completely off breast milk.
Hence, the ideal measure of weaning behavior accounts for both the timing (age of onset)
and rate of weaning, in addition to the chemical composition of childhood diet (Schurr
1997).
The consumption of supplementary foods has a complex effect on the total amount of
nutrient absorbed by infants, including 1) adding nutrients in a less bioavailable form; 2)
decreasing the bioavailability (impairing the absorption) of nutrients (e.g., iron) in human
milk; and 3) decreasing the intake of breast milk and other important factors in milk (see
Institute of Medicine 1991: 154).
Most importantly, the period and pattern of breastfeeding is both a biological and
culturally determined process. Cultural beliefs about weaning age can affect birth
spacing, maternal nutrition and the physiological and emotional health and growth of
young children. Understanding the cultural determinants of weaning is essential.

115
In practical terms, weaning depends on factors such as mother and infant health; rate
of physiological, neurodevelopmental and behavioral maturation; interest in food;
number of erupted teeth; available food types that are considered suitable for child
digestion; relative cost of supplementary foods; mothers’ perceptions of a child’s
developmental progress or delay (i.e., wasting); and, importantly, cultural beliefs about
appropriate age (see Ballabriga and Rey 1987; Dettwyler and Fishman 1992; Fildes 1986;
Konner and Worthmann 1980; Lawrence 1989; Martorell and O’Gara 1985; Stuart-
Macadam and Dettwyler 1995, among others).
Nursing practices are influenced significantly by living standards (socioeconomic
status), individual needs, tradition and religious beliefs, customs, structure of the
mother’s daily activities, and beliefs about where the child sleeps at night. Other
culturally-specific beliefs include issues of contamination of supplementary foods; the
nature of infancy; notions of “proper” relationships between mother and child, between
mother and father, and their perception by society; and ideas about personal
independence and autonomy.
Sex of the child can also play a role in determining weaning behavior. For instance,
mothers have been reported to nurse male infants longer than their female infants in
Canada, Sweden, Ireland, France, Guatemala, Ecuador, Brazil, Peru, Taiwan, India,
Jordan, Liberia and Botswana (see Fildes 1986; McKee 1984). In extreme situations, the
disparate weaning patterns play a direct role in disease incidence, infant growth and
development, and ultimately, child survival, i.e., death of malnourished female infants
(see Stuart-Macadam 1995a: Figure 1.1).

116
Indeed, weaning represents a nutritional and health transition in early childhood with
adaptive significance for later life. Other than fulfilling its immediate role of nourishing
a developing child, breast milk and its astounding array of bioactive ingredients bestow
numerous advantages. These are principally related to breast milk’s role in infant
immunological development, intestinal growth and maturation (Bines and Walker 1991)
and neurodevelopment.
Specific benefits of breastfeeding for human offspring include: reduced rate of food
and environmental allergies (Kramer 1988); lower risk of jaundice, ear, gastrointestinal
and respiratory infections (Cunningham 1995; Duffy et al. 1986; Mata et al. 1967; Glass
et al. 1983), cancer (Davis et al. 1988; Freudenheim et al. 1994; Micozzi 1995), Crohn’s
disease and colitis (Cunningham 1981, 1995; Institute of Medicine 1991; Stuart-
Macadam 1993); prevention of iron deficiency (Calvo and Gnazzo 1990; Oski and
Landaw 1980); and increased body activity and stronger arousal reaction, which are
related to enhanced cognitive and neurodevelopment (Bauer et al. 1991; Fergusson et al.
1982; Lucas et al. 1992; Rogan and Gladen 1993; Temboury et al. 1994). Altogether,
such advantages result in greater adulthood survival of breastfed infants who are not
genetically prone to disease.
A survey of developed countries between the period of 1850-1950 found distinctly
lower mortality rates among breastfeed infants, compared to bottle-fed infants (see
Institute of Medicine 1991: Figure 7-1). In developing countries, exclusive breastfeeding
is also associated with the lowest mortality and infectious disease rates among young
infants (Institute of Medicine 1991). In early life (i.e., first year), shortened breastfeeding
is clearly associated with high infant mortality (Knodel and Kintner 1977).

117
On the other hand, after the first year, the association between breastfeeding and
mortality is inconsistent, with some studies finding no protection against excess mortality
during the second and third years of life, while others indicate continued protection until
the third birthday (Briend et al. 1988; Institute of Medicine 1991).
Importantly, with respect to morbidity and mortality risk, besides providing
antimicrobial, anti-inflammatory and immunologic stimulating agents (Goldman and
Goldblum 1990; Goldman et al. 1990), breastfeeding protects young children against
infectious diseases by limiting exposure to potential pathogens in contaminated foods.
Other cultural factors associated with breastfeeding that reduce morbidity include the
lesser likelihood of breastfeeding mothers to engage in health-risky behavior such as
smoking, which reduces the risk for respiratory infections, and the reduced exposure to
infectious disease that accompanies increased contact with non-maternal caregivers when
an infant is not dependent on breast milk (and mom) for food. As a result of reduced risk
of infection and malnutrition among breastfed infants, in areas of poor sanitation in
developing countries (low income), such infants tend to grow more rapidly than formula-
fed infants in the first six months of life (see Institute of Medicine 1991).
In mothers, prolonged breastfeeding has the immediate effect of increasing the
duration of lactational amenorrhea, which results in increased birth spacing (Anderson
1983; Habicht et al. 1985; Hobcraft 1987; Lee 1980; Short 1983; Thapa et al. 1988).
This reduced fertility benefits both survival of the mother in subsequent pregnancies, as
well as survival of infants (Ellison 1995; Hobcraft 1987; Thapa et al. 1988, among
others). The effects of child spacing are dependent on socioeconomic status, however,

118
with only limited impact on survival in developed countries (Fedrick and Adelstein
1973), and further research is needed for disadvantaged groups.
In addition, breastfeeding appears to benefit the mother by reducing the risk of
reproductive cancers (Byers et al. 1985; Ing et al. 1977; Micozzi 1995; Newcomb et al.
1994; Post 1982; Schneider 1987, among others); and possibly increasing bone mineral
density and lowering the risk of fractures and osteoporosis (Aloia et al. 1983; Cumming
and Klineberg 1993; Feldblum et al. 1992; Hreshchyshyn et al. 1988, among others).
(Note, however, that the evidence for bone density and fracture incidence is not entirely
consistent [e.g., Chan et al. 1982; Hayslip et al. 1989; Institute of Medicine 1991].)
At the same time, while primarily protecting infants, breastfeeding can also be the
mechanism through which infants contract infectious diseases. Depending on hygiene
and pathogen load in the living environment, enteropathogens such as Shigella and
Salmonella can contaminate human milk through the areola and nipples of the breast,
consequently infecting nursing infants (Mata et al. 1967; Wyatt and Mata 1969).
Mothers infected with viruses such as cytomegalovirus, hepatitis B, rubella, human T
lymphocytotropic virus type 1 (HTLV-1) and possibly, human immunodeficiency virus
type I (HIV-1), can also produce breast milk containing such pathogens (Institute of
Medicine 1991).
Besides breast milk transmission of disease, extended nursing can also lead to
increased susceptibility of infants to infectious disease. In this way, exclusive
breastfeeding (or minimal food supplementation) after six months of age, when infants
require additional nutrients for growth and development, can result in malnutrition that
renders children vulnerable to infection.

119
At the same time, infectious disease and malnutrition can prompt mothers to
withdraw solid foods and resume intensive nursing because they often associate illness,
particularly gastrointestinal disorders, with solid food intake (see Dettwyler and Fishman
1992; Fildes 1986; Stuart-Macadam and Dettwyler 1995; Wood 1983). In fact,
undernourished and underdeveloped children are sometimes nursed longer than healthy
infants because mothers associate increased mortality risk with the period after weaning
(Caulfield et al. 1996; Simondon and Simondon 1998). A vicious cycle of chronic
malnutrition and infectious disease can result, often with fatal effects.
Like the synergistic cycle of infectious disease and malnutrition, the ability to
breastfeed adequately is negatively affected by illness (due to physical weakness and/or
separation from mothers), and directly leads to delayed infant development. Furthermore,
with prolonged avoidance of breastfeeding, breast milk production is reduced and it is
often difficult to resume nursing after recovery. Thus, it is often illness that results in
nursing interruption, rather than lack of breastfeeding that leads to infant ill health.
5.3 The Biological Bases for Breastfeeding Duration and Weaning Age
As there are various physiological consequences to changes in weaning patterns,
particularly repercussions that affect survival, the overriding question is “at what age
were most children weaned in the past?”
In a noteworthy study, Dettwyler (1995) carefully examined the primate and cross-
cultural human data to construct a “hominid blueprint” for breastfeeding and weaning
that ensures optimal physical, cognitive and emotional health for infants. Ultimately,

120
these inferences can shed light on the underlying biological basis for human weaning
patterns, irrespective of cultural beliefs. As she states, this question encompasses infant
nursing frequency, timing of solid food introduction, and the age at which breast milk
intake is completely ceased.
Biologically, there are several theories as to when human mothers normally wean
their children. In general, they are based in relation to developmental factors, namely: 1)
tripling or quadrupling of birth weight; 2) attainment of one-third adult weight; 3) 2.71 x
adult female body weight (Harvey and Clutton-Brock 1985); 4) length of gestation; and
5) age at eruption of first permanent molar (see Dettwyler 1995). In terms of
development, weaning is specifically dependent on factors such as mechanical ability
(chewing, swallowing), functional capacity of the gastrointestinal system (esophagus,
stomach, small and large intestines, pancreas) and child appetite.
From the primate data, it appears that compared to primates with small neonates,
those with large neonates tend to have relatively long gestation periods, late weaning age,
late sexual maturity age, large neonatal and adult brain sizes, and a long life span (Harvey
and Clutton-Brock 1985). As large primates with relatively large neonates, humans
would then be expected to have one of the latest weaning ages (Dettwyler 1995). For
instance, among great apes, orangutans breastfeed for 4.21 x gestation length in months;
gorillas, 6.18 x gestation; and chimpanzees, 6.4 x gestation length (Dettwyler 1995: Table
2.3). Drawing from this, Dettwyler (1995: Table 2.3) estimates a minimum weaning age
of 6 x Gestation Length for humans, or 54 months (4.5 years).
Harvey and Clutton-Brock (1985) have compiled various human data to arrive at a
regression formula for calculation of weaning age. Namely, weaning age in days equals

121
2.71 times the adult female body weight in grams. From this equation, a weaning age
range of approximately 2.8 to 3.7 years is suggested, accounting for variation in adult
female body weight. However, Dettwyler (1995: 52) advises that this range represents
the minimum ages at weaning since Harvey and Clutton-Brock used a relatively young
weaning age of 720 days for humans in developing their overall regression for primate
subfamilies (which results in underestimations of weaning age for all primates, including
humans). As it turns out, a late weaning age of around five to six years also coincides
with developmental achievements such as eruption of the first permanent molar (Smith
1989, 1991a,b, 1992), which facilitates food processing, and full adult immune
competence.
With respect to immunology, it is clear that components of breast milk are crucial.
While lactoferrin, an iron-binding protein, prevents the growth of iron-requiring toxigenic
bacteria such as Bacteroides, Clostridium, Escherichia, Salmonella and Staphylococcus
(Weinberg 1994), lymphokines prime the active immune response of children until they
achieve adult immune competence (IgA, IgG, IgM) (Hahn-Zoric et al. 1990; Pabst and
Spady 1990; D. Fredrickson, pers. comm. to Dettwyler 1995). Prolonged breastfeeding
then ensures that children are immunologically capable once weaning occurs. Beyond a
developmental coincidence, Dettwyler (1995: 56) believes that “the fact that children’s
immune systems do not become mature until six years of age is understandable if we
assume that the active immunities provided by breast milk were normally available to the
child until about this age”.
In total, accounting for one-third adult weight, first permanent molar eruption,
gestation length, adult female body weight and birth weight quadrupling, the range of

122
human weaning times is between two and a half and six years of age (see Dettwyler
1995: Figure 2.3). This is supported by a plethora of human studies. A wide variety of
ethnographic data point to weaning ages significantly longer than the current Western
standard, which, for the first time in human history, has replaced maternal breast milk
with artificial substitutes. For example, ancient Babylonians and Egyptians commonly
breastfed up to three years of age (Fildes 1986: Table 15.2); Wickes (1953) reported an
average weaning age of three to four years among various contemporary tribes; Konner
and Worthmann (1980) observed intensive breastfeeding for three to four years among
!Kung hunter-gatherers; a survey of forty-six non-industrialized societies found almost
75% weaned between two and three years of age, 25% at eighteen months and 1% at six
months (see Lawrence 1989); and in the pre-industrial West, children were routinely
breastfed until two to four years of age (see Dettwyler 1995; Fildes 1986, 1995; Stuart-
Macadam 1995). Three to four years is also the same period reported for self-weaning
among children who nurse for as long as they want (Dettwyler 1995).
For the Maya, early colonial and contemporary ethnographic evidence indicate that
pre-contact and later pre-industrial children were extensively breastfed, then weaned onto
atole, a watery maize gruel made by boiling ground (alkali-processed) maize with water
(Behar 1968; Benedict and Steggerda 1937; Landa, in Tozzer 1941). White (1986) notes
that there is no evidence that beans and/or meat supplements were given to children
during the weaning process. However, in addition to atole, which is commonly drunk by
Maya of all ages at breakfast, present-day Guatemalan Ladinos also wean children on
small quantities of softened corn tortillas and black bean broth after six months of age
(Izurieta and Larson-Brown 1995).

123
In general, it is believed that the food habits of rural Maya, principally their reliance
on maize (and atole for weaning infants), have remained more or less stable since ancient
times. Modern surveys indicate that the rural Mexican diet provides 70% of total energy
as carbohydrates (predominantly maize), 11% as protein (beans and some meat), and
19% as fat (see Villalpando et al. 1998). This accords well with Lamanai bone isotopic
evidence of maize consumption reaching 70% of total diet in the Postclassic (White 1986).
This continuity also applies to the extended period of breastfeeding that is
documented in both contemporary and contact period times. In particular, ethnohistorical
evidence from Friar Diego de Landa sheds some light on the childcare practices of 16th
century Maya in the Yucatan (see Tozzer 1941). Yucatec Maya at this time were
described as healthy and robust people who had an adequate and diverse diet. As such,
fecundity was high and mothers traditionally nursed their children for three to four
years (Landa, in Tozzer 1941). In contemporary times, the mean duration of
breastfeeding in indigenous Guatemalan (Maya) communities is two years (Wright and
Schwarcz 1998), while Guatemalan Ladinos breastfeed for an average of 15 months
(Izurieta and Larson-Brown 1995).
While breast milk is considered a nutritionally superior food for infants, compared to
low quality weanling foods, its nutritional value is unquestionably dependent on the
length of lactation. After six to nine postnatal months, breast milk quality significantly
declines. Anemia is a viable health risk when breast milk remains the primary (or
exclusive) food source after an extended period. Iron deficiency is a significant
nutritional problem globally, and even in contemporary times, it is estimated that the
worldwide prevalence of anemia in children between birth and four years of age is 43%,

124
with significant differences between economically developed and underdeveloped
countries (Institute of Medicine 1991: 159). Among Lamanai Maya, hard tissue evidence
suggests that well over one third of the population was anemic at some point in their lives
from the Postclassic period onward (White 1988: 15).
For the Maya, extended breastfeeding and supplementation with iron-poor maize
(which contain iron-binding phytates) may have been a significant factor in poor infant
health. Combined with parasitic and infectious diseases and ensuing diarrhea, an iron
and protein deficient diet fed a synergistic cycle that increasingly compromised infant
health and nutrition. For the Lamanai Maya, the interaction of such factors was
instrumental in making weaning age a critical period of childhood (White 1986; White et
al. 1994).
As stated in Chapter Two, White and colleagues (1994) believe that hard tissue
evidence of chronic infectious diseases, particularly during the Historical period, reflect a
common weaning age, between two and six years, and the period of highest risk for
infectious and parasitic diseases among Lamanai children. Lamanai bone chemical
evidence supports ethnohistorical data that children were weaned onto maize, and further
indicates that children consumed high levels of maize comparable to adults throughout
the Postclassic and Historical periods (White 1986).
All told, overwhelming ancient and contemporary evidence points to a range between
two and five years of age for children to be weaned (Stuart-Macadam and Dettwyler
1995; Fildes 1986; Lawrence 1989, among others). While this wide range is partially the
result of culturally-prescribed customs, it is also reflective of the various human
adaptations to different geographic zones, climates, dietary resources and pathogen loads.

125
5.4 Methods of Identifying Weaning Patterns in the Past
Owing to the importance of early childhood health in the overall health status and
survival of a population, numerous approaches have been adopted to recognize weaning
patterns in ancient populations. Traditional methods have examined infant mortality rate
and paleodemography (Clarke 1980; Cook 1976; Herring et al. 1998; Katzenberg et al.
1996; Knodel and Kintner 1977; Lallo 1973; Saunders and Herring 1995; Swedlund
1990); deciduous tooth wear (Farnum and Benfer 1995; Molleson 1995); and non-
specific hard tissue indicators of physiological stress, e.g., enamel hypoplasia (Corruccini
et al. 1985; Hillson 1979; Blakey et al. 1994; Farnum and Benfer 1995; Goodman et al.
1984; Goodman and Armelagos 1988, 1989; Lanphear 1990; Moggi-Cecchi et al. 1994;
Song 1997; Wood 1996; Wright 1990). However, the suitability of such studies has been
complicated by 1) methodological issues; 2) variations in hard tissue development; 3)
conflicting results, i.e., contradicting documented weaning age; and 4) the biased nature
of ancient burial samples (i.e., population misrepresentation, particularly of subadults [a
sample that has died prematurely], and true skeletal “health” [see Wood 1992]).
To better assess the effects of dietary behavior on hard tissues, chemical methods of
hard tissue analysis have been applied to trace breast milk intake and weaning. Such
studies have focused on isotopes of nitrogen (Dupras et al. 2001; Fogel et al. 1989;
Herring et al. 1998; Katzenberg and Pfeiffer 1995; Katzenberg et al. 1993, 1996;
Richards et al. 2002; Schurr 1997, 1998; Tuross and Fogel 1994; White and Schwarcz
1994), carbon and oxygen (Dupras et al. 2001; Richards et al. 2002; Wright and

126
Schwarcz 1998); relative levels of zinc (Farnum and Benfer 1995; Grupe 1986); and
strontium/calcium ratios (Huhne-Osterloh and Grupe 1989; Sillen and Smith 1984).
Stable nitrogen isotopes have been used to infer changes in infant dietary behavior
because proteins in breast milk and solid foods differ in their nitrogen isotopic
composition. Importantly, the nitrogen stable isotope values of an organism’s tissues
(15
N) have been demonstrated to reflect the nitrogen isotope values of its diet (DeNiro
and Epstein 1981). Like strontium, there is a trophic level effect for nitrogen isotope
ratios in animals within an ecosystem, so that a carnivore’s tissues will have protein
nitrogen values approximately 3‰ higher than its prey (Schoeninger and DeNiro 1984).
In the food web, breastfed infants exist one trophic level above their lactating mothers
because they are consuming their mother’s tissues in the form of breast milk, which is a
good source of nitrogen. (At birth, infants will have 15
N values comparable to their
mothers.) With nursing, protein in infant tissues should be enriched by 2-4‰ in the
stable isotope of nitrogen (15
N), compared to their mothers. With the cessation of breast
milk intake, weaning is identified by a decline in 15
N of bone collagen back to adult
levels (Fogel et al. 1989; Katzenberg et al. 1993, 1996; Katzenberg and Pfeiffer 1995;
White and Schwarcz 1994).
This hypothesis has been tested by Fogel, Tuross and Owsley (1989) using fingernails
of contemporary infants and their mothers, in addition to bones of ancient infants.
Fingernail evidence indicate that breast milk intake is indeed reflected as nitrogen
enrichment, and that this enrichment is still apparent several months after the onset of
food supplementation. Nitrogen enrichment (trophic level shift) averaged 2.4‰ more
than mothers’ values, and was evident throughout the period of exclusive breastfeeding.

127
With supplementation (infant formula, dairy products, cereals), however, there began a
decrease in infant 15
N values toward those of mothers (Fogel et al. 1989).
Furthermore, bone collagen from prehistoric/protohistoric Tennessee Valley infants
(one year olds) also display 15
N enrichment (Fogel et al. 1989). Later, between eighteen
and twenty months of age, 15
N values decline significantly, which the authors interpret
to be evidence of non-breast milk consumption (cessation of breast milk as the main
source of protein).
Nitrogen enrichment associated with breast milk intake has also been observed in the
bones of historic period children from the Sully Site, South Dakota (A.D. 1650 to 1733)
(Tuross and Fogel 1994). In children between three months and two years of age, 15
N
was on average 1.6‰ enriched relative to adult bone remains. The highest bone collagen
15
N values (12.8‰) characterize the three-month to two year old group, while children
thereafter, aged two to five years, have a decreased 15
N value of 10.2‰ (Tuross and
Fogel 1994). From their sample, the authors conclude that for at least the first year,
infants were exclusively breastfed without significant supplementation of other nitrogen
sources. Weaning is thought to have occurred between the ages of two and six years.
Katzenberg and colleagues (1993) similarly found enriched 15
N values in infants
when compared to their mothers in Ontario Iroquoian samples dated to A.D. 1530-1580.
In particular, infants younger than two years had 15
N values 2-3‰ higher than adults.
This parallels ethnohistorical accounts of Huron groups nursing infants until two to three
years of age (Katzenberg et al. 1993). (In contrast, 19th
century Canadian infants of

128
European descent were shown by 15
N values to cease breastfeeding at around one year
of age, which was supported by historical data [Katzenberg and Pfeiffer 1995]).
Like indigenous groups throughout the Americas, children were weaned on the staple
maize, which is reflected in high 13
C values among older infants and young children
(Katzenberg et al. 1993). Similarly, infants whose mothers could not breastfeed, or those
whose mothers died in childbirth, were likely mostly consuming maize, which would be
indicated by relatively high 13
C values and 15
N values similar to those of adults
(Katzenberg et al. 1993).
Combining data from bone, skin, muscle and hair of prehistoric Nubians, White and
Schwarcz (1994) also found age-related differences in 15
N values that are attributed to
an extended period of breast milk intake. With a significant negative correlation between
15
N and age, they found a 3‰ enrichment in children aged from birth to six years
compared to adults. With respect to weaning, their observations of a gradual decrease in
15
N over the first six years of childhood point to a slow, extended, process, rather than a
sudden shift from total breast milk reliance to complete non-milk food intake.
In the case of Middle Mississippian maize horticulturalists from the Ohio River
Valley, an increase in 15
N values commenced after birth until it reached the highest
value between one and three years of age (Schurr 1997). The author attributes the
nitrogen enrichment to substantial breast milk intake during at least the first two years of
life, while the subsequent decline in nitrogen values indicate increasing consumption of
non-breast milk proteins from a weanling diet. Furthermore, based on Schurr’s non-
linear model, which is more accurate in tracing changes (nitrogen decline) related to
supplementation and weaning, the weaning process was a gradual one that began before

129
the age of two years. For at least six months after the start of supplementation, breast
milk continued to provide the bulk of dietary protein (Schurr 1997).
As indicated by these nitrogen studies, bone chemical composition yields a rather
broad time frame for changes in dietary intake. Beyond identifying the start of
supplementation as before two years of age (Fogel et al. 1989; Katzenberg et al. 1993;
Schurr 1997; Tuross and Fogel 1994) and continued breast milk intake until five or six
years of age (Tuross and Fogel 1994; White and Schwarcz 1994), finer details of the
weaning process are imperceptible. It is maintained that such general indications are
inadequate, especially since ethnographic data can already account for such a wide range
(see above).
Additionally, in order to adequately sample the breadth of childhood that experiences
breast milk intake and weaning, i.e., to trace the subtle changes of the weaning process,
skeletons of more infant categories are required. Unfortunately, three obstacles challenge
this requirement: 1) the nature of post-depositional environments to deteriorate
undermineralized subadult skeletons; 2) the differential interment of children in the
archaeological record; and 3) the degree of error in traditional methods of ageing skeletal
remains.
Wright and Schwarcz (1998) have also experimented with stable isotopic signals of
carbon and oxygen in enamel to trace weaning. Their research represents the first
attempt to infer childhood dietary behavior from the chemistry of dental hard tissues.

130
Subsequent analyses by Richards and colleagues (2002) on the nitrogen and carbon
isotopic composition of tooth dentine reflect the current research shift to dental tissues.1
In Wright and Schwarcz’s (1998) investigation, the recognition of weaning is based
on the fact that the ratio of 18
O and 16
O in body tissues reflects the source of water
imbibed as a liquid, and to a lesser degree, the oxygen obtained from food. As human
breast milk is derived from the body water pool, it is more enriched in 18
O than the
water ingested by a lactating mother (but reflective of maternal diet). Oxygen isotope
values will reflect differences in water intake attributed to breast milk versus non-breast
milk liquid. Thus, hard tissues of breastfed infants will be heavier in 18
O than those of
bottle-fed infants (Wright and Schwarcz 1998). For breastfed infants, declining breast
milk intake and supplementation with different water sources will be exhibited as lower
18
O values in later developing enamel.
Conversely, relative values of 13
C in enamel reflect processes of dietary
supplementation, particularly maize, which is enriched in 13
C. The consumption of maize
gruels prior to complete weaning would be identified as an increase in 13
C in enamel
forming at the time of supplementation.
According to Wright and Schwarcz (1998), 13
C and 18
O values of enamel material
point to a long, gradual, transition from breast milk to solid foods for most children at
Kaminaljuyu. Enriched 13
C values of premolars (which develop between two and six
1 Their comparison of dentine and bone isotopic composition indicates that tooth dentine in children up to 11 years of age is
enriched in δ15N compared to adult rib values, with a significant portion of dentine formed during breastfeeding, with little, or no
turnover since (Richards et al. 2002). Juvenile rib collagen δ15N decreases to adult levels after the age of two years, suggesting that
weaning occurred at this age. After weaning, for several years, juvenile rib 15N is decreased compared to adult rib values, indicating that children’s diet was lower in animal protein than the diet of adults. Importantly, they maintain that the delay between the cessation
of breastfeeding and its chemical expression in bone collagen is still not fully understood (Richards et al. 2002: 206).

131
years of age) suggest that children two to six years of age were eating more maize foods
than they had eaten as infants, which the authors expect if maize gruel formed the
weanling diet (Wright and Schwarcz 1998). Based on their results, they propose that
Kaminaljuyu children began solid food supplementation by at least one year of age, with
substantial amounts of solid food ingested by two years. Nevertheless, breast milk intake
probably continued until five or six years of age, as indicated by 18
O values in premolars,
pointing to a relatively late weaning age (Wright and Schwarcz 1998).
Recognizing the investigative nature of such research, the authors also present several
factors unrelated to weaning that might cause changes in chemical composition (18
O).
These include higher metabolic rates of young children, changing activity patterns with
age, or more rapid water usage related to smaller body size. In this case, body size and
metabolic rate are known to affect the magnitude of isotopic fluxes and fractionation
(Wright and Schwarcz 1998: 15).
Unfortunately, due to the total sampling (dissolution) of enamel from teeth of
different developmental age (first molar, premolar, third molar), which Wright and
Schwarcz utilize to infer chemistry changes through time, a precise assessment of
weaning age is not possible. Their dental analysis controls for issues plaguing bone
chemical studies, i.e., bone turnover and hard tissue contamination, but this method does
not compare to the subtle time-depth sampling capabilities of LA-ICP-MS (see Chapter
Six).

132
5.5 Hard Tissue Strontium and the Inferences for Infant Nutrition
The present study addresses many challenges in paleonutritional chemistry by
analyzing time-sensitive dental hard tissues utilizing localized laser (micro)sampling
of enamel across known periods of development. Both features make this analysis
unique in reconstructing diet and weaning behavior of ancient children at specific ages of
development.
Particular attention is on strontium, in the form of strontium-calcium (Sr/Ca) ratios,
because this element is a reliable hard tissue indicator of dietary Sr/Ca (see Chapter
Four). Historically, such research has predominantly centered on the usefulness of
strontium to indicate degrees of herbivory or carnivory among human groups, but its
ability to also trace infant breast milk intake, relative to food supplementation, and
weaning is postulated in this study. The present study builds on the work of Sillen and
Smith (1984), who examined the Sr/Ca ratios of juvenile skeletons to determine the
timing of dietary supplementation until the age of weaning.
Strontium is used to infer the process of weaning because the Sr/Ca ratio of solid
foods is relatively high compared to the Sr/Ca ratio of calcium-rich human milk, where it
is practically non-existent (Schroeder et al. 1972; Sillen and Smith 1984). Comparatively,
breast milk exhibits Sr/Ca values ranging from 0.14-0.35 x 10-3
(Harrison et al. 1965;
Krachler et al. 1999; Rehnberg et al. 1969; Rosenthal 1981), while food such as maize
has significantly higher ratios of 1.129 x 10-3
(Wright 1994), 2 x 10-3
(LaVigne 2002) and
even 7.14 x 10-3
(Shacklette 1980; Watt and Merrill 1963), for example, which is
dependent on geological conditions.

133
In truth, strontium content varies minimally in dental tissues. Still, some variation in
strontium levels reflects dietary and age-related changes in humans. These are due to
variations in gastrointestinal discrimination, food and water sources, and most
importantly, for this study, its absence in human milk.
During fetal development, strontium is discriminated by the placental barrier. In
newborns, this fact combines with discrimination at the mother’s mammary gland to
produce the lowest Sr/Ca ratio in humans (Comar et al. 1957). This lower Sr/Ca ratio in
young mammals may be due to the high calcium demands during rapid growth in infants
(Sillen and Kavanagh 1982).
Among infants, Sr is increasingly discriminated against in favor of Ca due to
maturation of the kidneys and digestive system after the first year of life (Sillen and
Kavanagh 1982). This is reflected in high OR values (ratio of Sr/Ca in infant bones to
Sr/Ca in diet) among young infants that steadily decrease with age: 1.0 among neonates;
0.6 at one year; 0.25 at 3 years; 0.23 at 5 years; 0.18 at 10 years of age and adulthood
(Sillen and Smith 1984: Table 1).
During the process of dietary supplementation, infants consume solid foods and non-
breast milk liquids that have Sr/Ca ratios significantly higher than breast milk. Thus, low
Sr/Ca ratios in infant remains could be interpreted as evidence of exclusive breast milk
intake and lack of dietary supplementation. Increasing Sr/Ca ratios will reflect the entire
process of the introduction of dietary supplementation until weaning. (This differs from
nitrogen stable isotope ratios of hard tissue collagen, which reflect the cessation of
breastfeeding.)

134
Maximum bone Sr has also been found by Grupe (1986) to be associated with
minimum Zn levels among young children of weaning age, suggesting that Zn might be
an indicator of weaning stress. While Farnum and Benfer (1995) could find no statistical
change in Sr due to bone contamination, they found decreasing Zn levels over time in
young children, particularly in mid to late childhood. However, they could not determine
if low Zn levels were due to weaning stress due to prolonged breastfeeding and
inadequate protein supplementation, or stress due to other factors.
In their important study, Sillen and Smith (1983, 1984) found that Sr/Ca values of
medieval Arab bones gradually increase after birth, with individuals aged 1.5 to 3.5 years
having the highest values for the entire population (see Table 5.2). The authors note that
this is consistent with ethnographic data of traditional Palestinian Arabs who continue to
breastfeed until two to three years of age. Thereafter, Sr/Ca values decline to levels that
persist throughout adulthood (Sillen and Smith 1984). Assuming a consistent adult diet,
these lower values reflect the discrimination against strontium that occurs with age in the
mature (adult) gastrointestinal tract.
Table 5.2 Bone Sr/Ca changes from the Dor sample, Israel
(estimated Sillen and Smith 1984 data, in Katzenberg et al. 1996)
Age (years) N
Sr/Ca
(x 10-3
)
Range
Birth
2
1.50
1.4-1.6
0.5 3 1.75 1.5-2.0
1 10 2.25 1.5-3.0
2 2 2.50 1.7-2.9
3 5 2.25 1.8-2.8
8 11 1.60 1.3-2.3
Adult 1.80

135
Huhne-Osterloh and Grupe (1989) similarly examined strontium concentration in
ancient (German) bones to estimate weaning age. They found the highest Sr/Ca ratios in
children aged six months to two years, the period for which they postulate dietary
supplementation and eventual weaning. Like Sillen and Smith (1984), bone strontium
levels decline in subsequent age groups (see Table 5.3).
Table 5.3 Bone Sr/Ca changes from the Schleswig sample, Germany
(data from Grupe 1986; Huhne-Osterloh and Grupe 1989;
derived from Katzenberg et al. 1996: Table 2)
Age
(in years)
N
(individuals)
Sr/Ca
(x 10-3
)
S.D.
(total sample)
Birth-0.5
6
0.58
2 5 0.72
4 3 0.63
14 6 0.51
Adult 78 0.53
0.16
Most recently, Mays (2003) investigated bone Sr/Ca to determine the duration of
breastfeeding in an English Medieval sample. Likewise, bone Sr/Ca indicated significant
age-related changes that could be associated with breastfeeding, supplementation and
weaning. In particular, significant Sr/Ca increase between one and two years of age is
interpreted as representing the period of significant food supplementation (Mays 2003).
Reflecting immature digestive systems, infants and young children (under two years)
exhibit higher Sr/Ca ratios than older individuals and adults (Mays 2003: 734). In this
sample, a steady decline in Sr/Ca from about two to eight years of age, when values then
resemble adults, indicates that most children in this sample were fully weaned by two.

136
Preliminary analyses of archaeological teeth in the Hampshire College Natural
Science lab suggest that weaning can also be recognized in strontium levels quantified by
LA-ICP-MS analysis of enamel. Ablation results of several ancient Anasazi canines
indicate increased Sr/Ca ratios in the mid-crown regions (Perez 1998). This increase,
occurring around three to four years of age, may indicate a trophic level shift associated
with dietary changes during a relatively late weaning period, when solid food,
presumably maize, consumption increased relative to breast milk. This finding is also
observed by Wright and Schwarcz (1998) in stable carbon and oxygen isotope analyses
of dental enamel for other Maya samples.
The combined evidence suggests that hard tissue strontium analysis is useful for
tracking breastfeeding, food supplementation and weaning. But while Sillen and Smith
(1984) first postulated this model almost twenty years ago, it has generally been
overlooked in paleonutritional research, which has largely concentrated on isotopic
analysis. Mays’ (2003) recent findings are the most notable since Huhne-Osterloh and
Grupe (1989). Moreover, diet-related changes in hard tissue strontium have never been
examined in human dental remains. This fact makes this research exciting and
worthwhile to the field of paleonutrition.
5.6 Enamel Sr/Ca and Infant Nutrition at Lamanai
As indicated by ethnohistorical sources, hard tissue pathology and enamel isotopic
data, it is likely that ancient Maya mothers breastfed their infants for periods of up to four
or five years. While supplementation with non-breast milk foods (maize) likely began

137
after six months of age, due to diminished nutritional quality of breast milk, prolonged
nursing is within the norms of many past and present human populations.
Chemically, this pattern of infant feeding can be characterized by 1) initial low Sr/Ca
values reflecting exclusive breastfeeding, followed by 2) a significant increase signaling
the start of supplementation, that then assumes 3) a gradual increase over the extended
time period that represents prolonged breast milk consumption, concluded by 4)
stabilization and slight reduction of Sr/Ca values to levels comparable to adults. Figure
5.1 illustrates such a model of Sr/Ca change during early childhood, which incorporates
information on human strontium absorption (Comar 1963; Rivera and Harley 1965;
Sillen and Kavanagh 1982), general patterns of infant food supplementation and weaning
(see above), and the original premise of Sillen and Smith’s (1984) research.
Figure 5.1 Proposed model of Sr/Ca behavior during infancy and early childhood
0 1 2 3 4 5
Age in years
Sr/
Ca R
ati
o

138
Bone chemical evidence suggests that Historical Maya at Lamanai consumed the
same variety and proportion of foods as their Postclassic ancestors, with children
continuing to consume comparably high levels of maize (two-thirds of total diet) as their
parents (White 1986). In this case, nutritional stability might also extend to infant
feeding and weaning patterns.
However, while details of the Maya menu were maintained, it is also possible that
food availability and eating patterns were affected. Amidst an unsettled colonial social
climate and a significant increase in morbidity and mortality among Lamanai Maya, it is
postulated that Spanish demands for tribute negatively affected the health and nutrition of
Maya infants through economic pressures on their mothers (Chapter Two). Potential
physiological consequences include reduced breastfeeding, reduced weaning age (with
potential morbidity and mortality risks), and even, changes in population birth rates and
female survival, since extended nursing can reduce fertility and increase the time period
between conceptions.
Assuming colonial changes were significant enough to impact weaning behavior (i.e.,
breastfeeding duration) and children’s health, Historical period enamel Sr/Ca could
appear as a condensed version of Postclassic patterns. Several models are possible in this
case: 1) supplementation might have begun earlier, which would be reflected in earlier
enamel Sr/Ca increases, but was followed by continued long-term nursing; 2)
supplementation might have begun earlier, but was followed by a faster rate of breast
milk cessation (earlier weaning age), which would appear as a steeper slope graphically;
3) age of supplementation might have remained constant but the rate of food
supplementation increased (earlier weaning age).

139
In contrast, social conditions at Lamanai may have affected infant health by reducing
the consistency of available weanling foods (maize gruel). In this case, food
supplementation (Sr/Ca increase) may have been erratic or delayed because of seasonal
or occasional disruptions in maize production and distribution, as well as infant and
maternal illness.
In the colonial environment of epidemics and famines, it is likely that more Maya
mothers perished than during the Postclassic. In such cases, breast milk intake would
have ended abruptly and orphaned infants might not have had the fortune of a substitute
wet nurse. Immediate total reliance on weanling foods would characterize such infants
and would appear chemically as a substantial increase in Sr/Ca over a very brief period.
Such infants might be recognized as those with the steepest Sr/Ca slopes over time.
Considering that early nutrition has major repercussions for later survival, the ability
to reconstruct subtle dietary changes during weaning via LA-ICP-MS of enamel provides
an unprecedented detailed account of the transition from infant to adult diet. Application
of microsampling techniques preserves the general integrity of fossilized remains and
precisely captures the time-sensitive chemical data that is inherent in sequentially formed
enamel. More so than bone, teeth provide a hard tissue biography of ancient childhood
nutrition that is minimally affected by taphonomic processes and are rich reservoirs of
element composition clearly associated with dietary intake, which can be attributed to
known developmental age. Teeth are thus the ideal hard tissues for reconstructing the
weaning process among ancient populations.

140
CHAPTER 6
MATERIALS AND METHODS
6.1 Analytical Technique
This study broadens the range of bioarchaeological questions that can be asked of
human remains by utilizing Laser Ablation-Inductively Coupled Plasma-Mass
Spectrometry (LA-ICP-MS). Specifically, LA-ICP-MS analysis of ancient human
enamel is used to provide meaningful inferences for dietary intake during early
childhood, the period when teeth are formed. The ability of LA-ICP-MS to sample
enamel of known developmental age enables study of important life events such as the
timing of infant dietary transitions and their nutritional consequences. With this
technology, this research establishes a method of paleonutritional reconstruction that
addresses two key challenges in hard tissue analysis: 1) the isolation of chemical
composition to precise developmental age, and 2) minimal destruction of invaluable
human remains.
LA-ICP-MS involves the coupling of Laser Ablation (LA) sampling with Inductively
Coupled Plasma-Mass Spectrometry (or Spectroscopy) (ICP-MS), which is a technique
of multi-element analysis that quantifies element concentrations based on atomic mass to
charge (m/z) ratio. Laser ablation technology has been applied to two major areas of
research: 1) bulk analysis, which involves low spatial resolution (80-350 m crater
diameters) and 2) local analysis, such as this one, which has high spatial resolution (4-80
m crater diameters) (see Gunther et al. 2000).

141
In this study, enamel is microsampled by localized laser energy, rather than by total
acid dissolution. It offers several advantages over conventional methods of chemical
analysis, namely rapid, multielement and isotopic analyses, low detection limits (high
sensitivity), minimal sample preparation (and low cost), high spatial resolution, high
sample throughput capability, and importantly, for studies of archaeological specimens,
minimal sample destruction (Denoyer et al. 1991; Evans et al. 1995; Gray 1985; Kang et
al. 2004; Outridge 1996; Outridge et al. 1995; Perkins et al. 1991; Ward et al. 1992).
Like ICP-AES, ICP-MS is an ideal technique for the quantification of trace elemental
composition of biological tissues, environmental, geological and food samples.
To date, this technology has been effective in tracing environmental pollution, animal
habitat origins and migrations, human migration, hard tissue preservation, archaeological
materials provenance, element profiling, fetal and infant nutrition, and even forensic
science (Ward et al. 1992; Fuge et al. 1993; Wang et al. 1994; Evans et al. 1994, 1995;
Kang et al. 2004; Outridge et al. 1995; Cox et al. 1996; Outridge 1996; Watmough et al.
1997; Budd et al. 1998; Lee et al. 1999; Watling 1999; Belloto and Miekeley 2000;
Gratuze et al. 2000; Haverkort 2001; Prohaska et al. 2002; Goodman et al. 2003; Neff
2003, among others). Utilized materials for such studies include animal and human
bones and teeth, tusks and soft tissues, mollusks, fish otoliths (inner ear stones), scales,
tree rings, ancient pottery, gold, cannabis leaves and crime scene artifacts.
Briefly, laser ablation sampling entails directing a high energy pulsed laser beam
(LA) onto the surface of a solid sample housed in a vacuum-sealed chamber. Upon
contact, the solid sample becomes vaporized, and this ablated material is then swept with
a flow of argon gas into an inductively coupled argon plasma (ICAP) torch. Here, single

142
charged ions are produced from the vaporized sample by excitation, at temperatures
between 8000K and 10,000K, and then extracted by the mass spectrometer (MS) for
detection. A quadrupole mass filter in the mass spectrometer separates analytes
according to atomic mass-to-charge (m/z) ratios (see Denoyer et al. 1991; Gray 1985;
Ward et al. 1992). Visually, element abundance across the sampled transect can be
reported as a line graph with a y-axis of “element intensity”, and an x-axis of “reading #”
or “time in seconds”, which can be correlated to distance on the tooth (see Figure 6.1).
Figure 6.1 Tooth cross-section indicating correlation of laser ablation position,
development age and enamel Sr/Ca intensity ratio

143
While ICP-MS has been widely used to analyze samples prepared by acid digestion,
its coupling with LA technology represents a novel approach to sampling. In total,
combining these technologies takes advantage of the benefits and capabilities of both:
localized microsampling and surface profiling by LA and extremely sensitive,
quantitative, multielement analyses by ICP-MS.
Firstly, laser ablation sampling eliminates the difficulties involved with enamel
separation that characterize “crude” techniques such as scraping and drilling, which often
also gouge dentine material. As well, it avoids questions of quantity and developmental
age of sampled enamel that plague acid digestion techniques. Further, both physical and
acid removal of enamel can result in combined sampling of both contaminated surface
enamel and more intact interior enamel. In creating an ablation trench only 50 m wide
and deep, any limitations with enamel collection are strictly due to operator error or
problems with dental material identification.
Sampling of enamel by laser ablation requires minimal sample preparation, as the
material can be analyzed practically intact. The main challenge with human dental
material involves preparing an internal enamel plane that is as level and developmentally
complete as possible, which is generally along the central axis of teeth. Compared to
methods of chemical analysis that involve acid digestion, the preparation for laser
ablation sampling entails minimal contamination risk.
LA-ICP-MS also diminishes the degree of polyatomic interference that can arise in
conventional solution nebulization (SN)-ICP-MS. “Dry (argon) plasma” used in LA-
ICP-MS reduces the likelihood of the formation of 40
Ar16
O+,
15N
16O
+ and
16O2H
+, which
can result from the interaction of water and HNO3 acid (Durrant 1999).

144
Significantly, the application of LA microsampling technology reveals a level of
intra-tooth variation that is not captured by other methods. The coupling of laser ablation
sampling to ICP-MS is highly amenable to dental analysis as it can “read” chemical
markers across the tooth surface, producing a “map” of chemical signatures specific to
tooth location (see Figure 6.1). Due to the tree-ring-like nature of teeth, which allows
association of tooth location with developmental age (see Massler et al. 1941), chemical
profiles produced by LA-ICP-MS disclose a wealth of data documenting health and
nutrition that can be attributed to specific periods, e.g., fetal development or infant
weaning. The nature of health and nutrition during these periods of childhood are
particularly significant and predictive of future adult survival (see Cook and Buikstra
1979; Goodman and Armelagos 1988; Goodman et al. 1980, among others).
In the words of Outridge and co-workers (1995: 167):
LA-ICP-MS and other microprobe techniques are thus revealing that the
heterogeneity of metal distribution at microspatial scales is no less of a sampling
design problem than for macroscale entities such as forests, oceans, etc., and entails
similar types of statistical considerations, data management, and visual
representation.
6.2 Technological and Methodological Challenges
While LA-ICP-MS has great potential, there remain several limitations to its effective
use on biological material. To date, (LA) ICP-MS is only semi-quantitative due to the
unavailability of matrix-matched standards (see Amarasiriwardena et al. 1997; Evans et
al. 1994; Outridge et al. 1995). Results of quantitative analyses with a range (or
accuracy) of +/- 30-50% are considered semi-quantitative (Amarasiriwardena et al. 1997),

145
while determinations with accuracy of less than +/-10% are quantitative (Ahrens and
Taylor 1961; Wang et al. 1972).
Lacking a suitable reference standard for enamel, external standards (e.g., glass NIST
610, 612, 614) have been used to calibrate LA-ICP-MS results of teeth and carbonates
(e.g., Durrant 1999), but this is still limited by a matrix mismatch between hard tissue
apatite and glass (see below).
Difficulty with calibration led Cox and colleagues (1996: 256) to conclude:
As no suitable standard reference materials are currently available for teeth, and in
view of the requirement for qualitative data and for inter-sample comparison rather
than absolute quantitation, we suggest that the principle of element/element ratioing
be adopted.
Likewise, Ward, Durrant and Gray (1992: 1145) state: “as it is not known in advance
whether or not internal standardization is necessary, its use may be advisable as a matter
of policy”. In this case, calcium is an appropriate major element. Budd and colleagues
(1998) maintain that, owing to the slight effect of diagenetic substitutions on the
comparatively elevated level of calcium in hard tissues, calcium provides the best internal
standard for such chemical analyses.
According to Denoyer (1992), if matrix-matched, or internal standards are used, an
improved accuracy, or range, of +/-20-50% is possible for chemical determinations with
LA-ICP-MS. Outridge and colleagues (1995) estimate that with internal standards and
optimal instrumental conditions, the analytical precision of LA-ICP-MS can be less than
10% for elements between 0.1 and 1 ppm, and less than 5% for elements with abundance
exceeding 1 ppm, such as strontium. While still semi-quantitative, its application to hard
tissue analysis remains worthwhile because normalizing with internal elements provides

146
invaluable relative concentration data. In this study, strontium levels will be interpreted
relative to simultaneously collected calcium abundance. As calcium is present in the
enamel matrix at high, relatively stable levels, it is an excellent choice for an internal
standard (see Cox et al. 1996).
Recent research utilizing LA-ICP-MS has focused on improving quantification
procedures, such as matrix-independent calibration methods using external solid
reference materials, which have increased in number (see Gunther et al. 2000). Gunther
and colleagues (2000: 4) note that with improvements in calibration and internal
standardization, LA-ICP-MS can yield accurate results between +/- <5 to 25% for
element concentrations.
The use of external standards is hampered by matrix differences, however. In
particular, the efficiency of laser ablation sampling is poor for glass discs (NIST), which
are often used for dental calibration, compared to pressed powder carbonate materials
(Bellotto and Miekeley 2000; Durrant 1999). Under the same laser conditions, crater
shape, depth and morphology will differ significantly between glass and carbonates,
resulting in distinct ablated masses and element results (Bellotto and Miekeley 2000).
Glass standard calibration curves will have lower sensitivity and there will be an
overestimation of trace element concentrations in carbonate matrix samples (Bellotto and
Miekeley 2000: 638).
Quantification of LA-ICP-MS results remains a research concern because of several
factors: uneven sample ablation, non-representative sub-sampling, differences in
transport efficiencies of material components to the plasma and, importantly, lack of

147
suitable matrix-matched standards for calibration (Durrant 1999; Bellotto and Miekeley
2000; Gunther et al. 2000)
Additional challenges include lack of homogeneity within an increment of analyte
(Outridge et al. 1995) and non-uniform ablation sampling (Evans et al. 1995). Non-
uniformity of samples is either due to variable amounts of tooth material vaporization
(due to tissue density or laser intensity), or differential amounts entering the sample
stream (Evans et al. 1995). Watmough and colleagues (1997) observed non-uniformity
for various types of wood, calling for standards specific to the analyte. It is assumed such
differential ablation is due to variations in material composition and density, and if so,
this has significant implications for LA-ICP-MS of human teeth. However, in this case,
normalizing with calcium resolves some issues of differential sampling (variation).
Fortunately, preliminary results of laser ablations done within growth bands of an
ancient canine suggest that there is general elemental homogeneity (Goodman and Song,
unpublished data), which has also been observed in deciduous teeth (Dolphin et al., in
press; Goodman et al. 2003; Kang et al. 2004). Nonetheless, it is believed that the
advantages of minimal contamination, tooth preservation and precise enamel sampling
far outweigh the possibility of sampling anomalous enamel. In general, it is widely
believed that enamel within the same growth band exhibit compositional homogeneity.
Recent developments in LA-ICP-MS research include use of laser systems with
shorter wavelengths (1064 nm, as opposed to 266 nm used in this study and many
others); UV gas (excimer) lasers; changes in laser pulse to pulse stability and the optical
arrangement of beam transfer optics; experiments with sampling instrumentation (e.g.,
ablation cells); mechanisms of sample transport (e.g., mixed gas sample introduction);

148
ICP-MS measurement (detector) efficiency; direct solution ablation; and improvements
in quantitative analysis, among others (see Gunther et al. 2000). Such research has been
geared toward improving sensitivity and sample transport, stability of ICP conditions,
reducing element fractionation (“non-sample related variation of the analyte response
during ablation” [Gunther et al. 2000: 4]) and simplifying instrumentation set-up and
operation.
As a result, LA-ICP-MS results (i.e., analyte sensitivity and limits of detection) have
been found to be influenced by crater dimensions (diameter to depth ratio), energy
density, repetition rate, laser focal point (rather than wavelength) and background noise,
which all affect the amount of ablated and transported material to the ICP and are
important factors in elemental fractionation (see Durrant 1999; Gratuze et al. 2001;
Gunther et al. 2000).
Recent research suggests that variation in ablation yield, decreased signal and
elemental fractionation can be limited by 1) applying raster ablations, rather than single
spots; coupled with 2) relatively large crater size (100 μm) (Neff 2003); 3) collecting data
in three or more different ablation runs (Gratuze et al. 2001); and 4) use of a 213 nm
wavelength laser instead of 266 nm (Gunther and Hattendorf 2001; Jackson 2001). Still,
it is a challenge to distinguish whether the sources of fractionation are related to the
ablation process, the transportation process and/or the ICP excitation and ionization
process (Gunther et al. 2000).

149
In surveying the literature, tremendous variation exists in the preparation and
collection of dental material for chemical analysis. This variation includes: 1) the type
of mechanical separation and localization of dental tissues (e.g., burr drilling, chipping,
scraping); 2) choice of teeth, and whether they are analyzed individually or together; 3)
region of dental tissue sampled, since surface enamel and root dentine exhibit higher
levels of contaminating elements such as lead; 4) duration and concentration of acid
solutions used to clean teeth; and 5) analytical technique employed, which has resulted in
diverse findings.
A significant source of variation concerns sample decontamination. Preparation of
hard tissue samples requires utmost care because of the present understanding of
diagenesis. Evidence by researchers such as Ambrose (1990), DeNiro and Epstein
(1981), DeNiro and Hastorf (1985), Farnsworth et al. (1985), Kyle (1986), and Lambert
et al. (1979, 1982, 1985), among many others, have demonstrated that post-mortem
contamination of hard tissue elements and stable isotopes can significantly alter chemical
values, and consequently, dietary reconstruction. Diagenetic mechanisms include
deposition, leaching or substitution of elements and other components (lipids, carbonates,
humic acid), as well as extraneous factors that degrade overall tissue composition (soil
pH, water solution, temperature).
An initial assessment of chemical integrity can be based on overall appearance (see
Chapter Three), but such discrimination should only be applied site-specifically, since the
gross, morphological appearance of bone is not necessarily a good indicator of chemical
integrity (e.g., Klepinger et al. 1986). Diagenetic change also tends not to be a

150
predictable function of duration of interment (Klepinger et al. 1986) (although see
below).
In teeth, gross structural integrity, as reflected in intact crowns, consistent ivory color,
minimal attrition and disease (caries, hypocalcification) will be more likely associated
with good chemical preservation. Post-mortem alteration of dental organic material is
more time-dependent than apatite crystallite diagenesis (Carlson 1990: 545). As dental
enamel is predominantly inorganic, it is the least susceptible to post-burial contamination,
particularly regarding aluminum, iron, strontium and barium (Kyle 1986; Parker and
Toots 1970, 1980). The only limitation with dental enamel is that its composition will
only reflect trace elements circulating at the time of enamel formation (childhood).
In order to circumvent the effects of diagenesis on fossil remains, especially for the
purposes of chemical analysis, numerous methods have been devised to remove
contaminating substances prior to analysis. The simplest method, as described by
Szpunar et al. (1978) involves simple brushing and washing with distilled/deionized
water. This fundamental process only removes clinging surface contaminants, as does
ultrasonic cleaning in high-purity water (e.g., Millipore Alpha Q H2O) (Budd et al. 1998;
Wright and Schwarcz 1998).
Chemical methods have been developed to cope with pervasive contamination of hard
tissues. Extraneous elements have been removed with washes of 1N acetic acid (Krueger
and Sullivan 1984; Lambert et al. 1990; Sillen 1986; Sullivan and Krueger 1981); ethyl
alcohol (Wright 1990); hydrochloric acid (HCl) (Gulson and Wilson 1994; Rose 1977,
1979); nitric acid (HNO3) (Gulson and Wilson 1994; Song and Stillman 1998); and

151
hydrogen peroxide solutions (H2O2) (Budd et al. 1998; Gulson and Wilson 1994; Song,
present analysis), among others.
These chemical procedures remove substantial diagenetic material from hard tissues,
but they also entail several consequences, such as: 1) the removal of biogenic Na, Mg and
K that are present on the surface of bone crystals (see Lambert et al. 1990); 2) deposition
of calcium salts on the surface of tooth sections, as the result of hydrochloric acid washes
(see Wright 1990); and 3) removal of significant biogenic bone (by 1N acetic acid)
(Krueger and Sullivan 1984; Sullivan and Krueger 1981).
This can be contrasted to the physical abrasion method, which does not affect
biogenic levels of aluminum, iron, magnesium, potassium, sodium or zinc. Overall, a
combination of physical abrasion and mild chemical washings (e.g., acetone, hydrogen
peroxide) is probably the best procedure for removing diagenetic additions (see Lambert
et al. 1990). Fortunately, none of the abovementioned methods were found to deplete
biogenic calcium or strontium, which are important elements with respect to diet.
Despite appeals for a reliable standardized preparation methodology (Boyde et al.
1978; Carlson 1990; Fergusson and Purchase 1987), there is as much variation as ever
regarding cleaning procedures. For histological studies, in particular, non-standardized
variations in preparation methods have notable consequences. Significantly, there can be
confusion with comparing histology and structural patterns of teeth, which can appear
different or similar depending on preparation technique (see Carlson 1990; Wright 1990).

152
6.3 Materials and Methods
The teeth were collected by the author from the Department of Anthropology at Trent
University, Peterborough, Ontario, Canada, where the bulk of Lamanai human remains
are currently housed (others remain in storage in Belize). Sample collection involved
sorting through all Postclassic and Historic period individuals at the university, which
amounts to approximately 106 Postclassic and 131 Historic period individuals. The
Lamanai human remains are in good condition for Lowland Maya burials, particularly the
dental remains.
Based on several criteria, one intact permanent mandibular or maxillary canine was
collected from each suitable individual, and the relative macroscopic “health” of each
tooth was assessed for the purposes of data completeness (see Appendix A). Permanent
canines are utilized because they are among the longest developing teeth in the human
dentition, with enamel forming shortly after birth to approximately five or six years of
age. As a result, they record life history for almost the entire period of breast milk
consumption and provide useful comparative data for tracing the subtle changes of a
prolonged dietary transition.
The teeth derive from adults of both sexes, as well as children of varying ages who
did not survive to adulthood, but whose canine crowns are complete. The sample is rich
with the dental chemistry histories of individuals with varying adaptive abilities, dietary
intakes and development. Note, however, that due to the requirement for: 1) a
completely developed canine crown (with cemento-enamel junction present), and 2)
minimal to moderate dental attrition, children who died younger than five to six years of
age and older adults with highly attritioned teeth are not included in this study.

153
Obviously, this limitation excludes individuals from two demographic extremes:
1) the least “successful” individuals who were not fit enough to survive early childhood
(younger than 5-6 years), and 2) the most advantaged individuals, or those older adults
with heavily worn teeth who probably experienced the greatest benefits to health and diet
in early life.
Yet, inclusion of complete canines from individuals who died in later childhood, i.e.,
older than 5 years of age (25% of total sample), means that some individuals who
experienced greater health and/or dietary stress are still considered in this analysis. (One
must remember that these disadvantaged children may have had atypical diets associated
with their poor health.) Overall, it is felt that the sample adequately represents the
general health and dietary conditions of the population in prehistoric and colonial times.
Age and sex information for each individual was derived from the Lamanai database
at Trent University [evaluations by Drs. H. Helmuth (Trent University) and C.D. White
(University of Western Ontario)], as well as the author’s general assessment of the dental
and skeletal remains during sample collection.
To maintain an adequate representation of dental remains for the Lamanai population,
canines were only collected from individuals with at least one other permanent canine, so
that such teeth are still available for future research. Collected canines are both
mandibular and maxillary. Only teeth with non-existent or minimal surface cracks
(overall good structural integrity) were chosen. Consequently, deeply filed or inlaid teeth
were not considered, as were many teeth that were fragmented due to poor post-
depositional and post-excavation preservation.

154
In total, only canines from 26 Postclassic and 34 Historic period individuals were
deemed appropriate for this analysis (see Table 6.1).
Table 6.1 Sample Breakdown
Time Period
Sub-Adults
Females
Males
Indeterminate
Adult
Total
Postclassic
6 (5 maxillary,
1 mandibular)
12 (5 maxillary,
7 mandibular)
7 (2 maxillary,
5 mandibular)
1 (1 mandibular)
26
Historical 9 (4 maxillary,
5 mandibular)
13 (7 maxillary,
6 mandibular)
12 (7 maxillary,
5 mandibular)
0 34
Once in Amherst, the teeth were further examined and prepared at the School of
Natural Science, Hampshire College. Macroscopic analyses of the teeth were undertaken
with the unaided eye, using an electronic sliding caliper for measurements of tooth
dimension and surface defect location. Observations include total crown height, degree
of dental attrition and calculus build-up, caries prevalence and the presence of any
developmental defects of enamel, namely hypocalcification discoloration and enamel
hypoplasia (see Appendix A). Enamel defects and the time of their formation are also
compared by temporal period, age (children vs. adults) and sex in Appendix A.
Macroscopic dental features were examined for the purpose of assessing dental health
patterns related to diet, health and hygiene. At this stage, surface calculus was carefully
removed to facilitate crown and defect measurements and to prepare the teeth for
cleaning (see below). Considering the potential for future chemical analyses of the
calculus, all such residue was retained and stored in labeled Ziploc bags.

155
6.4 Sample Preparation
In preparation for LA-ICP-MS, the teeth underwent a four stage cleaning process
according to the specifications of Drs. D. Amarasiriwardena and A.H. Goodman
(Hampshire College). This process is believed to maximize organic and inorganic
contaminant removal, while also preserving the innate chemical composition of the
material. The decision not to use certain strong acids (e.g., acetic, nitric, hypochloric) to
rinse or wash the teeth was based on the potential risks for ionic interference, i.e.,
leaching and substitutions.
Additionally, in order to minimize contamination, all glassware utilized for cleaning
was washed and rinsed in 18 M. cm (distilled and deionized) water, as were plastic
utensils. Other utilized materials were new, and in the case of kimwipes, a cellulose-
based, nonabrasive, lint- and static-free “delicate task wipe”, are deemed non-
contaminating for the purposes of ICP-MS analysis.
Initial cleaning involved manual brushing of the teeth with a new medium bristle
nylon toothbrush and 18 M. cm water. The teeth were subsequently soaked in 10 mL
disposable plastic micro beakers filled with 18 M. cm water for two days, so that any
adhering residue was loosened from the teeth. Two days later, the teeth were soaked in
new beakers filled with a 1% (w/w) papain solution for two days. Papain is a proteolytic
enzyme derived from papaya that breaks down protein and lipids. The beakers were
covered with plastic wrap to protect the teeth from air-borne contaminants.
Once removed from the papain with plastic tweezers, the teeth were thoroughly
rinsed several times with 18 M. cm water. They were then given a 30 second bath in a
3% (v/v) H2O2 solution for additional removal of organic material. Each dip was

156
carefully timed to avoid etching of the teeth and the hydrogen peroxide solution was
changed after every fifth tooth. Following each dip, the teeth were thoroughly rinsed
with 18 M. cm water. They were soaked a second time in new plastic beakers filled
with 18 M. cm water for one day. After removal from the water, the teeth were dried in
new beakers covered with kimwipes for one day.
After the four-stage, six-day, cleaning process, the teeth were prepared for
embedding. Using plastic tweezers, and holding the teeth by their roots, the teeth were
positioned in new plastic embedding containers so that they stood upright on their root,
with the occlusal surfaces facing up. The root tips were adhered to the base of the
container by heated glue. To facilitate subsequent cutting of the resin block, the teeth
were oriented with the labial-lingual plane parallel to the sides of the square container.
Once set, the containers were filled with Buehler Epoxide Resin (Lakebluff, IL, USA)
(five parts Epoxide Resin [bisphenol-A epoxy resin, N-butyl glycidyl ether] to one part
Epoxide Hardener [alkyl ether amine, diethylenetriamine, phenol, triethylenetetramine]),
a plastic resin that is considered non-contaminating for trace chemical analytical
purposes. The containers were then placed in a vacuum for approximately twenty
minutes to minimize air pockets in the resin as it set. They were left for two days to cure
and harden.
The last stage of tooth preparation involved cutting the resin blocks. Here, an Isomet
slow-speed saw was used with a new Buehler copper diamond-tipped disk blade. To
remove contaminants from manufacturing and packaging, the blade was rinsed with
acetone and 18 M. cm water prior to use. 18 M. cm water was also used throughout
the cutting process to lubricate the blade. The resin block was positioned so that the teeth

157
were cut at the midline along the labial-lingual plane. However, in some cases, when
there was uneven occlusal wear or the presence of a surface crack, the tooth halves were
divided unevenly, i.e., not along the midline. In such cases, as for all teeth in general, the
teeth were cut at the best position to ensure maximum enamel exposure in the sectioned
block.
At this time, one thin section of tooth and resin was also removed adjacent to the first
dividing cut and glued to a microscopic glass slide. According to the calibration on the
saw, this thin section is approximately 250 m thick. Thin sections of each tooth were
cut for the purposes of comparative histological analysis, which might aid in assessments
of nutritional status (see Marks and Rose [1985] for methodology).
Prior to laser ablation sampling, the cut surface of one block half underwent another
rinse with 3% (v/v) H2O2, followed by a rinse with 18 M. cm water, to eliminate
surface contaminants. To prevent spreading and admixture of dissolved material on the
cut tooth surface, the hydrogen peroxide was carefully poured so as to just cover the
tooth, and left to set for approximately one minute. The peroxide was then briskly
shaken off the tooth and the block rinsed in 18 M. cm water.
Importantly, the potentials for contamination in the use of manual brushing, papain
soaks, hydrogen peroxide dips and embedding in epoxy resin are minimal. ICP-MS
analysis of resin also indicates that there is minimal potential for contamination of the
embedded tooth. In general, the resin contains a thousand times less concentration of
relevant analytes present in dental material (A.H. Goodman, personal communication,
2000).

158
6.5 A Protocol for LA-ICP-MS Analyses of Human Dental Enamel
The operation of the LA-ICP-MS entails: 1) placing the embedded tooth section in
the laser ablation sampling chamber (“sample cell”), which is secured to a rotating
platform with double-sided tape; 2) orienting the tooth so that the ablation path is
horizontal at 180o; 3) determining the appropriate settings for the laser and mass
spectrometer in the computer program that operates the device; and finally 4) setting the
coordinates for the laser's path along the tooth surface. Analyses are carried out using the
time resolved analysis (TRA) mode, in which data is collected in discrete time segments
(ion counts/second) while the sample is continuously ablated at a constant rate (Lee et al.
1999; Montgomery et al. 1998).
As the laser can only sample material across a straight line path, several separate
transects angled to follow the tooth’s curve from occlusal tip to cervix, near the cemento-
enamel junction, were ablated (see Figure 6.2). Thus, ablated trenches vary in length
based on the morphology of each tooth, but most teeth have two to three connecting
ablation lines. Due to ionic incorporation of surface elements during life and/or post-
mortem, particular care was taken to ablate samples from inner enamel only.
Figure 6.2 Position of laser ablation trenches along secondary canine crown
(with approximate ages of enamel development indicated)

159
For this analysis, laser ablation sampling was the “line” ablation type, which samples
enamel across a continuous line at a constant rate (20 m/sec). All canines were ablated
across the entire length of labial enamel to ensure that enamel was collected from all
periods of crown development, from approximately six months to five or six years of age.
Labial enamel was ablated because of two factors: 1) enamel is thicker in this region
and thus, easier to sample and observe histologically, and 2) surface absorption of ions
(contamination) is less likely. As noted previously, despite its inorganic composition,
surface enamel is prone to ionic exchange with several elements in the oral environment,
most notably lead and fluorine in contemporary times. Surface absorption of contaminants
is most pronounced on the lingual surface of teeth and surrounding the neck due to their
increased contact with saliva and plaque (Carroll et al. 1972; Purchase and Fergusson
1986). Additionally, with respect to lead, higher levels characterize the amelodentinal
region and root dentine compared to coronal dentine (Malik and Fremlin 1974).
In archaeological teeth affected by diagenesis, strontium values in dentine are also
inflated relative to enamel, which tends to be largely homogeneous in strontium
concentration (Montgomery et al. 1998). In total, owing to the thicker nature of labial
enamel, sampling within an inner region considerably distant from (less) potentially
contaminated surface enamel is more feasible and preferable.
Table 6.2 outlines the LA-ICP-MS operating parameters utilized in this investigation.
The instrumentation includes a Perkin Elmer Sciex, Elan 6000 ICP-MS (Perkin Elmer
Instruments, Shelton, CT, USA) coupled to a CETAC LSX-100 Laser Ablation System
(frequency quadrupled Nd:YAG [neodymium-yttrium aluminum garnet] laser) (CETAC
Technologies, Omaha, NE), operating at 266nm in (UV) wavelength. For adequate

160
sampling of enamel material, an energy level averaging around 0.9 -1.0 mJ was required
for the laser (S. Keydel, personal communication, 2000).
Table 6.2 LA-ICP-MS Operating Parameters
Laser Ablation and ICP-MS Operating Parameters
Laser Ablation Operating Parameters:
Laser Type Nd: YAG
Laser Mode Frequency quadrupled 266 nm UV,
Q –switched mode
Repetition Rate/Hz 10
Laser Energy/mJ 0.6-1.1
Sampling Scheme Raster
Scanning speed/μm.s-1
20
ICP-MS Operating Parameters:
Forward power/ kW 1
Ar gas flow rates/L. min-1
Coolant 15
Auxiliary 1.2
Nebulizer gas 0.9
Measurement conditions:
Dwell time/ms 25
Resolution high
Sweeps/reading 1
Readings/replicate 5600
Number of replicates 1
Isotopes measured (m/z) 137
Ba, 138
Ba, 43
Ca, 44
Ca, 63
Cu, 65
Cu, 57
Fe, 24
Mg, 25
Mg,
55Mn,
206Pb,
207Pb,
208Pb,
78Se,
82Se,
86Sr,
88Sr,
64Zn,
66Zn
Internal standard (m/z) 43
Ca

161
Considering the operating parameters, the energy level chosen for the laser ablation
device was 13, with a pulse rate of 10 and an ablation speed of 20 m/second. These
settings produced an ablated trench measuring approximately 50 m wide and deep.
Only microgram quantities of enamel were sampled from each tooth.
Throughout, the ablation process (and element behavior) was viewed in real time
through a 20x video monitoring system (see Figure 6.3). This attached monitor and
microscopic camera enabled identification of different dental tissues and establishment of
the ablation path prior to analysis. Observations were made during each ablation process
and were recorded on a standard form prepared by the author (see Figure 6.4). Recorded
details include operating parameters, fluctuating energy levels, ICP reading numbers for
each ablation, relative location, ablation lengths (measured by the LA device and
computer software), and reading locations of distinct enamel features (e.g., defects,
cracks, proximity to dentine and cemento-enamel junction). A sketch of each tooth
section and ablation locations accompanied each data sheet on the reverse side.
Figure 6.3 Schematic of instrumentation used in LA-ICP-MS
(from Denoyer et al. 1991: Fig. 1)

162
Figure 6.4 Example of LA-ICP-MS recording sheet used in this analysis

163
Once ablated, the vaporized enamel was swept from the sample chamber to the
plasma source with a flow of argon gas. In the plasma, the sample was atomized, ionized
and then transported to the mass spectrometer for quantification. As befits the
capabilities and advantages of ICP-MS technology, multiple elements were quantified.
In addition to strontium (86
Sr, 88
Sr), these elements include barium (137
Ba, 138
Ba), calcium
(43
Ca, 44
Ca), copper (63
Cu, 65
Cu), iron (57
Fe), lead (206
Pb, 207
Pb, 208
Pb), magnesium (24
Mg,
25Mg), manganese (
55Mn), selenium (
78Se,
82Se) and zinc (
64Zn,
66Zn). Quantification of
these isotopes were deemed appropriate for the purposes of dietary and nutritional
reconstruction, reduced polyatomic interference (D. Amarasiriwardena, S. Keydel, P.
Outridge, personal communications, 2000, 2001), as well as future studies on pollution
(for instance, lead may be useful in tracing possible environmental exposure during the
production of lead-glazed ceramics [e.g., Tunstall and Amarasiriwardena 2002]).
As stated earlier, the LA-ICP-MS analytical approach is ideally suited to dental
analyses as it produces a “map” of chemical signatures specific to tooth location and
developmental age. Thus, interpretation of the chemical blueprint of sampled enamel is
the final stage of LA-ICP-MS analysis. This involves statistical comparison of strontium
levels relative to calcium over time (tooth location). Calcium (e.g., 43
Ca) is an
appropriate normalizing element since it is present in the enamel matrix at relatively
stable, high, levels.
Upon completion of this investigation, dental remains will be returned to Trent
University either in embedded form, or disengaged from the resin mold and thin section.
In this case, analyzed teeth can be returned to their original state by heat removal of the
plastic resin and gluing of the thin section within the cut tooth halves (it is suggested,

164
however, that the thin section is maintained for the purposes of a histological database
and future analyses). Other than a minuscule trench of ablated enamel (approximately 50
m wide) in the center of the tooth, glued teeth appear complete and remain available for
future research.
6.6 LA-ICP-MS Observations
In light of the sparse application of laser ablation sampling to human tissues, several
features of LA-ICP-MS are worth mentioning. First, in an analysis that involves raster
(continuous) laser sampling of enamel along entire crown lengths, substantial numerical
data is generated. For instance, with a canine crown measuring 11 mm, the approximate
average unworn crown height of Lamanai canines (Appendix A: Table A.1), a continuous
laser ablation produces around 800 readings per element (see Appendix B Tables).
Quantification of multiple elements and several isotopes of each element results in large
data files for every ablation.
During the process of laser ablation, ICP-MS quantification of elements is recorded
for both the periods of sample ablation as well as during non-sampling, when argon gas
continues to flow. Graphically, each phase of operating the LA-ICP-MS can be
distinguished by distinct Sr/Ca patterns (see Figure 6.5).
The periods of closed chamber non-sampling when argon gas continues to flow
through the ICP-MS torch are clearly recognizable by the significantly higher Sr/Ca
ratios (Figure 6.5: phase “A”). This ratio is the result of relatively low Ca readings,
rather than elevated Sr values. (With enamel ablation sampling, significantly higher Ca
values dramatically reduce the Sr/Ca ratio.) Although the argon gas used in ICP-MS is of

165
a high purity, it is clear from Figure 6.5 that the gas blanks are not completely uniform,
with some variation evident. Slight variations in the very low total Ca intensity relative
to Sr intensity in argon gas account for the fluctuations.
Figure 6.5 Graph illustrating distinct patterns in Sr/Ca during LA-ICP-MS
Periods of actual enamel ablation are distinguished by a substantial drop in Sr/Ca due
to significantly higher Ca values (phase “B”). As a result, the readings during initial
enamel sampling are often significantly higher than Sr/Ca values of subsequent enamel,
since they are actually residual “argon blank” readings. Care has been taken to eliminate
unusually high initial readings for the data analysis and presentation, but in some cases,
the initial periods of enamel development are still marked by high Sr/Ca values that
diminish shortly thereafter (see Chapter Seven).
A
Fluctuations in Sr/Ca during the entire process of LA-ICP-MS
0.00
0.50
1.00
1.50
2.00
2.50
0 200 400 600 800 1000 1200
Reading #
Sr/
Ca
ra
tio
------------- Dental Enamel ---------------
Argon Blank Argon Blank
Sample Chamber Open
A
B C

166
A third phase of activity during LA-ICP-MS can also be identified in Figure 6.5
(phase “C”). When the sample chamber is open to re-position tooth sections for
additional ablations, and argon gas continues to flow, Sr/Ca ratios appear slightly
enhanced compared to readings when enamel is being ablated (“B”). However, they are
still significantly reduced compared to periods of non-sampling when the chamber is
closed (“A”).
Factors that affect the sampling of material to the ICP torch (i.e., intensity of the
element signal) and quantification of elements include sample hardness, sample position
in the laser chamber, whether or not it is level, as well as instrumental drift in the ICP-
MS over several hours or days (see Gratuze et al. 2001; Neff 2003). Instrumental factors,
such as decreased signal (sensitivity) and elemental fractionation over time, are attributed
to thermal processes at the ablation site, as well as changes in the aspect ratio of the
ablation crater (Gunther and Hattendorf 2001; Jackson 2001).
To diminish such effects, Gratuze and colleagues (2001: 646) suggest collecting data
in three or more different runs, but this was not possible in this study due to the thinness
of enamel in sections of labial crown near the CEJ. Neff (2003: 24) also suggests that
raster, or continuous lines ablations, rather than single spots, and ablations of relatively
wide crater width (100 μm) can reduce the problem of decreased signal and fractionation.
This study adopts the raster mode of sampling, with crater widths restricted to
approximately 50 μm (microns), because a wider ablation risks sampling of dentine in
areas of thin labial enamel (i.e., CEJ third). A crater width of 50 μm is appropriate for
this paleodietary analysis, however, and ICP-MS results of Lamanai canines indicate that
signal strength was not compromised during ablations.

167
6.7 Laser Ablation Details for Lamanai Canines
The following photograph illustrates an ablated Lamanai canine section with multiple
ablations indicated. A superimposed micrometer scale provides measurements of each
ablation.
Figure 6.6 Photograph of ablated tooth section and micrometer scale
(extrapolated crown tip and separate ablations indicated in black)
Periods of crown development covered by each ablation are based on the relative
lengths of laser ablations and enamel not ablated near the cusp tip and cemento-enamel
junction, compared to dental standards outlined in Appendix A. As illustrated in Figure
6.6, measurements are based on photomicrographs of ablated teeth and a micrometer
glass scale calibrated to 20 μm (0.02 mm), which were obtained with a NIKON 9000

168
digital camera and LEICA binocular microscope at 16x magnification. These figures are
generally greater (some are lower) than measurements obtained during laser ablation with
CETAC Laser Ablation operating software (CETAC, Omaha, NB), but they are not
significantly different, i.e., less than 5 % higher on average (3.64%). Similarly,
Haverkort (2001) has found slight discrepancies between measurements of laser ablation
tracks made from SEM images and software-generated distances based on scan duration
and scan speed, which is attributed to slight distortions in the SEM images.
Photographic evidence was used since measurements for unablated enamel near the
cemento-enamel junction and at the cusp tip could not be measured by the laser ablation
device and software. In particular, enamel missing from the cusp tip (the initial period of
crown development) had to be estimated in worn teeth by extrapolating a cusp curve on
the actual photograph (see Figure 6.6).
Based on photographic measurements, Tables B.1-4 in Appendix B outline the
ablation details for all Lamanai teeth analyzed in this study. Tables are grouped into
maxillary and mandibular teeth for consistency of enamel developmental period and the
“ages” (in years) represented in each ablation. Mandibular and maxillary teeth are
generally equally distributed in both samples and between age and sex cohorts:
Postclassic maxillary canines = 5 subadults, 5 females, 2 males; Postclassic mandibular
canines = 1 subadult, 7 females, 5 males, 1 adult unknown sex; Historical maxillary
canines = 4 subadults, 7 females, 7 males; Historical mandibular canines = 5 subadults, 6
females, 5 males.

169
6.8 Data Analysis
Initial organization of the data involved conversion of the raw data (intensity vs.
reading number) from PE Elan Software to Microsoft Excel files. Background gas 86
Sr
values (averaged for each ablation) were then deducted from the original 86
Sr intensity
readings for each tooth. High purity argon gas acts as the medium in which ablated
enamel is delivered to the ICP-MS for semi-quantification and it contains minor amounts
of trace elements. Argon gas blank values were recorded when the laser was not firing,
i.e., in between ablation runs, and range from approximately 2-20% of 86
Sr values
detected in enamel during laser sampling.
Next, as there is no suitable matrix-matched standard, 86
Sr readings were normalized
against 43
Ca values (minus argon gas 43
Ca) at each count of the ablation raster, producing
a Sr/Ca ratio, which is the basic unit of analysis in this investigation. The majority of
86Sr/
43Ca values for permanent canine enamel lie between 0.02-0.14 in this study (see
Chapter Seven). [While not available to this study, LA-ICP-MS data processing can be
simplified by implementation of the LADA (Laser Ablation Data Analysis) program
created by B. Hazes (see Haverkort 2001). Among other functions, this Fortran program
subtracts gas blank values from sample measurement values, calculates Ca ratios and
provides basic statistics of select data sets (e.g., correlations between elements,
logarithms).]
Unfortunately, as this study examined specific isotopes of strontium and calcium, the
Sr/Ca values calculated in this analysis have no directly comparable data in the chemical
and paleodietary literature, which have either examined total Sr/Ca or 90
Sr.

170
One is reminded of Sillen and Kavanagh’s (1982: 69) statement:
This raises a difficulty with most of the available literature and reviews on
strontium. Since researchers were primarily concerned about fallout, they tended to
emphasize the distribution of 90
Sr and other radioactive isotopes1. While this
information is, in many cases, extremely useful, it should be kept in mind that these
studies were not designed with the intent of developing a method for the
investigation of paleodiets. Indeed, apart from any applications, a great deal of
basic research on the use of Sr/Ca ratios for paleodietary research remains to be
conducted.
Even now, much of what is known about strontium’s distribution, behavior and
incorporation in hard tissues is based on early studies of radioactive fallout in the 1950’s
and 1960’s. Among many diverse studies, important data often referenced in hard tissue
strontium research include Beninson and colleagues (1964); Bryant and Loutit (1964);
Comar and co-authors (Comar 1963; Comar et al. 1957), Lengemann (1960, 1963;
Lengemann et al. 1957); Lough and colleagues (1963); Rosenthal (1981); Spencer and
colleagues (1961); Thompson (1963); Turekian and Kulp (1956); Steadman and co-
authors (1958); and Wasserman (1963; Wasserman and Comar 1960; Wasserman et al.
1958, 1977).
The majority of strontium reference data concerns 90
Sr, but all radioactive and stable
isotopes of Sr have the same physical properties, including the accumulation in human
hard tissues as a substitute for calcium (see Chapter Four). The four stable isotopes of
strontium occur in the following natural abundance: 88
Sr (82.56%), 86
Sr (9.86%), 87
Sr
(7.02%) and 85
Sr (0.56%).
1 Radioactive 90Sr was discovered in the 1940’s during nuclear experiments for the development of the atomic bomb, which resulted
in the atmospheric dispersal of large amounts of 90Sr worldwide. It is one of fourteen radioactive isotopes of strontium and is a by-product of the nuclear fission of uranium and plutonium (Faure and Powell 1972). With a half-life of 29 years and a widespread
distribution due to weapons experiments, nuclear reactor waste, and industrial accidents (Chernobyl), it is an important radioactive
isotope in the environment.

171
In this study, 86
Sr is the focus of investigation, with 43
Ca used as an internal standard.
Mass spectroscopic interferences and mass abundances are important factors in
considering which isotopes to analyze. Here, 43
Ca is used because it is the least abundant
Ca isotope (0.14%), is not subject to polyatomic interferences and is also free from
isobaric interferences (D. Amarasiriwardena, personal communication, 2002). The
reduced abundance of this isotope provides a manageable, yet adequate, signal for
calcium.
Relative signal strength and (reduced) risk of interferences are also important factors
in choosing 86
Sr, though one notes that there can be an isobaric interference of this
isotope with 86
Kr if it is present as a trace contaminant in argon gas (D.
Amarasiriwardena, personal communication, 2004). Still, 86
Sr is considered a suitable
isotope for strontium analysis in teeth.
Permanent canine enamel 86
Sr/43
Ca ratios for 0.25 year intervals range between
0.014-0.179, with the majority lying between 0.02-0.14. Compared to hard tissue total
Sr/Ca data of other studies (Mays 2003; Sillen and Smith 1984), these values are higher
by orders of magnitude due to the relatively higher abundance of 86
Sr against 43
Ca,
compared to total strontium versus calcium (see Chapter Seven).
However, total Sr/Ca ratios can be derived from the 86
Sr/43
Ca values presented in this
analysis since the abundance of all naturally occurring isotopes of strontium and calcium
are known. Calcium has six naturally occurring isotopes, with the following natural
abundance in percentages: 40
Ca (96.941%), 42
Ca (0.647%), 43
Ca (0.135%), 44
Ca
(2.086%), 46
Ca (0.004%) and 48
Ca (0.187%). Taking into account the differential

172
abundance of 43
Ca (0.135% of total Ca) and 86
Sr (9.86% of total Sr) isotopes, one can
extrapolate total Sr/Ca values that are comparable to reference data (see Chapter Seven).
This analysis has focused on 86
Sr and 43
Ca isotopes for dietary reconstruction, but
other human hard tissue strontium studies have utilized different naturally occurring
isotopes, albeit for different reasons (dietary vs. environmental exposure). These include
88Sr (Goodman et al. 2003; Haverkort 2001) and
90Sr (Bryant and Loutit 1961; Rivera
and Harley 1965; Thompson 1963). For calcium, 43
Ca has also been used as an internal
standard by Imai (1992), Goodman et al. (2003) and Kang and colleagues (2004), while
Haverkort (2001) and Montgomery and others (1998) utilized 46
Ca in LA-ICP-MS
analyses of human teeth.
Comparing Sr/Ca ratios of different Sr and Ca isotopes reflects the dramatic extent to
which ratios can differ in absolute value. Based on a set of ablations from a single day
(5600 readings), including six full ablations of canine enamel, different ratio
combinations using 86
Sr, 88
Sr, 43
Ca and 44
Ca were compared. In ablated enamel, with
44Ca as the internal standard,
86Sr/
44Ca was, on average, approximately 14.5 times lower
than the standard ratio used in this study (86
Sr/43
Ca), reflecting the higher abundance of
44Ca. With
88Sr instead of
86Sr, normalized with
43Ca, the opposite is true, with the
88Sr/
43Ca ratio approximately 8.7 times, on average, higher than the standard ratio (due to
increased 88
Sr abundance). Using different isotopes for both elements (88
Sr and 44
Ca)
produced a more analogous ratio to 86
Sr/43
Ca, but even in this case, 88
Sr/44
Ca was
approximately 1.7 times lower than the ratio used in this analysis.
Figure 6.7 illustrates the four combinations using 86
Sr, 88
Sr, 43
Ca and 44
Ca from one
enamel ablation run. Each set of Sr/Ca data have isotope-specific argon blank values

173
deducted. The values for 88
Sr/43
Ca are noticeably higher than the three other isotopic
ratios due to greater abundance of 88
Sr. In contrast, greater abundance of 44
Ca reduces
the ratio of 86
Sr/44
Ca dramatically.
Figure 6.7 Graphical comparison of different Sr/Ca isotope ratios in enamel
Additionally, Figure 6.8 is an adjusted comparison of the four combinations with
86Sr/
44Ca values multiplied by 5 and
88Sr/
43Ca divided by 10 to produce more comparable
distributions. Notably, all four patterns mimic each other closely, particularly in areas of
significant peaks and valleys. This expected consistency of strontium and calcium ratios
demonstrates the capacity of laser sampling and ICP-MS (semi-) quantification of enamel
composition and confirms the utility of any Sr/Ca isotope ratio in reflecting important
relative information regarding element behavior over development time.
Logarithmic Comparison of Various Sr/Ca Isotope Ratios
0.00
0.01
0.10
1.00
10.00
0 100 200 300 400 500 600 700 800
Reading #
Lo
g S
r/C
a
86Sr/44Ca
88Sr/44Ca
86Sr/43Ca
88Sr/43Ca

174
As expected, other alkaline elements are also comparable, namely 137
Ba, 138
Ba, 24
Mg
and 25
Mg. Between the two elements, Mg parallels Sr and Ca better than Ba, although
25Mg sometimes appears stable when
88Sr and
43Ca increase. Barium generally follows
the other alkaline elements, but also can differ at times.
Figure 6.8 Adjusted comparison of different Sr/Ca isotope ratios
Since results of LA-ICP-MS are in the form of element values per reading number or
counts, in order to assess Sr/Ca change over developmental time, the reading numbers
were converted to relative ages of crown formation. The exact period of enamel
development sampled for each canine is based on the relative lengths of laser ablations
and unablated enamel compared to established models of crown formation time.
Comparison of Various Sr/Ca Isotope Ratios
0
0.02
0.04
0.06
0.08
0.1
0.12
0.14
0 100 200 300 400 500 600 700 800
Reading #
Sr/
Ca R
ati
o
86Sr/44Ca x 5
88Sr/44Ca
86Sr/43Ca
88Sr/43Ca /10

175
In this study, permanent maxillary canines are attributed to begin enamel
development at around birth and finish at 5.0 years, while permanent mandibular canines
are delayed by 0.5 years (see Appendix A.4). As outlined in Appendix A, approximately
one year of initial crown development is buried in inner enamel of permanent canines
(Reid and Dean 2000), and thus not recognized macroscopically. Determination of the
timing of surface enamel defects accounts for this year of cuspal enamel (Appendix A).
For LA-ICP-MS, however, which samples inner enamel, some buried cuspal enamel
is analyzed. In this case, it is appropriate to assume a shorter period of “hidden” enamel
that is not examined. Only 0.5 years is considered missing from the initial period of
crown development so that the periods of enamel development sampled by laser ablation
in this study are 0.5-5.0 years in maxillary canines and 1.0-5.5 years in mandibular
canines.
However, not all teeth have enamel dating to the first age interval (0.5-0.75 years)
that could be sampled because of cuspal wear and the need to sample non-surface
(potentially contaminated) enamel. No enamel is available from the latest period of
development either, due to the thinness of enamel near the CEJ, but it is represented by
averaged values for the last interval (5.25-5.5 years).
Due to the large number of element readings spanning the period from 0.5-5.5 years,
Sr/Ca values for ablated enamel were reduced to averages for 0.25 year (3 month)
intervals. Three month intervals could be differentiated since reading numbers can easily
be converted to age in years based on the total period of enamel development ablated in
each tooth (see Section 6.7). The 0.25 year interval is the main unit of analysis in this
study. Scatter plot distributions are presented in Chapter Seven.

176
Statistical analyses were undertaken with SPSS (version 11.0) software. Inter- and
intra-comparisons of Sr/Ca values are provided by one-way ANOVA’s with level set at
p>0.05 (two-tailed) (see Appendix C). Comparisons are made at the level of overall
Sr/Ca change; temporal period; age (children vs. adults) and adult sex. Importantly,
temporal comparisons indicate that the total Postclassic and Historical samples differ
significantly at many age intervals. Throughout the entire period of permanent canine
enamel formation, Postclassic mean Sr/Ca is elevated compared to Historical values (see
Section 7.2).
Some distinctions are also noted between Postclassic and Historical adults (Section
7.3). The nature of the difference and the implications for dietary supplementation
timing are illustrated by adjusted scatter plots in Section 7.3. Adjusted graphs are also
presented for comparisons that are not statistically significant, but which might have
implications for delays in attaining Sr/Ca levels in certain cohorts.

177
CHAPTER 7
RESULTS OF ANALYSIS
7.1 Major Findings
The primary focus of this investigation is to assess whether Sr/Ca composition in
sequentially calcified enamel can be observed to reflect changes in infant diet from
exclusive breastfeeding to complete food dependence (weaning). This dietary transition
represents one of the most significant developmental stages in human life.
Ancient Maya teeth are used in this LA-ICP-MS analysis to specifically examine the
nature of infant feeding through the transition from indigenous prehistory to Spanish
colonial history. This historic period offers a dynamic cultural context to the analysis
that can be reflected in the human remains. Age, sex and temporal cohorts are compared
to provide insight into varying dietary practices and differences in infant care.
The results suggest that enamel Sr/Ca does record the process of infant dietary
supplementation, from predominant breast milk intake to predominant solid food
reliance. This is reflected in rising Sr/Ca values from 0.5 to 5.5 years of age, as solid
food supplementation is introduced and augmented over time. The mean Sr/Ca pattern
over time for the entire Lamanai sample is outlined in Section 7.2, which generally
follows the proposed model in Section 5.6. From the data, the weaning process entails an
approximately two-fold increase in enamel Sr/Ca (see Table 7.15).
It should be stressed, however, that hard tissue Sr/Ca can only reliably be used to
identify the introduction and consumption of solid foods among infants. Currently, only

178
nitrogen isotopes can be used to identify the cessation of breast milk intake (weaning) in
hard tissues (see Chapter Five). One can infer that peak Sr/Ca represents the age of
weaning, i.e., maximum solid food intake, but it is complicated by two opposing realities:
1) weaning may have occurred earlier than the age of peak Sr/Ca, which may only reflect
the (later) age of efficient Sr discrimination by mature digestive systems; 2) increased
discrimination of Sr by the gut and reduced Sr/Ca values thereafter does not rule out the
continued intake of some breast milk after attaining peak Sr/Ca. Nevertheless, in this
case, when nursing is extended (i.e., beyond two years), breast milk consumption likely
reflects a minimal (insignificant) dietary contribution (see Section 8.2).
As discussed in Section 6.8, utilization of LA-ICP-MS in this analysis results in the
ratioing of specific isotopes of strontium and calcium, which does not directly compare
with total Sr/Ca data. Nevertheless, comparison of various Sr/Ca isotope patterns in
Chapter Six indicates that 86
Sr/43
Ca provides highly reliable relative information about
the nature of strontium change during enamel development. Total Sr/Ca ratios can be
derived from the specific isotope ratios and are presented in Section 7.2, but the primary
unit of analysis in this investigation is the 86
Sr/43
Ca ratio, which is considerably larger
than total Sr/Ca values. Here, permanent canine enamel 86
Sr/43
Ca ratios vary from 0.014-
0.179, with the majority between 0.02-0.14, while total Sr/Ca values range from 0.199-
2.454 x 10-3
(0.000199-0.002454).
Temporally, the Postclassic and Historical samples differ significantly in average
Sr/Ca values of permanent canine enamel, with Historical individuals displaying lower
Sr/Ca ratios at all age intervals (Figure 7.2). It is suggested that the Historical pattern
reflects a process of food supplementation and weaning that is delayed compared to

179
Postclassic Lamanai Maya. The mean age for peak Sr/Ca is 5 years among Postclassic
individuals and no earlier than 5.25 years, and possibly as late as 6.25 years, in Historical
individuals (see discussion of Figure 7.7).
Section 7.3 describes averaged Sr/Ca data for age, sex and temporal cohorts.
Comparisons are made between time periods for adult males, females and children and
hypotheses provided for differences in mean Sr/Ca values and overall pattern. Most
differences are not statistically significant, but Historical adults consistently exhibit lower
mean Sr/Ca than Postclassic adults at the same age interval. Lower mean Sr/Ca values
are interpreted to reflect a possible delay in acquiring a comparable level of food
supplementation. Shifting Sr/Ca distributions between cohorts demonstrates the potential
consequences of distinctions that are not necessarily statistically significant (see below).
Specific Sr/Ca details for all individuals are tabulated and illustrated graphically in
Section 7.4 according to age, sex and temporal context. Individual graphs demonstrate
the extent of variation in Sr/Ca patterns over development time (Figures 7.15-7.20).
While most individuals display an overall increase in Sr/Ca over time, the total sample
pattern between 0.5 and 5.5 years of age includes many subtle fluctuations, which is
thought to reflect intermittent changes in relative breast milk and solid food intake.
Four patterns can be discerned from the Sr/Ca data: 1) gradual increase in food
supplementation (Sr/Ca) until complete weaning; 2) delay in significant food
supplementation followed by a rapid transition to weaning; 3) fluctuations in relative
breast milk and solid food intake over time; 4) generally stable enamel Sr/Ca over time
(see Section 7.5).

180
Within each pattern, enamel Sr/Ca suggests that there is tremendous variation
between and within individuals. The transition to complete food dependence can be
recognized in rising Sr/Ca values over time, but many children experience alternating
periods of changing relative breast milk and food intake. These fluctuations likely reflect
the interaction of changing socioeconomic conditions (e.g., access to food resources),
maternal health and infant illnesses.
7.2 The Lamanai Sr/Ca Pattern
As postulated in this study, Sr/Ca values increase significantly over time during the
development of secondary canine enamel (0.5-5.5 years of age). Figure 7.1 illustrates
standard error bars for mean 86
Sr/43
Ca values of the entire sample at each age interval,
which are summarized in Table 7.1. Two standard deviations on either side of the mean
are included in each bar.
The greatest variation appears at both extremes of the distribution, where Sr/Ca
values often exhibit significant fluctuations. This is likely due to instrumentation and
sampling factors: for example, the start of enamel sampling after a period of argon gas
blank readings is often marked by a considerable peak in Sr/Ca, and the end of enamel
ablation sampling is also characterized by erratic element readings (see Chapter Six).
Notably, the significant increase visible at 0.75 years of age (9 months) suggests that
most Lamanai infants began food supplementation at around this age. This is followed
by gradual food supplementation, which seems to accelerate after the age of two years
(see further below). Peak Sr/Ca values at 5-5.25 years of age suggest that Lamanai

181
children may not have been completely dependent on solid foods until this late age.
Alternatively, it may reflect the age when digestive systems fully mature and efficiently
discriminate against strontium, which thereafter diminishes in hard tissue concentration.
Figure 7.1 Standard Error graph of mean 86
Sr/43
Ca for all Lamanai individuals
Age in Years
5.5
5 4.5 4 3.5 3 2.5 2 1.5 1 0.5
Mean S
r/C
a +
- 2 S
E
.10
.08
.06
.04
.02
0.00
.12

182
Below, Table 7.1 summarizes the mean 86
Sr/43
Ca ratios for 0.25 year intervals
between 0.5 and 5.5 years of age for Postclassic and Historical cohorts, as well as the
total sample. It is clear that mean Sr/Ca values increase with developmental age.
Graphically, the Postclassic and Historical data from this table are compared in Figure
7.2, which reflects distinctions in mean Sr/Ca at every age interval (see below for
elaboration). Additionally, Figure 7.2 is a standard error graph of the mean Sr/Ca data
for the two cohorts.
Table 7.1 Summary of mean 86
Sr/43
Ca for 0.25 year intervals
Age Interval Postclassic Historical Total Sample
0.50-0.74 0.05914 0.03653 0.04557
0.75-0.99 0.05843 0.04384 0.04968
1.00-1.24 0.05665 0.04661 0.05083
1.25-1.49 0.05674 0.04898 0.05227
1.50-1.74 0.05794 0.04999 0.05344
1.75-1.99 0.06021 0.05145 0.05525
2.00-2.24 0.06296 0.05248 0.05702
2.25-2.49 0.06472 0.05415 0.05873
2.50-2.74 0.06686 0.05579 0.06059
2.75-2.99 0.07034 0.05749 0.06306
3.00-3.24 0.07396 0.05992 0.06600
3.25-3.49 0.07628 0.06143 0.06787
3.50-3.74 0.07782 0.06357 0.06974
3.75-3.99 0.08006 0.06582 0.07199
4.00-4.24 0.08335 0.06810 0.07456
4.25-4.49 0.08542 0.06768 0.07520
4.50-4.74 0.08770 0.06954 0.07724
4.75-4.99 0.08860 0.07025 0.07828
5.00-5.24 0.09563 0.07217 0.08312
5.25-5.49 0.08718 0.07542 0.08154

183
Figure 7.2 Standard Error graph of mean Sr/Ca for Postclassic and
Historical individuals
Analyses of Variance (ANOVA) indicate significant differences between
Analyses of Variance (ANOVA) indicate significant differences between Postclassic
and Historical cohorts only between 2.25 to 4.75 years of age (see Table 7.2). During
this period, the rate of Sr/Ca increase among Postclassic individuals is slightly
accelerated compared to Historical period Maya at Lamanai. It appears that Postclassic
children after 2.25 years of age increase their intake of solid foods relative to breast milk,
while Historical children consume relatively less solid foods, i.e., maintain the same
proportions of food and breast milk intake as earlier.
Age in Years
5.2554.75
4.54.254
3.753.5
3.2532.75
2.52.252
1.751.5
1.2510.75
0.5
Mea
n +
- 2
SE
.12
.10
.08
.06
.04
.02
0.00
Postclassic
Historical
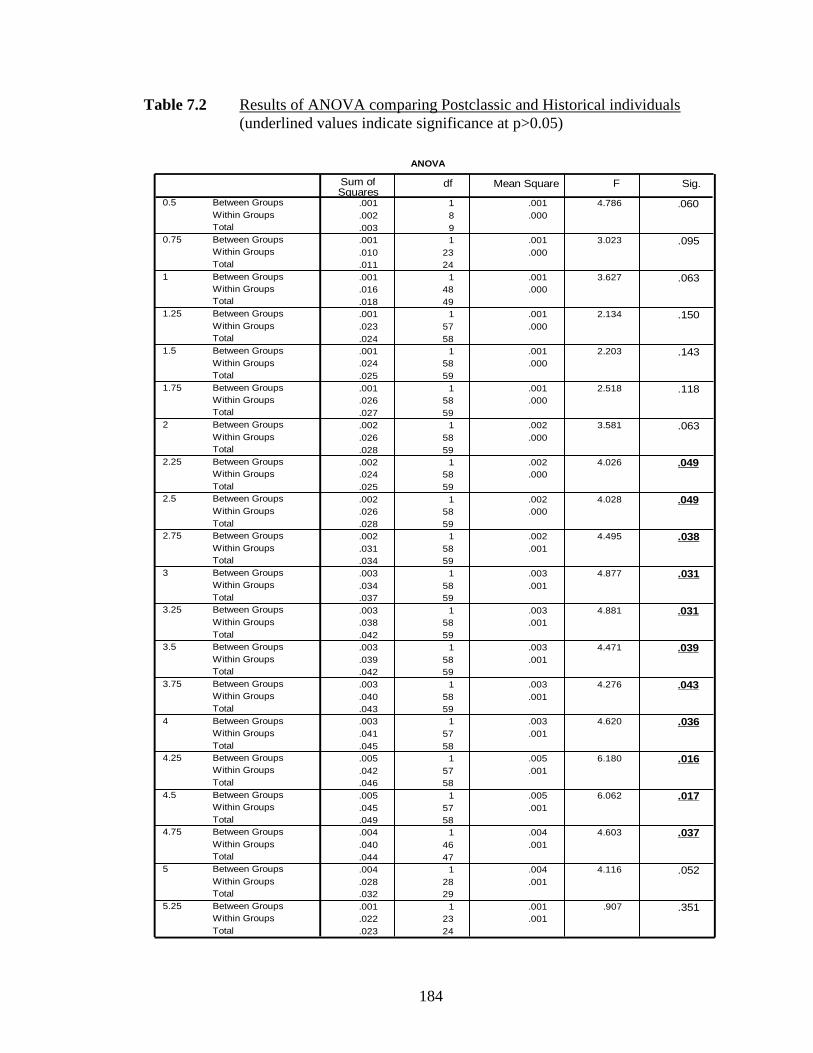
184
Table 7.2 Results of ANOVA comparing Postclassic and Historical individuals
(underlined values indicate significance at p>0.05)
ANOVA
.001 1 .001 4.786 .060
.002 8 .000
.003 9
.001 1 .001 3.023 .095
.010 23 .000
.011 24
.001 1 .001 3.627 .063
.016 48 .000
.018 49
.001 1 .001 2.134 .150
.023 57 .000
.024 58
.001 1 .001 2.203 .143
.024 58 .000
.025 59
.001 1 .001 2.518 .118
.026 58 .000
.027 59
.002 1 .002 3.581 .063
.026 58 .000
.028 59
.002 1 .002 4.026 .049
.024 58 .000
.025 59
.002 1 .002 4.028 .049
.026 58 .000
.028 59
.002 1 .002 4.495 .038
.031 58 .001
.034 59
.003 1 .003 4.877 .031
.034 58 .001
.037 59
.003 1 .003 4.881 .031
.038 58 .001
.042 59
.003 1 .003 4.471 .039
.039 58 .001
.042 59
.003 1 .003 4.276 .043
.040 58 .001
.043 59
.003 1 .003 4.620 .036
.041 57 .001
.045 58
.005 1 .005 6.180 .016
.042 57 .001
.046 58
.005 1 .005 6.062 .017
.045 57 .001
.049 58
.004 1 .004 4.603 .037
.040 46 .001
.044 47
.004 1 .004 4.116 .052
.028 28 .001
.032 29
.001 1 .001 .907 .351
.022 23 .001
.023 24
Between Groups Within Groups Total Between Groups Within Groups Total Between Groups Within Groups Total Between Groups Within Groups Total Between Groups Within Groups Total Between Groups Within Groups Total Between Groups Within Groups Total Between Groups Within Groups Total Between Groups Within Groups Total Between Groups Within Groups Total Between Groups Within Groups Total Between Groups Within Groups Total Between Groups Within Groups Total Between Groups Within Groups Total Between Groups Within Groups Total Between Groups Within Groups Total Between Groups Within Groups Total Between Groups Within Groups Total Between Groups Within Groups Total Between Groups Within Groups Total
0.5
0.75
1
1.25
1.5
1.75
2
2.25
2.5
2.75
3
3.25
3.5
3.75
4
4.25
4.5
4.75
5
5.25
Sum of Squares
df Mean Square F Sig.

185
In light of the significant difference in mean Sr/Ca ratios between Postclassic and
Historical cohorts, it should be noted that the low Historical values are partially
influenced by two individuals (YDL-85/50B and YDL-85/68B) with consistently very
low values throughout enamel development (see Section 7.5 and Figure 7.29). Several
explanations are presented in Section 7.5, but they are excluded from mean values in the
following tables (7.3-7.4) and figures.
First, for the purposes of developmental and nutritional significance and research
comparisons, the mean Sr/Ca data is condensed into fewer age intervals of greater length
(in years) in Table 7.3 below. Number of values (N) refers to the number of averaged
0.25 year intervals in the new age range available from each cohort. (These figures
exclude two Historical individuals (YDL-85/50B and /68B) with very low values.)
Table 7.3 Mean 86
Sr/43
Ca ratios for intervals between 0.5 and 5.5 years of age
Age Interval
Postclassic S.D.
N Historical S.D.
N Total Sample
S.D.
0.5-1.0
0.05863
0.01854
14
0.04610
0.01739
18
0.05158
0.01870
1.0-1.5 0.05670 0.01942 46 0.05006 0.01775 59 0.05297 0.01871
1.5-2.0 0.05907 0.02006 52 0.05287 0.01989 64 0.05565 0.02012
2.0-3.0 0.06622 0.02139 104 0.05731 0.01977 128 0.06131 0.02095
3.0-4.0 0.07703 0.02365 104 0.06543 0.02499 128 0.07063 0.02502
4.0-5.0 0.08617 0.02754 96 0.07189 0.02541 121 0.07821 0.02725
5.0-5.5 0.09156 0.02565 27 0.07356 0.03512 28 0.08240 0.03187
From these data, total Sr/Ca values can be calculated to contextualize this study in the
field of hard tissue strontium research. This is possible by considering the known
abundances of 86
Sr and 43
Ca relative to total strontium and calcium. 86
Sr is 9.86% of the

186
total naturally occurring abundance of strontium, while 43
Ca is 0.135% of total calcium.
With these constants, 86
Sr/43
Ca ratios are converted to total Sr/Ca values in Table 7.4.
The values range from 0.199 - 2.454 x 10-3
(0.000199-0.002454), with mean values
ranging from 0.631 - 1.254 x 10-3
(0.000631-0.001254). These ratios accord well with
other nutritional research: Sillen and Smith (1984) calculated bone Sr/Ca values ranging
from approximately 1.5 x 10-3
(0.0015) to 3.0 x 10-3
(0.003), while Mays (2003) has
reported more comparable bone Sr/Ca ratios in the range of 0.5 x 10-3
(0.0005) to 1.05 x
10-3
(0.00105), or a mean value of 0.855 x 10-3
(0.000855) among juveniles and 0.692 x
10-4
(0.000692) among adults. In particular, Mays’ (2003: Figure 1) mean ratio of
approximately 0.88 x 10-3
for infants less than a year old corresponds well with the total
Lamanai ratio of 0.706 x 10-3
for infants 0.5-1.0 years of age (Table 7.4).
The Sr/Ca ratios fall within the normal range of human enamel variation discussed in
Chapter Four, but there is considerable variation evident, even compared to other
research (Mays 2003; Sillen and Smith 1984). This can be attributed to numerous
factors: differences in food and water intake, and individual variation in strontium
absorption and hard tissue incorporation. Foremost, the continuous method of tissue
sampling in this investigation allows disclosure of much more previously undetected
variation.
Comparatively, the Postclassic sample can be distinguished by mean Sr/Ca values
higher than the total sample mean at all age intervals, while the reverse is true for the
Historical cohort, which is consistently lower than the total sample mean. This is clearly
apparent in Figure 7.3, which illustrates the distribution of mean Sr/Ca ratios for each
cohort presented in Table 7.4.
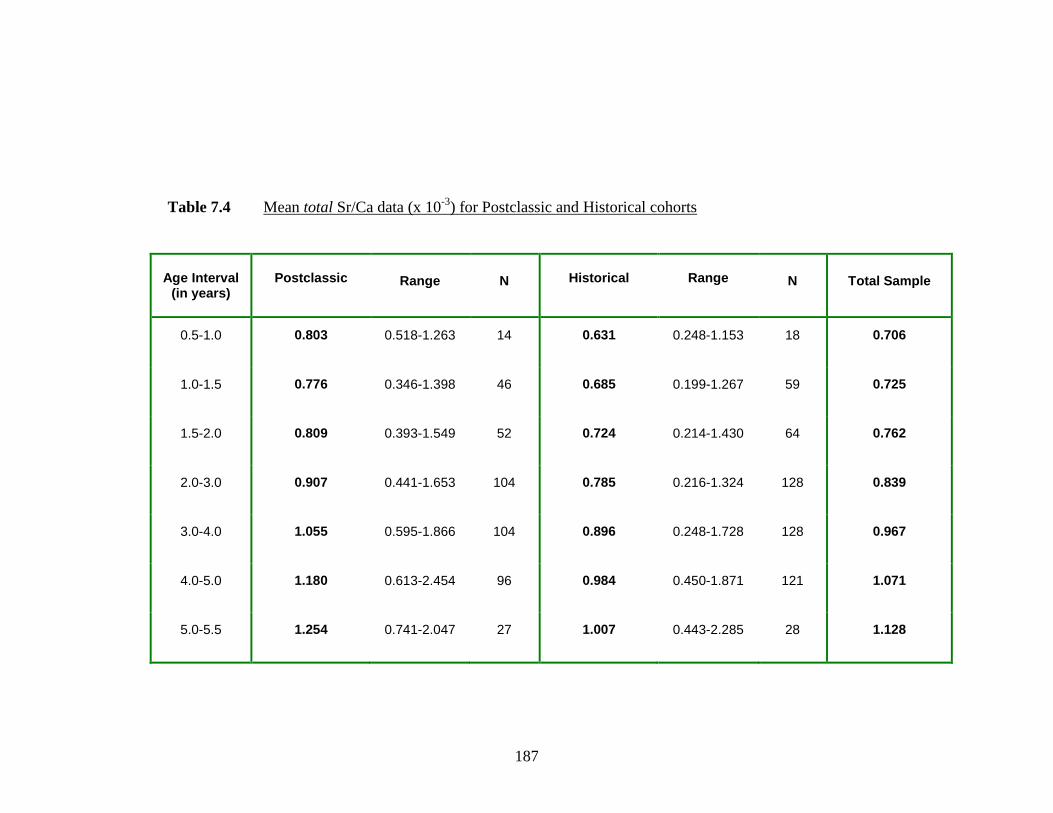
187
Table 7.4 Mean total Sr/Ca data (x 10-3
) for Postclassic and Historical cohorts
Age Interval (in years)
Postclassic
Range
N Historical
Range
N
Total Sample
0.5-1.0 0.803 0.518-1.263
14
0.631 0.248-1.153
18
0.706
1.0-1.5 0.776 0.346-1.398
46 0.685 0.199-1.267
59 0.725
1.5-2.0 0.809 0.393-1.549
52 0.724 0.214-1.430
64 0.762
2.0-3.0 0.907 0.441-1.653
104 0.785 0.216-1.324
128 0.839
3.0-4.0 1.055 0.595-1.866
104 0.896 0.248-1.728
128 0.967
4.0-5.0 1.180 0.613-2.454
96 0.984 0.450-1.871
121 1.071
5.0-5.5 1.254 0.741-2.047
27 1.007 0.443-2.285
28 1.128

188
Figure 7.3 Scatter plot distributions of total Sr/Ca for Lamanai cohorts
Distribution of Sr/Ca ratios in Historical Lamanai individuals
0.000
0.250
0.500
0.750
1.000
1.250
1.500
1.750
2.000
2.250
2.500
0 0.5-1.0 1.0-1.5 1.5-2.0 2.0-3.0 3.0-4.0 4.0-5.0 5.0-5.5 Age Interval (in years)
Sr/
Ca R
ati
o (
x 1
0-3
)
Distribution of Sr/Ca ratios in Postclassic Lamanai individuals
0.000
0.250
0.500
0.750
1.000
1.250
1.500
1.750
2.000
2.250
2.500
0 0.5-1.0 1.0-1.5 1.5-2.0 2.0-3.0 3.0-4.0 4.0-5.0 5.0-5.5 Age Interval (in years)
Sr/
Ca r
ati
o (
x 1
0-3
)

189
Significantly, like the specific 86
Sr/43
Ca pattern in Figure 7.1, the distribution of mean
total Sr/Ca ratios in Figure 7.3 reflects a pattern of increase and slight stabilization that
supports the hypothesis of this investigation. In fact, after the age of four years, it is
likely that Sr/Ca stabilizes at a greater rate than mean values would suggest; it appears
that mean values after this age are being inflated by several very high Sr/Ca ratios.
As also observed by Haverkort (2001: 183), the non-Gaussian distribution of
elements in teeth is not completely suited to the use of the sample mean as an indicator of
central tendency. Nonetheless, it remains a useful indicator of relative similarities or
distinctions. In this study, where single teeth can amass 800-900 element readings,
averaging counts over 0.25 year intervals for age, sex and temporal cohorts facilitates
data analysis and recognition of inter- and intra-sample variation.
Examining the total Sr/Ca pattern provides meaningful inferences for infant nutrition
of Lamanai Maya as a whole, particularly the significant increase in Sr/Ca at around nine
months that represents the start of food supplementation, but importantly, cohort
comparisons and extensive individual variation can also be observed with LA-ICP-MS.
The variation between and among temporal, age and sex cohorts is explored in the
following sections. Rather than total Sr/Ca, the unit of analysis is the 86
Sr/43
Ca ratio,
since these particular isotopes were directly quantified by LA-ICP-MS in this
investigation. Unless specified, the “Sr/Ca” ratio refers to the comparison of these single
isotopes rather than total Sr/Ca.

190
7.3 The Sr/Ca Patterns of Lamanai Cohorts
The Lamanai sample was evaluated on the basis of age (children vs. adults), sex
(female vs. male) and temporal period (Postclassic vs. Historical). Each category is
presented graphically by averaging Sr/Ca values for each 0.25 year age interval between
0.5 and 5.25-5.5 years. The following graph demonstrates the mean Sr/Ca values for
children, females and males from the Postclassic (N10) and Historical (YDL) periods at
Lamanai.
Figure 7.4 Sr/Ca patterns of Lamanai cohorts
Average Sr/Ca values for 0.25 yr intervals among
Postclassic and Historical Lamanai Individuals
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6
Age in years
Sr/
Ca
ra
tio
N10 Children N10 Females N10 Males YDL Children YDL Females YDL Males
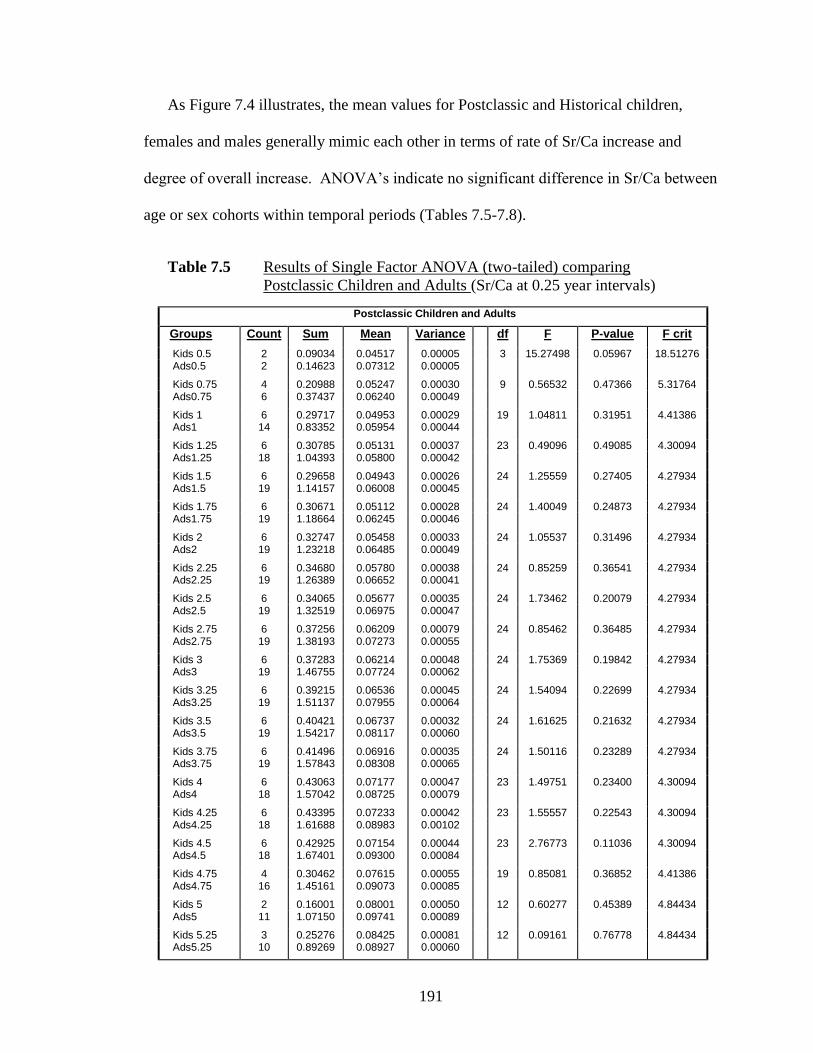
191
As Figure 7.4 illustrates, the mean values for Postclassic and Historical children,
females and males generally mimic each other in terms of rate of Sr/Ca increase and
degree of overall increase. ANOVA’s indicate no significant difference in Sr/Ca between
age or sex cohorts within temporal periods (Tables 7.5-7.8).
Table 7.5 Results of Single Factor ANOVA (two-tailed) comparing
Postclassic Children and Adults (Sr/Ca at 0.25 year intervals)
Postclassic Children and Adults
Groups Count Sum Mean Variance
df F P-value F crit
Kids 0.5 2 0.09034 0.04517 0.00005 3 15.27498 0.05967 18.51276 Ads0.5 2 0.14623 0.07312 0.00005
Kids 0.75 4 0.20988 0.05247 0.00030 9 0.56532 0.47366 5.31764 Ads0.75 6 0.37437 0.06240 0.00049
Kids 1 6 0.29717 0.04953 0.00029 19 1.04811 0.31951 4.41386 Ads1 14 0.83352 0.05954 0.00044
Kids 1.25 6 0.30785 0.05131 0.00037 23 0.49096 0.49085 4.30094 Ads1.25 18 1.04393 0.05800 0.00042
Kids 1.5 6 0.29658 0.04943 0.00026 24 1.25559 0.27405 4.27934 Ads1.5 19 1.14157 0.06008 0.00045
Kids 1.75 6 0.30671 0.05112 0.00028 24 1.40049 0.24873 4.27934 Ads1.75 19 1.18664 0.06245 0.00046
Kids 2 6 0.32747 0.05458 0.00033 24 1.05537 0.31496 4.27934 Ads2 19 1.23218 0.06485 0.00049
Kids 2.25 6 0.34680 0.05780 0.00038 24 0.85259 0.36541 4.27934 Ads2.25 19 1.26389 0.06652 0.00041
Kids 2.5 6 0.34065 0.05677 0.00035 24 1.73462 0.20079 4.27934 Ads2.5 19 1.32519 0.06975 0.00047
Kids 2.75 6 0.37256 0.06209 0.00079 24 0.85462 0.36485 4.27934 Ads2.75 19 1.38193 0.07273 0.00055
Kids 3 6 0.37283 0.06214 0.00048 24 1.75369 0.19842 4.27934 Ads3 19 1.46755 0.07724 0.00062
Kids 3.25 6 0.39215 0.06536 0.00045 24 1.54094 0.22699 4.27934 Ads3.25 19 1.51137 0.07955 0.00064
Kids 3.5 6 0.40421 0.06737 0.00032 24 1.61625 0.21632 4.27934 Ads3.5 19 1.54217 0.08117 0.00060
Kids 3.75 6 0.41496 0.06916 0.00035 24 1.50116 0.23289 4.27934 Ads3.75 19 1.57843 0.08308 0.00065
Kids 4 6 0.43063 0.07177 0.00047 23 1.49751 0.23400 4.30094 Ads4 18 1.57042 0.08725 0.00079
Kids 4.25 6 0.43395 0.07233 0.00042 23 1.55557 0.22543 4.30094 Ads4.25 18 1.61688 0.08983 0.00102
Kids 4.5 6 0.42925 0.07154 0.00044 23 2.76773 0.11036 4.30094 Ads4.5 18 1.67401 0.09300 0.00084
Kids 4.75 4 0.30462 0.07615 0.00055 19 0.85081 0.36852 4.41386 Ads4.75 16 1.45161 0.09073 0.00085
Kids 5 2 0.16001 0.08001 0.00050 12 0.60277 0.45389 4.84434 Ads5 11 1.07150 0.09741 0.00089
Kids 5.25 3 0.25276 0.08425 0.00081 12 0.09161 0.76778 4.84434 Ads5.25 10 0.89269 0.08927 0.00060

192
Table 7.6 Results of Single Factor ANOVA (two-tailed) comparing
Postclassic Females and Males (Sr/Ca at 0.25 year intervals)
Postclassic Females and Males
Groups* Count Sum Mean Variance df F P-value F crit
Fem0.75 5 0.33568 0.06714 0.00044 5 1.53018 0.28375 7.70865 Male0.75 1 0.03870 0.03870 N/A
Fem1 9 0.55501 0.06167 0.00048 13 0.24351 0.63059 4.74722 Male1 5 0.27851 0.05570 0.00044
Fem1.25 11 0.68828 0.06257 0.00049 17 1.43833 0.24787 4.49400 Male1.25 7 0.35565 0.05081 0.00028
Fem1.5 12 0.76424 0.06369 0.00052 18 0.92622 0.34934 4.45132 Male1.5 7 0.37733 0.05390 0.00034
Fem1.75 12 0.78660 0.06555 0.00056 18 0.67197 0.42371 4.45132 Male1.75 7 0.40005 0.05715 0.00030
Fem2 12 0.82220 0.06852 0.00055 18 0.88769 0.35930 4.45132 Male2 7 0.40998 0.05857 0.00038
Fem2.25 12 0.83502 0.06959 0.00045 18 0.72750 0.40555 4.45132 Male2.25 7 0.42886 0.06127 0.00037
Fem2.5 12 0.87157 0.07263 0.00055 18 0.56655 0.46193 4.45132 Male2.5 7 0.45361 0.06480 0.00035
Fem2.75 12 0.90650 0.07554 0.00066 18 0.45086 0.51095 4.45132 Male2.75 7 0.47543 0.06792 0.00040
Fem3 12 0.96391 0.08033 0.00079 18 0.48237 0.49673 4.45132 Male3 7 0.50364 0.07195 0.00038
Fem3.25 12 0.98693 0.08224 0.00077 18 0.35894 0.55700 4.45132 Male3.25 7 0.52444 0.07492 0.00045
Fem3.5 12 0.99102 0.08258 0.00077 18 0.10421 0.75077 4.45132 Male3.5 7 0.55116 0.07874 0.00036
Fem3.75 12 0.99497 0.08291 0.00076 18 0.00122 0.97255 4.45132 Male3.75 7 0.58346 0.08335 0.00056
Fem4 11 0.96493 0.08772 0.00105 17 0.00759 0.93165 4.49400 Male4 7 0.60549 0.08650 0.00049
Fem4.25 11 0.99639 0.09058 0.00160 17 0.01481 0.90465 4.49400 Male4.25 7 0.62049 0.08864 0.00023
Fem4.5 11 1.03343 0.09395 0.00110 17 0.02856 0.86793 4.49400 Male4.5 7 0.64057 0.09151 0.00053
Fem4.75 9 0.84098 0.09344 0.00102 15 0.16922 0.68703 4.60011 Male4.75 7 0.61062 0.08723 0.00073
Fem5 6 0.61787 0.10298 0.00078 10 0.43608 0.52556 5.11736 Male5 5 0.45363 0.09073 0.00114
Fem5.25 5 0.47264 0.09453 0.00071 9 0.43560 0.52779 5.31764 Male5.25 5 0.42005 0.08401 0.00056
* ANOVA was not possible at the 0.5 year interval due to insufficient data
(lack of earliest developing canine enamel among Postclassic males)

193
Table 7.7 Results of Single Factor ANOVA (two-tailed) comparing
Historical Children and Adults (Sr/Ca at 0.25 year intervals)
Historical Children and Adults
Groups Count Sum Mean Variance df F P-value F crit
Kids 0.5 2 0.08834 0.04417 0.00012 5 0.70560 0.44818 7.70865 Ads0.5 4 0.13081 0.03270 0.00029
Kids 0.75 4 0.15685 0.03921 0.00031 14 0.25278 0.62353 4.66719 Ads0.75 11 0.50077 0.04552 0.00051
Kids 1 9 0.42220 0.04691 0.00023 28 0.00377 0.95148 4.21001 Ads1 20 0.92937 0.04647 0.00036
Kids 1.25 9 0.45277 0.05031 0.00042 33 0.04976 0.82490 4.14909 Ads1.25 25 1.21257 0.04850 0.00044
Kids 1.5 9 0.47784 0.05309 0.00051 33 0.26645 0.60927 4.14909 Ads1.5 25 1.22192 0.04888 0.00042
Kids 1.75 9 0.48534 0.05393 0.00055 33 0.15423 0.69713 4.14909 Ads1.75 25 1.26390 0.05056 0.00047
Kids 2 9 0.48725 0.05414 0.00047 33 0.07166 0.79066 4.14909 Ads2 25 1.29715 0.05189 0.00047
Kids 2.25 9 0.49945 0.05549 0.00037 33 0.05010 0.82432 4.14909 Ads2.25 25 1.34172 0.05367 0.00046
Kids 2.5 9 0.51406 0.05712 0.00043 33 0.04641 0.83079 4.14909 Ads2.5 25 1.38267 0.05531 0.00048
Kids 2.75 9 0.52248 0.05805 0.00049 33 0.00741 0.93195 4.14909 Ads2.75 25 1.43209 0.05728 0.00054
Kids 3 9 0.52691 0.05855 0.00051 33 0.03772 0.84722 4.14909 Ads3 25 1.51049 0.06042 0.00065
Kids 3.25 9 0.54858 0.06095 0.00052 33 0.00381 0.95118 4.14909 Ads3.25 25 1.54020 0.06161 0.00082
Kids 3.5 9 0.58571 0.06508 0.00060 33 0.03528 0.85219 4.14909 Ads3.5 25 1.57552 0.06302 0.00086
Kids 3.75 9 0.62978 0.06998 0.00068 33 0.26277 0.61175 4.14909 Ads3.75 25 1.60799 0.06432 0.00085
Kids 4 9 0.67403 0.07489 0.00080 33 0.76154 0.38935 4.14909 Ads4 25 1.64150 0.06566 0.00072
Kids 4.25 9 0.67580 0.07509 0.00078 33 1.05428 0.31222 4.14909 Ads4.25 25 1.62540 0.06502 0.00059
Kids 4.5 9 0.69170 0.07686 0.00087 33 0.82536 0.37041 4.14909 Ads4.5 25 1.67280 0.06691 0.00077
Kids 4.75 9 0.69766 0.07752 0.00078 26 0.74953 0.39486 4.24170 Ads4.75 18 1.19914 0.06662 0.00103
Kids 5 5 0.39210 0.07842 0.00023 15 0.22539 0.64229 4.60011 Ads5 11 0.76260 0.06933 0.00167
Kids 5.25 4 0.31256 0.07814 0.00031 11 0.02911 0.86793 4.96459 Ads5.25 8 0.59250 0.07406 0.00204

194
Table 7.8 Results of Single Factor ANOVA (two-tailed) comparing
Historical Females and Males (Sr/Ca at 0.25 year intervals)
Historical Females and Males
Groups* Count Sum Mean Variance df F P-value F crit
Fem0.75 6 0.20677 0.03446 0.00024 10 4.18371 0.07114 5.11736
Male0.75 5 0.29401 0.05880 0.00057
Fem1 8 0.34199 0.04275 0.00043 19 0.49835 0.48926 4.41386 Male1 12 0.58739 0.04895 0.00033
Fem1.25 13 0.58623 0.04509 0.00049 24 0.71174 0.40756 4.27934 Male1.25 12 0.62634 0.05220 0.00039
Fem1.5 13 0.58174 0.04475 0.00045 24 1.10439 0.30422 4.27934 Male1.5 12 0.64018 0.05335 0.00039
Fem1.75 13 0.60493 0.04653 0.00053 24 0.93576 0.34343 4.27934 Male1.75 12 0.65897 0.05491 0.00040
Fem2 13 0.64215 0.04940 0.00061 24 0.34790 0.56105 4.27934 Male2 12 0.65500 0.05458 0.00035
Fem2.25 13 0.64362 0.04951 0.00055 24 1.01163 0.32498 4.27934 Male2.25 12 0.69810 0.05818 0.00036
Fem2.5 13 0.67428 0.05187 0.00055 24 0.65850 0.42541 4.27934 Male2.5 12 0.70839 0.05903 0.00041
Fem2.75 13 0.70098 0.05392 0.00061 24 0.55458 0.46400 4.27934 Male2.75 12 0.73111 0.06093 0.00049
Fem3 13 0.74855 0.05758 0.00083 24 0.32557 0.57381 4.27934 Male3 12 0.76193 0.06349 0.00050
Fem3.25 13 0.77394 0.05953 0.00111 24 0.13688 0.71478 4.27934 Male3.25 12 0.76626 0.06386 0.00057
Fem3.5 13 0.78107 0.06008 0.00120 24 0.26361 0.61255 4.27934 Male3.5 12 0.79445 0.06620 0.00055
Fem3.75 13 0.82460 0.06343 0.00127 24 0.02418 0.87777 4.27934 Male3.75 12 0.78339 0.06528 0.00046
Fem4 13 0.84387 0.06491 0.00114 24 0.02013 0.88840 4.27934 Male4 12 0.79763 0.06647 0.00033
Fem4.25 13 0.81894 0.06300 0.00088 24 0.18078 0.67465 4.27934 Male4.25 12 0.80646 0.06721 0.00031
Fem4.5 13 0.83177 0.06398 0.00109 24 0.29363 0.59311 4.27934 Male4.5 12 0.84103 0.07009 0.00047
Fem4.75 10 0.68550 0.06855 0.00134 17 0.07696 0.78501 4.49400 Male4.75 8 0.51364 0.06421 0.00077
Fem5 6 0.44710 0.07452 0.00219 10 0.19533 0.66895 5.11736 Male5 5 0.31550 0.06310 0.00135
Fem5.25 5 0.36461 0.07292 0.00186 7 0.00728 0.93477 5.98737 Male5.25 3 0.22789 0.07596 0.00341
* ANOVA was not possible at the 0.5 year interval due to insufficient data
(lack of earliest developing canine enamel among Historical males)

195
Lack of significant difference in the mean Sr/Ca ratios of Lamanai children, females
and males within temporal period reflects a general weaning pattern shared among
Lamanai infants in the Postclassic and Historical periods. According to enamel Sr/Ca,
this weaning process is long and protracted, lasting as much as five years.
Specifically, within temporal periods, Sr/Ca patterns exhibit comparable rates of
increase and degree of overall increase. This is not unexpected as the composition of
Postclassic and Historical samples is generally uniform in socioeconomic status and
burial context. Figures 7.5a and 7.5b display the mean Sr/Ca patterns of children, adult
females and males for the Postclassic and Historical periods.
Comparing individuals by time period reflects a slight difference in the feeding
experiences of Lamanai infants before and after Spanish contact. In the Postclassic graph
(Figure 7.5a), the average Sr/Ca values for children seem to parallel the rate of increase
among females, but progresses at a slower rate compared to adult males (whose slope is
steeper).
Though not significantly, Postclassic juvenile Sr/Ca values are clearly lower than
male and female adults (Figure 7.5a). If Sr/Ca ratio is indicative of relative solid food
intake, as proposed in this study, then this suggests that those who died in childhood were
slightly delayed in the supplementation of food compared to those who survived to
adulthood (also see Figure 7.8).
Early supplementation (i.e., prior to six postnatal months) has been widely linked to
increased morbidity and mortality (see Chapter Five), but these results also indicate that
delayed supplementation later in infancy, when toddlers experience rapid growth and
development, also has negative consequences. Extended reliance on breast milk and

196
Figure 7.5 Average Sr/Ca for Postclassic and Historical individuals
a)
b)
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6
Sr/
Ca r
ati
o
Age in years
Average Sr/Ca values for 0.25 yr intervals among Postclassic individuals
N10 Children
N10 Females
N10 Males
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6
Sr/
Ca r
ati
o
Age in years
Average Sr/Ca values for 0.25 yr intervals among Historical individuals
YDL Children
YDL Females
YDL Males

197
accompanied malnutrition due to inadequate food and nutritional supplementation in
these children may account for their premature mortality.
In contrast, Historical groups are strikingly similar in both Sr/Ca ratios and rate of
increase. (The slight difference being that Sr/Ca increase speeds up between the ages of
3.25 and 4 years in Historical children [Figure 7.5b].) The effect of colonization seems to
be an “homogenizing” of childhood nutritional experiences.
Furthermore, all Historical groups are slightly diminished (though not significantly)
in mean Sr/Ca compared to Postclassic adults. In Figure 7.6, the total Historical sample
is compared to Postclassic males and females. This graph clearly indicates that the rate
of food supplementation increase among Historical individuals is generally stable
throughout the entire weaning process.
Figure 7.6 Comparison of total Historical sample with Postclassic adults
All Historical vs. Postclassic Adults
(average Sr/Ca for 0.25 yr intervals)
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6
Age in years
Sr/
Ca
ra
tio
N10 Females
N10 M ales
All YDL

198
However, Postclassic males display a steeper Sr/Ca pattern, or accelerated rate of
solid food supplementation, after approximately two years of age. Prior to one year of
age, their Sr/Ca values are the lowest, suggesting a higher proportion of breast milk
intake compared to Postclassic females and Historical individuals. Postclassic females
and the Historical sample share a similar rate of Sr/Ca change, but higher Sr/Ca values
among Postclassic females suggest that they were supplemented with solid foods earlier
than Historical individuals, as well as Postclassic males.
Figure 7.6 suggests that all Historical individuals, including those who lived past
childhood, were delayed in reaching similar levels of food supplementation compared to
Postclassic individuals who survived to adulthood. The similarity among Historical
individuals may reflect a nutritional adaptation to the changing socio-economic
conditions of colonial Lamanai, i.e., access to food resources and/or disease prevalence.
In addition to re-settlement, all Lamanai Maya were “reduced” to a collective,
subordinate, social position in colonial times that might have affected their ability to
supplement infants with food, particularly those afflicted with illness.
In this study, it is proposed that the extent of differences between cohorts can be
interpreted by adjusting the graphical distributions of mean Sr/Ca. Rates of Sr/Ca change
(slope) and total increase are generally comparable between groups (see Figures 7.2 and
7.4), so it is postulated that “shifting” the graphs can reflect tangible differences in the
infant feeding patterns of different cohorts.
Two sets of Sr/Ca data are compared in this way, i.e., Postclassic vs. Historical
individuals, adults vs. children, males vs. females (see Figures 7.7-7.14). Based on
analogous rates of change and overall increase, it is assumed that differences in mean

199
Sr/Ca reflect a time delay in one group reaching comparable Sr/Ca values as the other.
Between two groups, the greater Sr/Ca pattern is horizontally shifted forward in
development time (to the right) to “synchronize” with the lower Sr/Ca pattern (see
Figures 7.7-7.14). The extent required to equalize mean Sr/Ca values reflects the
possible delay in time of the lower Sr/Ca group reaching comparable levels of Sr/Ca, or
food supplementation, as the higher Sr/Ca group. This approach provides a visual
indication of the potential differences between cohorts that may, or may not, be
statistically significant (e.g., Figures 7.7-7.14).
Comparing Lamanai Maya in this way indicates that Historical individuals may have
been delayed by up to 1.25 years in reaching comparable levels of Sr/Ca as Postclassic
individuals (Figure 7.7). An adjustment of 1.25 years results in two distributions that are
generally equal in Sr/Ca values, as well as rate of change (although a small discrepancy
occurs in Historical Sr/Ca after 4 years of age, when Sr/Ca values diminish slightly).
While mean Sr/Ca peaks at 5-5.25 years of age among Postclassic Maya, Historical Maya
may not have reached peak Sr/Ca until 6.25-6.5 years of age. This delay is considered a
maximum difference between samples and assumes that mean Sr/Ca values exhibit
minimal variation, which is known not to be the case (see Chapter Four).

200
Figure 7.7 Comparison of Postclassic and Historical cohorts
a)
b) Time shifted by 1.25 years
Adjusted Postclassic and Historical Lamanai Individuals
(average Sr/Ca for 0.25 yr intervals)
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6
Age in years
Sr/
Ca
ra
tio
Postclassic
Historical
Comparison of all Postclassic and Historical Individuals
(average Sr/Ca for 0.25 yr intervals)
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6
Age in years
Sr/
Ca r
ati
o
Postclassic
Historical

201
Beyond gross temporal comparisons, the Lamanai sample was also compared by
cohorts based on age and sex. In many cases, ANOVA’s indicate lack of statistical
difference (Appendix C). However, some slight discrepancies still exist between
Lamanai groups. As stated earlier, “shifting” the distribution of two sets of data to
“synchronize” mean Sr/Ca values can provide an indication of the extent of time
differences in reaching comparable levels of Sr/Ca, or food supplementation.
Figures 7.8-7.14 illustrate comparative Sr/Ca distributions of Postclassic children vs.
adults; Postclassic males vs. females; Historical children vs. adults; Historical males vs.
females; Postclassic vs. Historical children; Postclassic vs. Historical females and
Postclassic vs. Historical males. They are accompanied by “adjusted” graphs reflecting
the possible time delay in the lower Sr/Ca group. To reiterate, the higher Sr/Ca pattern is
shifted to the right to match up with the lower Sr/Ca values, but the extent of the
adjustment (in years) reflects the delay of the lower group.
In Figure 7.8, teeth from individuals who died in childhood are compared to deceased
adults to ascertain the effects of nutritional status on relative lifespan. Diet, weaning age,
infectious disease and socio-economic factors are clearly associated with rates of
morbidity and mortality in archaeological and living populations (Cook 1981; Goodman
and Armelagos 1988, 1989; Herring et al. 1998; Katzenberg et al. 1996; Stuart-Macadam
and Dettwyler 1995; Swedlund and Ball 1998, among many others). Individuals who
died in childhood are particularly burdened in life with ill health, as reflected in hard
tissue pathologies such as enamel hypoplasia, Wilson bands and porotic hyperostosis
(Blakey and Armelagos 1985; Cook and Buikstra 1979; Goodman and Armelagos 1988,

202
Figure 7.8 Comparison of Postclassic Children and Adults
a)
b) Time shifted by 1.5 years
Postclassic Children vs. Adults
(average Sr/Ca for 0.25 yr intervals)
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6
Age in years
Sr/
Ca r
ati
o
N10 Children N10 Adults
Adjusted Postclassic
Children and Adults
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6
Age in years
Sr/
Ca r
ati
o
N10 Children N10 Adults

203
1989; Huhne-Osterloh and Grupe 1989; Katzenberg et al. 1996; Song 1997; Stuart-
Macadam 1985; Wright 1990).
Figure 7.8a reflects the differences in average Sr/Ca ratio between Postclassic
children and adults, with children averaging lower values at every age interval. While
not statistically significant for any age interval (see Appendix C), Figure 7.8b is an
adjusted scatter plot that demonstrates the possible “delay” in children’s Sr/Ca compared
to adults.
Since Sr/Ca patterns in both cohorts mimic each other in the rate of increase,
adjusting the graphs can reflect a quantifiable difference in the infant feeding patterns of
individuals who died in childhood versus adulthood. In order to synchronize the Sr/Ca
patterns between the two groups, the adult pattern is staggered to begin later in
development age, but in fact, the graph illustrates an approximately 1.5 year “delay”
among Postclassic individuals who died in childhood (in obtaining a comparable level of
solid food supplementation of Postclassic individuals who survived to adulthood).
In comparing Postclassic adults (Figure 7.9), there is no significant difference in
Sr/Ca values at any age interval (Appendix C). Graphically, Postclassic males appear
slightly “delayed”, which is reflected in Figure 7.9b, as well as Figure 7.6.
Synchronizing the two graphs indicates that Postclassic males may have only been
supplemented with comparable levels of solid food 0.25-0.5 years after Postclassic
females. A slightly steeper rate of increase (accelerated food supplementation) in males
may compensate for the delay.

204
Figure 7.9 Comparison of Postclassic Females and Males
a)
b) Time shifted by 0.25 years
Postclassic Females vs. Males
(average Sr/Ca for 0.25 yr intervals)
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6
Age in years
Sr/
Ca r
ati
o
N10 FemalesN10 Males
Adjusted Postclassic Adults
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6
Age in years
Sr/
Ca r
ati
o
N10 FemalesN10 Males

205
If one assumes that male children were preferentially treated compared to females, as
archaeological, ethnohistorical and bone chemical evidence suggest greater adult male
social status in Maya society, e.g., increased access to meat and seafood (White 1986,
1999; White et al. 1993), then the Sr/Ca data reflects such increased investment of
maternal care on male infants. In many patriarchal societies, e.g., Canada, Sweden,
Ireland, France, Guatemala, Ecuador, Brazil, Peru, Taiwan, India, Jordan, Liberia and
Botswana, infant boys have been observed to nurse longer than females (see Fildes 1986;
McKee 1984). Delayed food supplementation in infancy, i.e., after six months of age,
when children require increased nutrients to sustain energy and growth demands, has
potentially negative consequences and is discussed further in Chapter Eight.
In terms of socioeconomic status, the Postclassic sample as a whole can be
characterized as an advantaged group (Pendergast 1981; White 1986). Two adult male
individuals are particularly recognized elites: N10-2/20 and N10-4/46A. But while N10-
4/46A exhibits a generally increasing Sr/Ca pattern over time, values for N10-2/20 do not
significantly start to increase until 2.5 years of age (see Figure 7.17), which suggests a
delay period much longer than the 0.25-0.5 year period suggested above. (Remember,
though, that this individual does not have enamel sampled from before one year of age,
when Sr/Ca [food supplementation] might have already started to rise).

206
Figure 7.10 Comparison of Historical Children and Adults
As stated earlier, and presented graphically above, all Historical individuals likely
shared a common pattern of breast milk consumption and delayed food supplementation
compared to Postclassic individuals. ANOVA’s indicate no significant difference in
Sr/Ca at any age interval (Appendix C).
Figure 7.11 illustrates the mean Sr/Ca patterns of Historical adults. There is no
statistical difference in Sr/Ca patterns between males and females (Appendix C).
Adjusting the scatter plots, however, suggests that Historical females may be delayed in
reaching similar levels of food supplementation (Sr/Ca) by approximately one year
compared to males (Figure 7.11b). This contrasts the prehistoric pattern, when it is the
males who show slight delays in reaching comparable Sr/Ca levels.
Historical Children vs. Adults
(average Sr/Ca for 0.25 yr intervals)
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6
Age in years
Sr/
Ca r
ati
o
YDL ChildrenYDL Adults

207
Figure 7.11 Comparison of Historical Females and Males
a)
b) Time shifted by 1.0 year
Historical Females vs. Males
(average Sr/Ca for 0.25 yr intervals)
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6
Age in years
Sr/
Ca r
ati
o
YDL FemalesYDL Males
Adjusted Historical Adults
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6
Age in years
Sr/
Ca r
ati
o
YDL FemalesYDL Males

208
Figure 7.12 Comparison of Postclassic and Historical Children
Despite differing social conditions, Sr/Ca patterns suggest that Postclassic and
Historical Lamanai infants who died in later childhood shared similar patterns of infant
nutrition, with no statistical difference in Sr/Ca over time (Figure 7.12). As it has been
shown above that both Postclassic and Historical children are “delayed” compared to
Postclassic adults (who also exhibit higher Sr/Ca values than Historical adults), the
parallel graphs of Lamanai individuals who died in childhood reflect a pattern of Sr/Ca
increase and food supplementation that is likely sub-optimal for development and, in the
long term, detrimental to survival.
Postclassic vs. Historical Children
(average Sr/Ca for 0.25 yr intervals)
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6
Age in years
Sr/
Ca r
ati
o
N10 Children
YDL Children

209
Figure 7.13 Comparison of Postclassic and Historical Females
a)
b) Time shifted by 2.25 years
Postclassic vs. Historical Females
(average Sr/Ca for 0.25 yr intervals)
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6
Age in years
Sr/
Ca r
ati
o
N10 FemalesYDL Females
Adjusted Females
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6
Age in years
Sr/
Ca r
ati
o
N10 FemalesYDL Females

210
The comparisons between Postclassic and Historical females show the greatest
number of differences in Sr/Ca, with N10 females having higher Sr/Ca ratios at every age
interval. Specifically, ANOVA’s highlight significance at the following age intervals:
0.50-0.749 (p=0.037); 0.75-0.99 (p=0.016); 1.50-1.749 (p=0.042); 2.25-2.49 (0.035);
2.50-2.749 (0.037); 2.75-2.99 (0.043); and 4.50-4.749 (0.038) (Appendix C: Table C.13).
In the adjusted graph (Figure 7.13b), Historical women seem to be delayed by as much as
2.25 years in reaching comparable Sr/Ca levels as Postclassic women. This suggests that
Postclassic females were supplemented with relatively more weanling foods than
Historical female infants of the same age and likely began the weaning process earlier as
well.
Postclassic and Historical males start off with comparable Sr/Ca values and rate of
increase, but noticeably differ after the age of two (Figure 7.14). After 2-2.25 years,
Postclassic males consume supplementary foods at an accelerated rate compared to
Historical males, whose rate of supplementation continues increasing at the same rate
prior to 2 years of age. This acceleration of solid food intake after 2 years of age (most
likely comprising the predominant food source) is also apparent in graphs for Lamanai
individuals with a pattern of “steady increase” in Sr/Ca (see Figures 7.22 and 7.23).
Figure 7.14b suggests that after 2.25 years of age, Historical males are “delayed” in
reaching comparable Postclassic Sr/Ca values by approximately 1.5 years. In total,
Historical male toddlers were relying on more breast milk / less solid food intake than
their prehistoric counterparts. Statistically, however, Postclassic and Historical males
only differ in average Sr/Ca at the 4.0-4.249 age interval (p=0.047) and 4.25-4.49 age
interval (p=0.016) (Appendix C: Table C.14).
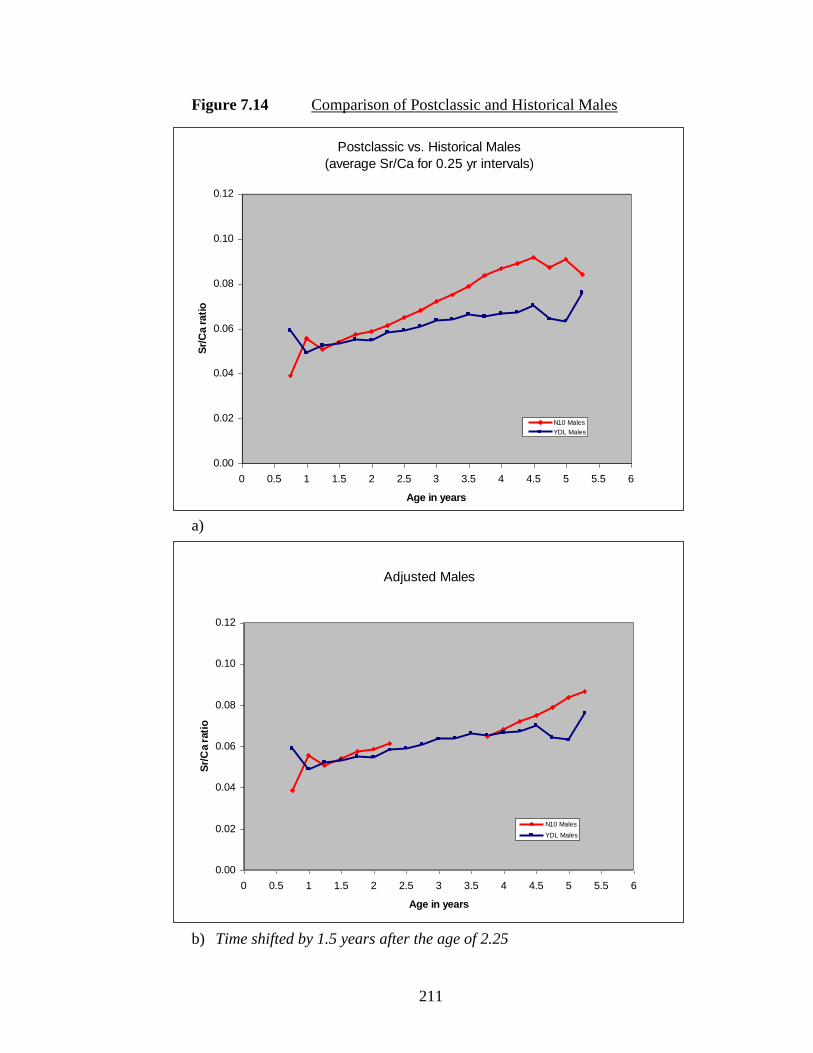
211
Figure 7.14 Comparison of Postclassic and Historical Males
a)
b) Time shifted by 1.5 years after the age of 2.25
Postclassic vs. Historical Males
(average Sr/Ca for 0.25 yr intervals)
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6
Age in years
Sr/
Ca r
ati
o
N10 Males
YDL Males
Adjusted Males
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6
Age in years
Sr/
Ca r
ati
o
N10 Males
YDL Males

212
7.4 Sr/Ca Patterns among Lamanai Individuals
The following tables display average Sr/Ca values for 0.25 year intervals among
Postclassic and Historical period children, females and males. Age intervals are in years
and represent a 0.25 year period (3 months). Blank values in each table indicate areas in
which enamel is either missing, or could not be ablated due to poor preservation.
Underlined values represent the highest Sr/Ca reading for each individual. Values for
“Greatest Increase” are based on the highest mean Sr/Ca interval and the first mean Sr/Ca
reading for the individual. In some cases, when the lowest value is not the first (earliest)
Sr/Ca reading, but rather, is the second interval reading, then this value is used to derive
the degree of “greatest increase”. Accompanying the tables are scatter plots of the mean
Sr/Ca values for each age interval among Postclassic and Historical children, females and
males (Figures 7.15-7.20).

213
Table 7.9 Average Sr/Ca values for 0.25 year intervals of Postclassic Children
Age Interval
N10-2/16
N10-2/21
N10-2/44
N10-2/49
N10-2/50
N10-4/2A
AVERAGE
S.D.
N
RANGE
0.50-0.74 - 0.040 0.050 - - - 0.045 0.007 2 0.040 - 0.050
0.75-0.99 - 0.039 0.058 - 0.038 0.075 0.052 0.017 4 0.038 - 0.075
1.00-1.24 0.052 0.043 0.059 0.025 0.042 0.076 0.050 0.017 6 0.025 - 0.076
1.25-1.49 0.058 0.042 0.056 0.027 0.041 0.083 0.051 0.019 6 0.027 - 0.083
1.50-1.74 0.055 0.041 0.053 0.029 0.043 0.076 0.049 0.016 6 0.029 - 0.076
1.75-1.99 0.057 0.043 0.053 0.031 0.043 0.080 0.051 0.017 6 0.031 - 0.080
2.00-2.24 0.060 0.044 0.066 0.032 0.044 0.083 0.055 0.018 6 0.032 - 0.083
2.25-2.49 0.062 0.043 0.073 0.035 0.047 0.086 0.058 0.019 6 0.035 - 0.086
2.50-2.74 0.064 0.044 0.052 0.041 0.049 0.092 0.057 0.019 6 0.041 - 0.092
2.75-2.99 0.064 0.046 0.051 0.045 0.048 0.118 0.062 0.028 6 0.045 - 0.118
3.00-3.24 0.065 0.054 0.051 0.047 0.052 0.105 0.062 0.022 6 0.047 - 0.105
3.25-3.49 0.071 0.056 0.048 0.051 0.061 0.105 0.065 0.021 6 0.048 - 0.105
3.50-3.74 0.078 0.057 0.048 0.059 0.064 0.098 0.067 0.018 6 0.048 - 0.098
3.75-3.99 0.082 0.057 0.045 0.065 0.068 0.098 0.069 0.019 6 0.045 - 0.098
4.00-4.24 0.091 0.059 0.045 0.064 0.068 0.104 0.072 0.022 6 0.045 - 0.104
4.25-4.49 0.085 0.061 0.045 0.065 0.075 0.103 0.072 0.020 6 0.045 - 0.103
4.50-4.74 0.085 0.058 0.051 0.057 0.073 0.106 0.072 0.021 6 0.051 - 0.106
4.75-4.99 0.095 - 0.056 0.056 - 0.098 0.076 0.023 4 0.056 - 0.098
5.00-5.24 0.096 - - 0.064 - - 0.080 0.022 2 0.064 - 0.096
5.25-5.49 0.088 - - 0.054 - - 0.071 0.024 2 0.054 - 0.088
Greatest increase
1.830
1.522
1.442
2.569
1.990
1.576
1.821
0.420
Figure 7.15 Graph of Sr/Ca change among Postclassic children
Sr/Ca change among Postclassic Lamanai children
(3 month intervals)
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
0.16
0.18
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6
Age in Years
Sr/
Ca R
ati
o
N10-2/16
N10-2/21
N10-2/44
N10-2/49
N10-2/50
N10-4/2A
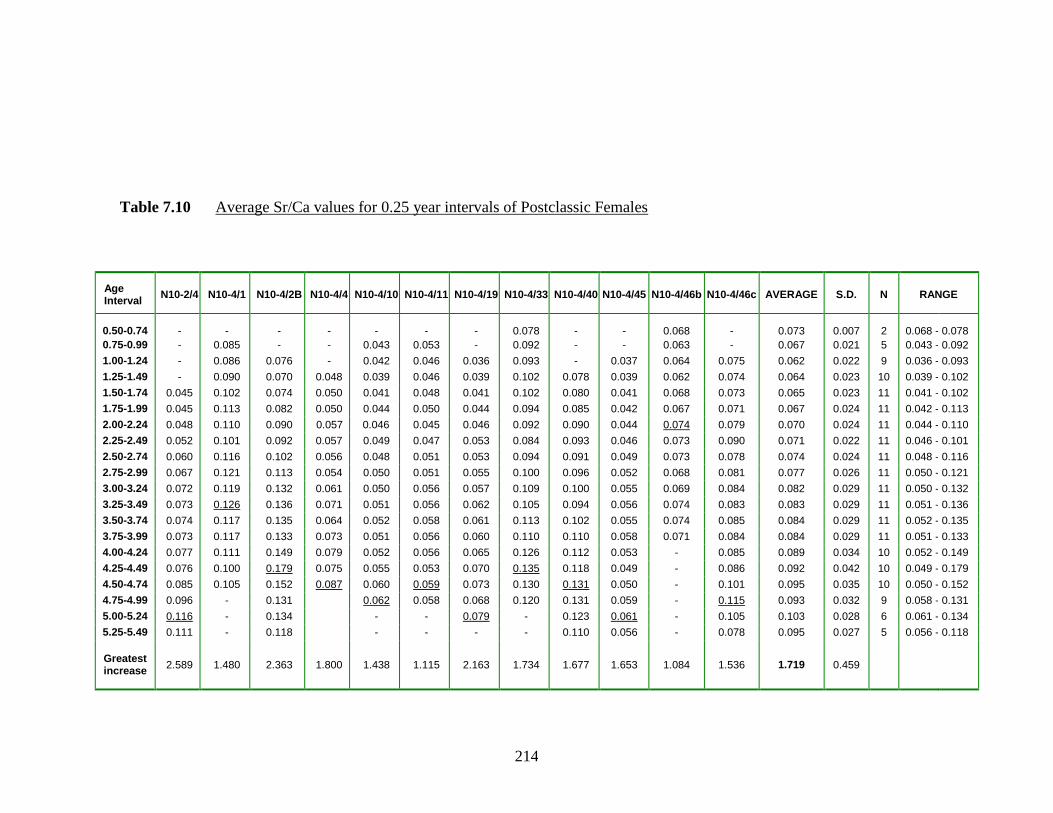
214
Table 7.10 Average Sr/Ca values for 0.25 year intervals of Postclassic Females
Age Interval
N10-2/4 N10-4/1 N10-4/2B
N10-4/4 N10-4/10 N10-4/11 N10-4/19 N10-4/33 N10-4/40 N10-4/45 N10-4/46b N10-4/46c AVERAGE S.D. N RANGE
0.50-0.74 - - - - - - - 0.078 - - 0.068 - 0.073 0.007 2 0.068 - 0.078
0.75-0.99 - 0.085 - - 0.043 0.053 - 0.092 - - 0.063 - 0.067 0.021 5 0.043 - 0.092
1.00-1.24 - 0.086 0.076 - 0.042 0.046 0.036 0.093 - 0.037 0.064 0.075 0.062 0.022 9 0.036 - 0.093
1.25-1.49 - 0.090 0.070 0.048 0.039 0.046 0.039 0.102 0.078 0.039 0.062 0.074 0.064 0.023 10 0.039 - 0.102
1.50-1.74 0.045 0.102 0.074 0.050 0.041 0.048 0.041 0.102 0.080 0.041 0.068 0.073 0.065 0.023 11 0.041 - 0.102
1.75-1.99 0.045 0.113 0.082 0.050 0.044 0.050 0.044 0.094 0.085 0.042 0.067 0.071 0.067 0.024 11 0.042 - 0.113
2.00-2.24 0.048 0.110 0.090 0.057 0.046 0.045 0.046 0.092 0.090 0.044 0.074 0.079 0.070 0.024 11 0.044 - 0.110
2.25-2.49 0.052 0.101 0.092 0.057 0.049 0.047 0.053 0.084 0.093 0.046 0.073 0.090 0.071 0.022 11 0.046 - 0.101
2.50-2.74 0.060 0.116 0.102 0.056 0.048 0.051 0.053 0.094 0.091 0.049 0.073 0.078 0.074 0.024 11 0.048 - 0.116
2.75-2.99 0.067 0.121 0.113 0.054 0.050 0.051 0.055 0.100 0.096 0.052 0.068 0.081 0.077 0.026 11 0.050 - 0.121
3.00-3.24 0.072 0.119 0.132 0.061 0.050 0.056 0.057 0.109 0.100 0.055 0.069 0.084 0.082 0.029 11 0.050 - 0.132
3.25-3.49 0.073 0.126 0.136 0.071 0.051 0.056 0.062 0.105 0.094 0.056 0.074 0.083 0.083 0.029 11 0.051 - 0.136
3.50-3.74 0.074 0.117 0.135 0.064 0.052 0.058 0.061 0.113 0.102 0.055 0.074 0.085 0.084 0.029 11 0.052 - 0.135
3.75-3.99 0.073 0.117 0.133 0.073 0.051 0.056 0.060 0.110 0.110 0.058 0.071 0.084 0.084 0.029 11 0.051 - 0.133
4.00-4.24 0.077 0.111 0.149 0.079 0.052 0.056 0.065 0.126 0.112 0.053 - 0.085 0.089 0.034 10 0.052 - 0.149
4.25-4.49 0.076 0.100 0.179 0.075 0.055 0.053 0.070 0.135 0.118 0.049 - 0.086 0.092 0.042 10 0.049 - 0.179
4.50-4.74 0.085 0.105 0.152 0.087 0.060 0.059 0.073 0.130 0.131 0.050 - 0.101 0.095 0.035 10 0.050 - 0.152
4.75-4.99 0.096 - 0.131 0.062 0.058 0.068 0.120 0.131 0.059 - 0.115 0.093 0.032 9 0.058 - 0.131
5.00-5.24 0.116 - 0.134 - - 0.079 - 0.123 0.061 - 0.105 0.103 0.028 6 0.061 - 0.134
5.25-5.49 0.111 - 0.118 - - - - 0.110 0.056 - 0.078 0.095 0.027 5 0.056 - 0.118
Greatest increase
2.589
1.480
2.363
1.800
1.438
1.115
2.163
1.734
1.677
1.653
1.084
1.536
1.719
0.459

215
Table 7.11 Average Sr/Ca values for 0.25 year intervals of Postclassic Males
Age Interval
N10-2/5 N10-2/20 N10-2/22 N10-2/40 N10-2/42 N10-4/43 N10-4/46A AVERAGE S.D. N RANGE
0.50-0.74 - - - - - - - - - 0 - -
0.75-0.99 - - - 0.039 - - - 0.039 n/a 1 0.039 -
1.00-1.24 0.092 0.046 - 0.041 0.043 - 0.055 0.056 0.021 5 0.041 - 0.092
1.25-1.49 0.085 0.044 0.033 0.042 0.045 0.049 0.058 0.051 0.017 7 0.033 - 0.085
1.50-1.74 0.091 0.046 0.032 0.047 0.046 0.055 0.061 0.054 0.019 7 0.032 - 0.091
1.75-1.99 0.090 0.048 0.035 0.049 0.055 0.059 0.063 0.057 0.017 7 0.035 - 0.090
2.00-2.24 0.095 0.048 0.033 0.050 0.052 0.066 0.066 0.059 0.020 7 0.033 - 0.095
2.25-2.49 0.095 0.050 0.033 0.057 0.057 0.071 0.066 0.061 0.019 7 0.033 - 0.095
2.50-2.74 0.097 0.056 0.036 0.059 0.064 0.076 0.066 0.065 0.019 7 0.036 - 0.097
2.75-2.99 0.103 0.060 0.037 0.064 0.068 0.077 0.066 0.068 0.020 7 0.037 - 0.103
3.00-3.24 0.109 0.070 0.043 0.068 0.069 0.079 0.065 0.072 0.020 7 0.043 - 0.109
3.25-3.49 0.118 0.071 0.050 0.070 0.068 0.082 0.064 0.075 0.021 7 0.050 - 0.118
3.50-3.74 0.119 0.073 0.061 0.079 0.069 0.082 0.069 0.079 0.019 7 0.061 - 0.119
3.75-3.99 0.131 0.080 0.064 0.083 0.064 0.092 0.069 0.083 0.024 7 0.064 - 0.131
4.00-4.24 0.130 0.078 0.069 0.085 0.072 0.101 0.071 0.086 0.022 7 0.069 - 0.130
4.25-4.49 0.119 0.082 0.077 0.079 0.085 0.099 0.079 0.089 0.015 7 0.077 - 0.119
4.50-4.74 0.140 0.087 0.075 0.088 0.074 0.099 0.078 0.092 0.023 7 0.074 - 0.140
4.75-4.99 0.143 0.086 0.065 0.078 0.061 0.093 0.084 0.087 0.027 7 0.061 - 0.143
5.00-5.24 0.149 0.085 0.068 - 0.068 - 0.083 0.091 0.034 5 0.068 - 0.149
5.25-5.49 0.126 0.080 0.071 - 0.069 - 0.075 0.084 0.024 5 0.069 - 0.126
Greatest increase
1.624
1.874
2.334
2.263
1.957
2.038
1.519
1.944
0.303

216
Figure 7.16 Graph of Sr/Ca change among Postclassic females
Figure 7.17 Graph of Sr/Ca change among Postclassic males
Sr/Ca change among Postclassic Lamanai males
(3 month intervals)
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
0.16
0.18
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6
Age in Years
Sr/
Ca R
ati
o
N10-2/5
N10-2/20
N10-2/22
N10-2/40
N10-2/42
N10-4/43
N10-4/46A
Sr/Ca change among Postclassic Lamanai females
(3 month intervals)
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
0.16
0.18
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6
Age in Years
Sr/
Ca R
ati
o
N10-2/4 N10-4/1 N10-4/2B N10-4/10 N10-4/11 N10-4/19 N10-4/33 N10-4/40 N10-4/45 N10-4/46b N10-4/46c
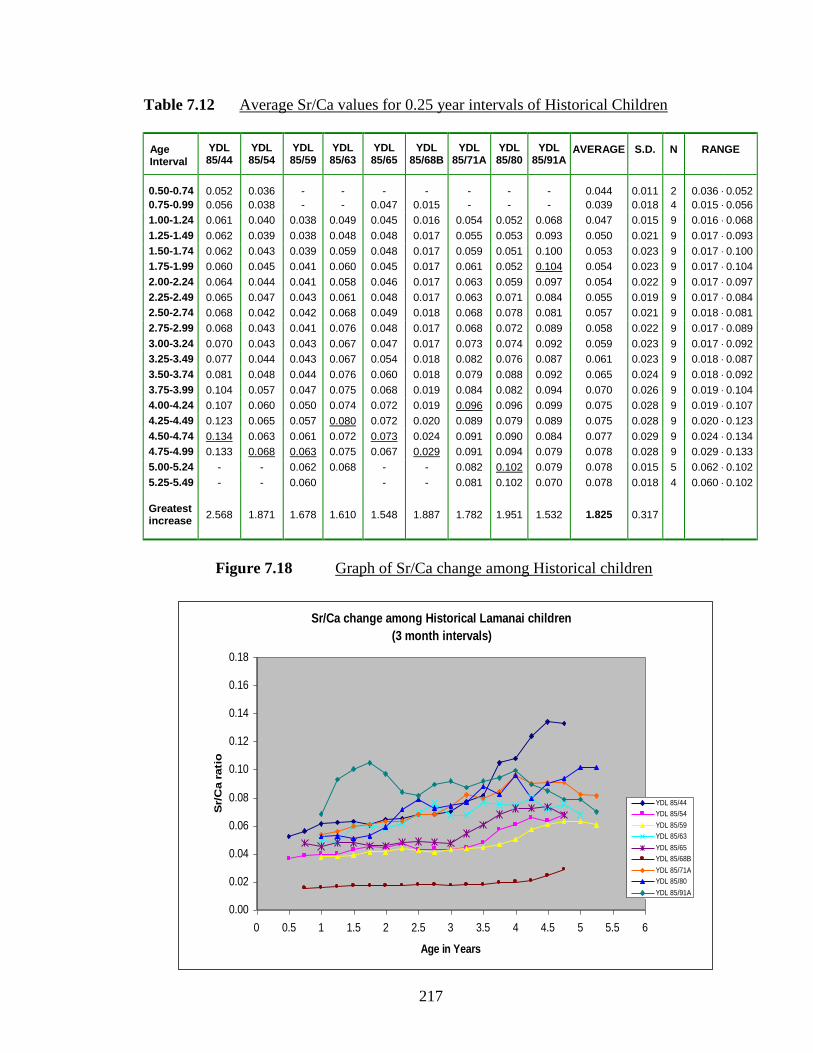
217
Table 7.12 Average Sr/Ca values for 0.25 year intervals of Historical Children
Age Interval
YDL 85/44
YDL 85/54
YDL 85/59
YDL 85/63
YDL 85/65
YDL 85/68B
YDL 85/71A
YDL 85/80
YDL 85/91A
AVERAGE
S.D.
N
RANGE
0.50-0.74 0.052 0.036 - - - - - - - 0.044 0.011 2 0.036 - 0.052
0.75-0.99 0.056 0.038 - - 0.047 0.015 - - - 0.039 0.018 4 0.015 - 0.056
1.00-1.24 0.061 0.040 0.038 0.049 0.045 0.016 0.054 0.052 0.068 0.047 0.015 9 0.016 - 0.068
1.25-1.49 0.062 0.039 0.038 0.048 0.048 0.017 0.055 0.053 0.093 0.050 0.021 9 0.017 - 0.093
1.50-1.74 0.062 0.043 0.039 0.059 0.048 0.017 0.059 0.051 0.100 0.053 0.023 9 0.017 - 0.100
1.75-1.99 0.060 0.045 0.041 0.060 0.045 0.017 0.061 0.052 0.104 0.054 0.023 9 0.017 - 0.104
2.00-2.24 0.064 0.044 0.041 0.058 0.046 0.017 0.063 0.059 0.097 0.054 0.022 9 0.017 - 0.097
2.25-2.49 0.065 0.047 0.043 0.061 0.048 0.017 0.063 0.071 0.084 0.055 0.019 9 0.017 - 0.084
2.50-2.74 0.068 0.042 0.042 0.068 0.049 0.018 0.068 0.078 0.081 0.057 0.021 9 0.018 - 0.081
2.75-2.99 0.068 0.043 0.041 0.076 0.048 0.017 0.068 0.072 0.089 0.058 0.022 9 0.017 - 0.089
3.00-3.24 0.070 0.043 0.043 0.067 0.047 0.017 0.073 0.074 0.092 0.059 0.023 9 0.017 - 0.092
3.25-3.49 0.077 0.044 0.043 0.067 0.054 0.018 0.082 0.076 0.087 0.061 0.023 9 0.018 - 0.087
3.50-3.74 0.081 0.048 0.044 0.076 0.060 0.018 0.079 0.088 0.092 0.065 0.024 9 0.018 - 0.092
3.75-3.99 0.104 0.057 0.047 0.075 0.068 0.019 0.084 0.082 0.094 0.070 0.026 9 0.019 - 0.104
4.00-4.24 0.107 0.060 0.050 0.074 0.072 0.019 0.096 0.096 0.099 0.075 0.028 9 0.019 - 0.107
4.25-4.49 0.123 0.065 0.057 0.080 0.072 0.020 0.089 0.079 0.089 0.075 0.028 9 0.020 - 0.123
4.50-4.74 0.134 0.063 0.061 0.072 0.073 0.024 0.091 0.090 0.084 0.077 0.029 9 0.024 - 0.134
4.75-4.99 0.133 0.068 0.063 0.075 0.067 0.029 0.091 0.094 0.079 0.078 0.028 9 0.029 - 0.133
5.00-5.24 - - 0.062 0.068 - - 0.082 0.102 0.079 0.078 0.015 5 0.062 - 0.102
5.25-5.49 - - 0.060 - - 0.081 0.102 0.070 0.078 0.018 4 0.060 - 0.102
Greatest increase
2.568
1.871
1.678
1.610
1.548
1.887
1.782
1.951
1.532
1.825
0.317
Figure 7.18 Graph of Sr/Ca change among Historical children
Sr/Ca change among Historical Lamanai children
(3 month intervals)
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
0.16
0.18
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6
Age in Years
Sr/
Ca r
ati
o
YDL 85/44
YDL 85/54
YDL 85/59
YDL 85/63
YDL 85/65
YDL 85/68B
YDL 85/71A
YDL 85/80
YDL 85/91A

218
Table 7.13 Average Sr/Ca values for 0.25 year intervals of Historical Females
Age Interval
YDL 85-17
YDL 85-21
YDL 85-23
YDL 85-27
YDL 85-31
YDL 85-41
YDL 85-46
YDL 85-50B
YDL 85-53
YDL 85-62
YDL 85-78C
YDL 85-101
YDL 85-102
AVERAGE
S.D.
N
RANGE
0.50-0.74 0.050 0.018 - - - - 0.045 0.018 - - - - - 0.033 0.017 4 0.018 - 0.050
0.75-0.99 0.054 0.020 - - - - 0.046 0.014 - 0.039 - 0.033 - 0.034 0.016 6 0.014 - 0.054
1.00-1.24 0.065 0.020 - - - 0.043 0.048 0.015 0.075 0.045 - 0.031 - 0.043 0.021 8 0.015 - 0.075
1.25-1.49 0.056 0.022 0.015 0.072 0.076 0.043 0.050 0.016 0.081 0.048 0.034 0.029 0.043 0.045 0.022 13 0.015 - 0.081
1.50-1.74 0.065 0.022 0.016 0.062 0.074 0.044 0.051 0.016 0.079 0.047 0.030 0.031 0.045 0.045 0.021 13 0.016 - 0.079
1.75-1.99 0.060 0.024 0.018 0.070 0.074 0.046 0.045 0.016 0.093 0.046 0.029 0.035 0.050 0.047 0.023 13 0.016 - 0.093
2.00-2.24 0.071 0.025 0.018 0.078 0.078 0.047 0.047 0.018 0.094 0.048 0.031 0.036 0.051 0.049 0.025 13 0.018 - 0.094
2.25-2.49 0.073 0.027 0.016 0.075 0.081 0.045 0.050 0.018 0.085 0.052 0.030 0.037 0.055 0.050 0.024 13 0.016 - 0.085
2.50-2.74 0.081 0.036 0.016 0.076 0.085 0.046 0.052 0.017 0.076 0.060 0.031 0.039 0.059 0.052 0.024 13 0.016 - 0.085
2.75-2.99 0.093 0.035 0.017 0.074 0.093 0.049 0.056 0.019 0.071 0.057 0.032 0.044 0.061 0.054 0.025 13 0.017 - 0.093
3.00-3.24 0.099 0.035 0.018 0.085 0.110 0.053 0.057 0.019 0.069 0.048 0.035 0.048 0.074 0.058 0.029 13 0.018 - 0.110
3.25-3.49 0.107 0.035 0.019 0.082 0.113 0.053 0.056 0.019 0.049 0.039 0.035 0.057 0.111 0.060 0.033 13 0.019 - 0.113
3.50-3.74 0.117 0.035 0.022 0.076 0.111 0.055 0.062 0.020 0.044 0.034 0.035 0.055 0.115 0.060 0.035 13 0.020 - 0.117
3.75-3.99 0.126 0.038 0.032 0.078 0.122 0.060 0.059 0.020 0.046 0.035 0.040 0.056 0.113 0.063 0.036 13 0.020 - 0.126
4.00-4.24 0.117 0.043 0.051 0.067 0.119 0.058 0.060 0.021 0.051 0.038 0.034 0.062 0.123 0.065 0.034 13 0.021 - 0.123
4.25-4.49 0.116 0.047 0.058 0.069 0.120 0.056 0.057 0.021 0.054 0.033 0.034 0.067 0.087 0.063 0.030 13 0.021 - 0.120
4.50-4.74 0.132 0.056 0.060 0.070 0.131 0.052 0.063 0.022 0.049 0.035 0.037 0.067 0.058 0.064 0.033 13 0.022 - 0.132
4.75-4.99 0.137 0.055 0.055 0.069 0.129 - 0.073 0.022 0.047 - 0.042 - 0.056 0.069 0.037 10 0.022 - 0.137
5.00-5.24 - - 0.065 0.071 0.167 - - - 0.050 - 0.036 - 0.059 0.075 0.047 6 0.036 - 0.167
5.25-5.49 - - 0.059 0.073 0.146 - - - 0.052 - 0.034 - - 0.073 0.043 5 0.034 - 0.146
Greatest increase
2.758
3.065
4.447
1.179
2.188
1.383
1.604
1.247
1.260
1.514
1.224
2.007
2.849
2.056
0.982

219
Table 7.14 Average Sr/Ca values for 0.25 year intervals of Historical Males
Age Interval
YDL I-68
YDL 85/32
YDL 85/33
YDL 85/35
YDL 85/47
YDL 85/73
YDL 85/78A
YDL 85/86
YDL 85/87B
YDL 85/97
YDL 85/98
YDL 85/98
AVERAGE
S.D.
N
RANGE
0.50-0.74 - - - - - - - - - - - - - - 0 - -
0.75-0.99 0.063 0.035 - - - - - 0.078 0.033 0.084 - - 0.059 0.024 5 0.033 - 0.084
1.00-1.24 0.062 0.037 0.026 0.026 0.049 0.046 0.051 0.079 0.033 0.080 0.058 0.042 0.049 0.018 12 0.026 - 0.080
1.25-1.49 0.071 0.038 0.032 0.026 0.055 0.046 0.051 0.082 0.033 0.088 0.059 0.046 0.052 0.020 12 0.026 - 0.088
1.50-1.74 0.070 0.041 0.031 0.027 0.058 0.048 0.055 0.083 0.032 0.088 0.060 0.048 0.053 0.020 12 0.027 - 0.088
1.75-1.99 0.072 0.042 0.031 0.027 0.058 0.054 0.059 0.085 0.035 0.089 0.060 0.048 0.055 0.020 12 0.027 - 0.089
2.00-2.24 0.074 0.043 0.029 0.028 0.059 0.051 0.057 0.081 0.038 0.082 0.065 0.048 0.055 0.019 12 0.028 - 0.082
2.25-2.49 0.073 0.043 0.032 0.031 0.059 0.057 0.064 0.089 0.047 0.089 0.062 0.052 0.058 0.019 12 0.031 - 0.089
2.50-2.74 0.077 0.042 0.031 0.034 0.053 0.051 0.068 0.090 0.045 0.093 0.062 0.062 0.059 0.020 12 0.031 - 0.093
2.75-2.99 0.083 0.045 0.032 0.036 0.053 0.054 0.073 0.095 0.038 0.094 0.065 0.066 0.061 0.022 12 0.032 - 0.095
3.00-3.24 0.090 0.050 0.032 0.043 0.053 0.057 0.073 0.094 0.037 0.099 0.061 0.074 0.063 0.022 12 0.032 - 0.099
3.25-3.49 0.089 0.052 0.040 0.042 0.049 0.051 0.075 0.107 0.034 0.094 0.054 0.079 0.064 0.024 12 0.034 - 0.107
3.50-3.74 0.094 0.056 0.046 0.044 0.052 0.049 0.084 0.103 0.038 0.098 0.053 0.077 0.066 0.023 12 0.038 - 0.103
3.75-3.99 0.098 0.054 0.049 0.041 0.051 0.049 0.084 0.104 0.050 0.086 0.054 0.064 0.065 0.022 12 0.041 - 0.104
4.00-4.24 0.099 0.056 0.053 0.041 0.059 0.048 0.093 0.082 0.065 0.078 0.056 0.067 0.066 0.018 12 0.041 - 0.099
4.25-4.49 0.097 0.059 0.052 0.046 0.052 0.053 0.094 0.077 0.066 0.084 0.052 0.074 0.067 0.018 12 0.046 - 0.097
4.50-4.74 0.104 0.058 0.060 0.045 0.057 0.040 0.097 0.090 0.064 0.094 0.053 0.079 0.070 0.022 12 0.040 - 0.104
4.75-4.99 0.112 - 0.058 0.048 0.055 0.036 0.103 - 0.055 - 0.046 - 0.064 0.028 8 0.036 - 0.112
5.00-5.24 - - 0.053 0.049 0.055 0.032 0.127 - - - - - 0.063 0.037 5 0.032 - 0.127
5.25-5.49 - - - 0.050 - 0.035 0.143 - - - - - 0.076 0.058 3 0.035 - 0.143
Greatest increase
1.762
1.689
2.316
1.965
1.209
1.241
2.828
1.363
2.004
1.173
1.126
1.872
1.712
0.524

220
Figure 7.19 Graph of Sr/Ca change among Historical females
Figure 7.20 Graph of Sr/Ca change among Historical males
Sr/Ca change among Historical Lamanai females
(3 month intervals)
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
0.16
0.18
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6
Age in Years
Sr/
Ca R
ati
o YDL 85-17
YDL 85-21
YDL 85-23
YDL 85-27
YDL 85-31
YDL 85-41
YDL 85-46
YDL 85-50B
YDL 85-53
YDL 85-62
YDL 85-78C
YDL 85-101
YDL 85-102
Sr/Ca change among Historical Lamanai males
(3 month intervals)
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
0.16
0.18
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6
Age in Years
Sr/
Ca R
ati
o
YDL I-68
YDL 85/32
YDL 85/33
YDL 85/35
YDL 85/47
YDL 85/73
YDL 85/78A
YDL 85/86
YDL 85/87B
YDL 85/97
YDL 85/98
YDL 85/98
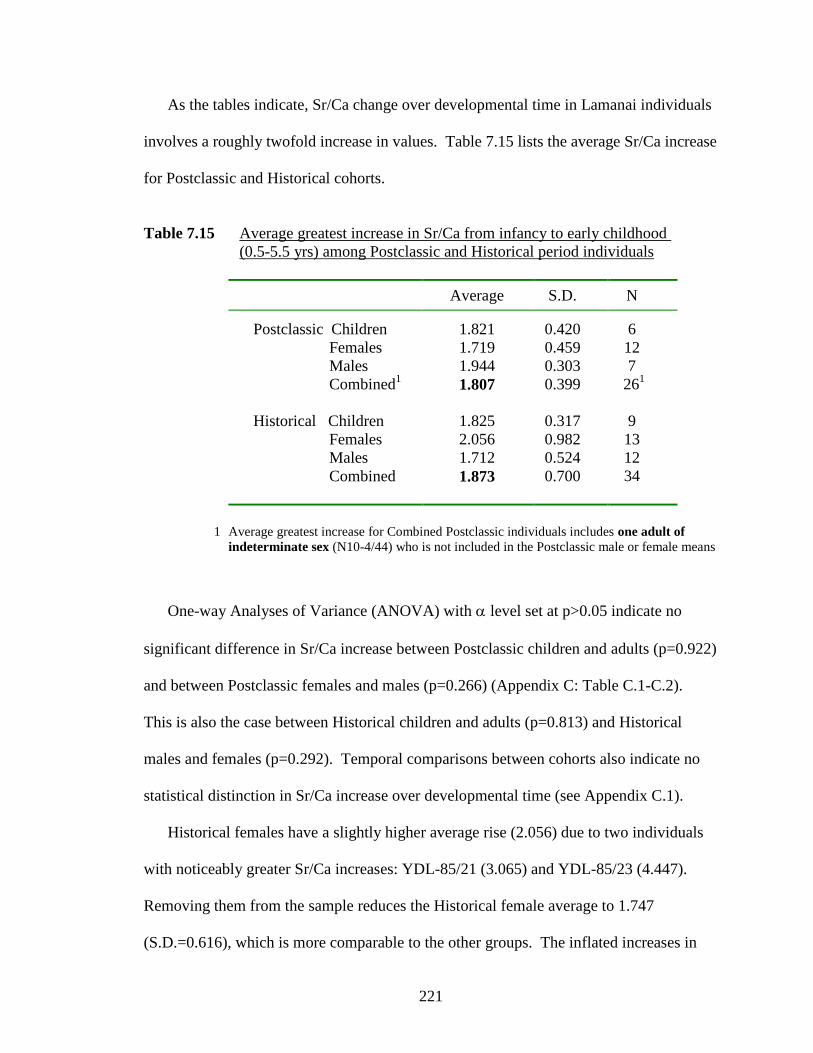
221
As the tables indicate, Sr/Ca change over developmental time in Lamanai individuals
involves a roughly twofold increase in values. Table 7.15 lists the average Sr/Ca increase
for Postclassic and Historical cohorts.
Table 7.15 Average greatest increase in Sr/Ca from infancy to early childhood
(0.5-5.5 yrs) among Postclassic and Historical period individuals
Average S.D. N
Postclassic Children
1.821
0.420
6
Females 1.719 0.459 12
Males 1.944 0.303 7
Combined1
1.807 0.399 261
Historical Children 1.825 0.317 9
Females 2.056 0.982 13
Males 1.712 0.524 12
Combined 1.873 0.700 34
1 Average greatest increase for Combined Postclassic individuals includes one adult of
indeterminate sex (N10-4/44) who is not included in the Postclassic male or female means
One-way Analyses of Variance (ANOVA) with level set at p>0.05 indicate no
significant difference in Sr/Ca increase between Postclassic children and adults (p=0.922)
and between Postclassic females and males (p=0.266) (Appendix C: Table C.1-C.2).
This is also the case between Historical children and adults (p=0.813) and Historical
males and females (p=0.292). Temporal comparisons between cohorts also indicate no
statistical distinction in Sr/Ca increase over developmental time (see Appendix C.1).
Historical females have a slightly higher average rise (2.056) due to two individuals
with noticeably greater Sr/Ca increases: YDL-85/21 (3.065) and YDL-85/23 (4.447).
Removing them from the sample reduces the Historical female average to 1.747
(S.D.=0.616), which is more comparable to the other groups. The inflated increases in

222
these two women may reflect one of several circumstances: individual variation
(reflecting true dietary increase); diagenetically altered enamel, or instrumentation
variables.
The data in Tables 7.9-7.14 also contain important information regarding the ages of
peak Sr/Ca per individual (underlined values). In this case, one can infer that the highest
Sr/Ca values represent the point when maximum solid food intake is attained. Figure
7.21 is a histogram detailing the frequency of peak Sr/Ca values according to 0.25 year
age intervals. In individuals with the same maximum values at different age intervals, the
earliest period is considered to be the age of attaining peak Sr/Ca.
Figure 7.21 Distribution of peak Sr/Ca values per age interval
It is evident from the bar graph that the majority of Lamanai children did not achieve
maximum enamel Sr/Ca ratios until after the age of four years, with many in the range of
4.25 to 5.25 years. Prior to four years of age, 23.1% (6 of 26) of Postclassic individuals
Comparison of Peak Sr/Ca Age
0
1
2
3
4
5
6
7
8
1.75 2 2.25 2.5 2.75 3 3.25 3.5 3.75 4 4.25 4.5 4.75 5 5.25
Age Interval
Nu
mb
er
of
Ind
ivid
uals
Postclassic
Historical
15.4% Postclassic
29.4% Historical

223
and 38.2% (13 of 34) of Historical individuals exhibit peak Sr/Ca values, while after four
years of age, 77.9% (20 of 26) of Postclassic and 61.8% (21 of 34) of Historical
individuals display peak enamel Sr/Ca values.
For the most part, Postclassic and Historical children resemble each other in the
distribution of peak Sr/Ca, but the Historical sample can also be distinguished by more
children attaining maximum Sr/Ca at earlier ages (1.75-3.5 years). Unlike the Postclassic
sample, the Historical cohort assumes a slightly bimodal distribution in the timing of
peak Sr/Ca values (Figure 7.21). If one assumes that peak values represent the age of
maximum solid food intake, then it appears that more Historical children were weaned at
earlier ages than Postclassic individuals. Between 1.75 and 3.5 years of age, Historical
individuals are distinguished for having almost twice the incidence of peak Sr/Ca values:
29.4% of the Historical sample (10 of 34 individuals), while only 15.4% of Postclassic
individuals (4 of 26) exhibit peak values at these earlier ages.
The peak Sr/Ca distribution reflects a Postclassic dietary pattern that is much more
uniform compared to the Historical period. In the Postclassic, maximum Sr/Ca values
infer that most children were completely weaned by 4.25 to 5.25 years of age. Relatively
stable socio-political-economic conditions, and greater socioeconomic status of the entire
Postclassic sample, may account for a weaning pattern that is more conventional in terms
of continued increase in food supplementation over time.
While many Historical children were also weaned at such late ages, the Historical
sample is also noteworthy for many individuals with peak Sr/Ca ratios between 1.75-3.5
years of age. This earlier weaning age might be expected in the Historical period due to
socio-economic, political and biological upheaval, as well as the presumed demands on
adult women for tribute products.

224
Remember though, that while rising Sr/Ca values can reflect the introduction of solid
foods, weaning age can only roughly be inferred by (maximum) hard tissue Sr/Ca values.
Importantly, this presumption is complicated by two opposing realities: 1) weaning may
have occurred earlier than the age of peak Sr/Ca, but Sr/Ca continued to rise due to
inefficient Sr discrimination (thus, peak Sr/Ca may only reflect the age of efficient Sr
discrimination by mature digestive systems); 2) increased discrimination of Sr by the gut
and reduced Sr/Ca values thereafter does not rule out the continued intake of some breast
milk after attaining peak Sr/Ca (see below).
7.5 The Nature of Enamel Sr/Ca Change in Lamanai Individuals
Plotting of Sr/Ca values in Lamanai canines produces graphs generally indicating
positive unidirectional change (Figures 7.22-7.27). Initial enamel development (0.5-1.0
year) is characterized by Sr/Ca values mostly in the range of 0.02-0.08, while the
majority of values at the end of canine crown formation (5-5.5 years) range from 0.04-
0.12. (These ranges reflect the tremendous variation in Sr found in hard tissues [see
Section 4.4]). While mostly increasing over time, Sr/Ca change in Lamanai canines can
be grouped into four patterns: 1) general increase; 2) fluctuating changes; 3) delayed
increase (i.e., period of stability followed by a rise); and 4) general stability.
Figures 7.22-7.27 and 7.29 illustrate such patterns in Postclassic and Historical
Lamanai. Of note, Sr/Ca values plotted at each 0.25 year age (x-axis) reflect average
values for the subsequent interval (e.g., Sr/Ca value at 0.5 years = average for the period
0.50-0.749 years; Sr/Ca value at 5.25 years = average for the period 5.25-5.49 years).
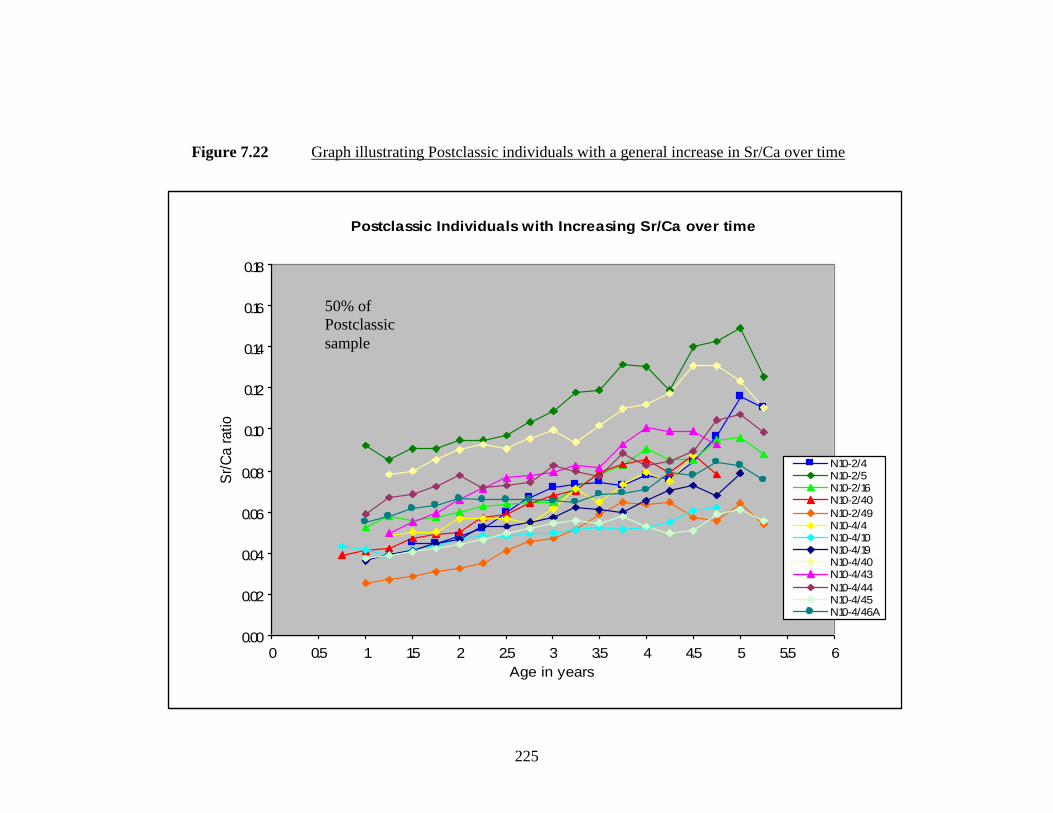
225
Figure 7.22 Graph illustrating Postclassic individuals with a general increase in Sr/Ca over time
Postclassic Individuals with Increasing Sr/Ca over time
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
0.16
0.18
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6
Age in years
Sr/
Ca r
atio
N10-2/4 N10-2/5 N10-2/16 N10-2/40
N10-2/49 N10-4/4N10-4/10 N10-4/19 N10-4/40 N10-4/43
N10-4/44 N10-4/45 N10-4/46A
50% of
Postclassic
sample

226
Figure 7.23 Graph illustrating Historical individuals with a general increase in Sr/Ca over time
Historical Individuals with Increasing Sr/Ca over time
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
0.16
0.18
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6
Age in years
Sr/
Ca r
atio
YDL I-68
YDL 85-17
YDL 85-21
YDL 85-31
YDL 85/32
YDL 85-46 YDL 85/63
YDL 85/71A
YDL 85/78A
YDL 85/80
YDL 85-101
32% of
Historical
sample

227
Generally Increasing Sr/Ca
As postulated in this study, enamel Sr/Ca significantly increases during the period of
permanent canine enamel formation. Figures 7.22 and 7.23 clearly illustrate an
approximately twofold increase between the ages of 0.5 and 5.5 years in 13 Postclassic
(50% of sample) and 11 Historical individuals (32.35% of sample). The degree of overall
Sr/Ca increase is not statistically significant between all Postclassic and Historical
individuals (p= 0.668).
Though not consistently, many individuals do display the pattern of Sr/Ca change
predicted in Chapter Five (Figure 5.1), albeit without the initial period of very low,
stable, Sr/Ca when young infants are exclusively breastfeeding (since enamel from
between 0-0.5 years of age is not available) (see Figures 7.22, 7.23, 7.26, 7.27). Since it
is likely that infants were supplemented with food as early as 6 months of age, due to
inadequate breast milk nutrients, it is not surprising to find Sr/Ca values rising from the
start of canine enamel sampled in this study (0.5-0.75 years).
In most of these individuals, it appears that weanling foods were consumed in a
steady, gradual, manner after their introduction. (Some individuals exhibit Sr/Ca values
that fluctuate slightly, but their overall trend is to increase over time.) After 2-2.5 years
of age, however, the rate of solid food supplementation seems to escalate, most likely to
accommodate increased growth and development (see Figures 7.22-23). Combined with
the declining nutritional quality of breast milk, it is likely that after two years of age,
most Lamanai children were consuming solid foods for the bulk of their energy and
protein requirements, with breast milk comprising a minor nutritional component.

228
As illustrated in Figure 7.22, Sr/Ca values of Postclassic individuals reach their peak
and stabilize between the ages of 3.75-5, or predominantly between 4.5-5 years of age.
After nine months to one year of age, or the period of exclusive/predominant breast milk
intake, the time it takes to reach peak Sr/Ca values ranges from 2.75 (N10-2/49) to 4 years
(N10-2/5) (see Figure 7.22). Notably, Individual N10-4/46A, who is identified as possibly
the last pre-Hispanic ruler of Lamanai (Pendergast 1991b), exhibits an enamel Sr/Ca
pattern that resembles the model of gradual increase in food supplementation over time,
peaking at 4.75 years of age (see Figure 7.22). Assuming that erratic enamel Sr/Ca change
over time reflects extended periods of poor health or nutrition (see below), his strontium
pattern indicates that greater social status might have shielded him from such stressors.
Importantly, it suggests that even children of Maya elites nursed extensively in ancient
times.
Among Historical individuals, Sr/Ca reaches its peak at 3.5-5.25 years of age, or a
period ranging from 3 (YDL-85/71A) to 4.25 years (YDL-85/17) (see Figure 7.23).
Overall, Sr/Ca reaches its peak between 4.5-5 years in most Lamanai individuals. In each
graph, a subsequent drop in Sr/Ca reflects the more efficient discrimination of Sr by
mature digestive systems (see Chapter Four).

229
Fluctuating Sr/Ca
Individuals with fluctuating Sr/Ca over time demonstrate the variable nature of Sr/Ca
in terms of environmental availability, strontium discrimination by the gut, hard tissue
incorporation and solid food intake. Eight of 26 Postclassic (30.77%) and 13 of 34
Historical (38.24%) individuals display variable changes in Sr/Ca between the age of 0.5
and 5.5 years. As noted above, additional individuals listed as having “generally
increasing” Sr/Ca over time can also be described as “fluctuating” in Sr/Ca.
The fluctuations likely reflect true variations in Sr/Ca associated with disruptions or
variations in weanling diet, in addition to changing Sr discrimination and hard tissue
incorporation.
On one hand, shifts in the direction of Sr/Ca over time can be interpreted to reflect
alternating periods of high and low solid food intake, with levels of breastfeeding
dependent on such consumption. To effect such compositional changes, these episodes
are assumed to be several weeks or months in duration, according to the period of Sr/Ca
change evident across enamel (and developmental age). Varying the relative proportion
of breast milk and solid food consumption (as opposed to a positive trajectory of
increased solid food intake) reflects the tenuous nature of the transition from nursing to
solid food dependence. As stated elsewhere, this can be determined by child preference,
illness (maternal and infant), food availability, and the socioeconomic obligations of the
mother.

230
Figure 7.24 Graph illustrating Postclassic individuals with fluctuations in Sr/Ca over time
Postclassic Individuals with Fluctuating Change in Sr/Ca
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
0.16
0.18
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6
Age in years
Sr/
Ca r
atio
N10-2/42
N10-2/44
N10-4/1
N10-4/2A
N10-4/2B
N10-4/33
N10-4/46b
N10-4/46c
31% of
Postclassic
sample

231
Figure 7.25 Graph illustrating Historical individuals with fluctuations in Sr/Ca over time
Historical Individuals with Fluctuating Sr/Ca over time
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
0.16
0.18
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6
Age in years
Sr/
Ca r
atio YDL 85-27
YDL 85/47
YDL 85-53
YDL 85-62
YDL 85/73
YDL 85-78C
YDL 85/86
YDL 85/87B
YDL 85/91A
YDL 85/97
YDL 85/98
YDL 85-102
YDL 85/103
38% of
Historical
sample

232
Alternatively, changing Sr/Ca ratios may reflect variations in the supplementary diet
prior to weaning. Changes in infant solid foods (of differing Sr/Ca abundance) may be
due to seasonal availability of resources, crop failure and other ecological factors, as well
as withholding of taboo foods, or treatment with therapeutic foods/herbs, during illness.
While not recognized ethnographically, possible alternative weanling foods include
mashed immature (not dry) maize (high in Sr/Ca), meat (low in Sr/Ca), and various raw
fruit, nuts or vegetables (high in Sr/Ca).
Beyond dietary availability, a significant factor affecting food intake and the relative
levels of Sr/Ca in these individuals is childhood disease. Gastrointestinal conditions,
which were particularly common in Historical times (White 1986; White et al. 1994),
would have particularly affected an infant’s ability and desire to eat solid foods. Cultural
taboos about diet (e.g., food “contamination”) during illness may also determine feeding
patterns at such times (see Stuart-Macadam and Dettwyler 1995).
Related to this, obviously, is the role of maternal health in infant nutrition. The
ability of mothers to breastfeed and/or properly supplement infants can be severely
compromised by their ill health and/or malnutrition. For both mothers and infants,
periods of illness and poor nutrition that could impact enamel Sr/Ca probably lasted
longer than several weeks, i.e., months. Historical period women and their children
would have been particularly affected by the poor epidemiological conditions of
colonialism. The slightly elevated rate of Historical individuals with fluctuating Sr/Ca
patterns (38.2% compared to 30.8% of Postclassic individuals) is thought to reflect this.
Thus, examining enamel Sr/Ca patterns can provide potential indicators of individual
illness history.

233
In the Lamanai population, among Postclassic adults with fluctuating Sr/Ca, it is
mostly females who display irregular patterns compared to males, who generally exhibit
the expected pattern of increasing Sr/Ca over time (see Figures 7.16 and 7.17). This
suggests that female children may have been more prone to dietary inconsistency than
boys, i.e., shifts in breast milk intake and food supplementation during times of economic
uncertainty or childhood illness. Dietary stability and nutritional advantages among
Lamanai males is also suggested above in comparing the averaged Sr/Ca values for
Postclassic males and females (see Section 7.3).
During the Postclassic period, when Maya elites maintained socioeconomic
dominance over the Lamanai population (Pendergast 1986, 1991a, 1992), gender
differences were more pronounced than in Spanish colonial times, when all Maya were
reduced to a shared, subordinate, position. The Postclassic sample examined in this
analysis is advantaged socio-economically, as a whole, but gender distinctions prevail. It
is, therefore, not surprising that Postclassic males and females display distinctions that
are manifest in hard tissue composition (reflecting varied health and nutrition).
On the other hand, in the Historical period, fluctuating Sr/Ca patterns appear in
comparable frequency among both males and females (see Figures 7.19 and 7.20). This
“equality” in Historical times may reflect the shared predicament of “reduced” Maya
interred in the Lamanai cemetery, and it is also demonstrated in the total mean Sr/Ca
values for Historical males and females (see Section 7.3).
Fluctuating Sr/Ca patterns may also reflect the varied origins of Maya refugees from
the Yucatan and elsewhere. As children, they may have experienced famine, infectious
disease, maternal death and dietary instability related to the demographic upheaval of the

234
colonial period (see Chapter Two).
Among “fluctuating” cases, several Postclassic and Historical individuals show peak
Sr/Ca values comparable to the late ages among “generally increasing” individuals. Four
Postclassic individuals and three Historical individuals have peak values at 4-4.75 years
of age. Many individuals in this category also exhibit patterns indicating that they may
have completed weaning much earlier. These include individuals with peak Sr/Ca values
in the mid-range of the graphs, i.e. 1.75-3.5 years of age (Figures 7.24-25). Three
Postclassic individuals and ten Historical individuals reach maximum Sr/Ca values in this
age range (also see Figure 7.21 for a peak Sr/Ca distribution of the entire sample).
Fluctuations, and in some cases, progressive decline in Sr/Ca after peak values may
reflect developmental variations in Sr discrimination (i.e., increased efficiency), hard
tissue incorporation, changes in weanling diet (e.g., meat, fruits and vegetables) or,
alternatively, subsequent reductions in food intake and resumption of nursing at
intermittent periods. In this instance, peak Sr/Ca values do not necessarily represent the
age at which children were fully dependent on solid foods.
As stated earlier, the association of maximum Sr/Ca ratios with total solid food
reliance is complicated by other factors that can change relative Sr/Ca values. Due to the
immaturity of infant digestive systems, it is possible that if weaning occurred early in
infancy (i.e., first year of life), Sr/Ca might continue to rise due to inefficient
discrimination. However, clinical data appears to refute this after the first year of life,
when Sr discrimination increases, and particularly after the second year, when Sr
absorption is less than one third of the newborn level (more than three times reduced)
(Rivera and Harley 1965). Data by Rivera and Harley (1965) on the Observed Ratio, or

235
physiological discrimination, of strontium indicates that the greatest absorption of Sr
occurs between birth and one year of age, when the OR is approximately 1.00. At one
year of age, this value drops to 0.60, reflecting greater discrimination of Sr, followed by
0.24 at 2 years of age (see Sillen and Smith 1984: Table 1; and Table 7.16 below). By
approximately 9 years of age, the OR reduces to 0.18, which continues to be the average
level of Sr discrimination throughout adulthood (Rivera and Harley 1965).
In fairness, the extent of strontium change at a more micro level is not fully
understood. Even at two, three and four years of age, Rivera and Harley observed some
fluctuations in the observed ratio: 0.24 at two years, 0.25 at three years and 0.24 at four
years of age, followed by a gradual reduction thereafter. In this study, laser ablation
microsampling may be capturing a much greater level of Sr absorption fluctuation, or,
even possibly, compositional inconsistencies at the enamel prism (or prism layer) level.
Related to factors of ill health and poor nutrition, fluctuations in enamel Sr/Ca might
be associated with macroscopic developmental defects of enamel such as hypoplasia and
hypocalcification. However, with enamel hypoplasias, which are deficiencies in the
amelogenesis (enamel matrix formation) stage, periods of Sr/Ca fluctuation do not
necessarily coincide with either the position or presence of defects in teeth, though 62.5%
of hypoplastic Postclassic teeth (5 out of 8) reflect fluctuating Sr/Ca (compared to 46.2%
of hypoplastic Historical teeth [6 out of 13]). Likewise, canines with hypocalcifications,
or disruptions in enamel mineralization (which are more appropriately associated with
element data quantified by ICP-MS) are not likely to exhibit fluctuating Sr/Ca patterns.
In this case, 1 out of 8 (12.5%) hypocalcified Postclassic teeth and 2 out of 13 (15.4%)
Historical teeth have enamel Sr/Ca patterns that fluctuate over time.

236
In total, it appears that hard tissue incorporation of strontium is much more complex
than simply associating enamel composition with dietary intake. The findings can only
suggest that Lamanai infants commonly experienced fluctuations in: 1) the relative intake
of solid foods versus breast milk, and/or the types of weanling foods; 2) Sr discrimination
by the digestive system; or, at least, 3) hard tissue Sr incorporation.
Delayed Increase in Sr/Ca
As illustrated in Figures 7.26 and 7.27, “delayed” individuals are those with initial
Sr/Ca values that appear generally stable for some time prior to increasing. The period of
highest Sr/Ca ratio remains consistent, however, generally peaking between four and five
years of age. The “delayed” pattern closely follows the predicted behavior of strontium
postulated in Chapter Five (Figure 5.1), but in this case, one cannot assume that the initial
low values represent an extended period of exclusive breastfeeding. Insufficient breast
milk nutrients for infants after six months of age implies that the initial low Sr/Ca values
represent an extended period of combined breast milk/low solid food intake. Quantifying
the exact proportion of breast milk and solid food intake in these individuals is beyond
the scope of this study, but relative to other patterns, one could state that these individuals
were consuming considerably less solid food for much longer than other Lamanai infants.
Four Postclassic individuals (2 children, 2 adults; or 15.38% of sample) and nine
Historical individuals (5 children, 4 adults; or 26.47% of sample) are characterized by a
delayed Sr/Ca increase. Thus, almost twice as many Historical individuals exhibit a
delayed Sr/Ca pattern compared to Postclassic individuals. Beyond poor epidemiological

237
Figure 7.26 Graph illustrating Postclassic individuals with a delayed increase in Sr/Ca over time
15% of
Postclassic
sample
Postclassic Individuals with Delayed Sr/Ca Increase over time
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
0.16
0.18
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6
Age in years
Sr/
Ca r
atio
N10-2/20
N10-2/21
N10-2/22
N10-2/50
15% of
Postclassic
sample

238
Figure 7.27 Graph illustrating Historical individuals with a delayed increase in Sr/Ca over time
27% of
Historical
sample
Historical Individuals with Delayed Sr/Ca Increase over time
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
0.16
0.18
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6
Age in years
Sr/
Ca r
atio
YDL 85-23
YDL 85/33 YDL 85/35
YDL 85-41
YDL 85/44 YDL 85/54
YDL 85/59
YDL 85/65YDL 85/68B
27% of
Historical
sample

239
conditions and widespread famine in the early colonial period, this discrepancy may be
related to historical changes at Lamanai regarding women’s roles (see Chapter Eight).
Notably, sex is not a factor in the likelihood of a delayed increase. Social status also
does not exclude the likelihood of delayed Sr/Ca increase: Individual N10-2/20, an elite
male adult, clearly exhibits a delayed pattern (Figure 7.26). However, it should be noted
that as this individual, as well as N10-2/22, are “missing” ablated enamel prior to 1.0 and
1.25 years of age, it is possible that they experienced an earlier increase in Sr/Ca (food
supplementation increase) that is not recognized here and may belong in the “generally
increasing” Sr/Ca category. Strontium data for early enamel development is also absent
for several Historical individuals (Figure 7.27), and should be considered.
Importantly, if it is the case that these “delayed” individuals actually experienced an
earlier rise in Sr/Ca that is not captured in the LA-ICP-MS data, then such patterns
suggest a different model of Sr/Ca behavior in young children. In this case, Sr/Ca
undergoes two phases of positive growth (food supplementation increase), alternating
between two phases of stability: the first reflecting an extended period of consistent food
intake relative to breast milk and the second phase reflecting total reliance on solid food.
Nevertheless, while interesting to consider, the patterns illustrated above do not seem
to indicate an earlier rise in Sr/Ca that might have been excluded from the analysis unless
enamel development times are incorrect. For instance, underestimating the sampling of
buried cuspal enamel might attribute earlier development times to “older” enamel.
(Additional analysis of permanent first molars, to sample enamel from birth to six months
of age, might elucidate the neonatal Sr/Ca pattern.)

240
Based on Figures 7.26 and 7.27, more than half are individuals who died in
childhood: two Postclassic, aged 7-8 and 9 years; and five Historical, aged 6-7 years
(three children) and 8-12 years (2 children). Yet, children only comprise 25% of the total
sample in this study (6 Postclassic and 9 Historical, out of 60 individuals). It appears that
the effect of delaying substantial food supplementation in growing infants may be
increased childhood mortality.
In addition, it appears that Historical individuals are even further delayed than
Postclassic individuals for the age at which Sr/Ca starts to rise considerably. The
following histogram (Figure 7.28) demonstrates the apparent delay among Historical
individuals for the point at which Sr/Ca starts to rise after a period of general stability.
(The “time lag” is not related to dental development time associated with maxillary vs.
mandibular canine sampling. The “delayed” sample is composed of 2 maxillary and 2
mandibular Postclassic teeth and 5 maxillary and 4 mandibular Historical teeth.)
Figure 7.28 Histogram of age of significant Sr/Ca change in “delayed” individuals
Age of Significant Sr/Ca change
in "Delayed" Individuals
0
1
2
3
4
2 2.25 2.5 2.75 3 3.25 3.5 3.75 4 4.25 4.5
Age Interval
Nu
mb
er
of
Ind
ivid
uals
Postclassic
Historical

241
All “delayed” Postclassic individuals display noticeable Sr/Ca increases between 2.5-
3 years of age (2.5, 3, 3, 3), but among Historical individuals, the majority only reflect
significant Sr/Ca change after 3-3.25 years of age (2.25, 2.75, 3.25, 3.25, 3.25, 3.5, 3.5,
3.75, 4.5). One individual, YDL-85/68B is particularly late in Sr/Ca change, not rising
until 4.5 years of age, and is also included in the “stable” pattern below. It is possible
that the consistent Sr/Ca readings reflect: 1) diagenesis, which can result in homogeneous
composition; 2) poor LA-ICP-MS sensitivity (resulting in consistently low values); or 3)
stability after an early rise in enamel Sr/Ca that is not captured by LA-ICP-MS (i.e., from
enamel shortly after birth), which could be attributed to infants who did not breastfeed
(due to maternal illness or death), and who were weaned with maize gruel as neonates.
Once strontium starts to rise, the period required to reach peak Sr/Ca values is
truncated compared to individuals with increasing Sr/Ca from the start of laser sampling
(Figures 7.22-7.23). In the Postclassic sample, this period of rapid food supplementation
and complete weaning ranges from 0.5 years (N10-2/21) to 2.25 years (N10-2/20).
“Delayed” Historical children are completely weaned in 1.0 (most individuals) to 1.75
years (YDL 85/23) after food supplementation increases significantly. (Remember, that
the extended period of low Sr/Ca at the start of each individual probably reflects some
solid food supplementation.) These periods are noticeably shorter than those observed in
individuals with Sr/Ca values increasing from the start of ablation sampling: 2.75 to 4
years among Postclassic individuals and 3 to 4.25 years among Historical individuals (see
above). Obviously, infants who are delayed in substantial solid food supplementation
(i.e., consuming more foods than breast milk) are subject to a faster dietary transition to
solid foods.

242
Stable Sr/Ca
The individuals represented in the “stable” category are distinguished for having low
Sr/Ca ratios throughout the entire ablation period (Figure 7.29). There are some
noticeable fluctuations and slight increases over time, but overall, these individuals
exhibit relatively minimal total change. One Postclassic individual (N10-4/46B) in the
“fluctuating” category (Figure 7.24) also exhibits very low Sr increase (1.084), but is not
included here as enamel could not be sampled from after 3.75 years of age, when further
changes may have occurred.
Similarly, N10-4/11 (Figure 7.29) exhibits some fluctuations, but they reflect only
slight shifts in relative solid food and breast milk intake, with a total increase of just
1.115. It is possible, however, that this female individual experienced a more significant
increase (e.g., 1.5-2.0) in Sr/Ca (food supplementation) earlier in infancy, as enamel prior
to 0.75 years could not be sampled.
Figure 7.29 Graph illustrating Lamanai individuals with stable Sr/Ca over time
Lamanai Individuals with Stable Sr/Ca over time
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
0.16
0.18
0 0.5 1 1.5 2 2.5 3 3.5 4 4.5 5 5.5 6
Age in years
Sr/
Ca r
ati
o
N10-4/11 YDL 85/50B YDL 85/68B
4% of Postclassic
3% of Historical

243
The two Historical individuals (YDL-85/50B and YDL-85/68B) are noteworthy for
having the consistently lowest Sr/Ca ratios in the entire study sample. In YDL-85/50B,
this is the result of very low Sr readings throughout the entire enamel, rather than high
Ca. This is also the case for the first ablation of YDL-85/68B (approximately 2/3 of the
total crown length), but the remaining ablation of YDL-85/68B (cervical 1/3) exhibits a
low Sr/Ca ratio due to significantly higher Ca values, rather than lower Sr values.
(N.B. Individual YDL-85/68B is also included in the “delayed increase” category since a
late rise in Sr/Ca can be observed after 4.25 years of age [see Figure 7.27].)
It is possible that these three teeth were altered diagenetically, particularly the
cervical third of YDL-85/68B, which is thinner in enamel thickness and may have
undergone calcium enrichment. Nevertheless, White (1986) has previously determined
that diagenesis is not a significant factor for Lamanai skeletons in terms of geological
conditions and burial contexts, particularly regarding calcium: if anything, Lamanai
bones reflect some leaching of calcium.
It is also possible that ICP-MS sensitivity was sub-optimal during laser sampling of
these teeth, resulting in reduced strontium quantification. However, ablation conditions
do not distinguish these teeth from others sampled during the same periods, and other
teeth sampled before and after these teeth do not exhibit such distinctions in Sr and Ca.
Considering the extent of strontium variation in human populations, even within the
same geological locale, the low values of enamel Sr/Ca in these individuals may simply
reflect such diversity. Importantly, it is highly possible that earlier-forming enamel not
sampled by LA-ICP-MS in these individuals, i.e., birth to six months, exhibits an even
more reduced Sr/Ca pattern associated with exclusive breastfeeding. In this case, one can

244
propose that such infants’ mothers may have perished, resulting in their abrupt weaning
with solid food (maize gruel). This sudden transition may have occurred prior to the
period sampled in their enamel (< six months), so that the “stable” patterns captured by
LA-ICP-MS may actually reflect completely weaned infants.
7.6 Determination of Dietary Sr/Ca from Enamel Sr/Ca
In Chapter Four, the Observed Ratio (OR) was outlined, which is used to quantify the
differential distribution of strontium in the food chain (Comar et al. 1957). In this study,
it can express the Sr/Ca relationship between dental enamel and diet since the
discrimination against strontium (“biopurification”) can be quantified as the inverse of
the OR (Elias et al. 1982). To reiterate, Observed Ratio (OR) = Enamel Sr/Ca
Since we have quantified the total Sr/Ca ratio of enamel in this study, the Sr/Ca
signature of the diet at each age category can be extrapolated using Rivera and Harley’s
(1965) calculated OR values for children and adults (Table 7.16).
Table 7.16 Age-related bone-diet observed ratios (ORbone-diet) (Rivera and Harley 1965)
Age in years OR bone-diet
Fetal/newborn 1.00
1 0.60
2 0.24 + 0.08
3 0.25 + 0.09
4 0.24 + 0.09
5 0.23 + 0.08
6 0.21 + 0.07
7 0.20 + 0.06
8 0.19 + 0.05
9 0.18 + 0.05
10 0.18 + 0.04
Adults 0.18
Dietary Sr/Ca

245
Declining OR values (or increasing Discrimination Factor [DF]) over time reflects the
growing discrimination of Sr in favor of Ca as children’s kidneys and digestive systems
mature after the first year of life. While newborns exhibit little or no discrimination of Sr
(OR = 1.0 / DF = 0), due to placental discrimination, OR subsequently declines to 0.6 and
0.2 of the total dietary Sr/Ca thereafter, averaging around 0.18 in older children and
adults (Rivera and Harley 1965).
In this case, Dietary Sr/Ca (age x) = Enamel Sr/Ca
ORbone-diet (age x)
Using this formula, and the age-related OR values of Rivera and Harley (1965), the
dietary Sr/Ca of Lamanai children can be determined from the total Sr/Ca ratios of
enamel at each interval. These data are presented in Table 7.17 below, and graphically
illustrated in Figure 7.30, which includes the Postclassic, Historical and Total Sample
enamel and dietary Sr/Ca values for the period between 0.5 and 5.5 years of age.
Table 7.17 Enamel vs. Dietary Sr/Ca values for Lamanai cohorts at various ages
(OR values based on Rivera and Harley, 1965)
Enamel Sr/Ca (x 10-3
)
Dietary Sr/Ca (x 10-3
)
AGE ORbone-diet Postclassic Historical Total Postclassic Historical Total
0.5-1.0 0.60 0.80 0.63 0.71 1.34 1.05 1.18
1.0-1.5 0.60 0.78 0.69 0.73 1.29 1.14 1.21
1.5-2.0 0.24 0.81 0.72 0.76 3.37 3.02 3.17
2.0-3.0 0.25 0.91 0.78 0.84 3.63 3.14 3.36
3.0-4.0 0.24 1.05 0.90 0.97 4.39 3.73 4.03
4.0-5.0 0.23 1.18 0.98 1.07 5.13 4.28 4.66
5.0-5.5 0.23 1.25 1.01 1.13 5.45 4.38 4.91

246
Figure 7.30 Comparison of enamel Sr/Ca and dietary Sr/Ca in Lamanai cohorts
The pattern of dietary Sr/Ca derived from estimated ORbone-diet values from Rivera
and Harley (1965) indicate that at around two years of age, infant diet is markedly higher
in Sr/Ca content. After the age of two years, with increased strontium discrimination (DF
of approximately 4), inferred dietary Sr/Ca is more than four times higher than the ratio
of Sr/Ca incorporated in enamel. This four to five-fold increase from birth is mirrored in
Sillen and Smith’s (1984: Figure 4) age-specific dietary Sr/Ca values for medieval Arab
bones.
Enamel Sr/Ca vs. Dietary Sr/Ca of Postclassic and Historical cohorts
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
4.5
5.0
5.5
6.0
0 0.5-1.0 1.0-1.5 1.5-2.0 2.0-3.0 3.0-4.0 4.0-5.0 5.0-5.5 Age Interval
Sr/
Ca R
atio (
x 1
0-3
)
Postclassic Enamel Sr/Ca Historical Enamel Sr/Ca Total Sample Enamel Sr/Ca Postclassic Dietary Sr/Ca Historical Dietary Sr/Ca Total Sample Dietary Sr/Ca

247
It could be speculated that this is the age at which Lamanai children were
supplemented with substantial quantities of solid food. Despite increased gastrointestinal
discrimination of strontium after two years of age (approximately four times that at birth),
continued Sr/Ca rise in canine enamel developing at that age infers a considerable solid
food contribution, or the start of the weaning process. Sillen and Smith (1984) also found
an identical pattern of dietary Sr/Ca change based on bone Sr/Ca, leading them to
conclude that their sample was weaned at two years of age.
However, it should be kept in mind that this pattern is based on Rivera and Harley’s
(1965) estimation of increased radioactive 90
Sr discrimination, or considerably reduced
ORbone-diet, after the age of two years (see Table 7.16). 90
Sr is not a naturally occurring
isotope. This strontium isotope is created by the fission of uranium and plutonium and
was primarily introduced into the atmosphere during nuclear testing in the 1950’s and
1960’s. Ancient Lamanai hard tissues will not contain biogenic levels of 90
Sr. It is
possible that this distinction from 86
Sr may change the current understanding of age-
related strontium discrimination, but there is a lack of evidence in this regard.
In truth, Figure 7.30 only clearly reflects the increased efficiency of strontium
discrimination by the digestive system over time. If enamel Sr/Ca ratios remain stable
over time, or even decline slightly, reduced OR values after 1.5-2 years of age will still
reflect substantially higher dietary Sr/Ca ratios thereafter.
Theoretically, if substantial food supplementation begins after two years of age,
dietary Sr/Ca inferred from enamel values will not reflect the same level of significant
rise as at two years of age. The commencement of significant food intake after two years
of age would not be clearly reflected in dietary Sr/Ca patterns due to the stabilization of

248
(reduced) ORbone-diet after that age. Enamel Sr/Ca values at age three and older would
have to be more than four times higher than two-year-old values to reflect a significant
surge in dietary Sr/Ca that would exceed the rise at two years of age. Conversely, if food
supplementation begins prior to two years of age, it would not be evident. A
significantly reduced OR at two years of age will always indicate a substantial increase in
dietary Sr/Ca at this time.
At this time, however, Rivera and Harley’s (1965) observations on digestive system
discrimination are widely accepted. This implies that Lamanai children were mostly
weaned, or primarily dependent on food, at around two years of age. Inferred dietary
Sr/Ca (Table 7.16) indicates that food supplementation continues to increase steadily
after this age, even beyond five years of age. In general, this pattern does not contradict
other Maya hard tissue evidence (Wright and Schwarcz 1998), ethnohistorical accounts
of Maya weaning, nor ethnographic evidence (see Chapter Five). But, it is probable that
by three to four years of age, most Lamanai infants were eating the maximum amount of
solid food. Breast milk intake may have continued, but it was likely just a minor source
of liquid, and a nutritionally insignificant supplement at that.
Newborn data is not available in this study, but it is expected that the dietary Sr/Ca
ratio and enamel Sr/Ca ratio at this age are the same value, since the ORbone-diet of fetuses
and newborns is 1.00 according to Rivera and Harley (1965). For the Lamanai sample,
this lowest value is likely to be around 0.5 x 10-3
(or lower), and would be comparable to
contemporary values from British bones (0.22 x 10-3
) (United Kingdom Medical
Research Council 1959-1970).

249
The stabilization of high strontium discrimination after two years of age suggests that
hard tissue Sr/Ca thereafter should also level out (or decrease) unless food intake (relative
Sr/Ca) continues to increase significantly. This is not reflected in the Lamanai data.
Discounting diagenesis and instrumentation variables as significant factors influencing
canine enamel Sr/Ca, one can propose that possibly the nature (e.g., rate) of enamel
mineralization after the age of two years (i.e., from approximately mid-crown to CEJ)
amplifies Sr/Ca incorporation beyond levels controlled by gastrointestinal discrimination
(also see Chapter Eight).
Summary
The data indicates that permanent canine enamel does disclose a pattern of Sr/Ca
change that likely reflects the transition from breastfeeding to consumption of weaning
foods. It is posited that hard tissue diagenesis and instrumentation variables are not
significant factors in determining the patterns of Sr/Ca observed in this investigation.
These results represent the first extensive study to quantify Sr/Ca ratios in
sequentially calcifying areas of human enamel. The mean total Sr/Ca values compare
closely with other clinical and nutritional hard tissue strontium data. Importantly, use of
LA-ICP-MS has allowed identification of a continuous pattern of element change and
variation that is not captured in other analytical techniques. The findings are particularly
rich, providing inferences for subtle shifts in diet that can be scrutinized in relation to the
entire period of enamel development and early childhood.
The ability to track detailed life history within individuals is a major outcome of this
investigation, with the recognition of substantial intra-tooth variation in permanent canine

250
enamel. It has revealed that the process leading to complete weaning (breast milk
cessation) is much more complex and variable than the assumed pattern of steady
increase in food supplementation over time. Postclassic and Historical Lamanai
individuals exhibit variations in the ages of peak Sr/Ca values (weaning), rate and age of
food supplementation and patterns of alternating breast milk and solid food consumption.
Where there is significant difference, the consistent finding is that Historical Lamanai
infants experienced an extended period of breastfeeding and delayed food
supplementation compared to their prehistoric counterparts. This factor undoubtedly
contributed to the increased morbidity and mortality of Lamanai Maya during the
colonial period.
The slightly bimodal distribution of peak enamel Sr/Ca (by age) also indicates that
more Historical children may have been weaned much earlier, i.e., younger than three
years of age. The possible reasons for this alternative weaning pattern may be related to
the changing roles of colonial women at Lamanai. In the face of tribute demands, it is
possible that some women may have traded time-intensive childcare (prolonged nursing)
for greater economic production (e.g., cloth, food), resulting in a younger weaning age
for their children. Considering the disparate socioeconomic backgrounds of Postclassic
and Historical period Maya at Lamanai, and the contrasting socio-political climates in
which they lived, the distinctions are not surprising (see Chapter Eight).
Even in cases where significance is not indicated, comparing and shifting total
averaged Sr/Ca patterns for groups of individuals has been worthwhile in constructing
models of ancient infant nutrition. The “adjustment” in total Sr/Ca patterns is not entirely
justified, since most means do not differ significantly and there is considerable variation,

251
but this approach has recognized possible distinctions in cohorts based on age, sex and
time period that correspond with archaeological and ethnographic evidence. In the case
of individuals who died in childhood, a diminished Sr/Ca pattern suggesting delayed
(and/or reduced) food supplementation, reflects a disadvantageous model of infant
nutrition and health that can be associated with shortened lifespan.
The enamel Sr/Ca data presented in this chapter reflects the wealth of life history data
that can be captured by LA-ICP-MS. Dental material provides the ideal canvas for
paleodietary reconstruction and it reflects a high level of sensitivity that can be reliably
associated with developmental age. In Chapter Eight, the possible factors responsible for
the subtle dietary changes inferred by enamel strontium are investigated.

252
CHAPTER 8
DISCUSSION
The findings in Chapter Seven attest to the general pattern of increasing strontium
over time that was postulated in this study, and reflect variations in the rate and pattern of
Sr change that have multiple implications. Based on the results, five major issues are
elaborated in this chapter:
1) Comparative data regarding strontium composition of breast milk vs. weanling
foods is examined, including the effect of alkali processing on maize and hard
tissue Sr/Ca values.
2) The inferences for ancient Maya infant nutrition are discussed and evaluated
within the framework of known weaning behavior.
3) The implications for infant nutrition are then contextualized within the
Postclassic and colonial histories of Lamanai, to better understand the factors
that affect maternal childcare practices and infant health. These include
comparisons of enamel strontium between children and adults and between
males and females.
4) To broaden the perspective, the results are compared to other hard tissue
strontium data and the role of strontium analysis in paleodietary reconstruction
is assessed.
5) Finally, analytical issues of this study are examined to formulate a set of
recommendations and considerations for future enamel research using LA-ICP-
MS technology, as well as further research that can arise from this study.

253
8.1 The Implications of Maize Processing for Dietary Reconstruction
For the ancient Maya, examining the transition from breast milk to solid food intake
and weaning means that the relative Sr/Ca signatures of human breast milk, maize gruel
and other presumed weanling foods must be understood. Universally, non-milk weanling
foods, or beikost, are easily digestible forms (gruel or porridge) of starches such as maize,
wheat or rice. Children are also often supplemented with soft fruits and vegetables, eggs,
bread or tortillas, as well as regurgitated adult foods and even ale (Fildes 1986, 1988;
Stuart-Macadam and Dettwyler 1995).
Based on ethnographic analogy and historical documents at the time of contact, it is
probable that Lamanai Maya infants consumed alkali-processed maize gruel (atole) or
other softened maize products (e.g., soaked tortillas) during the weaning process (Behar
1968; Benedict and Steggerda 1937; Izurieta and Larson-Brown 1995; Landa in Tozzer
1941). Bone isotopic evidence (13
C) verifies this and indicates that children consumed
high levels of maize that are comparable with adults throughout the Postclassic and
Historical periods (White 1986).
Like Maya adults, and probably more so, maize comprised the bulk of their diet,
depending on relative breast milk intake, with possible intake of other foods such as
fruits, vegetables (beans or squash), and possibly even regurgitated meat from parents.
White (1986) notes that bone chemical evidence does not indicate the consumption of
beans and/or meat in Lamanai weanlings, but contemporary information on Guatemalan
Ladinos includes small quantities of softened corn tortillas and black bean broth as a
weanling supplement after six months of age (Izurieta and Larson-Brown 1995). As a
major Maya food staple even in ancient times, it is likely that Lamanai infants consumed

254
similar bean products, but it is reasonably assumed that maize products represent the
majority of the weanling diet.
On the surface, the nature of Sr/Ca patterns suggests that most Lamanai Maya infants
were weaned onto maize that was not processed with lime to produce a calcium-rich
atole. This verdict is based on Burton and Wright’s (1995) observations that alkali-
treated maize becomes so enriched with calcium that the Sr/Ca signature of raw maize
becomes “invisible” under the weight of added calcium. The appearance of rising Sr/Ca
values in enamel sampled in this study contradicts this finding.
Yet, it must be remembered that Lamanai mothers are also producing extremely low
Sr/Ca breast milk as a result of consuming reduced Sr/Ca lime-processed maize
throughout their life. With mammary gland discrimination, Maya breast milk Sr/Ca will
still be reduced compared to maize foods that are processed with lime.
In a study tracking the childhood introduction of maize in enamel Sr/Ca, several
fundamental details should ideally be known. These include the relative strontium and
calcium content of: 1) the breast milk of Maya mothers eating a traditional diet; 2)
ancient Lamanai raw maize; 3) traditional weanling foods, in this case, atole (lime-treated
maize gruel); and 4) local soil and water. Based on such data, one needs to assess the
nature of two main relationships: a) the difference in Sr/Ca between human breast milk
and processed maize atole (weanling gruel) (to track the start of supplementation after
exclusive breastfeeding); and b) the effect of consuming lime-treated atole on dental
enamel composition.
Essentially, in order to observe a trophic level effect indicative of solid food (maize)
intake after exclusive breastfeeding, the Sr/Ca ratio of weanling gruel, which was likely

255
treated with lime, would have to be significantly higher than the relative Sr/Ca of breast
milk. Maize atole elemental composition is not known, but one can apply the strontium
and calcium values of other treated maize products, i.e., tortillas (Wright 1994).
Unfortunately, because breast milk, hard tissue and dietary elemental composition
will be wholly dependent on the local water and food growing conditions (soil and
water), there is limited reference data that can be accounted for here. Firstly, there is no
comparable data for the Sr/Ca content of breast milk from women consuming a strictly
traditional Maya diet. There is also no available data for the composition of ancient
maize from Lamanai, but in this case, dietary elemental composition can be inferred from
other Maya sources.
Importantly, since Lamanai mothers would have consumed very low Sr/Ca processed
maize, which would be further fractionated in their breast milk due to mammary gland
discrimination, we can correctly assume that the lime-processed maize gruel consumed
by their infants will still be enhanced in Sr/Ca compared to breast milk. Strontium
concentration in human breast milk has been found to vary from 0.14-0.35 mg Sr / g Ca,
or a Sr/Ca ratio of 0.14 to 0.35 x 10-3
(Harrison et al. 1965; Krachler et al. 1999;
Rehnberg et al. 1969; Rosenthal 1981). The variation can predominantly be attributed to
differences in water and food composition, but it does not include Maya individuals
consuming a traditional diet. Lacking such specific data, this range nonetheless forms the
basis of comparison with maize Sr/Ca and hard tissue composition in this study.
Regarding maize, White (1986) has noted that, on average, raw maize is composed
of 7.5 mg of calcium per 100g, or 75 ppm. In contrast, lime-soaked, and lime-soaked and
boiled, maize returns calcium values ranging from 370-2404 ppm, in experiments

256
undertaken by Wright (1994: Appendix F). For this study, it would be useful to know the
Sr/Ca composition of ancient Lamanai maize, as well as the relative strontium and
calcium contents of maize samples before and after lime treatment, but this is not
available. Consequently, maize strontium composition is inferred by geological factors
(soil, water), experimental tortilla research, as well as extrapolation from enamel Sr/Ca
using known levels of strontium discrimination (inverse of OR).
An important consideration in maize composition and processing is water. As in
food, strontium varies widely in water supplies, depending on geology (see Chapter
Four). Maize Sr/Ca composition will be directly influenced by water and soil chemistry.
The relative Sr/Ca abundance of local water is particularly significant because strontium
is known to be one of the few elements that are absorbed by vegetables from cooking
water (Losee and Adkins 1968).
Alkaline elements are more completely absorbed from liquids, particularly milk, than
from solid food (Marcus and Lengemann 1962). So while the Sr composition of maize at
Lamanai must be accounted for, the Sr content of local water is equally important
because infants are mainly eating watery gruel as a supplement. Contemporary evidence
also indicates that maize atole is frequently cooled with fresh water prior to feeding,
which results in a further diluted (and possibly strontium-enriched) gruel (Izurieta and
Larson-Brown 1995). However, while boiling maize and adding cool water will result in
a potentially strontium-rich watery gruel, calcium is also enhanced in the same manner,
maintaining the relatively low Sr/Ca signature.
Unfortunately, no site-specific water values are available for Lamanai, but local soil
Sr/Ca can be used to infer the relative composition of water and give an indication of

257
maize values, which is assumed to generally resemble the soil. The weathering of
limestone bedrock at Lamanai would have supplied most of the Sr and Ca that entered the
water and soil systems at the site (Wasserman et al. 1977). However, despite having high
Sr concentration, limestone has a relatively low Sr/Ca ratio due to the high abundance of
Ca, while shales and granites have the highest Sr/Ca ratios (Turekian and Kulp 1956a).
White and Schwarcz (1989: Table 2) have reported on the element composition of
some soil samples from Early Classic to Historical archaeological contexts at Lamanai.
Soil Sr/Ca ratios range from 0.737 to 1.045 x 10-3
(see Table 8.1 below), with a mean
ratio of 0.880 x 10-3
. As expected, bone Sr/Ca values are lower and range from 0.417 to
0.752 x 10-3
, averaging 0.529 x 10-3
, according to White and Schwarcz (1989: Table 2).
Table 8.1 Relative Sr and Ca data for Lamanai bone and soil samples
(based on raw data of White and Schwarcz 1989: Table 2)
Time Pd.
Structure
No.
N
Sample
Type
Sr
(µg/g)
Ca
(g/kg)
Sr/Ca
(x 10-3
)
Postclassic
N10-2
5
X bone
112.2
203.99
0.550
1 Soil 48.3 50.62 0.954
N10-3 1 Bone 87.5 198.87 0.440
1 Soil 65.6 78.57 0.835
N10-9 1 Bone 99.1 204.11 0.486
1 Soil 57.3 77.79 0.737
N11-5 1 Bone 154.2 205.14 0.752
1 Soil 52.9 50.62 1.045
Historical
YDL
5
X bone
84.7
203.35
0.417
1 Soil 51.6 62.09 0.831

258
White and Schwarcz’ (1989: Table 2) element data for Lamanai human bone and soil
samples indicates that bone Sr/Ca values are 1.28 to 2.75 times lower than soil values. In
comparison, enamel strontium composition is generally five times lower than dietary
Sr/Ca in adults (see above). But these soil strontium values are similar to mean soil
values observed by Wright (1994) for ancient Maya sites in Guatemala, which are, on
average, 1.53 times lower than bone Sr/Ca (see Table 8.2).
Table 8.2 Mean bone and soil Sr/Ca of Pasion Maya sites in Guatemala
(derived from raw data of Wright 1994: Appendix G)
Sample
Type
N
Mean Sr/Ca
(x 10-3
)
S.D.
(x 10-3
)
Range
Bone Ash
117
0.413
0.226
0.134-1.357
Soil 84 0.630 0.503 0.254-2.676
Based on strontium and calcium values observed by Wright (1994: Appendix G) for
remains from various sites in the Pasion region, Guatemala, other ancient Maya bone
Sr/Ca ratios range from 0.134 to 1.357 x 10-3
(which excludes one sample with a Sr/Ca
ratio of 3.699 x 10-3
). Wright’s raw data reflects the enormous variation that
characterizes human hard tissue and soil composition, particularly from different
geological and temporal origins. At the same time, it resembles the range of Sr/Ca values
observed in this study for Lamanai teeth: average enamel Sr/Ca values range from 0.631
to 1.254 x 10-3
, depending on age (see Table 7.4).

259
Wright (1994: Appendix D) has reported on the composition of one raw maize cob
from Altar de Sacrificios (Guatemala), which has a Sr/Ca ratio of: 1.13 x 10-3
. No other
data for maize Sr/Ca from the Maya area has been located in the literature, but LaVigne
(2002) has determined a mean Sr/Ca value of approximately 2.51 x 10-3
for six modern
unprocessed maize (kernel) samples from central Mexico. By inference, Lamanai soil
Sr/Ca values ranging from 0.737 to 1.045 x 10-3
(White and Schwarcz 1989: Table 2)
suggest that unprocessed maize Sr/Ca is likely not higher than 1.0 x 10-3
at the site, but
these samples derive from cultural contexts and do not necessarily reflect the Sr/Ca
values of agricultural fields.
It should be noted that strontium values vary significantly between plant parts, due to
differential discrimination, i.e., greater accumulation in roots and stems (Runia 1987b).
Plant roots, root crops and cereal grains have the highest Sr/Ca ratio, with values 1.5-10
times higher than leaves or shoots, which usually resemble soil levels (Runia 1987b).
Maize kernels and the cob likely differ in Sr/Ca ratio, suggesting that LaVigne’s kernel
average may be closer to the composition of dietary maize at Lamanai, but the efficiency
of strontium discrimination in maize species and the extent of differences between maize
parts are not known (see Burton et al. 1999).
Assuming that alkali treatment will reduce the Sr/Ca ratio by approximately six to
twenty times, or greater, Wright’s maize value is reduced to a range of 0.056 to 0.188 x
10-3
for lime-processed maize. LaVigne (2002) has similarly reported on Sr/Ca values of
six lime-treated tortillas from Solis, Central Mexico, averaging 0.176 x 10-3
(compared to
2.51 x 10-3
for raw Solis maize).

260
However, in laboratory experiments simulating alkali-processing, using comparable
Solis maize and cal (lime solution), LaVigne found that processed maize Sr/Ca reached a
mean of 1.3 x 10-3
(n=6). This value derives from kernels softened with lime, after
removal of the pericarp. Based on this finding, one can only assume that the process of
tortilla-making (including toasting on a comal, or griddle) entails changes in chemical
composition, i.e., dramatically reduced Sr/Ca, that do not resemble the chemistry of
alkali-processed maize at an earlier stage of preparation.
This higher ratio (compared to tortilla Sr/Ca) reflects a potential difference in the
Sr/Ca values of other traditional maize foods such as weanling gruel. Furthermore, the
dilution of maize dough with water, to produce gruel, provides an additional source of
strontium that could result in a food enriched in Sr/Ca compared to tortillas. Without
analysis of such watery maize food, which was the primary Maya weanling supplement,
one cannot assume that tortilla Sr/Ca values will infer the relative Sr/Ca of weanling
gruel. Importantly, this value is significantly higher than the Sr/Ca range of human breast
milk: 0.14 to 0.35 x 10-3
(Krachler et al. 1999; Rehnberg et al. 1969; Rosenthal 1981).
Elsewhere, Sr/Ca values of some raw maize in the United States average around 7.14
x 10-3
(Shacklette 1980; Watt and Merrill 1963), which is significantly higher than the
values of the raw Altar de Sacrificios and Solis maize. If one refers to the North
American raw maize Sr/Ca value, one arrives at hard tissue Sr/Ca ratios ranging from
4.28 x 10-3
at 0.5 years, 1.78 x 10-3
at 3.0 years and 1.64 x 10-3
at 5.0 years of age, using
Rivera and Harley’s (1965) age-specific OR values for Sr discrimination.
For Lamanai Maya, mean enamel Sr/Ca ratios range from 0.63 x 10-3
(Historical
cohort at 0.5-1.0 years) to 1.25 x 10-3
(Postclassic cohort at 5-5.5 years), which infers

261
dietary Sr/Ca values of 1.05 to 5.45 x 10-3
based on discrimination factors (see Table
7.17). The low end of this range resembles LaVigne’s (2002) Sr/Ca values for lab-
simulated central Mexican maize: 1.3 x 10-3
, as well as unprocessed maize: 2.51 x 10-3
.
The inferred Lamanai dietary Sr/Ca values are reduced compared to the 7.14 x 10-3
ratio observed by Shacklette (1980) and Watt and Merrill (1963) for North American
maize. This may suggest that the maize consumed by Lamanai infants was indeed treated
with lime to reduce the Sr/Ca ratio. But compared to the maize Sr/Ca derived from
Wright (1994), and especially the Sr/Ca range of treated maize after calcium enrichment
(0.056 to 0.188 x 10-3
), extrapolated Lamanai maize values are noticeably higher.
Yet, Wright’s (1994) bone Sr/Ca data also infers higher dietary Sr/Ca values than
those of lime-treated maize, ranging from 0.223 to 2.262 x 10-3
at 0.5 years (OR=0.60) to
0.583 to 5.900 x 10-3
at 5.0 years (OR=0.23). The range reflects sample variation (bone
Sr/Ca of 0.134 to 1.357 x 10-3
) and, as age is not specified, also accounts for different
levels of strontium discrimination (OR value). Clearly, the Sr/Ca values suggest that
hard tissue composition is inflated compared to the hard tissue values that should
theoretically arise from dietary components such as maize tortillas, based on human
strontium discrimination (OR).
Since enamel Sr/Ca ratios observed in this study fall within the range of bone and
dental values outlined in Chapter 8.5, one can propose several explanations for the
discord between treated maize values and theoretical food values derived from hard
tissues. Firstly, it is possible that the discrimination factors observed for strontium and
human metabolism are inaccurate. It is noted that Rivera and Harley (1965) and others
measured such levels of discrimination using 90
Sr, which is not a naturally occurring

262
isotope, but one produced from the fission of uranium and plutonium (nuclear testing).
Human gastrointestinal discrimination of naturally-occurring isotopes such as 86
Sr may
differ from cited OR values because of this distinction.
Second, accompanying the reductions in rates of enamel secretion and mineralization
that have been observed in some permanent teeth in the mid-crown to cervical regions
(see Chapter Three and Appendix A), it is possible that such changes also enhance Sr/Ca
incorporation in such enamel, i.e., after 2-3 years of age. This would explain the
continued rise in enamel Sr/Ca after two years of age, despite stabilization of high
gastrointestinal discrimination, and result in a lowering of the inferred dietary Sr/Ca.
Unfortunately, the current state of enamel mineralization research cannot confirm this
supposition, but the evidence overwhelmingly indicates that enamel Sr/Ca does continue
to rise after maturation of digestive systems.
While the enamel and inferred dietary data suggest otherwise, it cannot be assumed
that Lamanai maize was not treated with alkali because we do not know the typical
strontium and calcium values for Lamanai water, breast milk, maize and weanling diet
(atole). The extent of strontium enrichment associated with boiling of maize with
Lamanai water and the addition of water prior to consumption would be informative in
this respect. Other sources of strontium that could contribute to total hard tissue Sr/Ca
include native salts (critical for adequate nutrition and biological function), as well as the
clay of cooking and serving vessels, which are additional strontium sources in Hopi diet
(Kuhnlein and Calloway 1979).
It is possible that consumption of other higher Sr/Ca foods (e.g., mashed raw maize,
beans, other raw fruits or vegetables) might account for an elevated strontium input.

263
Importantly, experiments by LaVigne (2002) suggest that tortilla Sr/Ca values may be
reduced compared to other alkali-processed maize foods (e.g., mashed maize, dough,
gruel). Weanling foods, in the form of watery gruel, likely have enriched Sr/Ca values
compared to experimentally tested (modern) tortillas. Additionally, the total dietary Sr
could be elevated if maize intake is temporarily withdrawn or dramatically reduced,
relative to consumption of other (higher Sr/Ca) foods.
It is highly likely that Lamanai children were weaned on a diet almost entirely
composed of maize products. Bone chemical evidence suggests that up to two-thirds or
more (70%) of the Lamanai adult diet consisted of maize (White 1986). Since adults
consume relatively more meat and non-maize foods than children, it is not implausible to
attribute a predominantly maize diet to children, who also probably consumed some high
strontium foods such as raw fruit and, possibly, untreated mashed maize.
In the end, enamel should reflect a clear Sr/Ca rise in regions developing after the
start of food supplementation and exclusive nursing because, despite alkali processing of
maize, and a reduced Sr/Ca signal, mothers are consuming the same low Sr/Ca maize.
Maternal tissues will reflect this very low Sr/Ca value, with mammary gland
discrimination actually producing breast milk that is further reduced in relative strontium
composition.

264
8.2 Sr/Ca Behavior in Lamanai Permanent Canine Enamel
Several factors support the assertion that enamel Sr/Ca captured in this investigation
faithfully records a biogenic signal associated with childhood dietary intake:
1) enamel is highly resistant to diagenetic processes and can be relied upon to reflect
strontium incorporation during mineralization
2) hard tissue strontium is a very reliable indicator of dietary intake of strontium
during tissue development
3) Sr and Ca counts (and relative Sr/Ca) of enamel at the termination and start of
consecutive rasters within a single tooth are consistent, therefore discounting
instrumentation variables as the source of rising Sr/Ca over time (see Section 8.5)
4) enamel Sr/Ca patterns and dietary behavior associated with strontium change
correspond well with known childcare traditions, particularly the significant
increase in enamel Sr/Ca (food initiation) after six to nine months of age, and
attainment of peak Sr/Ca (weaning) between three and five years of age
The results of this analysis suggest that relative strontium abundance can be
recognized in permanent canine enamel in patterns that likely reflect dietary intake during
early childhood. From an archaeological perspective, understanding the diet and weaning
patterns of children at this critical stage can provide valuable inferences for the impact of
social change on biology.
In respect of their study on Medieval British skeletons, Richards and co-authors
(2002) have stated that for hard tissue-based research on infant weaning to be valid,
several criteria must be demonstrated: 1) a culturally determined age of weaning that is
observed by all members of the sample; 2) weaning that is relatively abrupt (i.e., a few

265
months); and 3) children were weaned onto foods similar to the adult diet. The results of
their investigation confirm all of their assumptions.
Here, however, enamel Sr/Ca data reflects important deviations. An alternative
weaning model can be constructed from the LA-ICP-MS analysis of Lamanai enamel,
and it is characterized by 1) variations in the age of maximum Sr/Ca values, or presumed
weaning age; 2) an extended period of gradual food supplementation; and 3) periods of
fluctuating Sr/Ca reflecting alternating breast milk and solid food intake, or fluctuations
in the types of weanling foods, i.e., due to seasonal availability, crop failure, food taboos
during illness.
While most teeth in this study exhibit an increasing Sr/Ca signature over time, many
Postclassic (31%) and Historical individuals (38%) are noteworthy for Sr/Ca patterns that
fluctuate. Besides reflecting variations in absorption and hard tissue incorporation, the
prevalence of gross fluctuations in enamel Sr/Ca pattern also likely reflects significant
cultural and biological changes in Lamanai society. Fluctuations in food intake, relative
to breastfeeding, may be related to pervasive illness and/or disruptions in access to food.
Furthermore, children’s nursing may be interrupted by the addition of subsequent
siblings, since many cultures view the breast milk of pregnant mothers as “unhealthy”
and inappropriate for consumption (see Chavez and Martinez 1982; Institute of Medicine
1991; Stuart-Macadam and Dettwyler 1995; among others).
Beyond availability, cultural perceptions or taboos against “contaminated” solid food
intake during illness (which is often associated with gastrointestinal disorders) might also
account for occasional food withholding, as observed in many societies (Dettwyler and
Fishman 1992; Fildes 1986; Stuart-Macadam and Dettwyler 1995; Wood 1983). In this

266
context, Dettwyler and Fishman (1992) observed the delay of any solid food
supplementation for up to a year or more. In some cases, undernourished and
underdeveloped children are nursed longer because mothers associate increased mortality
risk with the period after weaning (Caulfield et al. 1996; Simondon and Simondon 1998).
In contrast to the general model of steady, gradual, food supplementation until
weaning, the variation in canine Sr/Ca in both the Postclassic and Historical periods
indicates that weaning behavior at Lamanai was highly flexible and subject to subtle
changes. Enamel strontium clearly reflects a combination of: a) the adaptability of
children to changing socio-economic and epidemiological conditions, which is dependent
on maternal childcare practices; b) different diet and health histories of colonial Maya
children originating from elsewhere in the Maya Lowlands, as well as local children; and
c) variations in hard tissue element incorporation.
Enamel isotopic values analyzed by Wright and Schwarcz (1998) also reflect
considerable variation in the duration of lactation, which they attribute to variations in
child growth, nutritional demands, “maternal competence”, socio-economic factors and
temporal shifts in diet. Undoubtedly, these fluctuations reflect human adaptations to
changing nutritional, ecological and socioeconomic conditions, particularly during
colonial times.
Although hard tissue strontium cannot directly indicate the cessation of breast milk
intake, if one surmises that peak Sr/Ca followed by stable and/or decreasing values can
infer the period of maximum solid food intake (weaning), the Lamanai data indicate that
this transition is relatively late in both Postclassic and Historical Maya. In general, the
Lamanai weaning pattern is characterized by 1) dietary supplementation starting in the

267
first year, particularly by 0.75 years (9 months) of age; 2) gradual food supplementation
that accelerates after two years of age; and 3) breast milk intake that continues up to five
years of age.
Based on clinical data (Buckley 2001), it is likely that breast milk intake remained
high for the first two years, and then significantly decreased after three years of age,
when breastfeeding typically decreases in frequency and length of feedings. Further, the
results suggest that Richards and colleagues’ condition for a culturally determined
weaning age is not entirely appropriate for the ancient Maya, where infant feeding varied
a great deal within a general model of extended nursing.
Time of weaning across human cultures is highly variable and specific to each
family’s circumstances. Mothers, fathers and children are all active agents in the process.
Factors that influence the termination of breastfeeding include cultural perceptions of
milk quality or quantity, maternal illness, resumption of menses, birth of another child,
anxiety and emotions of the mother, economic changes (work responsibilities), social
trends (e.g., female body image and fashion), socioeconomic status, seasonal time of
year, infant’s age, tooth eruption, child’s bowel development and general health (Carballo
and Pelto 1991; Fildes 1986).
An important factor in the timing of weaning is illness. Withholding of food and
exclusive nursing has been associated with childhood illness, but sick children may also
cease breastfeeding due to an inability to suckle, or separation from their mothers. In this
case, extended exclusion of breast milk often results in the weaning of the child, due to
the mother’s inability to produce milk without suckling stimulation (or manual
expression), and the infant’s adjustment to solid foods. Thus, child illness may lead to

268
weaning, rather than the cessation of breastfeeding causing illness (see Institute of
Medicine 1991).
Numerous lines of evidence support an extended breastfeeding pattern in ancient
times (see Chapter Five). Many cultures attribute extended nursing, i.e., more than two
years, to greater child survival (see Fildes 1986). Besides contact period ethnographic
observations of infant nursing until three to four years of age (Landa, in Tozzer 1941),
extended breastfeeding among the ancient Maya is substantiated by contemporary
evidence: the mean duration of breastfeeding in indigenous Guatemalan (Maya)
communities is two years (Wright and Schwarcz 1998), while Guatemalan Ladinos
breastfeed for an average of 15 months (Izurieta and Larson-Brown 1995). Hard tissue
evidence of chronic infectious diseases, in the form of porotic hyperostosis and enamel
defects, particularly during the Historical period at Lamanai, is likewise associated with a
late (common) weaning age between two and six years of age (White et al. 1994).
8.3 The Nature of Ancient Maya Health and Nutrition at Contact
Significantly, the enamel strontium data reflects important temporal differences
between the prehistoric and historical populations at Lamanai that serve to enrich the
archaeological understanding of the contact experience. Most notably, Historical
Lamanai children were more susceptible to dietary fluctuations (inconsistency), and
experienced delayed food supplementation compared to their Postclassic counterparts.
Throughout the colonized New World, a soaring mortality rate was the gravest
consequence of contact. Contributing factors include introduced epidemics, violence,
increased infant mortality, reduced birth rates, female infertility, forced resettlement,

269
socio-economic reorganization and the overall effects of a perilous disease environment
(see Chapter Two).
Reflecting this turbulent era, the Postclassic and Historical Lamanai samples differ in
the mean Sr/Ca ratio of enamel at all intervals between 0.5 and 5.5 years of age, as well
as the extent of variable Sr/Ca patterns. For instance, at all age intervals, Historical
males exhibit reduced Sr/Ca ratios compared to Postclassic males, particularly after the
age of 2.25 years of age (Section 7.3). After this age, Historical males may have been
delayed in reaching comparable Postclassic values by up to 1.5 years, suggesting that
Historical male toddlers consumed relatively less solid food than prehistoric boys of the
same age.
Postclassic and Historical females reflect the most differences in mean Sr/Ca, with
Postclassic girls having higher enamel Sr/Ca at every age (Figure 7.13). Adjusting
strontium patterns indicates that Historical individuals may have required up to 2.25
years to attain comparable Sr/Ca values as Postclassic individuals. It is proposed that the
reduced mean Sr/Ca pattern in Historical individuals, and delayed food supplementation
compared to prehistoric Maya, reflects important cultural responses to colonialism at
Lamanai.
Interestingly, Lamanai bone composition data supports this finding. White and
Schwarcz (1989: Table 1) report a mean Sr value of 111.7 ppm (0.1117 x 10-3
) for 17
Post Classic individuals (mixed adults and juveniles) and 83.1 ppm (0.0831 x 10-3
) from
10 mixed Historical individuals. (This is at the lower range of mean bone strontium
abundance: 150-250 ppm in bone ash [Aufderheide 1989].) Mean Sr/Ca ratios for
Lamanai bone reflect the reduction from Post Classic to Historical times: approximately

270
0.225 x 10-3
(0.000225) in the Postclassic (N=17) and 0.205 x 10-3
(0.000205) in the
Historical cohort (N=10).
With Spanish rule, Maya populations were forced to pay tribute in the form of
material products and labor. More so than men, whose domain was principally the milpa
and hunting grounds, Maya women fulfilled this role because most demanded goods were
produced in the domestic sphere, i.e., kitchen garden produce, hens, honey, beeswax, and
especially cotton mantas (Clark and Houston 1998; Clendinnen 1982, 1987; Restall
1995).
In the colonial Maya household, women assumed mounting economic and domestic
responsibilities. Besides the traditional female role of household maintenance (cooking,
hygiene), kitchen gardening and childcare, Maya women were also responsible for
weaving, flour-grinding, raising animals for consumption and exchange, market
commerce, tribute payments and even direct labor tribute, i.e., domestic services in
Spanish establishments and households (Clark and Houston 1998; Clendinnen 1982;
Hunt 1974; Landa in Tozzer 1941; Restall 1995; Schroeder et al. 1997; Vail and Stone
2002).
Since their duties were mostly situated in a domestic context, many colonial Lamanai
women were able to sustain time-intensive childcare practices such as extended nursing.
Inconsistent resource availability (see below) likely contributed to the maintenance of
extended nursing in most children, yet, at the same time, greater incidence of peak
enamel Sr/Ca at earlier ages in the colonial cohort also suggest that more Historical
women weaned their children earlier. This behavior would be an expected outcome of
increased female labor and economic responsibility.

271
Extending into contemporary times, female work obligations continue to have
significant repercussions for infant care and health. In many societies, a market economy
leads to reduced breastfeeding among working women, increased use of substitute foods
(formulas), earlier infant weaning, increased infant mortality and even reduced fertility
(e.g., Chandra and Lakshmiswaramma 1991; Swedlund 1990; Swedlund and Ball 1998).
Like colonial Lamanai families, the movement of contemporary populations from rural to
urban settings also results in the disintegration of extended kin networks and integral
support systems.
Women are not visible in the historical record at Lamanai, but Lamanai women likely
assumed these roles too, since the site continued to participate in the exchange of goods,
which now included European products, with Spanish authorities and Maya merchants
trading between the Yucatan and the southeastern frontier (Graham et al. 1989). Colonial
Lamanai women were burdened with more responsibilities compared to pre-contact times
and childcare was often modified to accommodate the change, entailing either delayed
food supplementation or earlier weaning.
As Graham (1991: 333) has eloquently stated, colonial period Maya women acted as:
communicators of cultural information to children, as filters of Spanish ideas and
processors of new information, as dietary planners and maintainers of health and
hygiene, and as wives, advisers, and companions of the first Spaniards-the primary
weavers of the first tapestry to include threads of Old and New World cultures.
With an increase in the domestic workload of Maya women during colonial times, it
was often their bodies that suffered the physiological consequence. Being bearers of
future generations was probably the greatest health risk. Based on historical accounts
(Maya testators) and rates of infant and adult mortality, it is likely that, on average, Maya

272
women delivered at least four children in the mid-17th
century and more than eight by the
end of the colonial period, with a child survival rate of 50% (see Restall 1995).
As evident throughout the New World after contact, indigenous women’s health often
deteriorated with the added stresses of famine, infectious disease, social breakdown and
economic hardship (Cook 1998; Farriss 1984; Graham 1991; Larsen and Milner 1994;
Stannard 1990, 1991; Verano and Ubelaker 1992; White et al. 1994). Introduced diseases
that were devastating in their effectiveness include smallpox, measles, influenza,
tuberculosis, bubonic plague, typhus, cholera and yellow fever.
Lamanai Maya were also increasingly anemic in colonial times, as reflected in greater
rates of porotic hyperostosis (White 1986; White et al. 1994). No gender or age (adult
vs. child) differentiation was noted. Among women, this may have arisen from a
combination of helminthic or other parasitic infections, heavy reliance on an iron-poor
maize diet, excessive fluid loss from the tropical climate, monthly blood loss and/or the
physiological demands of pregnancy and lactation (White 1986; White et al. 1994).
Among children, the primary causes were probably parasitic infections (e.g., dysentery,
malaria, hookworm) and weanling diarrhea, combined with an iron-poor maize diet
(White 1986), which synergistically interact to exacerbate each condition.
As the most vulnerable members of society, Lamanai children, particularly of
weaning age, best reflect the changing epidemiological conditions of the colonial period.
At Lamanai, children experienced significantly higher rates of infection after contact.
Hard tissue evidence of more severe, acute, and episodic health stress among children
during colonial times includes microscopic enamel defects, or Wilson bands, which are
three times more prevalent in colonial remains (Wright 1990).

273
An important contributing factor to rates of infection and public health is poor
hygiene in crowded or unsanitary living conditions. Records in 1637 suggest that
Lamanai was sparsely inhabited in later colonial times, with a population of only 72
Maya (Jones 1989), but this is not necessarily indicative of the community just before
and after contact (White et al. 1994). Remember too, that colonial Lamanai was a
community in flux, with settlement that was scattered and impermanent in nature, both in
terms of population composition and architecture (Graham et al. 1989; Pendergast 1982).
In this context, it is likely that public hygiene, in the form of waste and water
management, was not regularly maintained. Weather cycles, i.e., the rainy season, would
have also increased the environmental pathogen load for several months of each year.
These conditions and the congregation of Maya from other areas that were disease-
affected (e.g., Yucatan) would have nurtured the transmission of infectious illnesses. In
extreme cases, the combined effect of infectious disease and malnutrition is a failure to
meet the increased physiological requirements of growing children during the weaning
period (Mata 1990; Mata et al. 1977; Rivera and Martorell 1988a, b; Scrimshaw 1978;
Scrimshaw et al. 1968; White et al. 1994; Wood 1983; among others).
Notwithstanding epidemiological conditions, the weaning transition is undisputedly a
critical stage of childhood. A marginal diet is an important factor. Unfortunately,
weanling foods are commonly deficient in protein and low in total energy, as well as iron
content, in the case of maize. The process can be traumatic and stressful and children
often experience separation anxiety, wasting and gastrointestinal disorders. In the event
of abrupt weaning, either due to social trends (e.g., among British and American
colonies) (Fildes 1986), or maternal illness or death, children are especially affected.

274
Common ailments often associated with the weaning process include diarrhea,
undernourishment (e.g., protein deficiency), vomiting, rickets, scurvy and infection. This
is manifest as greater rates of hard tissue pathology, particularly enamel defects, during
weaning (i.e., 2-4 years of age) (Cohen et al. 1994; Danforth 1989, 1997; Saul 1972;
Song 1997; White 1986; Whittington 1992; Wright 1990).
“Weanling diarrhea” is an important disease of childhood that has extensive
ramifications for growth and development (Scrimshaw et al. 1968). The clinical data
overwhelmingly demonstrates that it is a major source of infant morbidity and its role in
infant health and survival cannot be overstressed (Brown et al. 1989; Chen and
Scrimshaw 1983; Martorell et al. 1975; Mata et al. 1967; Mølbak et al. 1994; Sabin 1963;
Scrimshaw 1978; Walker-Smith and McNeish 1986, among others).
Spanish domination also degraded the health of women and children in other ways
related to social cohesion. Concomitant with Christianizing efforts, the traditional Maya
family structure and (multi-) household organization, which was virilocal (centered on
the husband’s family), polygamous and extended, was forcibly reduced to a Spanish
model of single nuclear families (Landa, in Tozzer 1941; Clendinnen 1982; Restall
1997). This disruption removed the familial support system, with its extensive physical
and psychosocial benefits, and further taxed a women’s time, her ability to fulfill multiple
domestic and economic roles, her health, and the health and care of her children. With a
permanent Spanish presence at the site, such domestic turmoil was likely reproduced at
Lamanai (Graham 1991).
Nevertheless, despite their subordinate social position in Maya and Spanish society,
Maya women were known to actively and passively challenge Spanish and Christian

275
authorities in many ways (Clendinnen 1982; Restall 1995; Schroeder et al. 1997). In fact,
their significant role in cotton cloth production and the colonial economy actually
enhanced their value in society to an extent, even giving them credence with legal matters
(e.g., inheritance) and social complaints in the Yucatan (see Restall 1995).
At Lamanai, enamel Sr/Ca suggests that Maya women may have also resisted Spanish
demands on their time by maintaining intensive childcare practices such as prolonged
breastfeeding. Despite increased demand on their time, colonial Lamanai women were
probably able to breastfeed for extensive periods because their responsibilities remained
largely within the domestic sphere.
Resistance might also extend to maintaining a traditional Maya diet, which is
indicated by bone chemical (White 1986) and faunal evidence (Emery 1999). Cross-
culturally, many societies actively express their identity and resistance to hegemony and
social change through specific food consumption behavior (e.g., Brown and Mussell
1984). In the Yucatan, ethnohistorical documents reveal that the Spanish disapproved of
Maya food and eating habits (Landa, in Tozzer 1941). As observed by Emery (1999: 78),
the maintenance of Late Postclassic dietary traditions at Lamanai in colonial times,
particularly animal utilization, reflects “a conscious return to traditional Maya foodways
and identity”.
Faunal and bone isotopic evidence may reflect general dietary stability in 16th
and
17th
century Lamanai, but observable changes in the types of exploited animal resources
also characterize the Historical period. At Lamanai, the introduction of bow and arrow
and fish netting technology in the Postclassic accompanied intensive exploitation of

276
freshwater resources into the colonial period (Emery 1999; Graham 1991). In particular,
fish and birds were intensively exploited in lieu of mammals.
Significantly, species shifts in animal exploitation reflect a level of socio-economic
and political upheaval associated with colonization, as well as the vulnerability of
reduced communities to inconsistent and unpredictable food production (Graham et al.
1989; Pendergast 1991b). Historical references to famines at Lamanai (J.F. Chuckiak,
pers. communication, 2004) fortify the precarious food situation at the site. Thus, while
the colonial Maya menu generally resembled the pre-contact diet, relative food
proportions (e.g., maize vs. meat) in Historical meals were probably more variable within
a season.
Based on hard tissue evidence of increased health disturbances during colonial times
(White 1986; White et al. 1994; Wright 1990) and the enamel strontium data, it is
postulated that poor colonial health may have affected Lamanai adults’ ability to produce
or procure nutritious foods for weanling infants. Due to the fluid nature of reduced
communities, agricultural production is often disrupted and unreliable (Graham et al.
1989). Conversely, diminished food sources and poor nutrition will lead to ill health. In
either case, it is a synergistic cycle that can result in exacerbation of the original
predicament.
Inconsistent maize production, in addition to ecological crop failure, seasonal
availability of wild resources and childhood illness are important factors that would lead
to changes in infant diet and the Sr/Ca composition of enamel. While pre-contact Maya
would have been subject to similar factors, the prevalence of Historical individuals with
fluctuating enamel Sr/Ca suggests that they were more vulnerable to dietary uncertainties,

277
and this is attributed to demographic instability, political economic factors and
epidemiological conditions.
The variable infant dietary pattern of colonial times may also reflect the impermanent
situation of northern Yucatec Maya refugees, who migrated to the southeastern periphery
to escape the greater Spanish domination of the northern Lowlands (see Chapter Two).
Their presence at Lamanai in colonial times resulted in a more heterogeneous population,
and it is possible that the enamel data identifies them.
In the end, whether motivated by child illness, dietary instability or political defiance,
many Historical Maya women at Lamanai delayed the supplementation of solid food to
their infants and breastfed longer than pre-contact mothers, potentially with detrimental
effect. Biologically, despite the overall benefits of extended breastfeeding, several
negative consequences can arise in both mothers and children if adequate nutrition is not
maintained. For optimal nutrition and growth, medical experts traditionally advise that
the introduction of solid foods should begin at approximately six months of age, when
breast milk quality, notably iron content, declines and infants require additional nutrients
to sustain growing energy requirements and general development (AAP 1980; Riordan
and Auerbach 1993; WHO 1981; Whitehead 1983, and many others). Indeed, a
significant increase in mean Sr/Ca between 0.5 and 0.75 years of age (6-9 months) for the
entire Lamanai sample (see Figure 7.1) suggests that solid food supplementation did
commence at this time.
Exclusive nursing for an extended period beyond six months of age can readily result
in malnourishment (Mata 1990). Specifically, breastfeeding for more than a year has
been negatively associated with growth and nutritional status (Asenso-Okyere 1996; Rao

278
and Kanade 1992). Sultan and Zuberi (2003) also attribute a late weaning age (i.e., mean
of 7 months) to be the most significant risk factor (predictor) in the development of iron
deficiency anemia in toddlers aged 1-2 years.
The negative association between prolonged breastfeeding and growth and nutritional
status is not consistent, however, due to differences in the control of variables such as
cultural and socioeconomic factors in clinical studies (see Buckley 2001). Other research
reflects a positive relationship between extended nursing past one year and infant growth,
as well as a decrease in malnutrition risk (Cousens et al. 1993; Prentice 1994; Taren and
Chen 1993). Importantly, mothers must balance the increased risk of parasitic infection
associated with food supplementation, with the need for increased nutrition for adequate
growth.
With changing attitudes, the American Academy of Pediatrics (AAP 1996) has
recommended breast milk consumption as the optimal infant diet for at least the first 12
months, “and thereafter for as long as mutually desired”. In Western societies
particularly, women have revived the practice of “Stone Age mothering” that is
advocated by La Leche League, whereby breast milk is always available to infants, who
nurse on demand, and can involve prolonged night nursing, co-sleeping and late weaning
(also see Hausman 2003; Menon et al. 2003). Fortunately, this is starting to provide
important western clinical data that can be systematically researched to assess the
physiological consequences of extended breastfeeding up to four years of age (e.g.,
Buckley 2001).
In truth, when nursing is extended, i.e., beyond two years of age, breast milk
consumption usually reflects a minimal (insignificant) dietary contribution.

279
Contemporary mothers themselves recognize that breastfeeding after one year represents
a minor dietary supplement to total diet (Buckley 2001). Instead, the continuation of
nursing beyond two or three years of age primarily fulfils an emotional purpose (Buckley
2001). As Buckley (2001) found, long-term “comfort nursing” is a nurturing process that
promotes mother-child bonding and acts to soothe and pacify both children and mothers.
With extended nursing, mothers recognize that their children obtain essential nutrients
from foods other than breast milk (Buckley 2001).
Buckley (2001: 310) found that nursing children aged one to four years required an
average intake of 100-460 mL of breast milk per day to meet the RDA recommendations
for energy intake and nutrients that were lower in their diet compared to national surveys.
Long-term nursed children had complementary (non-breast milk) diets that met an
average of 77% of the RDA for energy intake and at least 70% of the RDA for protein,
vitamin C, thiamine, riboflavin, niacin, vitamin B6, phosphorous and magnesium
(Buckley 2001: Table 4). This compares to non-breastfed children aged 1-3 years who
had mean energy intakes that met 93-108% of the RDA, the standards of which generally
exceed most children’s physiologic requirements (Buckley 2001).
The mean weight of breastfed infants is also significantly lower than formula-fed
infants up to 18 months of age (Dewey et al. 1992). However, Buckley (2001) found that
although children who nursed after the first year had lower weight-for-age, length/height-
for-age and weight-for-length/height, their values fell within two standard deviations of
the median and were deemed indicative of normal growth. Thus, while there are slight
nutritional inadequacies among long-term nursed children, her data reflect the absence of
major nutritional deficiencies among such children. Buckley (2001) posits that breast

280
milk probably compensates for the lower levels of nutrients found in children’s
supplementary foods, but further research is needed to assess the degree to which breast
milk contributes to the daily energy and nutrient intakes of children who breastfeed in
later years.
Maternally, lactating mothers experience short-term and long-term effects of
breastfeeding. The nutritional stress on mothers during lactation actually exceeds their
nutritional needs during pregnancy (Institute of Medicine 1991). Common short-term
physiological changes include increased energy expenditure; declining body fat
composition in the first six months of lactation, followed by increased body fat afterward;
ovulation suppression; and nutrient deficiencies (Picciano and Lonnerdal 1992).
Most significantly, the calcium requirement of nursing mothers increases to supply
nutritionally balanced breast milk. Adequate milk calcium concentration is maintained
by decreasing renal excretion of calcium and increasing bone calcium resorption (Cross
et al. 1995; DeSantiago-Soledad et al. 2002; Kent et al. 1990; Krebs et al. 1997; Laskey
et al. 1998; Sowers et al. 1993, 1995). Even in well-nourished mothers, particularly those
on high fiber (maize) diets (De Santiago et al. 2002), negative calcium balance can result,
with consequent reductions in bone mineral density (Cross et al. 1995; DeSantiago et al.
2002; Kalkwarf and Specker 1995; Krebs et al. 1997; Laskey et al. 1998; Sowers et al.
1993, among others). This long-term effect is particularly evident during menopause and
commonly affects the spine, femoral neck and trochanter (e.g., Gur et al. 2003).

281
8.4 The Implications of Infant Weaning Behavior for Ancient Maya Society
As a patriarchal society, ancient Maya recognized gender distinctions favoring males
that were manifest in several ways. Socially, economically, and politically, males
dominated Maya society and these advantages can be recognized in human hard tissues
because of increased access to status foods and enhanced nutrition (White 1999; White et
al. 1993; Whittington and Reed 1997; Wright and White 1996; among others).
Like many women in patriarchal societies, Maya mothers deferred status foods to
husbands and male children as a result of long-standing “processes of enculturation that
link(ed) food, status and gender” (Ardren 2002: 73). In many such societies, the custom
to invest greater childcare on male infants can begin at birth (Saunders and Barrans 1999)
and often entails extended breastfeeding, or delayed weaning, in such children (see Fildes
1986; McKee 1984). In Europe, medieval writers and doctors specifically recommended
an extra 6-12 months of nursing for male infants (Fildes 1986).
The nature of Lamanai enamel Sr/Ca reflects this gender discrimination to an extent.
While not significantly different, the Postclassic male pattern appears slightly delayed
compared to female individuals (Figure 7.9). Specifically, males appear to receive breast
milk up to 0.25-0.5 years longer than female infants (or receive comparatively less solid
food supplementation than females), which is compensated by a subsequent acceleration
in food supplementation.
In fact, Postclassic female enamel strontium is slightly elevated compared to the
mean values of all Postclassic and Historical cohorts (Figure 7.4), possibly suggesting
their earlier food supplementation. While some female adults exhibit higher bone Sr/Ca
compared to males, which has been attributed to metabolic changes associated with

282
pregnancy and lactation (see Chapter 4.4), such a distinction obviously cannot account
for the differences observed here in enamel formed during childhood. No evidence exists
to indicate that infant girls differ in strontium discrimination or hard tissue incorporation.
The disparity would appear to reflect differences in dietary supplementation (and
maternal childcare) of Lamanai boys versus girls.
Furthermore, possible gender distinctions in the rate of enamel development may
affect dietary reconstructions based on enamel Sr/Ca between males and females.
Several human and primate studies have found that female dental development is
advanced compared to males (Demirjian 1986; Demirjian and Levesque 1980;
Kuykendall 1996; Moorrees et al. 1963). While the difference is not large, i.e., in the
range of 2-3% (see review in Smith 1991), the difference in mean enamel Sr/Ca observed
between the sexes suggests that female food supplementation and weaning is even further
advanced compared to boys.
If one assumes that the relative mean Sr/Ca patterns reflect tangible differences in the
time required to attain comparable values, as “shifting” sample distributions illustrate
(Chapter Seven), then one can recognize important distinctions in the level of food
supplementation between cohorts. In this case, one can assess the physiological
consequences of a 3-6 month delay among Postclassic male infants, which is apparent
from the start of enamel sampling, i.e., 6 months of age, but which may have begun
earlier. At this early age, increased nursing of male infants compared to females, or less
solid food supplementation of males compared to females, may have entailed nutritional
consequences such as reduced growth rate and possible deficiencies (e.g., iron).

283
However, subsequent accelerated food supplementation among males after two years
of age, which is evident as a steeper Sr/Ca slope (Figure 7.6), indicates that Postclassic
males may have recovered from the early delay in supplementation. Hard tissue evidence
supports this, with no gender differentiation in disease and undernutrition in Lamanai
adults (White 1986; Wright 1990). Rather than being nutritionally detrimental, it appears
that the expedited food supplementation of infant boys after a slightly longer intensive
nursing period, i.e., shorter transition period to total food reliance, compensates for the
early delay.
Postclassic females also reflect greater fluctuations in enamel Sr/Ca compared to the
number of males exhibiting the expected pattern of increasing strontium over time. This
suggests that female children may have been more susceptible to dietary inconsistency or
illness than boys and reflects the greater gender differentiation in the pre-contact period.
The gender distinction is less evident in Historical individuals. The general equality
that is reflected in Historical male vs. female enamel Sr/Ca (Figure 7.11a) indicates that
infants at this time shared a feeding pattern that was not significantly influenced by
gender. This is not unusual considering the egalitarian (yet subordinate) status of all
colonial Maya reduced under a Spanish system. The archaeological record at Lamanai,
particularly the nature of Maya interments, supports this shared social position (Graham
et al. 1989; Pendergast 1991b, 1993).
Yet, Historical females do have a slightly reduced mean Sr/Ca ratio at almost all age
intervals, indicating that there might have been some gender differences. In this case, it
is female children who are delayed, possibly by as much as one year, in attaining
comparable enamel Sr/Ca (food supplementation) as boys (see Figure 7.11b). Raw bone

284
strontium abundance quantified by White and Schwarcz (1989: Table 6) support the
gender difference. Bone strontium averages 96.6 ppm (N=15; s.d.=12.7) for combined
Postclassic and Historical females, while males from the same period average 105.3 ppm
(N=14; s.d.=26.2) (White and Schwarcz 1989: Table 6).
At many Maya sites, skeletal and dental evidence portray girls and women as
disadvantaged members of society, with poorer health and reduced access to resources
(Ardren 2002; Saul and Saul 1997; Whittington 1991; Whittington and Reed 1997). At
Lamanai, this is manifest in bone chemistry as greater meat and seafood consumption
among males, particularly elite males, in the Classic period (White 1986; White and
Schwarcz 1989).
The findings are not consistent, however, as other evidence indicates general equality
between the sexes. For instance, macroscopic and microscopic enamel defects, which
reflect physiological disturbances (dietary or disease-related), are equally distributed
among the sexes in many Maya samples (e.g., Danforth 1997; Song 1997; Whittington
1992), including Lamanai (White 1986). Porotic hyperostosis prevalence at Lamanai
similarly reflects gender equality (White et al. 1994).
Other dietary reconstructions based on bone composition refute gendered distinctions
in food access (Gerry and Chesson 2000). It could be implied that male and female Maya
infants probably shared the same weaning process (Danforth et al. 1997; White et al.
1994), but the inconsistent data suggests that there is no pan-Maya blueprint for gendered
weaning behavior, with differences possibly related to regional or even site-specific
socio-political and economic factors that determine gender relations.

285
Finally, the enamel strontium data reflects important distinctions between individuals
who died as children and those who survived to adulthood. Though not significantly, it
appears that Postclassic individuals who died in childhood may have been delayed in
obtaining comparable levels of solid food supplementation as surviving adults. Like
early supplementation, delayed supplementation later in infancy (i.e., after six months of
age, when children’s energy requirements increase) also has potentially detrimental
repercussions.
It is proposed that some Lamanai individuals who died as children may have been
“breast starved” (Orkney 1946), or exclusively breastfed for more than six months. This
can readily lead to malnutrition, particularly protein and iron deficiency, and may have
been a contributing factor to early mortality. In fact, growth faltering has been observed
as early as three months of age in exclusively breastfed (indigenous) infants in
Mesoamerica (Butte et al. 1992; Rivera and Ruel 1997).
Even in cases of extended nursing, breast milk’s anti-microbial and immunological
qualities are active and provide supplemental protection for growing children (Institute of
Medicine 1991). There is no clear consensus on the nature of breastfeeding and mortality
after the first year, however, with some research indicating continued safeguarding
against mortality up to three years of age (Briend et al. 1988), while others find no
protection associated with extended nursing in the second and third years of life
(Cantrelle and Leridon 1971). As discussed earlier, exclusion of solid foods from infants
can be attributed to several factors, mainly concerning cultural taboos about food
contamination and diet during illness.

286
8.5 The Comparative Hard Tissue Strontium Data
As there is no comparative incremental Sr/Ca data for human enamel, results of this
investigation can only be assessed by analogy to bone chemistry findings. Table 8.3
outlines the results of paleodietary investigations and also includes mean Sr/Ca ratios
from a clinical study of human bones from the United Kingdom between 1960-1968
(Sillen and Kavanagh 1982: Table 4, adapted from United Kingdom Medical Research
Council, 1959-1970). Total enamel Sr/Ca derived from 86
Sr/43
Ca ratios in Section 7.2
can be compared to such research.
Results from Sillen and Smith (1984) represent the first paleodietary study to
associate bone strontium with infant diet and weaning. As their report provides only
graphic distribution of Sr/Ca ratios (Figure 3), one refers to Katzenberg and colleagues’
(1996: Table 2) estimate of the values. Similarly, their organization of the results from
Grupe (1986) and Huhne-Osterloh and Grupe (1989) (see Katzenberg et al. 1996: Table
2) is referred to in the following table (Table 8.3).
Other bone strontium data in this comparison include estimates from Farnum and
Benfer’s (1995: Figure 4) report on ancient Peruvian infant bones and, most recently,
Mays’ (2003) analysis of Medieval British bones (estimates from Figure 1). Individual
tables in Chapter Five outline the supporting details of such calculations (when
available), such as sample numbers (N), standard deviation and range.

287
Table 8.3 Comparative age-related mean Sr/Ca (x 10-3
) values from various clinical and paleodietary investigations
Age
(in years)
United Kingdom
Medical Research
Council (1959-1970)
Sillen and
Smith 1984
Grupe 1986;
Huhne-Osterloh
& Grupe 1989
Farnum and
Benfer 1995
Mays 2003
Song 2004
Birth
0.22
1.50
0.52
0.5 0.22 1.75 0.58 0.87 0.71
1 0.29 2.25 0.54 0.73
2 0.27 2.50 0.72 0.89 0.76
3 0.23 2.25 0.84
4 0.30 0.63 0.85 0.97
5 0.29 1.07
6 0.32 0.84 1.13
8 0.27 1.60 0.75
14 0.27 0.51 0.79
Adult 0.27 1.80 0.53 0.69

288
As the table indicates, there is considerable variation in the total Sr/Ca abundance of
human hard tissues, which is not unexpected considering the geographic differences.
Notably, Sillen and Smith’s (1984) mean Sr/Ca values of ancient Arab bones is higher
than any other known data, with a range of approximately 1.5 x 10-3
(0.0015) to 3.0 x 10-3
(0.003). The authors do not specify whether these ratios are the result of relatively high
Sr values, or relatively low Ca abundance. Mays’ (2003) data is most comparable to this
study, ranging from approximately 0.50 x 10-3
(0.0005) to 1.05 x 10-3
(0.00105), or a
mean ratio of 0.855 x 10-3
(0.000855) among juveniles and 0.692 x 10-3
(0.000692)
among adults.
Farnum and Benfer (1995) have also examined Sr content in ancient Peruvian bones
to identify weaning, but in their opinion, the Sr/Ca values were too high not to rule out
diagenetic alteration. In their scatter plot of Sr abundance, the peak value, observed for
bones at one year of age, is approximately 540 ppm, or 0.54 x 10-3
(Farnum and Benfer
1995: Figure 4). However, placing their data in the context of other studies and clinical
data (Table 8.3) indicates that this mean ratio is actually within the normal range of
human findings, and is comparable to values in this analysis.
Besides absolute Sr/Ca values, two main differences can be recognized between the
Lamanai results and other studies, which can be attributed to the nature of sample
collection and analytical technique. Though not significantly, 1) the total mean Sr/Ca
increase in the Postclassic and Historical Lamanai samples is pronounced compared to
other studies, and 2) the Lamanai patterns also exhibit more variation that does not
necessarily adhere to the expected model of positive increase followed by decline and
stabilization.

289
Sr/Ca change over developmental time in Lamanai individuals involves a roughly
twofold increase in values. The mean increase in Sr/Ca is 1.807 among Postclassic Maya
and 1.873 in Historical Maya at Lamanai, which occurs over an extended period between
0.5 and 5.5 years of age (Table 7.15).
In comparison, Sillen and Smith (1983, 1984) found that Sr/Ca increases by a factor
of approximately 1.67 between birth and two years of age (see Katzenberg et al. 1996:
Table 2). Sr/Ca values of Medieval Arab bones gradually increase after birth, with
individuals aged 1.5 to 3.5 years having the highest values for the entire population,
which peaks at around 2 years of age (Sillen and Smith 1984). This is consistent with
ethnographic data on traditional Palestinian Arab communities, whose children consumed
breast milk until two to three years of age (Grinquist 1947). Afterward, with increased
gastrointestinal discrimination of strontium, Sr/Ca values decline to levels that persist
throughout adulthood.
Grupe (1986) and Huhne-Osterloh and Grupe (1989) also examined strontium
concentration in ancient German bones to estimate weaning age. They found the highest
Sr/Ca ratios in children aged six months to two years, the period they attribute to dietary
supplementation and weaning. In this case, Sr/Ca increased by a factor of 1.24 (see
Katzenberg et al. 1996: Table 2). Like Sillen and Smith (1984), bone strontium levels
declined in subsequent age groups.
Finally, Mays (2003) found that bone Sr/Ca increased most significantly between one
and two years of age in a Medieval skeletal sample, the period of significant food
supplementation. A steady decline in Sr/Ca from about two to eight years of age, when
values then resemble adults, indicates that most children in this sample were fully weaned

290
by two. The overall rate of increase between exclusive breastfeeding and weaning is not
explicitly stated, but graphical data suggest that infant bone Sr/Ca rises by a factor less
than 1.25 (Mays 2003: Figure 1). In this instance, Mays (2003) determined that the
transition from exclusive breastfeeding to an adult diet involved an increase in dietary
Sr/Ca by a factor of three to four.
In Mays’ (2002) study, bone strontium composition accords well with nitrogen
isotope results and Medieval documentation about infant diet. However, importantly, it
should be realized that in all cases, calculation of the dietary Sr/Ca based on Rivera and
Harley (1965) will indicate a significant rise in Sr/Ca at two years of age due to their
significantly reduced OR value at that age. Even in cases where hard tissue Sr/Ca is
stable or slightly diminishing between birth and two years of age (and older), enhanced
strontium discrimination at two years of age (more than three times) will return a
substantially higher dietary Sr/Ca value at that age. This pattern more correctly reflects
the increased ability to discriminate against strontium in more mature infant guts.
This fact reinforces the necessity for supporting contextual data in paleodietary and
paleonutritional investigations. In the reconstruction of ancient weaning based on hard
tissue chemistry, ethnographic, historical and archaeological inferences are imperative.
The hard tissue and dietary Sr/Ca data of Mays’ (2003) and Sillen and Smith’s (1984)
studies are validated by historical documentation, but here, in light of ethnographic
observations, the dietary Sr/Ca derived from enamel strontium must be scrutinized.
On one hand, inferred dietary Sr/Ca indicates that Lamanai Maya children
significantly increased their solid food intake at approximately two years of age. Stable
isotopic composition (δ15
N and δ13
C) of ancient Maya teeth from Kaminaljuyu support

291
this, with nitrogen data indicating that proteins from milk became less important in infant
diet by two years of age, when milk proteins were largely replaced by solid food proteins
(Wright and Schwarcz 1998, 1999). According to Wright and Schwarcz (1998, 1999),
Maya infants were nursed for at least one year or longer. While all children began
consuming significant quantities of solid food before two years of age, many children
were substantially supplemented by one year of age.
At the same time, enamel δ18
O suggests that children continued to rely on breast milk
for many years afterward, i.e., up to five or six years of age. Two different nursing
“strategies” are identified from the dental nitrogen and oxygen isotopic data: 1) many
children who imbibed breast milk up to five years, primarily as a water source; and 2)
few children who relied on breast milk to provide nutrients beyond one year of age
(Wright and Schwarcz 1999). Unfortunately, historical documentation does not reveal
when Maya infants were significantly supplemented with food, and contemporary
evidence in this regard is also lacking (Wright and Schwarcz 1998: 12).
Considering the various nutritional, physiological and social data, greater food
supplementation at two years of age can be associated with a combination of factors: 1)
growing developmental needs of children; 2) complete eruption of the deciduous
dentition, which is perceived to accommodate greater solid food intake; 3) better
developed digestive systems; and 4) increased child interaction with a wider social
environment.
On the other hand, as also found by Wright and Schwarcz (1998, 1999), it is likely
that many Maya infants were significantly supplemented earlier than two years of age,
but this is not evident in the Lamanai dietary Sr/Ca values due to the reduced OR factor

292
at two years (which will always indicate a substantial increase at this age). As well,
supplementation with substantial solid food after two years of age will not be visible in
the dietary Sr/Ca values due to the stabilization of hard tissue OR after that age. Enamel
Sr/Ca values at age three and older would have to be higher than two-year-old values by
an order of magnitude to reflect a significant difference with dietary Sr/Ca at two years of
age. This is not evident in the enamel strontium data presented here.
Nevertheless, based on the supporting data, it can be postulated that most Lamanai
children probably started consuming significant quantities of solid food at around two
years of age, with weaning taking place several years later. But while enamel strontium
can be used to infer the age of weaning, the nature of this investigation cannot clarify the
relative proportion of breast milk and solid food in later infancy.
In this study, among most Lamanai individuals, enamel Sr/Ca reaches its peak
between 4.5-5 years of age. Specifically, Postclassic Maya display peak values followed
by stabilization between the ages of 3.75-5 years, while the highest Historical Maya
values range from 3.5-5.25 years of age. In the entire sample, the time it takes to reach
peak Sr/Ca ranges from 2.75 to 4.25 years (see Section 7.3). In each distribution, a
subsequent drop in Sr/Ca reflects the more efficient discrimination of Sr by mature
digestive systems. These findings are significantly later than those found by Sillen and
Smith (1984), Huhne-Osterloh and Grupe (1989) and Mays (2003), which all attribute
peak bone Sr/Ca values to around two years of age.
However, it is also known that strontium levels are the most varied among children,
particularly between the ages of 4 and 13 (Lambert et al. 1979; Sillen and Kavanagh
1982). In fact, Rivera and Harley (1965) found that Sr/Ca values only reduced to

293
comparable adult levels by 8-10 years of age, when the digestive system is fully mature
and efficient at discriminating against strontium. Adolescence is also characterized by
high Sr values due to increased metabolic activity (Lambert et al. 1979; Loutit 1967). In
general, permanent enamel strontium is highly variable, with values as low as 13 ppm to
1400 ppm in teeth from dissimilar geographic regions and time periods (Chapter Four).
The comparison of bone Sr reference data to this dental study might account for some
differences in Sr/Ca, but bone Sr concentrations generally resemble enamel values: bone
Sr averages between 150-250 ppm (Aufderheide 1989), while enamel Sr commonly
ranges between 100-200 ppm (Brudevold & Steadman 1956). Nonetheless, in the event
that bone values are derived from diagenetically altered tissues, which is not uncommon
for bone Sr composition, the use of different hard tissues may be a significant factor.
Imprecise “ageing” of bone remains due to continuous tissue regeneration may also
account for some differences.
Two additional factors are also considered. Firstly, the late age of Sr/Ca stabilization
may simply reflect the extended breastfeeding behavior of ancient Maya children. The
populations examined by Sillen and Smith (1984), Huhne-Osterloh and Grupe (1989) and
Mays (2003) are known to have nursed for considerably shorter periods, including earlier
supplementation with animal milk in the Medieval British sample (Mays 2003). As
observed by Schroeder and colleagues (1972), children and adults from cultures that
consume animal milk after childhood tend to have reduced hard tissue Sr/Ca due to
higher calcium intake.
Second, instrumentation factors such as sample buildup and associated memory
effects in the ICP, or elemental fractionation over time, might account for a continued

294
rise in Sr/Ca over time, resulting in peak values at the end of the ablation sampling. The
ablation of calcified tissues can lead to significant intrainstrument buildup of calcium and
instrument drift (Outridge et al. 1995). But in such cases, particularly with carbonate-
based samples, calcium is accumulating in comparable abundance, so that any ratio, e.g.,
Sr/Ca, should remain indicative of true composition. Furthermore, in this study, as
terminal element values of an ablation correspond very well with initial values of the
subsequent ablation (facilitating “pasting” of multiple ablations to create one complete
graph), it does not appear that calcium accumulation is significant. In total, the
correspondence of enamel strontium patterns between ablations within an individual
tooth and the variability of enamel strontium patterns suggests that the increasing Sr/Ca
behavior observed in most Lamanai teeth are real and reflective of dietary
supplementation.
8.6 Recommendations for Future Research
This study represents the first intensive reconstruction of ancient infant diet using
LA-ICP-MS analysis of human enamel. As a result, there is a lack of comparable data,
but it is anticipated that these challenges will be resolved with additional research.
Like other avenues of bioarchaeological research, standardization of analytical
methodology would be invaluable, particularly in: 1) tooth sample preparation, including
the use of macroscopic or histological landmarks for tooth sectioning (e.g., to capture
consistent areas of enamel according to developmental age); 2) LA-ICP-MS operating
parameters; 3) data organization (e.g., software); and 4) use of specific isotopes for

295
element ratioing. Improvements in quantification, such as matrix-independent solid
reference materials, would also facilitate data comparisons.
Gunther and colleagues (2000: 13) predict that future LA-ICP-MS instrumentation
will be simplified in set-up and operation, with increased automation with sample
changers and data reduction. Continued investigation into laser wavelength, transport
efficiency and element fractionation (Gunther et al. 2000; Roy and Neufeld 2004) will
surely enhance the technology and fine-tune its application to bioarchaeological research.
Beyond this, as undertaken by Haverkort (2001), applying LA-ICP-MS to the hard
tissues of individuals with known dietary history will further evaluate: 1) the suitability of
the technology to paleochemistry and paleonutrition; 2) the complex nature of element
relationships between dietary components and the effect on total element intake; and
most fundamentally, 3) the relationship between dietary intake and enamel incorporation.
For this specific investigation into the nature of hard tissue strontium and solid food
supplementation, further quantification of strontium would be a fruitful avenue of
continued research. Less elements, i.e., only strontium and calcium, might be quantified
at a slower scan speed, which would reduce the “memory effect” common with LA-ICP-
MS, when ablated material is mixed with material from subsequent laser pulses due to the
delay during sample transport from the ablation chamber to the ICP (see Haverkort
2001). This would improve spatial resolution and sample quantification, as well as
reduce instrumentation error.
LA-ICP-MS analysis of other teeth could also contribute important supporting data.
In particular, since the earliest canine enamel (developing at birth) is not sampled due to
its interior (“hidden”) position, analysis of deciduous enamel developing before and at

296
birth, e.g., deciduous canines and molars, could provide an unequivocal non-food
supplemented Sr/Ca baseline from which permanent canine enamel could be compared.
However, in this case, analysis of deciduous enamel would be limited to individuals who
died as children and who retain complete crowns of such deciduous teeth, in addition to
secondary canines, i.e., those who died between approximately six and ten years of age.
To verify the findings in this research, i.e., the association of permanent canine
enamel Sr/Ca with infant diet and weaning, isotopic analysis would be a worthwhile
future endeavor. In this case, the same Lamanai canines could undergo additional
microsampling to quantify carbon (13
C), nitrogen (15
N) and oxygen (18
O) isotopic
composition. As described in Chapter Five, such isotopes can be used to infer the
introduction of non-breast milk water (oxygen) and maize foods (carbon), as well as the
cessation of nursing (nitrogen).
Drawing from this study’s dataset, other elements quantified by ICP-MS could also
be analyzed. To elucidate the weaning process, zinc may be another valuable element
that can be associated with changes in relative breast milk and solid food consumption.
Breast milk is known to be a good source of zinc (Institute of Medicine 1991; Lonnerdal
et al. 1981), but during the course of lactation, its zinc levels have been observed to
steadily decrease (Karra et al. 1988). Similarly, Farnum and Benfer (1995) observed
diminishing Zn levels in the bones of young children, but they could not differentiate
whether the low levels were due to changes in infant diet and weaning or other dietary
and health stresses (e.g., protein deficiency).
Like hard tissue elemental studies in general, which dwindled in light of diagenesis
research and better understanding of the preservation of bone elements, zinc has not been

297
investigated in the context of weaning studies since. However, in the context of dental
enamel research, it is possible that re-examination of zinc incorporation during infancy
and weaning, and use of microsampling techniques to provide continuous data at each
stage of the process, may provide useful comparative data for the strontium evidence.
Finally, beyond weaning, strontium isotope analysis would be an important extension
of this investigation to assess the nature of population heterogeneity at Lamanai.
Strontium isotope ratios (87
Sr/86
Sr) have been examined to identify the childhood origins
of individuals and groups, as well as the nature of group migrations among
archaeological samples (e.g., Budd et al. 1996, 1997; Goodman et al. 1999; Grupe et al.
1997; Gulson et al. 1997; Price et al. 1994; Prohaska et al. 2002; Sealy et al. 1995). It is
based on the premise that distinct ratios of the two isotopes reflect different geological
environments, most notably local water, soil and food.
This is an area of research with significant implications for population dynamics in
the Maya area as well, and particularly for Lamanai, which is known to have been a
frontier mission. Strontium isotope levels could shed light on questions of origin, and
provide a clearer picture of population movements incurred by Spanish reducciones.
Importantly, identification of immigrants in the Maya colonial cemetery at Lamanai
might explain the degree of variation in enamel Sr/Ca from this cohort. Colonial
Lamanai individuals raised elsewhere as children would have had varying experiences
with disease, famine, diet changes, violence, psychological trauma, and the stress of
migration. Thus, for instance, one might be able to associate a highly variable/fluctuating
enamel Sr/Ca pattern with individuals originating from regions that suffered the most
under colonialism.

298
CHAPTER 9
CONCLUDING REMARKS
As animals, what we eat determines the course of our biological and social evolution.
This realization has fueled the immense growth in paleonutritional studies that seek to
reconstruct subsistence from hard tissue chemistry. Advances in analytical chemistry
have greatly aided in the pursuit.
From this investigation, LA-ICP-MS results of enamel composition suggest that
strontium patterns do reflect changes in diet attributed to solid food supplementation,
waning breast milk intake and eventual weaning. This process begins before a Lamanai
child’s first birthday and ends three to four years later.
As illustrated by this study, the reconstruction of ancient diet and feeding behavior
must be an essentially holistic, anthropological, endeavor. Since the interactions
between food chemistry, processing techniques, human metabolism, health, nutrition and
hard tissue incorporation are complex and incompletely understood, diet must be
reconstructed within the context of diverse data. A critical biocultural approach accounts
for the interrelatedness of social and physical environments and it is a research paradigm
that clearly recognizes the diverse conditions that shape the health of society’s most
vulnerable members.
In this way, enamel development and diet-related chemical composition are viewed as
sensitive indicators of total environmental quality. Variations in enamel Sr/Ca pattern
within and between individuals reflect this sensitivity.

299
Teeth are proven to be the ideal human tissue for paleodietary reconstruction based
on chemical composition. Circumventing the main challenges of bone chemical analysis,
i.e., mineral preservation and diagenetic alteration, this dental study represents a return to
hard tissue element analysis. The kymographic nature of teeth, at a known rate of
development, is an additional advantage and it is perhaps its most unique feature. It
means that enamel represents a detailed chronicle of important milestones in early life
history. In this case, it has allowed observations of the dietary transition to weaning and
has provided an enriched understanding of indigenous health and nutrition.
For Lamanai, archaeological, osteological, ethnohistorical, ethnobotanical,
zooarchaeological and bone chemical evidence provide substantial inferences for the
nature of infant diet and health before and after contact. Invaluable contextual
information includes details of childcare, maternal health, female household and non-
domestic roles, and gender disparities. Altogether, the evidence elucidates the five
archaeological questions that were posed at the start of this analysis:
1) Are there temporal distinctions in breastfeeding and dietary supplementation between
pre-colonial and historical periods?
The data suggests that there are recognizable distinctions in subsistence pattern
between Postclassic and Historical Lamanai children. The primary differences
concern a) age of initial food supplementation, which is delayed in colonial children
compared to pre-contact children; and b) dietary inconsistency, which is more
prevalent in colonial children, who exhibit more fluctuations in alternating breast
milk/solid food intake.

300
2) Did Spanish control of economic activities, i.e., demands for agricultural products
and human labor, affect household patterns and childhood nutrition?
It is likely that Spanish demands for female-produced tribute products affected
mothers’ abilities to provide consistent solid food supplementation. However, as
tribute production was amenable to household settings, mothers were able to maintain
intensive childcare practices such as extended breastfeeding (see #3).
3) Did this demand for increased labor result in changes in weanling diet or duration?
The Sr/Ca evidence suggests that colonial period children consumed more breast milk
for longer periods than prehistoric children, i.e., they were weaned at a later age.
Maize gruel continues to comprise the bulk of weanling diet, but as in Lamanai
adults, the minor meat component of colonial children’s diet (when available)
probably shifted from land mammals to lagoon fish and birds.
4) Is the heterogeneous nature of the colonial mission population, including Maya
“refugees” from other areas, reflected in childhood diet and weaning patterns?
While identification of such sub-cohorts is not possible in this study, the extent of
individuals with variable Sr/Ca patterns in the colonial period may indeed reflect the
varied dietary patterns of the heterogeneous mission population. Fluctuating breast
milk and solid food intake of colonial children may also mirror the transitory nature
of colonial Maya refugees who settled at Lamanai, after fleeing from northern regions
(Yucatan) and epidemic disease. Dietary inconsistency would be an expected
outcome of their ecological, domestic and socioeconomic instability.

301
5) Are there distinctions in weaning age, duration or diet that could be related to
socioeconomic status or gender?
A socioeconomic distinction is recognized between temporal cohorts at Lamanai,
with Postclassic Maya having greater access to resources (including social power),
than Historical individuals. This is manifest as differences in enamel strontium and
diet that are outlined in #1 above, i.e., reduced Sr/Ca values (delayed supplementation
and weaning) in the disadvantaged colonial cohort. With gender, distinctions are only
observed in the Postclassic sample, when males appear to be slightly delayed in initial
food supplementation compared to females, reflecting greater maternal investment in
boys. This gender-based childcare may have had developmental consequences for
later health and survival, but it appears that the delay in boys was brief and was
subsequently compensated by a faster rate of food supplementation. Hard tissue
evidence of health, such as bone and dental pathology, portrays gender equality in the
health and survival of Lamanai males and females.
Methodologically, laser ablation sampling combined with ICP-MS is proven to be a
highly sensitive method of quantifying hard tissue elements across developmental age. It
enables the most detailed hard tissue reconstruction of dietary fluctuations over time.
Most significantly, it indicates that the weaning process is more variable and complex
than previously recognized by other methods.
Recognition of fluctuating Sr/Ca values over an extended developmental period
highlights the microsampling advantages of LA-ICP-MS. While other studies can
compare element values between cohorts of one to two years (e.g., Grupe 1986; Huhne-
Osterloh and Grupe 1989; Mays 2003; Sillen and Smith 1984), these results reflect a

302
level of intra-tooth variation and complexity that is not captured by macrosampling
techniques.
For bioarchaeological purposes, LA-ICP-MS offers a method of sampling that
broadens the questions that can be asked of human remains. For example, laser
microsampling can disclose greater variation in hard tissue composition across 1)
different tissues such as enamel and dentin (Haverkort 2003; Kang 2004); 2) different
teeth of comparable developmental age (Haverkort 2003); 3) pre-natal vs. post-natal
deciduous enamel (Goodman et al. 2003); 4) normal vs. defective enamel (enamel
hypoplasias or Wilson bands); and 5) surface vs. interior enamel or bone (for pollution
exposure and diagenetic diagnoses) (Budd et al., 1998; Cox et al. 1996; Montgomery et
al. 1998). Importantly, these investigations will enhance the understanding of childhood
nutrition and the nature of hard tissue incorporation under healthy, as well as less than
ideal, conditions.
As Outridge and colleagues (1995: 167) have stated:
LA-ICP-MS and other microprobe techniques are thus revealing that the
heterogeneity of metal distribution at microspatial scales is no less of a sampling
design problem than for macroscale entities such as forests, oceans, etc., and entails
similar types of statistical considerations, data management, and visual
representation.
Indeed, the data in this study have proven to be amenable to the same analytical
procedures as macrosampling techniques. While daunting at first, the ability to quantify
element ions at a micro scale provides a large reservoir of data that can be managed (i.e.,
reduced) in countless ways to suit the research question.

303
An important consequence of this study concerns the nutritional relevance of the
existing definition of weaning, i.e., the cessation of breast milk intake. Previously, as
noted by Stuart-Macadam (1995: 82), studies of ancient weaning were limited by a lack
of “contemporary longitudinal data on infants who have been breastfed in the ‘ancient
pattern’, that is, for longer than one year and with gradual supplementation of additional
foods.” This situation is changing, however, as women in both western and non-western
societies increasingly adopt the “traditional” pattern of long-term breastfeeding.
Buckley’s (2001) report on such mothers highlights the potential clinical data that
will clarify the ancient pattern. From her observations, it appears that breastfeeding after
the second year represents an insignificant nutritional contribution. Its primary role
thereafter is for emotional bonding and mother-child socialization. In ancient
populations, breast milk may also represent an important source of water (see Wright and
Schwarcz 1998), but its nutritional role is minor after the first year.
For all intents and purposes, children nursing beyond two years of age should be
considered “weaned” at approximately two years, when nursing tends to diminish
significantly. In fact, this is the age when many ancient and contemporary children stop
breastfeeding. In societies that advocate nursing for up to four or five years, like the
Maya, it is nutritionally inconsequential to stress a late weaning age because, by then,
children have already been primarily dependent on an adult diet for a few years.
Furthermore, attributing a late weaning age of four to five years to ancient populations
like the Maya is not a novel concept. Ethnographic accounts already affirm this.
Based on this study, it is perhaps more meaningful to track the age of significant food
supplementation and dietary adaptations to external stress, rather than the age of breast

304
milk cessation. In children breastfed for many years, health status and survival appear to
be better associated with the age of initial food consumption. Fortunately, enamel Sr/Ca
reflects the supplementation of solid food more confidently than the end of nursing
(weaning), and the patterns observed here have enriched the understanding of ancient
infant subsistence. Most importantly, the detailed and age-specific data provided by LA-
ICP-MS analysis of enamel documents a complex process leading to weaning. It
indicates that the process is highly sensitive to many external influences, which are
ultimately tied to interrelated ecological, political and economic factors.
In sum, the present research represents an evolution of the application of analytical
chemistry to paleodietary reconstruction. Several factors make this study exciting: 1) it
utilizes dental enamel, which is the least susceptible to post-mortem deterioration, and
thus, is more accurately reflective of intake and incorporation; 2) examination of
sequentially calcified, inert, dental tissue provides an indelible, detailed, history of intake
and nutritional status that can be captured by the localized sampling capabilities of laser
ablation; 3) employing LA-ICP-MS, it asks new, nutritional, questions of the dental
tissue analyzed by such technology. Sampling of permanent canine enamel, in particular,
allows reconstruction of dietary intake for the entire weaning process, which is a critical
period of childhood; 4) it applies minimally invasive microsampling technology to
multielement analysis that preserves the integrity of ancient remains; and 5) the Lamanai
sample presents a unique window into the dynamic processes that occurred at the
crossroads of Maya-Spanish contact. Above all, assessing the nature of infant diet by
LA-ICP-MS has provided invaluable inferences for the colonial consequences to Maya
childhood health and nutrition, an area that has been inadequately explored.

305
APPENDIX A
DENTAL HEALTH OF LAMANAI CANINES
This chapter includes macroscopic data for all canines used in this analysis. While
the sampling of only one tooth per individual precludes conclusions on dental health, raw
observations are provided as “osteobiographic” information for each individual. Such
reference data (e.g., defect measurements) might assist future research on dental defects
and enamel timing. For additional dental data, one is referred to more complete analyses
done by White (1986) on whole dentitions of Lamanai individuals.
A.1 Dental Attrition, Caries and Calculus
All sixty permanent canines used in this analysis were assessed for general indicators
of age and dental health (see Table A.3). This entailed the extent of dental attrition,
caries, calculus deposition and macroscopic enamel defects (hypoplasia and
hypocalcification).
Dental attrition was assessed based on a comparative scale, with teeth being judged as
complete (no dental wear) (“0”), slight (“1”), moderate (“2”), moderate-heavy (“3”) and
severe (“4”) in wear. Slight attrition was comparable to Brothwell's (1981: Fig. 3.9)
Stages 1 to 3. Moderately worn teeth had some cuspal enamel missing (up to half), with
some inner dentine exposed, and followed Brothwell's (1981: Fig. 3.9) stages 3+ and 4.
Moderate-heavy wear was characterized as those teeth with almost all cuspal enamel
absent and dentine exposed (with about half the complete crown height) (see Brothwell

306
1981: Fig. 3.9, stages 4+ and 5), while severely worn teeth were those with almost the
entire crown attritioned, sometimes down to the level of the root (see Brothwell 1981:
Fig. 3.9, stages 5+ to 6).
The canines were measured for complete crown height with sharpened electronic
calipers calibrated to 0.01 mm. Average unworn crown heights, specific to the Lamanai
population, are required for estimating the timing of enamel defects (see Table A.1
below). Crown heights of slightly worn teeth were also recorded to assess the degree of
missing enamel and the developmental period not recorded by such teeth (and not
sampled by laser ablation). Due to the requirement for intact, or mostly complete, teeth
for this study, samples with more than moderate-heavy wear were excluded. Obviously,
this biases the sample toward younger adults and children older than 5-6 years of age (the
age of permanent canine crown completion).
Carious lesions represent the progressive destruction of oral hard tissues by bacterial
activity and are assessed based on the number of carious lesions per tooth, as well as
lesion dimensions and tooth location. In ancient Maya samples, carious lesions
commonly afflict adult dentitions due to a high carbohydrate (maize) diet. Due to the
broad, fissured, surface of multi-cuspal teeth, premolars and molars will be the most
frequent site of carious activity. Conversely, single cusp teeth such as incisors and
canines are more rarely affected. Indeed, in this case, none of the utilized Lamanai
canines exhibit evidence of caries. Other reasons for this include the lack of older adults,
who tend to have more carious teeth, as well as the small sample size.
Finally, calculus deposits, or mineralized bacterial plaque containing calcium and
food debris, were rated following Brothwell (1981: Fig. 6.14b), with gradients of none

307
(0), slight (1), moderate (2), heavy (3) and very heavy (4). Generally, extensive calculus
deposits characterize adult dentitions, particularly older individuals. In the Maya area, a
high starchy maize diet means that calculus deposits often obscure the cervical margin
(crown base) of teeth. However, for this study, teeth with heavy or very heavy levels of
calculus were not included, either because of their permanent fixture on enamel surfaces,
and/or their association with heavily attritioned teeth. Canines used in this study are
either absent of calculus or have slight deposits.
A.2 Developmental Defects of Enamel
Macroscopic developmental defects of enamel are one of the most common
pathologies observed in archaeological populations and they include enamel hypoplasia
and enamel hypocalcification. Many researchers have examined enamel hypoplasia to
assess the health status of prehistoric and contemporary populations, particularly during
the weaning period, when the secondary dentition can be affected (see Chapter 5.4).
As a general term, enamel hypoplasia refers to all defects in enamel thickness, and is
thus, a quantitative defect. The enamel is usually deficient in quantity, but it may be fully
mineralized (Beynon 1986). On the other hand, hypocalcifications are simply areas of
opaque or discolored enamel that are smooth and normal in enamel thickness, except in
cases when they are combined with hypoplasia (FDI 1982). They are thought to be the
result of less severe stress, when compared to hypoplastic pits and bands (Blakey 1981).
Despite being the hardest substance in the body, with a rank of 3 on the Moh’s scale,
as well as the densest calcified tissue (about 3gm/cc) (Weatherell et al. 1967), enamel can
be subject to disturbances during its formation that alter its morphology and the physical

308
appearance of the crown. Enamel hypoplasia is the end result of disruptions in the
secretory/matrix formation phase of amelogenesis (Guita 1984; Shafer et al. 1983;
Yaeger 1980). Defective enamel can appear as 1) shallow or deep pits, or rows of pits,
arranged horizontally in a linear fashion across the tooth surface, or generally distributed
over the enamel surface; 2) shallow or deep, wide or narrow grooves or bands; or 3) as
regions of entirely missing enamel over small or large areas of dentine (FDI 1982: 160).
Three distinct factors contribute to the epidemiology of enamel defects. They include
hereditary, traumatic and systemic conditions. Generally, with archaeological samples,
the principal cause of enamel hypoplasia will be systemic factors (Goodman and Rose
1990; Goodman et al. 1984; Rose et al. 1985). Such defects are likely to be found on a
variety of teeth developing at the time of the stress, and their location on the crown
reflects the relative completeness of the enamel at the time of the disruption (Goodman et
al. 1980; Sarnat and Schour 1941; Pindborg 1982; Yaeger 1980).
Systemic conditions which can result in disturbed enamel formation include
malnutrition, infectious disease, exanthematous fevers, hypovitaminosis A, C and D,
hormone imbalance, fluorosis, neonatal tetany, hemolytic disease and prematurity, among
many others (Cutress and Suckling 1982; Beynon 1986; Blakey 1981; Moller 1982;
Newton et al. 1984; Purvis et al. 1973; Sweeney and Guzman 1966; Sweeney et al. 1969;
TenCate 1980).
Importantly, macroscopic enamel defects are relatively sensitive to growth
disturbances, and are nonspecific indicators of stress (Nikiforuk and Fraser 1981). It
should also be stressed that enamel hypoplasias indicate periods of stress that were
obviously followed by renewed enamel formation and recovery, representing an

309
individual’s survival of the stress. Populations experiencing severe, chronic malnutrition
and/or infections may not show any enamel defects due to a lack of recovery.
A.3 Scoring of Enamel Defects
Bioarchaeologists, dental anthropologists and dentists usually refer to either the
Federation Dentaire Internationale system (FDI 1982) or the Standards for Data
Collection from Human Skeletal Remains (Buikstra and Ubelaker 1994: Attachment 18a,
b) for recording developmental defects of enamel. These standards outline scoring
procedures for hypoplasias, opacities, combinations of both, and discolored enamel
(hypocalcifications).
In this study, hypocalcifications are described according to color and crown
distribution. Enamel discoloration, whether due to intrinsic or extrinsic factors, was
commonly found in the cervical region, i.e., accompanying calculus buildup. Calculus
staining therefore left the cause of enamel pigmentation in question, and the exact nature
of some discolorations cannot be identified. In truth, hypocalcifications are often not
scored in dental analyses as they frequently resemble extrinsic staining from either pre-
mortem (calculus), or post-mortem factors, such as the soil in the burial environment.
For measurements of enamel hypoplasia, the same sharpened electronic caliper was
utilized. The enamel defect was measured from the cemento-enamel junction to the
middle of the defect, if it was narrow, and to the superior and inferior borders if it was
broad. Thus, the entire period of defective enamel formation was recorded. At all stages,
enamel surfaces were visually examined under regular incandescent lighting, often
enhanced with natural light, and viewed with the unaided eye. Instances where defect

310
severity was questionable also involved the use of a hand-held magnifying glass.
However, consistent with Goodman and Rose (1990), if the defect could not be detected
without magnification, it was considered absent.
The present analysis varies slightly from the abovementioned standards (Buikstra and
Ubelaker 1994; FDI 1982), but it is closely comparable in the types of metric and
descriptive data that are outlined (if not more extensive in the recognition of defect
variations). Each tooth was assessed as descriptively as possible, making particular note
of the nature of defect types (shallow lines, grooves, bands, pits, patches of missing
enamel), location, color distinctions, and their measurements.
Figure A.1 Standards of enamel hypoplasia severity
(illustration of the labial surface of incisors)
According to Figure A.1, which follows the scoring of defects for Altun Ha (Song
1997), teeth are divided into two main categories, depending on defect width, or duration
of stress episode. “A” defects are narrow, or shorter in duration, and range from shallow
(1) to deep (3). “B” defects are broad, representing chronic stress, and are similarly

311
divided by depth (1-shallow to 3-deep). These degrees of defect depth (1, 2, 3) are
specified in Table A.3 (see below). Most defects observed in this sample are narrow, so
they are not specified in Table A.3, but in cases where the margins of a (wider) defect
could be measured, both distances from the CEJ are indicated. Multiple defects in a
single tooth are differentiated as “a”, “b” or “c”.
A.4 Estimation of Dental Defect Timing
In any study of dental material, especially one identifying a transition such as
weaning, an important consideration is the timing of enamel formation. While human
dental development is generally well understood, it is based on the primary assumption
that it is a regular or linear process. As a result, the standards outlined by Massler,
Schour and Poncher (1941) continue to form the bases of dental research involving
development time, as well as the determination of ancient subadult age (see review in
Goodman and Song 1999).
Current research, however, suggests that enamel development is far more complex.
Extensive findings indicate that enamel secretion is not linear across the entire period of
development, with a noticeably reduced rate of secretion at the cervical margin of
crowns, i.e., near the end of crown formation (see Beynon et al. 1998; Reid et al. 1998;
Reid and Dean 2000). Additionally, the delay between enamel secretion and maturation
(mineralization) is not fully understood, especially the extent of intratooth variation in
mineralization rates (L. Humphrey, pers. communication, 2004). For instance, the final
stages of enamel secretion, near the CEJ, may take several months to fully mineralize due
to a prolonged maturation process (Beynon et al. 1998; Reid and Dean 2000).

312
Even though Massler and coworkers’ (1941) ages of crown initiation and completion
(total duration) generally agree with (European) histological data (Liversidge 1994; Reid
and Dean 2000), deceleration of enamel formation in the CEJ third means that the period
between cusp tip and CEJ third will be truncated compared to their standards. For
enamel defects, which commonly occur in the mid-crown or cervical third region, the
outcome will be earlier ages of enamel and enamel defect formation (also see below).
The non-linear rate of enamel formation may represent the greatest source of
variation in standards of enamel development. However, as the extent of such variation
is not completely understood, and because of methodological issues and research
comparability, enamel development continues to be regarded as a linear process, with the
standards of Massler, Schour and Poncher (1941) being the primary reference data.
Other sources of variation in the estimation of enamel formation time include
population and sex differences, buried cuspal enamel and the choice of developmental
standard (see Goodman and Song 1999). Of these, “buried” or appositional enamel can
account for significant differences in the age of permanent enamel and (macroscopic)
enamel defect formation. This enamel represents the initial period of crown formation,
when Striae of Retzius form partial circles that begin and end in the inner enamel
(Goodman and Rose 1990: Fig. 1). Significant and observable time lapses exist between
this initial matrix formation and the appearance of Striae of Retzius on the outer enamel
surface, when they are visible macroscopically. Histological evidence suggests extensive
variation (e.g., Bullion 1987; Dean and Beynon 1991; FitzGerald 1995), but it is
reasonable to assume that little if any visible surface enamel develops before the end of
the first year in anterior permanent teeth, including canines (Reid and Dean 2000).

313
Additionally, timing of enamel development in this study accounts for population
differences in permanent canine crown completion. Numerous studies suggest that
permanent canines of North American indigenous populations complete crown formation
earlier than the white European standards of Massler and colleagues, i.e., 4.5 years
instead of 6.0 and 6.5 years for maxillary and mandibular canines, respectively (see
Anderson et al. 1976; Fanning and Brown 1971; Ubelaker 1989). Wright (1994) and
Song (1997) have applied this earlier age of canine completion to ancient Maya teeth,
which has resulted in slightly earlier mean ages of defect formation. However, as the
evidence has not been substantiated histologically, and 4.5 years is thought to be rather
early for permanent canines (A. Goodman, pers. communication, 2003), in this analysis,
they are considered complete at 5.0 years for maxillary teeth and 5.5 years for mandibular
teeth.
Regarding gender, some evidence indicates that females are advanced compared to
males in dental development (Demirjian 1986; Demirjian and Levesque 1980; Moorrees
et al. 1963). However, as noted by Smith (1991: 165), the patterned differences in rates
of enamel formation (population and sex-related) “may not be large”, i.e., 2-3%
(Demirjian and Levesque 1980; Garn et al. 1958; Gleiser and Hunt 1955).
The following regression formulae (Table A.1) are employed to arrive at the age at
which enamel defects developed in Lamanai canines. The equations inherently imply
that enamel growth is linear, and in this study, they account for 1) earlier canine crown
completion age for aboriginal populations; 2) one year of buried cuspal enamel; and
3) sample-specific mean unworn crown heights, which are 0.96 to 1.4 mm longer than the
Swedish standards (Swardstedt 1966) often used.
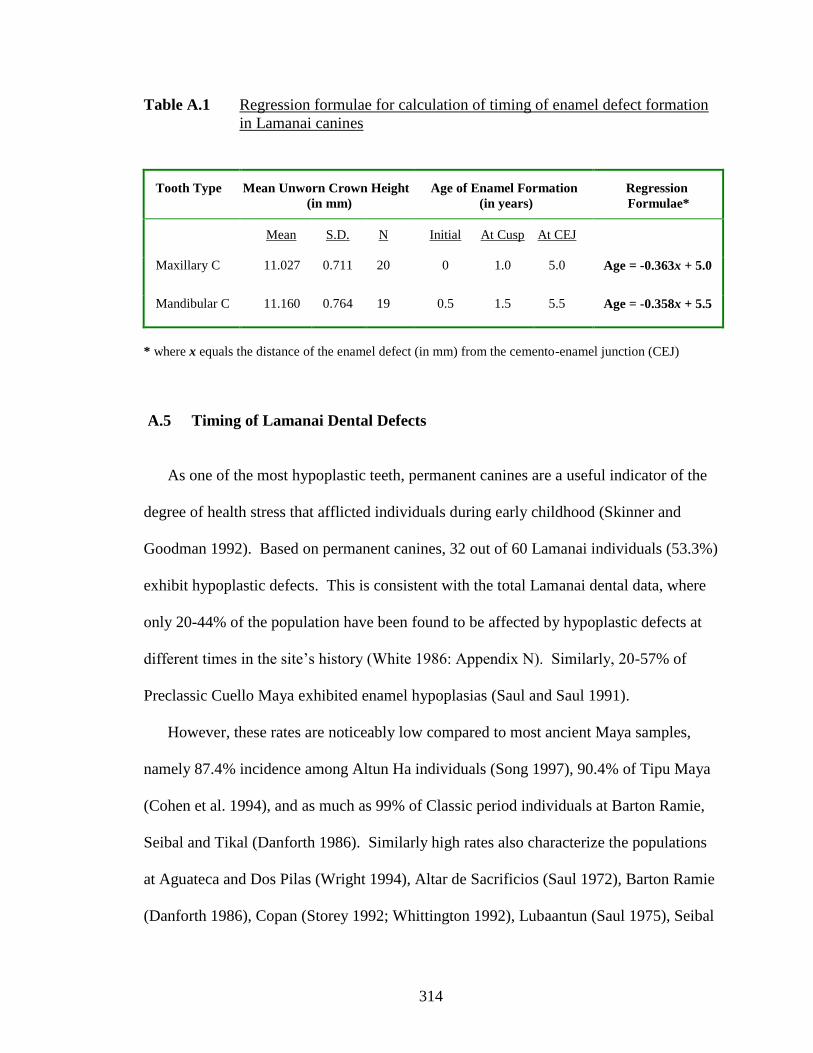
314
Table A.1 Regression formulae for calculation of timing of enamel defect formation
in Lamanai canines
Tooth Type
Mean Unworn Crown Height
(in mm)
Age of Enamel Formation
(in years)
Regression
Formulae*
Mean
S.D.
N
Initial
At Cusp
At CEJ
Maxillary C 11.027 0.711 20 0 1.0 5.0 Age = -0.363x + 5.0
Mandibular C 11.160 0.764 19 0.5 1.5 5.5 Age = -0.358x + 5.5
* where x equals the distance of the enamel defect (in mm) from the cemento-enamel junction (CEJ)
A.5 Timing of Lamanai Dental Defects
As one of the most hypoplastic teeth, permanent canines are a useful indicator of the
degree of health stress that afflicted individuals during early childhood (Skinner and
Goodman 1992). Based on permanent canines, 32 out of 60 Lamanai individuals (53.3%)
exhibit hypoplastic defects. This is consistent with the total Lamanai dental data, where
only 20-44% of the population have been found to be affected by hypoplastic defects at
different times in the site’s history (White 1986: Appendix N). Similarly, 20-57% of
Preclassic Cuello Maya exhibited enamel hypoplasias (Saul and Saul 1991).
However, these rates are noticeably low compared to most ancient Maya samples,
namely 87.4% incidence among Altun Ha individuals (Song 1997), 90.4% of Tipu Maya
(Cohen et al. 1994), and as much as 99% of Classic period individuals at Barton Ramie,
Seibal and Tikal (Danforth 1986). Similarly high rates also characterize the populations
at Aguateca and Dos Pilas (Wright 1994), Altar de Sacrificios (Saul 1972), Barton Ramie
(Danforth 1986), Copan (Storey 1992; Whittington 1992), Lubaantun (Saul 1975), Seibal

315
(Danforth 1986), Tancah (Saul 1982) and Tikal (Danforth 1986). For all samples,
including Lamanai, most hypoplasias are mild or moderate in severity.
On an individual level, 15 of 26 Postclassic individuals have a canine with enamel
defects (57.7%), which is comparable to the Historical sample: 17 of 34 individuals
(50%). Besides temporal correspondence, enamel hypoplasias also do not differ
significantly between Lamanai males and females and between children and adults
(White 1986; Wright 1990). Like comparisons with other Maya samples, the 50%
incidence among Historical Lamanai individuals is significantly lower than the 89% rate
of enamel hypoplasia among enslaved African Americans, who are one of the most
hypoplastic colonial populations (Blakey et al. 1994).
The incidence of longer-term, wide, enamel defects is also similar for both Lamanai
cohorts: 2 out of 15 Postclassic individuals and 3 out of 17 Historical individuals have a
single period of chronic stress (defect wider than 0.4 mm). These long-term stress
episodes (i.e., 2 months or more) occur at all ages of enamel development, but the
predominant period of broad defect formation is 4-4.5 years of age.
In terms of multiple defects, however, there is a significant temporal distinction.
While 1 out of 15 (6.7%) Postclassic individuals has more than one enamel defect, 6 out
of 17 (35.3%) Historical individuals with hypoplasia have multiple defects. A two
sample t-test indicates significance at the 0.05 level (p=0.047). This temporal distinction
is not unexpected considering the epidemiological context of the colonial period (see
Chapter Two).
Among the seven individuals who experienced multiple episodes of childhood stress,
three had two distinct episodes (42.9 %) and four had three periods of ill health (57.1%)

316
(also see Table A.3) . These seven recurrently stressed individuals are composed of 5
adults (out of 24), or 20.8% of all adults with enamel hypoplasia, and 2 children (out of
8), or 25% of all affected subadults, which are comparatively equal. Thus, it would seem
that experiencing multiple stress episodes did not necessarily determine whether an
individual died as a child, or survived into adulthood.
Nevertheless, with multi-stressed individuals only comprising 21.9% (7/32) of the
total sample of hypoplastic individuals (or 11.7% of the total sample examined in this
analysis), it appears that recurrent health or dietary insults were fairly uncommon in
Lamanai children. In comparison, Altun Ha Maya with multiple enamel defects make up
42.5% of the total number of hypoplastic individuals at the site (Song 1997).
With respect to age distribution, Figures A.2 and A.3 illustrate the unimodal
distribution of defects for each half-year age interval among Postclassic and Historical
individuals. All defect timings are represented, including multiple defects occurring in
single individuals. In individuals with an episode of long-term stress, only the initial 0.5
year interval (age at onset) was counted.
It is clear from the graphs that the vast majority of defects developed between the ages
of three to five years. This is echoed in the entire Lamanai dental sample (White 1986).
In terms of crown distribution, the majority of defects occur in the region of the mid-
crown to cervical third. The pattern of peak enamel defects between three and five years
of age is a consistent finding in lowland Maya samples from the Preclassic to Historical
period, namely at nearby Altun Ha (Song 1997), Aguateca and Dos Pilas (Wright 1994),
Altar de Sacrificios (Saul 1972), Barton Ramie, Seibal and Tikal (Danforth 1986), Cuello
(Saul and Saul 1991), Tancah (Saul 1982), and Tipu (Cohen et al. 1994).

317
Figure A.2 Distribution of defect timings in Lamanai maxillary canines
(5 Postclassic with 7 defects; 7 Historical with 14 defects)
Figure A.3 Distribution of defect timings in Lamanai mandibular canines
(10 Postclassic with 10 defects; 10 Historical with 12 defects)
Distribution of Enamel Defect Timings in Upper Canines
0
1
2
3
4
5
6
7
8
9
2.5-2.99 3.0-3.49 3.5-3.99 4.0-4.49 4.5-4.99
Age Category
Num
ber
of
Defe
cts
Postclassic
Historical
Distribution of Enamel Defect Timings in Lower Canines
0
1
2
3
4
5
6
7
8
9
2.5-2.99 3.0-3.49 3.5-3.99 4.0-4.49 4.5-4.99
Age Category
Num
ber
of
Defe
cts
Postclassic
Historical

318
Table A.2 outlines the mean age of defect formation for permanent maxillary and
mandibular canines. It appears that enamel defects recording health or dietary stress
occur most often between the ages of 3.5 and 4.5 years.
Table A.2 Average ages of enamel defect formation in the Lamanai sample
(including teeth with multiple episodes)
Tooth
Mean Age
S.D.
No. of Defects
Postclassic
Max. C 3.83 0.64 7
Mand. C 4.31 0.25 10
Historical
Max. C 3.68 0.35 14
Mand. C 4.27 0.27 12
The table and graphical data clearly reflect the significant difference in mean ages of
enamel defect formation between maxillary and mandibular canines (ANOVA result of
p=0.000012). This is due to differences in the age at which canine enamel is attributed to
commence and complete development, which is 0.5 years later in mandibular teeth
(Massler et al. 1941). Despite the slight delay, one expects defect timing between the
mandibular and maxillary teeth to be comparable. Dissimilar mean values suggests that
the existing understanding of canine enamel formation, with mandibular canines
developing at the same rate and duration 0.5 years later than maxillary teeth, should be
reconsidered.

319
Finally, as mentioned previously, enamel hypoplastic defects are valuable for gauging
the physiological consequences of infant weaning. Assuming that weaning stress can be
recognized in enamel defects, Skinner and Goodman (1992) suggest that the age of first
defect formation, which is based on Corruccini and coworkers (1985), would be the most
appropriate indicator. Specifically, if lactation provides health buffering from stress, one
would expect a narrow distribution of ages at the first occurrence of hypoplasia, which
might indicate a “culturally sanctioned age at weaning” (Skinner and Goodman 1992:
169).
Ages of first defect formation lower the mean ages of total defect timing outlined in
Table A.2. In this case, timing of maxillary canine defects average 3.61 years
(S.D.=0.70) in Postclassic individuals and 3.56 years (S.D.=0.40) in Historical
individuals, while mandibular defects develop at a mean age of 4.29 years (S.D.=0.27) in
Postclassic individuals and 4.18 years (S.D.=0.27) in Historical individuals. Although
the sample size is limited, these ages correspond well with the late peaking of enamel
Sr/Ca observed in this study, in addition to ethnohistorical evidence of weaning.
A.6 Dental Health Raw Data
Table A.3 includes all the macroscopic dental data that was collected in this study for
the sixty ablated Lamanai canines, which was used to assess general dental “health”.
These details include crown height, dental attrition, caries, degree of calculus deposits,
measurement of enamel hypoplasias, age of enamel defect formation and additional
comments. Scoring criteria are detailed in each heading, with CEJ referring to the
cemento-enamel junction.

320
Table A.3 Dental health details for Lamanai canines
Individual # Sex/Age Tooth Type Crown
Height
Dental
Attrition Caries Calculus
Enamel Defect
Measurement
Age of Defect
Formation Comments
N10-2 = Postclassic
N10-4 = Postclassic
YDL-85 = Historical
F=female
M=male
Ad.=adult
SubAd.=
subadult (ages in years)
Up=upper
Lo=lower
R=right
L=left
Measured
from cusp tip
to cemento
enamel
junction (CEJ)
in mm
0=none
1=slight
2=moderate
3=moderate-
heavy 4=severe
0=none
1-3 =
number of
carious
lesions
0=none
1=slight
2=moderate
3=heavy
4=very heavy
distance in mm
from middle of
defect to CEJ,
1-3 = defect severity
(see Song 1997) a-c=separate
defect occurrences
in years,
based
on Regression
Formulae
(Table A.1)
H=hypocalcification
(W-white, B-brown,
O-orange, C-cream)
LAM N10-2/4 F 14-18 Lo LC 9.63 2 0 1-2 2/3: 2.99 - 4.16 4.01-4.43 depression at CEJ 1/3
LAM N10-2/5 M Ad. Lo RC 9.65 1-2 0 0 0 0 crack across lingual enam / root
LAM N10-2/16 SubAd. Up LC 11.63 0 0 0 0 0 White scratched areas
LAM N10-2/20 M Ad. Lo RC 9.60 2 0 1-2 at CEJ approx. 2.21 or CEJ
1/4 4.71 Rough patch; elite individ.
LAM N10-2/21 10-12 Up RC 11.12 0 0 0 0 0 some H-OB stains
LAM N10-2/22 M Ad. Lo LC 10.56 2 0 2 1 shallow LEH at
CEJ 1/4-1/3 approx. 4.17 small vertical crack
LAM N10-2/40 Ad. M? Up RC 11.89 1-2 0 1-2 0 0 vertical midline crack
LAM N10-2/42 M 20-30 Lo RC 11.53 1-2 0 1-2 2: 3.95 4.09 -
LAM N10-2/44 7-8 Up LC 11.01 0 0 0 pit- mid is 3.80 3.62 vertical LEH coming from
pit- ends at 7.43
LAM N10-2/49 10-11 Lo RC 11.30 1 0 0 0 0 glued at CEJ 1/3 line; brown
staining all over (H?)
Continued, next page

321
Table A.3, continued
Individual # Sex/Age Tooth Type Crown
Height
Dental
Attrition Caries Calculus
Enamel Defect
Measurement
Age of Defect
Formation Comments
N10-2 = Postclassic
N10-4 = Postclassic
YDL-85 = Historical
F=female
M=male
Ad.=adult
SubAd.=
subadult (ages in years)
Up=upper
Lo=lower
R=right
L=left
Measured
from cusp tip
to cemento
enamel
junction (CEJ)
in mm
0=none
1=slight
2=moderate
3=moderate-
heavy 4=severe
0=none
1-3 =
number of
carious
lesions
0=none
1=slight
2=moderate
3=heavy
4=very heavy
distance in mm
from middle of
defect to CEJ,
1-3 = defect severity
(see Song 1997) a-c=separate
defect occurrences
in years,
based
on Regression
Formulae
(Table A.1)
H=hypocalcification
(W-white, B-brown,
O-orange, C-cream)
LAM N10-2/50 6-8 Up RC 11.36 0-1 0 0 a- 1: 1.37, b- 2: 2.40,
c- 2: 3.58 3.70; 4.13; 4.50 1/3 root formation
LAM N10-4/1 F >35 Up RC 10.42 1-2 0 2-3 on medial
side 1: 3.46 3.74
vertical crack at midline, H-
white
LAM N10-4/2A 14-18 Up LC 11.54 1 0 3 at CEJ 0 0 vertical crack at midline
LAM N10-4/2B F >40 Lo RC 9.96 2-3 0 2-3 2: 4.56 3.87 glued down midline crack
LAM N10-4/4 F >50 Lo LC 9.50 0 0 0 0 0 H-B/W all over
LAM N10-4/10 F 15-20 Up LC 9.91 1-2 0 2 at CEJ 2: 1.45 4.47 notched on med side
LAM N10-4/11 F? 30+ Up LC 9.04 2 0 1 at CEJ 2/3: 6.27 to 6.85 2.51-2.72 large vertical crack at
mid-line
LAM N10-4/19 F Ad+ Lo RC 10.82 1-3 0 2-3 pit at cervical 1/4 approx. 4.50 -
LAM N10-4/33 F 30-40 Up LC 9.68 1 0 1 at CEJ 0 0 -
LAM N10-4/40 F 25-30 Lo LC 10.64 2 0 1-2 1: 3.35 4.30 vertical midline crack
LAM N10-4/43 M Ad+ Up LC 8.98 2-3 0 3 at CEJ 0 0 -
Continued, next page

322
Table A.3, continued
Individual # Sex/Age Tooth Type Crown
Height
Dental
Attrition Caries Calculus
Enamel Defect
Measurement
Age of Defect
Formation Comments
N10-2 = Postclassic
N10-4 = Postclassic
YDL-85 = Historical
F=female
M=male
Ad.=adult
SubAd.=
subadult (ages in years)
Up=upper
Lo=lower
R=right
L=left
Measured
from cusp tip
to cemento
enamel
junction (CEJ)
in mm
0=none
1=slight
2=moderate
3=moderate-
heavy 4=severe
0=none
1-3 =
number of
carious
lesions
0=none
1=slight
2=moderate
3=heavy
4=very heavy
distance in mm
from middle of
defect to CEJ,
1-3 = defect severity
(see Song 1997) a-c=separate
defect occurrences
in years,
based
on Regression
Formulae
(Table A.1)
H=hypocalcification
(W-white, B-brown,
O-orange, C-cream)
LAM N10-4/44 Ad. Lo RC 10.61 1-2 0 1 1?: 2.57 4.58 slight depressed region
cervical 1/3
LAM N10-4/45 F 30+ Lo LC 11.28 1 0 1-2 depression around
2.94 4.45 -
LAM N10-4/46A? M? Ad. Lo LC 12.06 1-2 0 2-3 0 0 long tooth; elite individ.
LAM N10-4/46B F Ad.+ Up LC 11.76 1-2 0 2 0 0 -
LAM N10-4/46C F Ad. Lo LC 11.22 0-1 0 2 1: 3.51 4.24 small tooth
LAM YDL-85/17 F 20-40 Up LC 10.48 1-2 0 1-2 at CEJ
a- 2: 2.72,
b- 1/2: 4.06,
c- 1: 5.17
3.12; 3.53; 4.01 -
LAM YDL-85/21 F 20-30 Up LC 9.52 1-2 0 1 a- 1: 3.67, b- 1: 4.35,
c- 1: 5.22 3.11; 3.42; 3.67
H-B/C on sides, some H-O on
labial surface
LAM YDL-85/23 F 20-30 Lo RC 11.21 2 0 1 at CEJ 2: 2.93 4.45 -
LAM YDL-85/27 F 20-30 Lo LC 9.09 2 0 2-3 0 0 vertical midline crack
LAM YDL-85/31 F? 20-25 Lo RC 10.17 2-3 0 2-3 1: 3.69 4.18 Depressed region? CEJ 1/3
Continued, next page

323
Table A.3, continued
Individual # Sex/Age Tooth Type Crown
Height
Dental
Attrition Caries Calculus
Enamel Defect
Measurement
Age of Defect
Formation Comments
N10-2 = Postclassic
N10-4 = Postclassic
YDL-85 = Historical
F=female
M=male
Ad.=adult
SubAd.=
subadult (ages in years)
Up=upper
Lo=lower
R=right
L=left
Measured
from cusp tip
to cemento
enamel
junction (CEJ)
in mm
0=none
1=slight
2=moderate
3=moderate-
heavy 4=severe
0=none
1-3 =
number of
carious
lesions
0=none
1=slight
2=moderate
3=heavy
4=very heavy
distance in mm
from middle of
defect to CEJ,
1-3 = defect severity
(see Song 1997) a-c=separate
defect occurrences
in years,
based
on Regression
Formulae
(Table A.1)
H=hypocalcification
(W-white, B-brown,
O-orange, C-cream)
LAM YDL-85/32 Ad. M? Up LC 11.02 1-2 0 2-3 2: 3.61 3.69 some enamel missing on
lingual
LAM YDL-85/33 M 25-30 Lo LC 10.85 2 0 2-3 1: ~3.13 4.38 depression CEJ 1/3 ?
LAM YDL-85/35 M 30-40 Lo LC 10.25 2 0 3 0 0 -
LAM YDL-85/41 F 18-25 Up RC 9.25 1-2 0 2 0 0 crack along med-lateral axis
from tip to root
LAM YDL-85/44 5-7 Up RC 11.05 0 0 0 0 0 H - O/B along CEJ
LAM YDL-85/46 F 20-30 Up LC 9.93 2 0 1 1?: 6.74 0 vertical midline crack
LAM YDL-85/47 M 12-16 Lo LC 11.03 1-2 0 1-2
2: 2.01 to 3.86
(rough region with
lines within)
4.12-4.78
rough depressed region across
most of CEJ 1/3 (vert mid
crack)
LAM YDL-85/50B F Ad. Up LC 11.04 1-2 0 1 1?: 6.18 0 2nd individual
LAM YDL-85/53 F 16-23 Lo RC 11.43 1 0 1 0 0 H-W/C in CEJ 1/3 region
Continued, next page

324
Table A.3, continued
Individual # Sex/Age Tooth Type Crown
Height
Dental
Attrition Caries Calculus
Enamel Defect
Measurement
Age of Defect
Formation Comments
N10-2 = Postclassic
N10-4 = Postclassic
YDL-85 = Historical
F=female
M=male
Ad.=adult
SubAd.=
subadult (ages in years)
Up=upper
Lo=lower
R=right
L=left
Measured
from cusp tip
to cemento
enamel
junction (CEJ)
in mm
0=none
1=slight
2=moderate
3=moderate-
heavy 4=severe
0=none
1-3 =
number of
carious
lesions
0=none
1=slight
2=moderate
3=heavy
4=very heavy
distance in mm
from middle of
defect to CEJ,
1-3 = defect severity
(see Song 1997) a-c=separate
defect occurrences
in years,
based
on Regression
Formulae
(Table A.1)
H=hypocalcification
(W-white, B-brown,
O-orange, C-cream)
LAM YDL-85/54 5-7 Up LC 11.85 0 0 0 1?: 4.27 0 crown only
LAM YDL-85/59 6-7 Lo LC 11.41 0 0 0 1: 4.85 3.76 crown only
LAM YDL-85/62 F 20-25 Up LC 9.50 1 0 1 0 0 crack along medial-lateral
axis thru to root tip
LAM YDL-85/63 6 Lo RC 11.86 0 0 0 1: 3.91 line, (border
of CEJ 1/3 rough) 4.10 H-B/W in CEJ 1/3
LAM YDL-85/65 6-7 Up RC 10.40 0 0 0 0 0 H-B along CEJ
LAM YDL-85/68b 8-12 Up RC 11.12 1 0 1 pit/rough patch: 2.44 4.11 small midline crack
LAM YDL-I-68? M 20-30 Up LC 9.49 1-2 0 1 0 0 small midline crack,
glued in several places
LAM YDL-85/71A 7-8 Lo LC 10.92 0 0 0 a- 2: 2.74
b- 2: 4.02 4.06; 4.52 H-B(?) along CEJ
LAM YDL-85/73 M? 20-25 Lo LC 10.43 1-2 0 2 0 0 -
Continued, next page

325
Table A.3, continued
Individual # Sex/Age Tooth Type Crown
Height
Dental
Attrition Caries Calculus
Enamel Defect
Measurement
Age of Defect
Formation Comments
N10-2 = Postclassic
N10-4 = Postclassic
YDL-85 = Historical
F=female
M=male
Ad.=adult
SubAd.=
subadult (ages in years)
Up=upper
Lo=lower
R=right
L=left
Measured
from cusp tip
to cemento
enamel
junction (CEJ)
in mm
0=none
1=slight
2=moderate
3=moderate-
heavy 4=severe
0=none
1-3 =
number of
carious
lesions
0=none
1=slight
2=moderate
3=heavy
4=very heavy
distance in mm
from middle of
defect to CEJ,
1-3 = defect severity
(see Song 1997) a-c=separate
defect occurrences
in years,
based
on Regression
Formulae
(Table A.1)
H=hypocalcification
(W-white, B-brown,
O-orange, C-cream)
LAM YDL-85/78A M Ad. Lo RC 11.86 1-2 0 2 a- 1: 2.99
b- 2: 4.45 3.91; 4.43
small midline crack, enamel
missing from part of labial,
mark root for cut
LAM YDL-85/78C F Ad. Lo RC 10.02 1-2 0 1 2: 2.24 4.70 rough area and H-OB at CEJ 1/3
region, small midline crack
LAM YDL-85/80 8-9 Lo LC 12.87 0 0 0 0 0 midline crack
LAM YDL-85/86 M? 20-25 Up LC 9.95 2 0 1 a- 2: 2.99
b- 2: 4.46 3.38; 3.91 -
LAM YDL-85/87B M 30-40 Up C 10.79 1 0 1-2 0 0 small midline crack
LAM YDL-85/91A 8-10 Lo LC 10.46 0 0 0
a- 2: 3.24,
b- 2 (pit area): 3.69
(is same pd.)
4.18-4.34
midline crack & some labial
enamel missing, plus vertical
LEH (?) along lat. labial in
midcrown region (1.70mm long)
Continued, next page

326
Table A.3, continued
Individual # Sex/Age Tooth Type Crown
Height
Dental
Attrition Caries Calculus
Enamel Defect
Measurement
Age of Defect
Formation Comments
N10-2 = Postclassic
N10-4 = Postclassic
YDL-85 = Historical
F=female
M=male
Ad.=adult
SubAd.=
subadult (ages in years)
Up=upper
Lo=lower
R=right
L=left
Measured
from cusp tip
to cemento
enamel
junction (CEJ)
in mm
0=none
1=slight
2=moderate
3=moderate-
heavy 4=severe
0=none
1-3 =
number of
carious
lesions
0=none
1=slight
2=moderate
3=heavy
4=very heavy
distance in mm
from middle of
defect to CEJ,
1-3 = defect severity
(see Song 1997) a-c=separate
defect occurrences
in years,
based
on Regression
Formulae
(Table A.1)
H=hypocalcification
(W-white, B-brown,
O-orange, C-cream)
LAM YDL-85/97 M 25-30 Up LC 10.23 2 0 1 a- 1: 2.26, b- 2: 3.11,
c- 2: 4.17 3.49; 3.87; 4.18
some enam missing on labial
near CEJ; small midline crack
LAM YDL-85/98 M Ad. Up LC 9.49 2 0 1 0 0 midline crack
LAM YDL-85/101 F Ad. Up RC 11.48 1 0 1-2 0 0
glued midline crack, portion of
tooth at ling CEJ gone, glued
here
LAM YDL-85/102 F 30-35 Lo LC 8.95 1-2 0 2-3 0 0 small tooth
LAM YDL-85/103 M 30-40 Up RC 11.40 2 0 1 1-bit wide at CEJ 1/4-
1/5
mid at approx.
4.00
some enam gone around medial
side of labial

327
APPENDIX B
ABLATION DETAILS FOR LAMANAI CANINES
Tables B.1 – B.4 outline the ablation details for all Lamanai teeth analyzed in this
study, which are based on photographic measurements (see Chapter Six). They are
grouped into maxillary and mandibular teeth for consistency of enamel developmental
period and the “ages” (in years) represented in each ablation. The specific details are
described below:
Ablation detail
Explanation
1ST LA
Length of first ablation in micrometers (cusp tip end)
2ND LA Length of second ablation in micrometers
3RD LA Length of third ablation in micrometers (CEJ end)
Total Ablation Length Total length of all ablations (along axis of crown)
Total # Readings Total number of ICP-MS element readings
Microns per reading Mean number of micrometers per element reading
CEJ enamel not ablated Length of enamel at CEJ end not ablated
"Missing" cusp enamel (extrap.) Length of enamel extrapolated to be missing from cusp
Crown Height Total (microns) Total height of crown in micrometers (along central axis)
Microns Per half year Mean number of micrometers per half year period
Start Age with Cuspal enam (0.5/1 yr) Age of start of 1st ablation, accounting for cuspal enamel
time that is not sampled (approx. 0.5 years)
End Age of 5/5.5 years End age of last sampled enamel (at end of last ablation)
Total years covered by ablations Total number of years sampled by all ablations
LA1 Reading #'s Element reading numbers of first ablation
Years detailed in LA1 Number of years represented in first ablation
LA2 Reading #'s Element reading numbers of second ablation
Years detailed in LA2 Number of years represented in second ablation
LA3 Reading #'s Element reading numbers of third ablation
Years detailed in LA3 Number of years represented in third ablation

Table B.1 Ablation details for Postclassic Lamanai maxillary canines
Individual # N10-2/16 N10-2/21 N10-2/40 N10-2/44 N10-2/50 N10-4/1
1ST LA (microns) 9290 5540 6480 10500 6120 7130
2ND LA (microns) 1750 3000 4470 1320 4060 2550
3RD LA (microns) - 1580 - - - -
Total Ablation Length 11040 10120 10950 11820 10180 9680
Total # Readings 795 799 863 842 810 734
Microns per reading 13.887 12.666 12.688 14.038 12.568 13.188
CEJ enamel not ablated 420 750 470 420 780 500
"Missing" cusp enamel (extrap.) 380 300 1240 320 700 860
Crown Height Total (microns) 11840 11170 12660 12560 11660 11040
Microns Per half year 1315.556 1241.111 1406.667 1395.556 1295.556 1226.667
Start Age with Cuspal enam (0.5yr) 0.644 0.621 0.941 0.615 0.770 0.851
End Age of 5 years 4.840 4.698 4.833 4.850 4.699 4.796
# Readings per half year: 94.734 97.989 110.863 99.413 103.084 93.014
Total years covered by ablations 4.196 4.077 3.892 4.235 3.929 3.946
LA1 Reading #'s 1-665 1-436 1-511 1-746 1-493 1-533
Years detailed in LA1 0.644-4.175 0.621-2.853 0.941-3.244 0.615-4.377 0.770-3.132 0.851-3.757
LA2 Reading #'s 666-795 437-679 512-863 747-842 494-810 534-734
Years detailed in LA2 4.176-4.840 2.854-4.062 3.245-4.833 4.378-4.850 3.133-4.699 3.758-4.796
LA3 Reading #'s - 680-799 - - - -
Years detailed in LA3 - 4.063-4.698 - - - -
Continued, next page
328

Table B.1, continued
Individual # N10-4/2A N10-4/10 N10-4/11 N10-4/33 N10-4/43 N10-4/46B
1ST LA (microns) 6330 7400 5700 8045 6010 8880
2ND LA (microns) 3170 1160 2240 2225 2870 -
3RD LA (microns) 1220 1700 - - - -
Total Ablation Length 10720 10260 7940 10270 8880 8880
Total # Readings 780 774 630 755 673 627
Microns per reading 13.744 13.256 12.603 13.603 13.195 14.163
CEJ enamel not ablated 140 180 300 250 200 3030
"Missing" cusp enamel (extrap.) 1260 805 560 340 1900 620
Crown Height Total (microns) 12120 11245 8800 10860 10980 12530
Microns Per half year 1346.667 1249.444 977.778 1206.667 1220.000 1392.222
Start Age with Cuspal enam (0.5yr) 0.968 0.822 0.786 0.641 1.279 0.723
End Age of 5 years 4.948 4.928 4.847 4.896 4.918 3.912
# Half year periods: 7.960 8.212 8.120 8.511 7.279 6.378
# Readings per half year: 97.985 94.256 77.582 88.708 92.462 98.302
Total years covered by ablations 3.980 4.106 4.060 4.256 3.639 3.189
LA1 Reading #'s 1-451 1-544 1-441 1-571 1-442 1-627
Years detailed in LA1 0.968-3.318 0.822-3.783 0.786-3.701 0.641-3.974 1.279-3.742 0.723-3.912
LA2 Reading #'s 452-687 545-637 442-630 572-755 443-673 -
Years detailed in LA2 3.319-4.496 3.784-4.248 3.702-4.847 3.975-4.896 3.743-4.918 -
LA3 Reading #'s 688-780 638-774 - - - -
Years detailed in LA3 4.497-4.948 4.249-4.928 - - - -
329

Table B.2 Ablation details for Postclassic Lamanai mandibular canines
Individual # N10-2/4 N10-2/5 N10-2/20 N10-2/22 N10-2/42 N10-2/49 N10-4/2B
1ST LA (microns) 5990 6750 8580 5410 6120 4780 2620
2ND LA (microns) 2760 2030 2540 2550 3080 5420 4805
3RD LA (microns) 500 - - 1300 1090 1200 2650
Total Ablation Length 9250 8780 11120 9260 10290 11400 10075
Total # Readings 698 713 828 760 773 859 776
Microns per reading 13.252 12.314 13.430 12.184 13.312 13.271 12.983
CEJ enamel not ablated 460 300 280 600 310 575 505
"Missing" cusp enamel (extrap.) 1400 480 420 1150 380 300 525
Crown Height Total (microns) 11110 9560 11820 11010 10980 12275 11105
Microns Per half year 1234.444 1062.222 1313.333 1223.333 1220.000 1363.889 1233.889
Start Age with Cuspal enam (1yr) 1.567 1.226 1.160 1.470 1.156 1.110 1.213
End Age of 5.5 years 5.314 5.359 5.393 5.255 5.373 5.289 5.295
# Half year periods: 7.493 8.266 8.467 7.569 8.434 8.358 8.165
# Readings per half year: 93.151 86.260 97.791 100.403 91.648 102.770 95.037
Total years covered by ablations 3.747 4.133 4.234 3.785 4.217 4.179 4.083
LA1 Reading #'s 1-453 1-541 1-634 1-451 1-454 1-365 1-206
Years detailed in LA1 1.567-3.993 1.226-4.403 1.160-4.426 1.470-3.681 1.156-3.664 1.110-2.862 1.213-2.274
LA2 Reading #'s 454-646 542-713 635-828 452-656 455-676 366-768 207-549
Years detailed in LA2 3.994-5.112 4.404-5.359 4.427-5.393 3.682-4.724 3.665-4.927 2.863-4.850 2.275-4.222
LA3 Reading #'s 647-698 - - 657-760 677-773 769-859 550-776
Years detailed in LA3 5.113-5.314 - - 4.725-5.255 4.928-5.373 4.851-5.289 4.223-5.295
Continued, next page
330

Table B.2, continued
Individual # N10-4/4 N10-4/19 N10-4/40 N10-4/44 N10-4/45 N10-4/46A N10-4/46C
1ST LA (microns) 3850 7340 4860 7000 5700 8150 5650
2ND LA (microns) 2610 2200 4540 3480 5000 3550 5870
3RD LA (microns) 0 - - 800 - - -
Total Ablation Length 6460 9540 9400 11280 10700 11700 11520
Total # Readings 481 702 772 800 832 808 830
Microns per reading 13.430 13.590 12.176 14.100 12.861 14.480 13.880
CEJ enamel not ablated 1440 1180 450 530 300 560 325
"Missing" cusp enamel (extrap.) 580 275 1200 200 300 230 260
Crown Height Total (microns) 8480 10995 11050 12010 11300 12490 12105
Microns Per half year 942.222 1221.667 1227.778 1334.444 1255.556 1387.778 1345.000
Start Age with Cuspal enam (1yr) 1.308 1.113 1.489 1.075 1.119 1.083 1.097
End Age of 5.5 years 4.736 5.017 5.317 5.301 5.381 5.298 5.379
# Half year periods: 6.856 7.809 7.656 8.453 8.522 8.431 8.565
# Readings per half year: 70.156 89.896 100.835 94.641 97.628 95.840 96.905
Total years covered by ablations 3.428 3.905 3.828 4.226 4.261 4.215 4.283
LA1 Reading #'s 1-286 1-542 1-370 1-500 1-448 1-560 1-425
Years detailed in LA1 1.308-3.351 1.113-4.117 1.489-3.468 1.075-3.698 1.119-3.389 1.083-4.019 1.097-3.197
LA2 Reading #'s 287-481 543-702 371-772 501-746 449-832 561-808 426-830
Years detailed in LA2 3.352-4.736 4.118-5.017 3.469-5.317 3.699-5.003 3.39-5.381 4.020-5.298 3.198-5.379
LA3 Reading #'s - - - 747-800 - - -
Years detailed in LA3 - - - 5.004-5.301 - - -
331
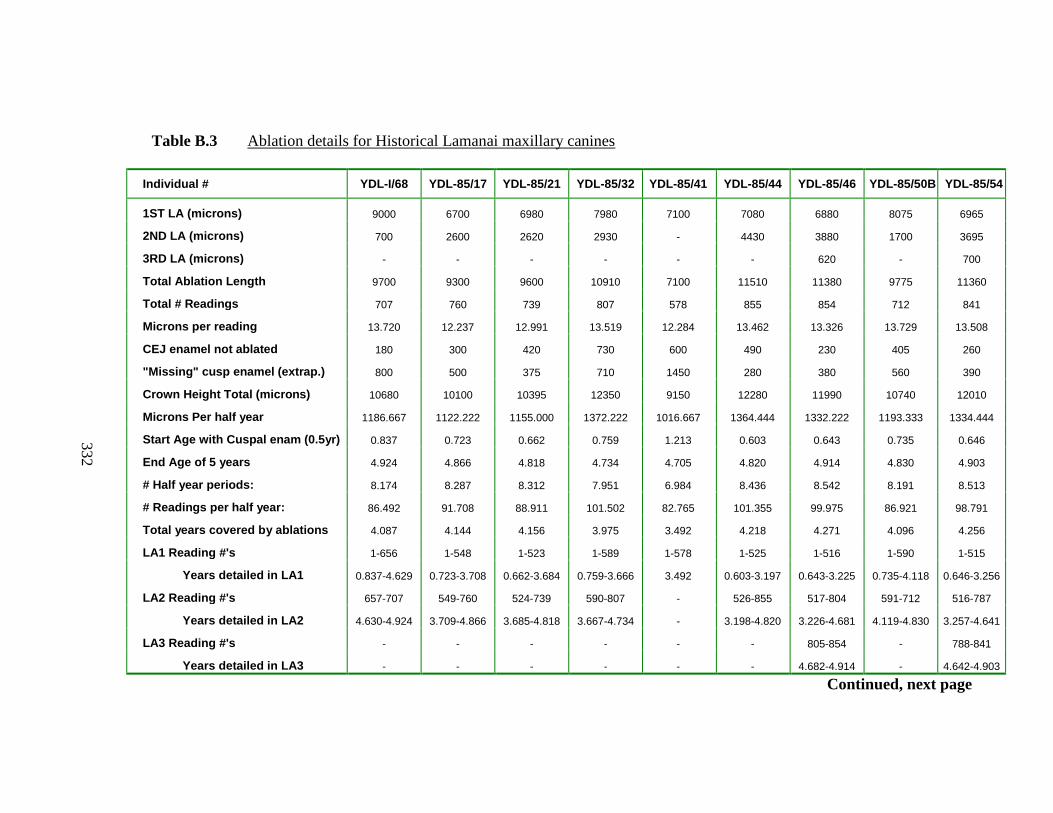
Table B.3 Ablation details for Historical Lamanai maxillary canines
Individual # YDL-I/68 YDL-85/17 YDL-85/21 YDL-85/32 YDL-85/41 YDL-85/44 YDL-85/46 YDL-85/50B YDL-85/54
1ST LA (microns) 9000 6700 6980 7980 7100 7080 6880 8075 6965
2ND LA (microns) 700 2600 2620 2930 - 4430 3880 1700 3695
3RD LA (microns) - - - - - - 620 - 700
Total Ablation Length 9700 9300 9600 10910 7100 11510 11380 9775 11360
Total # Readings 707 760 739 807 578 855 854 712 841
Microns per reading 13.720 12.237 12.991 13.519 12.284 13.462 13.326 13.729 13.508
CEJ enamel not ablated 180 300 420 730 600 490 230 405 260
"Missing" cusp enamel (extrap.) 800 500 375 710 1450 280 380 560 390
Crown Height Total (microns) 10680 10100 10395 12350 9150 12280 11990 10740 12010
Microns Per half year 1186.667 1122.222 1155.000 1372.222 1016.667 1364.444 1332.222 1193.333 1334.444
Start Age with Cuspal enam (0.5yr) 0.837 0.723 0.662 0.759 1.213 0.603 0.643 0.735 0.646
End Age of 5 years 4.924 4.866 4.818 4.734 4.705 4.820 4.914 4.830 4.903
# Half year periods: 8.174 8.287 8.312 7.951 6.984 8.436 8.542 8.191 8.513
# Readings per half year: 86.492 91.708 88.911 101.502 82.765 101.355 99.975 86.921 98.791
Total years covered by ablations 4.087 4.144 4.156 3.975 3.492 4.218 4.271 4.096 4.256
LA1 Reading #'s 1-656 1-548 1-523 1-589 1-578 1-525 1-516 1-590 1-515
Years detailed in LA1 0.837-4.629 0.723-3.708 0.662-3.684 0.759-3.666 3.492 0.603-3.197 0.643-3.225 0.735-4.118 0.646-3.256
LA2 Reading #'s 657-707 549-760 524-739 590-807 - 526-855 517-804 591-712 516-787
Years detailed in LA2 4.630-4.924 3.709-4.866 3.685-4.818 3.667-4.734 - 3.198-4.820 3.226-4.681 4.119-4.830 3.257-4.641
LA3 Reading #'s - - - - - - 805-854 - 788-841
Years detailed in LA3 - - - - - - 4.682-4.914 - 4.642-4.903
Continued, next page
332

Table B.3, continued
Individual # YDL-85/62 YDL-85/65 YDL-85/68 YDL-85/86 YDL-85/87B YDL-85/97 YDL-85/98 YDL-85/101 YDL-85/103
1ST LA (microns) 6335 8420 6840 7260 7730 6220 7500 6985 8890
2ND LA (microns) 3170 710 3220 2000 3450 3770 900 1500 2290
3RD LA (microns) - 1140 - - - - - 1540 -
Total Ablation Length 9505 10270 10060 9260 11180 9990 8400 10025 11180
Total # Readings 689 758 814 684 818 728 712 780 813
Microns per reading 13.795 13.549 12.359 13.538 13.667 13.723 11.798 12.853 13.752
CEJ enamel not ablated 825 620 200 640 370 715 320 625 1040
"Missing" cusp enamel (extrap.) 830 620 640 1050 1120 960 1500 725 1620
Crown Height Total (microns) 11160 11510 10900 10950 12670 11665 10220 11375 13840
Microns Per half year 1240.000 1278.889 1211.111 1216.667 1407.778 1296.111 1135.556 1263.889 1537.778
Start Age with Cuspal enam (0.5yr) 0.835 0.742 0.764 0.932 0.898 0.870 1.160 0.787 1.027
End Age of 5 years 4.667 4.758 4.917 4.737 4.869 4.724 4.859 4.753 4.662
# Half year periods: 7.665 8.030 8.306 7.611 7.942 7.708 7.397 7.932 7.270
# Readings per half year: 89.885 94.391 97.996 89.870 103.002 94.451 96.252 98.337 111.826
Total years covered by ablations 3.833 4.015 4.153 3.805 3.971 3.854 3.699 3.966 3.635
LA1 Reading #'s 1-464 1-627 1-550 1-535 1-554 1-462 1-623 1-534 1-652
Years detailed in LA1 0.835-3.389 0.742-4.034 0.764-3.588 0.932-3.915 0.898-3.643 0.870-3.270 1.160-4.463 0.787-3.550 1.027-3.917
LA2 Reading #'s 465-689 628-672 551-814 536-684 555-818 463-728 624-712 535-657 653-813
Years detailed in LA2 3.390-4.667 4.035-4.313 3.589-4.917 3.916-4.737 3.644-4.869 3.271-4.724 4.464-4.859 3.551-4.144 3.918-4.662
LA3 Reading #'s - 673-758 - - - - - 658-780 -
Years detailed in LA3 - 4.314-4.758 - - - - - 4.145-4.753 -
333

Table B.4 Ablation details for Historical Lamanai mandibular canines
Individual # YDL-85/23 YDL-85/27 YDL-85/31 YDL-85/33 YDL-85/35 YDL-85/47 YDL-85/53 YDL-85/59
1ST LA (microns) 5870 5580 5290 5380 6400 6550 7130 5480
2ND LA (microns) 4010 3370 4240 3860 2980 3670 4010 4420
3RD LA (microns) 1080 860 1400 - - - 790 700
Total Ablation Length 10960 9810 10930 9240 9380 10220 11930 10600
Total # Readings 817 687 817 742 749 741 868 850
Microns per reading 13.415 14.279 13.378 12.453 12.523 13.792 13.744 12.471
CEJ enamel not ablated 360 180 240 620 250 900 300 460
"Missing" cusp enamel (extrap.) 700 650 860 420 520 330 550 440
Crown Height Total (microns) 12020 10640 12030 10280 10150 11450 12780 11500
Microns Per half year 1335.556 1182.222 1336.667 1142.222 1127.778 1272.222 1420.000 1277.778
Start Age with Cuspal enam (1yr) 1.262 1.275 1.322 1.184 1.231 1.130 1.194 1.172
End Age of 5.5 years 5.365 5.424 5.410 5.229 5.389 5.146 5.394 5.320
# Half year periods: 8.206 8.298 8.177 8.089 8.317 8.033 8.401 8.296
# Readings per half year: 99.557 82.792 99.914 91.724 90.054 92.242 103.316 102.463
Total years covered by ablations 4.103 4.149 4.089 4.045 4.159 4.017 4.201 4.148
LA1 Reading #'s 1-429 1-411 1-399 1-429 1-511 1-475 1-523 1-431
Years detailed in LA1 1.262-3.460 1.275-3.635 1.322-3.300 1.184-3.539 1.231-4.068 1.130-3.704 1.194-3.704 0.646-3.256
LA2 Reading #'s 430-732 412-636 400-715 430-742 512-749 476-741 524-805 432-783
Years detailed in LA2 3.461-4.962 3.636-5.061 3.301-4.887 3.540-5.229 4.069-5.389 3.705-5.146 3.705-5.117 3.257-4.641
LA3 Reading #'s 733-817 637-687 716-817 - - - 806-868 784-850
Years detailed in LA3 4.963-5.365 5.062-5.424 4.888-5.410 - - - 5.118-5.394 4.642-4.903
Continued, next page
334

Table B.4, continued
Individual # YDL-85/63 YDL-85/71A YDL-85/73 YDL-85/78A YDL-85/78C YDL-85/80 YDL-85/91A YDL-85/102
1ST LA (microns) 5340 6410 6090 8180 7100 10640 9020 6950
2ND LA (microns) 5280 3640 4730 2060 2680 3200 1500 1410
3RD LA (microns) - - - - - - - -
Total Ablation Length 10620 10050 10820 10240 9780 13840 10520 8360
Total # Readings 782 758 810 833 709 969 769 605
Microns per reading 13.581 13.259 13.358 12.293 13.794 14.283 13.680 13.818
CEJ enamel not ablated 1030 380 540 540 520 400 210 620
"Missing" cusp enamel (extrap.) 640 290 580 540 700 100 475 820
Crown Height Total (microns) 12290 10720 11940 11320 11000 14340 11205 9800
Microns Per half year 1365.556 1191.111 1326.667 1257.778 1222.222 1593.333 1245.000 1088.889
Start Age with Cuspal enam (1yr) 1.234 1.122 1.219 1.215 1.286 1.031 1.191 1.377
End Age of 5.5 years 5.123 5.340 5.296 5.285 5.287 5.374 5.416 5.215
# Half year periods: 7.777 8.438 8.156 8.141 8.002 8.686 8.450 7.678
# Readings per half year: 100.552 89.837 99.316 102.317 88.605 111.556 91.008 78.801
Total years covered by ablations 3.889 4.219 4.078 4.071 4.001 4.343 4.225 3.839
LA1 Reading #'s 1-403 1-470 1-460 1-662 1-516 1-742 1-654 1-510
Years detailed in LA1 1.234-3.190 1.122-3.813 1.219-3.514 1.215-4.466 1.286-4.191 1.031-4.370 1.191-4.813 1.377-4.568
LA2 Reading #'s 404-782 471-758 461-810 663-833 517-709 743-969 655-769 511-605
Years detailed in LA2 3.191-5.123 3.814-5.340 3.515-5.296 4.467-5.285 4.192-5.287 4.371-5.374 4.814-5.416 4.569-5.215
LA3 Reading #'s - - - - - - - -
Years detailed in LA3 - - - - - - - -
335
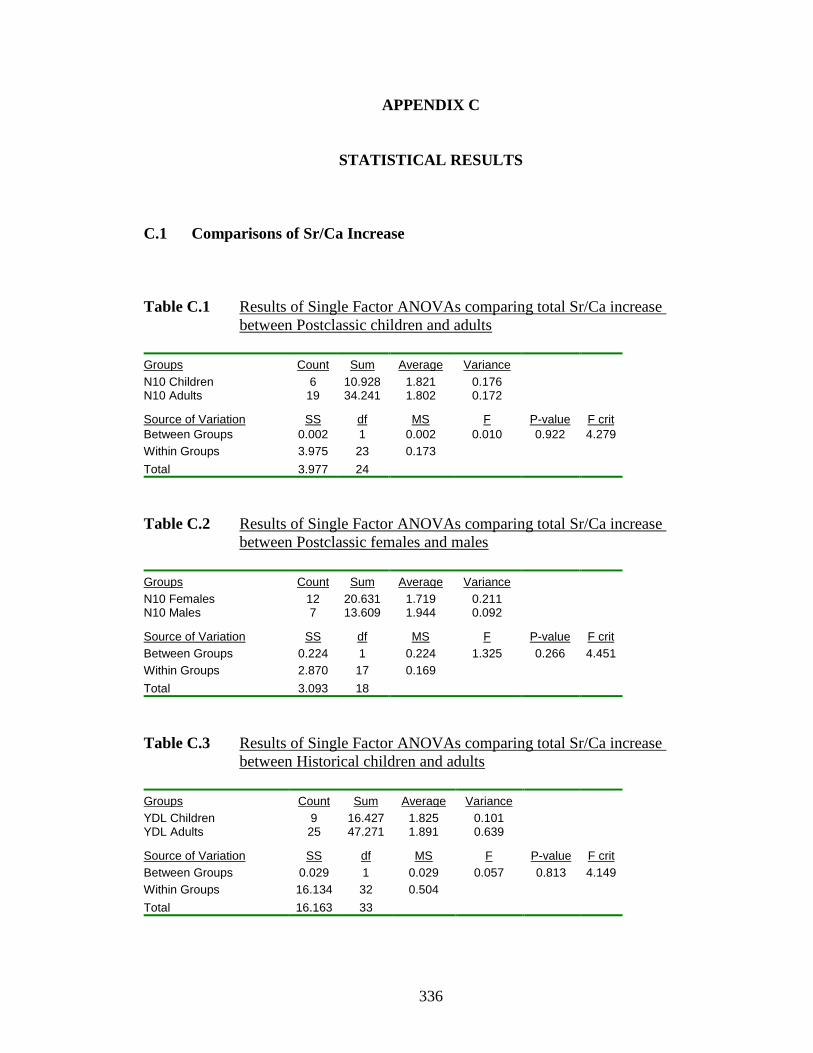
336
APPENDIX C
STATISTICAL RESULTS
C.1 Comparisons of Sr/Ca Increase
Table C.1 Results of Single Factor ANOVAs comparing total Sr/Ca increase
between Postclassic children and adults
Groups Count Sum Average Variance
N10 Children 6 10.928 1.821 0.176 N10 Adults 19 34.241 1.802 0.172
Source of Variation SS df MS F P-value F crit
Between Groups 0.002 1 0.002 0.010 0.922 4.279
Within Groups 3.975 23 0.173
Total 3.977 24
Table C.2 Results of Single Factor ANOVAs comparing total Sr/Ca increase
between Postclassic females and males
Groups Count Sum Average Variance
N10 Females 12 20.631 1.719 0.211 N10 Males 7 13.609 1.944 0.092
Source of Variation SS df MS F P-value F crit
Between Groups 0.224 1 0.224 1.325 0.266 4.451
Within Groups 2.870 17 0.169
Total 3.093 18
Table C.3 Results of Single Factor ANOVAs comparing total Sr/Ca increase
between Historical children and adults
Groups Count Sum Average Variance
YDL Children 9 16.427 1.825 0.101 YDL Adults 25 47.271 1.891 0.639
Source of Variation SS df MS F P-value F crit
Between Groups 0.029 1 0.029 0.057 0.813 4.149
Within Groups 16.134 32 0.504
Total 16.163 33

337
Table C.4 Results of Single Factor ANOVAs comparing total Sr/Ca increase
between Historical females and males
Groups Count Sum Average Variance
YDL Females 13 26.725 2.056 0.965 YDL Males 12 20.546 1.712 0.274
Source of Variation SS df MS F P-value F crit
Between Groups 0.737 1 0.737 1.161 0.292 4.279
Within Groups 14.592 23 0.634
Total 15.328 24
Table C.5 Results of Single Factor ANOVAs comparing total Sr/Ca increase
between Postclassic and Historical children
Groups Count Sum Average Variance
N10 Children 6 10.928 1.821 0.176 YDL Children 9 16.427 1.825 0.101
Source of Variation SS df MS F P-value F crit
Between Groups 0.000 1 0.000 0.000 0.984 4.667
Within Groups 1.688 13 0.130
Total 1.688 14
Table C.6 Results of Single Factor ANOVAs comparing total Sr/Ca increase
between Postclassic and Historical females
Groups Count Sum Average Variance
N10 Females 12 20.631 1.719 0.211 YDL Females 13 26.725 2.056 0.965
Source of Variation SS df MS F P-value F crit
Between Groups 0.707 1 0.707 1.170 0.291 4.279
Within Groups 13.893 23 0.604
Total 14.599 24
Table C.7 Results of Single Factor ANOVAs comparing total Sr/Ca increase
between Postclassic and Historical males
Groups Count Sum Average Variance
N10 Males 7 13.609 1.944 0.092 YDL Males 12 20.546 1.712 0.274
Source of Variation SS df MS F P-value F crit
Between Groups 0.238 1 0.238 1.134 0.302 4.451
Within Groups 3.569 17 0.210
Total 3.806 18

338
C.2 Comparisons of Sr/Ca at 0.25 year intervals
Table C.8 Results of Single Factor ANOVAs comparing Postclassic children
and adults (Sr/Ca at 0.25 year intervals)
Postclassic Children and Adults
Groups Count Sum Average Variance df F P-value F crit
Kids 0.5 2 0.09034 0.04517 0.00005 3 15.27498 0.05967 18.51276 Ads0.5 2 0.14623 0.07312 0.00005
Kids 0.75 4 0.20988 0.05247 0.00030 9 0.56532 0.47366 5.31764 Ads0.75 6 0.37437 0.06240 0.00049
Kids 1 6 0.29717 0.04953 0.00029 19 1.04811 0.31951 4.41386 Ads1 14 0.83352 0.05954 0.00044
Kids 1.25 6 0.30785 0.05131 0.00037 23 0.49096 0.49085 4.30094 Ads1.25 18 1.04393 0.05800 0.00042
Kids 1.5 6 0.29658 0.04943 0.00026 24 1.25559 0.27405 4.27934 Ads1.5 19 1.14157 0.06008 0.00045
Kids 1.75 6 0.30671 0.05112 0.00028 24 1.40049 0.24873 4.27934 Ads1.75 19 1.18664 0.06245 0.00046
Kids 2 6 0.32747 0.05458 0.00033 24 1.05537 0.31496 4.27934 Ads2 19 1.23218 0.06485 0.00049
Kids 2.25 6 0.34680 0.05780 0.00038 24 0.85259 0.36541 4.27934 Ads2.25 19 1.26389 0.06652 0.00041
Kids 2.5 6 0.34065 0.05677 0.00035 24 1.73462 0.20079 4.27934 Ads2.5 19 1.32519 0.06975 0.00047
Kids 2.75 6 0.37256 0.06209 0.00079 24 0.85462 0.36485 4.27934 Ads2.75 19 1.38193 0.07273 0.00055
Kids 3 6 0.37283 0.06214 0.00048 24 1.75369 0.19842 4.27934 Ads3 19 1.46755 0.07724 0.00062
Kids 3.25 6 0.39215 0.06536 0.00045 24 1.54094 0.22699 4.27934 Ads3.25 19 1.51137 0.07955 0.00064
Kids 3.5 6 0.40421 0.06737 0.00032 24 1.61625 0.21632 4.27934 Ads3.5 19 1.54217 0.08117 0.00060
Kids 3.75 6 0.41496 0.06916 0.00035 24 1.50116 0.23289 4.27934 Ads3.75 19 1.57843 0.08308 0.00065
Kids 4 6 0.43063 0.07177 0.00047 23 1.49751 0.23400 4.30094 Ads4 18 1.57042 0.08725 0.00079
Kids 4.25 6 0.43395 0.07233 0.00042 23 1.55557 0.22543 4.30094 Ads4.25 18 1.61688 0.08983 0.00102
Kids 4.5 6 0.42925 0.07154 0.00044 23 2.76773 0.11036 4.30094 Ads4.5 18 1.67401 0.09300 0.00084
Kids 4.75 4 0.30462 0.07615 0.00055 19 0.85081 0.36852 4.41386 Ads4.75 16 1.45161 0.09073 0.00085
Kids 5 2 0.16001 0.08001 0.00050 12 0.60277 0.45389 4.84434 Ads5 11 1.07150 0.09741 0.00089
Kids 5.25 3 0.25276 0.08425 0.00081 12 0.09161 0.76778 4.84434 Ads5.25 10 0.89269 0.08927 0.00060

339
Table C.9 Results of Single Factor ANOVAs comparing Postclassic females
and males (Sr/Ca at 0.25 year intervals)
Postclassic Females and Males
Groups Count Sum Average Variance df F P-value F crit
Fem0.75 5 0.33568 0.06714 0.00044 5 1.53018 0.28375 7.70865 Male0.75 1 0.03870 0.03870 N/A
Fem1 9 0.55501 0.06167 0.00048 13 0.24351 0.63059 4.74722 Male1 5 0.27851 0.05570 0.00044
Fem1.25 11 0.68828 0.06257 0.00049 17 1.43833 0.24787 4.49400 Male1.25 7 0.35565 0.05081 0.00028
Fem1.5 12 0.76424 0.06369 0.00052 18 0.92622 0.34934 4.45132 Male1.5 7 0.37733 0.05390 0.00034
Fem1.75 12 0.78660 0.06555 0.00056 18 0.67197 0.42371 4.45132 Male1.75 7 0.40005 0.05715 0.00030
Fem2 12 0.82220 0.06852 0.00055 18 0.88769 0.35930 4.45132 Male2 7 0.40998 0.05857 0.00038
Fem2.25 12 0.83502 0.06959 0.00045 18 0.72750 0.40555 4.45132 Male2.25 7 0.42886 0.06127 0.00037
Fem2.5 12 0.87157 0.07263 0.00055 18 0.56655 0.46193 4.45132 Male2.5 7 0.45361 0.06480 0.00035
Fem2.75 12 0.90650 0.07554 0.00066 18 0.45086 0.51095 4.45132 Male2.75 7 0.47543 0.06792 0.00040
Fem3 12 0.96391 0.08033 0.00079 18 0.48237 0.49673 4.45132 Male3 7 0.50364 0.07195 0.00038
Fem3.25 12 0.98693 0.08224 0.00077 18 0.35894 0.55700 4.45132 Male3.25 7 0.52444 0.07492 0.00045
Fem3.5 12 0.99102 0.08258 0.00077 18 0.10421 0.75077 4.45132 Male3.5 7 0.55116 0.07874 0.00036
Fem3.75 12 0.99497 0.08291 0.00076 18 0.00122 0.97255 4.45132 Male3.75 7 0.58346 0.08335 0.00056
Fem4 11 0.96493 0.08772 0.00105 17 0.00759 0.93165 4.49400 Male4 7 0.60549 0.08650 0.00049
Fem4.25 11 0.99639 0.09058 0.00160 17 0.01481 0.90465 4.49400 Male4.25 7 0.62049 0.08864 0.00023
Fem4.5 11 1.03343 0.09395 0.00110 17 0.02856 0.86793 4.49400 Male4.5 7 0.64057 0.09151 0.00053
Fem4.75 9 0.84098 0.09344 0.00102 15 0.16922 0.68703 4.60011 Male4.75 7 0.61062 0.08723 0.00073
Fem5 6 0.61787 0.10298 0.00078 10 0.43608 0.52556 5.11736 Male5 5 0.45363 0.09073 0.00114
Fem5.25 5 0.47264 0.09453 0.00071 9 0.43560 0.52779 5.31764 Male5.25 5 0.42005 0.08401 0.00056

340
Table C.10 Results of Single Factor ANOVAs comparing Historical children
and adults (Sr/Ca at 0.25 year intervals)
Historical Children and Adults
Groups Count Sum Average Variance df F P-value F crit
Kids 0.5 2 0.08834 0.04417 0.00012 5 0.70560 0.44818 7.70865 Ads0.5 4 0.13081 0.03270 0.00029
Kids 0.75 4 0.15685 0.03921 0.00031 14 0.25278 0.62353 4.66719 Ads0.75 11 0.50077 0.04552 0.00051
Kids 1 9 0.42220 0.04691 0.00023 28 0.00377 0.95148 4.21001 Ads1 20 0.92937 0.04647 0.00036
Kids 1.25 9 0.45277 0.05031 0.00042 33 0.04976 0.82490 4.14909 Ads1.25 25 1.21257 0.04850 0.00044
Kids 1.5 9 0.47784 0.05309 0.00051 33 0.26645 0.60927 4.14909 Ads1.5 25 1.22192 0.04888 0.00042
Kids 1.75 9 0.48534 0.05393 0.00055 33 0.15423 0.69713 4.14909 Ads1.75 25 1.26390 0.05056 0.00047
Kids 2 9 0.48725 0.05414 0.00047 33 0.07166 0.79066 4.14909 Ads2 25 1.29715 0.05189 0.00047
Kids 2.25 9 0.49945 0.05549 0.00037 33 0.05010 0.82432 4.14909 Ads2.25 25 1.34172 0.05367 0.00046
Kids 2.5 9 0.51406 0.05712 0.00043 33 0.04641 0.83079 4.14909 Ads2.5 25 1.38267 0.05531 0.00048
Kids 2.75 9 0.52248 0.05805 0.00049 33 0.00741 0.93195 4.14909 Ads2.75 25 1.43209 0.05728 0.00054
Kids 3 9 0.52691 0.05855 0.00051 33 0.03772 0.84722 4.14909 Ads3 25 1.51049 0.06042 0.00065
Kids 3.25 9 0.54858 0.06095 0.00052 33 0.00381 0.95118 4.14909 Ads3.25 25 1.54020 0.06161 0.00082
Kids 3.5 9 0.58571 0.06508 0.00060 33 0.03528 0.85219 4.14909 Ads3.5 25 1.57552 0.06302 0.00086
Kids 3.75 9 0.62978 0.06998 0.00068 33 0.26277 0.61175 4.14909 Ads3.75 25 1.60799 0.06432 0.00085
Kids 4 9 0.67403 0.07489 0.00080 33 0.76154 0.38935 4.14909 Ads4 25 1.64150 0.06566 0.00072
Kids 4.25 9 0.67580 0.07509 0.00078 33 1.05428 0.31222 4.14909 Ads4.25 25 1.62540 0.06502 0.00059
Kids 4.5 9 0.69170 0.07686 0.00087 33 0.82536 0.37041 4.14909 Ads4.5 25 1.67280 0.06691 0.00077
Kids 4.75 9 0.69766 0.07752 0.00078 26 0.74953 0.39486 4.24170 Ads4.75 18 1.19914 0.06662 0.00103
Kids 5 5 0.39210 0.07842 0.00023 15 0.22539 0.64229 4.60011 Ads5 11 0.76260 0.06933 0.00167
Kids 5.25 4 0.31256 0.07814 0.00031 11 0.02911 0.86793 4.96459 Ads5.25 8 0.59250 0.07406 0.00204

341
Table C.11 Results of Single Factor ANOVAs comparing Historical females
and males (Sr/Ca at 0.25 year intervals)
Historical Females and Males
Groups Count Sum Average Variance df F P-value F crit
Fem0.75 6 0.20677 0.03446 0.00024 10 4.18371 0.07114 5.11736
Male0.75 5 0.29401 0.05880 0.00057
Fem1 8 0.34199 0.04275 0.00043 19 0.49835 0.48926 4.41386 Male1 12 0.58739 0.04895 0.00033
Fem1.25 13 0.58623 0.04509 0.00049 24 0.71174 0.40756 4.27934 Male1.25 12 0.62634 0.05220 0.00039
Fem1.5 13 0.58174 0.04475 0.00045 24 1.10439 0.30422 4.27934 Male1.5 12 0.64018 0.05335 0.00039
Fem1.75 13 0.60493 0.04653 0.00053 24 0.93576 0.34343 4.27934 Male1.75 12 0.65897 0.05491 0.00040
Fem2 13 0.64215 0.04940 0.00061 24 0.34790 0.56105 4.27934 Male2 12 0.65500 0.05458 0.00035
Fem2.25 13 0.64362 0.04951 0.00055 24 1.01163 0.32498 4.27934 Male2.25 12 0.69810 0.05818 0.00036
Fem2.5 13 0.67428 0.05187 0.00055 24 0.65850 0.42541 4.27934 Male2.5 12 0.70839 0.05903 0.00041
Fem2.75 13 0.70098 0.05392 0.00061 24 0.55458 0.46400 4.27934 Male2.75 12 0.73111 0.06093 0.00049
Fem3 13 0.74855 0.05758 0.00083 24 0.32557 0.57381 4.27934 Male3 12 0.76193 0.06349 0.00050
Fem3.25 13 0.77394 0.05953 0.00111 24 0.13688 0.71478 4.27934 Male3.25 12 0.76626 0.06386 0.00057
Fem3.5 13 0.78107 0.06008 0.00120 24 0.26361 0.61255 4.27934 Male3.5 12 0.79445 0.06620 0.00055
Fem3.75 13 0.82460 0.06343 0.00127 24 0.02418 0.87777 4.27934 Male3.75 12 0.78339 0.06528 0.00046
Fem4 13 0.84387 0.06491 0.00114 24 0.02013 0.88840 4.27934 Male4 12 0.79763 0.06647 0.00033
Fem4.25 13 0.81894 0.06300 0.00088 24 0.18078 0.67465 4.27934 Male4.25 12 0.80646 0.06721 0.00031
Fem4.5 13 0.83177 0.06398 0.00109 24 0.29363 0.59311 4.27934 Male4.5 12 0.84103 0.07009 0.00047
Fem4.75 10 0.68550 0.06855 0.00134 17 0.07696 0.78501 4.49400 Male4.75 8 0.51364 0.06421 0.00077
Fem5 6 0.44710 0.07452 0.00219 10 0.19533 0.66895 5.11736 Male5 5 0.31550 0.06310 0.00135
Fem5.25 5 0.36461 0.07292 0.00186 7 0.00728 0.93477 5.98737 Male5.25 3 0.22789 0.07596 0.00341

342
Table C.12 Results of Single Factor ANOVAs comparing Postclassic and Historical
children (Sr/Ca at 0.25 year intervals)
Postclassic and Historical Children
Groups Count Sum Average Variance df F P-value F crit
N10 0.5 2 0.09034 0.04517 0.00005 3 0.01122 0.92531 18.51276 YDL 0.5 2 0.08834 0.04417 0.00012
N10 0.75 4 0.20988 0.05247 0.00030 7 1.15284 0.32423 5.98737 YDL 0.75 4 0.15685 0.03921 0.00031
N10 1 6 0.29717 0.04953 0.00029 14 0.09634 0.76118 4.66719 YDL 1 9 0.42220 0.04691 0.00023
N10 1.25 6 0.30785 0.05131 0.00037 14 0.00898 0.92594 4.66719 YDL 1.25 9 0.45277 0.05031 0.00042
N10 1.5 6 0.29658 0.04943 0.00026 14 0.11738 0.73737 4.66719 YDL 1.5 9 0.47784 0.05309 0.00051
N10 1.75 6 0.30671 0.05112 0.00028 14 0.06351 0.80497 4.66719 YDL 1.75 9 0.48534 0.05393 0.00055
N10 2 6 0.32747 0.05458 0.00033 14 0.00167 0.96798 4.66719 YDL 2 9 0.48725 0.05414 0.00047
N10 2.25 6 0.34680 0.05780 0.00038 14 0.05124 0.82444 4.66719 YDL 2.25 9 0.49945 0.05549 0.00037
N10 2.5 6 0.34065 0.05677 0.00035 14 0.00106 0.97456 4.66719 YDL 2.5 9 0.51406 0.05712 0.00043
N10 2.75 6 0.37256 0.06209 0.00079 14 0.09678 0.76066 4.66719 YDL 2.75 9 0.52248 0.05805 0.00049
N10 3 6 0.37283 0.06214 0.00048 14 0.09344 0.76469 4.66719 YDL 3 9 0.52691 0.05855 0.00051
N10 3.25 6 0.39215 0.06536 0.00045 14 0.14263 0.71177 4.66719 YDL 3.25 9 0.54858 0.06095 0.00052
N10 3.5 6 0.40421 0.06737 0.00032 14 0.03841 0.84765 4.66719 YDL 3.5 9 0.58571 0.06508 0.00060
N10 3.75 6 0.41496 0.06916 0.00035 14 0.00433 0.94853 4.66719 YDL 3.75 9 0.62978 0.06998 0.00068
N10 4 6 0.43063 0.07177 0.00047 14 0.05192 0.82330 4.66719 YDL 4 9 0.67403 0.07489 0.00080
N10 4.25 6 0.43395 0.07233 0.00042 14 0.04310 0.83876 4.66719 YDL 4.25 9 0.67580 0.07509 0.00078
N10 4.5 6 0.42925 0.07154 0.00044 14 0.14477 0.70973 4.66719 YDL 4.5 9 0.69170 0.07686 0.00087
N10 4.75 4 0.30462 0.07615 0.00055 12 0.00716 0.93408 4.84434 YDL 4.75 9 0.69766 0.07752 0.00078
N10 5 2 0.16001 0.08001 0.00050 6 0.01267 0.91475 6.60788 YDL 5 5 0.39210 0.07842 0.00023
N10 5.25 3 0.25276 0.08425 0.00081 6 0.12529 0.73780 6.60788 YDL 5.25 4 0.31256 0.07814 0.00031

343
Table C.13 Results of Single Factor ANOVAs comparing Postclassic and Historical
females (Sr/Ca at 0.25 year intervals)
(* denotes significant difference)
Postclassic and Historical Females
Groups Count Sum Average Variance df F P-value F crit
N10 0.5 2 0.14623 0.07312 0.00005 5 9.48862 *0.03692 7.70865 YDL 0.5 4 0.13081 0.03270 0.00029
N10 0.75 5 0.33568 0.06714 0.00044 10 8.84065 *0.01562 5.11736 YDL 0.75 6 0.20677 0.03446 0.00024
N10 1 9 0.55501 0.06167 0.00048 16 3.30189 0.08923 4.54307 YDL 1 8 0.34199 0.04275 0.00043
N10 1.25 11 0.68828 0.06257 0.00049 23 3.73040 0.06641 4.30094 YDL 1.25 13 0.58623 0.04509 0.00049
N10 1.5 12 0.76424 0.06369 0.00052 24 4.65515 *0.04164 4.27934 YDL 1.5 13 0.58174 0.04475 0.00045
N10 1.75 12 0.78660 0.06555 0.00056 24 4.14578 0.05341 4.27934 YDL 1.75 13 0.60493 0.04653 0.00053
N10 2 12 0.82220 0.06852 0.00055 24 3.92022 0.05980 4.27934 YDL 2 13 0.64215 0.04940 0.00061
N10 2.25 12 0.83502 0.06959 0.00045 24 4.98177 *0.03564 4.27934 YDL 2.25 13 0.64362 0.04951 0.00055
N10 2.5 12 0.87157 0.07263 0.00055 24 4.89554 *0.03713 4.27934 YDL 2.5 13 0.67428 0.05187 0.00055
N10 2.75 12 0.90650 0.07554 0.00066 24 4.57797 *0.04322 4.27934 YDL 2.75 13 0.70098 0.05392 0.00061
N10 3 12 0.96391 0.08033 0.00079 24 4.00118 0.05741 4.27934 YDL 3 13 0.74855 0.05758 0.00083
N10 3.25 12 0.98693 0.08224 0.00077 24 3.39603 0.07828 4.27934 YDL 3.25 13 0.77394 0.05953 0.00111
N10 3.5 12 0.99102 0.08258 0.00077 24 3.17937 0.08778 4.27934 YDL 3.5 13 0.78107 0.06008 0.00120
N10 3.75 12 0.99497 0.08291 0.00076 24 2.30252 0.14279 4.27934 YDL 3.75 13 0.82460 0.06343 0.00127
N10 4 11 0.96493 0.08772 0.00105 23 2.82204 0.10713 4.30094 YDL 4 13 0.84387 0.06491 0.00114
N10 4.25 11 0.99639 0.09058 0.00160 23 3.75095 0.06573 4.30094 YDL 4.25 13 0.81894 0.06300 0.00088
N10 4.5 11 1.03343 0.09395 0.00110 23 4.88964 *0.03771 4.30094 YDL 4.5 13 0.83177 0.06398 0.00109
N10 4.75 9 0.84098 0.09344 0.00102 18 2.46219 0.13504 4.45132 YDL 4.75 10 0.68550 0.06855 0.00134
N10 5 6 0.61787 0.10298 0.00078 11 1.63537 0.22984 4.96459 YDL 5 6 0.44710 0.07452 0.00219
N10 5.25 5 0.47264 0.09453 0.00071 9 0.90683 0.36884 5.31764 YDL 5.25 5 0.36461 0.07292 0.00186

344
Table C.14 Results of Single Factor ANOVAs comparing Postclassic and Historical
males (Sr/Ca at 0.25 year intervals)
(* denotes significant difference)
Postclassic and Historical Males
Groups Count Sum Average Variance df F P-value F crit
N10 0.75 1 0.03870 0.03870 N/A 5 0.59267 0.48433 7.70865 YDL 0.75 5 0.29401 0.05880 0.00057
N10 1 5 0.27851 0.05570 0.00044 16 0.44552 0.51461 4.54307 YDL 1 12 0.58739 0.04895 0.00033
N10 1.25 7 0.35565 0.05081 0.00028 18 0.02406 0.87855 4.45132 YDL 1.25 12 0.62634 0.05220 0.00039
N10 1.5 7 0.37733 0.05390 0.00034 18 0.00368 0.95235 4.45132 YDL 1.5 12 0.64018 0.05335 0.00039
N10 1.75 7 0.40005 0.05715 0.00030 18 0.06110 0.80773 4.45132 YDL 1.75 12 0.65897 0.05491 0.00040
N10 2 7 0.40998 0.05857 0.00038 18 0.19576 0.66374 4.45132 YDL 2 12 0.65500 0.05458 0.00035
N10 2.25 7 0.42886 0.06127 0.00037 18 0.11595 0.73764 4.45132 YDL 2.25 12 0.69810 0.05818 0.00036
N10 2.5 7 0.45361 0.06480 0.00035 18 0.37445 0.54869 4.45132 YDL 2.5 12 0.70839 0.05903 0.00041
N10 2.75 7 0.47543 0.06792 0.00040 18 0.47558 0.49974 4.45132 YDL 2.75 12 0.73111 0.06093 0.00049
N10 3 7 0.50364 0.07195 0.00038 18 0.68924 0.41793 4.45132 YDL 3 12 0.76193 0.06349 0.00050
N10 3.25 7 0.52444 0.07492 0.00045 18 1.02062 0.32654 4.45132 YDL 3.25 12 0.76626 0.06386 0.00057
N10 3.5 7 0.55116 0.07874 0.00036 18 1.43544 0.24732 4.45132 YDL 3.5 12 0.79445 0.06620 0.00055
N10 3.75 7 0.58346 0.08335 0.00056 18 2.90750 0.10637 4.45132 YDL 3.75 12 0.78339 0.06528 0.00046
N10 4 7 0.60549 0.08650 0.00049 18 4.59187 *0.04689 4.45132 YDL 4 12 0.79763 0.06647 0.00033
N10 4.25 7 0.62049 0.08864 0.00023 18 7.10591 *0.01630 4.45132 YDL 4.25
12 0.80646 0.06721 0.00031
N10 4.5 7 0.64057 0.09151 0.00053 18 4.11598 0.05844 4.45132 YDL 4.5 12 0.84103 0.07009 0.00047
N10 4.75 7 0.61062 0.08723 0.00073 14 2.64189 0.12807 4.66719 YDL 4.75 8 0.51364 0.06421 0.00077
N10 5 5 0.45363 0.09073 0.00114 9 1.53145 0.25098 5.31764 YDL 5 5 0.31550 0.06310 0.00135
N10 5.25 5 0.42005 0.08401 0.00056 7 0.08040 0.78628 5.98737 YDL 5.25 3 0.22789 0.07596 0.00341

345
BIBLIOGRAPHY
AAP (American Academy of Pediatrics)
1980 Committee on Nutrition: On the Feeding of Supplemental Foods to Infants.
Pediatrics 65: 1178.
1996 (Work Group on Breastfeeding). Breastfeeding and the use of human Milk.
Pediatrics 100: 1035-1039.
Adair, L.S. and E. Pollitt
1985 Outcome of maternal nutritional supplementation: A comprehensive review of
the Bacon Chow study. Am. J. Clin. Nutr. 41: 948-978.
Ahlberg, M. and R. Akelsson
1976 Proton induced X-ray emission in the trace analysis of human tooth enamel and
dentine. Int. J. Appl. Radiat. Biol. 27: 279-90.
Ahrens, L.H. and S.R. Taylor
1961 Spectrochemical Analysis, 2nd
ed., Chapter 13. Addison-Wesley, Reading, MA.
Alexander, G.V., R.E. Nusbaum and N.S. MacDonald
1956 The relative retention of strontium and calcium in bone tissue. J. Biol. Chem.
218: 911-919.
Allen, L.H.
1994 Maternal micronutrient malnutrition: Effects on breast milk and infant nutrition,
and priorities for intervention. SCN News No. 11: 21-24.
1998 Women’s dietary calcium requirements are not increased by pregnancy or
lactation. Am. J. Clin. Nutr. 67: 591-592.
Allen, L.H., J.C. King and B. Lonnerdal (eds)
1994 Nutrient Regulation During Pregnancy, Lactation and Infant Growth. Plenum,
New York.
Aloia, J.F., A.N. Vaswani, J.K. Yeh, P. Ross, K. Ellis and S.H. Cohn
1983 Determinants of bone mass in postmenopausal women. Archives of Internal
Medicine 143: 1700-1704.
Alt, K.W., F.W. Rosing and M. Teschler-Nicola
1998 Dental Anthropology: Fundamentals, Limits and Prospects. Springer-Verlag,
New York.

346
Amarasiriwardena, D., S.F. Durrant, A. Laszitity, A. Krushevska, M.D. Argentine, and
R.M. Barnes
1997 Evaluation of semi-quantitative analysis of biological materials by inductively
coupled plasma mass spectrometry. Microchemical Journal 56: 352-372.
Ambrose, S.H.
1986 Stable carbon and nitrogen isotope analysis of human and animal diet in Africa.
J. Human Evol. 15: 707-731.
1990 Preparation and characterization of bone and tooth collagen for stable carbon
and nitrogen isotope analysis. J. of Archaeological Science 17: 431-51.
1991 Effects of diet, climate and physiology on nitrogen isotope abundances in
terrestrial foodwebs. J. Archaeological Science 18: 293-317.
1993 Isotopic analysis of paleodiets: Methodological and interpretive considerations.
In Investigations of Ancient Human Tissue: Chemical Analyses in
Anthropology, ed. by MK Sandford, 59-130. Gordon and Breach Science,
Langhorne, PA.
Anderson, D.L., G.W. Thompson and F. Popovich
1976 Age of attainment of mineralization stages of the permanent dentition. J.
Forensic Science 21: 191-200.
Anderson, P.
1983 The reproductive role of the human breast. Current Anthropology 24(1): 25-45.
Ardren, T. (ed.)
2002 Ancient Maya Women. Altamira Press, New York.
Armelagos, G.J., B. Brenton, M. Alcorn, D. Martin and D.P. van Gerven
1989 Factors affecting elemental and isotopic variation in prehistoric human
skeletons. In The Chemistry of Prehistoric Human Bone, ed. by T.D. Price, 230-
244. Cambridge University Press, England.
Asenso-Okyere, N.M.
1996 Large differences in nutritional status between fully weaned and partially breast
fed children beyond the age of 12 months. Eur. J. Clin. Nutr. 50: 171-177.
Asper, H.
1916 Uber die braune Retzius’sche Parallelstreifung im Schmelz der Menschlichen
Zahne. Schweiz Vjschr Zahnheilk 26: 275-314.
Aufderheide, A.C.
1989 Chemical analysis of skeletal remains. In Reconstruction of Life from the
Skeleton, ed. by M.Y. Iscan and K.A.R. Kennedy, 237-260. Alan R. Liss, New
York.

347
Avioli, L.V.
1988 Calcium and phosphorus. In Modern Nutrition in Health and Disease, ed. by
M.E. Shils and V.R. Young, 7th
ed., 142-158. Lea and Febiger, Philadelphia,
PA.
Ballabriga, A. and J. Rey (eds)
1987 Weaning: Why, What, and When? Nestle Nutrition Workshop Series Volume
10. Raven Press, New York.
Bauer, G., L.S. Ewald, J. Hoffman and R. Dubanoski
1991 Breastfeeding and cognitive development of three-year old children.
Psychological Reports 68: 1218.
Beck, L.A.
1985 Bivariate analysis of trace elements in bone. J. Human Evolution 14: 493-502.
Bedford, J., G.E. Harrison, W.H.A. Raymond and A. Sutton
1960 The metabolism of strontium in children. Brit. Med. J. I: 589-592.
Behar, M.
1968 Food and nutrition of the Maya before the conquest and at the present time. In
Biomedical Challenges Presented to the American Indians. Scientific
Publication No. 165, 114-119. Pan American Health Organization, Washington,
DC
Belloto, V.R. and N. Miekeley
2000 Improvements in calibration procedures for the quantitative determination of
trace elements in carbonate material (mussel shells) by laser ablation ICP-MS.
Fresenius J. Anal. Chem. 367: 635-640.
Bender, M.M., D.A. Baerris and R.L. Steventon
1981 Further light on carbon isotopes and Hopewell agriculture. American Antiquity
46: 346-353.
Benedict, F.G. and M. Steggerda
1937 The Food of the Present-day Maya Indians of the Yucatan. Contributions to
American Archaeology No. 18. Publication 456. Carnegie Institution of
Washington, Washington, DC.
Benfer, R.A.
1984 The challenges and rewards of sedentism: The preceramic village of Paloma,
Peru. In Paleopathology at the Origins of Agriculture, ed. by M.N. Cohen and
G.J. Armelagos, 531-38. Academic Press, Orlando.

348
Beninson, D., E. Ramos and R. Touzet
1964 Sr-90 levels in the diets and bones of children. US Atomic Energy Commission
HASL Report no. 149.
Bennett, S.
1986 Trace Element Evidence of the Colonial Mayan Diet at Tipu. Paper presented at
the 51st Ann. Meeting of the Society for American Archaeology, New Orleans.
Beynon, A.D.
1986 Tooth growth and structure in living and fossil hominoids. In Teeth and
Anthropology, ed. by E. Cruwys and R.A. Foley, 23-30. BAR International
Series 291. Oxford.
1992 Circaseptan rhythms in enamel development in modern humans and Plio-
Pleistocene hominids. In Structure, Function and Evolution of Teeth, ed. by P.
Smith and E. Tchernov, 295-310. Freund Publishing House, London.
Beynon, A.D., C.B. Clayton, F.V. Ramirez Rozzi and D.J. Reid
1998 Radiographic and histological methodologies in estimating the chronology of
crown development in modern humans and great apes: A review, with some
applications for studies on juvenile hominids. J. Human Evolution 35 (4-5):
351-370.
Bines, J. and W.A. Walker
1991 Growth factors and the development of neonatal host defense. In Immunology of
Milk and the Neonate, ed. By J. Mestecky, C. Blair and P. Ogra, 31-39. Plenum
Press, New York.
Blakely R.L.
1989 Bone strontium in pregnant and lactating females from archaeological samples.
Am. J. of Phys. Anthropol. 80: 173-185.
Blakey, M.L.
1981 An analysis of hypoplasia and hypocalcification in deciduous dentition from
Dickson Mounds. In Biocultural Adaptation: Comprehensive Approaches to
Skeletal Analysis, ed. by D.L. Martin and M.P. Bumsted, 24-34. Research
Reports No. 20, Dept. of Anthropology, Univ. of Massachusetts, Amherst.
1994 Psychophysiological stress disorders of industrial society: A critical theoretical
formulation of biocultural research. In Diagnosing America: Anthropology and
Public Engagement, ed. by S. Forman, 149-92. University of Michigan Press,
Ann Arbor.
Blakey, M.L. and G.J. Armelagos
1985 Deciduous enamel defects in prehistoric Americans from Dickson Mounds:
Prenatal and postnatal stress. Am. J. Phys. Anthropol. 66: 371-380.

349
Blakey, M.L., T.E. Leslie and J.P. Reidy
1994 Frequency and chronological distribution of dental enamel hypoplasia in
enslaved African Americans: A test of the weaning hypothesis. Am. J. Phys.
Anthropol. 95: 371-383.
Blanusa, M., M. Harmut and K. Kostial
1970 Comparative strontium and calcium metabolism in lactation. Arch. Hig. Rad.
Toksik. 21: 125-127.
Boyde, A.
1976 Amelogenesis and the development of teeth. In Scientific Foundations of
Dentistry, ed. by B. Cohen and I.R.H. Kramer. Heinemann, London.
1989 Enamel. In Teeth, ed. by B.K.B. Berkovitz, A. Boyde, R.M. Frank, H.J.
Hohling, B.J. Moxham, J. Nalbandian and C.H. Tonge, 309-473. Springer-
Verlag, Berlin.
Boyde, A., S.J. Jones and P.S. Reynolds
1978 Quantitative and qualitative studies of enamel etching with acid and EDTA.
Scanning Electron Microscopy 1978/II: 991-1002.
Boyde, J., W.F. Neumann and H.C. Hodge
1958 On the mechanism of skeletal fixation of strontium. Atomic Energy Project UR
513: 1-16, University of Rochester.
Brace, C.L.
1995 Biocultural interaction and the mechanism of mosaic evolution in the
emergence of ‘modern’ morphology. American Anthropologist 97: 711-721.
Brace, C.L. and P.E. Mahler
1971 Post-Pleistocene changes in the human dentition. Am. J. Phys. Anthropol. 34:
191-204.
Bressani, R.
1990 Chemistry, technology, and nutritive value of maize tortillas. Food Reviews
International 6(2): 225-264.
Bressani, R., V. Benavides, E. Acevedo and M.A. Ortiz
1990 Changes in selected nutrient contents and in protein quality of common and
quality-protein maize during rural tortilla preparation. Cereal Chemistry 67(6):
515-518.
Bressani, R., J.C. Turcios and A.S.C. de Ruiz
2002 Nixtamalization effects on the contents of phytic acid, calcium, iron and zinc in
the whole grain, endosperm and germ of maize. Food Science and Technology
International 8(2): 81-86.

350
Bressani, R., R. Paz and N.S. Scrimshaw
1958 Effect of lime treatment on in vitro availability of essential amino acids and
solubility of protein fractions in corn. J. Agric. Food Chem. 6: 770-774.
Briend, A., B. Wojtyniak and M.G. Rowland
1988 Breast feeding, nutritional state, and child survival in rural Bangladesh. Br.
Med. J. 296: 879-882.
Bromage, T.G. and M.C. Dean
1985 Re-evaluation of the age at death of immature fossil hominids. Nature 317: 525-
527.
Brothwell, D.
1963 Dental Anthropology (ed.). Pergamon Press, New York.
1981 Digging Up Bones. Cornell University Press, Ithaca.
Brown, A.B.
1973 Bone Strontium Content as a Dietary Indicator in Human Skeletal Populations.
Ph.D. Dissertation, University of Michigan, Ann Arbor, MI.
1974 Bone strontium as a dietary indicator in human skeletal populations.
Contributions to Geology 13: 47-48.
Brown, K.H., R.E. Black, G. Lopez de Romana and H. Creed de Kanashiro
1989 Infant-feeding practices and their relationship with diarrheal and other disease
in Huascar (Lima), Peru. Pediatrics 83: 31-40.
Brown, L.K. and K. Mussell (eds.)
1984 Ethnic and Regional Foodways in the United States: The Performance of Group
Identity. University of Tennessee Press, Knoxville.
Bruce-Chwatt, L.J.
1965 Paleogenesis and paleoepidemiology of primate malaria. Bulletin of the World
Health Organization 32: 363-387.
Brudevold, F. and L.T. Steadman
1956 The distribution of lead in human enamel. J. of Dental Research 35: 420.
Bryant, F.J. and J.F. Loutit
1964 The entry of strontium-90 into human bone. Proceedings of the Royal Society of
London 159B: 449-465.
Buckley, K.M.
2001 Long-term breastfeeding: Nourishment or nurturance? J. Human Lactation
17(4): 304-312.

351
Budd, P., B.L. Gulson, J. Montgomery, P. Rainbird, R.G. Thomas and S.M.M. Young
1996 Pb- and Sr-isotope composition of human dental enamel: An indicator of Pacific
Islander population dynamics. In The Western Pacific 5000 to 2000 BP, 3rd
Archaeological Conference, ed. by J.C. Galipaud. Noumea, Port Vila, Vanuatu.
Budd P., J. Montgomery, B. Barreiro and R.G. Thomas
2000 Differential diagenesis of strontium in archaeological human dental tissues.
Applied Geochemistry 15(5): 687-694.
Budd, P., J. Montgomery, A. Cox, P. Krause, B. Barreiro and R.G. Thomas
1998 The distribution of lead within ancient and modern human teeth: Implications
for long-term and historical exposure monitoring. The Science of the Total
Environment 220: 121-136.
Buikstra, J.E., S. Frankenburg, J.B. Lambert and L. Xue
1989 Multiple elements: Multiple expectations. In The Chemistry of Prehistoric
Human Bone, ed. by T.D. Price, 155-210. Cambridge University Press,
Cambridge.
Buikstra, J.E. and D. Ubelaker (eds.)
1994 Standards for Data Collection from Human Skeletal Remains. Arkansas
Archaeological Survey Research Series, 44.
Bullion, S.K.
1987 Incremental Structures of Enamel and their Applications to Archaeology. Ph.D.
dissertation, University of Lancaster, United Kingdom.
Bumsted, M.P.
1985 Past human behavior from bone chemical analysis - respects and prospects. J.
Human Evolution 14: 539-551.
Burton, J.H. and T.D. Price
1990 The ratio of barium to strontium as a paleodietary indicator of consumption of
marine resources. J. of Archaeological Science 17: 547-557.
1999 Evaluation of bone strontium as a measure of seafood consumption. Int. J.
Osteoarchaeology 9: 233-236.
Burton, J.H. and L.E. Wright
1995 Nonlinearity in the relationship between bone Sr/Ca and diet: Paleodietary
Implications. Am. J. Phys. Anthropol. 96: 273-282.
Burton, J.H., T. D. Price and W.D. Middleton
1999 Correlation of bone Ba/Ca and Sr/Ca due to biological purification of calcium.
J. Arch. Science 26: 609-616.

352
Butte, N.F., S. Villalpando, W.W. Wong, S. Flores-Huerta, M.J. Hernandez-Beltran,
E.O.B. Smith and C. Garza
1992 Human milk intake and growth faltering of rural Mesoamerindian infants. Am.
J. Clin. Nutr. 55: 1109-1116.
Byers, T., S. Graham, T. Rzepka and J. Marshall
1985 Lactation and breast cancer: Evidence for a negative association in
premenopausal women. Am. J. of Epidemiology 12: 664-674.
Calvo, E.B. and N. Gnazzo
1990 Prevalence of iron deficiency in children aged 9-24 mo from a large urban area
of Argentina. Am. J. Clinical Nutrition 52: 534-540.
Cantrelle, P. and H. Leridon
1971 Breast feeding, mortality in childhood and fertility in a rural zone of Senegal.
Population Studies 25: 505-533.
Carballo, M. and G.H. Pelto
1991 Social and psychological factors in breastfeeding. In Infant and Child Nutrition
Worldwide: Issues and Perspectives, ed. by F. Falkner, 175-190. CRC Press,
Boston.
Carlson, S.J.
1990 Vertebrate dental structures. In Skeletal Biomineralization: Patterns, Processes
and Evolutionary Trends, Volume 1, edited by J.G. Carter, Chapter 21 (531-
556). Van Nostrand Reinhold, New York.
Carroll, K.G., H.L. Needleman, O.C. Tuncay and I.M. Shapiro
1972 The distribution of lead in human deciduous teeth. Experientia 28: 434-5.
Caulfield, L.E., M.E. Bentley and S. Ahmed
1996 Is prolonged breastfeeding associated with malnutrition? Evidence from
nineteen demographic and health surveys. Int. J. Epidemiol. 25: 693-703.
Chan, G.M., N. Ronald, P. Slater, J. Hollis and M.R. Thomas
1982 Decreased bone mineral status in lactating adolescent mothers. J. Pediatrics
101: 767-770.
Chandra, S.K. and M. Lakshmiswaramma
1991 Working Women and Infant Care (Problems of Breast Feeding and Infant
Weaning faced by Women Working in the Organized and Unorganized Sectors).
Mittal Publications, New Delhi, India.
Chaudhri, M.A. and T. Ainsworth
1981 Applications of PIXE to studies in dental and mental healths. Nuclear Instrum.
Methods 181: 333-336.

353
Chavez, A. and C. Martinez
1982 Growing Up in a Developing Community: A Bio-Ecologic Study of the
Development of Children from Poor Peasant Families in Mexico. INCAP,
Mexico.
Chen, L.C. and N.S. Scrimshaw (eds.)
1983 Diarrhea and Malnutrition. Plenum Press, New York.
Chisholm, B.S.
1989 Variation in diet reconstructions based on stable carbon isotopic evidence. In
The Chemistry of Prehistoric Human Bone, ed. by T.D. Price, 10-37.
Cambridge University Press, England.
Chisholm, B.S., D.E. Nelson and H.P. Schwarcz
1982 Stable isotope ratios as a measure of marine versus terrestrial protein in ancient
diets. Science 216: 1131-32.
Clark, J.E. and S.D. Houston
1998 Craft specialization, gender and personhood among the post-conquest Maya of
Yucatan, Mexico. In Craft and Social Identity, ed. by C.L. Costin and R.P.
Wright, 31-46. Archaeological papers of the American Anthropological
Association #8. Arlington, VA.
Clarke, S.K.
1980 Early childhood morbidity trends in prehistoric populations. Human Biology 52:
79-85.
Clendinnen, I.
1982 Yucatec Maya women and the Spanish Conquest: Role and ritual in historical
reconstruction. J. Social History (summer): 427-442.
1987 Ambivalent Conquests: Maya and Spaniard in Yucatan, 1517-1570. Cambridge
University Press, England.
Cohen, M.N. and G.J. Armelagos (eds)
1984 Paleopathology at the Origins of Agriculture. Academic Press, Orlando.
Cohen, M.N., K. O’Connor, M. Danforth, K. Jacobi and C. Armstrong
1994 Health and death at Tipu. In In the Wake of Contact: Biological Responses to
Conquest, ed. by C.S. Larsen and G.R. Milner, 121-133. Wiley-Liss, New York.
Comar, C.L.
1963 Some overall aspects of strontium-calcium discrimination. In The Transfer of
Calcium and Strontium Across Biological Membranes, ed. by R.H., Wasserman,
405-417. Academic Press, New York.

354
Comar, C.L., R.S. Russell and R.H. Wasserman
1957 Strontium-calcium movement from soil to man. Science 126: 3272.
Comar, C.L., I.B. Whitney and F.W. Lengemann
1955 Comparative utilization of dietary Sr90
and calcium by developing rat fetus and
growing rat. Proc. Soc. Exp. Biol. Med. 88: 232-236.
Comar, C.L., R.H. Wasserman and A.R. Twardock
1961 Secretion of calcium and strontium into milk. Health Physics 7: 69-80.
Connor, M. and D. Slaughter
1984 A diachronic study of Inuit diets using trace element analysis. Arctic
Anthropology 21: 121-134.
Cook, D.C.
1976 Pathologic States and Disease Process in Illinois Woodland Populations: An
Epidemiologic Approach. Ph.D. Dissertation, Dept. of Anthropology,
University of Chicago, Chicago.
1981 Mortality, age-structure and status in the interpretation of stress indicators in
prehistoric skeletons: A dental example from the Lower Illinois Valley, in The
Archaeology of Death, ed. by R. Chapman, I. Kinnes and K. Randsborg, 133-
144. Cambridge University Press.
Cook, D.C. and J.E. Buikstra
1979 Health and differential survival in prehistoric populations: Prenatal dental
defects. Am. J. Phys. Anthropol. 51: 649-664.
Cook, N.D.
1998 Born to Die: Diseases and New World Conquest, 1492-1650. Cambridge:
Cambridge Univ. Press.
Corruccini, R.S., J.S. Handler and K.P. Jacobi
1985 Chronological distribution of enamel hypoplasias and weaning in a Caribbean
slave population. Human Biology 57: 699-711.
Cotzias, G.C.
1967 Trace substances in environmental health. Bioscience 17 (12): 920.
Cousens, S., B. Nacro, V. Curtis
1993 Prolonged breast-feeding: No association with increased risk of clinical
malnutrition in young children in Burkina Faso. Bull. World Health
Organization 71: 713-722.
Cox, A., F. Keenan, M. Cooke and J. Appleton
1996 Trace element profiling of dental tissues using laser ablation-inductively
coupled plasma-mass spectrometry. J. of Analytical Chemistry 334: 254-258.

355
Coyston, S., C.D. White and H.P. Schwarcz
1999 Dietary carbonate analysis of bone and enamel for two sites in Belize. In
Reconstructing Ancient Maya Diet, ed. by C.D. White, 221-243. University of
Utah Press, Salt Lake City.
Cross N.A., L.S. Hillman, S.H. Allen and G.F. Krause
1995 Changes in bone mineral density and markers of bone remodeling during
lactation and postweaning in women consuming high amounts of calcium. J
Bone Miner Res 10: 1312–20.
Cross N.A., L.S. Hillman, S.H. Allen, G.F. Krause and E.N. Vieira
1995 Calcium homeostasis and bone metabolism during pregnancy, lactation and
postweaning: A longitudinal study. Am. J. Clin. Nutr. 61: 514–23.
Cruwys, E. and R.A. Foley (eds)
1986 Teeth and Anthropology. British Archaeological Reports, International Series,
291. Oxford.
Cumming, R.G. and R.J. Klineberg
1993 Breastfeeding and other reproductive factors and the risk of hip fractures in
elderly women. International J. of Epidemiology 22(4): 684-691.
Cummings, S.R., M.C. Nevitt, W.S. Browner
1995 Risk factors for hip fracture in white women: Study of Osteoporotic Fractures
Research Group. New England J. Med. 332: 767-773.
Cunningham, A.S.
1981 Breastfeeding and morbidity in industrialized countries: An update. In Advances
in Maternal and Child Health, Vol. I, ed. By D.B. Jelliffe and E.F.P. Jelliffe,
128-168. Oxford University Press, Oxford.
1995 Breastfeeding: Adaptive behavior for child health and longevity. In
Breastfeeding: Biocultural Perspectives, ed. by P.L. Stuart-Macadam and K.A.
Dettwyler, 243-264. Aldine de Gruyter, New York.
Curzon, M.E.J. and T.W. Cutress (eds.)
1983 Trace Elements in Dental Disease. John Wright Press, New York.
Curzon, M.E.J. and J.D.B. Featherstone
1983 Chemical composition of enamel. In CRC Handbook of Experimental Aspects of
Oral Biochemistry, ed. by E.P. Lazzari, 123-135. CRC Press, Boca Raton, FLA.
Curzon, M.E.J. and F.L. Losee
1977 Strontium content of enamel and dental caries. Caries Research 11: 321-326.

356
Curzon, M.E.J. and P.C. Spector
1977 Enamel mottling in a high strontium area of the USA. Community Dent. Oral
Epidemiol. 5: 243-247.
Cutress, T.W.
1972 The inorganic composition and solubility of dental enamel from several specific
population groups. Arch. Oral Biol. 17: 93.
Cutress, T.W. and G.W. Suckling
1982 The assessment of non-carious defects of enamel. Int’l Dentistry Journal 2:
117-122.
Dallman, P.R.
1986 Iron deficiency in the weanling: A nutritional problem on the way to resolution.
Acta Paediatr. Scand. Suppl. 323: 59-67.
Danforth, M.E.
1986 Health patterns in the Classic Maya: Dental evidence. Paper presented at the
Annual Meeting of the American Anthropological Association, Philadelphia.
1989 Comparison of Health Patterns in Late Classic and Colonial Maya Populations
Using Enamel Microdefects. Ph.D. Dissertation, Indiana University,
Bloomington.
Danforth, M.E.
1997 Late Classic Maya health patterns: Evidence from enamel microdefects. In
Bones of the Maya Studies of Ancient Skeletons, ed. by S. L. Whittington and
D.M. Read, 127-137. Smithsonian Institution Press, Washington.
Danforth, M.E., K.P. Jacobi and M.N. Cohen
1997 Gender and health among the Maya of Tipu, Belize. Ancient Mesoamerica 8:13-
22.
Davis, M.K., D.A. Savitz and B.I. Graubard
1988 Infant feeding and childhood cancer. Lancet 2: 365-368.
Dean, M.C.
1987 Growth layers and incremental markings in hard tissues: A review of the
literature and some preliminary observations about enamel structure in
Paranthropus boisei. J. Human Evolution 16: 157-172.
Dean, M.C. and A.D. Beynon
1991 Histological reconstruction of crown formation times and initial root formation
times in a modern human child. Am. J. Phys. Anthropol. 86: 215-228.

357
Dedhiya, M.G., F. Young and W.I. Higuchi
1974 Mechanism of hydroxyapatite dissolution. The synergistic effects of solution
fluoride, strontium and phosphate. J. Phys. Chem 78: 1273-1279.
Delgado, H.L., V.E. Valverde, R. Martorell and R.E. Klein
1982 Relationship of maternal and infant nutrition to infant growth. Early Human
Development 6: 273-286.
Demirjian, A.
1986 Dentition. In Human Growth, A Comprehensive Treatise, ed. by F. Falkner and
J.M. Tanner, 2nd
ed., 269-298. Plenum Press, New York.
Demirjian, A. and G.-Y. Levesque
1980 Sexual differences in dental development and prediction of emergence. J. Dent.
Research 59: 1110-1122.
Denevan, W.M. (ed.)
1992 The Native Population of the Americas in 1492. Second ed. Madison: Univ. of
Wisconsin Press.
DeNiro, M.J.
1985 Postmortem preservation and alteration of in vivo bone collagen isotope ratios
in relation to paleodietary reconstruction. Nature 317: 806-809.
DeNiro, M.J. and S. Epstein
1978 Influence of diet on the distribution of carbon isotopes in animals. Geochimica
et Cosmochimica Acta 42: 495-506.
1981 Influence of diet on the distribution of nitrogen isotopes in animals. Geochimica
et Cosmochimica Acta 45: 341-351.
DeNiro, M.J. and C. Hastorf
1985 Alteration of 15
N/14
N and 13
C/12
C ratios of plant matter during the initial stages
of diagenesis: Studies using archaeological specimens from Peru. Geochim. et
Cosmochimica Acta 49: 97-115.
Denoyer, E.R., K.J. Fredeen and J.W. Hager
1991 Laser solid sampling for inductively coupled plasma-mass spectrometry.
Analytical Chemistry 63(8): 445-457.
DeSantiago, S., L. Alonso, A. Halhali, F. Larrea, F. Isoard and H. Bourges
2002 Negative calcium balance during lactation in rural Mexican women. Am. J.
Clin. Nutrition 76(4): 845-851.

358
Dettwyler, K.A.
1995 A time to wean: The hominid blueprint for the natural age of weaning in
modern human populations. In Breastfeeding: Biocultural Perspectives, ed. by
P.L. Stuart-Macadam and K.A. Dettwyler, 39-73. Aldine de Gruyter, New
York.
Dettwyler, K.A. and C. Fishman
1992 Infant feeding practices and growth. Annual Review of Anthropology 21: 171-
204.
Dewey, K.G., M.J. Heinig, L.A. Nommsen, J.M. Peerson and B. Lonnerdal
1992 Growth of breast-fed and formula-fed infants from 0 to 18 months: The
DARLING study. Pediatrics 89: 1035-1041.
Dobyns, H.F.
1983 Their Number Became Thinned: Native American Population Dynamics in
Eastern North America. Knoxville: Univ. of Tennessee Press.
Dolphin, A., A.H. Goodman and D. Amarasiriwardena
in press Variation in elemental intensities among teeth and between pre- and postnatal
regions of enamel. Submitted to American Journal of Physical Anthropology.
Dorea, J.G.
1999 Calcium and phosphorus in human milk. Nutrition Research 19(5): 709-739.
2000 Breast milk calcium and phosphorus concentrations. Nutrition 16(2): 146-147.
Duffy, L.C., M. Riepenhoff-Talty, T.E. Byers, L.J. La Scolea, M.A. Zielezny, D.M.
Dryja and P.L. Ogra
1986 Modulation of rotavirus enteritis during breastfeeding. Am. J. Dis. Child. 140:
1164-1168.
Dunn, F.L.
1965 On the antiquity of malaria in the western hemisphere. Human Biology 37: 385-
393.
Dupras, T.L., H.P. Schwarcz and S.I. Fairgrieve
2001 Infant feeding and weaning practices in Roman Egypt. Am. J. Phys. Anthropol.
115(3): 204-212.
Durrant, S.F.
1999 Laser ablation inductively coupled plasma mass spectrometry: Achievements,
problems, prospects. J. Analytical Atomic Spectrometry 14: 1385-1403.

359
Elias, R.W., Y. Hirao and C.C. Patterson
1982 The circumvention of the natural biopurification of calcium nutrient pathways
by atmospheric inputs of industrial lead. Geochemica Cosmochimica Acta 46:
2561-2580.
Ellison, P.T.
1995 Breastfeeding, fertility, and maternal condition. In Breastfeeding: Biocultural
Perspectives, ed. by P.L. Stuart-Macadam and K.A. Dettwyler, 305-346. Aldine
de Gruyter, New York.
Emery, K.
1990 Postclassic and Colonial Period Subsistence Strategies in the Southern Maya
Lowlands: Faunal Analyses from Lamanai and Tipu, Belize. M.A. thesis, Dept.
of Anthropology, University of Toronto, ON.
1999 Continuity and variability in Postclassic and Colonial animal use at Lamanai
and Tipu, Belize. In Reconstructing Ancient Maya Diet, ed. by C.D. White, 61-
82. University of Utah Press, Salt Lake City.
Evans, R.D., P. Richner and P.M. Outridge
1994 Applications of laser ablation inductively coupled mass spectrometry to the
determination of environmental contaminants in calcified biological structures.
J. of Analytical Atomic Spectrometry 9: 985-989.
Evans, R.D., P. Richner and PM Outridge
1995 Micro-spatial variations of heavy metals in the teeth of walrus as determined by
laser ablation ICP-MS: The potential for reconstructing a history of metal
exposure. Archives of Environmental Contamination and Toxicology 28: 55-60.
Ezzo, J.A.
1994 Putting the chemistry back into archaeological bone chemistry analysis:
Modeling potential paleodietary indicators. J. of Anthropological Archaeology
13: 1-34
FDI (Federation Dentaire Internationale)
1982 An epidemiological index of developmental defects of dental enamel (DDE).
Int’l Dental Journal 32: 159-167.
Fanning, E.A. and T. Brown
1971 Primary and permanent tooth development. Australian Dentistry Journal 67:
41-43.
Farnsworth, P., J.E. Brady, M.J. DeNiro and R.S. MacNeish
1985 A re-evaluation of the isotopic and archaeological reconstruction diet in the
Tehuacan Valley. American Antiquity 50: 102-116.

360
Farnum, J.F. and R.A. Benfer Jr.
1995 Age of pregnancies and age of weaning from prehistoric bone: Are Sr and Zn
indicators? Paper presented at the 64th
Annual Meeting of the Am. Assoc. of
Physical Anthropologists, Oakland, CA, March 30.
Farriss, N. M.
1984 Maya Society Under Colonial Rule: The Collective Enterprise of Survival.
Princeton University Press, Princeton, NJ.
1993 Persistent Maya resistance and cultural retention in Yucatan. In The Indian in
Latin American History: Resistance, Resilience, and Acculturation, ed. by J.E.
Kicza, 51-68. Scholarly Resources Inc, Wilmington, DE.
Featherstone, J.D.B. and D.G.A. Nelson
1980 The effect of F, Zn, Sr, Mg and Fe on the crystal-structural disorder in synthetic
carbonated apatites. Aust. J. Chem. 33: 2363.
Featherstone, J.D.B., D.G.A. Nelson and J.D. McLean
1981 An electron microscope study of modifications to defect regions in dental
enamel and synthetic apatites. Caries Res. 15: 278.
Fedrick, J. and P. Adelstein
1973 Influence of pregnancy spacing on outcome of pregnancy. Br. Med. J. 4: 753-
756.
Feldblum, P.J., J. Zhang, L.E. Rich, J.A. Fortney and R.V. Talmage
1992 Lactation history and bone mineral density among premenopausal women.
Epidemiology 3(6): 527-531.
Fergusson, D.M., A.L. Beautrais and P.A. Silva
1982 Breastfeeding and cognitive development in the first seven years of life. Soc.
Science and Med. 16: 1705-1708.
Fergusson, J.E. and N.G. Purchase
1987 The analysis and levels of lead in human teeth: A review. Environmental
Pollution 46: 11-44.
Fildes, V.A.
1986 Breasts, Bottles and Babies. Edinburgh University Press, Edinburgh, Scotland.
1988 Wet Nursing: A History from Antiquity to the Present. Basil Blackwell, Oxford.
1995 The culture and biology of breastfeeding: An historical review of western
Europe. In Breastfeeding: Biocultural Perspectives, ed. by P.L. Stuart-
Macadam and K.A. Dettwyler, 101-126. Aldine de Gruyter, New York.
FitzGerald, C.M.
1995 Tooth Crown Formation and the Variation of Enamel Microstructural Growth
Markers in Modern Humans. PhD. Dissertation, University of Cambridge.

361
FitzGerald, C.M.
1998 Do enamel microstructures have regular time dependency? Conclusions from
the literature and a large scale study. J. Human Evolution 35: 371-386.
FitzGerald, C.M., R.A. Foley and M.C. Dean
1996 Variation of circaseptan cross striation repeat intervals in the tooth enamel of
three modern human populations (abstract). Am. J. Phys. Anthropol., Suppl. 22:
104-105.
Fogel, M., N. Tuross and D. Owsley
1989 Nitrogen isotope tracers of human lactation in modern and archaeological
populations. Annual Report of the Director, Geophysical Laboratory 1988-
1989, 2150: 111-117.
Fosse, G., N.P.B. Justesen and G.B.R. Wesenberg
1981 Micro structure and chemical composition of fossil mammalian teeth. Calcified
Tissue International 33(5): 521-528.
Francalacci, P. and S.B. Tarli
1988 Multielementary analysis of trace elements and preliminary results on stable
isotopes in two Italian prehistoric sites: Methodological apects. In Trace
elements in Environmental History, ed. by G. Grupe and B. Herrmann, 41-52.
Springer-Verlag, Berlin.
Freudenheim, J.L., J.R. Marshall, S. Graham, R. Laughlin, J.E. Vena, E. Bandera, P.
Muti, M. Swanson and T. Nemoto
1994 Exposure to breast milk in infancy and the risk of breast cancer. Epidemiology
5(3): 324-331.
Friedlander, J.
1977 Malaria and demography in the lowlands of Mexico: An ethnohistorical
approach. In Culture, Disease and Healing, ed. by D. Landy, 113-119.
Macmillan Press, New York.
Fuchs, A.
1978 Strontium/calcium ratios in bone: An assessment of their use in paleodietary
research. MASCA Journal: 10-11. Univ. of Pennsylvania, Philadelphia.
Fuge, R., T.J. Palmer, N.J.G. Pearce and W.T. Perkins
1993 Minor and trace element chemistry of modern shells: A laser ablation
inductively coupled plasma mass spectrometry study. Appl. Geochem., 2
(suppl. issue): 111-116.
Furley, P.A. and A.J. Crosbie
1976 Geography of Belize. 2nd
Edition. Collins, London.

362
Garn, S.M., A.B. Lewis, K. Koski and D.L. Polacheck
1958 The sex differences in tooth calcification. J. Dent. Research 37: 561-567.
Gerhard, P.
1979 The Southeast Frontier of New Spain. Princeton University Press, Princeton, NJ.
Gerry, J.P. and M.S. Chesson
2000 Classic Maya diet and gender relationships. In Gender and Material Culture in
Archaeological Perspective, ed. by M. Donald and L. Hurcombe, 250-265. St.
Martin’s Press, New York.
Gibson, J.
1983 Summary of Hurricanes Recorded as Affecting Belize. RN & A Manuscript.
Gilbert, R.
1975 Trace elements analysis of three skeletal Amerindian populations at Dickson
Mounds. Ph.D. dissertation, Dept. of Anthropology, University of
Massachusetts, Amherst, MA.
1977 Applications of trace element research to problems in archeology. In
Biocultural Adaptation in Prehistoric America, ed. by R.L. Blakey, 85-100.
University of Georgia Press, Athens.
1985 Stress, paleonutrition, and trace elements. In The Analysis of Prehistoric Diets,
ed. by R.I. Gilbert and J.H. Mielke, 339-358. Academic Press, Orlando.
Gilbert, R.I. and J.H. Mielke (eds.)
1985 The Analysis of Prehistoric Diets. Academic Press, Orlando, FL.
Gilbert, C., J. Sealy and A. Sillen
1994 An investigation of barium, calcium and strontium as palaeodietary indicators in
the Southwestern Cape, South Africa. J. Arch. Science 21: 173-184.
Glass, R.I., A.M. Svennerholm, B.J. Stoll, M.R. Khan, K.M.B. Hossain, M.I. Huq and J.
Holmgren
1983 Protection against cholera in breast-fed children by antibodies in breast milk.
New England Journal of Medicine 308: 1389-1392.
Gleiser, I. and E.E. Hunt
1955 The permanent mandibular first molar: Its calcification, eruption and decay. Am.
J. Phys. Anthropol. 13: 253-283.
Goldman, A.S. and R.M. Goldblum
1990 Human milk: Immunologic-nutritional relationships. Ann. New York Academy
of Science 587: 235-245.

363
Goldman, A.S., R.M. Goldblum and L.A. Hanson
1990 Anti-inflammatory systems in human milk. In Antioxidant Nutrients and
Immune Functions, ed. By A. Bendich, M. Phillips and R.P. Tengerdy, 69-76.
Plenum Press, New York.
Goodman, A.H. and G.J. Armelagos
1988 Childhood stress and decreased longevity in a prehistoric population. American
Anthropologist 90: 936-44.
Goodman, A.H. and G.J. Armelagos
1989 Infant and childhood morbidity and mortality risks in archaeological
populations. World Archaeology 21(2): 225-243.
Goodman, A.H., G.J. Armelagos and J.C. Rose
1980 Enamel hypoplasias as indicators of stress in three prehistoric populations from
Illinois. Human Biology 52: 515-528.
1984 The chronological distribution of enamel hypoplasias from prehistoric Dickson
Mounds populations. Am. J. Phys. Anthropol. 65: 259-266.
Goodman, A.H. and L.L. Capasso (eds)
1992 Recent Contributions to the Study of Enamel Developmental Defects. Journal of
Paleopathology, Monographic Publications, 2.
Goodman, A.H., A.E. Dolphin, D.D. Amarasiriwardena, R. Klein, J.R. Backstrand, J.B.
Reid Jr. and P. Outridge
2003 Tooth rings: Dental enamel as a chronological biomonitor of elemental
absorption from pregnancy to adolescence. J. Children’s Health vol. 1, no. 2:
203-214.
Goodman, A.H. and J.C. Rose
1990 Assessment of systemic physiological perturbations from dental enamel
hypoplasias and associated histological structures. Yearbook of Physical
Anthropology 33: 59-110.
Goodman, A.H. and R.J. Song
1999 Sources of variation in estimated ages at formation of linear enamel
hypoplasias. In Human Growth in the Past: Studies from Bones and Teeth, ed.
by R. Hoppa and C. FitzGerald, 210-239. Cambridge University Press, England.
Graham, E.
1991 Archaeological insights into colonial period Maya life at Tipu, Belize. In
Columbian Consequences, vol. 3, The Spanish Borderlands in Pan-American
Perspective, ed. by D.H. Thomas, 319-335. Smithsonian Institution Press,
Washington, DC.

364
Graham, E., D.M. Pendergast and G.D. Jones
1989 On the fringes of conquest: Maya-Spanish contact in colonial Belize. Science
246: 1254-1259.
Gratuze, B., M. Blet-Lemarquand and J.-N. Barrandon
2000 Mass spectrometry with laser sampling: A new tool to characterize
archaeological materials. J. of Radioanalytical and Nuclear Chemistry 247(3):
645-656.
Gray, A.L.
1985 Solid sample introduction by laser ablation for inductively coupled plasma
source mass spectrometry. Analyst May, vol. 110: 551-556.
Grine, F.E. and R.F. Kay
1988 Early hominid diets from quantitative image analysis of dental microwear.
Nature (London) 333: 765-768.
Grinquist, H.
1947 Birth and Childhood Among the Arabs. Soderstrom and Company, Helsingfors.
Grupe, G.
1986 Rekonstruktion bevolkerungsbiologischer parameter aus dem elementgehalt
boldengelagerter Knochen. In Innovative Trends in Prehistoric Anthropology,
ed. by B. Herrmann 7:39-44. Mitt Bert Ges Anthrop Ethnol Urgesch.
1995 Reconstructing migration in the Bell Beaker period by 87Sr/86Sr isotope ratios
in teeth and bones. In Proceedings of the 10th
International Symposium on
Dental Morphology, ed. by R.J. Radlandski and H. Renz, 338-342. ‘M’
Marketing Services, C & M Brunne GbR, Berlin.
Grupe, G. and B. Herrmann (eds)
1988 Trace Elements in Environmental History. Springer-Verlag, Berlin.
Grupe, G., T.D. Price, P. Schroter, F. Sollner, C.M. Johnson and B.L. Beard
1997 Mobility of bell beaker people revealed by strontium isotope ratios of tooth and
bone: A study of Southern Bavarian skeletal remains. Appl. Geochem 12: 517-
525.
Guita, J.L.
1984 Oral Pathology, 2nd
edition. Williams and Wilkins, Baltimore.
Gulson, B.L., C.W. Jameson and B.R. Gillings
1997 Stable lead isotopes in teeth as indicators of past domicile: A potential new tool
in forensic science? Journal Forensic Science 42: 787-791.

365
Gulson, B.L. and D. Wilson
1994 History of lead exposure in children revealed from isotopic analyses of teeth.
Archives of Environmental Health 49: 279-283.
Gunther, D. and B. Hattendorf
2001 Elemental fractionation in LA-ICP-MS. In Laser-Ablation-ICPMS in the Earth
Sciences: Principles and Applications, ed. by P. Sylvester, 83-91. Short Course
Series, Vol. 29. Mineralogical Association of Canada, St. Johns, Newfoundland.
Gunther, D., I. Horn and B. Hattendorf
2000 Recent trends and developments in laser ablation-ICP-mass spectrometry.
Fresenius J. Anal. Chem 368: 4-14.
Gur, A., R. Cevik, K. Nas, A.J. Sarac, S. Ataoglu, M. Karakoc, A. Can and F. Gurkan
2003 The influence of duration of breastfeeding on bone mass in postmenopausal
women of different age groups. Int’l J. Clin. Practice 57(2): 82-6.
Gysi, A.
1931 Metabolism in adult enamel. Dental Digest 37: 661-668.
Habicht, J.-P., J. DaVanzo, W.P. Butz and L. Meyers
1985 The contraceptive role of breastfeeding. Population Studies 39: 213-232.
Hahn-Zoric, M., F. Fulconis, I. Minoli, G. Moro, B. Carlsson, M. Bottiger, N. Raiha and
L.A. Hanson
1990 Antibody responses to parenteral and oral vaccines are impaired by
conventional and low protein formulas as compared to breast-feeding. Acta
Paediatrica Scandinavica 79: 1137-1142.
Hanihara, K.
1979 Dental traits in Ainu, Australian Aborigines and New World populations. In The
First Americans: Origins, Affinities and Adaptations, ed. by W.S. Laughlin and
A.B. Harper, 125-134. Gustav Fischer, New York.
Hanson, D.B. and J.E. Buikstra
1987 Histomorphological alteration in buried human bone from the Lower Illinois
Valley: Implications for paleodietary research. J. Arch. Science 14: 549-563.
Harmon, A.M. and J.C. Rose
1988 The role of dental microwear analysis in the reconstruction of prehistoric diet.
In Diet and Subsistence: Current Archaeological Perspectives, ed. by B.V.
Kennedy and G.M. LeMoine, 267-272. Archaeological Proceedings of the 19th
Conference, University of Calgary, AB.
Harrison, G.E., A. Sutton, H. Shepherd and E.M. Widdowson
1965 Strontium balance in breast-fed babies. British J. Nutrition 19: 111-117.

366
Hartshorn, G., L. Nicolait, L. Harshorn, G. Bevier, R. Brightman, J. Cal, A. Cawich, W.
Davidson, R. DuBois, C. Dyer, J. Gibson, W. Hawley, J. Leonard, R. Nicolait, D. Weyer,
H. White and C. Wright
1984 Belize: Country Environmental Profile A Field Study (USAID). R. Nicolait and
Associates, Belize City.
Harvey, P.H. and T.H. Clutton-Brock
1985 Life history variation in primates. Evolution 39(3): 559-581.
Hausman, B.L.
2003 Mother’s Milk: Breastfeeding Controversies. Routledge, New York.
Haverkort, C.M.
2001 Enamel trace element composition and palaeodietary studies-an exploratory
model. Ph.D. dissertation, University of Alberta, Edmonton.
Hayslip, C.C., T.A. Klein, H.L. Wray and W.E. Duncan
1989 The effects of lactation on bone mineral content in healthy postpartum women.
Obstet. Gynecol. 73: 588-592.
Heaney, R.P. and T.G. Skillman
1971 Calcium metabolism in normal human pregnancy. J. Clin Endocrinol. 33: 661-
670.
Heaton, T.H.E., J.C. Vogel, G. von la Chevallerie and G. Collet
1986 Climatic influence on the isotopic composition of bone nitrogen. Nature 322:
822-823.
Hellmuth, N.
1977 Cholti-Lacandon (Chiapas) and Peten-Ytza agriculture, settlement pattern and
population. In Social Process in Maya Prehistory, ed. by N. Hammond , 421-
448. Academic Press, New York.
Helmuth, H. and D.M. Pendergast
1989 Lamanai tomb N9-56/1: Analysis of the skeletal evidence. Ossa 13: 109-117.
Herrera, M.G., J.O. Mora, B. de Paredes and M. Wagner
1980 Maternal weight/height and the effect of food supplementation during
pregnancy and lactation. In Maternal Nutrition During Pregnancy and
Lactation, ed. by H. Aebi and R. Whitehead, 252-263. Hans Huber Publishers,
Bern.
Herring, D.A., S.R. Saunders and M.A. Katzenberg
1998 Investigating the weaning process in past populations. Am. J. Phys. Anthropol.
105: 425-439.

367
Hess, A.F., J.M. Lewis and B. Roman
1932 A radiographic study of calcification of the teeth from birth to adolescence.
Dent. Cosmos 74: 1053-1061.
Hillson, S.
1979 Diet and dental disease. World Archaeology 11: 147-162.
1986 Teeth. Cambridge University Press, Cambridge, England.
1996 Dental Anthropology. Cambridge University Press, Cambridge, England.
Hobcraft, J.
1987 Does family planning save children’s lives? Technical background paper
prepared for the International Conference on Better Health for Women and
Children Through Family Planning, Nairobi, Kenya, October, 1987. Population
Council, New York.
Hodges, R.M., N.S. MacDonald, R. Nusbaum, R. Stearns, F. Ezmirlain, P. Spain and C.
MacArthur
1950 The strontium content of human bones. J. Biol. Chem. 185: 519-524.
Hreshchyshyn, M.M., A. Hopkins, S. Zylstra and M. Anbar
1988 Associations of parity, breast-feeding, and birth control pills with lumbar spine
and femoral neck bone densities. Am. J. of Obstetrics and Gynecology 159(2):
318-322.
Huhne-Osterloh, G. and G. Grupe
1989 Causes of infant mortality in the Middle Ages revealed by chemical and
palaeopathological analyses of skeletal remains. Zeitschrift für Morphologie
und Anthropologie 77: 247-258.
Hunt, M. Espejo-Ponce
1974 Colonial Yucatan: Town and Region in the Seventeenth Century. PhD.
Dissertation, University of California, Los Angeles.
Huo, D., D.S. Lauderdale and L. Li
2003 Influence of reproductive factors on hip fracture risk in Chinese women.
Osteoporosis Int’l 14(8): 694-700.
Imai, N.
1992 Microprobe analysis of geological materials by laser ablation inductively
coupled plasma mass spectrometry. Anal. Chimica Acta 269: 263-268.
Ing, R., J.H.C. Ho and N.L. Petrakis
1977 Unilateral breast-feeding and breast cancer. Lancet 2: 124-127.

368
Ingersoll, R.J. and R.H. Wasserman
1971 Vitamin D3-induced calcium-binding protein: Protein characteristics,
confirmational effects, and other properties. J. Biol. Chem. 246: 2808-2814.
Institute of Medicine
1991 Nutrition During Lactation. National Academy Press, Washington, DC.
Izurieta, L.M. and L.B. Larson-Brown
1995 Child feeding practices in Guatemala. Ecol. Food Nutr. 33: 249-262.
Jackson, S.
2001 The application of Nd: YAG lasers in LA-ICP-MS. In Laser-Ablation-ICPMS
in the Earth Sciences: Principles and Applications, ed. by P. Sylvester, 29-46.
Short Course Series, Vol. 29. Mineralogical Association of Canada, St. Johns,
Newfoundland.
Jacobi, K.P.
2003 Last Rites of the Tipu Maya: Genetic Structuring in a Colonial Cemetery.
University of Alabama Press, Tuscaloosa.
Jenkins, G.N.
1978 The Physiology and Biochemistry of the Mouth, 4th
ed. Blackwell Scientific,
Oxford, England.
Johansson, S.A.E., J.L. Campbell and K.G. Malmqvist (eds.)
1995 Particle-Induced X-ray Emission Spectrometry (PIXE). Wiley, New York.
Jones, G.D.
1977 Anthropology and History in Yucatan (editor). University of Texas Press,
Austin.
1989 Maya Resistance to Spanish Rule: Time and History on a Colonial Frontier.
University of New Mexico Press, Albuquerque.
1998 The Conquest of the Last Maya Kingdom. Stanford University Press, Stanford,
CA.
Jones, G.D., R.R.Kautz and E. Graham
1986 Tipu: A Maya town on the Spanish colonial frontier. Archaeology 39: 40-47.
Kalkwarf H.J. and B.L. Specker
1995 Bone mineral loss during lactation and recovery after weaning. Obstet Gynecol
86: 26–32.

369
Kang, D., D. Amarasiriwardena and A.H. Goodman
2004 Application of laser ablation–inductively coupled plasma-mass spectrometry
(LA–ICP–MS) to investigate trace metal spatial distributions in human tooth
enamel and dentine growth layers and pulp. Analytical and Bioanalytical
Chemistry 378(6): 1608-1615.
Karra, M.V., A. Kirksey, O. Galal, N.S. Bassily, G.G. Harrison and N.W. Jerome
1988 Zinc, calcium and magnesium concentrations in milk from American and
Egyptian women throughout the first 6 months of lactation. Am. J. Clin Nutr 47:
642-8.
Katz, S.H., M.L. Hediger and L.A. Valleroy
1974 Traditional maize processing techniques in the New World. Science 184: 765-
763.
1975 The anthropological and nutritional significance of traditional maize processing
techniques in the New World. In Biosocial Interrelations in Population
Adaptation, ed. by E.S. Watts, F.E. Johnston and G.W. Lasker, 195-232.
Aldine, Chicago, IL.
Katzenberg, M.A.
1984 Chemical Analysis of Pre-Historic Bone from Five Temporally Distinct
Populations in Southern Ontario. National Museum of Man Mercury Series No.
129, National Museums of Canada.
Katzenberg, M.A. and S. Pfeiffer
1995 Nitrogen isotope evidence for weaning age in a nineteenth century Canadian
skeletal sample. In Bodies of Evidence, ed. by A.L. Grauer, 221-235. John
Wiley and Sons, New York.
Katzenberg, M.A., D.A. Herring and S.R. Saunders
1996 Weaning and infant mortality: Evaluating the skeletal evidence. Yearbook of
Phys. Anthropol. 39: 177-199.
Katzenberg, M.A., S. Saunders and W.R. Fitzgerald
1993 Age differences in stable carbon and nitrogen isotope ratios in a population of
prehistoric maize horticulturalists. Am. J. Phys. Anthropol. 90(3): 267-281.
Keeley, H, G. Hudson and J. Evans
1977 Trace element content of human bones in various states of preservation. J.
Archaeological Science 4:19-24.
Kelley, M.A. and C.S. Larsen (eds)
1991 Advances in Dental Anthropology. Wiley-Liss, New York.

370
Kent, G.N., R.I. Price, D. Gutterdige
1990 Human lactation: Forearm trabecular bone loss, increased bone turnover, and
renal conservation of calcium and inorganic phosphate with recovery of bone
mass following weaning. J Bone Mineral Research 5: 361–9.
King J.C., B.P. Halloran, N. Huq, T. Diamond and P.E. Buckandahl
1992 Calcium metabolism during pregnancy and lactation. In Mechanisms Regulating
Lactation and Infant Nutrition Utilization, ed. by M.F. Picciano and B.
Lonnerdal, 129-146. Wiley-Liss, New York.
Klepinger, L.L.
1993 Culture, health, and chemistry: A technological approach to discovery. In
Investigations of Ancient Human Tissue, ed. By M.K. Sandford, 167-180.
Gordon and Breach, Langhorne, PA.
Klepinger, L.L., J.K. Kuhn and W.S. Williams
1986 An elemental analysis of archaeological bone from Sicily as a test of
predictability of diagenetic change. Am. J. Phys. Anthropol. 70: 325-331.
Knizhnikov, V.A. and A.N. Marei
1967 Strontium metabolism in Man. In Strontium Metabolism, ed. By J.M.A.
Lenihan, J.F. Loutit and J.H. Martin, 71-82. Academic Press, New York.
Knodel, J. and H. Kintner
1977 The impact of breast feeding patterns on the biometric analysis of infant
mortality. Demography 14: 391-409.
Koetting, C.A. and G.M. Wardlaw
1988 Wrist, spine, and hip bone density in women with variable histories of lactation.
Am. J. Clin. Nutr. 48: 1479-1481.
Kohn, M.J., M.J. Schoeninger and W.W. Barker
1999 Altered states: Effects of diagenesis on fossil tooth chemistry. Geochimica et
Cosmochimica Acta 63(18): 2737-2747.
Konner, M. and C. Worthman
1980 Nursing frequency, gonadal function, and birth spacing among !Kung. In
Kalahari Hunter-Gatherers, ed. by R.B. Lee and I. DeVore, 218-299. Harvard
University Press, Boston, MA.
Kostial, K., N. Gruden and A. Durakovic
1969 Intestinal absorption of calcium 47 and strontium 85 in lactating rats. Calc. Tis.
Tes. 4: 13-19.

371
Krachler, M., E. Rossipal and D. Micetic-Turk
1999 Trace element transfer from the mother to the newborn-investigations of triplets
of colostrums, maternal and umbilical cord sera. European Journal of Clinical
Nutrition 53: 486-494.
Kramer, M.S.
1988 Does breast feeding help protect against atopic disease? Biology, methodology,
and a golden jubilee of controversy. J. Pediatr. 112: 181-190.
Krebs N.F., C.J. Reidinger, A.D. Robertson and M. Brenner
1997 Bone mineral density changes during lactation: Maternal, dietary, and
biochemical correlates. Am. J. Clin. Nutr. 65: 1738–46.
Krueger, H.W. and C.H. Sullivan
1984 Models for carbon isotope fractionation between diet and bone. In Stable
Isotopes in Nutrition. ACS Symposium, ed. by J.F. Turnland and P.E. Johnson,
205-220. American Chemical Society, Washington, DC.
Kuhnlein, H.V.
1981 Dietary mineral ecology of the Hopi. J. Ethnobiology 1: 84-94.
Kuhnlein, H.V. and D.H. Calloway
1979 Adventitious mineral elements in Hopi Indian diets. J. Food Science 44(1): 282.
Kyle, J.H.
1986 Effect of post-burial contamination on the concentrations of major and minor
elements in human bones and teeth: The implications for palaeodietary research.
J. of Archaeological Science 13: 403-416.
Lallo, J.
1973 The Skeletal Biology of Three Prehistoric American Indian Societies from
Dickson Mounds. Ph.D. Dissertation, Dept. of Anthropology, University of
Massachusetts, Amherst.
Lalueza Fox, C., J. Juan and R.M. Albert
1996 Phytolith analysis on dental calculus, enamel surface and burial soil:
Information about diet and paleoenvironment. Am. J. Phys. Anthropol. 101:
101-114.
Lambert, J.B., S.V. Simpson, C.B. Szpunar and J.E. Buikstra
1984 Copper and barium as dietary discriminants: the effect of diagenesis.
Archaeometry 26: 131-38.
1985 Bone diagenesis and dietary analysis. J. of Human Evolution 14: 477-482.

372
Lambert, J.B., C.B. Szpunar and J.E. Buikstra
1979 A biochemical analysis of excavated bone from Middle and Late Woodland
sites. Archaeometry 21: 115-29.
Lambert, J.B., S.M. Vlasak, A.C. Thometz and J.E. Buikstra
1982 A comparative study of the chemical analysis of ribs and femurs in Woodland
populations. Am. J. Phys. Anthropol. 59: 289-94.
Lambert, J.B., J.M. Weydert, S.R. Williams and J.E. Buikstra
1990 Comparison of methods for the removal of diagenetic material in buried bone. J.
of Archaeological Science 17: 453-468.
Lambert, J.D.H.
1985 The ecological consequences of ancient Maya agricultural practices in Belize.
In Prehistoric Agriculture in the Tropics, ed. by I.S. Farrington, 113-125.
B.A.R. International Series, no. 232, pt. 1.
Lambert, J.D.H. and J.T. Arnason
1978 Distribution of vegetation on Maya ruins and its relationship to ancient land use.
Turrialba 28 (1): 33-41.
1979 Drained fields in the New River flood plain and their agricultural potential-past
and present. Ms. on file, Dept. of Biology, Carleton University, Ottawa, ON.
Lambert, J.D.H., J.T. Arnason and A.H. Siemens
1984 Ancient Maya drained field agriculture: Its possible application today in the
New River flood-plain, Belize, C.A. Agri. Ecosys. and Environ. 11: 67-84.
Landa, D. de
1566 Relacion de las Cosas de Yucatan. Edited and translated by A.M. Tozzer
(1941). Peabody Museum of American Archaeology and Ethnology, Harvard
University, Cambridge, MA.
1978 Yucatan Before and After the Conquest. Translated by W. Gates, Dover
Publications, New York.
Lane, D.W. and D.F. Peach
1997 Some observations on the trace element concentrations in human dental enamel.
Biological Trace Element Research 60: 1-11.
Lang, C.A.
1990 The Dental Morphology of the Ancient Maya from Lamanai and Tipu, Belize:
A Study of Population Movement. Unpublished Master's Thesis, Dept. of
Anthropology, Trent University, Peterborough, ON.
Lanphear, K.M.
1990 Frequency and distribution of enamel hypoplasias in a historic skeletal sample.
Am. J. Phys. Anthropol. 81: 35-43.

373
Larsen, C.S.
1997 Bioarchaeology: Interpreting Behavior from the Human Skeleton. Cambridge
University Press, Cambridge, England.
Larsen, C.S. and G.R. Milner (eds.)
1994 In the Wake of Contact: Biological Responses to Conquest. Wiley-Liss, New
York.
Laskey, M.A., A. Prentice, LA. Hanratty, L.M.A. Jarjou, B. Dibba, S.R. Beavan and T.J.
Cole
1998 Bone changes after 3 mo of lactation: Influence of calcium intake, breast-milk
output, and vitamin D-receptor genotype. Am. J. Clin. Nutr. 67(4): 685-692.
LaVigne, M.
2002 Sr/Ca Ratios in Modern Mexican Diets Dominated by Alkali-Processed Maize:
Implications for paleodietary reconstructions. Unpublished BA thesis,
Hampshire College, Amherst, MA.
Lawrence, R.A.
1989 Breastfeeding: A Guide for the Medical Profession, 3rd
ed. C.V. Mosby, St
Louis, MO.
Lechtig, A. and R.E. Klein
1980 Maternal food supplementation and infant health: Results of a study in rural
areas of Guatemala. In Maternal Nutrition During Pregnancy and Lactation, ed.
By H. Aebi and R. Whitehead, 285-313. Hans Huber Publishers, Bern.
Lee, K.M., J. Appleton, M. Cooke, F. Keenan, K. Sawicka-Kapusta
1999 Use of laser ablation inductively coupled plasma mass spectrometry to provide
element versus time profiles in teeth. Analytica Chimica Acta 395: 179-185.
Lee, R.
1980 Lactation, ovulation, infanticide, and women’s work: A study of hunter-gatherer
regulation. In Biosocial Mechanisms of Population Regulation, ed. By M.
Cohen, R.S. Malpass and H.G. Klein, 321-348. Yale Univ. Press, New Haven,
CT.
Lee-Thorp, J.A.
1989 Stable Carbon Isotopes in Deep Time, The Diets of Fossil Fauna and Hominids.
Ph.D. Dissertation, Archaeology Dept., University of Cape Town, South Africa.
LeGeros, R.Z., M.A. Miravite and G. Bonel
1976 CO3 in apatites: Influence of other ions on carbonate incorporation. IADR Abst.
#955, J. Dental Research 55: 300.

374
LeGeros, R.Z., M.A. Miravite, G.B. Quirologico and M.E.J. Curzon
1977 The effect of some trace elements on the lattice parameters of human and
synthetic apatites. Calcified Tissue Research 22: 362-367.
Lengemann, F.W.
1960 Studies on the discrimination against strontium by bone grown in vitro. J. Biol.
Chem. 235: 1859-1961.
1963 Overall aspects of calcium and strontium absorption. In The Transfer of
Calcium and Strontium Across Biological Membranes, ed. by R.H. Wasserman,
85-96. Academic Press, New York.
Lengemann, F.W., C.L. Comar and R.H. Wasserman
1957 Absorption of calcium and strontium from milk and non-milk diets. J. Nutrition
61: 571.
Lentz, D.L.
1999 Plant resources of the ancient Maya: The paleoethnobotanical evidence. In
Reconstructing Ancient Maya Diet, ed. by C.D. White, 3-18. University of Utah
Press, Salt Lake City.
Lindsay, W.L.
1979 Chemical Equilibria in Soils. Wiley, New York.
Little, M.F. and K. Barrett
1976 Strontium and fluoride content of surface and inner enamel versus caries
prevalence in the Atlantic coast of the USA. Caries Research 10: 297-307.
Liversidge, H.M.
1994 Accuracy of age estimation from developing teeth of a population of known age
(0-5.4 years). Int. J. Osteoarchaeology 4: 37-45.
Liversidge, H.M.
1995 Crown formation times of the permanent dentition and rooth extension rate in
humans. In Aspects of Dental Biology: Paleontology, Anthropology and
Evolution, ed. by J. Moggi-Cecchi, 267-275. International Institute for the Study
of Man, Florence, Italy.
Lizana, B. de
1633 Historia de Yucatan: Devocionario de Nuestra Senora de Izamal y Conquista
Espiritual, 2nd Ed. Mexico: Museo Nacional, 1893.
Locke, S.E. and M. Hornig-Rohan
1983 Mind and Immunity: Behavioral Immunology, An Annotated Bibliography,
1976-1982. Praeger, New York.

375
Lonnerdal, B.
1986a Effects of maternal dietary intake on human milk composition. J. Nutr. 116:
499-513.
1986b Effects of maternal nutrition on human lactation. In Human Lactation 2:
Maternal and Environmental Factors, ed. By M. Hamosh and A.S. Goldman,
301-323. Plenum Press, NY.
Lonnerdal, B., C.L. Keen and L.S. Hurley
1981 Iron, copper, zinc and manganese in milk. Annu. Rev. Nutr. 1: 149-174.
Lopez de Cogolludo, D.
1654 Historia de Yucatan. J. Garcia Infanzon, Madrid, Spain 1688.
1688 Los Tres Siglos de la Dominacion Espanola en Yucatan, o sea Historia de esta
Provincia. 1st edition Madrid 1688 (5
th edition, 2 vols: Graz 1971).
Losee, F.L. and B.L. Adkins
1968 Anti-cariogenic effect of minerals in food and water. Nature 219: 630-631.
Losee, F.L., M.F. Little-McClellan and G.M. Orbell
1971 Strontium content of teeth related to geologic environment. IADR Abst. # 638,
J. Dent. Res 50: 123.
Loten, H.S.
1985 Lamanai Postclassic. In The Lowland Maya Postclassic, ed. by A.F. Chase and
P.M. Rice, 85-90. University of Texas Press, Austin.
Lough, S.A., J. Rivera and C.L. Comar
1963 Retention of strontium, calcium, and phosphorus in human infants. Society of
Experimental Biology and Medicine: New York Proceedings 112: 631-636.
Loutit, J.F.
1967 Strontium-90 from fall-out in human bone. In Strontium Metabolism, ed. by
J.M.A. Lenihan, J.F. Loutit and J.G. Martin, 41-45. Academic Press, New York.
Lovell, W.G.
1985 Conquest and Survival in Colonial Guatemala: A Historical Geography of the
Cuchumatan Highlands, 1500-1821. McGill-Queen’s University Press,
Montreal, QC.
Lucas, A., R. Morley, T.J. Cole, G. Lister and C. Leeson-Payne
1992 Breast milk and subsequent intelligence quotient in children born preterm.
Lancet 339: 261-264.

376
Lukacs, J.R.
1989 Biological affinities from dental morphology: The evidence from Neolithic
Mehrgarh. In Old Problems and New Perspectives in the Archaeology of South
Asia, ed. by J.M. Kenoyer, 75-88. Wisconsin Archaeological Reports, 2.
Lukacs, J.R.
1992 Dental paleopathology and agricultural intensification in South Asia: New
evidence from Bronze Age Harappa. Am. J. Phys. Anthropol. 87: 133-150.
Lukacs, J.R. and B.E. Hemphill
1991 The dental anthropology of prehistoric Baluchistan: A morphometric approach
to the peopling of South Asia. In Advances in Dental Anthropology, ed. by M.A.
Kelley and C.S. Larsen, 77-119. Wiley-Liss, New York.
1993 Odontometry and biologic affinities among South Asians: An analysis of three
ethnic groups from northwest India. Human Biology 65: 279-325.
Lukacs, J.R. and J.N. Pal
1993 Mesolithic subsistence in north India: Inferences from dental attributes. Current
Anthropology 34: 745-765.
Lynott, M.J., T.W. Boutton, J.E. Price and D.E. Nelson
1986 Stable carbon isotopic evidence from maize agriculture in southeast Missouri
and Northeast Arkansas. American Antiquity 51: 51-65.
Malik, S.R. and J.H. Fremlin
1974 A study of lead distribution in human teeth, using charged particle activation
analysis. Caries Research 8: 283-292.
Manter, H.W.
1967 Some aspects of the geographical distribution of parasites. J. of Parasitology
53: 3-9.
Marcus, C.S. and F.W. Lengemann
1962 Absorption of Ca45
and Sr85
from solid and liquid food at various levels of the
alimentary tract of the rat. J. Nutr. 66: 155-160.
Marcus, C.S. and R.H. Wasserman
1965 Comparison of intestinal discrimination between calcium 47, strontium 85 and
barium 133. Amer. J. Physiol. 209: 973-977.
Marcus, J.
1982 The plant world of the sixteenth- and seventeenth- century Lowland Maya. In
Maya Subsistence: Studies in Memory of Dennis E. Puleston, ed. by K.
Flannery, PAGES. Academic Press, New York.

377
Marks, M.K. and J.C. Rose
1985 Scanning electron microscopy of Wilson bands and enamel hypoplasias. Am. J.
Phys. Anthropol. 66(2): 202.
Martorell, R., C. Yarbrough, A. Lechtig, J.P. Habicht and R.E. Klein
1975 Diarrheal diseases and growth retardation in preschool Guatemalan children.
Am. J. Phys. Anthropol. 43: 341-346.
Martorell, R. and C. O'Gara
1985 Breastfeeding, infant health and socioeconomic status. Medical Anthropology 9:
173-181.
Massler, M. and I. Schour
1946 The appositional life span of the enamel and dentin-forming cells. I. Human
deciduous teeth and first permanent molars. J. of Dental Research 25: 145-150.
Massler, M., I. Schour and H.G. Poncher
1941 Developmental pattern of the child as reflected in the calcification pattern of the
teeth. American Journal of Diseases of Children 62: 33-67.
Mata, L.J.
1990 Breast-feeding, infections and infant outcomes: An international perspective. In
Breastfeeding, Nutrition, Infection and Infant Growth in Developed and
Emerging Countries, ed. by S.A. Atkinson, L.A. Hanson and R.K. Chandra.
ARTS Bio-medical Publisher, St. John’s, Nfld
Mata, L.J., J.J. Urrutia and J.E. Gordon
1967 Diarrhoeal disease in a cohort of Guatemalan village children observed from
birth to age two years. Trop. Geogr. Med. 19: 247-257.
Mata, L.J., J.J. Urrutia and B. Garcia
1977 Effects of infection on food intake and the nutritional state: Perspectives as
viewed from the village. Am. J. Clin. Nutr. 30: 1215.
Mays, S.
2003 Bone strontium: calcium ratios and duration of breastfeeding in a Mediaeval
skeletal population. J. Arch. Science 30(6): 731-741.
McCaslin F.E. and J.M. Janes
1959 The Effect of strontium lactate in the treatment of osteoporosis. Proc Staff
Meeting Mayo Clin. 34: 329-334.
1981 Effects of stable strontium in treatment of osteoporosis. In Handbook of Stable
Strontium, ed. by S.K. Skoryna. Plenum Press, New York.

378
McKee, L.
1984 Sex differentials in survivorship and the customary treatment of infants and
children. Medical Anthropology 8(2): 91-103.
McKinley, J.I.
1994 The Anglo-Saxon Cemetery at Spong Hill, North Elmham. Part VIII: The
Cremations. East Anglian Archaeology Report No. 69. Field Archaeology
Division, Norfolk Museums Service, Dereham, England.
Menczel, J. and E. Mor
1972 The effect of thyrocalcitonin on calcium binding protein and on strontium
binding protein. In Proceedings of the 2nd
International Symposium on
Strontium Metabolism. Glasgow, Scotland.
Menon, L., F. Anwar, S. Amin and S. Siew
2003 The Breastfeeding Movement: A Source Book. World Alliance for Breastfeeding
Action (WABA), Penang, Malaysia.
Micozzi, M.S.
1995 Breast cancer, reproductive biology, and breastfeeding. In Breastfeeding:
Biocultural Perspectives, ed. by P.L. Stuart-Macadam and K.A. Dettwyler, 347-
384. Aldine de Gruyter, New York.
Miksicek, C.H., E.S. Wing and S.J. Scudder
1991 The ecology and economy of Cuello. In Cuello: An Early Maya Community in
Belize, ed. by N. Hammond, 70-84. Harvard University Press, Cambridge, MA.
Miller, A.G. and N.M. Farriss
1979 Religious syncretism in colonial Yucatan: The archaeological and
ethnohistorical evidence from Tancah, Quintana Roo. In Maya Archaeology and
Ethnohistory, ed. by N. Hammond and G.R. Willey, 223-240. Univ. of Texas
Press, Austin, TX.
Mitchell, R.L.
1955 Trace elements in soil. In Trace Elements in Soil, ed. by F.E. Bear, 253-285.
Reinhold, New York.
1957 The trace element content of plants. Research 10: 357-362.
Moggi-Cecchi, J. (ed.)
1995 Aspects of Dental Biology: Palaeontology, Anthropology and Evolution.
International Institute for the Study of Man, Florence, Italy.
Moggi-Cecchi, J., E. Pacciani and J. Pinto-Cisternas
1994 Enamel hypoplasia and age at weaning in 19th
century Florence, Italy. Am. J.
Phys. Anthropol. 93: 299-306.

379
Mølbak, K., A. Gottschau, P. Aaby, N. Højlyng, L. Ingholt and A.P.J. da Silva
1994 Prolonged breast feeding, diarrhoeal disease, and survival of children in Guinea-
Bissau. British Medical Journal 308: 1403-1406.
Moller, I.J.
1982 Fluorides and dental fluorosis. Int’l Dentistry Journal 32: 135-147.
Molleson, T.
1995 The importance of porridge. In Nature et Culture, ed. by M. Otte, 479-486.
Colloque de Liege, Vol. 68. ERAUK, Liege.
Molnar, S.
1971 Human tooth wear, tooth function and cultural variability. Am. J. Phys.
Anthropol. 34: 175-189.
Molnar, S. and I.M. Molnar
1990 Dental arch shape and tooth wear variability. Am. J. Phys. Anthropol. 82: 385-
395.
Montgomery, J.P. Budd, A. Cox, P. Krause and R.G. Thomas
1998 LA-ICP-MS evidence for the distribution of lead and strontium in ancient and
modern human teeth: Implications for trace element and isotope analysis. Ms.,
University of Bradford.
Moorrees, C.F.A., E.A. Fanning and E.E. Hunt
1963 Age variation of formation stages for ten permanent teeth. J. Dental Research
42: 1490-1502.
Murray, M.J., A.B. Murray, N.J. Murray and M.B. Murray
1978 The effect of iron status of Nigerian mothers on that of their infants at birth and
6 months, and on the concentration of Fe in breast milk. Br. J. Nutr. 39: 627-
630.
Nations, J.D.
1979 Snail shells and maize preparation: A Lacandon Maya analogy. Am. Antiquity
44: 568-571.
Neel, J.V.
1977 Health and disease in unacculturated Amerindian populations. In Health and
Disease in Tribal Societies, Ciba Foundation Symposium No. 49, n.s., 155-168.
Elsevier/Excerpta Medica, Amsterdam.
Neff, H.
2003 Analysis of Mesoamerican plumbate pottery surfaces by laser ablation-
inductively coupled plasma-mass spectrometry (LA-ICP-MS). J. of
Archaeological Science 30: 21-35.

380
Nelson, B., M.J. DeNiro, M.J. Schoeninger and D.J. DePaolo
1983 Strontium isotope evidence for diagenetic alteration of bone: Consequences for
diet reconstruction. Geological Soc. of America Bulletin 95: 652.
Nelson, D.G.A.
1981 The influence of carbonate on the atomic structure and reactivity of
hydroxyapatite. J. Dent. Res. 60(C): 1621.
Nelson, P.A. and N.J. Sauer
1984 An evaluation of postdepositional changes in the trace element content of
human bone. Am. Antiquity 49: 141-147.
Newcomb, P.A., B.E. Storer, M.P. Longnecker, R. Mittendorf, E.G. Greenberg, R.W.
Clapp, K.P. Burke, W.C. Willett and B. MacMahon
1994 Lactation and a reduced risk of premenopausal breast cancer. New England J. of
Medicine 330(2): 81-87.
Newman, H.N. and D.F.G. Poole
1974 Observations with scanning and transmission electron microscopy on the
structure of human surface enamel. Archives Oral Biology 19: 1135-1143.
Newton, R.W., R.S. Levine, E.P. Turner and A.J. Barson
1984 The sensitivity of the developing tooth germ to systemic disturbances. Early
Human Development 9: 269-273.
Nikiforuk, G. and D. Fraser
1981 The etiology of enamel hypoplasia: A unifying concept. Journal of Pediatrics
98: 888-893.
Norr, L.
1981 Prehistoric Costa Rican diet as determined from stable carbon isotope ratios in
bone collagen. Am. J. Phys. Anthropol. 54: 258-259.
Odum, J.T.
1957 Strontium in natural waters. Texas Univ. Inst. Marine Sci. Publ. 4: 22-37.
Orams, H.J., J.J. Zybert, P.P. Phakey and W.A. Rachinger
1976 Ultrastructural study of human dental enamel using selected area argon-beam
thinning. Archives of Oral Biology 21: 663.
Orkney, J.M.
1946 The influence of feeding on infant mortality. Indian Medical Gazette 81: 150-
154.

381
Oski, F.A. and S.A. Landaw
1980 Inhibition of iron absorption from human milk by baby food. J. of Diseases of
Children 134: 459-460.
Outridge, P.M., G. Veinott and R.D. Evans
1995 Laser ablation ICP-MS analysis of incremental biological structures: Archives
of trace-element accumulation. Environmental Review 3: 160-170.
Outridge, P.M.
1996 Potential applications of laser ablation ICP-MS in forensic biology and
exploration geochemistry. Spectroscopy 11(4): 21-26.
Oviedo y Valdes, G.F. de
1535-47 Historia General y Natural de las Indias (1535-1547). 4 vols. Real Academia
de la Historia, Madrid.
Pabst, H.F. and D.W. Spady
1990 Effect of breast-feeding on antibody response to conjugate vaccine. Lancet 336:
269-270.
Parker, R.B. and H. Toots
1970 Minor elements in fossil bone. Geological Society of America Bulletin 81: 925-
32.
1974 Minor elements in fossil bone: application to Quaternary samples. Geological
Survey of Wyoming, Report of Investigations 10:74-77.
1980 Trace elements in bones as paleobiological indicators. In Fossils in the Making,
ed. by A.K. Behrensmeyer and A.P. Hill, 197-207. University of Chicago Press,
Chicago.
Pate, FD and KA Brown
1985 The stability of bone strontium in the geochemical environment. Journal of
Human Evolution 14:483-91.
Pate, F.D. and J.T. Hutton
1988 The use of soil chemistry data to address post-mortem diagenesis in bone
mineral. J. of Archaeological Science 15: 729-739.
Pendergast, D.M.
1981a Lamanai, Belize: Summary of excavation results, 1974-1980. Journal of Field
Archaeology 8 (1): 29-53.
1981b Lamanai, Belize: 1981 excavations. Mexicon 3(4):62-63.
1986 Stability through change: Lamanai, Belize, from the ninth to the seventeenth
century. In Late Lowland Maya Civilization: Classic to Postclassic, ed. by J.A.
Sabloff and E. Wyllys Andrews V, 223-249. University of New Mexico Press,
Albuquerque.

382
Pendergast, D.M.
1991a Up from the dust: The Central Lowlands Postclassic as seen from Lamanai and
Marco Gonzalez, Belize. In Vision and Revision in Maya Studies, ed. by F.
Clancy and P.D. Harrison, 169-177. University of New Mexico Press,
Albuquerque.
1991b The southern Maya lowlands contact experience: The view from Lamanai,
Belize. In Columbian Consequences, Vol. 3: The Spanish Borderlands in Pan-
American Perspective, ed. by D.H. Thomas, 337-354. Smithsonian Institution
Press, Washington, DC.
1992 Noblesse oblige: The elites of Altun Ha and Lamanai, Belize. In Mesoamerican
Elites: An Archaeological Assessment, ed. by D.Z. Chase and A.F. Chase, 61-
79. University of Oklahoma Press, Norman.
1993 Worlds in collision: The Maya/Spanish encounter in Sixteenth and Seventeenth
Century Belize. In The Meetings of Two Worlds: Europe and the Americas,
1492-1650, ed. by W. Bray, 105-143. Proceedings of The British Academy No.
81. Oxford University Press, Oxford, England.
Perez, V.R.
1998 Reconstruction of Prehistoric Weaning Practices from Sr and Ba: An Elemental
Analysis of Human Tooth Enamel. Unpublished Ms., in possession of the
author.
Perkins, W.T., R. Fuge and N.J. Pearse
1991 Quantitative analysis of trace elements in carbonates using laser ablation
inductively coupled plasma mass spectrometry. J. Anal. Atomic Spectrometry 6:
445-449.
Picciano, M.F. and B. Lonnerdal (eds)
1992 Mechanisms Regulating Lactation and Infant Nutrient Utilization. Wiley-Liss,
New York.
Piepenbrink, H.
1986 Two examples of biogenous dead bone decomposition and their consequences
for taphonomic interpretation. J. Arch. Science 13: 417-430.
Pindborg, J.J.
1982 Aetiology of developmental enamel defects not related to fluorosis. Int’l
Dentistry Journal 32: 123-134.
Pohl, M. (ed.)
1990 Ancient Maya Wetland Agriculture: Excavations on Albion Island, Northern
Belize. Westview Press, Boulder, CO.
Post, R.H.
1982 Breast cancer, lactation, and genetics. Social Biology 29: 357-386.

383
Powell, M.L.
1985 The analysis of dental wear and caries for dietary reconstruction. In The
Analysis of Prehistoric Diets, ed. by R.I. Gilbert and J.H. Mielke, 307-338.
Academic Press, Orlando.
Powis, T.G.
2001 The Preclassic whole vessels of Lamanai, Belize: A final report. Report
submitted to the Foundation for the Advancement of Mesoamerican Studies,
Inc. (FAMSI), April 2001.
Prentice, A.
1994 Extended breast-feeding and growth in rural China. Nutr. Review 52: 144-146.
Prentice, A. and D.V. Barclay
1991 Breast-milk calcium and phosphorus concentrations of mothers in rural Zaire.
Eur. J. Clin. Nutr. 45(12): 611-617.
Prentice, A., B. Dibba, L.M.A. Jarjou, M.A. Laskey and A.A. Paul
1994 Is breast-milk calcium-concentration influenced by calcium intake during
pregnancy? Lancet 344 (8919): 411-412.
Price, T.D.
1985 Late Archaic subsistence in the Midwestern United States. J. of Human
Evolution 14: 449-459.
Price, T.D.
1989a The Chemistry of Prehistoric Human Bone (ed.). Cambridge University Press,
Cambridge, England.
1989b Multi-element studies of diagenesis in prehistoric bone. In The Chemistry of
Prehistoric Human Bone, ed. by T.D. Price, 126-154. Cambridge University
Press, Cambridge.
Price, T.D., R.A. Bentley, J. Luning, D. Gronenborn and J. Wahl
2001 Prehistoric human migration in the Linearbandkeramik of Central Europe.
Antiquity 75: 593-603.
Price, T.D., J. Blitz, J. Burton and J.A. Ezzo
1992 Diagenesis in prehistoric bone: Problems and solutions. J. of Archaeological
Science 19: 513-529.
Price, T.D., M. Conner and J.D. Parsen
1985 Bone chemistry and the reconstruction of diet: Strontium discrimination in
white-tailed deer. J. of Archaeological Science 12: 419-442.

384
Price, T.D., C.M. Johnson, J.A. Ezzo, J.H. Burton and J. Ericson
1994 Residential mobility in the prehistoric southwest: A preliminary study using
strontium isotope analysis. J. of Archaeological Science 21: 315-330.
Price, T.D. and M. Kavanagh
1982 Bone composition and the reconstruction of diet: Examples from the
Midwestern United States. Journal of Archaeology 7: 61-80.
Price, T.D., L. Manzanilla and W.D. Middleton
2000 Immigration and the ancient city of Teotihuacan in Mexico: A study using
strontium isotope ratios in human bone and teeth. J. of Archaeological Science
27(10): 903-913.
Price, T.D., M.J. Schoeninger and G.J. Armelagos
1985 Bone chemistry and past behaviour: An overview. J. of Human Evolution 14:
419-47.
Price, T.D., R.W. Swick and E.P. Chase
1986 Bone chemistry and prehistoric diet: Strontium studies of laboratory rats. Am. J.
Phys. Anthropol. 70: 365-375.
Prohaska, T., C. Latkoczy, G. Schultheis, M. Teschler-Nicola and G. Stingeder
2002 Investigation of Sr isotope ratios in prehistoric human bones and teeth using
laser ablation ICP-MS and ICP-MS after Rb/Sr separation. J. of Analytical
Atomic Spectrometry 17(8): 887-891.
Puech, P.F., P. Dhamelincourt, M. Taieb and C. Serratrice
1986 Laser Raman microanalysis of fossil tooth enamel. J. Human Evolution 15(1):
13-20.
Puleston, D.E.
1982 The role of ramon in Maya subsistence. In Maya Subsistence: Studies in
Memory of Dennis E. Puleston, ed. by K.V. Flannery, 353-366. Academic
Press, New York.
Purvis, R.J., G.S. MacKay, F. Cockburn, W.J.M. Barrie, E.M. Wilkinson, N.R. Belton
and J.O. Forfar
1973 Enamel hypoplasia of the teeth associated with neonatal tetany: A manifestation
of maternal vitamin D deficiency. Lancet 2: 811.
Radosevich, S.C.
1993 The six deadly sins of trace element analysis: A case of wishful thinking in
science. In Investigations of Ancient Human Tissue: Chemical Analyses in
Anthropology, ed. by M.K. Sandford, 269-332. Gordon and Breach Science
Publishers, Langhorne, PA.

385
Rao, S. and A.N. Kanade
1992 Prolonged breast-feeding and malnutrition among rural Indian children below 3
years of age. Eur. J. Clin. Nutr. 46: 187-195.
Reed, D.M.
1994 Ancient Maya diet at Copan, Honduras, as determined through the analysis of
stable carbon and nitrogen isotopes. In Paleonutrition: The Diet and Health of
Prehistoric Americans, ed. By K.D. Sobolik, 210-221. Occasional Papers
Series, Center for Archaeological Investigations, Southern Illinois University,
Carbondale.
Rehnberg, G.L., A.B. Strong, C.R. Porter and M.W. Carter
1969 Levels of stable strontium in milk and the total diet. J. Environ. Sci. Technol. 3:
171-173.
Reid, D.J., A.D. Beynon and F.V.R. Rozzi
1998 Histological reconstruction of dental development in four individuals from a
medieval site in Picardie, France. J. Human Evol. 35: 463-477.
Reid, D.J. and M.C. Dean
2000 Brief communication: The timing of linear hypoplasias on human anterior teeth.
Am. J. Phys. Anthropol. 113(1): 135-139.
Reid, J.B. Jr., A.H. Goodman, J. Jones, D. Coleman, D. Walker, M. Blakey, M. Mack and
C. DeCorse
2001 Using 87Sr/86Sr in teeth as clues to the life histories of enslaved Africans
buried in New York City. Paper presented at the Geological Society of America
(GSA), 2001 Annual Meeting, Boston, MA.
Reitznerová, E., D. Amarasiriwardena, M. Kopčáková and R.M. Barnes
2000 Determination of some trace elements in human tooth enamel. Fresenius J.
Anal. Chem. 367: 748-754.
Restall, M.
1995 “He Wished it in Vain”: Subordination and resistance among Maya women in
post-Conquest Yucatan. Ethnohistory 42(4): 577-594.
1997 The Maya World: Yucatec Culture and Society, 1550-1850. Stanford University
Press, CA.
Rheingold, A.L., S. Hues and M.N. Cohen
1983 Strontium and zinc content in bones as an indication of diet. J. Chem. Ed. 60(3):
233-234.
Richards, M.P., S. Mays and B.T. Fuller
2002 Stable carbon and nitrogen isotope values of bone and teeth reflect weaning age
at the Medieval Wharram Percy site, Yorkshire, UK. Am. J. Phys. Anthropol.
119(3): 205-210.

386
Riordan, J. and K.G. Auerbach
1993 Breastfeeding and Human Lactation. Jones and Bartlett, Boston.
Ritchie, L.D., E.B. Fung, B.P. Halloran et al.
1998 A longitudinal study of calcium homeostasis during human pregnancy and
lactation and after the resumption of menses. Am. J. Clin. Nutr. 67: 693-701.
Rivera, J. and J.H. Harley
1965 The HASL bone program: 1961-1964. US Atomic Energy Commission Health
and Safety Lab Report, No. 163.
Rivera, J. and R. Martorell
1988a Nutrition, infection, and growth part I: Effects of infection on growth. Clinical
Nutrition 7(4): 156-162.
Rivera, J. and R. Martorell
1988b Nutrition, infection, and growth part II: Effects of malnutrition on infection and
general conclusions. Clinical Nutrition 7(4): 163-167.
Rivera, J. and M.T. Ruel
1997 Growth retardation starts in the first three months of life among rural
Guatemalan children. Eur. J. Clinical Nutrition 51: 92-96.
Rogan, W.J. and B.C. Gladen
1993 Breast-feeding and cognitive development. Early Human Development 31: 181-
193.
Rose, J.C.
1977 Defective enamel histology of prehistoric teeth from Illinois. Am. J. Phys.
Anthropol. 46: 439-446.
1979 Morphological variations of enamel prisms within abnormal striae of Retzius.
Human Biology 51: 139-151.
Rose, J.C., K.W. Condon and A.H. Goodman
1985 Diet and dentition: Developmental disturbances. In The Analysis of Prehistoric
Diets, ed. by R.I. Gilbert and J.H. Mielke, 281-305. Academic Press, Orlando.
Rosenthal, H.L.
1981 Content of stable strontium in man and animal biota. In Handbook of Common
Strontium, ed. by C. Skoryna, 503-514. Plenum, New York.
Rowles, S.L.
1967 Chemistry of the mineral phase of dentine. In Structural and Chemical
Organization of Teeth, vol. 2, ed. by A.E.W. Miles, 201-246. Academic Press,
London.

387
Roy, J. and L. Neufeld
2004 Laser ablation solid sampling for plasma spectrochemistry: The importance of
matching the hardware to the application. Spectroscopy 19(1): 16-28.
Roys, R.L.
1933 The Book of Chilam Balam of Chumayel. Carnegie Institution of Washington
Publication 438, Washington, DC.
1954 The Maya Katun Prophecies of the Books of Chilam Balam. Carnegie Institution
of Washington Publication 606, Washington, DC.
Runia, L.T.
1987a Analysis of bone from the Bronze-Age site Bovenkarspel-Het Valkje, The
Netherlands: A preliminary report. Archaeometry 1987: 221-232.
Runia, L.T.
1987b Strontium and calcium distribution in plants: Effect on palaeodietary studies. J.
of Archaeological Science 14: 599-608.
Rygh, O.
1949 Recherches sur les oligo-elements. I. De l’importance du strontium, du beryum
et du zinc. Bull Soc. Chim. Biol. 31: 1052-1057.
Sabin, A.B.
1963 Cause and control of fatal, infantile diarrheal diseases. Am. J. Tropical Medicine
and Hygiene 12(4): 556-579.
Sandford, M.K.
1993a Investigations of Ancient Human Tissue: Chemical Analyses in Anthropology
(editor). Gordon and Breach Science Publishers, Langhorne, PA
1993b Understanding the biogenic-diagenetic continuum: Interpreting elemental
concentrations of archaeological bone. In Investigations of Ancient Human
Tissue: Chemical Analyses in Anthropology, ed. by M.K. Sandford, 3-57.
Gordon and Breach Science Publishers, Langhorne, PA.
Sarnat, B.G. and I. Schour
1941 Enamel hypoplasias (chronologic enamel aplasia) in relation to systemic
disease: A chronologic, morphologic and etiologic classification. J. Am. Dental
Assoc. 28: 1989-2000.
Saul, F.P.
1972 The Human Skeletal Remains of Altar de Sacrificios. Papers of the Peabody
Museum of Archaeology and Ethnology Vol. 63, No. 2. Harvard University,
Cambridge, MA.
1975 Human remains from Lubaantun. In Lubaantun, ed. by N. Hammond, 389-410.
Peabody Museum of Archaeology and Ethnohistory, Monograph No. 2. Harvard
University, Cambridge, MA.

388
Saul, F.P.
1982 The human skeletal remains from Tancah, Mexico. Appendix II, in On the Edge
of the Sea: Mural Painting at Tancah-Tulum, Quintana Roo, Mexico, ed. by
A.G. Miller, 115-128. Dumbarton Oaks, Washington, D.C.
Saul, J.M. and F.P. Saul
1991 The Preclassic population of Cuello. In Cuello, An Early Maya Community in
Belize, ed. by N. Hammond, 134-158. Cambridge University Press, Cambridge.
1997 The Preclassic skeletons from Cuello. In Bones of the Maya: Studies of Ancient
Skeletons, ed. by S.L. Whittington and D.M. Reed, 28-50. Smithsonian
Institution Press, Washington.
Saunders, S.R. and L. Barrans
1999 What can be done about the infant category in skeletal samples? In Human
Growth in the Past: Studies from Bones and Teeth, ed. by R.D. Hoppa and C.M.
FitzGerald, 183-209. Cambridge University Press, Cambridge.
Saunders, S.R. and D.A. Herring (eds.)
1995 Grave Reflections: Portraying the Past through Cemetery Studies. Canadian
Scholars’ Press, Toronto, ON.
Schmidt, G.D. and L.S. Roberts
1981 Foundations of Parasitology. CV Mosby, St. Louis, MO.
Schneider, A.P.
1987 Risk factor for ovarian cancer. New England J. of Medicine 317(8): 508-509.
Schoeninger, M.J.
1979a Diet and status at Chalcatzingo: Some empirical and technical aspects of
strontium analysis. Am. J. Phys. Anthropol. 51: 295-310.
1979b Dietary Reconstruction at Chalcatzingo, A Formative Period Site in Morelos,
Mexico. Museum of Anthropology, Technical Reports, No. 9. University of
Michigan, Ann Arbor.
1981 The agricultural “revolution”: Its effect on human diet in prehistoric Iran and
Israel. Paleorient 7: 73-91.
1982 Diet and the evolution of modern human form in the Middle East. Am. J. Phys.
Anthropol. 58: 37-52.
Schoeninger, M.J. and M.J. DeNiro
1984 Nitrogen and carbon isotopic composition of bone collagen from marine and
terrestrial animals. Geochimica et Cosmochimica Acta 48: 625-639.
Schoeninger, M.J., M.J. DeNiro and H. Tauber
1983 Stable nitrogen isotope ratios of bone collagen reflect marine and terrestrial
components of prehistoric human diet. Science 220: 1381-1383.

389
Schoeninger, M.J. and C.S. Peebles
1981 Effect of mollusc eating on human bone strontium levels. J. Arch. Science 8:
391-397.
Schoeninger, M.J., N.J. van der Merwe, K. Moore, J. Lee-Thorp and C.S. Larsen
1990 Decrease in diet quality between the prehistoric and contact periods. In The
Archaeology of Mission Santa Catalina de Guale: 2. Biocultural Interpretations
of a Population in Transition, ed. by C.S. Larsen, 78-93. Anthropological
Papers of the American Museum of Natural History, 68.
Schour, I. And M.M. Hoffman
1939 Studies in tooth development. II. The rate of apposition of enamel and dentin in
man and other animals. J. of Dental Research 18: 161-175.
Schour, I. and M. Massler
1940 Studies in tooth development: The growth pattern of human teeth, Part II. J. Am.
Dental Assoc. 27: 1918-1931.
Schroeder, H.A., I.H. Tipton and A.P. Nason
1972 Trace metals in man: Strontium and barium. J. of Chronic Diseases 25: 491-
517.
Schroeder, S., S. Wood and R. Haskett (eds)
1997 Indian Women of Early Mexico. University of Oklahoma Press, Norman.
Schurr, M.R.
1997 Stable nitrogen isotopes as evidence for the age of weaning at the Angel site: A
comparison of isotopic and demographic measures of weaning age. J. of
Archaeological Science 24: 919-927.
1998 Using stable nitrogen isotopes to study weaning behavior in past populations.
World Archaeology 30: 327-342.
Schwartz, G.T., D.J. Reid and M.C. Dean
1999 Dental development and canine dimorphism in the common chimpanzee, Pan
troglodytes. Am. J. Phys. Anthropol. Suppl. 28: 246.
2001 Developmental aspects of sexual dimorphism in hominoid canines. Int’l J. of
Primatology 22(5): 837-860.
Scott, G.R. and C.G. Turner II
1997 The Anthropology of Modern Human Teeth: Dental Morphology and its
Variation in Recent Human Populations. Cambridge University Press,
Cambridge, England.
Scott, J.H. and N.B.B. Symons
1974 Introduction to Dental Anatomy, 7th
ed. Churchill Livingston, Edinburgh.

390
Scrimshaw, N. S.
1978 Synergism of malnutrition and infection. Evidence from field studies in
Guatemala. J. Am. Med. Assoc. 212: 1685.
Scrimshaw, N.S., C.E. Taylor and J.E. Gordon
1968 Interactions of nutrition and infection. World Health Organization, Geneva.
Sealy, J.C., R. Armstrong and C. Schrire
1995 Beyond lifetime averages: Tracing life histories through isotopic analysis of
different calcified tissues from archaeological human skeletons. Antiquity 69:
290-300.
Sealy, J.C. and A. Sillen
1988 Sr and Sr/Ca in marine and terrestrial foodwebs in the Southwestern Cape,
South Africa. J. Archaeological Science 15: 425-438.
Sealy, J.C., N.J. van der Merwe, J.A. Lee Thorp and J.L. Lanham
1987 Nitrogen isotopic ecology in southern Africa: Implications for environmental
and dietary tracing. Geochimica et Cosmochimica Acta 51: 2701-17.
Shacklette, H.T.
1980 Elements in Fruits and Vegetables From Areas of Commercial Production in
the Conterminous United States. US Geological Survey Professional Paper No.
1178. U.S. Government Printing Office, Washington, D.C.
Shafer, W.G., M.K. Hines and B.M. Levy
1983 A Textbook of Oral Pathology, 4th
edition. Saunders, Philadelphia.
Sharon, I.M.
1988 The significance of teeth in pollution detection. Perspectives Biol. Med. 24:
124-131.
Shaw, L.C.
1991 The Articulation of Social Inequality and Faunal Resource Use in the Preclassic
Community of Colha, Northern Belize. Ph.D. dissertation, University of
Massachusetts, Amherst.
Shipman, P., G. Foster and M. Schoeninger
1984 Burnt bones and teeth: An experimental study of color, morphology, crystal
structure and shrinkage. J. of Archaeological Science 11: 307-325.
Shorr E and A.C. Carter
1952 The usefulness of Strontium as an adjuvant to calcium in the remineralization of
the skeleton in man. Bull Hosp Joint Dis Apr 13(1): 59-66.

391
Short, R.
1983 The biological basis for the contraceptive effects of breastfeeding. In Advances
in International Maternal and Child Health, Vol. 3, ed. by D.B. Jelliffe and
E.F.P. Jelliffe, 27-39. Oxford University Press, Oxford, England.
Siimes, M.A., L. Salmenpera and J. Perheentupa
1984 Exclusive breast-feeding for 9 months: Risk of iron deficiency. J. Pediatr. 104:
196-199.
Sillen, A.
1981a Strontium and diet at Hayonim Cave. Am. J. Phys. Anthropol. 56: 131-137.
Sillen, A.
1981b Post-depositional changes in Natufian and Aurignacian faunal bones from
Hayonim Cave. Paleorient 7:81-85.
1986 Biogenic and diagenic Sr/Ca in Plio-Pleistocene fossils of the Omo Shungura
Formation. Paleobiology 12: 311-323.
1989 Diagenesis of the inorganic phase of cortical bone. In The Chemistry of
Prehistoric Bone, ed. by T.D. Price, 211-229. Cambridge University Press,
Cambridge.
1992 Strontium-calcium ratios (Sr/Ca) of Australopithecus robustus and associated
fauna from Swartkrans. J. Human Evolution 23: 495-516.
Sillen, A. and M. Kavanagh
1982 Strontium and paleodietary research: A review. Yearbook of Physical
Anthropology 25: 67-90.
Sillen, A. and J.A. Lee-Thorp
1994 Trace element and isotope aspects of predator-prey relationships in terrestrial
foodwebs. Palaeogeography, Palaeoclimatology, Palaeocology 107: 243-255.
Sillen, A. and J.C. Sealy
1995 Diagenesis of strontium in fossil bone: A reconsideration of Nelson et al.
(1986). J. of Archaeological Science 22: 313-320.
Sillen, A., J.C. Sealy and N.J. van der Merwe
1989 Chemistry and paleodietary research: No more easy answers. American
Antiquity 54: 504-512.
Sillen, A. and P. Smith
1983 Strontium calcium ratios reveal weaning age in a skeletal population. Am. J.
Phys. Anthropol. 60(2): 253-254.
1984 Weaning patterns are reflected in strontium-calcium ratios of juvenile skeletons.
J. of Archaeological Science 11: 237-245.

392
Simondon, K.B. and F. Simondon
1998 Mothers prolong breastfeeding of undernourished children in rural Senegal. Int.
J. Epidemiol. 27: 490-494.
Skinner, M.F. and A.H. Goodman
1992 Anthropological uses of developmental defects of enamel. In Skeletal Biology of
Past Peoples: Research Methods, ed. by S.R. Saunders and M.A.
Katzenberg,153-174. Wiley-Liss, New York.
Skougstad, M.W. and C.A. Horr
1960 Occurrence and distribution of strontium in natural water. Geological Survey
Circular 420. United States Geological Service, Washington, DC.
1963 Occurrence and distribution of strontium in natural water. United States
Geological Survey Water Supply Paper no. 1496-D.
Smith, B.H.
1989 Dental development as a measure of life history in primates. Evolution 43(3):
683-688.
1991a Age of weaning approximates age of emergence of the first permanent molar in
nonhuman primates (abstract). Am. J. Phys. Anthropol. Supplement 12: 163-
164.
1991b Dental development and the evolution of life history in the Hominidae. Am. J.
Phys. Anthropol. 86:157-174.
1992 Life history and the evolution of human maturation. Evolutionary Anthropology
1(4): 134-142.
Smith, D.T. and W.F. Panton
1986 On the climate of Belize and some implications for national economic
development. Belcast J. Belizean Affairs 3(1 and 2): 47-51.
Song, R.J.
1997 Developmental Defects of Enamel in the Maya of Altun Ha, Belize:
Implications for Ancient Maya Childhood Health. M.A. Thesis, Trent
University, Peterborough, ON.
Song, R.J. and A.H. Goodman
1999 Inferences for childhood nutrition from laser ablation inductively coupled
plasma-mass spectrometry (LA-ICP-MS) of dental hard tissues. Paper presented
at the 68th Annual Meetings of the American Association of Physical
Anthropologists, Columbus, OH.
Song, R.J. and T. Stillman
1998 Apples and oranges? Testing acid dissolution and laser ablation sampling ICP-
MS and the implications for chemical analyses of teeth (Research paper, Natural
Science, Hampshire College). Ms. in possession of the author.

393
Sowden, E.M. and S.R. Stitch
1957 Trace elements in human tissue. Biochem. J. 67: 104-109.
Sowers M.F., G. Corton and B. Shapiro
1993 Changes in bone density with lactation. J. Am. Med. Assoc. 269: 3130–5.
Sowers M.F., D. Eyre and B.W. Hollis
1995 Biochemical markers of bone turnover in lactating and nonlactating postpartum
women. J. Clin. Endocrinol. Metab. 80: 2210–6.
Spencer, H., M. Brothers, E. Berger, H.E. Hart and D. Laszlo
1956 Strontium85
metabolism in man and effect of calcium on strontium excretion.
Society of Experimental Biology and Medicine: New York Proceedings 91: 155-
157.
Spencer, H., M. Li, J. Samachson and D. Laszlo
1960 Metabolism of strontium-85 and calcium-45 in man. Metabolism 9: 916-925.
Spencer, H., M. Li and J. Samachson
1961 Effect of high calcium intake on strontium85
metabolism in man. J. Clin. Invest.
40: 1339-1344.
Spencer, H., J.M. Warren, L. Kramer and J. Samachson
1973 Passage of calcium and strontium across the intestine in man. Clinical
Orthopaedics and Related Research 91: 225-234.
Standley, P.C.
1961 Trees and Shrubs of Mexico. Contribution from the US National Herbarium,
Vol. 23. Smithsonian Institute, Washington, DC.
Stannard, D.E.
1990 Disease and infertility: A new look at the demographic collapse of native
populations in the wake of western contact. J. of American Studies 24(3): 1-26.
1991 The consequences of contact: Toward an interdisciplinary theory of native
responses to biological and cultural invasion. In Columbian Consequences, vol.
3, The Spanish Borderlands in Pan-American Perspective, ed. by D.H. Thomas,
519-540. Smithsonian Institution Press, Washington, DC.
Stead, I.M., J.B. Bourke and D.R. Brothwell (eds)
1986 Lindow Man. The Body in the Bog. British Museum Publications, London.
Steadman, L.T., F. Brudevold and F.A. Smith
1958 Distribution of strontium in teeth from different geographic areas. J. Am. Dent.
Assoc. 57: 340.

394
Storey, E.
1961 Strontium rickets, bone calcium and strontium changes. Aust. Ann. Med. 10:
213-333.
1992 Patterns of susceptibility to dental defects in the deciduous dentition of a
Precolumbian skeletal population. In Recent Contributions to the Study of
Enamel Developmental Defects, ed. by A.H. Goodman and L.L. Capasso, 171-
183. J. of Paleopathology, Monographic Publication 2. Associazione
Antropologica Abruzzese, Chieti, Italy.
Straub, C.P., B. Kahn and N.C. Telles
1961 Health Physics Society Meetings, 1961.
Stuart-Macadam, P.L.
1985 Porotic hyperostosis: Representative of a childhood condition. Am. J. Phys.
Anthropol. 66: 391-398.
1993 Replacing breasts with bottles: The biological consequences (abstract).
Abstract from the 13th
International Congress of Anthropological and
Ethnological Sciences. The Cultural and Biological Dimensions of Global
Change, p. 438. IUAES meetings, Mexico City, August.
1995a Biocultural perspectives on breastfeeding. In Breastfeeding: Biocultural
Perspectives, ed. by P.L. Stuart-Macadam and K.A. Dettwyler, 1-38. Aldine de
Gruyter, New York.
1995b Breastfeeding in prehistory. In Breastfeeding: Biocultural Perspectives, ed. by
P.L. Stuart-Macadam and K.A. Dettwyler, 75-100. Aldine de Gruyter, New
York.
Stuart-Macadam, P.L. and K.A. Dettwyler (eds)
1995 Breastfeeding: Biocultural Perspectives. Aldine de Gruyter, New York.
Suga 1982
1982 Progressive mineralization pattern of developing enamel during the maturation
stage. J. Dental Research 61: 1532-1542.
Sullivan, C.H. and H.W. Krueger
1981 Carbon isotope analysis of separate chemical phases in modern and fossil bone.
Nature 292: 333-335.
Sultan, A.N. and R.W. Zuberi
2003 J. of Ayub Medical College (JAMC) 15(2): 3-7.
Swardstedt, T.
1966 Odontological Aspects of a Medieval Population in the Province of
Jamtland/Mid-Sweden. Tiden-Barnangen AB, Tryckerier/Stockholm.

395
Swedlund, A.C.
1990 Infant mortality in Massachusetts and the United States in the nineteenth
century. In Disease in Populations in Transition, ed. by A.C. Swedlund and G.J.
Armelagos, 161-182. Bergin and Garvey, New York.
Swedlund, A.C. and H. Ball
1998 Nature, nurture, and the determinants of infant mortality: A case study from
Massachusetts, 1830-1920. In Building a New Biocultural Synthesis: Political-
Economic Perspectives on Human Biology, ed. by A.H. Goodman and T.L.
Leatherman, 191-228. University of Michigan Press, Ann Arbor.
Sweeney, E.A. and M. Guzman
1966 Oral conditions in children from three highland villages in Guatemala. Archives
of Oral Biology 11: 687-698.
Sweeney, E.A., J. Cabrera, J. Urritia and L. Mata
1969 Factors associated with linear hypoplasia of human deciduous incisors. J.
Dental Research 48: 1275-1279
Szpunar, C.
1977 Atomic Absorption of Archaeological Remains: Human Ribs from Woodland
Mortuary Sites. Ph.D. Dissertation. Northwestern University, Evanston, IL.
Szpunar, C., J.B. Lambert and J.E. Buikstra
1978 Analysis of excavated bone by atomic absorption. Am. J. Phys. Anthropol. 48:
199-202.
Tanaka, G., H. Kawamura and E. Nomura
1981 Reference Japanese man II: Distribution of strontium in the skeleton and in the
mass of mineralized bone. Health Physics 40: 601-614.
Taren, D. and J. Chen
1993 A positive relationship between extended breastfeeding and nutritional status in
rural Hubei Province, People’s Republic of China. Am. J. Clin. Nutr. 58: 862-
867.
Taylor, D.M., P. Bligh and M. Duggan
1962 The absorption of calcium, strontium, barium, and radium from the
gastrointestinal tract of the rat. Biochem. J. 83: 25-29.
Teaford, M.F.
1991 Dental microwear: What can it tell us about diet and dental function? In
Advances in Dental Anthropology, ed. by M.A. Kelley and C.S. Larsen, 341-
356. Wiley-Liss, New York.

396
Temboury, M.C., A. Otero, I. Polanco and E. Arribas
1994 Influence of breastfeeding on the infant’s intellectual development. J. of
Pediatric Gastroenterology and Nutrition 18: 32-36.
TenCate, A.R.
1980 Oral Histology: Development, Structure and Function. CV Mosby, St. Louis.
Thapa, S., R.V. Short and M. Potts
1988 Breast feeding, birth spacing and their effects on child survival. Nature 335:
679-682.
Thompson, J.E.S.
1938 Sixteenth- and Seventeenth-Century Reports on the Chol Mayas. American
Anthropologist 40: 584-604.
Thompson, R.C.
1963 Factors and conditions modifying the absorption and retention of chronically
ingested radiostrontium. In The Transfer of Calcium and Strontium Across
Biological Membranes, ed. by R.H. Wasserman, 393-404. Academic Press,
New York.
Thornthwaite, C.W.
1948 An approach toward a rational classification of climate. Geographical Review
38: 55-94.
Thurber, D.L., J.L. Kulp, E. Hodges, P.W. Gast and J.M. Wampler
1958 Common strontium content of the human skeleton. Science 128: 256-257.
Toots, H. and M.R. Voorhies
1965 Strontium in fossil bones and the reconstruction of food chains. Science 149:
854-855.
Tozzer, A.M. (ed.)
1941 Landa’s Relacion de las Cosas de Yucatan. Papers of the Peabody Museum of
Archaeology and Ethnology, Harvard University, Vol. 18. Harvard University
Press, Cambridge, MA.
Tremlett, G.
2003 Tracing Adam. The Guardian Newspaper (U.K.), Aug. 7, 2003.
Tunstall, S.D. and D. Amarasiriwardena
2002 Characterization of lead and lead leaching properties of lead glazed ceramics
from the Solis Valley, Mexico, using inductively coupled plasma-mass
spectrometry (ICP-MS) and diffuse reflectance infrared Fourier transform
spectroscopy (DRIFT). Microchemical Journal 73: 335-347.

397
Turekian, K.K. and J.L. Kulp
1956 Strontium content of human bone. Science 128: 405-407.
Turner, B.L. II and P.D. Harrison (eds.)
1983 Pulltrouser Swamp: Ancient Maya Habitat, Agriculture and Settlement in
Northern Belize. University of Texas Press, Austin.
Turner, B.L. II and C.H. Miksicek
1984 Economic plant species associated with prehistoric agriculture in the Maya
lowlands. Economic Botany 38: 179-193.
Turner, C.G., II, C.R. Nichol and G.R. Scott
1991 Scoring procedures for key morphological traits of the permanent dentition: The
Arizona State University Dental Anthropology System. In Advances in Dental
Anthropology, ed. by M.A. Kelley and C.S. Larsen, 13-31. Wiley-Liss, New
York.
Tuross, N. and M. Fogel
1994 Stable isotope analysis and subsistence patterns at the Sully Site. In Skeletal
Biology in the Great Plains: Migration, Warfare, Health, and Subsistence, ed.
by D.W. Owsley and R.L. Jantz, 283-289. Smithsonian Institution Press,
Washington, DC.
Twardock, A.R.
1963 Studies on the movement of calcium and strontium across the bovine mammary
gland. In The Transfer of Calcium and Strontium Across Biological Membranes,
ed. by R.H. Wasserman, 327-340. Academic Press, New York.
Ubelaker, D.H.
1989 Human Skeletal Remains: Excavation, Analysis, Interpretation, 2nd
edition.
Manuals on Archeology 2. Taraxacum, Washington, D.C.
Underwood, E.J.
1977 Trace Elements in Human and Animal Nutrition. Academic Press, New York.
United Kingdom Medical Research Council (1959-1970)
1959-70 Assay of strontium-90 in human bone in the United Kingdom. Monitoring
Reports no. 1-19. HMSO, London.
Vail, G. and A. Stone
2002 Representations of women in Postclassic and Colonial Maya literature and art.
In Ancient Maya Women, ed. by T. Ardren, 203-228. Altamira Press, New
York.

398
van der Merwe, N.J.
1989 Natural variation in 13
C concentration and its effect on environmental
reconstruction using 13
C/12
C ratios in animal bones. In The Chemistry of
Prehistoric Human Bone, ed. by T.D. Price, 105-125. Cambridge University
Press, Cambridge.
van der Merwe, N.J. and J.C. Vogel
1978 13
C content of human collagen as a measure of prehistoric diet in Woodlan
North America. Nature 276: 815-816.
Varner, J.G. and J.J. Varner
1983 Dogs of the Conquest. University of Oklahoma Press, Norman.
Verano, J.W. and D.H. Ubelaker (eds.)
1992 Disease and Demography in the Americas. Smithsonian Institution Press,
Washington, DC.
Villagutierre Soto-Mayor, J. de
1983 History of the Conquest of the Province of the Itza. Culver City, CA:
Labyrinthos.
Villalpando, S., N.F. Butte, S. Flores-Huerta and M. Thotathuchery
1998 Qualitative analysis of human milk produced by women consuming a maize-
predominant diet typical of rural Mexico. In Ann. Nutr. Metab 42: 23-32.
Vogel, J.C. and N.J. van der Merwe
1977 Isotopic evidence for early maize cultivation in New York State. American
Antiquity 42: 238-42.
Vrbič, V., J. Stupar and A.R. Byrne
1987 Trace element content of primary and permanent tooth enamel. Caries Research
21: 37-39.
Vuorinen H.S., S. Pihlman, H. Mussalo Rauhamaa, U. Tapper and T. Varrela
1996 Trace and heavy metal analyses of a skeletal population representing the town
people in Turku (Abo) Finland in the 16th-17th centuries: With special
reference to gender, age and social background. Science of the Total
Environment 177: 145-160.
WHO (World Health Organization)
1981 Contemporary Patterns of Breast-Feeding. Report on the WHO Collaborative
Study on Breast-Feeding. World Health Organization, Geneva.
Waldron, H.A.
1981 Postmortem absorption of lead by the skeleton. Am. J. Phys. Anthropol. 55:
395-398.

399
Waldron, H.A.
1983 On the post-mortem accumulation of lead by skeletal tissues. J. Arch. Science
10: 35-40.
Walker-Smith, J.A. and A.S. McNeish (eds.)
1986 Diarrhea and Malnutrition in Childhood. Butterworth, London.
Walser, M. and B. Robinson
1963 Renal excretion and tubular reabsorption of calcium and strontium. In The
Transfer of Calcium and Strontium Across Biological Membranes, ed. by R.H.
Wasserman, 305-326. Academic Press, New York.
Wang, M.S., W.T. Cave and W.S. Coakley
1972 Chapt. 8, in Analytical Emission Spectroscopy, Vol. 1, Part II, ed. by E.L.
Grove. Dekker, New York.
Wang, S., R. Brown and D.J. Gray
1994 Application of laser ablation-ICPMS to the spatially resolved micro-analysis of
biological tissue. Applied Spectroscopy 48(11): 1321-1325.
Ward, N.I., S.F. Durrant and A.L. Gray
1992 Analysis of biological standard reference materials by laser ablation inductively
coupled plasma mass spectrometry. J. of Analytical Atomic Spectrometry 7:
1139-1146.
Wasserman, R.H. (ed.)
1963 The Transfer of Calcium and Strontium Across Biological Membranes.
Academic Press, New York.
Wasserman, R.H. and C.L. Comar
1960 Effects of dietary calcium and phosphorous levels on body burdens of ingested
radiostrontium. Proc. Soc. Exper. Biol. Med. 103: 124-129.
Wasserman, R.H., C.L. Comar and M.M. Nold
1956 The influence of amino acids and other organic compounds on the
gastrointestinal absorption of calcium45
and strontium89
in the rat. Journal of
Nutrition 59: 371-383.
Wasserman, R.H., F.W. Lengeman and C.L. Comar
1958 Comparative metabolism of calcium and strontium in lactation. J. of Dairy
Science 41: 812-821.
Wasserman, R.H., E.M. Romney, M.W. Skougstad and R. Siever
1977 Strontium in Geochemistry and the Environment, Vol. II. The Relation of Other
Selected Trace Elements and Disease. National Academy of Sciences,
Washington, D.C.

400
Watling, R.J.
1999 Novel application of laser ablation inductively coupled plasma mass
spectrometry in forensic science and forensic archaeology. Spectroscopy 14(6):
16-34.
Watt, B.K. and A.L. Merril
1963 Composition of Foods. U.S. Department of Agriculture Handbook No. 8. U.S.
Government Printing Office, Washington, D.C.
Watmough, S.A., T.C. Hutchinson and R.D. Evans
1997 Application of laser ablation inductively coupled plasma-mass spectrometry in
dendrochemical analysis. Environmental Science Technology 31: 114-118.
Weatherell, J.A., S.M. Weidmann and S.M. Hamm
1967 Density patterns in enamel. Caries Research 1: 42-51.
Weinberg, E.D.
1994 Role of iron in sudden infant death syndrome. J. of Trace Elements in
Experimental Medicine 7: 47-51.
Wessen, G., F.H. Ruddy, C.E. Gustafson and H. Irwin
1978 Trace element analysis in the characterization of archaeological bone. In
Archaeological Chemistry, ed. By G.F. Carter, 99-108. American Chemical
Society, Pullman, WA.
White, C.D.
1985 Paleoepidemiology of the Pre-Columbian Maya. Ms on file, Dept. of
Anthropology, Trent University, Peterborough, ON.
1986 Paleodiet and Nutrition of the Ancient Maya at Lamanai, Belize: A Study of
Trace Elements, Stable Isotopes, Nutritional and Dental Pathology. M.A. thesis,
Dept. of Anthropology, Trent University, Peterborough, ON.
1988 The ancient Maya from Lamanai, Belize: Diet and health over 2,000 years. Can.
Rev. of Phys. Anthropol. 6(2): 1-21.
1997 Ancient diet a Lamanai and Pacbitun: Implications for the ecological model of
collapse. In Bones of the Maya: Studies of Ancient Skeletons, ed. by S.L.
Whittington and D.M. Reed, 171-180. Smithsonian Institution Press,
Washington.
White, C.D. (ed.)
1999 Reconstructing Ancient Maya Diet. University of Utah Press, Salt Lake City.
White, C.D., P.F. Healy and H.P. Schwarcz
1993 Intensive agriculture, social status and Maya diet at Pacbitun, Belize. Journal of
Anthropological Research 49: 347-375.

401
White, C.D. and H.P. Schwarcz
1989 Ancient Maya diet: As inferred from isotopic and elemental analysis of human
bone. Journal of Archaeological Science 16: 451-474.
1994 Temporal trends in stable isotopes for Nubian mummy tissue. Am. J. Phys.
Anthropol. 93: 165-187.
White, C.D., L.E. Wright and D.M. Pendergast
1994 Biological disruption in the early colonial period at Lamanai. In In the Wake of
Contact: Biological Responses to Conquest, ed. by C.S. Larsen and G.R.
Milner, 135-145. Wiley Liss, New York.
White, E.M. and L.A. Hannus
1983 Chemical weathering of bone in archaeological soils. Am. Antiquity 48: 316-
322.
Whitehead, R.G. (ed.)
1983 Maternal Diet, Breastfeeding Capacity and Lactational Infertility. Report of a
joint UNU/WHO workshop held in Cambridge, UK, 9-11 March, 1981. United
Nations University, Food and Nutrition Bulletin, Suppl. 6, Tokyo.
Whitehead, R.G. and A.A. Paul
1984 Growth charts and the assessment of infant feeding practices in the western
world and in developing countries. Early Human Development 9: 187-207.
Whittington, S.L.
1991 Detection of significant demographic differences between subpopulations of
Prehispanic Maya from Copan, Honduras, by survival analysis. Am. J. Phys.
Anthropol. 85: 167-184.
1992 Enamel hypoplasia in the low status Maya population of prehispanic Copan,
Honduras. In Recent Contributions to the Study of Enamel Developmental
Defects, ed. by A.H. Goodman and L.L. Capasso, 185-205. J. of
Paleopathology, Monographic Publications 2. Edigrafital Teramo, Chieti, Italy.
Whittington, S.L. and D.M. Reed
1997 Bones of the Maya: Studies of Ancient Skeletons. Smithsonian Institution Press,
Washington, D.C.
Wickes, I.G.
1953 A history of infant feeding. Archives of Diseases of Childhood Part I, 28: 151-
158, Part III: 332-340.
Wing, E.S. and S.J. Scudder
1991 The exploitation of animals. In Cuello: An Early Maya Community in Belize,
ed. by N. Hammond, 84-97. Cambridge University Press, Cambridge, England.

402
Winkler, L.A.
1995 A comparison of radiographic and anatomic evidence of tooth development in
infant apes. Folia Primatol. 65: 1-13.
Wolf, N., I. Gedalia and S. Yariv and H. Zuckermann
1973 The strontium content of bones and teeth of human fetuses. Arch. Oral Biology
18: 233-238.
Wolfsperger, M.
1993 Trace element analysis of Medieval and early modern skeletal remains from
western Austria for reconstruction of diet. Homo 43(3): 278-294.
Wood, C.S.
1983 Early childhood, the critical stage in human interactions with disease and
culture. Soc. Sci. Med. 17(2): 79-85.
Wood, J.W., G.R. Milner, H.C. Harpending and K.M. Weiss
1992 The osteological paradox: Problems of inferring prehistoric health from skeletal
samples. Current Anthropology 33: 343-370.
Wood, L.
1996 Frequency and chronological distribution of linear enamel hypoplasia in a North
American colonial skeletal sample. Am. J. Phys. Anthropol. 100: 247-260.
Wright, A.C.S., D.H. Romney, R.H. Arbuckle and V.E. Vial
1959 Land in British Honduras. Report of the British Honduras Land Use Survey
Team, ed. by D.H. Romney. Colonial Research Publications No. 24. H.M.
Stationary Office, London.
Wright, L.E.
1990 Stresses of conquest: A study of Wilson bands and enamel hypoplasias in the
Maya of Lamanai, Belize. Am. J. of Human Biology 2: 25-35.
1994 The Sacrifice of the Earth? Diet, Health, and Inequality in the Pasion Maya
Lowlands. PhD dissertation, Dept. of Anthropology, University of Chicago,
Chicago.
1997 Ecology or society? Paleodiet and the collapse of the Pasion Maya Lowlands. In
Bones of the Maya: Skeletal Studies of an Ancient People, ed. by S. Whittington
and D. Reed, 181-195. Smithsonian Institution Press, Washington, DC.
1999 The elements of Maya diets: Alkaline earth baselines and paleodietary
reconstruction in the Pasion region. In Reconstructing Ancient Maya Diet, ed.
By C.D. White, 197-220. University of Utah Press, Salt Lake City, UT.
Wright, L.E. and H.P. Schwarcz
1998 Stable carbon and oxygen isotopes in human tooth enamel: Identifying
breastfeeding and weaning in prehistory. Am. J. Phys. Anthropol. 106: 1-18.

403
Wright, L.E. and C.D. White
1996 Human biology in the Classic Maya collapse: Evidence from paleopathology
and paleodiet. Journal of World Prehistory 10 (2): 147-198.
Wyatt, R.G. and L.J. Mata
1969 Bacteria in colostrum and milk of Guatemalan Indian women. J. Trop. Pediatr.
15: 159-162.
Wyckoff, R.W.G. and A.R. Doberenz
1968 The strontium content of fossil teeth and bones. Geochim. Cosmochim. Acta 32:
109-115.
Yaeger, J.
1980 Enamel. In Orban’s Oral Histology and Embryology, 9th
edition, ed. by S.N.
Bhaskar, 46-106. CV Mosby, St. Louis.

404




















